
Studies on Trypanosomatid Actin I. Immunochemical and Biochemical Identification
6
8 J. PROTOZOOL., VOL. 36. NO. 1, JANUARY-FEBRUARY 1989 in Tetrahymena thermophila and Tetrahymetlapyrifortnis. J. Protozool., 32: 640-644. 28. Van Bell, C. T. 1986. 5s and 5.8s ribosomal evolution in the suborder Tetrahymenina (Ciliophora: Hymenostomatida). J. Mol. Evol., 22: 231-236. 29. Vaughn, J. C., Sperbech, S. J., Ramsey, W. J. & Lawrence, C. B. 1984. A universal model for the secondary structure of 5.8s ribosomal RNA molecules, their contact sites with 28s ribosomal RNA molecules and their prokaryotic equivalent. Nucierc Acids Res., 12: 7479-7502. 30. Williams, N. E., Buhse, H. E., Jr. & Smith, M. G. 1984. Protein similarities in the genus Tetrahymena and a description of Tetrahymena leucophrys n. sp. J. Protozool., 31: 313-321. 3 I. Woese, C. & Fox, G. E. 1977. Phylogenetic studies of the pro- karyotic domain: the primary kingdoms. Proc. Natl. Acad. Sci. USA, 74: 5088-5090. Received 3-22-88; accepted 6-27-88 J. Protozoo/.. 36(1), 1989. pp. 8-13 C 1989 by the Society of Protozoolousts Studies on Trypanosomatid Actin I. Immunochemical and Biochemical Identification RENATO A. MORTARA Department of Microbiology. Immunology and Parasitology, Escola Paulista de Medicina, Rua Botucatu, 862 6" andar, SGo Paul0 04023, Brad ABSTRACT. In this study, the presence of actin in cultured trypanosomatids was investigated using polyclonal antibodies to heter- ologous actin. Polyclonal antisera to rabbit muscle actin and a monospecific anti-actin antibody react with a 43-kDa polypeptide in extracts of Trypanosoma cruzi, Herpetomonas samuelpessoai and Leishmania mexicana amazonensis on protein immunoblots. The 43-kDa polypeptide co-migrates with skeletal muscle actin and is retained within trypanosomatid cytoskeletons. Attempts to isolate H. samuelpessoai actin through DNase I affinity chromatography showed that the 43-kDa polypeptide did not bind to the column. Instead, low yields of a 47-kDa polypeptide were obtained indicating that the trypanosomatid actin displays unusual DNase I binding behavior when compared to actins from higher eukaryotes. Immunofluorescence studies confirmed that cytoskeletons retain the actin- like protein. In H. sarnuelpessoai, actin is localized in the region close to the flagellum, whereas in T. cruzi it is more homogeneously distributed. The data presented here show that trypanosomatid actin displays biochemical characteristics similar to actins of other protozoa. Key words. DNase I binding, Herpetomonas samuelpessoai, immunoblot, immunofluorescence, Leishmania mexicana amazonenis, polyclonal antibodies, Trypansoma cruzi. CTIN is a highly conserved and ubiquitous protein essential A for the maintenance of complex structural and dynamic functions of eukaryotic cells. Typically, actin is localized in the cytoplasm, assembled in the filamentous (F) form usually found in equilibrium with the monomeric G form 11, 10, 271. The presence of actin in trypanosomatids was first demonstrated a few years ago by De Souza et al. [5]. Using heterologous anti- actin antibodies, they showed that T. cruzi epimastigotes react positively by indirect immunofluorescence IS]. In spite of nu- merous ultrastructural studies on different trypanosomatids [2, 6, 12, 20, 251, however, typical 5-7-nm microfilaments have not yet been described in these flagellates, suggesting that try- panosomatid actin might possess unique structural and assem- bling properties. Furthermore, the available data on the molec- ular nature of the immuno-reactive actin analogue from T. cruzi epimastigotes and the putative actin protein of other trypano- somatids is limited. In the current study the presence of actin in several trypanosomatids was examined using a combination of immunochemical and biochemical techniques. Evidence is presented that trypanosomatid actin has an apparent molecular weight of 43 kDa and has some similarities with actin isolated from other protozoa. MATERIALS AND METHODS Trypanosomatids. Leishmania mexicana amazonensis pro- mastigotes (strain MHOM/BR/73/M2269)and T. cruzi (G strain) epimastigotes and metacyclic trypomastigotes [22, 301 were ob- tained from cultures in liver infusion tryptose (LIT) medium [4]. Metacyclic trypomastigotes were purified as previously de- scribed [22]. Herpetomonas samuelpessoai (ATCC 30252) pro- mastigotes were grown in FYTS medium [ 181, and for experi- ments with microfilament-interfering drugs, 10 pg/ml cytochalasin B or 2 pM phalloidin (both drugs from Sigma Chem. Co.) were added to the medium. Preparation of cytoskeletons. Trypanosomatid cytoskeletons were prepared through minor modifications of the method de- scribed by Ben-Ze'ev et al. 131. Cells were harvested from cultures by centrifugation at 5,000 g for 5 min and washed in phosphate- buffered saline solution (PBS) 3 times; the cell pellet was cooled to 0" C and lysed in the following buffer: 150 mM NaCI, 30 rnM piperazine-N,N'-bis[2-ethane sulfonic acid] (PIPES), pH 7.3, 1 mM MgCl,, 0.1 mM ethyleneglycol-bis-(P-amino-ethyl ether)N,N'-tetra acetic acid (EGTA), 1 .O% (v/v) Nonidet-P40 and the following protease inhibitors: 1 m M phenylmethylsul- fonyl fluoride (PMSF), 3 mM benzamidine, 3 mM o-phenan- throline, and 1 &ml of leupeptin, aprotinin and antipain. The lysate was left at 0" C for 10 min and examined by phase-contrast microscopy to check for the integrity of the cytoskeletons. These cellular lysates containing intact cytoskeletons were diluted 10- to 50-fold in ice-cold PBS and the insoluble components col- lected by centrifugation onto a 55% sucrose cushion at 10,000 g for 30 min at 4" C. Antibodies. Rabbit polyclonal antisera to skeletal muscle actin were from Miles-Yeda (a gift from Wanderley de Souza, Insti- tuto de Biofisica, Rio de Janeiro) and prepared by S. Avrameas (Pasteur Institute, Paris). The latter was provided as an affinity- purified 5.6 mg/ml IgG solution in PBS/30% glycerol. These antisera were further affinitypurified as follows: 1 mg of rabbit skeletal muscle F-actin prepared as described 1171was adsorbed onto 1 -cm2pieces of nitrocellulose and air dried the pieces were
Transcript of Studies on Trypanosomatid Actin I. Immunochemical and Biochemical Identification

8 J . PROTOZOOL., VOL. 36. NO. 1, JANUARY-FEBRUARY 1989
in Tetrahymena thermophila and Tetrahymetlapyrifortnis. J . Protozool., 32: 640-644.
28. Van Bell, C. T. 1986. 5s and 5.8s ribosomal evolution in the suborder Tetrahymenina (Ciliophora: Hymenostomatida). J. Mol. Evol., 22: 231-236.
29. Vaughn, J. C., Sperbech, S. J., Ramsey, W. J. & Lawrence, C. B. 1984. A universal model for the secondary structure of 5.8s ribosomal RNA molecules, their contact sites with 28s ribosomal RNA molecules and their prokaryotic equivalent. Nucierc Acids Res., 12: 7479-7502.
30. Williams, N. E., Buhse, H. E., Jr. & Smith, M. G. 1984. Protein similarities in the genus Tetrahymena and a description of Tetrahymena leucophrys n. sp. J. Protozool., 31: 313-321.
3 I . Woese, C. & Fox, G. E. 1977. Phylogenetic studies of the pro- karyotic domain: the primary kingdoms. Proc. Natl. Acad. Sci. USA, 74: 5088-5090.
Received 3-22-88; accepted 6-27-88
J. Protozoo/.. 36(1), 1989. pp. 8-13 C 1989 by the Society of Protozoolousts
Studies on Trypanosomatid Actin I. Immunochemical and Biochemical Identification
RENATO A. MORTARA Department of Microbiology. Immunology and Parasitology, Escola Paulista de Medicina, Rua Botucatu,
862 6" andar, SGo Paul0 04023, B r a d
ABSTRACT. In this study, the presence of actin in cultured trypanosomatids was investigated using polyclonal antibodies to heter- ologous actin. Polyclonal antisera to rabbit muscle actin and a monospecific anti-actin antibody react with a 43-kDa polypeptide in extracts of Trypanosoma cruzi, Herpetomonas samuelpessoai and Leishmania mexicana amazonensis on protein immunoblots. The 43-kDa polypeptide co-migrates with skeletal muscle actin and is retained within trypanosomatid cytoskeletons. Attempts to isolate H. samuelpessoai actin through DNase I affinity chromatography showed that the 43-kDa polypeptide did not bind to the column. Instead, low yields of a 47-kDa polypeptide were obtained indicating that the trypanosomatid actin displays unusual DNase I binding behavior when compared to actins from higher eukaryotes. Immunofluorescence studies confirmed that cytoskeletons retain the actin- like protein. In H. sarnuelpessoai, actin is localized in the region close to the flagellum, whereas in T. cruzi it is more homogeneously distributed. The data presented here show that trypanosomatid actin displays biochemical characteristics similar to actins of other protozoa.
Key words. DNase I binding, Herpetomonas samuelpessoai, immunoblot, immunofluorescence, Leishmania mexicana amazonenis, polyclonal antibodies, Trypansoma cruzi.
CTIN is a highly conserved and ubiquitous protein essential A for the maintenance of complex structural and dynamic functions of eukaryotic cells. Typically, actin is localized in the cytoplasm, assembled in the filamentous (F) form usually found in equilibrium with the monomeric G form 11, 10, 271. The presence of actin in trypanosomatids was first demonstrated a few years ago by De Souza et al. [ 5 ] . Using heterologous anti- actin antibodies, they showed that T. cruzi epimastigotes react positively by indirect immunofluorescence IS]. In spite of nu- merous ultrastructural studies on different trypanosomatids [2, 6, 12, 20, 251, however, typical 5-7-nm microfilaments have not yet been described in these flagellates, suggesting that try- panosomatid actin might possess unique structural and assem- bling properties. Furthermore, the available data on the molec- ular nature of the immuno-reactive actin analogue from T. cruzi epimastigotes and the putative actin protein of other trypano- somatids is limited. In the current study the presence of actin in several trypanosomatids was examined using a combination of immunochemical and biochemical techniques. Evidence is presented that trypanosomatid actin has an apparent molecular weight of 43 kDa and has some similarities with actin isolated from other protozoa.
MATERIALS AND METHODS Trypanosomatids. Leishmania mexicana amazonensis pro-
mastigotes (strain MHOM/BR/73/M2269)and T. cruzi (G strain) epimastigotes and metacyclic trypomastigotes [22, 301 were ob- tained from cultures in liver infusion tryptose (LIT) medium [4]. Metacyclic trypomastigotes were purified as previously de- scribed [22]. Herpetomonas samuelpessoai (ATCC 30252) pro-
mastigotes were grown in FYTS medium [ 181, and for experi- ments with microfilament-interfering drugs, 10 pg/ml cytochalasin B or 2 pM phalloidin (both drugs from Sigma Chem. Co.) were added to the medium.
Preparation of cytoskeletons. Trypanosomatid cytoskeletons were prepared through minor modifications of the method de- scribed by Ben-Ze'ev et al. 131. Cells were harvested from cultures by centrifugation at 5,000 g for 5 min and washed in phosphate- buffered saline solution (PBS) 3 times; the cell pellet was cooled to 0" C and lysed in the following buffer: 150 mM NaCI, 30 rnM piperazine-N,N'-bis[2-ethane sulfonic acid] (PIPES), pH 7.3, 1 mM MgCl,, 0.1 mM ethyleneglycol-bis-(P-amino-ethyl ether)N,N'-tetra acetic acid (EGTA), 1 .O% (v/v) Nonidet-P40 and the following protease inhibitors: 1 mM phenylmethylsul- fonyl fluoride (PMSF), 3 m M benzamidine, 3 mM o-phenan- throline, and 1 &ml of leupeptin, aprotinin and antipain. The lysate was left at 0" C for 10 min and examined by phase-contrast microscopy to check for the integrity of the cytoskeletons. These cellular lysates containing intact cytoskeletons were diluted 10- to 50-fold in ice-cold PBS and the insoluble components col- lected by centrifugation onto a 55% sucrose cushion at 10,000 g for 30 min at 4" C.
Antibodies. Rabbit polyclonal antisera to skeletal muscle actin were from Miles-Yeda (a gift from Wanderley de Souza, Insti- tuto de Biofisica, Rio de Janeiro) and prepared by S. Avrameas (Pasteur Institute, Paris). The latter was provided as an affinity- purified 5.6 mg/ml IgG solution in PBS/30% glycerol. These antisera were further affinity purified as follows: 1 mg of rabbit skeletal muscle F-actin prepared as described 1171 was adsorbed onto 1 -cm2 pieces of nitrocellulose and air dried the pieces were

MORTARA - TRY PANOSOMATID ACTIN 9
blocked in blot buffer (1 50 mM NaC1, 1 mM ethylene diamine- tetra acetic acid [EDTA], 30 mM Tris-HC1 [pH 7.31, 0.25% gelatin and 0.05% NaN,) and rinsed in PBS before incubation for 2 h with the rabbit antisera diluted 1:lO in blot buffer. The immunoadsorbent pieces were washed with several changes of PBS and the bound immunoglobulins eluted with 1 ml of ice- cold 0.1 M glycine, pH 2.8, for 5 min; the solution was then neutralized with 30 pl of 2 M Tris, and 100 pl of 10 x blot buffer was added.
Anti-tubulin monoclonal antibody, YOL 1/34, was kindly provided by John Kilmartin [9]. Phalloidin coupled to rhoda- mine was a generous gift from Dr. H. Faulstich [7].
Immunofluorescence. For indirect immunofluorescence, sam- ples were fixed in freshly made 4% paraformaldehyde/PBS for 30 min at room temperature. After 3 washes with PBS and a 15-min incubation in 50 mM NH,Cl/PBS to quench excess aldehyde groups, the specimens were adsorbed onto multi-well slides, air-dried and stored dry at -20" C. Alternatively, cells were adsorbed onto poly-L-lysine-coated coverslips and lysed as described above before fixation. The slides (or coverslips) were soaked in blot buffer containing 0.05% saponin (Sigma) for at least 1 h and incubated for 1 h with the antibodies in a wet chamber. Subsequently, samples were washed 3 times by soaking in blot buffer with saponin and treated with the appro- priate FITC conjugates (Dako Corporation). After washing the conjugate as above, slides were mounted in 90% glycerol/0.2 M glycine, pH 9.2. Photographs were then taken using black and white Kodak Tri-X 400 ASA on a Zeiss standard WL micro- scope (Neophluor 100 x N.A. objective) or with color Fujicolor Super HR 400 ASA and Fujichrome 1600 ASA films on a Nikon UFX microscope (Planapo 100 x objective). Both microscopes were equipped for epifluorescence with selective FITC and rho- damine filters.
Sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE). For SDS-PAGE [ l 11 samples were dissolved in boiling SDS-PAGE sample buffer containing 10% SDS and 10% P-mercaptoethanol and electrophoresed in 6-1 6% polyacryl- amide linear gradient microslab gels prepared according to Mat- sudaira & Burgess [13]. Gels were stained for proteins with Coomassie brilliant blue or by the silver stain method [29]. Iodination of surface polypeptides from T. cruzi G strain meta- cyclics was carried out as described in detail elsewhere [22, 301. Radioactive polypeptides were visualized by autoradiography of dried gels exposed to X-ray films at -70" C in the presence of intensifying screens.
Mouse skeletal muscle extract. Mouse skeletal muscle extracts used as a source of skeletal muscle actin were prepared by ho- mogenization of Balb/c skeletal leg muscles followed by 5 wash- es in PBS [ 171.
Immunoblotting. This was camed out essentially as described by Towbin et al. [23]. After electrophoresis, proteins were trans- ferred to nitrocellulose sheets (Millipore, 0.45 pm pore size) for at least 2 h at 200 mA. The transferred polypeptides and mo- lecular weight markers were visualized by staining the sheets with 0.lo/o (w/v) Ponceau S in 10% acetic acid. After incubation with the antibodies for 1 h at room temperature, the sheets were subjected to 3 washes of 15 min each under constant motion in blot buffer containing 0.01% (v/v) Tween 20. Bound immu- noglobulins were visualized after incubation for 1 h with the appropriate anti-immunoglobulin (diluted 1 :200 in blot buffer) coupled to horseradish peroxidase (Dako) following 3 washes of 10 min each in PBS and reaction with diaminobenzidine (0.2 mg/ml) and 5 pl of a 30% H202 solution in 30 ml PBS.
DNase I affinity chromatography. The isolation of actin from cutured trypanosomatids was based on the DNase I-affinity chromatography method described by Zechel [3 11. Herpeto-
Fig. 1. Identification of immuno-reactive actin analogue in cultured trypanosomatids with polyclonal antisera to actin. Immunoblotting re- action of polyclonal anti-actin antiserum against cultured trypanoso- matid extracts (2 x lo6 cells): reaction of anti-rabbit muscle actin IgG (250 pg/ml, provided by S. Avrameas) with mouse skeletal muscle (lane 1) and Herpetomonas samuelpessoai extracts (lane 2); reaction of the same serum after affinity purification as described in Materials and Methods (unless otherwise stated, this serum is referred to as the mono- specific anti-actin antibody) with mouse muscle (lane 3), H . samuel- pessoai (lane 4), Trypanosoma cruzi epimastigotes (G strain) (lane 5) , and Leishmania mexicana amazonensis (lane 6 ) extracts. Note the in- creased reaction with H. samuelpessoai 43-kDa band after affinity pu- rification by comparing lanes 2 and 4. Molecular weight standards in- dicated on the right are: phosphorylase B 94 kDa, bovine serum albumin 67 kDa, egg white ovalbumin 43 kDa, carbonic anhydrase 30 kDa, soybean trypsin inhibitor 20 kDa and a-lactalbumin 14 kDa. Arrowhead indicates the migration of the 43-kDa mouse actin band. Control strips incubated with normal rabbit IgG (25 pg/ml) under identical conditions gave no reaction.
monas samuelpessoai (3.9 5 x 1 O I o cells, approximately 19 5 mg total protein) were homogenized in the buffer described by Ze- chel [31], also containing 1% 0-octyl-glucoside (Sigma) to im- prove membrane disruption and the protease inhibitor mixture described above, The homogenate was incubated for 20 min at room temperature and centrifuged at 10,000 g for 30 min; the supernatant fluid was applied to a 6-ml DNase I-Sepharose CL- 4B column containing 20 mg of coupled DNase I (DN-CS grade, Sigma). The elution was camed out as described [30] and the fractions ( I .5 ml) recovered from the column with 40% form- amide were pooled, dialyzed against 10 mM Tris-HCI, pH 7.4, lyophilized and subsequently analyzed by SDS-PAGE. The ef- ficiency of the column was previously checked by the effective isolation of mouse skeletal muscle actin from muscle homog- enates.
RESULTS Detection of actin in trypanosomatid extracts by protein im-
munoblotting. The presence of an actin-like protein was inves- tigated through the reaction of a polyclonal antiserum to rabbit muscle actin. The reactivity pattern of the antiserum towards skeletal muscle and H. samuelpessoai extracts shown in Fig. 1 (lanes 1 and 2) reveals that it recognizes actin and a 33-kDa polypeptide in muscle extracts (lane 1); however, several poly- peptides in H . samuelpessoai extracts gave positive reactions, including those around 43 kDa (47, 43 and 40 kDa). Further

10 J. PROTOZOOL., VOL. 36, NO. 1, JANUARY-FEBRUARY 1989
Fig. 2. DNase I affinity chromatography of Herpetomonas samuelpessoai proteins. The H . samuelpessoai homogenate, processed as described in Materials and Methods, was chromatographed in a DNase ISepharose column. (A) Elution profile of the fractions monitored at OD,,, nm. The position at which 40% formamide was added to the elution buffer is indicated. (B) SDS-PAGE of samples from the DNase I column stained for protein by the silver stain method [29]. Lane m, mouse skeletal muscle extract; lane h, a sample of H. samuelpessoai homogenate applied to the column; lane rt, material not bound to the column; lane b, a fraction equivalent to 10% of the material eluted with 40% formamide representing the components bound to the column. The position of mouse muscle actin is indicated, and an arrowhead also indicates the predominant band of the bound material that migrates with apparent molecular mass of 47 kDa; (C) immunoblotting analysis of the samples as indicated in 2B, with monospecific rabbit anti-actin antibodies. Arrowhead indicates the 43-kDa polypeptide recognized by the monospecific anti-actin antibodies.
affinity purification of this antiserum with rabbit muscle F-actin yielded a monospecific anti-actin that specifically recognizes mouse muscle actin (Fig. 1, lane 3). This monospecific serum reacts with a 43-kDa component in H. samuelpessoai promas- tigotes, T. cruzi epimastigotes and L. mexicana amazonensis promastigotes (Fig. 1, lanes 4, 5 and 6). Since this component is also present in muscle extracts and co-migrates with actin, it was concluded that trypanosomatids contain an actin protein of 43 kDa. Similar experiments camed out after affinity puri- fication of another polyclonal antiserum to rabbit muscle actin (from Miles-Yeda) gave essentially the same results (data not shown). The above immunochemical studies show that cultured trypanosomatids express a 43-kDa actin polypeptide that is rec- ognized by polyclonal monospecific antibodies to heterologous actin and also co-migrates with skeletal muscle actin. The pos- sibility remains, however, that other trypanosomatid polypep- tides (e.g. a 94-kDa polypeptide in L. mexicana amazonensis) may contain epitopes common to vertebrate actins.
DNase I affinity chromatography of Herpetomonas samuel- pessoui extracts. Taking advantage of its ease of handling and growth in relatively large quantities, H. samuelpessoai was cho- sen for the following experiments. In order to isolate actin from H . samuelpessoai, the DNase I affinity chromatography method [31] was employed. The elution profile of H. samuelpessoai proteins on DNase I-Sepharose shows that a peak of material is specifically eluted with 40% formamide (Fig. 2A). The SDS- PAGE analysis revealed that the bound material migrates with an apparent molecular weight of 47 kDa (Fig. 2B). It was also found that this fraction contained nucleic acids, which probably
contributed to the increase in the measured OD,,,. The unbound material contains a 43-kDa component that co-migrates with mouse muscle actin and reacts with the monospecific anti-actin antibodies (lane rt in Fig. 2C), indicating that the 43-kDa actin polypeptide did not bind to the immobilized DNase I.
Retention of trypanosomatid actin in cytoskeletons. Cytoskel- etons are biochemically defined as the detergent-resistant cel- lular components. Under phase-contrast microscopy, cytoskel- etons closely resemble cells that have lost their characteristic membrane refringency and resemble intact cell “ghosts.” In order to establish whether the 43-kDa actin protein is associated with the trypanosomatid cytoskeleton, SDS-PAGE and im- munoblots were camed out to check for the presence of actin after non-ionic detergent cell lysis.
Analysis by SDS-PAGE shows that several H. samuelpessoai and T. cruzi polypeptides are selectively extracted in the lysate, whereas others are concentrated in the cytoskeletal fraction (Fig. 3, lanes L and C). Immunoblotting identification with mono- specific anti-actin antibodies revealed that most of the 43-kDa actin band is retained in the cytoskeletal fraction after lysis of both H . samuelpessoai promastigotes and T. cruzi metacyclics (Fig. 3, right panel). The fact that no major 43-kDa band on the protein-stained gel displayed similar behavior suggests that actin is probably a minor component among the proteins of both trypanosomatids. Immunochemical experiments also showed that tubulin retention parallels that of actin in that it remains associated with the cytoskeletal fraction (Fig. 4, left panel). The surface profile from T. cruzi metacyclics is well characterized and comprises polypeptides of molecular mass
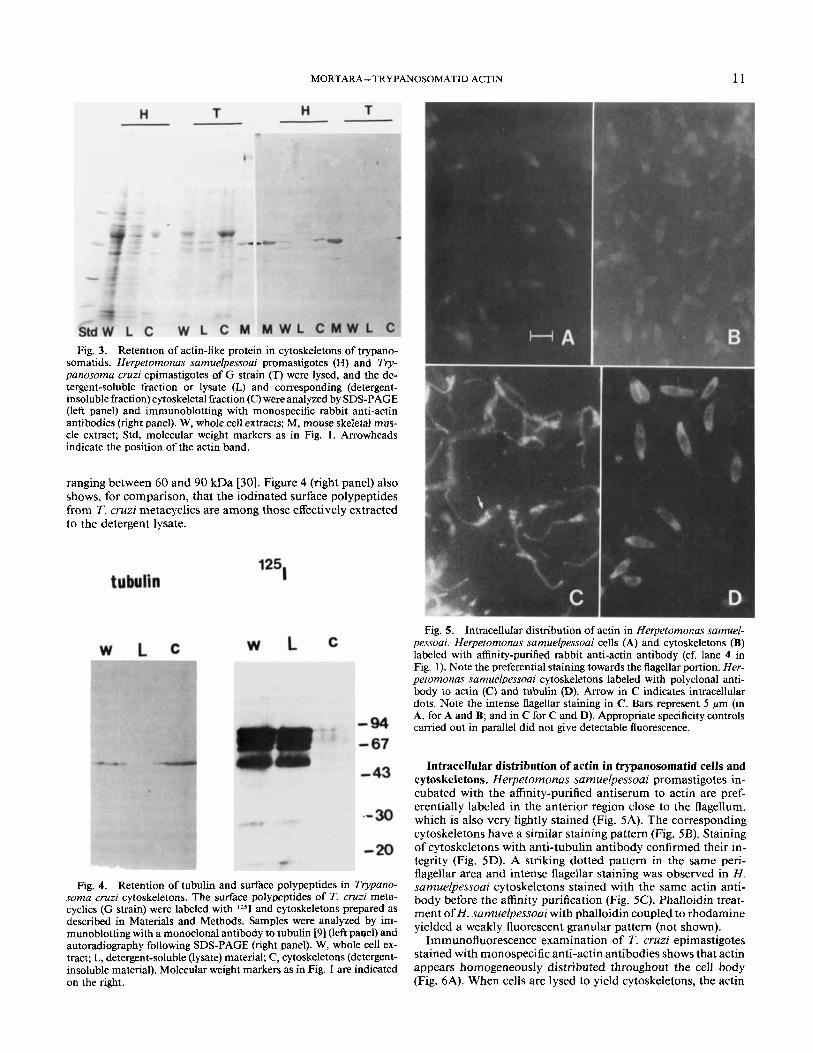
MORTARA- TRYPANOSOMATID ACTIN 11
Fig. 3. Retention of actin-like protein in cytoskeletons of trypano- somatids. Herpetomonas samuelpessoai promastigotes (H) and Try- panosoma cruzi epimastigotes of G strain (T) were lysed, and the de- tergent-soluble fraction or lysate (L) and corresponding (detergent- insoluble fraction) cytoskeletal fraction (C) were analyzed by SDS-PAGE (left panel) and immunoblotting with monospecific rabbit anti-actin antibodies (right panel). W, whole cell extracts; M, mouse skeletal mus- cle extract; Std, molecular weight markers as in Fig. 1. Arrowheads indicate the position of the actin band.
ranging between 60 and 90 kDa [30]. Figure 4 (right panel) also shows, for comparison, that the iodinated surface polypeptides from T. cruzi metacyclics are among those effectively extracted to the detergent lysate.
Fig. 5 . Intracellular distribution of actin in Herpetomonas samuel- pessoai. Herpetomonas samuelpessoai cells (A) and cytoskeletons (B) labeled with affinity-purified rabbit anti-actin antibody (cf. lane 4 in Fig. 1). Note the preferential staining towards the flagellar portion. Her- petomonas samuelpessoai cytoskeletons labeled with polyclonal anti- body to actin (C) and tubulin (D). Arrow in C indicates intracellular dots. Note the intense flagellar staining in C. Bars represent 5 Mm (in A, for A and B; and in C for C and D). Appropriate specificity controls carried out in parallel did not give detectable fluorescence.
Fig. 4. Retention of tubulin and surface polypeptides in Trypano- soma cruzi cytoskeletons. The surface polypeptides of T. cruzi meta- cyclics (G strain) were labeled with 1251 and cytoskeletons prepared as described in Materials and Methods. Samples were analyzed by im- munoblotting with a monoclonal antibody to tubulin [9] (left paqel) and autoradiography following SDS-PAGE (right panel). W, whole cell ex- tract; L, detergent-soluble (lysate) material; C, cytoskeletons (detergent- insoluble material). Molecular weight markers as in Fig. 1 are indicated on the right.
Intracellular distribution of actin in trypanosomatid cells and cytoskeletons. Herpetomonas sarnuelpessoai promastigotes in- cubated with the affinity-purified antiserum to actin are pref- erentially labeled in the anterior region close to the flagellum, which is also very lightly stained (Fig. 5A). The corresponding cytoskeletons have a similar staining pattern (Fig. 5B). Staining of cytoskeletons with anti-tubulin antibody confirmed their in- tegrity (Fig. 5D). A striking dotted pattern in the same peri- flagellar area and intense flagellar staining was observed in H. samuelpessoai cytoskeletons stained with the same actin anti- body before the affinity purification (Fig. 5C). Phalloidin treat- ment of H . samuelpessoai with phalloidin coupled to rhodamine yielded a weakly fluorescent granular pattern (not shown).
Immunofluorescence examination of T. cruzi epimastigotes stained with monospecific anti-actin antibodies shows that actin appears homogeneously distributed throughout the cell body (Fig. 6A). When cells are lysed to yield cytoskeletons, the actin

12 J. PROTOZOOL., VOL. 36, NO. 1, JANUARY-FEBRUARY 1989
Fig. 6 . Intracellular localization of actin in Trypuno~ornu cruzi. Sus- pensions of T. cruzi (G strain) epimastigotes, containing a few cells already differentiated into metacyclic trypomastigotes, were processed for immunofluorescence as described in Materials and Methods. (A) Trypanosoma cruzi cells labeled with monospecific anti-actin antibody. (B) Trypanosoma cruzi cytoskeletons labeled with monospecific anti- actin antibodies. (C) Trypanomma cruzi cells labeled with rhodamine- phalloidin. Arrows in A, B and C indicate granular structures. Trypa- nosoma cruzi cells (D) and cytoskeletons (E) labeled with monoclonal antibody to tubulin. Note metacyclic trypornastigotes at the left of the field in D and E. Bar represents 5 pm. Appropriate specificity controls camed out in parallel did not give detectable immunofluorescence.
distribution is not significantly changed (Fig. 6B). It was also noticed that some cells exhibited a discrete granular pattern that was more evident after staining with rhodamine-phalloidin (Fig. 6C). The staining pattern of tubulin in cells is also preserved in cytoskeletons (Fig. 6D. E), thus confirming the integrity of the preparation. These observations confirm the immunochemical evidence presented above that actin and tubulin are retained in trypanosomatid cytoskeletons.
DISCUSSION Evidence has been presented that the cultured trypanoso-
matids H . samuelpessoai, T. cruzi and L. me-xicana amazonen-
sis express a 43-kDa protein that co-migrates with skeletal mus- cle actin and reacts with a monospecific antiserum to heterologous actin.
DNase I binding of Herpetomonas samuelpessoai actin. The 43-kDa actin protein from H. samuelpessoai does not bind to immobilized DNase I, a behavior previously observed with Entamoeba histofytica [ 141 actin. The DNase I chromatography of H. samuelpessoai proteins resulted in the isolation of a 47- kDa polypeptide that did not react with the monospecific anti- actin antiserum to heterologous actin (Fig. 2). A polypeptide of identical molecular weight-47 kDa-reacts quite strongly with the polyclonal anti-actin antibody before its affinity purification (Fig. 1, lane 2). Interestingly, both Tetrahymena pyriformis [8, 151 and E. histolytica [ 141 actins migrate with a higher molecular weight than skeletal muscle actin on SDS-PAGE. Considering the immunological reactivity of the H . samuelpessoai 47-kDa polypeptide with polyclonal anti-actin antibodies and the bind- ing to DNase I, it is tempting to speculate that H. samuelpessoai promastigotes might synthesize 2 distinct forms of actin-43 kDa and 47 kDA. This diversity could be brought about through the processing of a 47-kDa precursor gene product into the 43- kDa protein with concomitant appearance of the reactive epi- tope. Another possibility is that H. samuelpessoai has (at least) 2 distinct actin genes encoding for different forms of actin. The actin gene from T. brucei has recently been cloned and sequenced [cf. ref. 191. The results indicate that T. brucez actin is one of the most divergent among the actin proteins. Considering the availability of DNA probes for protozoan actins [8, 16,241, and now for a trypanosomatid actin [ 191, it should soon be possible to elucidate these possibilities.
Intracellular distribution and possible function of trypanoso- matid actin. A reliable method for the preparation of well-pre- served trypanosomatid cytoskeletons is described in this report (Fig. 3-5). The immunochemical evidence that both actin and tubulin are retained within the cytoskeleton is significant since it suggests that the pool of soluble microtubular and microfil- amentous components is rather small. These observations were confirmed after immunofluorescence of the corresponding cy- toskeletons. Although the observation of tubulin retention was not unexpected [e.g. ref. 2 11, the parallel association of actin not organized in its typical filamentous form is intriguing. Actin distribution is different in H. samuelpessoai and T. cruzi. In H . samuelpessoai it is concentrated near the flagellar pocket, sug- gesting that it may be related to endocytic and exocytic functions [26]. In T. cruzi such distribution is not evident, but there is a clear indication that the trypanosomatid actin may be concen- trated in some areas (Fig. 5). Both results, however, would be compatible with its involvement in an as yet undefined form of intracellular compartmentalization or transport.
It was previously reported that heterologous anti-actin anti- bodies heavily stain flagella of T. cruzi epimastigotes, and it was suggested that microfilaments could be involved in the paraxial rod structural organization [5]. The results presented in the pres- ent study indicate that heavy flagellar staining (in H . samuel- pessoai) can be observed with heterologous polyclonal antisera, but becomes almost undetectable after affinity purification of the reagent. These observations suggest that there is very little of the 43-kDa protein in the flagellum of this trypanosomatid.
It is widely accepted that in non-muscle cells, actin plays key structural and dynamic roles through the constant modification of microfilament length and cross-linking modulated by acces- sory proteins [ I , 10, 271. Thus, cytochalasin and phalloidin, drugs which interfere with the assembly and disassembly of actin filaments, are highly toxic and may eventually kill treated cells [ i , 281. In preliminary experiments, it was observed that cy- tochalasin B (25 clg/ml) or phalloidin (2 &M), when present in

MORTARA- TRYPANOSOMATID ACTIN 13
culture medium at concentrations capable of inhibiting the growth of mouse 3T3 fibroblasts, did not affect the growth rate, mobility or the morphology of H . samuefpessoai promastigotes for a t least 3 days. The suggestion that these flagellates may not require actin polymerization/depolymerization for locomotion, growth and division indicates that there may be other unknown functions of trypanosomatid actin.The isolation of trypanoso- matid actin in sufficient amounts for further biochemical and immunochemical studies, and the precise definition by electron microscopy of the intracellular location of actin with specific antibodies should provide useful clues to these questions.
ACKNOWLEDGMENTS Supported by grants from FundagBo de Amparo ti Pesquisa
do Estado de SBo Paulo (FAPESP) and Conselho Nacional d e Desenvolvimento Cientifico e Tecnol6gico (CNPq) from Brazil. I am indebted to Erney Camargo for initial encouragement. I thank Nobuko Yoshida for helpful discussions and support throughout the work and J. Franco da Silveira for critically reading the manuscript. I am grateful to Mari Armelin for the access to the fluorescence microscope. I also thank Thom See- beck for his interest and for supplying a preprint of the work of his group.
LITERATURE CITED 1. Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K. & Watson,
J. D., ed. 1983. The cytoskeleton. In: The molecular biology of the cell. Garland Publishing Inc., New York, pp. 549-609.
2. Angelopoulos, E. V. 1970. Pellicular microtubules in the family Trypanosomatidae. J. Protozool., 17: 39-5 1 .
3. Ben-Ze’ev, A., Duerr, A., Solomon, F. & Penman, S. 1979. The outer boundary of the cytoskeleton: a lamina derived from plasma mem- brane proteins. Cell, 17: 859-865.
4. Camargo, E. 1964. Growth and differentiation in Trypanosoma cruzi I. Origin of metacyclic trypanosomes in liquid media. Rev. Znst. Med. Trop. Siio Paulo, 6: 93-100.
5. De Souza, W., Meza, I., Martinez-Palomo, A., Souto-Padron, T. & Meirelles, M. N. L. 1983. Trypanosoma cruzi: distribution of flu- orescently labeled tubulin and actin in epimastigotes. J. Parasitol., 69:
6. De Souza, W. & Benchimol, M. 1984. High voltage electron microscopy of critical-point dried trypanosomatids. J. Submicrosc. Cy-
7. Faulstich, H., Trischmann, H. & Mayer, D. 1983. Preparation oftetramethyl rhodaminyl-phalloidin and uptake of the toxin into short- term cultured hepatocytes by endocytosis. Exp. Cell Rex, 144: 63-72.
8. Hirono, M., Endoh, H., Okada, N., Numata, 0. & Watanabe, Y. 1987. Tetrahymena actin. Cloning and sequencing of the Tetrahymena actin gene and identification of its gene product. J. Mol. Biol., 194: 18 1- 192.
9. KiImartin, J. V., Wright, B. &Milstein,C. 1982. Rat monoclonal antitubulin antibodies derived by using a new nonsecreting rat cell line. J. Cell Biol., 93: 576-582.
10. Korn, E. D. 1978. Biochemistry of actomyosin-dependent cell motility (a review). Proc. Natl. Acad. Sci. USA, 75: 588-599.
11. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (Lond.), 227: 680- 685.
138-142.
tol., 16: 237-242.
12. Linder, J. C. & Staehlin, L. A. 1977. Plasma membrane spe- cializations in a trypanosomatid flagellate. J. Ultrastruc. Res., 60: 246- 262.
13. Matsudaira, P. T. & Burgess, D. R. 1978. SDS-microslab linear gradient polyacrylamide gel electrophoresis. Anal. Biochem., 87: 386- 396.
14. Meza, I., Sabanero, M., Cazares, F. & Bryan, J. 1983. Isolation and characterization of actin from Entamoeba histolytica. J. Biol. Chem.,
15. Mitchell, E. J. & Zimmerman, A. M. 1985. Biochemical evi- dence for the presence of an actin protein in Tetrahymena pyriformis. J. Cell Sci., 73: 279-297.
16. Nellen, W. & Gallwitz, D. 1982. Actin genes and actin mes- senger in Acanthamoeba castellanii. Nucleotide sequence of the split actin gene I. J. Mol. Biol., 159: 1-18.
17. Pardee, J. & Spudich, J. D. 1982. Purification of muscle actin. Methods Enzymol., 85: 164-1 8 1.
18. Roitman, C., Roitman, I. & Azevedo, H. P. 1972. Growth of an insect trypanosomatid at 37°C in a defined medium. J. Protozool.,
19. Seebeck, T., Schneider, A., Kueng, V., Schlaeppi, K. & Hemphill, A. 1988. The cytoskeleton of Trypanosoma brucei-the beauty of simplicity. Protoplasma, 145: 188-1 94.
20. Souto-Padron, T., De Souza, W. & Heuser, J. E. 1984. Quick- freeze deep-etch rotary replication of Trypanosoma cruzi and Herpe- tomonas megaseliae. J . Cell Sci., 69: 167-1 78.
21. Steiger, J., Wyler, T. & Seebeck, T. 1984. Partial purification and characterization of microtubular protein from Trypanosoma brucei. J. Biol. Chem., 259: 4596-4602.
22. Teixeira, M. M. G. & Yoshida, N. 1986. Stage-specific surface antigens of metacyclic trypomastigotes of Trypanosoma cruzi identified by monoclonal antibodies. Mol. Biochem. Parasitol., 18: 27 1-282.
23. Towbin, H., Staehlin, T. & Gordon, J. 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. USA, 76: 4350- 4354.
24. Vanderkerckhove, J. & Weber, K. 1980. Vegetative Dictyoste- lium cells containing 17 actin genes express a single major actin. Nature (Lond.), 284: 475-477.
25. Vickerman, K. 1969. The fine structure of Trypanosoma con- golense in its bloodstream phase. J. Protozool., 16: 54-69.
26. Vickerman, K. & Preston, T. M. 1976. Comparative cell biology of kinetoplastid flagellates. In: Lumsden, W. H. R. & Evans, D. A,, ed., Biology of the Kinetoplastida. Academic Press, London, pp. 35-1 30.
27. Weeds, A. G. 1982. Actin-binding proteins: regulators of cell architecture and mobility. Nature (Lond.), 296: 81 1-8 16.
28. Wieland, T. 1977. Modification of actins by phallotoxins. Na- turwissenschaften, 64: 303-309.
29. Wray, W., Boulikas, T., Wray, U. P. & Hancock, R. 198 1. Silver staining of proteins in polyacrylamide gels. Anal. Biochem.. 118: 197- 203.
30. Yoshida, N. 1983. Surface antigens of metacyclic trypomasti- gotes of Trypanosoma cruzi. Infect. Immun., 40: 836-839.
3 1. Zechel, K. 1980. Isolation of polymerization-competent cyto- plasmic actin by affinity chromatography on immobilized DNase I using formamide as eluant. Eur. J. Biochem., 110: 343-348.
258: 3936-3941.
19: 346-349.
Received 10-29-87; accepted 6-30-88




















