
7+( %/,1' %221( 5$*7,0( )(67,9$/ - West Coast Ragtime Society
Upload
khangminh22Category
view
1download
0
MiR-221 Influences Effector Functions and ActinCytoskeleton in Mast CellsRamon J. Mayoral1., Lorenzo Deho1., Nicole Rusca1., Nenad Bartonicek2, Harpreet Kaur Saini2, Anton J.
Enright2, Silvia Monticelli1*
1 Institute for Research in Biomedicine, Bellinzona, Switzerland, 2 EMBL - European Bioinformatics Institute, Wellcome Trust Genome Campus, Hinxton, Cambridge, United
Kingdom
Abstract
Mast cells have essential effector and immunoregulatory functions in IgE-associated allergic disorders and certain innateand adaptive immune responses, but the role of miRNAs in regulating mast cell functions is almost completely unexplored.To examine the role of the activation-induced miRNA miR-221 in mouse mast cells, we developed robust lentiviral systemsfor miRNA overexpression and depletion. While miR-221 favored mast cell adhesion and migration towards SCF or antigenin trans-well migration assays, as well as cytokine production and degranulation in response to IgE-antigen complexes,neither miR-221 overexpression, nor its ablation, interfered with mast cell differentiation. Transcriptional profiling of miR-221-overexpressing mast cells revealed modulation of many transcripts, including several associated with the cytoskeleton;indeed, miR-221 overexpression was associated with reproducible increases in cortical actin in mast cells, and with alteredcellular shape and cell cycle in murine fibroblasts. Our bioinformatics analysis showed that this effect was likely mediated bythe composite effect of miR-221 on many primary and secondary targets in resting cells. Indeed, miR-221-induced cellularalterations could not be recapitulated by knockdown of one of the major targets of miR-221. We propose a model in whichmiR-221 has two different roles in mast cells: in resting cells, basal levels of miR-221 contribute to the regulation of the cellcycle and cytoskeleton, a general mechanism probably common to other miR-221-expressing cell types, such as fibroblasts.Vice versa, upon induction in response to mast cell stimulation, miR-221 effects are mast cell-specific and activation-dependent, contributing to the regulation of degranulation, cytokine production and cell adherence. Our studies providenew insights into the roles of miR-221 in mast cell biology, and identify novel mechanisms that may contribute to mast cell-related pathological conditions, such as asthma, allergy and mastocytosis.
Citation: Mayoral RJ, Deho L, Rusca N, Bartonicek N, Saini HK, et al. (2011) MiR-221 Influences Effector Functions and Actin Cytoskeleton in Mast Cells. PLoSONE 6(10): e26133. doi:10.1371/journal.pone.0026133
Editor: Jose Alberola-Ila, Oklahoma Medical Research Foundation, United States of America
Received May 10, 2011; Accepted September 20, 2011; Published October 12, 2011
Copyright: � 2011 Mayoral et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: RJM was the recipient of a pre-doctoral fellowship from the San Raffaele University (Italy). This work was supported by the Swiss National ScienceFoundation grant 31003A_121991 to SM and by a contribution from the Ceresio Foundation (Switzerland) to SM. The funders had no role in study design, datacollection and analysis, decision to publish, or preparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist.
* E-mail: [email protected]
. These authors contributed equally to this work.
Introduction
Mast cells are cells of the innate immune system that reside in
most tissues and are characterized by cytoplasmic granules
containing active mediators such as histamine and proteases,
which can be rapidly released upon activation. A broad panel of
cytokines and chemokines is also rapidly synthesized upon acute
stimulation, although some cytokines (such as TNFa) can be pre-
stored in the granules [1,2]. On the surface, mast cells are
characterized by the expression of KIT and FceRI (high-affinity
IgE receptor). KIT, the receptor for the stem cell factor (SCF), is a
critical regulator of mast cell differentiation, proliferation,
activation and survival. Although mast cells have been considered
for a long time detrimental to the host, mainly because of their
major role as master effector cells in allergic responses, it is now
becoming clear that depending on the context, mast cells can
either positively or negatively regulate innate or adaptive immune
responses to pathogens or allergens (for a recent review see [3]).
MicroRNAs (miRNAs) are small, non-coding RNAs that are
emerging as major players in the regulation of endogenous gene
expression in immune homeostasis [4]. Specifically, miRNAs
regulate the expression of target genes by interacting with sites in
the 39 untranslated region (39UTR) of their target mRNAs.
MiRNAs regulate all aspects of a cell, including differentiation,
function, proliferation, survival, metabolism, and responses to
environmental changes. A single miRNA can potentially modulate
the expression of hundreds of transcripts in a cell, both by direct
and indirect effects, and as many as 90% of human genes may be
regulated by miRNAs [5]. MiRNAs are implicated in various
pathologic conditions as well as in tumorigenesis [4,6], and within
the immune system, deletion or overexpression of specific miRNAs
can impair innate or adaptive immune responses [7].
While the importance of miRNAs in various types of normal
and diseased cellular processes is by now well established, very
little is known about the role of miRNAs in mast cell development,
function and disease. Our lab identified miR-221/-222 as a family
of miRNAs that is transcriptionally induced upon mast cell
activation, and we showed that expression of miR-221 and/or
miR-222 to levels similar to the endogenous of activated mast cells,
led to reduced mast cell proliferation [8]. MiR-221 and miR-222
PLoS ONE | www.plosone.org 1 October 2011 | Volume 6 | Issue 10 | e26133

derive from the same primary transcript and share the same seed
sequence, implying that they should recognize the same targets
[9]. Following up on our previous studies, we investigated the role
of miR-221 in mast cell differentiation and function. Specifically,
we found that although miR-221 does not seem to affect mast cell
differentiation, it has important roles in regulating multiple
processes in differentiated mast cells, such as degranulation,
adhesion and migration, some of which may be linked to a
dysregulation in the actin cytoskeleton. Indeed, we found that
alteration of miR-221 expression in mast cells and fibroblasts led
not only to a reduction in cell proliferation similar to what we
previously described using a different expression system, but also
to an alteration of actin content and overall cellular shape in both
cell types. Transcriptional profiling and bioinformatics analysis
using the Sylamer algorithm [10], indicated that miR-221 effects
in mast cells were mediated by the alterations of the level of
expression of many primary and secondary targets. Importantly,
such miR-221-mediated alterations of the cell phenotype could not
be recapitulated by knockdown of one of the most prominent
target for miR-221, suggesting that the observed effect of miR-221
in resting mast cells and fibroblasts is likely to be composite, due to
the alteration of many genes. Moreover, we also observed mast
cell-specific, activation-dependent effects of miR-221. Since miR-
221 is expressed at basal level in mast cells, but it is also inducible
upon stimulation, we propose a model in which miR-221 has a
dual roles in these cells: at resting state, it contributes to the
regulation of the cell cycle and cytoskeleton, a housekeeping effect
that can be observed also in different cell types expressing this
miRNA. However, in response to stimulation through IgE-antigen
complexes, miR-221 effects are mast cell-specific and activation-
dependent, contributing to the regulation of degranulation,
cytokine production and cell adherence. Overall, our studies
provide insights on the role of miRNAs in mast cells, and lay the
groundwork for understanding some of the mechanisms underly-
ing pathological conditions caused by mast cells, such as allergy
and mastocytosis.
Materials and Methods
Ethics statementAll animal studies were performed in accordance with the Swiss
Federal Veterinary Office guidelines and were approved by the
Dipartimento della Sanita’ e della Socialita’, authorization number
18/2010.
PlasmidsAbout 400bp of the mouse miR-221 or miR-222 genomic
sequences were cloned into the pAPM lentiviral vector [11]. Point
mutations in the seed sequence of miR-221 were introduced using
the Quick Change Site-Directed Mutagenesis kit (Stratagene) and
the following primers (mutations underlined): miR-221mFW: 59-
GTTTGTTAGGCAACATCGCGATTGTCTGCTGGGTTTC-
AGG; miR-221mRV: 59- CCTGAAACCCAGCAGACAATCGC-
GATGTTGCCTAACAAAC. The control vector pAPM-shLuc,
expressing a miR-30-based shRNA against luciferase, was obtained
from Thomas Pertel and Jeremy Luban and contained the following
sequence: 59- CACAAACGCTCTCATCGACAAG. The miRNA
target (miRT) vectors containing four sequences fully complemen-
tary to miR-221 and/or miR-222, were provided by Bernhard
Gentner and Luigi Naldini [12].
Cell culturesFor bone marrow-derived mast cells (BMMCs) differentiation,
bone marrow cells from C57Bl/6 mice (6-8 weeks old) were
differentiated and maintained in IMDM with 10% FBS, 2 mM L-
glutamine, 0.1 mM non essential amino acids, 50 mMb-mercap-
toethanol, antibiotics and 50% WEHI-3 conditioned supernatant
as a source of IL-3 [13,14]. The IL-3-dependent mast cell line
MC/9 (ATCC) was cultured as for primary BMMCs. 3T3 cells
were cultured in DMEM supplemented with 10% FBS, 2 mM L-
glutamine, 0.1 mM non-essential amino acids, 50 mMb-mercap-
toethanol and antibiotics.
Transductions and transfectionsLentiviral transductions of mast cells were performed exactly as
described [8,13]. 3T3 cells were transduced with concentrated
lentiviruses prepared exactly as described [8,13] and selected with
2 mg/mL puromycin for several days before performing experi-
ments. For transient transfections with oligonucleotides, 3T3 cells
were transfected with Lipofectamine-2000 (Invitrogen) and 20pmol
of siRNAs against p27Kip1 (siGENOME Cdkn1b) or control siRNAs
(siGENOME non-targeting siRNA #2 and/or siGLO) (all from
Thermo Scientific) following manufacturer’s instructions. Cells were
analyzed 48–72 h after transfection. Efficiency of transfection was
assessed in each experiment by transfecting a fluorescent oligonu-
cleotide (siGLO) and analyzing the percentage of siGLO+ cells by
FACS, which usually ranged between 50 and 70%. The efficiency of
protein knockdown was assessed by Western blot.
RNA extraction and RT-PCRTotal RNA was extracted using TRIzol (Invitrogen) according
to manufacturer’s instructions. To analyze miRNA expression,
qRT-PCR was performed using a miRNA reverse transcription kit
and TaqMan miRNA assays from Applied Biosystems, following
exactly manufacturer’s instructions.
Immunostainings and microscopyAnti-KIT-APC, anti-FceRIa-PE and anti-CD25-PE were
purchased from eBioscience. For microscopy, 105 BMMCs were
labeled with CFSE (Invitrogen) and added to a monolayer of 3T3
cells on coverslips. After 12 hours, coverslips were washed to
eliminate non-adherent cells, and cells were fixed with 3.7% p-
formaldehyde, permeabilized with 0.1% Triton-X100 and stained
with 160nM phalloidin-AlexaFluor-594 (Invitrogen). Glass slides
were mounted using Gelvatol (20% polyvinyl alcohol, 100mM
Tris-HCl pH 8.5, 2.5% DABCO). The same protocol was used to
stain 3T3 cells grown directly on coverslips. Bright field images
were captured with a Nikon Eclipse E800 and analyzed with the
Openlab software (Improvision). For FACS-based quantification
of F-Actin, 3?105 cells were fixed 15–30 min in 4–5% p-
formaldehyde, permeabilized 2min with 0.1% Triton-X100, and
stained with phalloidin at a 1:100 dilution.
Degranulation, adhesion and migration assaysFor degranulation assay, BMMCs (5?104) were resuspended in
50 mL OptiMEM, 1% FBS and stimulated for 1 h with 1.5 mg/mL
IgE-anti-DNP (clone SPE-7, Sigma) and 0.2 mg/mL DNP-HSA
(Sigma). After stimulation, cell pellets were lysed in 50 mL of 0.5%
Triton-X100 in OptiMEM, 1% FBS, and 50 mL of a 3.8 mM
solution of the b-hexosaminidase substrate 4-nitrophenyl N-acetyl-
b-D-glucosaminide (Sigma) were added to both cell lysates and
supernatants. After incubation for 2 h at 37uC, the reaction was
stopped with 90 mL glycine 0.2 M, pH 10.7, and absorbance was
read at 405 nm. The percentage of degranulation was calculated as
the ratio between the absorbance of supernatants and the total
absorbance of supernatants and cell lysates [15]. Alternatively,
degranulation was assessed using the same annexin V-PE kit used to
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 2 October 2011 | Volume 6 | Issue 10 | e26133

detect apoptosis (BD-Pharmingen) [16,17]. Briefly, cells were
stimulated for 30 min with 1.5 mg/mL IgE-anti-DNP and 0.2 mg/
mL DNP-HSA, in the presence or absence of 20 ng/mL SCF
(Peprotech). Cells were then washed and stained with annexin V-PE
following manufacturer’s instructions. For adhesion experiments,
105 BMMCs were added to a monolayer of 3T3 cells in 24-well
plates. Floating cells were harvested 8h later, and adherent cells
were detached using 10mM EDTA. In order to distinguish BMMCs
from fibroblasts, cells were stained for KIT, and numbers of KIT+cells were evaluated by FACS. The percentage of adherent cells was
calculated as the ratio between adherent cells versus total. Cell
migration was assayed using 24-well transwell chambers (Corning)
with 8.0 mm pores in polycarbonate membranes. 2?105 BMMCs
were seeded in the upper chambers and allowed to migrate for 2 h.
In some experiments, 20 ng/mL of SCF was used as chemoat-
tractant, while in others BMMCs were first sensitized with 300 ng/
mL IgE-anti-DNP for 12 h and then allowed to migrate towards
0.2 mg/mL DNP-HSA. The percentage of migrated cells was
calculated as the ratio between the number of cells in the lower
chamber versus total.
Cell-cycle analysis and Western blotsFor propidium iodide staining and DNA content analysis, non-
confluent 3T3 cells were fixed in 70% ethanol for 45 min on ice,
followed by incubation for 30 min at 37uC with 100 mg/mL
RNaseA and 40 mg/ml propidium iodide. Cells were analyzed by
FACS immediately afterwards. Total protein extracts for Western
blot were prepared by cell lysis in Laemmli sample buffer. Samples
were run on 12% SDS-polyacrylamide gels and immunodetection
was performed with p27 C-19 and b-tubulin H-235 antibodies
(Santa Cruz Biotechnologies).
Microarrays and Sylamer analysisGene arrays were performed at Miltenyi Biotec using Agilent
dual-color whole-genome oligo arrays and total RNA from three
independent biological replicas. Genes found to be commonly
regulated were functionally annotated. Sylamer analysis was
performed through a Sylarray web server [18,19]. Specifically, the
Agilent probes were mapped to the 39zUTRs of genes stored in the
Sylarray database. The sequences were previously masked from low
complexity regions and redundant UTR sequences with DUST
algorithm and RSAT purge-sequence interface to Vmatch [20,21].
As a control we also analyzed the mutant seed sequence to check for
any evidence of its influence in the experiment and no significant
association was found. Array data are MIAME compliant and are
deposited in the MIAME compliant database Gene Expression
Omnibus (GEO) with accession number GSE24462.
Statistical analysisResults are expressed as mean 6 standard deviation. Compar-
isons were made using the Student’s unpaired t-test and the
GraphPad Prism Software.
Supporting InformationAdditional information can be found in the Supplementary
Methods S1.
Results
A robust lentiviral system for manipulating microRNAexpression in mast cells
As we previously described for acute stimulation of mast cells
[8], BMMCs stimulated with IgE-antigen complexes upregulated
miR-221 expression (Figure 1A). While stimulation-dependent
upregulation of this miRNA could be favored by SCF co-
stimulation, SCF alone had no effect on miR-221 expression (not
shown). To investigate the role of miR-221 in regulating primary
mast cell functions, we developed a lentiviral system to manipulate
miRNA expression in primary BMMC and used it to alter miR-
221 expression. The pAPM/pAGM vectors were used to
overexpress miR-221 or miR-222; as control, we used a mutant
version of miR-221 (miR-221m), containing mutations in the seed
region to abrogate target recognition, as well as a vector expressing
an irrelevant hairpin (shLuc) (Figure 1B). The miR-221m mature
sequence had no predicted targets as assessed by TargetScan [9].
The ‘miRNA target’ (miRT) vectors contain four miRNA binding
sites (miR-bs) cloned downstream a GFP reporter gene, and they
were used to functionally ablate miR-221/-222 [12]. Transcrip-
tion from such vectors results in accumulation of decoy mRNAs
that divert miRNAs from their physiological targets [22].
To assess expression from these vectors, BMMCs were
transduced with the indicated vectors, and miRNA expression
was assessed by qRT-PCR (Figures 1C and 1D). Compared to
untransduced, unstimulated cells (expression set to 1 in Figures 1C
and 1D), transduction of primary mast cells with pAGM/pAPM-
miR-221 increased miR-221 expression by ,60-fold, whereas
transduction with miRT-221 decreased expression by ,10-fold
(Figure 1D). Transduction with the mutant miR-221m had no
effect (Figure 1C). Initial experiments were performed using a
vector (Tween) that induced only modest overexpression (,4-fold),
similar to the levels of endogenous miR-221 observed upon cell
stimulation (Figure 1A) [8]. However, both types of vectors
(weaker and stronger expression) gave similar results qualitatively,
although the stronger vector provided bigger quantitative
differences, and was therefore used in most of the subsequent
experiments. To assess the functional effects of miRNA overex-
pression/ablation, the mast cell line MC/9 was transduced to
overexpress miR-221 or the mutant miR-221m. Transduced cells
were selected with puromycin, subjected to a second round of
transduction with the miRT vectors, and monitored for GFP
expression (Figure 1E). As a result of binding of the overexpressed
miRNAs to their cognate sites in the 39 UTR of the GFP reporter
mRNA expressed from the miRT, GFP expression was strongly
reduced specifically in cells expressing miR-221 but not the
mutant miR-221m. We therefore used both validated systems
(overexpression and ablation) to study mast cell differentiation in
the presence or absence of miR-221.
MiR-221/-222 as well as the transcriptional repressor PLZF are
both known important regulators of hematopoietic cell differen-
tiation [23,24,25]. We previously showed that binding sites for
PLZF were enriched in mast cell-specific DNaseI hypersensitive
sites found upstream of the miR-221-222 genomic sequence [8].
To address the possible relation between PLZF and miR-221, we
analyzed expression of both Plzf mRNA and miR-221 during mast
cell differentiation (Supplementary Figure S1). We observed an
inverse relation between Plzf and miR-221 expression during mast
cell differentiation, and ectopic expression of PLZF in mast cells
diminished miR-221 expression in response to acute stimulation,
suggesting that PLZF is able to repress miR-221-222 induction
either directly or indirectly, and possibly through PLZF-binding
regulatory elements in the miR-221-222 locus [8]. However,
ectopic expression of PLZF in differentiated mast cells had no
effect on the basal levels of endogenous miR-221, indicating that
other factors regulate basal expression of this miRNA in mast cells.
To assess whether miR-221/-222 may have a direct role in
regulating the differentiation process in mast cells, we transduced
bone marrow-derived hematopoietic progenitors with lentiviruses
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 3 October 2011 | Volume 6 | Issue 10 | e26133
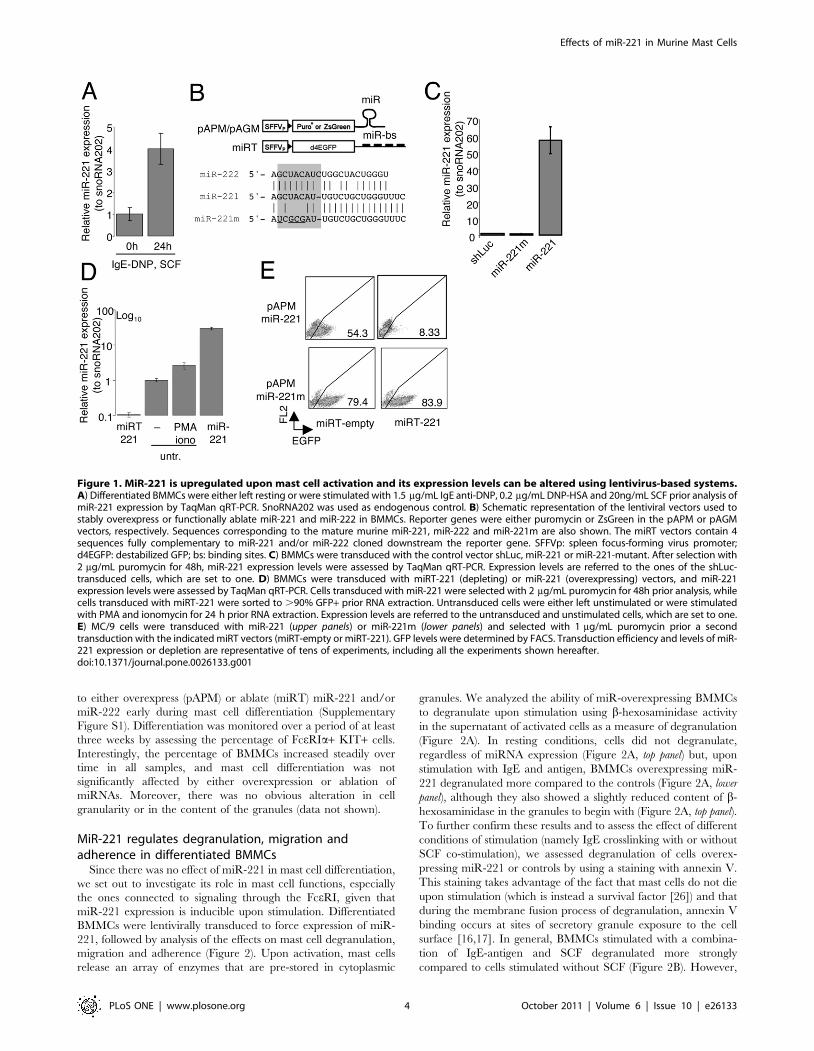
to either overexpress (pAPM) or ablate (miRT) miR-221 and/or
miR-222 early during mast cell differentiation (Supplementary
Figure S1). Differentiation was monitored over a period of at least
three weeks by assessing the percentage of FceRIa+ KIT+ cells.
Interestingly, the percentage of BMMCs increased steadily over
time in all samples, and mast cell differentiation was not
significantly affected by either overexpression or ablation of
miRNAs. Moreover, there was no obvious alteration in cell
granularity or in the content of the granules (data not shown).
MiR-221 regulates degranulation, migration andadherence in differentiated BMMCs
Since there was no effect of miR-221 in mast cell differentiation,
we set out to investigate its role in mast cell functions, especially
the ones connected to signaling through the FceRI, given that
miR-221 expression is inducible upon stimulation. Differentiated
BMMCs were lentivirally transduced to force expression of miR-
221, followed by analysis of the effects on mast cell degranulation,
migration and adherence (Figure 2). Upon activation, mast cells
release an array of enzymes that are pre-stored in cytoplasmic
granules. We analyzed the ability of miR-overexpressing BMMCs
to degranulate upon stimulation using b-hexosaminidase activity
in the supernatant of activated cells as a measure of degranulation
(Figure 2A). In resting conditions, cells did not degranulate,
regardless of miRNA expression (Figure 2A, top panel) but, upon
stimulation with IgE and antigen, BMMCs overexpressing miR-
221 degranulated more compared to the controls (Figure 2A, lower
panel), although they also showed a slightly reduced content of b-
hexosaminidase in the granules to begin with (Figure 2A, top panel).
To further confirm these results and to assess the effect of different
conditions of stimulation (namely IgE crosslinking with or without
SCF co-stimulation), we assessed degranulation of cells overex-
pressing miR-221 or controls by using a staining with annexin V.
This staining takes advantage of the fact that mast cells do not die
upon stimulation (which is instead a survival factor [26]) and that
during the membrane fusion process of degranulation, annexin V
binding occurs at sites of secretory granule exposure to the cell
surface [16,17]. In general, BMMCs stimulated with a combina-
tion of IgE-antigen and SCF degranulated more strongly
compared to cells stimulated without SCF (Figure 2B). However,
Figure 1. MiR-221 is upregulated upon mast cell activation and its expression levels can be altered using lentivirus-based systems.A) Differentiated BMMCs were either left resting or were stimulated with 1.5 mg/mL IgE anti-DNP, 0.2 mg/mL DNP-HSA and 20ng/mL SCF prior analysis ofmiR-221 expression by TaqMan qRT-PCR. SnoRNA202 was used as endogenous control. B) Schematic representation of the lentiviral vectors used tostably overexpress or functionally ablate miR-221 and miR-222 in BMMCs. Reporter genes were either puromycin or ZsGreen in the pAPM or pAGMvectors, respectively. Sequences corresponding to the mature murine miR-221, miR-222 and miR-221m are also shown. The miRT vectors contain 4sequences fully complementary to miR-221 and/or miR-222 cloned downstream the reporter gene. SFFVp: spleen focus-forming virus promoter;d4EGFP: destabilized GFP; bs: binding sites. C) BMMCs were transduced with the control vector shLuc, miR-221 or miR-221-mutant. After selection with2 mg/mL puromycin for 48h, miR-221 expression levels were assessed by TaqMan qRT-PCR. Expression levels are referred to the ones of the shLuc-transduced cells, which are set to one. D) BMMCs were transduced with miRT-221 (depleting) or miR-221 (overexpressing) vectors, and miR-221expression levels were assessed by TaqMan qRT-PCR. Cells transduced with miR-221 were selected with 2 mg/mL puromycin for 48h prior analysis, whilecells transduced with miRT-221 were sorted to .90% GFP+ prior RNA extraction. Untransduced cells were either left unstimulated or were stimulatedwith PMA and ionomycin for 24 h prior RNA extraction. Expression levels are referred to the untransduced and unstimulated cells, which are set to one.E) MC/9 cells were transduced with miR-221 (upper panels) or miR-221m (lower panels) and selected with 1 mg/mL puromycin prior a secondtransduction with the indicated miRT vectors (miRT-empty or miRT-221). GFP levels were determined by FACS. Transduction efficiency and levels of miR-221 expression or depletion are representative of tens of experiments, including all the experiments shown hereafter.doi:10.1371/journal.pone.0026133.g001
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 4 October 2011 | Volume 6 | Issue 10 | e26133
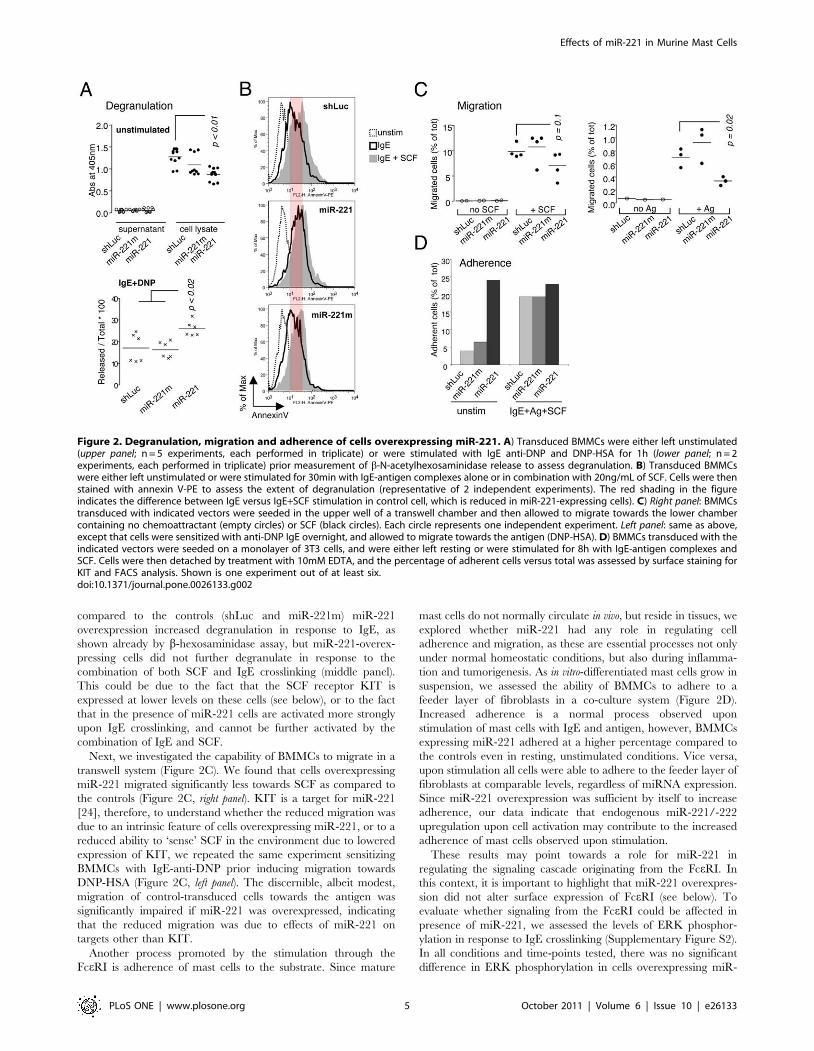
compared to the controls (shLuc and miR-221m) miR-221
overexpression increased degranulation in response to IgE, as
shown already by b-hexosaminidase assay, but miR-221-overex-
pressing cells did not further degranulate in response to the
combination of both SCF and IgE crosslinking (middle panel).
This could be due to the fact that the SCF receptor KIT is
expressed at lower levels on these cells (see below), or to the fact
that in the presence of miR-221 cells are activated more strongly
upon IgE crosslinking, and cannot be further activated by the
combination of IgE and SCF.
Next, we investigated the capability of BMMCs to migrate in a
transwell system (Figure 2C). We found that cells overexpressing
miR-221 migrated significantly less towards SCF as compared to
the controls (Figure 2C, right panel). KIT is a target for miR-221
[24], therefore, to understand whether the reduced migration was
due to an intrinsic feature of cells overexpressing miR-221, or to a
reduced ability to ‘sense’ SCF in the environment due to lowered
expression of KIT, we repeated the same experiment sensitizing
BMMCs with IgE-anti-DNP prior inducing migration towards
DNP-HSA (Figure 2C, left panel). The discernible, albeit modest,
migration of control-transduced cells towards the antigen was
significantly impaired if miR-221 was overexpressed, indicating
that the reduced migration was due to effects of miR-221 on
targets other than KIT.
Another process promoted by the stimulation through the
FceRI is adherence of mast cells to the substrate. Since mature
mast cells do not normally circulate in vivo, but reside in tissues, we
explored whether miR-221 had any role in regulating cell
adherence and migration, as these are essential processes not only
under normal homeostatic conditions, but also during inflamma-
tion and tumorigenesis. As in vitro-differentiated mast cells grow in
suspension, we assessed the ability of BMMCs to adhere to a
feeder layer of fibroblasts in a co-culture system (Figure 2D).
Increased adherence is a normal process observed upon
stimulation of mast cells with IgE and antigen, however, BMMCs
expressing miR-221 adhered at a higher percentage compared to
the controls even in resting, unstimulated conditions. Vice versa,
upon stimulation all cells were able to adhere to the feeder layer of
fibroblasts at comparable levels, regardless of miRNA expression.
Since miR-221 overexpression was sufficient by itself to increase
adherence, our data indicate that endogenous miR-221/-222
upregulation upon cell activation may contribute to the increased
adherence of mast cells observed upon stimulation.
These results may point towards a role for miR-221 in
regulating the signaling cascade originating from the FceRI. In
this context, it is important to highlight that miR-221 overexpres-
sion did not alter surface expression of FceRI (see below). To
evaluate whether signaling from the FceRI could be affected in
presence of miR-221, we assessed the levels of ERK phosphor-
ylation in response to IgE crosslinking (Supplementary Figure S2).
In all conditions and time-points tested, there was no significant
difference in ERK phosphorylation in cells overexpressing miR-
Figure 2. Degranulation, migration and adherence of cells overexpressing miR-221. A) Transduced BMMCs were either left unstimulated(upper panel; n = 5 experiments, each performed in triplicate) or were stimulated with IgE anti-DNP and DNP-HSA for 1h (lower panel; n = 2experiments, each performed in triplicate) prior measurement of b-N-acetylhexosaminidase release to assess degranulation. B) Transduced BMMCswere either left unstimulated or were stimulated for 30min with IgE-antigen complexes alone or in combination with 20ng/mL of SCF. Cells were thenstained with annexin V-PE to assess the extent of degranulation (representative of 2 independent experiments). The red shading in the figureindicates the difference between IgE versus IgE+SCF stimulation in control cell, which is reduced in miR-221-expressing cells). C) Right panel: BMMCstransduced with indicated vectors were seeded in the upper well of a transwell chamber and then allowed to migrate towards the lower chambercontaining no chemoattractant (empty circles) or SCF (black circles). Each circle represents one independent experiment. Left panel: same as above,except that cells were sensitized with anti-DNP IgE overnight, and allowed to migrate towards the antigen (DNP-HSA). D) BMMCs transduced with theindicated vectors were seeded on a monolayer of 3T3 cells, and were either left resting or were stimulated for 8h with IgE-antigen complexes andSCF. Cells were then detached by treatment with 10mM EDTA, and the percentage of adherent cells versus total was assessed by surface staining forKIT and FACS analysis. Shown is one experiment out of at least six.doi:10.1371/journal.pone.0026133.g002
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 5 October 2011 | Volume 6 | Issue 10 | e26133
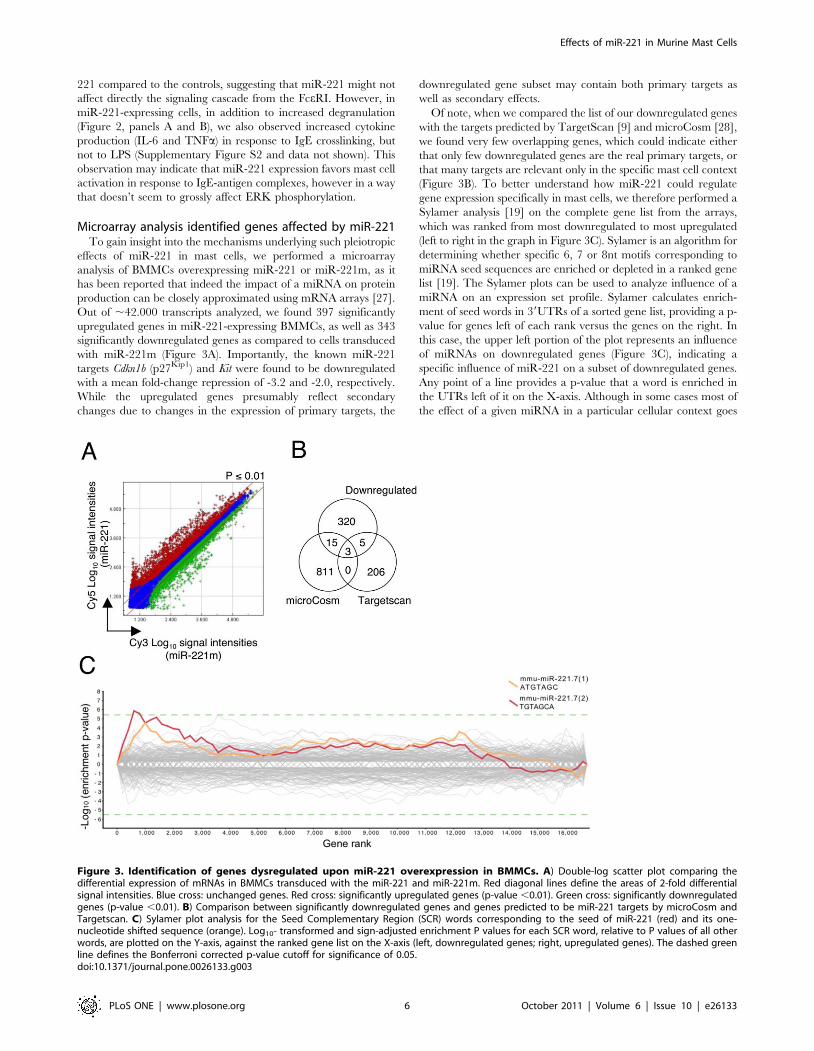
221 compared to the controls, suggesting that miR-221 might not
affect directly the signaling cascade from the FceRI. However, in
miR-221-expressing cells, in addition to increased degranulation
(Figure 2, panels A and B), we also observed increased cytokine
production (IL-6 and TNFa) in response to IgE crosslinking, but
not to LPS (Supplementary Figure S2 and data not shown). This
observation may indicate that miR-221 expression favors mast cell
activation in response to IgE-antigen complexes, however in a way
that doesn’t seem to grossly affect ERK phosphorylation.
Microarray analysis identified genes affected by miR-221To gain insight into the mechanisms underlying such pleiotropic
effects of miR-221 in mast cells, we performed a microarray
analysis of BMMCs overexpressing miR-221 or miR-221m, as it
has been reported that indeed the impact of a miRNA on protein
production can be closely approximated using mRNA arrays [27].
Out of ,42.000 transcripts analyzed, we found 397 significantly
upregulated genes in miR-221-expressing BMMCs, as well as 343
significantly downregulated genes as compared to cells transduced
with miR-221m (Figure 3A). Importantly, the known miR-221
targets Cdkn1b (p27Kip1) and Kit were found to be downregulated
with a mean fold-change repression of -3.2 and -2.0, respectively.
While the upregulated genes presumably reflect secondary
changes due to changes in the expression of primary targets, the
downregulated gene subset may contain both primary targets as
well as secondary effects.
Of note, when we compared the list of our downregulated genes
with the targets predicted by TargetScan [9] and microCosm [28],
we found very few overlapping genes, which could indicate either
that only few downregulated genes are the real primary targets, or
that many targets are relevant only in the specific mast cell context
(Figure 3B). To better understand how miR-221 could regulate
gene expression specifically in mast cells, we therefore performed a
Sylamer analysis [19] on the complete gene list from the arrays,
which was ranked from most downregulated to most upregulated
(left to right in the graph in Figure 3C). Sylamer is an algorithm for
determining whether specific 6, 7 or 8nt motifs corresponding to
miRNA seed sequences are enriched or depleted in a ranked gene
list [19]. The Sylamer plots can be used to analyze influence of a
miRNA on an expression set profile. Sylamer calculates enrich-
ment of seed words in 39UTRs of a sorted gene list, providing a p-
value for genes left of each rank versus the genes on the right. In
this case, the upper left portion of the plot represents an influence
of miRNAs on downregulated genes (Figure 3C), indicating a
specific influence of miR-221 on a subset of downregulated genes.
Any point of a line provides a p-value that a word is enriched in
the UTRs left of it on the X-axis. Although in some cases most of
the effect of a given miRNA in a particular cellular context goes
Figure 3. Identification of genes dysregulated upon miR-221 overexpression in BMMCs. A) Double-log scatter plot comparing thedifferential expression of mRNAs in BMMCs transduced with the miR-221 and miR-221m. Red diagonal lines define the areas of 2-fold differentialsignal intensities. Blue cross: unchanged genes. Red cross: significantly upregulated genes (p-value ,0.01). Green cross: significantly downregulatedgenes (p-value ,0.01). B) Comparison between significantly downregulated genes and genes predicted to be miR-221 targets by microCosm andTargetscan. C) Sylamer plot analysis for the Seed Complementary Region (SCR) words corresponding to the seed of miR-221 (red) and its one-nucleotide shifted sequence (orange). Log10- transformed and sign-adjusted enrichment P values for each SCR word, relative to P values of all otherwords, are plotted on the Y-axis, against the ranked gene list on the X-axis (left, downregulated genes; right, upregulated genes). The dashed greenline defines the Bonferroni corrected p-value cutoff for significance of 0.05.doi:10.1371/journal.pone.0026133.g003
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 6 October 2011 | Volume 6 | Issue 10 | e26133

primarily through one or few molecular targets, in other cases this
does not provide the complete picture of miRNA effect on the
transcriptome and proteome of a cell [27,29,30]. Our array
analysis showed that the transcriptomes of miR-221-overexpress-
ing cells differed by the expression of 740 genes compared to the
controls, and the Sylamer analysis indicated that many of the
downregulated genes in mast cells were likely to be directly
affected by miR-221 expression, suggesting that the effect of this
miRNA goes through the fine modulation of a multitude of targets.
Some of the targets that were up- or downregulated were also
confirmed at protein level; among others, we selected two of the
already known targets for miR-221, Kit and Cdkn1b [8,24,31], as
well as one upregulated gene (Il2Ra) that is particularly relevant in
mast cell biology as a marker for systemic mastocytosis [32,33]. As
assessed by surface staining, levels of KIT, but not of FceRI, were
significantly diminished (,2-fold) in BMMCs overexpressing miR-
221 (Figure 4A). In contrast, as we previously described [8], levels
of p27Kip1 protein were especially decreased in stimulated cells
(Figure 4B, compare lane 4 with lanes 2 and 6). While CD25
(Il2Ra) is normally not expressed by human mast cells in non-
pathogenic conditions [32,33], it is normally expressed by a subset
of in vitro-differentiated murine BMMCs (Figure 4C) as well as by a
subset of peritoneal and tissue mast cells in the mouse (Deho’ and
Monticelli, unpublished observation). The mean fold increase for
Il2Ra from the arrays was 2.1, which strongly correlated with the
increased surface expression of this marker in cells overexpressing
miR-221 (Figure 4C).
Although some of the targets for miR-221 were individually
confirmed, our arrays experiments and Sylamer analysis indicated
that miR-221 determines the downregulation of more than 200
primary targets and the subsequent secondary alterations of many
genes in the transcriptome. Therefore, to gain insights into the
function of the genes that showed altered expression in the presence
of miR-221, we performed functional grouping and gene ontology
Figure 4. MiR-221 primary and secondary targets are altered also at protein level. A) Differentiated BMMCs were either left untransducedor were transduced with the indicated vectors. After selection with puromycin, surface expression of KIT (left) and FceRI (right) was assessed by FACS.The mean fluorescence intensity (MFI) for KIT expression in miR-221- (red) and shLuc- (black) expressing cells is also indicated. B) DifferentiatedBMMCs as in A) were either left untreated or were stimulated with 20nM PMA and 1 mM ionomycin for 24h prior lysis and Western blot analysis ofp27Kip1 expression. b-tubulin was used as loading control. C) BMMCs as in A) were stained and FACS-analyzed to assess surface expression of KIT andCD25. The horizontal dotted line was added as a reference to appreciate the expected KIT dowregulation in miR-221 expressing cells.doi:10.1371/journal.pone.0026133.g004
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 7 October 2011 | Volume 6 | Issue 10 | e26133

(GO) analysis, and found that many basic biological processes were
affected (Table 1). Among the downregulated genes, the categories
that were statistically significant were surprisingly few, and
belonging for the most part to very general metabolic pathways,
such as protein folding, transcription, lipid metabolism and cell
differentiation. This finding may suggest that in resting, unstimu-
lated cells, miR-221/-222 regulate basic metabolic processes that
are common to many different cell types and/or species, and
correlates with the fact that these miRNAs belong to a family very
conserved in evolution, down to at least zebrafish [8]. Although they
are most likely due to secondary changes in the transcriptome, the
GO categories of the upregulated genes included several pathways
that we also observed altered experimentally, such as cell
proliferation, cell adhesion and cell migration. Among these latter
GO categories, we focused our attention on one of the most
statistically significant one, namely the cytoskeleton (which was also
present in the GO list of terms for the downregulated genes,
although with a p-value of 0.1), as we hypothesized that an
alteration in the ability of the cells to properly regulate the
cytoskeleton could to some extent explain why these miRNAs affect
many different biological processes, at least in unstimulated cells.
MiR-221-dependent alterations of the cytoskeleton andactin content in mast cells and 3T3 cells
To assess whether miR-221 expression could affect the
cytoskeleton, we stained transduced BMMCs with phalloidin
(Figure 5A). We used the same co-culture system with fibroblasts
used in Figure 2D to assess cell adherence, and to distinguish mast
cells from the underlying layer of feeder cells (also stained by
phalloidin), BMMCs were first labeled with CFSE. Strikingly, not
only miR-221-overexpressing cells showed increased numbers of
adherent cells (as shown also in Figure 2D), but while the actin ring
underneath the plasma membrane was barely visible in control
cells, cells overexpressing miR-221 (or miR-222, not shown)
showed the presence of a much thicker ring (Figure 5A).
Moreover, when we quantified the overall cellular amount of F-
actin in cells depleted for miR-221 (using the miRT-depleting
vectors), we observed a small but reproducible decrease in the
amount of F-actin present in these cells (Figure 5B), further
indicating that these miRNAs might be important regulators of the
actin organization in mast cells.
To independently confirm these results, and to investigate
whether the observed effect was a general feature of this miRNA
or a cell type-specific effect due to alterations of targets relevant
only in the mast cell context, we transduced 3T3 fibroblasts with
the same lentiviral vectors used on mast cells. 3T3 cells expressed
low levels of endogenous miR-221 that were increased ,20-folds
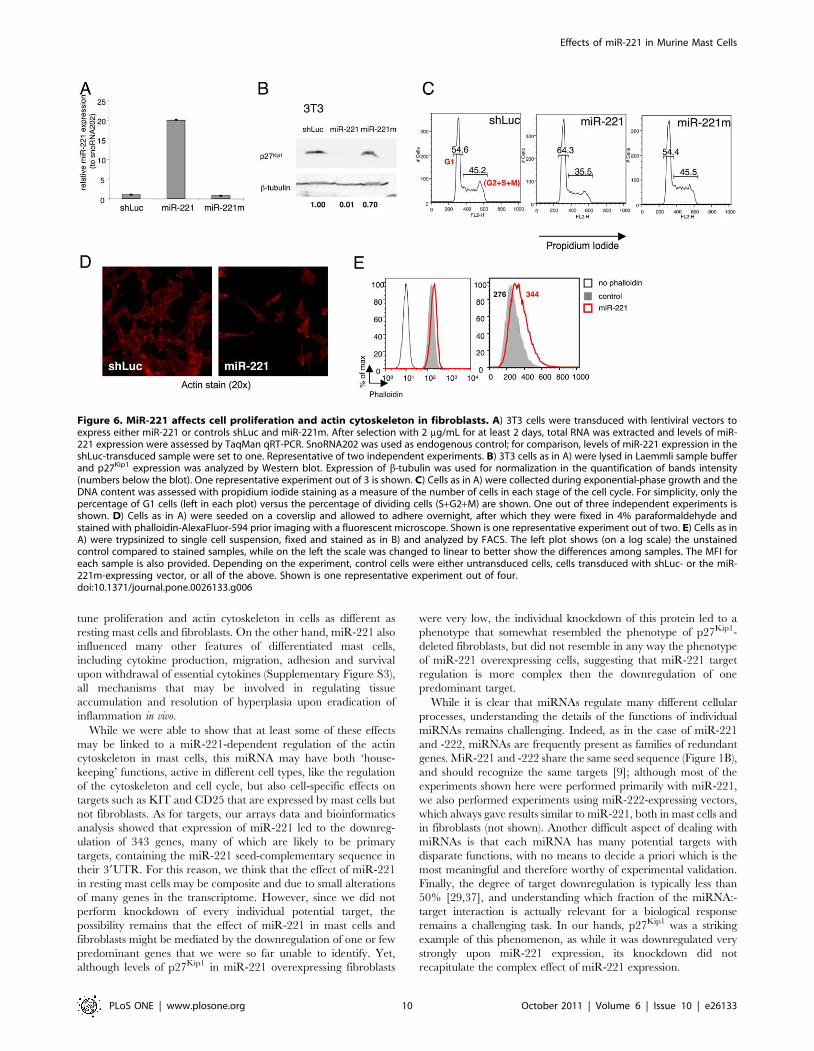
upon transduction with a miR-221 expressing vector (Figure 6A).
MiR-221 overexpression in 3T3 cells led to a strong downregu-
lation of endogenous p27Kip1, even more remarkable than the one
observed in mast cells (Figure 6B). Despite such strong
downregulation of the cell-cycle inhibitor p27Kip1, 3T3 cells
overexpressing miR-221 showed the same reduced proliferation
that we previously described for mast cells (Figure 6C) [8].
Moreover, 3T3 cells overexpressing miR-221 showed overall
altered morphology, with odd, elongated and/or irregular shapes
(Figure 6D), as well as a slightly increased content of F-actin
(Figure 6E), indicating that the miR-221-dependent effects on the
cytoskeleton and cell cycle observed in resting mast cells are likely
to be due to the dysregulation of targets that are ubiquitously
expressed and are therefore cell type-independent. However,
FceRI stimulation led to mast cell-specific (or at least not present in
fibroblasts) effects of miR-221, with increased degranulation and
cytokine production.
Analyzing the data from our transcriptome profiling, we found
that in the ‘cytoskeleton’ group of downregulated genes, the top
candidate, most downregulated gene was Cdkn1b (p27Kip1), and
specifically the one splice variant that can be regulated by miR-
221/-222 (as we previously described in [8]). While p27Kip1 is a
cell cycle inhibitor with a well established role in cell cycle
progression at the G1-S transition, it has also been shown that
cytoplasmic p27Kip1 plays an important role in cell motility and
migration, and that p27Kip1-deficient fibroblasts fail to form long
cellular protrusions, assume an overall rounded shape [34] and
show reduced migration [35]. To assess whether miR-221-
dependent down-regulation of p27Kip1 may have a role in
regulating 3T3 and mast cells shape and cytoskeleton, we therefore
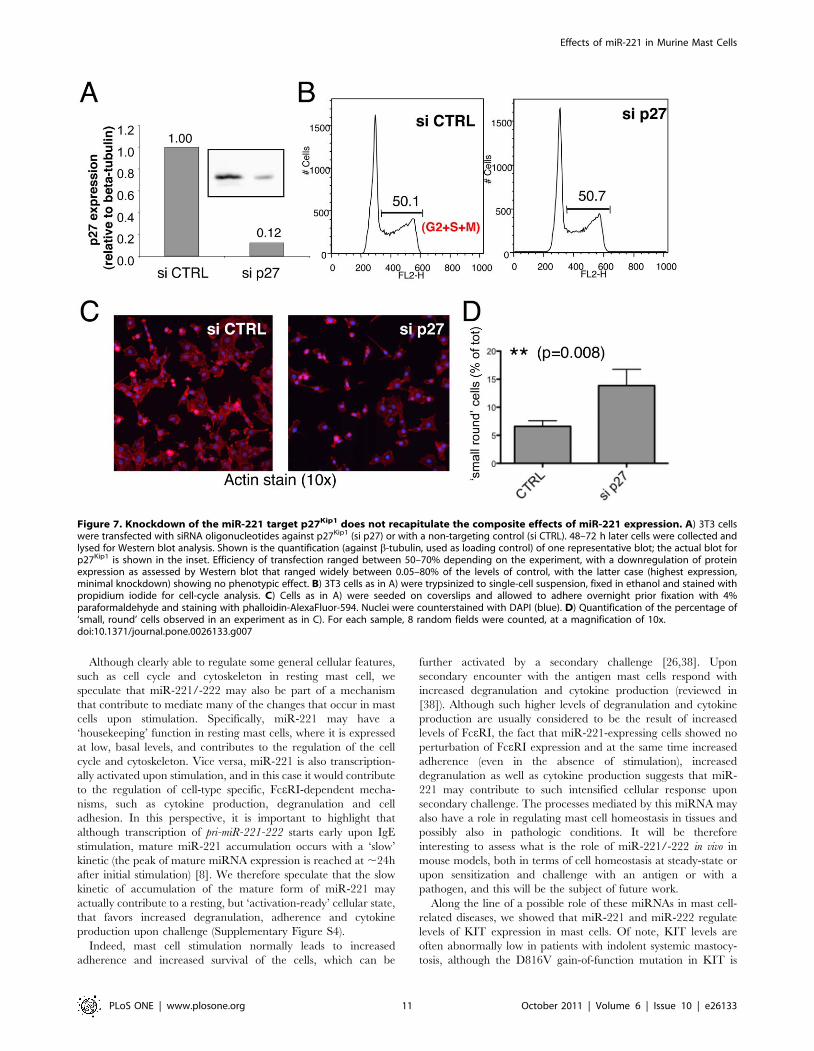
performed a knockdown of p27Kip1 in 3T3 cells using siRNAs
(Figure 7). Efficiency of transfection and p27Kip1 knockdown were
evaluated by transfection and FACS analysis of a fluorescent ds-
oligo (siGLO) and by Western blot, respectively (data not shown
and Figure 7A). It has to be noted that the efficiency of transfection
was at the most ,70%, so that the residual protein observed in
Western blot may in part be due to the fact that some cells still
expressed significant levels of p27Kip1. However, the knockdown of
p27Kip1 did not alter the overall cell-cycle profile of 3T3 cells
(Figure 7B), and the cells did not show any particularly altered
shape, apart from a slight increase in the percentage of cells that
were smaller and more rounded (Figure 7, panels C and D).
Although this effect was fairly modest (even in the experiments
Table 1. Gene Ontology (GO) frequency distribution fordownregulated and upregulated genes.
DOWNREGULATED –Categories Times Observed Probability
Cell migration 15 5.30E-05
Protein folding/modification 30 1.80E-04
Transcription 38 5.70E-04
Lipid metabolism 17 2.30E-03
Cell differentiation 28 3.20E-03
Other metabolism 29 8.00E-03
UPREGULATED – Categories Times Observed Probability
Receptor signaling 73 9.40E-09
Cell proliferation 26 4.60E-08
Cytoskeleton 35 4.70E-08
Cell cycle 27 6.40E-08
Cell adhesion 26 3.30E-07
Cell differentiation 37 1.10E-05
Kinase/Phosphatase signaling 23 1.30E-05
Other metabolism 39 1.90E-05
Protein folding/modification 35 2.50E-05
Development 46 2.90E-05
G-protein signaling 43 8.80E-05
Cell migration 15 3.10E-04
Nucleotide metabolism 26 3.70E-04
Inflammation 11 1.00E-03
Intracellular trafficking 20 5.08E-03
Gene ontology (Bonferroni corrected) of mRNAs that were downregulated orupregulated in the microarrays. A cutoff of P = 0.01 was used for the categoriesto be considered statistically significant.doi:10.1371/journal.pone.0026133.t001
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 8 October 2011 | Volume 6 | Issue 10 | e26133
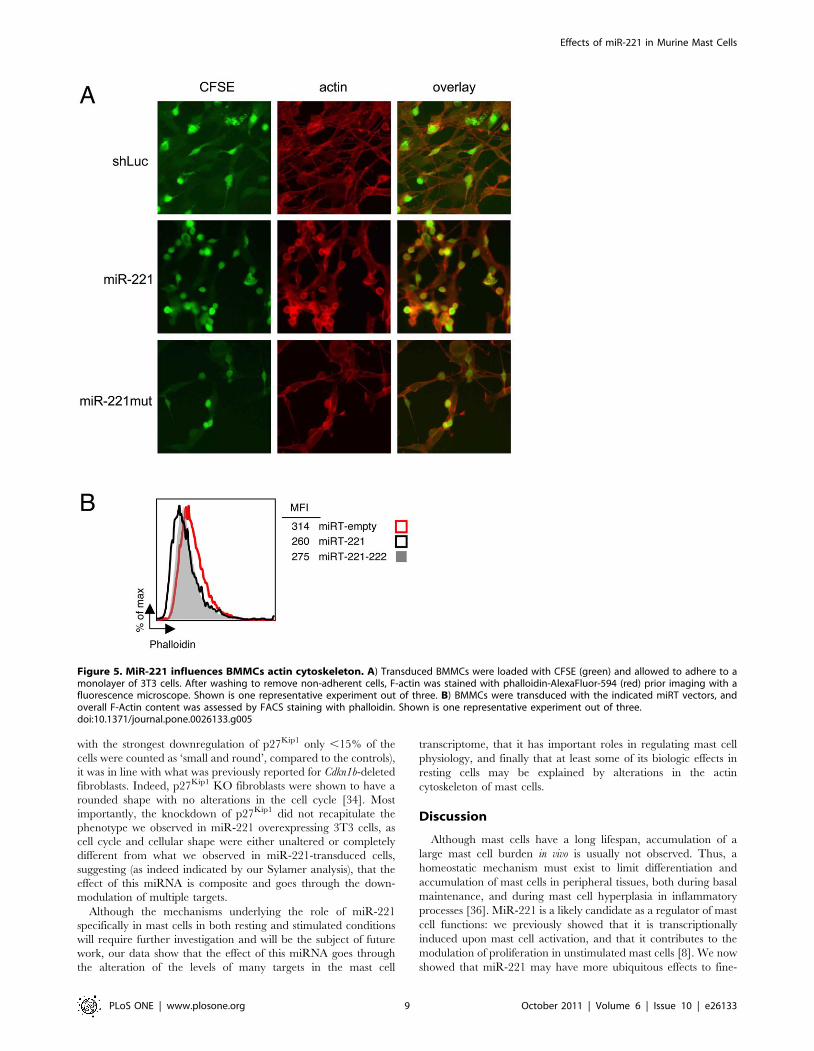
with the strongest downregulation of p27Kip1 only ,15% of the
cells were counted as ‘small and round’, compared to the controls),
it was in line with what was previously reported for Cdkn1b-deleted
fibroblasts. Indeed, p27Kip1 KO fibroblasts were shown to have a
rounded shape with no alterations in the cell cycle [34]. Most
importantly, the knockdown of p27Kip1 did not recapitulate the
phenotype we observed in miR-221 overexpressing 3T3 cells, as
cell cycle and cellular shape were either unaltered or completely
different from what we observed in miR-221-transduced cells,
suggesting (as indeed indicated by our Sylamer analysis), that the
effect of this miRNA is composite and goes through the down-
modulation of multiple targets.
Although the mechanisms underlying the role of miR-221
specifically in mast cells in both resting and stimulated conditions
will require further investigation and will be the subject of future
work, our data show that the effect of this miRNA goes through
the alteration of the levels of many targets in the mast cell
transcriptome, that it has important roles in regulating mast cell
physiology, and finally that at least some of its biologic effects in
resting cells may be explained by alterations in the actin
cytoskeleton of mast cells.
Discussion
Although mast cells have a long lifespan, accumulation of a
large mast cell burden in vivo is usually not observed. Thus, a
homeostatic mechanism must exist to limit differentiation and
accumulation of mast cells in peripheral tissues, both during basal
maintenance, and during mast cell hyperplasia in inflammatory
processes [36]. MiR-221 is a likely candidate as a regulator of mast
cell functions: we previously showed that it is transcriptionally
induced upon mast cell activation, and that it contributes to the
modulation of proliferation in unstimulated mast cells [8]. We now
showed that miR-221 may have more ubiquitous effects to fine-
Figure 5. MiR-221 influences BMMCs actin cytoskeleton. A) Transduced BMMCs were loaded with CFSE (green) and allowed to adhere to amonolayer of 3T3 cells. After washing to remove non-adherent cells, F-actin was stained with phalloidin-AlexaFluor-594 (red) prior imaging with afluorescence microscope. Shown is one representative experiment out of three. B) BMMCs were transduced with the indicated miRT vectors, andoverall F-Actin content was assessed by FACS staining with phalloidin. Shown is one representative experiment out of three.doi:10.1371/journal.pone.0026133.g005
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 9 October 2011 | Volume 6 | Issue 10 | e26133
tune proliferation and actin cytoskeleton in cells as different as
resting mast cells and fibroblasts. On the other hand, miR-221 also
influenced many other features of differentiated mast cells,
including cytokine production, migration, adhesion and survival
upon withdrawal of essential cytokines (Supplementary Figure S3),
all mechanisms that may be involved in regulating tissue
accumulation and resolution of hyperplasia upon eradication of
inflammation in vivo.
While we were able to show that at least some of these effects
may be linked to a miR-221-dependent regulation of the actin
cytoskeleton in mast cells, this miRNA may have both ‘house-
keeping’ functions, active in different cell types, like the regulation
of the cytoskeleton and cell cycle, but also cell-specific effects on
targets such as KIT and CD25 that are expressed by mast cells but
not fibroblasts. As for targets, our arrays data and bioinformatics
analysis showed that expression of miR-221 led to the downreg-
ulation of 343 genes, many of which are likely to be primary
targets, containing the miR-221 seed-complementary sequence in
their 39UTR. For this reason, we think that the effect of miR-221
in resting mast cells may be composite and due to small alterations
of many genes in the transcriptome. However, since we did not
perform knockdown of every individual potential target, the
possibility remains that the effect of miR-221 in mast cells and
fibroblasts might be mediated by the downregulation of one or few
predominant genes that we were so far unable to identify. Yet,
although levels of p27Kip1 in miR-221 overexpressing fibroblasts
were very low, the individual knockdown of this protein led to a
phenotype that somewhat resembled the phenotype of p27Kip1-
deleted fibroblasts, but did not resemble in any way the phenotype
of miR-221 overexpressing cells, suggesting that miR-221 target
regulation is more complex then the downregulation of one
predominant target.
While it is clear that miRNAs regulate many different cellular
processes, understanding the details of the functions of individual
miRNAs remains challenging. Indeed, as in the case of miR-221
and -222, miRNAs are frequently present as families of redundant
genes. MiR-221 and -222 share the same seed sequence (Figure 1B),
and should recognize the same targets [9]; although most of the
experiments shown here were performed primarily with miR-221,
we also performed experiments using miR-222-expressing vectors,
which always gave results similar to miR-221, both in mast cells and
in fibroblasts (not shown). Another difficult aspect of dealing with
miRNAs is that each miRNA has many potential targets with
disparate functions, with no means to decide a priori which is the
most meaningful and therefore worthy of experimental validation.
Finally, the degree of target downregulation is typically less than
50% [29,37], and understanding which fraction of the miRNA:-
target interaction is actually relevant for a biological response
remains a challenging task. In our hands, p27Kip1 was a striking
example of this phenomenon, as while it was downregulated very
strongly upon miR-221 expression, its knockdown did not
recapitulate the complex effect of miR-221 expression.
Figure 6. MiR-221 affects cell proliferation and actin cytoskeleton in fibroblasts. A) 3T3 cells were transduced with lentiviral vectors toexpress either miR-221 or controls shLuc and miR-221m. After selection with 2 mg/mL for at least 2 days, total RNA was extracted and levels of miR-221 expression were assessed by TaqMan qRT-PCR. SnoRNA202 was used as endogenous control; for comparison, levels of miR-221 expression in theshLuc-transduced sample were set to one. Representative of two independent experiments. B) 3T3 cells as in A) were lysed in Laemmli sample bufferand p27Kip1 expression was analyzed by Western blot. Expression of b-tubulin was used for normalization in the quantification of bands intensity(numbers below the blot). One representative experiment out of 3 is shown. C) Cells as in A) were collected during exponential-phase growth and theDNA content was assessed with propidium iodide staining as a measure of the number of cells in each stage of the cell cycle. For simplicity, only thepercentage of G1 cells (left in each plot) versus the percentage of dividing cells (S+G2+M) are shown. One out of three independent experiments isshown. D) Cells as in A) were seeded on a coverslip and allowed to adhere overnight, after which they were fixed in 4% paraformaldehyde andstained with phalloidin-AlexaFluor-594 prior imaging with a fluorescent microscope. Shown is one representative experiment out of two. E) Cells as inA) were trypsinized to single cell suspension, fixed and stained as in B) and analyzed by FACS. The left plot shows (on a log scale) the unstainedcontrol compared to stained samples, while on the left the scale was changed to linear to better show the differences among samples. The MFI foreach sample is also provided. Depending on the experiment, control cells were either untransduced cells, cells transduced with shLuc- or the miR-221m-expressing vector, or all of the above. Shown is one representative experiment out of four.doi:10.1371/journal.pone.0026133.g006
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 10 October 2011 | Volume 6 | Issue 10 | e26133
Although clearly able to regulate some general cellular features,
such as cell cycle and cytoskeleton in resting mast cell, we
speculate that miR-221/-222 may also be part of a mechanism
that contribute to mediate many of the changes that occur in mast
cells upon stimulation. Specifically, miR-221 may have a
‘housekeeping’ function in resting mast cells, where it is expressed
at low, basal levels, and contributes to the regulation of the cell
cycle and cytoskeleton. Vice versa, miR-221 is also transcription-
ally activated upon stimulation, and in this case it would contribute
to the regulation of cell-type specific, FceRI-dependent mecha-
nisms, such as cytokine production, degranulation and cell
adhesion. In this perspective, it is important to highlight that
although transcription of pri-miR-221-222 starts early upon IgE
stimulation, mature miR-221 accumulation occurs with a ‘slow’
kinetic (the peak of mature miRNA expression is reached at ,24h
after initial stimulation) [8]. We therefore speculate that the slow
kinetic of accumulation of the mature form of miR-221 may
actually contribute to a resting, but ‘activation-ready’ cellular state,
that favors increased degranulation, adherence and cytokine
production upon challenge (Supplementary Figure S4).
Indeed, mast cell stimulation normally leads to increased
adherence and increased survival of the cells, which can be
further activated by a secondary challenge [26,38]. Upon
secondary encounter with the antigen mast cells respond with
increased degranulation and cytokine production (reviewed in
[38]). Although such higher levels of degranulation and cytokine
production are usually considered to be the result of increased
levels of FceRI, the fact that miR-221-expressing cells showed no
perturbation of FceRI expression and at the same time increased
adherence (even in the absence of stimulation), increased
degranulation as well as cytokine production suggests that miR-
221 may contribute to such intensified cellular response upon
secondary challenge. The processes mediated by this miRNA may
also have a role in regulating mast cell homeostasis in tissues and
possibly also in pathologic conditions. It will be therefore
interesting to assess what is the role of miR-221/-222 in vivo in
mouse models, both in terms of cell homeostasis at steady-state or
upon sensitization and challenge with an antigen or with a
pathogen, and this will be the subject of future work.
Along the line of a possible role of these miRNAs in mast cell-
related diseases, we showed that miR-221 and miR-222 regulate
levels of KIT expression in mast cells. Of note, KIT levels are
often abnormally low in patients with indolent systemic mastocy-
tosis, although the D816V gain-of-function mutation in KIT is
Figure 7. Knockdown of the miR-221 target p27Kip1 does not recapitulate the composite effects of miR-221 expression. A) 3T3 cellswere transfected with siRNA oligonucleotides against p27Kip1 (si p27) or with a non-targeting control (si CTRL). 48–72 h later cells were collected andlysed for Western blot analysis. Shown is the quantification (against b-tubulin, used as loading control) of one representative blot; the actual blot forp27Kip1 is shown in the inset. Efficiency of transfection ranged between 50–70% depending on the experiment, with a downregulation of proteinexpression as assessed by Western blot that ranged widely between 0.05–80% of the levels of control, with the latter case (highest expression,minimal knockdown) showing no phenotypic effect. B) 3T3 cells as in A) were trypsinized to single-cell suspension, fixed in ethanol and stained withpropidium iodide for cell-cycle analysis. C) Cells as in A) were seeded on coverslips and allowed to adhere overnight prior fixation with 4%paraformaldehyde and staining with phalloidin-AlexaFluor-594. Nuclei were counterstained with DAPI (blue). D) Quantification of the percentage of‘small, round’ cells observed in an experiment as in C). For each sample, 8 random fields were counted, at a magnification of 10x.doi:10.1371/journal.pone.0026133.g007
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 11 October 2011 | Volume 6 | Issue 10 | e26133

present in most of these patients and is sufficient to cause indolent
mastocytosis [39]. This downregulation of KIT expression is even
more pronounced in poor prognosis systemic mastocytosis (i.e.
systemic mastocytosis associated with a clonal hematopoietic
disease, aggressive systemic mastocytosis and mast cell leukemia)
[39], suggesting that multiple layers of regulation of this receptor
may be acting in diseased cells. In this context, it will be interesting
to assess whether miR-221/-222 could be part of a molecular
mechanism involved in KIT regulation specifically in mastocytosis.
Indeed, although the D816V mutation may be sufficient to cause
indolent mastocytosis, other additional defects, that remain to be
identified, are required to induce aggressive mast cell disorders,
and may very well include dysregulated miRNA expression
[40,41,42]. This is even more intriguing considering that CD25,
which is so far the best available diagnostic marker for systemic
mastocytosis with bone marrow involvement [32,33], is also
regulated by these miRNAs. While miR-221/-222 were already
implicated in various human cancers for their effect on
proliferation, we show here for the first time that these miRNAs
also regulate mast cell adhesion, migration, and survival, all
processes that may have implications in mastocytosis.
In summary, although miR-221 doesn’t seem to affect mast cell
differentiation, it influences many features of the biology of
differentiated cells. Specifically, at basal levels, in resting
conditions, these effects are likely to be linked, at least in part,
to the regulation of the actin cytoskeleton and cell cycle, two
features that are regulated by miR-221 independently of the cell
type. However, upon mast cell stimulation, miR-221 may have
some more cell type-specific, activation-dependent effects, influ-
encing the extent of degranulation, adherence and cytokine
production in response to IgE-antigen complexes. Although still
speculative, we propose a model in which miR-221 would have
two different roles in mast cells: in resting cells, it contributes to
normal cell homeostasis through the regulation of the cell cycle
and cytoskeleton, while upon induction following acute stimula-
tion, it contributes to increase the strength of the response to
antigenic challenge. Overall, our work provides new insights into
previously unknown effects of miR-221 in mast cell biology, and
may have important implications for our understanding of the
molecular mechanisms underlying normal and pathologic mast
cell conditions.
Supporting Information
Figure S1 MiR-221 expression can be regulated by thetranscriptional repressor PLZF, but it has no role inBMMC differentiation. A) Lineage depleted (Lin–, lacking
surface expression of CD5, CD45R, CD11b, Gr-1, 7-4 and Ter-
119) and Lin+ bone marrow cells were either immediately used for
RNA extraction or differentiated to mast cells in IL-3 containing
medium [43]. Total RNA from Lin– derived mast cells was used to
assess Plzf mRNA expression (upper panel) and miR-221 (lower panel).
B) Differentiated BMMCs were lentivirally transduced to
ectopically express PLZF. After puromycin selection for 48h, cells
were either left untreated or were stimulated with 20nM PMA and
1 mM ionomycin for 24h, prior RNA extraction and analysis of
Plzf and miR-221 expression. C) Lin– cells were transduced with
the indicated vectors to either force (pAPM) or ablate (miRT)
miR-221 expression, and were cultured for three weeks in the
presence of IL-3 to allow mast cell differentiation. Cultures were
analyzed weekly for the presence of mast cells (FceRIa+ KIT+) by
surface staining. Each point represents one independent experi-
ment. Cells transduced with shLuc, miR-221 and miR-221m
vectors were selected with 2 mg/mL puromycin, while cells
transduced with the miRT vectors (empty, T-221 and T-221-
222) were FACS-sorted for GFP expression. D) Total RNA was
extracted from cells treated as in C) at the end of the
differentiation period (percentage of FceRIa+ KIT+ cells was
greater than 90%), and expression of miR-221 was assessed by
TaqMan qRT-PCR. SnoRNA202 was used as endogenous
control, with levels of miR-221 expression set to one in the
shLuc-transduced sample. Cells transduced with shLuc and miR-
221 vectors were selected with 2 mg/mL puromycin, while cells
transduced with the miRT vectors (empty, T-221 and T-221-222)
were FACS-sorted for GFP expression.
(TIF)
Figure S2 MiR-221 expression does not significantlyalter ERK phosphorylation in mast cells, but favorscytokine production. A) Differentiated BMMCs transduced
with the indicated vectors were sensitized with 1.5 mg/mL of IgE-
anti-DNP for 15min on ice. After washing to remove unbound
IgE, 200ng/mL of DNP-HSA were added, and the cells were
immediately moved to a 37uC water bath for 5, 15 and 45min.
Cells were subsequently fixed, permeabilized and stained with
biotinylated anti-phospho-p44/42 MAPK (Erk1/2). The mean
fluorescence intensity for each sample is indicated on the left. B)
Cells as in A) were either left untreated or were stimulated for
5min with 1 mM ionomycin and 20nM PMA at 37uC, after which
cells were fixed, permeabilized and stained with an anti-Erk1/2
antibody. The mean fluorescence intensity for each sample is
indicated next to the histograms. Shown is one experiment out of
two. C) BMMCs as in A) were stimulated with 1.5 mg/mL IgE-
anti-DNP and 200ng/mL DNP-HSA for 3.5h at 37uC. To block
export from the Golgi, brefeldin A (10 mg/mL) was added in the
last two hours of stimulation. Cells were subsequently fixed,
permeabilized and stained with anti-IL-6-PE and anti-TNF-a-PE-
Cy7. One representative experiment out of three is shown.
(TIF)
Figure S3 MiR-221 expression favors mast cell survivalin response to withdrawal of essential cytokines. BMMCs
were transduced with the indicated vectors and differentiated in
the presence of IL-3 only (top panel) or IL-3 + 10ng/mL SCF (bottom
panel) for three weeks, after which all cytokines were washed out of
the culture medium for at least 24h prior evaluation of early cell
death with annexin V and 7AAD staining. Shown is the
percentage of cells in early apoptosis (annexin V+ 7AAD–).
(TIF)
Figure S4 A ‘dual’ role for miR-221 in mast cells.Speculative model of the possible roles of miR-221 in mast cells.
At resting state, basal levels of miR-221 expression would regulate
homeostatic mechanisms such as the cell cycle and cytoskeleton.
These effects are not necessarily cell type-specific, as they can be
active also in fibroblasts, which also express miR-221. Upon mast
cell activation, ‘early’ effects include the release of preformed
mediators from the cytoplasmic granules and the de novo synthesis
of other mediators, including a broad panel of cytokines. The peak
of accumulation of mature miR-221 is instead a ‘late’ event upon
cell stimulation, and we speculate that it may contribute to the
strength of the response upon secondary challenge, with increased
degranulation, cytokine production and cell adherence.
(TIF)
Methods S1
(DOC)
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 12 October 2011 | Volume 6 | Issue 10 | e26133

Acknowledgments
We thank Anjana Rao and Federica Sallusto for critical reading of the
manuscript. Special thanks also to Michele Proietti, Fabio Grassi,
Mariagrazia Uguccioni and Marcus Thelen for sharing reagents.
Author Contributions
Conceived and designed the experiments: SM. Performed the experiments:
RJM LD NR SM. Analyzed the data: SM. Wrote the paper: SM.
Performed Sylamer analysis: NB HKS AJE.
References
1. Gordon JR, Galli SJ (1990) Mast cells as a source of both preformed and
immunologically inducible TNF-alpha/cachectin. Nature 346: 274–276.2. Kunder CA, St John AL, Li G, Leong KW, Berwin B, et al. (2009) Mast cell-
derived particles deliver peripheral signals to remote lymph nodes. J Exp Med206: 2455–2467.
3. Galli SJ, Tsai M (2010) Mast cells in allergy and infection: versatile effector and
regulatory cells in innate and adaptive immunity. Eur J Immunol 40:1843–1851.
4. Kanellopoulou C, Monticelli S (2008) A role for microRNAs in the developmentof the immune system and in the pathogenesis of cancer. Semin Cancer Biol 18:
79–88.5. Navarro F, Lieberman J (2010) Small RNAs guide hematopoietic cell
differentiation and function. J Immunol 184: 5939–5947.
6. Calin GA, Croce CM (2006) MicroRNA signatures in human cancers. Nat RevCancer 6: 857–866.
7. Xiao C, Rajewsky K (2009) MicroRNA control in the immune system: basicprinciples. Cell 136: 26–36.
8. Mayoral RJ, Pipkin ME, Pachkov M, van Nimwegen E, Rao A, et al. (2009)
MicroRNA-221-222 regulate the cell cycle in mast cells. J Immunol 182:433–445.
9. Lewis BP, Burge CB, Bartel DP (2005) Conserved Seed Pairing, Often Flankedby Adenosines, Indicates that Thousands of Human Genes are MicroRNA
Targets. Cell 120: 15–20.10. Bartonicek N, Enright AJ (2010) SylArray: A web-server for automated detection
of miRNA effects from expression data. Bioinformatics.
11. Bernasconi R, Pertel T, Luban J, Molinari M (2008) A dual task for the Xbp1-responsive OS-9 variants in the mammalian endoplasmic reticulum: inhibiting
secretion of misfolded protein conformers and enhancing their disposal. J BiolChem 283: 16446–16454.
12. Gentner B, Schira G, Giustacchini A, Amendola M, Brown BD, et al. (2009)
Stable knockdown of microRNA in vivo by lentiviral vectors. Nat Methods 6:63–66.
13. Mayoral RJ, Monticelli S (2010) Stable Overexpression of miRNAs in BoneMarrow-Derived Murine Mast Cells Using Lentiviral Expression Vectors.
Methods Mol Biol 667: 205–214.
14. Razin E, Ihle JN, Seldin D, Mencia-Huerta JM, Katz HR, et al. (1984)Interleukin 3: A differentiation and growth factor for the mouse mast cell that
contains chondroitin sulfate E proteoglycan. J Immunol 132: 1479–1486.15. Blank U, Rivera J (2006) Assays for regulated exocytosis of mast cell granules.
Curr Protoc Cell Biol Chapter 15: Unit 15 11.16. Demo SD, Masuda E, Rossi AB, Throndset BT, Gerard AL, et al. (1999)
Quantitative measurement of mast cell degranulation using a novel flow
cytometric annexin-V binding assay. Cytometry 36: 340–348.17. Liu S, Nugroho AE, Shudou M, Maeyama K (2011) Regulation of mucosal mast
cell activation by short interfering RNAs targeting syntaxin4. Immunol Cell Biol.18. Bartonicek N, Enright AJ (2010) SylArray: a web-server for automated detection
of miRNA effects from expression data. Bioinformatics In press.
19. van Dongen S, Abreu-Goodger C, Enright AJ (2008) Detecting microRNAbinding and siRNA off-target effects from expression data. Nat Methods 5:
1023–1025.20. Morgulis A, Gertz EM, Schaffer AA, Agarwala R (2006) A fast and symmetric
DUST implementation to mask low-complexity DNA sequences. J Comput Biol13: 1028–1040.
21. Thomas-Chollier M, Sand O, Turatsinze JV, Janky R, Defrance M, et al. (2008)
RSAT: regulatory sequence analysis tools. Nucleic Acids Res 36: W119–127.22. Brown BD, Naldini L (2009) Exploiting and antagonizing microRNA regulation
for therapeutic and experimental applications. Nat Rev Genet 10: 578–585.23. Doulatov S, Notta F, Rice KL, Howell L, Zelent A, et al. (2009) PLZF is a
regulator of homeostatic and cytokine-induced myeloid development. Genes
Dev 23: 2076–2087.
24. Felli N, Fontana L, Pelosi E, Botta R, Bonci D, et al. (2005) MicroRNAs 221 and
222 inhibit normal erythropoiesis and erythroleukemic cell growth via kit
receptor down-modulation. Proc Natl Acad Sci U S A 102: 18081–18086.
25. Spinello I, Quaranta MT, Pasquini L, Pelosi E, Petrucci E, et al. (2009) PLZF-
mediated control on c-kit expression in CD34(+) cells and early erythropoiesis.
Oncogene 28: 2276–2288.
26. Xiang Z, Ahmed AA, Moller C, Nakayama K, Hatakeyama S, et al. (2001)
Essential role of the prosurvival bcl-2 homologue A1 in mast cell survival after
allergic activation. J Exp Med 194: 1561–1569.
27. Guo H, Ingolia NT, Weissman JS, Bartel DP (2010) Mammalian microRNAs
predominantly act to decrease target mRNA levels. Nature 466: 835–840.
28. Griffiths-Jones S, Grocock RJ, van Dongen S, Bateman A, Enright AJ (2006)
miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids
Res 34: D140–144.
29. Baek D, Villen J, Shin C, Camargo FD, Gygi SP, et al. (2008) The impact of
microRNAs on protein output. Nature 455: 64–71.
30. Selbach M, Schwanhausser B, Thierfelder N, Fang Z, Khanin R, et al. (2008)
Widespread changes in protein synthesis induced by microRNAs. Nature 455:
58–63.
31. le Sage C, Nagel R, Egan DA, Schrier M, Mesman E, et al. (2007) Regulation of
the p27(Kip1) tumor suppressor by miR-221 and miR-222 promotes cancer cell
proliferation. Embo J 26: 3699–3708.
32. Hollmann TJ, Brenn T, Hornick JL (2008) CD25 expression on cutaneous mast
cells from adult patients presenting with urticaria pigmentosa is predictive of
systemic mastocytosis. Am J Surg Pathol 32: 139–145.
33. Sotlar K, Horny HP, Simonitsch I, Krokowski M, Aichberger KJ, et al. (2004)
CD25 indicates the neoplastic phenotype of mast cells: a novel immunohisto-
chemical marker for the diagnosis of systemic mastocytosis (SM) in routinely
processed bone marrow biopsy specimens. Am J Surg Pathol 28: 1319–1325.
34. Belletti B, Pellizzari I, Berton S, Fabris L, Wolf K, et al. (2010) p27kip1 controls
cell morphology and motility by regulating microtubule-dependent lipid raft
recycling. Mol Cell Biol 30: 2229–2240.
35. Besson A, Gurian-West M, Schmidt A, Hall A, Roberts JM (2004) p27Kip1
modulates cell migration through the regulation of RhoA activation. Genes Dev
18: 862–876.
36. Shelburne CP, Ryan JJ (2001) The role of Th2 cytokines in mast cell
homeostasis. Immunol Rev 179: 82–93.
37. Inui M, Martello G, Piccolo S (2010) MicroRNA control of signal transduction.
Nat Rev Mol Cell Biol 11: 252–263.
38. Kawakami T, Galli SJ (2002) Regulation of mast-cell and basophil function and
survival by IgE. Nat Rev Immunol 2: 773–786.
39. Teodosio C, Garcia-Montero AC, Jara-Acevedo M, Sanchez-Munoz L, Alvarez-
Twose I, et al. (2010) Mast cells from different molecular and prognostic
subtypes of systemic mastocytosis display distinct immunophenotypes. J Allergy
Clin Immunol 125: 719–726, 726 e711-726 e714\.
40. Deho L, Monticelli S (2010) Human mast cells and mastocytosis: harnessing
microRNA expression as a new approach to therapy? Arch Immunol Ther Exp
(Warsz) 58: 279–286.
41. Mayerhofer M, Gleixner KV, Hoelbl A, Florian S, Hoermann G, et al. (2008)
Unique effects of KIT D816V in BaF3 cells: induction of cluster formation,
histamine synthesis, and early mast cell differentiation antigens. J Immunol 180:
5466–5476.
42. Zappulla JP, Dubreuil P, Desbois S, Letard S, Hamouda NB, et al. (2005)
Mastocytosis in mice expressing human Kit receptor with the activating
Asp816Val mutation. J Exp Med 202: 1635–1641.
43. Kondo M, Wagers AJ, Manz MG, Prohaska SS, Scherer DC, et al. (2003)
Biology of hematopoietic stem cells and progenitors: implications for clinical
application. Annu Rev Immunol 21: 759–806.
Effects of miR-221 in Murine Mast Cells
PLoS ONE | www.plosone.org 13 October 2011 | Volume 6 | Issue 10 | e26133
Copyright © 2022 FDOKUMEN

















![[2016] JMSC Civ. 221 - Supreme Court of Jamaica](https://static.fdokumen.com/doc/165x107/63245e343a06c6d45f0688d3/2016-jmsc-civ-221-supreme-court-of-jamaica.jpg)


