
Stuttering: a dynamic motor control disorder
23
Journal of Fluency Disorders 28 (2003) 273–295 Stuttering: a dynamic motor control disorder Christy L. Ludlow ∗ , Torrey Loucks Laryngeal and Speech Section, Clinical Neurosciences Program, National Institute of Neurological Disorders and Stroke, 10 Center Drive MSC 1416, Bldg. 10 Rm. 5D38, Bethesda, MD 20892-1416, USA Received 22 July 2003; accepted 29 July 2003 Abstract The purpose of this review is to determine what neural mechanisms may be dysfunc- tional in stuttering. Three sources of evidence are reviewed. First, studies of dynamic inter-relationships among brain regions during normal speech and in persons who stut- ter (PWS) suggest that the timing of neural activity in different regions may be abnormal in PWS. Second, the brain lesions associated with acquired stuttering are reviewed. These indicate that in a high percentage of cases, the primary speech and language regions are not affected but lesions involve other structures, such as the basal ganglia, which may modulate the primary speech and language regions. Third, to characterize the motor control disorder in stuttering, similarities and differences from focal dystonias such as spasmodic dysphonia (SD) and Tourette’s syndrome (TS) are reviewed. This review indicates that the central control abnormalities in stuttering are not due to disturbance in one particular brain region but rather a system dysfunction that interferes with rapid and dynamic speech processing for production. Educational objectives: The reader will be able to describe: (1) the similarities and differences between stuttering and other speech motor control disorders, (2) which brain lesions are most likely to produce acquired stuttering in adults, and (3) what type of brain abnormality most likely underlies stuttering. © 2003 Elsevier Inc. All rights reserved. Keywords: Brain lesions; Dystonia; Task specific; Spasmodic dysphonia ∗ Corresponding author. Tel.: +1-301-496-9366; fax: +1-301-480-0803. E-mail address: [email protected] (C.L. Ludlow). 0094-730X/$ – see front matter © 2003 Elsevier Inc. All rights reserved. doi:10.1016/j.jfludis.2003.07.001
Transcript of Stuttering: a dynamic motor control disorder

Journal of Fluency Disorders28 (2003) 273–295
Stuttering: a dynamic motor control disorder
Christy L. Ludlow∗, Torrey LoucksLaryngeal and Speech Section, Clinical Neurosciences Program, National Institute
of Neurological Disorders and Stroke, 10 Center Drive MSC 1416,Bldg. 10 Rm. 5D38, Bethesda, MD 20892-1416, USA
Received 22 July 2003; accepted 29 July 2003
Abstract
The purpose of this review is to determine what neural mechanisms may be dysfunc-tional in stuttering. Three sources of evidence are reviewed. First, studies of dynamicinter-relationships among brain regions during normal speech and in persons who stut-ter (PWS) suggest that the timing of neural activity in different regions may be abnormalin PWS. Second, the brain lesions associated with acquired stuttering are reviewed. Theseindicate that in a high percentage of cases, the primary speech and language regions are notaffected but lesions involve other structures, such as the basal ganglia, which may modulatethe primary speech and language regions. Third, to characterize the motor control disorderin stuttering, similarities and differences from focal dystonias such as spasmodic dysphonia(SD) and Tourette’s syndrome (TS) are reviewed. This review indicates that the centralcontrol abnormalities in stuttering are not due to disturbance in one particular brain regionbut rather a system dysfunction that interferes with rapid and dynamic speech processingfor production.
Educational objectives: The reader will be able to describe: (1) the similarities anddifferences between stuttering and other speech motor control disorders, (2) which brainlesions are most likely to produce acquired stuttering in adults, and (3) what type of brainabnormality most likely underlies stuttering.© 2003 Elsevier Inc. All rights reserved.
Keywords: Brain lesions; Dystonia; Task specific; Spasmodic dysphonia
∗ Corresponding author. Tel.:+1-301-496-9366; fax:+1-301-480-0803.E-mail address: [email protected] (C.L. Ludlow).
0094-730X/$ – see front matter © 2003 Elsevier Inc. All rights reserved.doi:10.1016/j.jfludis.2003.07.001

274 C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295
1. Purpose
Our purpose is to identify whether particular neural mechanisms are implicatedby the current literature for characterizing stuttering as a neurodevelopmental mo-tor control disorder. Because speech is a dynamic motor function process, it re-quires a highly synchronized and adaptive network of neural activity in order tofunction without disruption. We will review the following topics in attemptingto fashion an improved understanding of the underlying mechanisms involved instuttering.
1. Recent evidence on the rapid interplay between multiple systems duringspeech processing
2. How lesions in many different locations in the brain can disrupt fluency3. The similarity and differences between stuttering and other speech motor
control disorders
2. The rapid interplay between multiple systems required for fluent speech
Speech is a rapid motor control task; movements must occur within a fewmilliseconds in order for the listener to perceive the correct message. For example,vocal fold opening and closing for voice onset and offset during voiceless andvoiced consonant distinctions are less than 20 ms, a time period which is less thana quarter of the time of any other laryngeal movements (Ludlow & Lou, 1996).To produce plosive consonants, the airflow must be sealed long enough for thepressure to build up and then a rapid ballistic opening gesture is needed to providethe necessary burst of air (Borden & Harris, 1984).
To study brain mechanisms involved in the rapid processing of speech, hightemporal resolution is needed. Previous studies of brain mechanisms involved instuttering have used PET scanning which has spatial resolution but poor temporalresolution; brain activity is studied over a minute or more. These studies haveshown that many different brain regions are associated with speech production;the left frontal operculum, the right and left sensory and motor regions, the rightand left primary and auditory association areas and several subcortical regionsincluding the cerebellum, anterior cingulate, thalamus and the putamen (Braunet al., 1997; Fox et al., 2000). PET scanning, however, cannot provide informationconcerning the rapid inter-relationships between neural events in different brainregions prior to and during speech gestures or stuttered interruptions. Magnetoen-cephalography (MEG), on the other hand, has the necessary temporal resolution.This technique can also have some spatial resolution when the recording pointsare mapped onto brain images obtained using magnetic resonance imaging (MRI).Because of high temporal resolution, within milliseconds, this technique can ex-amine the dynamic relationships between different brain regions during normalspeech and language (Helenius, Salmelin, Service, & Connolly, 1998; Levelt,

C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295 275
Praamstra, Meyer, Helenius, & Salmelin, 1998; Numminen, Salmelin, & Hari,1999).
For example, investigators have examined the dynamic interplay between re-sponses in different brain regions during single word reading (Salmelin, Helenius,& Service, 2000; Salmelin, Schnitzler, Schmitz, & Freund, 2000). When neuralevents were examined within a 400 ms time period between word presentation andmotor execution, predominant processing occurred in the left hemisphere in bothpersons who stutter (PWS) and controls. However, the sequence of activation dif-fered between groups indicating that the dynamic interplay between brain regionsis altered in persons with life-long stuttering. The normal speakers had earlierresponses in the left inferior region processing prior to left central motor regionactivation. The sequence was reversed in the PWS group; these speakers activatedthe left central MI region for motor execution before the left inferior region forarticulation programming.
Reduced activations in auditory association regions in PWS were also foundby Fox et al. (2000)andBraun et al. (1997)using PET scanning. This could beinterpreted as being an abnormal suppression of auditory feedback while speaking.Using MEG, Numminen et al. were able to examine responses to auditory stimuliduring oral reading and found that they were normally suppressed in control sub-jects by 44–71% in comparison with responses during silent reading (Numminenet al., 1999). A subsequent study by Curio, found that cortical responses to one’sown voice transiently activate the right hemisphere around 100 ms after their utter-ance by a speaker (Curio, Neuloh, Numminen, Jousmaki, & Hari, 2000). However,on the left side, this was delayed by 11 ms in normal speakers only during speak-ing and not during listening (Curio et al., 2000). It was suggested that speakingnormally both dampens and delays responses in the auditory cortex in the lefthemisphere. A similar study was conducted in PWS and controls contrasting audi-tory responses during silent reading, reading aloud and chorus reading (Salmelinet al., 1998). Responses to auditory stimuli differed during the different conditionsin the PWS. The authors suggested that responses to auditory stimuli were mostaltered during self-paced reading in PWS, when an abnormal pattern of greaterauditory suppression occurred on the right and reduced suppression on the left.Surprisingly, the normal pattern of greater auditory suppression on the left andreduced suppression on the right, occurred when the subjects were stuttering. Theresults implicate different excitatory and inhibitory inter-relationships betweenbrain regions in normal and PWS groups rather than one particular region beingdysfunctional.
Subtle fine motor deficits can be found in PWS in other motor systems (Borden,1983; Forster & Webster, 1991, 2001; Webster, 1990). In addition, sensory functiondifferences have been found in PWS; the effects of sensory deprivation reducesfluency (Hutchinson & Ringel, 1975) while the ability of PWS to use sensoryfeedback is affected for oral movements but not for hand movements (De Nil &Abbs, 1991; Fucci, Petrosino, Schuster, & Belch, 1991; Petrosino, Fucci, Gorman,& Harris, 1987). Although differences in sensory motor functioning may affect

276 C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295
other motor systems in some PWS, most often the oral-motor system is selectivelyaffected and this is most evident during speech.
Symptom occurrence is intermittent, however — fluent speech occurs the vastmajority of the time in PWS, albeit with considerable effort. Stuttering occursaround 10% of the time, although subjects report they consciously monitor theirspeech to prevent disruptions most of the time. The system seems most easilydisrupted when the demands for rapid and dynamic processing increase (Bosshardt,2002). It may be the functioning of the integrated system that is altered not theindividual components per se.
Some have suggested that left hemisphere dominance for the control of speech/language is altered in PWS. Neuroimaging studies demonstrate that both the leftand right motor and sensory regions are active in normal speakers during speechproduction (Braun et al., 1997). However, WADA testing, where a barbiturate isinfused into the right or left cerebral arterial system, has demonstrated that theleft hemisphere is essential for speech and language expression in most personswho have not sustained a left hemisphere lesion in early childhood (Rasmussen &Milner, 1977). Similarly, the occurrence of aphasia following left hemisphere le-sions is much more frequent than following right hemisphere lesions (Mazzocchi &Vignolo, 1979). For speech production, rather than language processing, however,it is unclear to what degree speech production is lateralized (Rasmussen & Milner,1977). One study used the WADA technique to demonstrate bilateral control ofspeech in four persons who stuttered following brain injury (Jones, 1966). Anotherstudy, however, found that speech was disrupted on the right side only in a patientwho had previously had brain injury and aphasia as well as stuttering. The otherpersons with idiopathic stuttering, only had speech disruption with amytal infusionon the left side, similar to normal (Andrews, Quinn, & Sorby, 1972). Similarly, astudy of three PWS without brain injury found all had left hemisphere dominancefor speech on WADA testing (Luessenhop, Boggs, LaBorwit, & Walle, 1973).There is no evidence to suggest reversed laterality in PWS, rather the data suggestdifferences in the functional inter-relationships between brain regions on the rightand left sides during speech and language processing in PWS.
Phonemic expression has been suggested to depend upon intact neural mecha-nisms in the left insula, based on injury data (Dronkers, 1996) and neuroimagingin normal speakers (Wise, Greene, Buchel, & Scott, 1999). That is, left insulafunctioning is likely involved but must interact with other regions in both the leftand right hemispheres occurs during speech. The evidence thus far suggests thatspeech production may have a different degrees of involvement of right hemi-sphere mechanisms for speech during stuttering (Braun et al., 1997; Fox et al.,1996, 2000). Wood found inadequate left frontal activation during stuttering usingzenon blood flow measures. This normalized after administration of haloperidolto induce fluency (Wood, Stump, McKennhan, Sheldon, & Proctor, 1980). Stud-ies are needed to examine the intricate sequence and timing relationships amongthese neural mechanisms during ongoing speech both in normally fluent speakersand PWS during both fluent and stuttered utterances. As some of the MEG data

C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295 277
suggest, the ability to effect the dynamic interplay between neural mechanismsmay be altered in PWS.
3. Location of brain injury that induces speech dysfluency
A review of the location and types of brain lesions that can induce acquiredstuttering can be informative if one hypothesizes that a particular brain regionmay be essential for fluent speech. To test this hypothesis, the studies reporting onacquired stuttering were reviewed.
Acquired stuttering is qualitatively different from developmental stuttering,possibly because stuttering during development alters emotional and psycholog-ical factors associated with speaking. As a result, anxiety, fear and embarrass-ment may become associated with speech communication in those who have stut-tered since childhood. Acquired stuttering is different from uncontrolled rapidfestinating syllable repetitions that occur during palilalia (LaPointe & Horner,1981). Rather, acquired stuttering involves repetitions, vowel prolongations andoccasional blocks (Ludlow, Rosenberg, Salazar, Grafman, & Smutok, 1987). Thesymptoms alone cannot be easily differentiated from developmental stuttering (VanBorsel & Taillieu, 2001). The similarities in the speech symptoms of the develop-mental and acquired forms, lends credence to the need for analysis of which brainlesions can induce stuttering in adults.
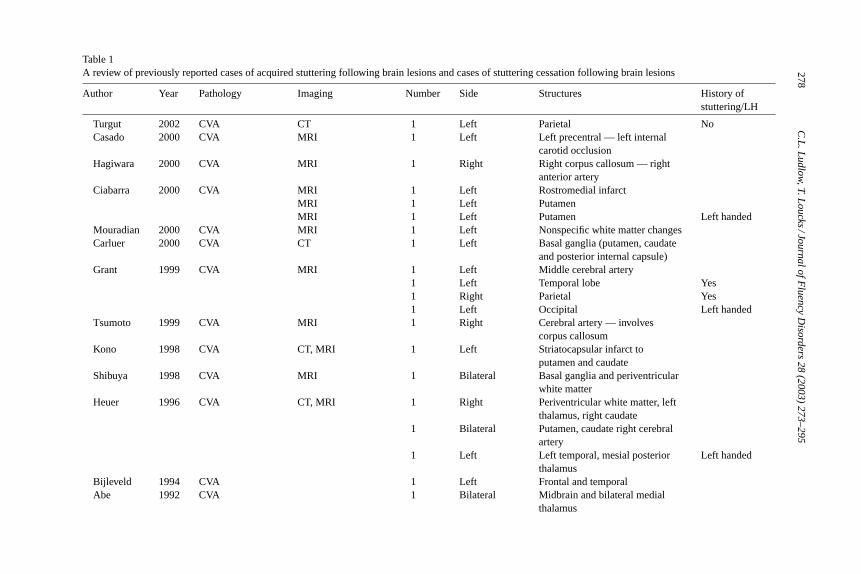
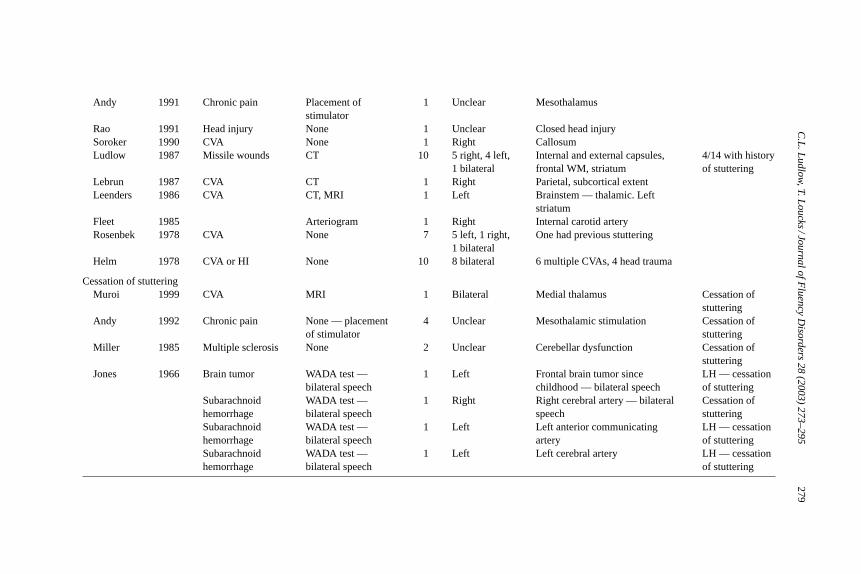
Early studies of acquired stuttering were case reports that lacked imaging dataand were unable to determine the location and extent of the brain lesions. More re-cent reports, however, were able to assess the lesion location (Table 1). These lesiondata do not demonstrate that any one brain region is more likely involved in the ac-quisition of stuttering. Many of the more recent studies, which used neuroimaging,reported involvement of the basal ganglia, with the putamen being listed most often(Abe, Yokoyama, & Yorifji, 1992; Andy & Bhatnagar, 1991; Carluer et al., 2000;Ciabarra, Elkind, Roberts, & Marshall, 2000; Heuer, Sataloff, Mandel, & Travers,1996; Kono, Hirano, Ueda, & Nakajima, 1998; Lebrun, Leleux, & Retif, 1987;Leenders et al., 1986; Ludlow et al., 1987; Shibuya, Wakayama, Murahashi, Aoki,& Ozasa, 1998). A few other studies have reported injuries involving the corpuscallosum (Hagiwara, Takeda, Saito, Shimizu, & Bando, 2000; Soroker, Bar-Israel,Schechter, & Solzi, 1990; Tsumoto, Nishioka, Nakakita, Hayashi, & Maeshima,1999). Only a small number of imaging studies suggest that primarily corticalregions are involved (Bijleveld, Lebrun, & van Dongen, 1994; Franco et al., 2000;Grant, Biousse, Cook, & Newman, 1999; Turgut, Utku, & Balci, 2002) with mostof these indicating diffuse cortical injury (Mouradian, Paslawski, & Shuaib, 2000;Rao, 1991). Other older studies do not provide imaging data (Fleet & Heilman,1985; Helm, Butler, & Benson, 1978; Rosenbek, Messert, Collins, & Wertz, 1978).On the other hand, studies documenting a cessation of stuttering following braininjury involve either diffuse lesions (Miller, 1985) or thalamic injury or stimulation(Andy & Bhatnagar, 1992). One study documented the cessation of stuttering fol-
278C
.L.L
udlow,T.L
oucks/JournalofF
luencyD
isorders28
(2003)273–295
Table 1A review of previously reported cases of acquired stuttering following brain lesions and cases of stuttering cessation following brain lesions
Author Year Pathology Imaging Number Side Structures History ofstuttering/LH
Turgut 2002 CVA CT 1 Left Parietal NoCasado 2000 CVA MRI 1 Left Left precentral — left internal
carotid occlusionHagiwara 2000 CVA MRI 1 Right Right corpus callosum — right
anterior arteryCiabarra 2000 CVA MRI 1 Left Rostromedial infarct
MRI 1 Left PutamenMRI 1 Left Putamen Left handed
Mouradian 2000 CVA MRI 1 Left Nonspecific white matter changesCarluer 2000 CVA CT 1 Left Basal ganglia (putamen, caudate
and posterior internal capsule)Grant 1999 CVA MRI 1 Left Middle cerebral artery
1 Left Temporal lobe Yes1 Right Parietal Yes1 Left Occipital Left handed
Tsumoto 1999 CVA MRI 1 Right Cerebral artery — involvescorpus callosum
Kono 1998 CVA CT, MRI 1 Left Striatocapsular infarct toputamen and caudate
Shibuya 1998 CVA MRI 1 Bilateral Basal ganglia and periventricularwhite matter
Heuer 1996 CVA CT, MRI 1 Right Periventricular white matter, leftthalamus, right caudate
1 Bilateral Putamen, caudate right cerebralartery
1 Left Left temporal, mesial posteriorthalamus
Left handed
Bijleveld 1994 CVA 1 Left Frontal and temporalAbe 1992 CVA 1 Bilateral Midbrain and bilateral medial
thalamus
C.L
.Ludlow
,T.Loucks
/JournalofFluency
Disorders
28(2003)
273–295279
Andy 1991 Chronic pain Placement ofstimulator
1 Unclear Mesothalamus
Rao 1991 Head injury None 1 Unclear Closed head injurySoroker 1990 CVA None 1 Right CallosumLudlow 1987 Missile wounds CT 10 5 right, 4 left,
1 bilateralInternal and external capsules,frontal WM, striatum
4/14 with historyof stuttering
Lebrun 1987 CVA CT 1 Right Parietal, subcortical extentLeenders 1986 CVA CT, MRI 1 Left Brainstem — thalamic. Left
striatumFleet 1985 Arteriogram 1 Right Internal carotid arteryRosenbek 1978 CVA None 7 5 left, 1 right,
1 bilateralOne had previous stuttering
Helm 1978 CVA or HI None 10 8 bilateral 6 multiple CVAs, 4 head trauma
Cessation of stutteringMuroi 1999 CVA MRI 1 Bilateral Medial thalamus Cessation of
stutteringAndy 1992 Chronic pain None — placement
of stimulator4 Unclear Mesothalamic stimulation Cessation of
stutteringMiller 1985 Multiple sclerosis None 2 Unclear Cerebellar dysfunction Cessation of
stutteringJones 1966 Brain tumor WADA test —
bilateral speech1 Left Frontal brain tumor since
childhood — bilateral speechLH — cessationof stuttering
Subarachnoidhemorrhage
WADA test —bilateral speech
1 Right Right cerebral artery — bilateralspeech
Cessation ofstuttering
Subarachnoidhemorrhage
WADA test —bilateral speech
1 Left Left anterior communicatingartery
LH — cessationof stuttering
Subarachnoidhemorrhage
WADA test —bilateral speech
1 Left Left cerebral artery LH — cessationof stuttering

280 C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295
lowing unilateral brain injury in four persons who had previously stuttering sincechildhood and had bilateral speech representation based on the WADA test (Jones,1966).
The articles listed inTable 1demonstrated some left hemispheric preferencefor inducing stuttering: 47% were left sided lesions, 26% were right and 26% werebilateral. Of those with imaging findings reported, 75% involved subcortical re-gions (basal ganglia, thalamus, internal capsule and corpus callosum). Similar todevelopmental stuttering, there was a 5:3 male to female ratio when sex was ex-amined across these studies. Further, a normal handedness distribution was found;80% were right handed and 19% were left handed. Few authors reported whetherpatients had previously stuttered during childhood; however of 57 cases, 14% re-ported childhood stuttering. This is a higher frequency than the 1% incidence ofstuttering in adults. Apparently, those who recovered from childhood stutteringhave a greater vulnerability for acquired stuttering after brain lesions.
The only conclusion that can be made from these studies are that the lesionsassociated with acquired stuttering rarely involve the primary speech and lan-guage regions in the left hemisphere (Broca’s area, the temporal planum, insulaor Wernicke’s area). When these regions are involved, patients usually exhibitaphasia, which might mask stuttering symptoms, however, such patients do notexhibit stuttering when they recover (Ludlow et al., 1986). Therefore, the lesionsleading to acquired stuttering seem to be those that impinge upon structures thatsupport and modulate rapid communication between brain regions such as thecorpus callosum, or areas that exert influence on multiple brain regions such asthe basal ganglia and thalamus. The lesions in either the corpus callosum or thala-mus could interfere with rapid communication of distributed areas during speechproduction. On the other hand, they were not disconnection lesions within thespeech and language system in the left hemisphere which result in different typesof aphasic syndromes (Geschwind, 1965a, 1965b). Rather, these lesions disturbrapid coordinated functioning of the brain networks. Acquired stuttering did notoccur when brain regions involved in speech and language functions were irrevo-cably damaged; rather it occurred when the speech and language areas were notable to inter-relate with other brain regions in a rapid dynamic fashion.
Stuttering may reflect a poorly coordinated speech system possibly becauseof deficient development of white matter tracts (Sommer, Koch, Paulus, Weiller,& Buchel, 2002) or reduced neural pruning (Foundas, Bollich, Corey, Hurley, &Heilman, 2001). The former was further supported by the recent results of Som-mer et al., who found higher thresholds for hand muscle responses to transcranialmagnetic stimulation over the motor cortex in stuttering adults (Sommer, Wischer,Tergau, & Paulus, 2003). Stuttering may reflect an instability or loss of controlin brain function rather than a loss of function — which makes stuttering simi-lar to some other motor control disorders such as tremor, dystonia or Gilles de laTourette’s syndrome (TS). To expand this view, a review of the some of the similar-ities and differences between stuttering and other speech motor control disordersis helpful.

C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295 281
4. The similarity and differences between stuttering and other speechmotor control disorders
On first approximation, the phenomenology of stuttering seems commensuratewith a neurologically based developmental motor control disorder (Brin, Blitzer, &Stewart, 1998; Kiziltan & Akalin, 1996). If the sensorimotor differences identifiedin stuttering have more similarities than differences relative to other speech motorcontrol disorders, then this initial approximation is supported.
Speech production takes place during human communication, except in speechscience laboratories where subjects are tested while producing meaningless phrasesmodeled by the tester. Such well controlled speech research tasks rarely elicitstuttering — stuttering is more frequent when subjects are required to formulatemeaningful communication (Ratner, Gawronski, & Rice, 1964; Ratner & Sih,1987). This augmentation of symptoms during communication is more pronouncedin stuttering, but is a common feature of most speech and voice motor controldisorders (Caligiuri, 1989; Sarno, 1968). In dysarthria and voice disorders, patientsare often able to produce the target behavior in the clinical setting when attentionis focused on production. After they leave the clinic, however, they tend to revert toimpaired performance during conversation with others — a phenomenon referredto as carry-over (Klaff, 1976). In TS, patients can control symptoms for shortperiods, only to have them emerge in a more frequent pattern when they are nolonger monitoring or controlling them.
Stuttering is a task specific disorder — that is, oral-motor dysfunction becomesapparent only during speech and not during humming, singing or chewing. In spas-modic dysphonia (SD), which is a laryngeal dystonia, the voice disruptions are mostmanifest during speech and much less evident during simple vowel prolongation(Sapienza, Murry, & Brown, 1998) and singing (Bloch, Hirano, & Gould, 1985).In TS, symptoms occur at rest but are increased by 30% during speech (Ludlow,1993). The separation mentioned earlier, between non-meaningful speech produc-tion which is usually symptom free, and linguistic communication with enhancedstuttering (Perkins, Kent, & Curlee, 1991) is also a task dependent attribute. Taskrelated differences also occur in oral-mandibular dystonia where some patients mayonly have oral-motor muscle abnormalities during speech but not during chewing,while others only have chewing affected with no oral-motor abnormalities duringspeech (Rosenbaum & Jankovic, 1988).
The difference in the occurrence of symptoms during speech communicationand emotional expression is also task related. Laughter and crying are usually nor-mal in both PWS and persons with SD (Bloch et al., 1985). In SD, this differencepreviously led to the assumption that the disorder was psychogenic. Now this dif-ference is understood as evidence of different vocalization systems being presentin human and non-human primates. Singing is thought to involve different brainregions from speech, particularly in the right hemisphere (Riecker, Ackermann,Wildgruber, Dogil, & Grodd, 2000) and simple vocalization and emotional ex-pression may involve the cingulate and periaquaductal grey (Jurgens, 2002). On

282 C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295
the other hand, speech communication involves both cortical and subcorticalsystems.
Stuttering is more frequent when central processing demands increase withgreater speech complexity or dual processing tasks (Bosshardt, 2002; Hall,Yamashita, & Aram, 1993; Starkweather, 2002). Similarly, in SD, motor functionand electromyographic abnormalities are more impaired on complex processingtasks (Schaefer et al., 1992).
Both disorders are focal, that is they involve oral-motor or laryngeal musculaturebut not other motor systems. Reich, for example, conducted similar studies in bothdisorders, comparing laryngeal and manual reaction times with similar results, thelaryngeal or phonatory systems were affected while the manual systems were not(Reich & Till, 1983; Reich, Till, & Goldsmith, 1981).
One difference between these disorders, of course, is the age of onset. Given thisdifference it is surprising that both groups have similar reactions to their speechdisorder. Persons with SD, which onsets in adulthood, develop an adverse reactionto telephone use, become socially isolated and often develop a reactive depres-sion (Cannito, 1991) which resolves with treatment (Murry, Cannito, & Woodson,1994). Developmental stuttering often involves the development of similar fearand avoidance reactions to the telephone and feelings of social isolation (Miller &Watson, 1992). Persons with either disorders may develop a social anxiety disordersecondary to their voice and speech impairments (Moutier & Stein, 1999).
The task specific attributes of stuttering and SD implicate two possible selectiveaspects of these disorders. First, they only appear during speech sound productionwhich has more rapid and precise motor control demands on vocal tract motorcontrol (Ludlow & Lou, 1996). Second they implicate that one system is affected,that required for speech production during communication and language, while an-other is not. The vocal spasms of SD and the repetitions, prolongations and pausesin stuttering point to the intermittent nature of both disorders during speech pro-duction. The intermittent nature of these disorders support a serious considerationof whether the central control abnormalities may be similar.
Another disorder with intermittent vocal symptom occurrence is Tourette’ssyndrome, a developmental disorder with onset most often between 5 and 12years of age. This disorder involves multiple and one or more vocal tics occur-ring either many times a day or intermittently in a year, the tic characteristicscan change over time, have onset before 21 years and are idiopathic, that is, notexclusively due to psychoactive substance intoxication or due to known centralnervous system disease (Diagnostic and statistical manual of mental disorders,1987). Speech and vocal tics occur at rest in TS, are exacerbated during speech(Ludlow, 1993; Ludlow, Polinsky, Caine, Bassich, & Ebert, 1982) but occurprimarily at the beginning of speech or during speech pauses (Frank, 1978;Martindale, 1976, 1977). No differences in tic production occur with delayed audi-tory feedback or white noise or reduced speaking rate, which differs from stutteringsymptom characteristics (Ludlow, 1993). Different authors have noted, however,that a high proportion of persons affected with TS report stuttered as a child (45%

C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295 283
reported developmental stuttering in one report,Ludlow, 1993), and an associa-tion between stuttering and TS in families was reported by one group (Comings& Comings, 1994; Comings, Comings, & Knell, 1989; Comings et al., 1996), al-though not supported by others (Pauls, Leckman, & Cohen, 1993). Some intriguingsimilarities, however, are that both disorders are benefited by dopamine receptorblockade (Kossoff & Singer, 2001; Ludlow & Braun, 1993), may be geneticallyrelated, wax and wane in severity, start in childhood, and spontaneously remit laterin a large proportion of cases. Recent functional neuroimaging has demonstratedthat TS involves aberrant activity in the precentral gyrus, insula, anterior cingulateand basal ganglia coupled with hypoactivity in the sensorimotor cortices (Sternet al., 2000). Increased activity in some of the same regions have been associatedwith the production of stuttering (Braun et al., 1997; Fox et al., 2000). SeveralTS patients report that tics are preceded by premonitions or urges to tics. Oth-ers emphasize that sensations often precede tics and that the tic is a responseto a sensory experience. In stuttering, similar premonitions can occur prior todysfluencies.
TS, however, is not a task specific disorder or focal to the speech musculatureas is the case for both SD and stuttering, although vocal and speech tics are anecessary part of the syndrome. Therefore, there are more similarities betweenstuttering and SD than between stuttering and TS in motor symptoms.
4.1. Sensory deviations
A distinctive attribute of the focal dystonias are sensory anomalies that involvesensory tricks, deviant reflexes, sensory perception differences and abnormal sen-sory evoked potentials (Hallett, 1995). Sensory tricks are used by individuals withfocal dystonias to control symptoms, such as, touching the face in Meige syndromeor chewing tooth-picks in oromandibular dystonia (Rosenbaum & Jankovic, 1988).In SD, a similar reduction in symptoms can occur during fiberoptic nasolaryn-goscopy, presumably due to altered sensation during placement of the nasolaryn-goscope in the oropharynx. Compensatory speech patterns, such as whispering,that are often observed clinically (Bloch et al., 1985) also alter sensory feedback.
We have studied laryngeal closure reflexes in the thyroarytenoid muscle in re-sponse to electrical stimulation of the laryngeal afferents contained in the superiorlaryngeal nerve in normal speakers and persons with SD. When paired electricalstimuli are presented at less than 1 s intervals for conditioning the late R2 responses(>60 ms latency) are normally suppressed reflecting central inhibitory processesmodulating these responses. Conditioning abnormalities of the R2 response of thelaryngeal adductor reflex have been found in both the adductor and abductor typesof SD suggesting an abnormality in central suppression of muscle responses tosensory feedback (Deleyiannis, Gillespie, Bielamowicz, Yamashita, & Ludlow,1999; Ludlow, Schulz, Yamashita, & Deleyiannis, 1995). Decreased or altered af-ferent feedback reduce spasmodic bursts in some of the dystonias (Kaji, Rothwell,et al., 1995; Yoshida et al., 1998).

284 C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295
As mentioned earlier, a number of studies point to corresponding sensory func-tion differences in stuttering. Psychophysical studies indicate PWS have higherperceptual somatosensory thresholds for minimal movements of the jaw, lowerlip and tongue and lingual vibrotactile stimuli. Persons who stutter also show de-viant kinesthetic perceptions of jaw position on movement accuracy tasks (De Nil& Abbs, 1991). Certain differences relative to SD are noted though. A bilateralblockade of oral sensation was associated with increases in the frequency of stutter-ing symptoms (Hutchinson & Ringel, 1975), unlike the effects of lidocaine in someof the dystonias (Kaji, Kohara, et al., 1995). Although an influential hypothesissuggested exaggerated reflex gains could lead to stuttering (Zimmermann, Smith,& Hanley, 1981), evidence for marked oral reflex deviations have not been found,although labial reflex gains may be slightly altered in PWS (McClean, 1987).
The observations of so-called secondary or compensatory behaviors in stutter-ing show a broad similarity to the sensory tricks and compensatory speech adjust-ments used by persons with focal dystonias (Kiziltan & Akalin, 1996; Mulligan,Anderson, Jones, Williams, & Donaldson, 2001). Reductions in stuttering symp-toms are associated with altered speech patterns (whisper, choral speech) and un-usual oral gestures or bodily movements. Such movement changes potentially alterauditory and somatosensory inputs in turn helping to avoid or suppress stuttering— and could be interpreted as sensory tricks.
The most profound sensory effects, however, are those of auditory feedbackmanipulations (delayed auditory feedback, masking, etc.) that significantlysuppress stuttering symptoms. The role of auditory feedback has also had a partic-ular role in speech fluency: the paradoxical effects of chorus reading and delayedor altered feedback is pronounced in this disorder (Adams & Ramig, 1980; Stager,Denman, & Ludlow, 1997), although also present in dysarthria in Parkinsonism(Hanson & Metter, 1983). However, extended exposure to altered auditory feed-back leads to less dramatic effects (Armson & Stuart, 1998) and it may not be theauditory modality per se but rather any significant change in the speaking con-text (Kalinowski, Stuart, Rastatter, Snyder, & Dayalu, 2000) that can alter fluency.One small study found no effect of white noise of delayed auditory feedback onsymptom occurrence in TS (Ludlow, Nauton, & Bassich, 1984).
Even if sensory abnormalities in either the larynx or orofacial structures areidentified in either disorder, the role of sensory abnormalities in these disordersstill requires elucidation. It has been long recognized that suppression of sensoryinformation that is possibly redundant or that could interfere with ongoing move-ment is possibly just as critical for the coordination of movement as the receipt ofsensory information. Inappropriate or a lack of inhibition in SD or stuttering couldindeed be a central factor in the emergence of vocal spasm or stuttering events, as issuggested for SD by the reduction in R2 conditioning. Studies of abnormal sensoryinhibition of brain responses to afferent feedback in stuttering are an avenue thatcan be addressed with rapid neurophysiological recording using MEG as has beendone for auditory feedback (Salmelin, Helenius, et al., 2000; Salmelin, Schnitzler,et al., 2000).

C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295 285
4.2. Muscle interference
Earlier ideas about the physiological basis for dysfluencies in both disorders in-volved speculation that tonically increased activity in the laryngeal muscles causedspeech disruptions in SD and stuttering. In the adductor type of SD,Nash andLudlow (1996)identified intermittent increases in thyroarytenoid activity duringvoice breaks, but that the overall level of muscle activity was not abnormal (VanPelt, udlow, & Smith, 1994). Thus, spasmodic bursts in the thyroarytenoid mus-cles intruded upon otherwise unremarkable muscle recruitment (Nash & Ludlow,1996). Muscle bursts in the abductor type of SD are more variable across subjects(Cyrus, Bielamowicz, Evans, & Ludlow, 2001), but again tonically elevated mus-cle tone has not been observed (Van Pelt et al., 1994). The mechanism involved inmuscle bursts during voice breaks may arise from deficient suppression of laryn-geal sensorimotor reflexes in both SD types. In SD, abnormal central modulationof brain stem mechanisms may result in reduced suppression of laryngeal reflexesin response to the sensory feedback due to speech and voice production, in anotherwise normal neural circuit for phonation.
In stuttering, evidence for elevated tonic activity in the laryngeal or orofacialmuscles has not been found during speech disruptions (Smith et al., 1993; Smith,Denny, Shaffer, Kelly, & Hirano, 1996). Tonic co-contraction of the laryngeal mus-cles during stuttering was first reported by Freeman (Freeman & Ushijima, 1978),but the study was flawed, because of the lack of a control group. Subsequent con-trolled studies of laryngeal muscle and orofacial muscles have not found tonicover-activity or abnormal co-contraction during fluent speech or dysfluencies inPWS (Smith et al., 1996). Instead, approximately 50% of adults and teens whostutter display high frequency oscillations (6–15 Hz range) in the orofacial andlaryngeal muscles during stuttering dysfluencies but not during fluent speech. Theoscillations occurred simultaneously in multiple vocal tract muscles and have beenlikened to tremorogenic activity (Smith et al., 1993). The 50% incidence of theoscillations prevents them from being considered characteristic of stuttering anddistances stuttering from SD, which shows a nearly universal incidence for burst-ing abnormalities in a limited set of muscles during speech breaks. Finer resolutionanalyses of muscle activity during stuttering may identify other aberrations in mus-cle activity, but it remains likely that stuttering dysfluencies may involve normalmuscle electromyographic amplitudes and spectral characteristics. Although mus-cle activity during a dysfluency is not characterized by spasms, tremor or excessiveco-contraction, questions remain whether the coordination of muscle activation isaberrant or if there is an absence of normal muscle inhibition during stuttering.
Increased cortical excitability in motor regions has been implicated in bothSD and stuttering by functional neuroimaging during symptom production (Braunet al., 1997; Fox et al., 1996, 2000; Hirano et al., 2001). Whether cortical ex-citability is due to decreased cortical inhibition can be examined using transcra-nial magnetic stimulation using a conditioning paradigm. First, motor thresholdsare determined for eliciting a motor evoked potential (MEP) in a hand muscle in

286 C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295
response to cortical stimulation in the primary motor area. Next, cortical inhibitioncan be assessed using a conditioning paradigm where a sub-threshold conditioningstimulus (at 90% of motor threshold amplitude) precedes a supra-threshold teststimulus at time intervals between 1 and 30 ms. The degree to which the MEPresponse to the test stimulus is suppressed relative to an MEP to an unconditionedstimulus of the same strength is measured (Kujirai et al., 1993). Increased exci-tation of hand muscle responses using this paradigm was found in various focaldystonias such as writer cramp and blepharospasm (Sommer, Ruge, et al., 2002).In a recent study, using the same conditioning paradigm to study hand muscleresponse inhibition, no abnormalities in inhibition were found in PWS (Sommeret al., 2003). The PWS group, however, had significantly higher motor thresh-olds for eliciting hand muscle responses than the controls, which was unexpected.These findings may suggest a different pathophysiology in stuttering, that is, areduced cortico-spinal tract excitability to hand muscles in PWS. Further studyis needed to examine whether similar findings occur in SD. Also, conditioningstudies of cortical excitability for oral and laryngeal musculature is needed in bothSD and TS.
Because stuttering dysfluencies are not specific to a small set of muscles, thismay explain why botulinum toxin injections have not and will not likely be appro-priate treatment for stuttering (Stager & Ludlow, 1994) even though it is clearlythe most effective treatment to date for SD (Truong, Rontal, Rolnick, Aronson, &Mistura, 1991). Botulinum toxin injection, however, suppresses muscle spasms bydenervating both the muscle fibers and muscle spindles. Current behavioral treat-ments for stuttering may also predominantly involve muscle suppression. Treat-ments that instead target the central mechanisms may be most effective in bothdisorders, because any abnormal muscle activity in either disorder is secondary tothe central pathological mechanisms.
4.3. Central abnormalities
Along with arguments for deviancies in sensori-motor processing and muscleactivation in PWS and SD, abnormalities in cortical and/or subcortical processingare thought to be present in both disorders. Only a few studies of brain functionhave been conducted in either disorder.
A case study of a person with SD using positron emission tomography (H215O
PET) indicated reduced regional cerebral blood flow (RCBF) responses in thesupplementary motor area (SMA) and increased RCBF in the left superior tem-poral gyrus during phonation relative to six control subjects (Hirano et al., 2001).Findings for a single subject are difficult to interpret however, due to the high vari-ability of RCBF both between participants and across tasks. Studies in other formsof dystonia have shown deficient central activation in response to sensory feed-back (Tempel & Perlmutter, 1990, 1993). Similarly, patterns of hyperactivation andhypoactivation have been found in PWS. Hypoactivation of the left post-centralsensory and auditory association regions were found in PWS during dysfluency

C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295 287
evoking tasks byWu et al. (1995), Fox et al. (1996, 2000), andBraun et al. (1997).At the same time increased activation in premotor and motor areas have been foundin both stuttering and dystonia (Braun et al., 1997; Odergren, Stone-Elander, &Ingvar, 1998). Firm conclusions regarding similar patterns of brain dysfunctionin the two disorders cannot be made given the limited data, particularly for SD.However, the corresponding patterns of sensory hypoactivation and frontal motorand premotor hyperactivation make a potential link between the disorders morecompelling.
4.4. Basal ganglia abnormalities
Basal ganglia dysfunction has been viewed as the most probable dysfunctionalregion in primary dystonias, in which the failure of the basal ganglia circuitry to in-hibit inappropriate cortical motor control regions may lead to pathological muscleactivation and dystonic posturing (Galardi et al., 1996). Given that basal gangliaoutput modulates cortical motor areas, the logic behind hypotheses of basal gangliadysfunction in dystonia is attractive. The neuroimaging studies of stuttering alsopoint to atypical basal ganglia activation or abnormal activity in areas receivingbasal ganglia output.Wu et al. (1995)reported hypoactivation in the left caudatenucleus during both fluency and dysfluency evoking tasks. In the Braun et al. study(Braun et al., 1997), PWS showed higher activity in the right caudate nucleus andgenerally lower activation in SMA compared to controls in both the dysfluentand fluent conditions. Consequently, the still limited evidence from neuroimag-ing indicates atypical activity in the basal ganglia or in areas receiving efferentoutput from basal ganglia nuclei. The brain lesion data on acquired stuttering re-viewed earlier, suggest that basal ganglia dysfunction can induce stuttering in fluentadults.
To date, the focal dystonias have not been found to be responsive to neurophar-macological intervention (Brin et al., 1998) and stuttering is only benefited bydopamine receptor blockade although the side effects usually outweigh the ben-efits (Ludlow & Braun, 1993). Paroxetine has been reported to have serious sideeffects (Bloch, Stager, Braun, & Rubinow, 1995) and risperidone has fewer side ef-fects but limited long term benefits (Lee et al., 2001; Maguire et al., 1999; Maguire,Riley, Franklin, & Gottschalk, 2000).
4.5. Vulnerable systems
Stuttering and SD are both idiopathic disorders; that is they are not normallysecondary to any know disease or disorder but emerge spontaneously, one duringdevelopment and the other during adulthood. Both disorders can emerge duringstressful events, not because stress is a cause of the disorder but rather because thespeech motor system in such persons may be more vulnerable to breakdown whenstress is increased. No studies have addressed the rate of co-occurrence of thesedisorders in families.

288 C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295
Our consideration of the similarities and differences between stuttering andother speech motor control disorders such as SD, is aimed at stimulating newavenues for hypothesis-driven research into the mechanisms of stuttering. Sensoryprocessing irregularities in stuttering that are similar to focal dystonias could beinvestigated using whole head magnetoencephalography to further assess whetherthe time courses, amplitudes and source locations of oral somatosensory and/orauditory evoked potentials are deviant in stuttering. Application of functional MRI(fMRI) to neurocognitive and neuromotor processes in stuttering (and SD) havebarely been initiated, but the success of fMRI in understanding brain activationfor such processes indicates it holds promise for understanding central processesin stuttering. The possibility that cortical pryramidal tract excitation is affected instuttering (Sommer et al., 2003) needs to be investigated further. Certainly, PETtechniques could be used to measure neurotransmitter uptake and to assess basalganglia function, techniques which have already added improved understandingof the pathogenesis in Parkinson’s disease (Ito et al., 2002). Integrating theseresearch questions with good data on the simultaneous activity in different brainmechanisms should help to parse out the dynamic inter-relationships and how thesediffer in stuttering both between tasks and from normal.
The major difference between stuttering and focal dystonia is the age of on-set. The period of vulnerability for developing stuttering is between 2 years ofage and puberty. The neurobiology of development may underlie the bases forthe emergence of stuttering in childhood. Developmental neurobiology involvestwo overall processes, the initial growth stages of cellular proliferation, migra-tion, differentiation and synaptogenesis which occur in the prenatal and earlypost-natal periods (Purves, 1994) and the later stages of selective refinement.Early childhood, the period of vulnerability for the development of stuttering,may include processes involved in refinement of the neurological system. Re-duction and refinement occur through dendritic pruning, programmed cell deathand selective enhancement of networks of synaptic connections (Purves, 1994).Reduction and refinement in neural processes may be essential for the develop-ment of normal speech motor control based in part, on activity and use. That is,as some synaptic connections are enhanced through use, others are lost throughinactivity, shaping the emergence of efficient circuitry for speech function withinthe central nervous system. Although little information is available on the refine-ment process within the speech motor control system during childhood, it canbe assumed that it follows the same neurodevelopmental processes as the rest ofthe central nervous system. The recent finding of increased size in the right andleft planum temporali in stuttering adults and additional gyri in perisylvian re-gion compared to normal (Foundas et al., 2001) may suggest abnormal pruning.On the other hand, reduced white matter tracts in central areas (Sommer, Koch,et al., 2002) might suggest abnormal growth and refinement processes duringdevelopment.
In summary, the evidence to date suggests that stuttering is a neurodevelop-mental motor control disorder. It has some intriguing similarities to other motor

C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295 289
control disorders, the focal dystonias, in particular. This suggests that conceptualframeworks based on the dynamic interplay between different components of themotor control system and its interface with language processing can be helpfulfor the study of the pathophysiology of both disorders. Although the etiologiesare most likely quite different, the functional abnormalities appear to be relatedand investigations into the rapid dynamic inhibitory and excitatory processes forspeech production may be beneficial to understanding both disorders.
References
Abe, K., Yokoyama, R., & Yorifji, S. (1992). On stuttering-like hesitation resulting from infarct in themidbrain and the mesial thalami.Rinsho Shinkeigaku, 32, 822–825.
Adams, M. R., & Ramig, P. (1980). Vocal characteristics of normal speakers and stutterers duringchoral reading.Journal of Speech and Hearing Research, 23(2), 457–469.
Andrews, G., Quinn, P. T., & Sorby, W. A. (1972). Stuttering: An investigation into cerebral dominancefor speech.Journal of Neurology, Neurosurgery, and Psychiatry, 35(3), 414–418.
Andy, O. J., & Bhatnagar, S. C. (1991). Thalamic-induced stuttering (surgical observations).Journalof Speech and Hearing Research, 34, 796–800.
Andy, O. J., & Bhatnagar, S. C. (1992). Stuttering acquired from subcortical pathologies and itsalleviation from thalamic perturbation.Brain and Language, 42(4), 385–401.
Armson, J., & Stuart, A. (1998). Effect of extended exposure to frequency-altered feedback on stutteringduring reading and monologue.Journal of Speech, Language, and Hearing Research, 41(3), 479–490.
Bijleveld, H., Lebrun, Y., & van Dongen, H. (1994). A case of acquired stuttering.Folia Phoniatricaet Logopaedica, 46(5), 250–253.
Bloch, C. S., Hirano, M., & Gould, W. J. (1985). Symptom improvement of spastic dysphonia inresponse to phonatory tasks.The Annals of Otology, Rhinology and Laryngology, 94, 51–54.
Bloch, M., Stager, S. V., Braun, A. R., & Rubinow, D. R. (1995). Severe psychiatric symptoms associatedwith paroxetine withdrawal [letter].Lancet, 346, 57.
Borden, G. J. (1983). Initiation versus execution time during manual and oral counting by stutterers.Journal of Speech and Hearing Research, 26(3), 389–396.
Borden, G. J., & Harris, K. S. (1984).Speech science primer: Physiology, acoustics, and perception ofspeech (2nd ed.). Baltimore, MD: Williams and Wilkins.
Bosshardt, H. G. (2002). Effects of concurrent cognitive processing on the fluency of word repetition:Comparison between persons who do and do not stutter.Journal of Fluency Disorders, 27(2),93–113 (quiz 113–114).
Braun, A. R., Varga, M., Stager, S., Schulz, G., Selbie, S., Maisog, J. M., Carson, R. E., & Ludlow,C. L. (1997). Altered patterns of cerebral activity during speech and language production indevelopmental stuttering. An H215O positron emission tomography study.Brain, 120, 761–784.
Brin, M. F., Blitzer, A., & Stewart, C. (1998). Laryngeal dystonia (spasmodic dysphonia): Observationsof 901 patients and treatment with botulinum toxin.Advances in Neurology, 78, 237–252.
Caligiuri, M. P. (1989). The influence of speaking rate on articulatory hypokinesia in Parkinsoniandysarthria.Brain and Language, 36, 493–502.
Cannito, M. P. (1991). Emotional considerations in spasmodic dysphonia: Psychometric quantification.Journal of Communication Disorders, 24, 313–329.
Carluer, L., Marie, R. M., Lambert, J., Defer, G. L., Coskun, O., & Rossa, Y. (2000). Acquired andpersistent stuttering as the main symptom of striatal infarction.Movement Disorders, 15(2), 343–346.

290 C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295
Ciabarra, A. M., Elkind, M. S., Roberts, J. K., & Marshall, R. S. (2000). Subcortical infarction resultingin acquired stuttering.Journal of Neurology, Neurosurgery, and Psychiatry, 69(4), 546–549.
Comings, D. E., & Comings, B. G. (1994). TS, learning, and speech problems [letter comment].Journalof American Academy of Child Adolescent Psychiatry, 33(3), 429–430.
Comings, D. E., Comings, B. G., & Knell, E. (1989). Hypothesis: Homozygosity in Tourette syndrome.American Journal of Medical Genetics, 34, 413–421.
Comings, D. E., Wu, S., Chiu, C., Ring, R. H., Gade, R., Ahn, C., MacMurray, J. P., Dietz, G., &Muhleman, D. (1996). Polygenic inheritance of Tourette syndrome, stuttering, attention deficithyperactivity, conduct, and oppositional defiant disorder: The additive and subtractive effect of thethree dopaminergic genes DRD2, D beta H, and DAT1.American Journal of Medical Genetics,67(3), 264–288.
Curio, G., Neuloh, G., Numminen, J., Jousmaki, V., & Hari, R. (2000). Speaking modifies voice-evokedactivity in the human auditory cortex.Human Brain Mapping, 9(4), 183–191.
Cyrus, C. B., Bielamowicz, S., Evans, F. J., & Ludlow, C. L. (2001). Adductor muscle activityabnormalities in abductor spasmodic dysphonia.Otolaryngology - Head and Neck Surgery, 124(1),23–30.
De Nil, L. F., & Abbs, J. H. (1991). Kinaesthetic acuity of stutterers and non-stutterers for oral andnon-oral movements.Brain, 114(Pt. 5), 2145–2158.
Deleyiannis, F. W., Gillespie, M., Bielamowicz, S., Yamashita, T., & Ludlow, C. L. (1999). Laryngeallong latency response conditioning in abductor spasmodic dysphonia.The Annals of Otology,Rhinology and Laryngology, 108(6), 612–619.
Diagnostic and statistical manual of mental disorders. (1987).Diagnostic and statistical manual ofmental disorders, r. (3rd ed.). Washington, DC: American Psychiatric Association.
Dronkers, N. F. (1996). A new brain region for coordinating speech articulation.Nature, 384, 159–161.Fleet, W. S., & Heilman, K. M. (1985). Acquired stuttering from a right hemisphere lesion in a
right-hander.Neurology, 35(9), 1343–1346.Forster, D. C., & Webster, W. G. (1991). Concurrent task interference in stutterers: dissociating
hemispheric specialization and activation.Canadian Journal of Psychology, 45, 321–335.Forster, D. C., & Webster, W. G. (2001). Speech-motor control and interhemispheric relations in
recovered and persistent stuttering.Developmental Neuropsycholgy, 19(2), 125–145.Foundas, A. L., Bollich, A. M., Corey, D. M., Hurley, M., & Heilman, K. M. (2001). Anomalous
anatomy of speech-language areas in adults with persistent developmental stuttering.Neurology,57(2), 207–215.
Fox, P. T., Ingham, R. J., Ingham, J. C., Hirsch, T. B., Downs, J. H., Martin, C., Jerabek, P., Glass, T.,& Lancaster, J. L. (1996). A PET study of the neural systems of stuttering.Nature, 382, 158–162.
Fox, P. T., Ingham, R. J., Ingham, J. C., Zamarripa, F., Xiong, J. H., & Lancaster, J. L. (2000). Braincorrelates of stuttering and syllable production. A PET performance-correlation analysis.Brain,123(Pt. 10), 1985–2004.
Franco, E., Casado, J. L., Lopez Dominguez, J. M., Diaz Espejo, C., Blanco, A., & Robledo, A. (2000).Stuttering as the only manifestation of a cerebral infarct.Neurologia, 15(9), 414–416.
Frank, S. M. (1978). Psycholinguistic findings in Gilles de la Tourette syndrome.Journal ofCommunication Disorders, 11, 349–363.
Freeman, F. J., & Ushijima, T. (1978). Laryngeal muscle activity during stuttering.Journal of Speechand Hearing Research, 21, 538–562.
Fucci, D., Petrosino, L., Schuster, S., & Belch, M. (1991). Lingual vibrotactile thresholdshift differences between stutterers and normal speakers during magnitude-estimation scaling.Perceptual and Motor Skills, 73(1), 55–62.
Galardi, G., Perani, D., Grassi, F., Bressi, S., Amadio, S., Antoni, M., Comi, G. C., Canal, N., &Fazio, F. (1996). Basal ganglia and thalamo-cortical hypermetabolism in patients with spasmodictorticollis. Acta Neurologica Scandinavica, 94(3), 172–176.
Geschwind, N. (1965a). Disconnexion syndromes in animals and man. I.Brain, 88, 237–294.Geschwind, N. (1965b). Disconnexion syndromes in animals and man. II.Brain, 88, 585–644.

C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295 291
Grant, A. C., Biousse, V., Cook, A. A., & Newman, N. J. (1999). Stroke-associated stuttering.Archivesof Neurology, 56(5), 624–627.
Hagiwara, H., Takeda, K., Saito, F., Shimizu, T., & Bando, M. (2000). A case of callosal apraxiawithout agraphia and acquired stuttering associated with callosal infarction.Rinsho Shinkeigaku,40(6), 605–610.
Hall, N. E., Yamashita, T. S., & Aram, D. M. (1993). Relationship between language and fluency inchildren with developmental language disorders.Journal of Speech and Hearing Research, 36(3),568–579.
Hallett, M. (1995). Is dystonia a sensory disorder?Annals of Neurology, 38(2), 139–140.Hanson, W. R., & Metter, E. J. (1983). Speech rate modification in Parkinson’s disease: A report of
two cases. In W. R. Berry (Ed.),Clinical dysarthria (pp. 231–251). San Diego, CA: College-HillPress.
Helenius, P., Salmelin, R., Service, E., & Connolly, J. F. (1998). Distinct time courses of word andcontext comprehension in the left temporal cortex.Brain, 121(Pt. 6), 1133–1142.
Helm, N. A., Butler, R. B., & Benson, D. F. (1978). Acquired stuttering.Neurology, 28(11), 1159–1165.Heuer, R. J., Sataloff, R. T., Mandel, S., & Travers, N. (1996). Neurogenic stuttering: Further
corroboration of site of lesion.Ear, Nose, and Throat Journal, 75(3), 161–168.Hirano, S., Kojima, H., Naito, Y., Tateya, I., Shoji, K., Kaneko, K., Inoue, M., Nishizawa, S., & Konishi,
J. (2001). Cortical dysfunction of the supplementary motor area in a spasmodic dysphonia patient.American Journal of Otolaryngology, 22(3), 219–222.
Hutchinson, J. M., & Ringel, R. L. (1975). The effect of oral sensory deprivation on stuttering behavior.Journal of Communication Disorders, 8(3), 249–258.
Ito, K., Nagano-Saito, A., Kato, T., Arahata, Y., Nakamura, A., Kawasumi, Y., Hatano, K., Abe, Y.,Yamada, T., Kachi, T., & Brooks, D. J. (2002). Striatal and extrastriatal dysfunction in Parkinson’sdisease with dementia: A 6-[18F]fluoro-L-dopa PET study.Brain, 125(Pt. 6), 1358–1365.
Jones, R. K. (1966). Observations on stammering after localized cerebral injury.Journal of Neurology,Neurosurgery, and Psychiatry, 29, 192–195.
Jurgens, U. (2002). Neural pathways underlying vocal control.Neuroscience and BiobehavioralReviews, 26(2), 235–258.
Kaji, R., Kohara, N., Katayama, M., Kubori, T., Mezaki, T., Shibasaki, H., & Kimura, J. (1995). Muscleafferent block by intramuscular injection of lidocaine for the treatment of writer’s cramp.Muscleand Nerve, 18(2), 234–235.
Kaji, R., Rothwell, J. C., Katayama, M., Ikeda, T., Kubori, T., Kohara, N., Mezaki, T., Shibasaki, H.,& Kimura, J. (1995). Tonic vibration reflex and muscle afferent block in writer’s cramp.Annals ofNeurology, 38(2), 155–162.
Kalinowski, J., Stuart, A., Rastatter, M. P., Snyder, G., & Dayalu, V. (2000). Inducement of fluentspeech in persons who stutter via visual choral speech.Neuroscience Letters, 281(2/3), 198–200.
Kiziltan, G., & Akalin, M. A. (1996). Stuttering may be a type of action dystonia.Movement Disorders,11(3), 278–282.
Klaff, A. (1976). Mass generalised learning applied to the problem of carry-over in articulation therapy.Journal of South African Speech Hearing Association, 23, 30–43.
Kono, I., Hirano, T., Ueda, Y., & Nakajima, K. (1998). A case of acquired stuttering resulting fromstriatocapsular infarction.Rinsho Shinkeigaku, 38(8), 758–761.
Kossoff, E. H., & Singer, H. S. (2001). Tourette syndrome: Clinical characteristics and currentmanagement strategies.Paediatric Drugs, 3(5), 355–363.
Kujirai, T., Caramia, M. D., Rothwell, J. C., Day, B. L., Thompson, P. D., Ferbert, A., Wroe, S.,Asselman, P., & Marsden, C. D. (1993). Corticocortical inhibition in human motor cortex.Journalof Physiology, 471, 501–519.
LaPointe, L. L., & Horner, J. (1981). Palilalia: A descriptive study of pathological reiterative utterances.Journal of Speech and Hearing Disorders, 46, 34–38.
Lebrun, Y., Leleux, C., & Retif, J. (1987). Neurogenic stuttering.Acta Neurochirurgica (Wien), 85(3/4),103–109.

292 C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295
Lee, H. J., Lee, H. S., Kim, L., Lee, M. S., Suh, K. Y., & Kwak, D. I. (2001). A case ofrisperidone-induced stuttering.Journal of Clinical Psychopharmacology, 21(1), 115–116.
Leenders, K. L., Frackowiak, R. S., Quinn, N., Brooks, D., Sumner, D., & Marsden, C. D. (1986).Ipsilateral blepharospasm and contralateral hemidystonia and Parkinsonism in a patient with aunilateral rostral brainstem-thalamic lesion: Structural and functional abnormalities studied withCT, MRI, and PET scanning.Movement Disorders, 1(1), 51–58.
Levelt, W. J., Praamstra, P., Meyer, A. S., Helenius, P., & Salmelin, R. (1998). An MEG study of picturenaming.Journal of Cognitive Neuroscience, 10(5), 553–567.
Ludlow, C. L. (1993). Speech tics in Tourette’s syndrome. In G. Blanken, J. Dittman, H. Grimm,J. Marshall, & C.-W. Wallesch (Eds.),Linguistic disorders and pathologies: An internationalhandbook (pp. 504–509). Berlin: Walter de Gruyter.
Ludlow, C. L., & Braun, A. R. (1993). Research evaluating the use of neuropharmacological agentsfor treating stuttering: Possibilities and problems.Journal of Fluency Disorders, 18, 169–182.
Ludlow, C. L., & Lou, G. (1996). Observations on human laryngeal muscle control. In N. Fletcher& P. Davis (Eds.),Controlling complexity and chaos: 9th vocal fold physiology symposium(pp. 201–218). San Diego, CA: Singular Press.
Ludlow, C. L., Nauton, R. F., & Bassich, C. J. (1984). Procedures for the selection of spastic dysphonicpatients for recurrent laryngeal nerve section.Otolaryngology - Head and Neck Surgery, 92, 24–31.
Ludlow, C. L., Polinsky, R. J., Caine, E. D., Bassich, C. J., & Ebert, M. H. (1982). Language and speechabnormalities in Tourette syndrome. In A. J. Friedhoff & T. N. Chase (Eds.),Gilles de la Tourettesyndrome (pp. 351–361). New York: Raven Press.
Ludlow, C. L., Rosenberg, J., Fair, C., Buck, D., Schesselman, S., & Salazar, A. (1986). Brain lesionsassociated with nonfluent aphasia fifteen years following penetrating head injury.Brain,109, 55–80.
Ludlow, C. L., Rosenberg, J., Salazar, A., Grafman, J., & Smutok, M. (1987). Site of penetrating brainlesions causing chronic acquired stuttering.Annals of Neurology, 22, 60–66.
Ludlow, C. L., Schulz, G. M., Yamashita, T., & Deleyiannis, F. W. (1995). Abnormalities in long latencyresponses to superior laryngeal nerve stimulation in adductor spasmodic dysphonia.The Annals ofOtology, Rhinology, and Laryngology, 104, 928–935.
Luessenhop, A. J., Boggs, J. S., LaBorwit, L. J., & Walle, E. L. (1973). Cerebral dominance in stutterersdetermined by Wada testing.Neurology, 23(11), 1190–1192.
Maguire, G. A., Gottschalk, L. A., Riley, G. D., Franklin, D. L., Bechtel, R. J., & Ashurst, J. (1999).Stuttering: Neuropsychiatric features measured by content analysis of speech and the effect ofrisperidone on stuttering severity.Comprehensive Psychiatry, 40(4), 308–314.
Maguire, G. A., Riley, G. D., Franklin, D. L., & Gottschalk, L. A. (2000). Risperidone for the treatmentof stuttering.Journal of Clinical Psychopharmacology, 20(4), 479–482.
Martindale, C. (1976). The grammar of the tic in Gilles de la Tourette’s syndrome.Language andSpeech, 19, 1–20.
Martindale, C. (1977). Syntactic and semantic correlates of verbal tics in Gilles de la Tourette’ssyndrome: A quantitative case study.Brain and Language, 4, 231–247.
Mazzocchi, F., & Vignolo, L. A. (1979). Localization of lesions in aphasia: Clinical CT scan correlationsin stroke patients.Cortex, 15, 627.
McClean, M. D. (1987). Surface EMG recording of the perioral reflexes: Preliminary observations onstutterers and nonstutterers.Journal of Speech and Hearing Research, 30(2), 283–287.
Miller, A. E. (1985). Cessation of stuttering with progressive multiple sclerosis.Neurology, 35(9),1341–1343.
Miller, S., & Watson, B. C. (1992). The relationship between communication attitude, anxiety, anddepression in stutterers and nonstutterers.Journal of Speech and Hearing Research, 35(4), 789–798.
Mouradian, M. S., Paslawski, T., & Shuaib, A. (2000). Return of stuttering after stroke.Brain andLanguage, 73(1), 120–123.
Moutier, C. Y., & Stein, M. B. (1999). The history, epidemiology, and differential diagnosis of socialanxiety disorder.Journal of Clinical Psychiatry, 60(Suppl. 9), 4–8.

C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295 293
Mulligan, H. F., Anderson, T. J., Jones, R. D., Williams, M. J., & Donaldson, I. M. (2001). Dysfluencyand involuntary movements: A new look at developmental stuttering.International Journal ofNeuroscience, 109(1/2), 23–46.
Murry, T., Cannito, M. P., & Woodson, G. E. (1994). Spasmodic dysphonia. Emotional status andbotulinum toxin treatment.Archives of Otolaryngology - Head & Neck Surgery, 120(3), 310–316.
Nash, E. A., & Ludlow, C. L. (1996). Laryngeal muscle activity during speech breaks in adductorspasmodic dysphonia.Laryngoscope, 106, 484–489.
Numminen, J., Salmelin, R., & Hari, R. (1999). Subject’s own speech reduces reactivity of the humanauditory cortex.Neuroscience Letters, 265(2), 119–122.
Odergren, T., Stone-Elander, S., & Ingvar, M. (1998). Cerebral and cerebellar activation in correlationto the action-induced dystonia in writer’s cramp.Movement Disorders, 13(3), 497–508.
Pauls, D. L., Leckman, J. F., & Cohen, D. J. (1993). Familial relationship between Gilles de la Tourette’ssyndrome, attention deficit disorder, learning disabilities, speech disorders, and stuttering [seecomments].Journal of the American Academy of Child and Adolescent Psychiatry, 32, 1044–1050.
Perkins, W. H., Kent, R. D., & Curlee, R. F. (1991). A theory of neuropsycholinguistic function instuttering.Journal of Speech and Hearing Research, 34, 734–752.
Petrosino, L., Fucci, D., Gorman, P., & Harris, D. (1987). Midline and off-midline tongue and right-and left-hand vibrotactile thresholds of stutterers and normal-speaking individuals.Perceptual andMotor Skills, 65(1), 253–254.
Purves, D. (1994). Neural activity and the growth of the brain. Cambridge: Cambridge University Press.Rao, P. R. (1991). Neurogenic stuttering as a manifestation of stroke and a mask of dysphonia.Clinical
Communication Disorders, 1, 31–37.Rasmussen, T., & Milner, B. (1977). The role of early left-brain injury in determining lateralization of
cerebral speech functions.Annals of the New York Academy of Sciences, 299, 355–369.Ratner, S. C., Gawronski, J. J., & Rice, F. E. (1964). The variable conconcurrent action in the language
of children: Effects of delayed speech feedback.The Psychological Record, 14, 47–56.Ratner, N. B., & Sih, C. C. (1987). Effects of gradual increases in sentence length and complexity on
children’s dysfluency.Journal of Speech and Hearing Disorders, 52(3), 278–287.Reich, A., & Till, J. (1983). Phonatory and manual reaction times of women with idiopathic spasmodic
dysphonia.Journal of Speech and Hearing Research, 26(1), 10–18.Reich, A., Till, J., & Goldsmith, H. (1981). Laryngeal and manual reaction times of stuttering and
nonstuttering adults.Journal of Speech and Hearing Research, 24(2), 192–196.Riecker, A., Ackermann, H., Wildgruber, D., Dogil, G., & Grodd, W. (2000). Opposite hemispheric
lateralization effects during speaking and singing at motor cortex, insula and cerebellum.Neuroreport, 11(9), 1997–2000.
Rosenbaum, F., & Jankovic, J. (1988). Task-specific focal tremor and dystonia: Categorization ofoccupational movement disorders.Neurology, 38, 522–527.
Rosenbek, J., Messert, B., Collins, M., & Wertz, R. (1978). Stuttering following brain damage.Brainand Language, 6, 82–96.
Salmelin, R., Helenius, P., & Service, E. (2000). Neurophysiology of fluent and impaired reading:A magnetoencephalographic approach [in process citation].Journal of Clinical Neurophysiology,17(2), 163–174.
Salmelin, R., Schnitzler, A., Schmitz, F., & Freund, H. (2000). Single word reading in developmentalstutterers and fluent speakers.Brain, 123(Pt. 6), 1184–1202.
Salmelin, R., Schnitzler, A., Schmitz, F., Jancke, L., Witte, O. W., & Freund, H. J. (1998). Functionalorganization of the auditory cortex is different in stutterers and fluent speakers.Neuroreport, 9(10),2225–2229.
Sapienza, C. M., Murry, T., & Brown, W. S. J. (1998). Variations in adductor spasmodic dysphonia:Acoustic evidence.Journal of Voice, 12(2), 214–222.
Sarno, M. T. (1968). Speech impairment in Parkinson’s disease.Archives of Physical Medicine andRehabilitation, 49(5), 269–275.
Schaefer, S. D., Roark, R. M., Watson, B. C., Kondraske, G. V., Freeman, F. J., Butsch, R. W., & Pohl, J.(1992). Multichannel electromyographic observations in spasmodic dysphonia patients and normalcontrol subjects.The Annals of Otology, Rhinology, and Laryngology, 101(1), 67–75.

294 C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295
Shibuya, S., Wakayama, Y., Murahashi, M., Aoki, K., & Ozasa, Y. (1998). A management for severeacquired stuttering in a case of pure akinesia syndrome.Rinsho Shinkeigaku, 38(1), 54–56.
Smith, A., Denny, M., Shaffer, L. A., Kelly, E. M., & Hirano, M. (1996). Activity of intrinsic laryngealmuscles in fluent and disfluent speech.Journal of Speech and Hearing Research, 39(2), 329–348.
Smith, A., Luschei, E., Denny, M., Wood, J., Hirano, M., & Badylak, S. (1993). Spectral analyses ofactivity of laryngeal and orofacial muscles in stutterers.Journal of Neurology, Neurosurgery, andPsychiatry, 56(12), 1303–1311.
Sommer, M., Koch, M. A., Paulus, W., Weiller, C., & Buchel, C. (2002). Disconnection ofspeech-relevant brain areas in persistent developmental stuttering.Lancet, 360(9330), 380–383.
Sommer, M., Ruge, D., Tergau, F., Beuche, W., Altenmuller, E., & Paulus, W. (2002). Intracorticalexcitability in the hand motor representation in hand dystonia and blepharospasm.MovementDisorders, 17(5), 1017–1025.
Sommer, M., Wischer, S., Tergau, F., & Paulus, W. (2003). Normal intracortical excitability indevelopmental stuttering.Movement Disorders, 18(7), 826–830.
Soroker, N., Bar-Israel, Y., Schechter, I., & Solzi, P. (1990). Stuttering as a manifestation ofright-hemispheric subcortical stroke.European Neurology, 30(5), 268–270.
Stager, S. V., Denman, D. W., & Ludlow, C. L. (1997). Modifications in aerodynamic variables bypersons who stutter under fluency-evoking conditions.Journal of Speech, Language, and HearingResearch, 40, 832–847.
Stager, S. V., & Ludlow, C. L. (1994). Responses of stutterers and vocal tremor patients to treatment withbotulinum toxin. In J. Jankovic & M. Hallett (Eds.),Therapy with botulinum toxin (pp. 481–490).New York: Marcel Dekker, Inc.
Starkweather, C. W. (2002). The epigenesis of stuttering.Journal of Fluency Disorders, 27(4), 269–288(quiz 287–288).
Stern, E., Silbersweig, D. A., Chee, K. Y., Holmes, A., Robertson, M. M., Trimble, M., Frith, C. D.,Frackowiak, R. S., & Dolan, R. J. (2000). A functional neuroanatomy of tics in Tourette syndrome.Archives of General Psychiatry, 57(8), 741–748.
Tempel, L. W., & Perlmutter, J. S. (1990). Abnormal vibration-induced cerebral blood flow responsesin idiopathic dystonia.Brain, 113, 691–707.
Tempel, L. W., & Perlmutter, J. S. (1993). Abnormal cortical responses in patients with writer’s cramp.Neurology, 43, 2252–2257.
Truong, D. D., Rontal, M., Rolnick, M., Aronson, A. E., & Mistura, K. (1991). Double-blind controlledstudy of botulinum toxin in adductor spasmodic dysphonia.Laryngoscope, 101, 630–634.
Tsumoto, T., Nishioka, K., Nakakita, K., Hayashi, S., & Maeshima, S. (1999). Acquired stutteringassociated with callosal infarction: A case report.No Shinkei Geka, 27(1), 79–83.
Turgut, N., Utku, U., & Balci, K. (2002). A case of acquired stuttering resulting from left parietalinfarction.Acta Neurologica Scandinavica, 105(5), 408–410.
Van Borsel, J., & Taillieu, C. (2001). Neurogenic stuttering versus developmental stuttering: Anobserver judgement study.Journal of Communication Disorders, 34(5), 385–395.
Van Pelt, F., Ludlow, C. L., & Smith, P. J. (1994). Comparison of muscle activation patterns in adductorand abductor spasmodic dysphonia.The Annals of Otology, Rhinology, and Laryngology, 103, 192–200.
Webster, W. G. (1990). Evidence in bimanual finger-tapping of an attentional component to stuttering.Behavioral Brain Research, 37, 93–100.
Wise, R. J., Greene, J., Buchel, C., & Scott, S. K. (1999). Brain regions involved in articulation.Lancet,353(9158), 1057–1061.
Wood, F., Stump, D., McKennhan, A., Sheldon, S., & Proctor, J. (1980). Patterns of regional cerebralblood flow during attempted reading aloud by stutterers both on and off haloperidol medication:Evidence for inadequate left frontal activation during stuttering.Brain and Language, 9, 141–144.
Wu, J. C., Maguire, G., Riley, G., Fallon, J., LaCasse, L., Chin, S., Klein, E., Tang, C., Cadwell, S., &Lottenberg, S. (1995). A positron emission tomography [18F]deoxyglucose study of developmentalstuttering.Neuroreport, 6, 501–505.

C.L. Ludlow, T. Loucks / Journal of Fluency Disorders 28 (2003) 273–295 295
Yoshida, K., Kaji, R., Kubori, T., Kohara, N., Iizuka, T., & Kimura, J. (1998). Muscle afferent blockfor the treatment of oromandibular dystonia.Movement Disorders, 13(4), 699–705.
Zimmermann, G. N., Smith, A., & Hanley, J. M. (1981). Stuttering: In need of a unifying conceptualframework.Journal of Speech and Hearing Research, 24(1), 25–31.
CONTINUING EDUCATION
Stuttering: a dynamic motor control disorder
QUESTIONS
1. The brain function abnormalities implicated in stuttering are:a. Right language dominanceb. Left hemisphere injuryc. Basal ganglia diseased. Abnormalities in the function of the neural network for speech
2. Brain injuries which cause stuttering in adults involve:a. Lesions only in Broca’s areab. Lesions only in Wernicke’s areac. Lesions only in the basal gangliad. Lesions outside Broca’s and Wernicke’s area
3. The similarities between stuttering and dystonias are:a. Both are task specific and involve emotional expressionb. Both are task specific, focal and intermittentc. Both are affected by auditory feedbackd. Both affect other disorders besides speech
4. Muscle activity in both stuttering and spasmodic dysphonia:a. Is abnormal only intermittently in both disordersb. Is constantly too high in spasmodic dysphonia but not in stutteringc. Is constantly too high in stuttering but not in spasmodic dysphoniad. Is normal all of the time in both disorders
5. Neurophysiological studies of persons who stutter indicates that stuttering in-volves:a. Language abnormalitiesb. Hearing abnormalitiesc. Sensory lossd. None of the above




















