
Structure of an Ultraweak Protein-Protein Complex and Its Crucial Role in Regulation of Cell...
11
Molecular Cell, Vol. 17, 513–523, February 18, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.molcel.2004.12.031 Structure of an Ultraweak Protein-Protein Complex and Its Crucial Role in Regulation of Cell Morphology and Motility to the limitations of most of the conventional techniques, such as coimmunoprecipitation, pull-down assay, and X-ray crystallography, which become ambiguous or in- effective when analyzing weak binary PPIs, especially those with K D 10 4 M. On the other hand, because Julia Vaynberg, 1,2,4 Tomohiko Fukuda, 3,4 Ka Chen, 3,4 Olga Vinogradova, 1 Algirdas Velyvis, 1 Yizeng Tu, 3 Lily Ng, 2 Chuanyue Wu, 3, * and Jun Qin 1,2, * 1 Structural Biology Program Lerner Research Institute cellular protein concentrations calculated based on The Cleveland Clinic Foundation whole-cell volumes are typically low (nM), weak binary 9500 Euclid Avenue PPIs, as detected in vitro, are often thought to be non- Cleveland, Ohio 44195 specific and are therefore disregarded for further investi- 2 Department of Chemistry gations. This inevitably leads to a biased view of how Cleveland State University proteins function in cells since weak PPIs, when locally 2121 Euclid Avenue enriched or involved in large multisubunit assemblies, Cleveland, Ohio 44115 may play pivotal roles in many important cellular events 3 Department of Pathology (Nooren and Thornton, 2003). For example, the interac- University of Pittsburgh tions between lymphocyte cell surface molecules are Pittsburgh, Pennsylvania 15261 known to be multivalent and avidity driven. The affinity between each individual contacting pair is very low, between 2 10 4 and 10 6 M, and such low affinity is Summary favorable for reversible cell-cell adhesion process (van der Merwe and Davis, 2003). Many enzyme-substrate Weak protein-protein interactions (PPIs) (K D 10 6 M) interactions are also weak, which is advantageous for are critical determinants of many biological pro- broad substrate specificity and rapid turnover during cesses. However, in contrast to a large growing num- catalysis (Schomburg and Schomburg, 2001). Different ber of well-characterized, strong PPIs, the weak PPIs, combinations of transcription factor NFB subunits re- especially those with K D 10 4 M, are poorly explored. sult in different weak affinities, which may differentially Genome wide, there exist few 3D structures of weak regulate gene expression (Huxford et al., 2002). The mo- PPIs with K D 10 4 M, and none with K D 10 3 M. lecular understanding of weak protein-protein recogni- Here, we report the NMR structure of an extremely tion remains elusive and is hampered largely by the lack weak focal adhesion complex (K D 3 10 3 M) be- of thorough functional characterization and structural tween Nck-2 SH3 domain and PINCH-1 LIM4 domain. data. A survey in the Protein Data Bank (PDB) revealed The structure exhibits a remarkably small and polar very few 3D structures of weak PPIs with K D 10 4 M interface with distinct binding modes for both SH3 and (Wang et al., 2001; Kang et al., 2003; Sundquist et al., LIM domains. Such an interface suggests a transient 2004), and none with K D 10 3 M. Nck-2/PINCH-1 association process that may trigger As part of an investigation for elucidating the mecha- rapid focal adhesion turnover during integrin signaling. nisms of integrin-mediated signaling during cell adhe- Genetic rescue experiments demonstrate that this in- sion and migration, we have examined the structural terface is indeed involved in mediating cell shape basis of an ultraweak interaction between adaptors change and migration. Together, the data provide a Nck-2 (also called Nck or Grb4) and PINCH-1. This molecular basis for an ultraweak PPI in regulating fo- interaction, important as part of integrin-mediated focal cal adhesion dynamics during integrin signaling. adhesions (Velyvis et al., 2003), was originally identified by yeast two-hybrid analysis (Tu et al., 1998), but it is Introduction not readily detectable by coimmunoprecipitation experi- ments due to an extremely weak affinity between the The sequencing of complete genomes followed by two proteins (K D 3 10 3 M). This involves the third large-scale proteomics has revealed tens of thousands SH3 domain (SH3-3) of Nck-2 and the fourth LIM domain of protein-protein interactions (PPIs) (Gavin et al., 2002; (LIM4) of PINCH-1 (Tu et al., 1998; Velyvis et al., 2003). Ho et al., 2002). These PPIs, varying in specificity and An NMR-based chemical shift mapping study has sug- affinity, form a web of intricate communication networks gested that PINCH LIM4 might bind to Nck-2 SH3-3 in that convey a variety of biological information for the a mode that is different from that of PINCH-1 LIM1 (Vel- maintenance and development of living organisms. Un- yvis et al., 2003); however, a complete 3D view of the derstanding how individual PPIs function in time and LIM4/SH3-3 complex has been missing, which prohibits space requires detailed genetic, biochemical, and struc- a thorough understanding of how the Nck-2/PINCH-1 tural characterizations. However, while a large number interaction functions at the molecular level. In this study, of strong PPIs (K D 10 6 M) have been characterized we have succeeded in determining the NMR structure to date (Nooren and Thornton, 2003), very limited studies of the complex involving the SH3-3 and LIM4 domains. have directly addressed the role of weak PPIs in various The structure provides the first snapshot of an ultraweak cellular processes. On one hand, this is probably due PPI with distinct binding modes for both SH3 and LIM domains, the two widely distributed protein interaction motifs. To examine how such a weak PPI controls cellu- *Correspondence: [email protected] (J.Q.); [email protected] (C.W.) 4 These authors contributed equally to this work. lar behavior, we further performed genetic small interfer-
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Structure of an Ultraweak Protein-Protein Complex and Its Crucial Role in Regulation of Cell...

Molecular Cell, Vol. 17, 513–523, February 18, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.molcel.2004.12.031
Structure of an Ultraweak Protein-Protein Complexand Its Crucial Role in Regulationof Cell Morphology and Motility
to the limitations of most of the conventional techniques,such as coimmunoprecipitation, pull-down assay, andX-ray crystallography, which become ambiguous or in-effective when analyzing weak binary PPIs, especiallythose with KD � 10�4 M. On the other hand, because
Julia Vaynberg,1,2,4 Tomohiko Fukuda,3,4 Ka Chen,3,4
Olga Vinogradova,1 Algirdas Velyvis,1 Yizeng Tu,3
Lily Ng,2 Chuanyue Wu,3,* and Jun Qin1,2,*1Structural Biology ProgramLerner Research Institute
cellular protein concentrations calculated based onThe Cleveland Clinic Foundationwhole-cell volumes are typically low (�nM), weak binary9500 Euclid AvenuePPIs, as detected in vitro, are often thought to be non-Cleveland, Ohio 44195specific and are therefore disregarded for further investi-2 Department of Chemistrygations. This inevitably leads to a biased view of howCleveland State Universityproteins function in cells since weak PPIs, when locally2121 Euclid Avenueenriched or involved in large multisubunit assemblies,Cleveland, Ohio 44115may play pivotal roles in many important cellular events3 Department of Pathology(Nooren and Thornton, 2003). For example, the interac-University of Pittsburghtions between lymphocyte cell surface molecules arePittsburgh, Pennsylvania 15261known to be multivalent and avidity driven. The affinitybetween each individual contacting pair is very low,between 2 � 10�4 and 10�6 M, and such low affinity isSummaryfavorable for reversible cell-cell adhesion process (vander Merwe and Davis, 2003). Many enzyme-substrateWeak protein-protein interactions (PPIs) (KD � 10�6 M)interactions are also weak, which is advantageous forare critical determinants of many biological pro-broad substrate specificity and rapid turnover duringcesses. However, in contrast to a large growing num-catalysis (Schomburg and Schomburg, 2001). Differentber of well-characterized, strong PPIs, the weak PPIs,combinations of transcription factor NF�B subunits re-especially those with KD � 10�4 M, are poorly explored.sult in different weak affinities, which may differentiallyGenome wide, there exist few 3D structures of weakregulate gene expression (Huxford et al., 2002). The mo-PPIs with KD � 10�4 M, and none with KD � 10�3 M.lecular understanding of weak protein-protein recogni-Here, we report the NMR structure of an extremelytion remains elusive and is hampered largely by the lackweak focal adhesion complex (KD�3 � 10�3 M) be-of thorough functional characterization and structuraltween Nck-2 SH3 domain and PINCH-1 LIM4 domain.data. A survey in the Protein Data Bank (PDB) revealedThe structure exhibits a remarkably small and polarvery few 3D structures of weak PPIs with KD � 10�4 Minterface with distinct binding modes for both SH3 and(Wang et al., 2001; Kang et al., 2003; Sundquist et al.,LIM domains. Such an interface suggests a transient2004), and none with KD � 10�3 M.Nck-2/PINCH-1 association process that may trigger
As part of an investigation for elucidating the mecha-rapid focal adhesion turnover during integrin signaling.nisms of integrin-mediated signaling during cell adhe-Genetic rescue experiments demonstrate that this in-sion and migration, we have examined the structuralterface is indeed involved in mediating cell shapebasis of an ultraweak interaction between adaptorschange and migration. Together, the data provide aNck-2 (also called Nck� or Grb4) and PINCH-1. Thismolecular basis for an ultraweak PPI in regulating fo-interaction, important as part of integrin-mediated focalcal adhesion dynamics during integrin signaling.adhesions (Velyvis et al., 2003), was originally identifiedby yeast two-hybrid analysis (Tu et al., 1998), but it is
Introductionnot readily detectable by coimmunoprecipitation experi-ments due to an extremely weak affinity between the
The sequencing of complete genomes followed by two proteins (KD�3 � 10�3 M). This involves the thirdlarge-scale proteomics has revealed tens of thousands SH3 domain (SH3-3) of Nck-2 and the fourth LIM domainof protein-protein interactions (PPIs) (Gavin et al., 2002; (LIM4) of PINCH-1 (Tu et al., 1998; Velyvis et al., 2003).Ho et al., 2002). These PPIs, varying in specificity and An NMR-based chemical shift mapping study has sug-affinity, form a web of intricate communication networks gested that PINCH LIM4 might bind to Nck-2 SH3-3 inthat convey a variety of biological information for the a mode that is different from that of PINCH-1 LIM1 (Vel-maintenance and development of living organisms. Un- yvis et al., 2003); however, a complete 3D view of thederstanding how individual PPIs function in time and LIM4/SH3-3 complex has been missing, which prohibitsspace requires detailed genetic, biochemical, and struc- a thorough understanding of how the Nck-2/PINCH-1tural characterizations. However, while a large number interaction functions at the molecular level. In this study,of strong PPIs (KD � 10�6 M) have been characterized we have succeeded in determining the NMR structureto date (Nooren and Thornton, 2003), very limited studies of the complex involving the SH3-3 and LIM4 domains.have directly addressed the role of weak PPIs in various The structure provides the first snapshot of an ultraweakcellular processes. On one hand, this is probably due PPI with distinct binding modes for both SH3 and LIM
domains, the two widely distributed protein interactionmotifs. To examine how such a weak PPI controls cellu-*Correspondence: [email protected] (J.Q.); [email protected] (C.W.)
4 These authors contributed equally to this work. lar behavior, we further performed genetic small interfer-

Molecular Cell514
ing RNA (siRNA) experiments by altering the specific (199–202), �2 (221–226), �3 (234–239), �4 (244–248), and�5 (252–254), arranged into two orthogonal antiparallelLIM4/SH3-3 interface in vivo and show that the Nck-2/� sheets. Two loops for the SH3-3 domain, the RT loopPINCH-1 interaction, albeit extremely weak, is indis-(18 residues) and the n-Src loop (7 residues), are alsopensable for regulating cell shape change and motilityseen in other SH3 domains (Figure 1C) (for recent re-as part of the integrin-mediated cytoskeletal network.views, see Mayer, 2001; Cesareni et al., 2002); (b) SimilarOur results not only reveal a molecular basis as to howto free LIM4 (Velyvis et al., 2003), the bound LIM4 domainthe specific Nck-2 SH3/PINCH-1 LIM4 interaction mightconsists of eight � strands (four � sheets) and a single,transiently regulate the cytoskeleton dynamics, but theyshort C-terminal � helix. The first four � strands, �1also demonstrate that NMR spectroscopy, combined(192–193), �2 (198–199), �3 (204–207), and �4 (210–213),with siRNA, is a powerful tool in potentially characteriz-are involved in the first zinc finger, whereas �5 (218–219),ing a large number of ultraweak PPIs genome wide.�6 (224–225), �7 (232–234), and �8 (237–239) are in-volved in the second zinc finger. Residues 242–247 formResults and Discussiona C-terminal � helix.
Structure of the Nck-2 SH3-3/PINCH-1 LIM4 ComplexDistinct Binding InterfaceAs compared to other biophysical techniques, includingfor the LIM4/SH3-3 ComplexX-ray crystallography, NMR is known to be a particularlyThe binding interface between SH3-3 and LIM4 is re-powerful tool in studying weak macromolecular interac-markably small, with a total buried surface area of 480 A2
tions (Qin et al., 2001); however, the technique has been(240 A2 on each domain) (Figure 1D). Such a small con-limited to chemical shift mapping in the past when ap-tact surface, in comparison with large contact areas inplied to study very weak PPIs, and no structure of anyhigh-affinity complexes (usually � 1000 A2; for review,ultraweak protein-protein complex with KD � 10�3 Msee Nooren and Thornton, 2003), is consistent with itshas been reported. To investigate if it is theoreticallyultraweak affinity. The interface is highly electrostaticpossible to determine the NMR structure of a complexand contains two salt bridges involving the LIM4 R197/with ultralow affinity, we undertook an equilibrium-SH3-3 D257 pair and the LIM4 R198/SH3-3 E233 pair,based calculation to estimate if there is a sufficient pop-and one hydrogen bond involving LIM4 R198/SH3-3 andulation of a complex with KD � 3 � 10�3 M that wouldN250 (Figure 1D). R197 and R198 in LIM4 are responsibleallow us to detect intermolecular NOEs, the key struc-for most of the interactions with SH3-3. These interac-tural constraints for NMR structure determination of thetions are mainly electrostatic; however, their aliphaticcomplex. Assuming a 1 mM:1 mM mixture of SH3-3 andside chains make some hydrophobic contacts withLIM4 in a fast-exchange equilibrium (KD � 3 � 10�3 M),SH3-3 V253 and V254 (Figures 1A–1D). Some other hy-we found that a yield of �21% 1:1 ratio complex (0.21drophobic interactions involve the pairs SH3 V253/LIM4mM) should be formed. We then prepared 1.0 mMI192 and SH3 V253/LIM4 P199 (Figure 1D).15N/13C-labeled LIM4 mixed with a slight excess of unla-
Examination of the binding interface revealed a dis-beled SH3-3 (1.5 mM), which should in principle producetinct recognition mode for SH3-3. First, for the typical�0.3 mM of the complex. A 15N/13C filtered, 3D NOESYSH3 domain, the majority of known ligands contain aexperiment (mixing time � 150 ms) successfully de-conserved PxxP motif preceded or followed by a posi-tected 14 intermolecular NOEs. Similarly, a sample oftive residue such as Arg, which adopts polyproline (PPII)15N/13C-labeled SH3-3 (0.6 mM) mixed with 2.5 mM unla-helix conformation upon binding and can bind in twobeled LIM4 detected 17 intermolecular NOEs. In total,opposite orientations—class I (RxxPxxP) and class II
we obtained 31 experimental intermolecular NOE con-(PxxPxxR). Both types of ligands bind in the same PPII
straints (Figure 1A). The assignment of the intermolecu-binding groove located between the RT and n-Src loops
lar NOEs was fully consistent with the chemical shift (Mayer, 2001; Cesareni et al., 2002). The conventionalmapping data (data not shown). Importantly, although PxxP sequences are clearly absent in LIM4 (Figure 2A).chemical shift changes for perturbed residues are small Consistently, DOCK180, a known SH3-3 ligand, con-upon complex formation, which corresponds to the taining the conventional polyproline SH3 recognition se-weak and fast exchange interaction (Velyvis et al., 2003), quence (1819DLLGSPTPPPPPPHQRHL1836) (Tu et al., 2001),only one set of signals was observed for the complex. binds SH3-3 differently from LIM4 (see SupplementalThis finding indicates a single time-averaged structure. Figure S1 at http://www.molecule.org/cgi/content/full/We performed the structure determination in two steps: 17/4/513/DC1/). Second, although numerous sequences(a) calculations of individual structures of SH3-3 and lacking the classical PxxP motif were reported to bindLIM4 in their bound forms; (b) calculations of complex by to SH3 domains (see Table 2), none of them have anyusing the bound SH3-3 and LIM4 structures as starting similarity to those of LIM4. Further, most of the non-templates and by including the intermolecular NOEs (see PxxP ligands still have proline in their sequence andthe Experimental Procedures). These two steps resulted bind directly to the canonical PPII ligand binding pocketin a well-defined structure of the complex (Figure 1B or at the sites overlapping with the PPII binding grooveand Table 1). (Figure 2B). In contrast, LIM4 binds to the SH3-3 domain
in a region that has minimal overlap with any knownStructural Features of the Bound binding sites, including the classical binding pocket (Fig-SH3-3 and LIM4 Domains ure 2B). A summary of representative ligand bindingAs expected, SH3-3 and LIM4 structures in the bound sites and the LIM4 binding site is shown in Figure 2B.forms both adopt a conserved scaffold (Figure 1C): (a) It is clear that the SH3 domain can adopt various binding
epitopes, including the distinct LIM4 binding site.the SH3-3 structure is represented by five � strands, �1
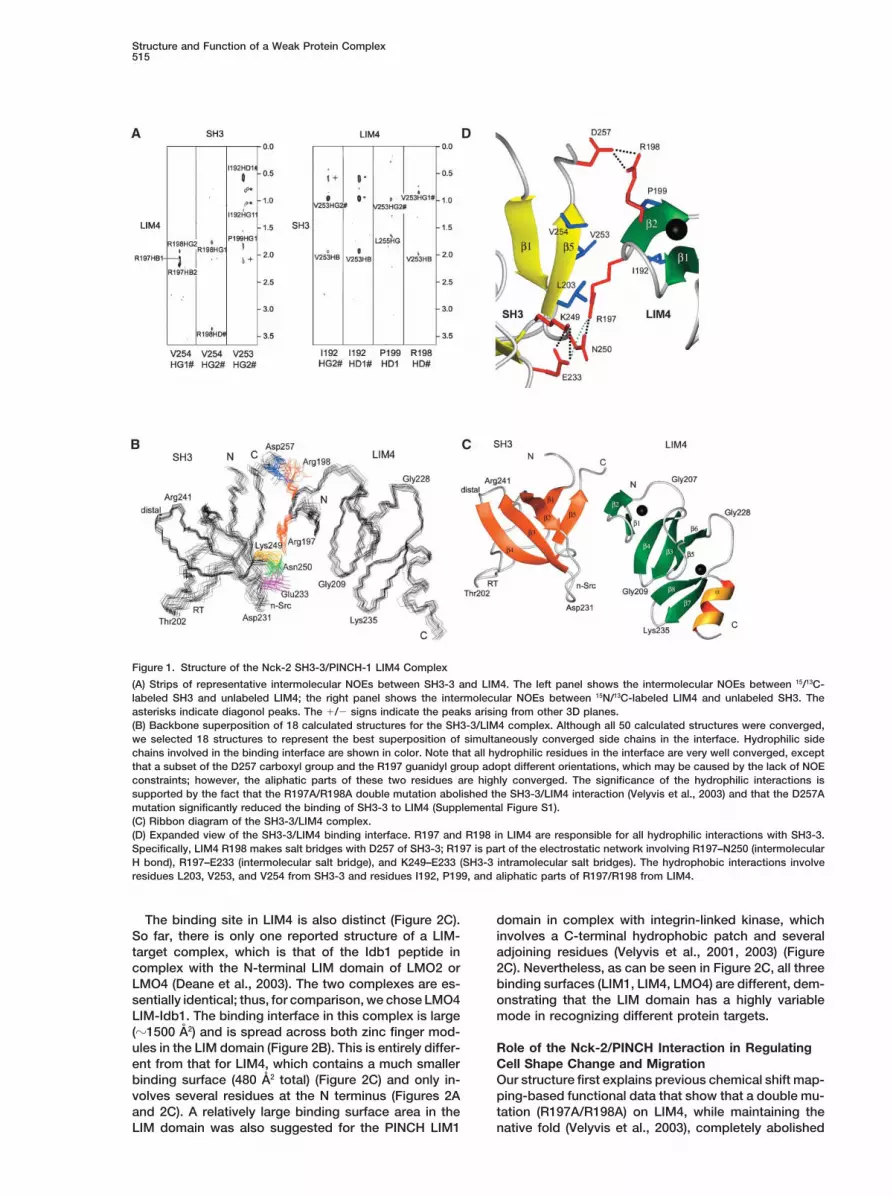
Structure and Function of a Weak Protein Complex515
Figure 1. Structure of the Nck-2 SH3-3/PINCH-1 LIM4 Complex
(A) Strips of representative intermolecular NOEs between SH3-3 and LIM4. The left panel shows the intermolecular NOEs between 15/13C-labeled SH3 and unlabeled LIM4; the right panel shows the intermolecular NOEs between 15N/13C-labeled LIM4 and unlabeled SH3. Theasterisks indicate diagonol peaks. The /� signs indicate the peaks arising from other 3D planes.(B) Backbone superposition of 18 calculated structures for the SH3-3/LIM4 complex. Although all 50 calculated structures were converged,we selected 18 structures to represent the best superposition of simultaneously converged side chains in the interface. Hydrophilic sidechains involved in the binding interface are shown in color. Note that all hydrophilic residues in the interface are very well converged, exceptthat a subset of the D257 carboxyl group and the R197 guanidyl group adopt different orientations, which may be caused by the lack of NOEconstraints; however, the aliphatic parts of these two residues are highly converged. The significance of the hydrophilic interactions issupported by the fact that the R197A/R198A double mutation abolished the SH3-3/LIM4 interaction (Velyvis et al., 2003) and that the D257Amutation significantly reduced the binding of SH3-3 to LIM4 (Supplemental Figure S1).(C) Ribbon diagram of the SH3-3/LIM4 complex.(D) Expanded view of the SH3-3/LIM4 binding interface. R197 and R198 in LIM4 are responsible for all hydrophilic interactions with SH3-3.Specifically, LIM4 R198 makes salt bridges with D257 of SH3-3; R197 is part of the electrostatic network involving R197–N250 (intermolecularH bond), R197–E233 (intermolecular salt bridge), and K249–E233 (SH3-3 intramolecular salt bridges). The hydrophobic interactions involveresidues L203, V253, and V254 from SH3-3 and residues I192, P199, and aliphatic parts of R197/R198 from LIM4.
The binding site in LIM4 is also distinct (Figure 2C). domain in complex with integrin-linked kinase, whichinvolves a C-terminal hydrophobic patch and severalSo far, there is only one reported structure of a LIM-
target complex, which is that of the Idb1 peptide in adjoining residues (Velyvis et al., 2001, 2003) (Figure2C). Nevertheless, as can be seen in Figure 2C, all threecomplex with the N-terminal LIM domain of LMO2 or
LMO4 (Deane et al., 2003). The two complexes are es- binding surfaces (LIM1, LIM4, LMO4) are different, dem-onstrating that the LIM domain has a highly variablesentially identical; thus, for comparison, we chose LMO4
LIM-Idb1. The binding interface in this complex is large mode in recognizing different protein targets.(�1500 A2) and is spread across both zinc finger mod-ules in the LIM domain (Figure 2B). This is entirely differ- Role of the Nck-2/PINCH Interaction in Regulating
Cell Shape Change and Migrationent from that for LIM4, which contains a much smallerbinding surface (480 A2 total) (Figure 2C) and only in- Our structure first explains previous chemical shift map-
ping-based functional data that show that a double mu-volves several residues at the N terminus (Figures 2Aand 2C). A relatively large binding surface area in the tation (R197A/R198A) on LIM4, while maintaining the
native fold (Velyvis et al., 2003), completely abolishedLIM domain was also suggested for the PINCH LIM1
Molecular Cell516
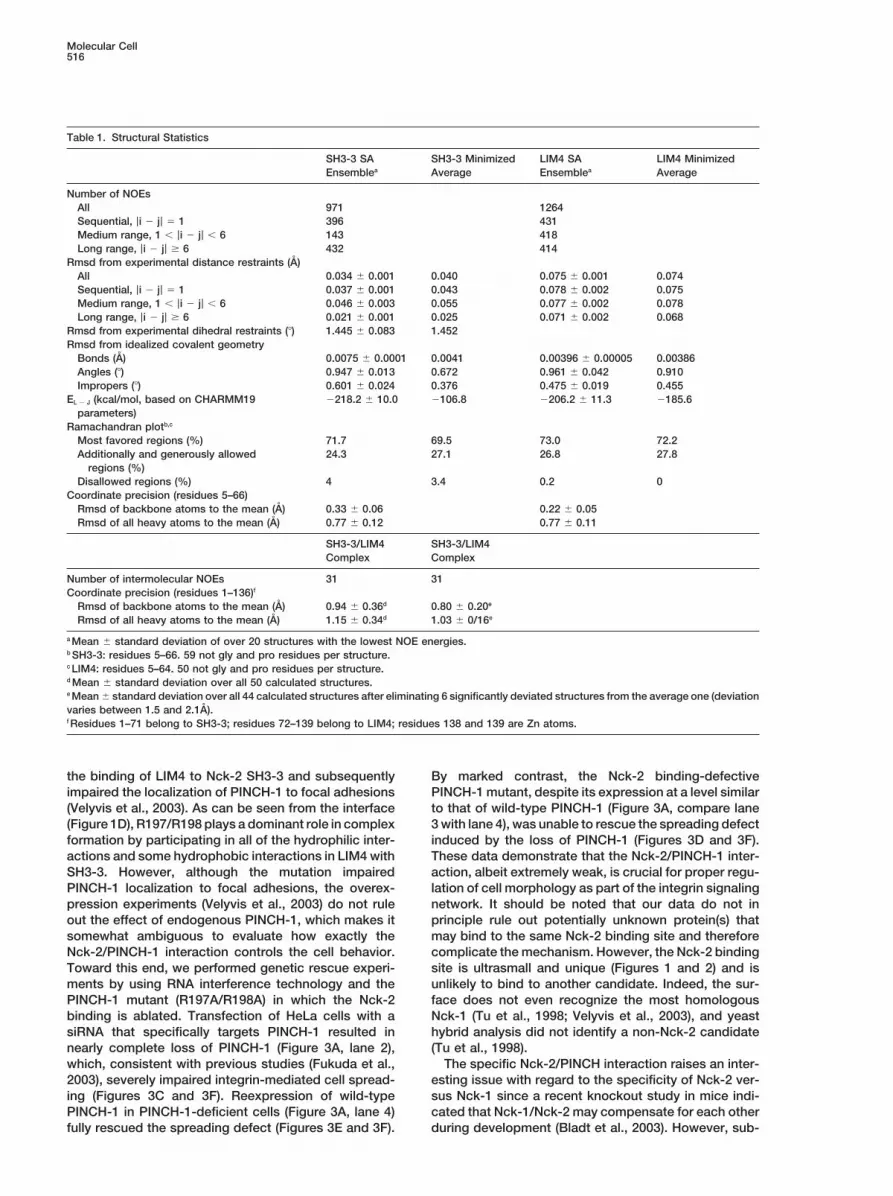
Table 1. Structural Statistics
SH3-3 SA SH3-3 Minimized LIM4 SA LIM4 MinimizedEnsemblea Average Ensemblea Average
Number of NOEsAll 971 1264Sequential, |i � j| � 1 396 431Medium range, 1 � |i � j| � 6 143 418Long range, |i � j| 6 432 414
Rmsd from experimental distance restraints (A)All 0.034 � 0.001 0.040 0.075 � 0.001 0.074Sequential, |i � j| � 1 0.037 � 0.001 0.043 0.078 � 0.002 0.075Medium range, 1 � |i � j| � 6 0.046 � 0.003 0.055 0.077 � 0.002 0.078Long range, |i � j| 6 0.021 � 0.001 0.025 0.071 � 0.002 0.068
Rmsd from experimental dihedral restraints (�) 1.445 � 0.083 1.452Rmsd from idealized covalent geometry
Bonds (A) 0.0075 � 0.0001 0.0041 0.00396 � 0.00005 0.00386Angles (�) 0.947 � 0.013 0.672 0.961 � 0.042 0.910Impropers (�) 0.601 � 0.024 0.376 0.475 � 0.019 0.455
EL � J (kcal/mol, based on CHARMM19 �218.2 � 10.0 �106.8 �206.2 � 11.3 �185.6parameters)
Ramachandran plotb,c
Most favored regions (%) 71.7 69.5 73.0 72.2Additionally and generously allowed 24.3 27.1 26.8 27.8
regions (%)Disallowed regions (%) 4 3.4 0.2 0
Coordinate precision (residues 5–66)Rmsd of backbone atoms to the mean (A) 0.33 � 0.06 0.22 � 0.05Rmsd of all heavy atoms to the mean (A) 0.77 � 0.12 0.77 � 0.11
SH3-3/LIM4 SH3-3/LIM4Complex Complex
Number of intermolecular NOEs 31 31Coordinate precision (residues 1–136)f
Rmsd of backbone atoms to the mean (A) 0.94 � 0.36d 0.80 � 0.20e
Rmsd of all heavy atoms to the mean (A) 1.15 � 0.34d 1.03 � 0/16e
a Mean � standard deviation of over 20 structures with the lowest NOE energies.b SH3-3: residues 5–66. 59 not gly and pro residues per structure.c LIM4: residues 5–64. 50 not gly and pro residues per structure.d Mean � standard deviation over all 50 calculated structures.e Mean � standard deviation over all 44 calculated structures after eliminating 6 significantly deviated structures from the average one (deviationvaries between 1.5 and 2.1A).f Residues 1–71 belong to SH3-3; residues 72–139 belong to LIM4; residues 138 and 139 are Zn atoms.
the binding of LIM4 to Nck-2 SH3-3 and subsequently By marked contrast, the Nck-2 binding-defectivePINCH-1 mutant, despite its expression at a level similarimpaired the localization of PINCH-1 to focal adhesions
(Velyvis et al., 2003). As can be seen from the interface to that of wild-type PINCH-1 (Figure 3A, compare lane3 with lane 4), was unable to rescue the spreading defect(Figure 1D), R197/R198 plays a dominant role in complex
formation by participating in all of the hydrophilic inter- induced by the loss of PINCH-1 (Figures 3D and 3F).These data demonstrate that the Nck-2/PINCH-1 inter-actions and some hydrophobic interactions in LIM4 with
SH3-3. However, although the mutation impaired action, albeit extremely weak, is crucial for proper regu-lation of cell morphology as part of the integrin signalingPINCH-1 localization to focal adhesions, the overex-
pression experiments (Velyvis et al., 2003) do not rule network. It should be noted that our data do not inprinciple rule out potentially unknown protein(s) thatout the effect of endogenous PINCH-1, which makes it
somewhat ambiguous to evaluate how exactly the may bind to the same Nck-2 binding site and thereforecomplicate the mechanism. However, the Nck-2 bindingNck-2/PINCH-1 interaction controls the cell behavior.
Toward this end, we performed genetic rescue experi- site is ultrasmall and unique (Figures 1 and 2) and isunlikely to bind to another candidate. Indeed, the sur-ments by using RNA interference technology and the
PINCH-1 mutant (R197A/R198A) in which the Nck-2 face does not even recognize the most homologousNck-1 (Tu et al., 1998; Velyvis et al., 2003), and yeastbinding is ablated. Transfection of HeLa cells with a
siRNA that specifically targets PINCH-1 resulted in hybrid analysis did not identify a non-Nck-2 candidate(Tu et al., 1998).nearly complete loss of PINCH-1 (Figure 3A, lane 2),
which, consistent with previous studies (Fukuda et al., The specific Nck-2/PINCH interaction raises an inter-esting issue with regard to the specificity of Nck-2 ver-2003), severely impaired integrin-mediated cell spread-
ing (Figures 3C and 3F). Reexpression of wild-type sus Nck-1 since a recent knockout study in mice indi-cated that Nck-1/Nck-2 may compensate for each otherPINCH-1 in PINCH-1-deficient cells (Figure 3A, lane 4)
fully rescued the spreading defect (Figures 3E and 3F). during development (Bladt et al., 2003). However, sub-

Structure and Function of a Weak Protein Complex517
Figure 2. Distinct Binding Modes of SH3-3and LIM4 Domains
(A) Sequence alignment for SH3-3 and LIM4with other representative homologs. For rep-resentative SH3 domains, CSK (Ghose et al.,2001), p67phox (Kami et al., 2002), andScPex13P (Pires et al., 2003) were chosenbased on the fact that their crystal structuresare available for comparison and that the for-mer two recognize conventional PXXP motifsand the last recognizes a nonconventionalsequence. Note that the recognition se-quences for Csk and p67phox are not in Table2 (only non-PxxP binding sequences are inTable 2). Also, although residues marked inblue are essential for PXXP binding, otherneighboring residues in the sequence couldbe potentially involved in binding to immedi-ate regions of the N or C terminus of the PXXPligand. For the LIM domain, PINCH LIM1 andLMO4 were chosen because their bindingmodes have been published (Velyvis et al.,2001, 2003; Deane et al., 2003). The bindinginterface residues for Nck-2 SH3-3 aremarked in red and are contrasted to conven-tional PxxP binding sites in CSK_Hum andp67phox_Hum (blue), nonconventional bindingsites in CSK_Hum and p67phox_Hum (orange),and ScPex13P (magenta). The binding inter-face residues for PINCH-1 LIM4 are markedin red and are contrasted to those in PINCH-1LIM1 and LMO4 (green).(B) The conventional and nonconventionalbinding site residues for SH3 domains in (A)are projected to the SH3-3 surface. PPII: con-ventional PxxP ligand binding site. Both CSKand p67phox SH3 domains have conven-tional and nonconventional binding sites.ScPex13P SH3 has an entirely nonconven-
tional binding mode (magenta), which is better seen by rotating the surface 90� counterclockwise around the z axis. However, all of thesebinding sites are different from the Nck-2 SH3-3 binding site by LIM4.(C) The binding site residues for the LIM domains in (A) are projected to the PINCH-1 LIM4 surface, showing that the LIM4 binding site forSH3-3 is distinct from those of LMO4 and PINCH-1 LIM1.
stantial evidence indicates that the two proteins may tural and genetic data, strongly supports the notion thatthe specific Nck-2/PINCH-1 interaction is important forfunction differently in some somatic cells (Tu et al., 1998;
Chen et al., 2000; Jahn et al., 2001; Cowan and Henke- integrin-mediated cell shape modulation. To further in-vestigate the functional significance of the Nck-2/meyer, 2001; Velyvis et al., 2003). Since depletion of
PINCH-1 causes a severe spreading defect in HeLa cells PINCH-1 interaction, we expressed FLAG-tagged wild-type Nck-2 and the PINCH-1 binding-defective D257A(Figures 3C and 3F), we anticipated that a similar phe-
nomenon occurs for Nck-2 in these cells. To test this, Nck-2 point mutant, respectively, in Nck1�/� Nck2�/�
mouse embryo fibroblasts (MEFs) (Figure 5A). Thewe transfected HeLa cells with a siRNA that specificallytargets Nck-2 and observed a significant reduction of Nck1�/� Nck2�/� cells exhibited significantly impaired
cell motility (Bladt et al., 2003). We repeated the cellNck-2 (Figure 4, lane 2). The suppression of Nck-2 ledto a significant cell spreading defect (Figures 4C–4E) migration experiment with the Nck1�/� Nck2�/� MEFs
under conditions similar to those used by Bladt et al.that is similar to that of PINCH-1 (Figures 3C and 3F).This result, together with the above-mentioned struc- (2003) and obtained almost identical results (Figures 5C
Table 2. Reported Non-PxxP Recognition Sequences for the SH3 Domain
Recognition Sequence Found in Target Proteins by the SH3 Domain
PXXDY e3b1/Abi-1, RN-tre proteins Esp8 (Mongiovi et al., 1999)Px(V/I)(D/N)RxxKP Gab1, AMSH, SLP-76 proteins Hbp, Grb2, STAM2, Gads (Lewitzky et al., 2001;
Kato et al., 2000; Liu et al., 2003)RKxxYxxY SKAP55 Fyb, Fyn, Lsk (Kang et al, 2000)WxxxFxxLE Pex5p Pex13p (Barnett et al., 2000)VPMRLR YAP p53BP2 (Espanel and Sudol, 2001)PPPVIAPRPETKS Pak Pix (Manser et al., 1998)
Molecular Cell518
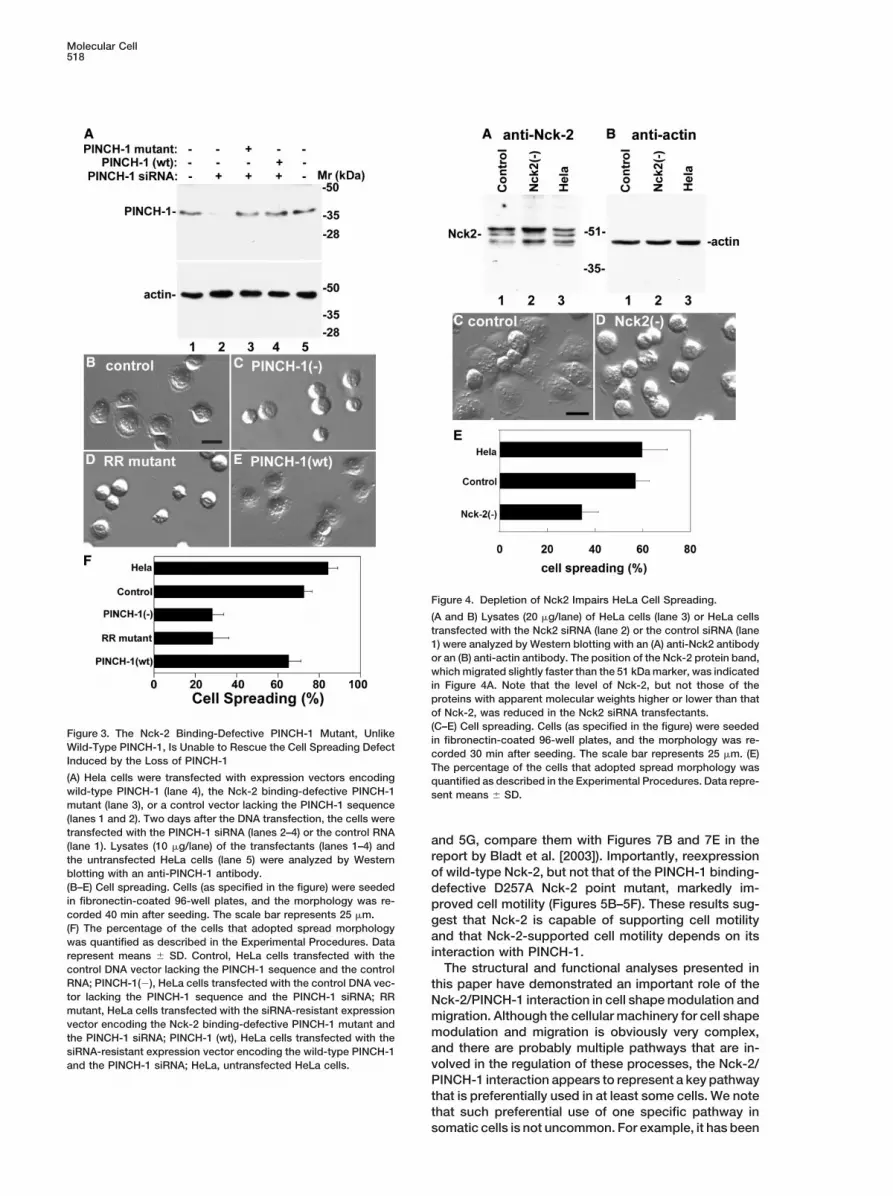
Figure 4. Depletion of Nck2 Impairs HeLa Cell Spreading.
(A and B) Lysates (20 g/lane) of HeLa cells (lane 3) or HeLa cellstransfected with the Nck2 siRNA (lane 2) or the control siRNA (lane1) were analyzed by Western blotting with an (A) anti-Nck2 antibodyor an (B) anti-actin antibody. The position of the Nck-2 protein band,which migrated slightly faster than the 51 kDa marker, was indicatedin Figure 4A. Note that the level of Nck-2, but not those of theproteins with apparent molecular weights higher or lower than thatof Nck-2, was reduced in the Nck2 siRNA transfectants.(C–E) Cell spreading. Cells (as specified in the figure) were seeded
Figure 3. The Nck-2 Binding-Defective PINCH-1 Mutant, Unlikein fibronectin-coated 96-well plates, and the morphology was re-
Wild-Type PINCH-1, Is Unable to Rescue the Cell Spreading Defectcorded 30 min after seeding. The scale bar represents 25 m. (E)
Induced by the Loss of PINCH-1The percentage of the cells that adopted spread morphology was
(A) Hela cells were transfected with expression vectors encoding quantified as described in the Experimental Procedures. Data repre-wild-type PINCH-1 (lane 4), the Nck-2 binding-defective PINCH-1 sent means � SD.mutant (lane 3), or a control vector lacking the PINCH-1 sequence(lanes 1 and 2). Two days after the DNA transfection, the cells weretransfected with the PINCH-1 siRNA (lanes 2–4) or the control RNA
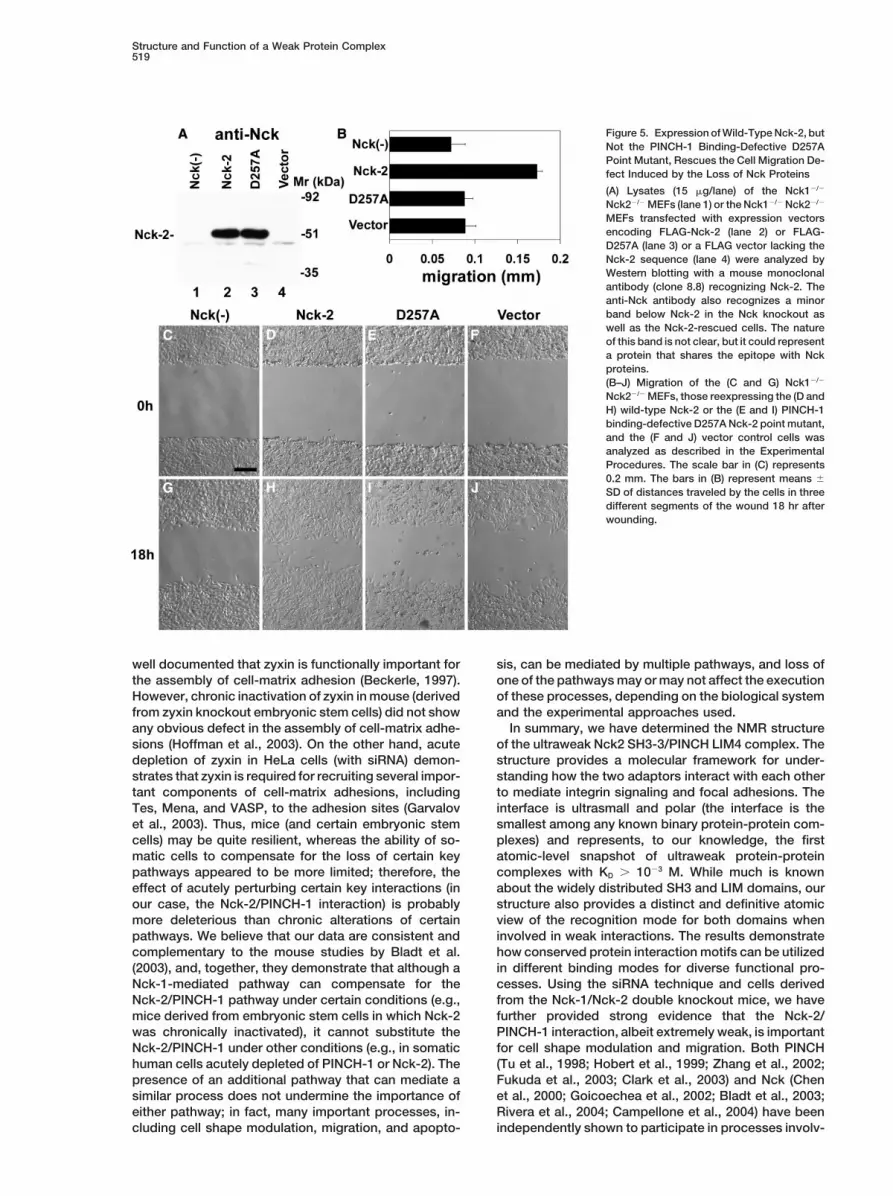
and 5G, compare them with Figures 7B and 7E in the(lane 1). Lysates (10 g/lane) of the transfectants (lanes 1–4) andreport by Bladt et al. [2003]). Importantly, reexpressionthe untransfected HeLa cells (lane 5) were analyzed by Westernof wild-type Nck-2, but not that of the PINCH-1 binding-blotting with an anti-PINCH-1 antibody.
(B–E) Cell spreading. Cells (as specified in the figure) were seeded defective D257A Nck-2 point mutant, markedly im-in fibronectin-coated 96-well plates, and the morphology was re- proved cell motility (Figures 5B–5F). These results sug-corded 40 min after seeding. The scale bar represents 25 m. gest that Nck-2 is capable of supporting cell motility(F) The percentage of the cells that adopted spread morphology
and that Nck-2-supported cell motility depends on itswas quantified as described in the Experimental Procedures. Datainteraction with PINCH-1.represent means � SD. Control, HeLa cells transfected with the
The structural and functional analyses presented incontrol DNA vector lacking the PINCH-1 sequence and the controlRNA; PINCH-1(�), HeLa cells transfected with the control DNA vec- this paper have demonstrated an important role of thetor lacking the PINCH-1 sequence and the PINCH-1 siRNA; RR Nck-2/PINCH-1 interaction in cell shape modulation andmutant, HeLa cells transfected with the siRNA-resistant expression migration. Although the cellular machinery for cell shapevector encoding the Nck-2 binding-defective PINCH-1 mutant and
modulation and migration is obviously very complex,the PINCH-1 siRNA; PINCH-1 (wt), HeLa cells transfected with theand there are probably multiple pathways that are in-siRNA-resistant expression vector encoding the wild-type PINCH-1volved in the regulation of these processes, the Nck-2/and the PINCH-1 siRNA; HeLa, untransfected HeLa cells.PINCH-1 interaction appears to represent a key pathwaythat is preferentially used in at least some cells. We notethat such preferential use of one specific pathway insomatic cells is not uncommon. For example, it has been
Structure and Function of a Weak Protein Complex519
Figure 5. Expression of Wild-Type Nck-2, butNot the PINCH-1 Binding-Defective D257APoint Mutant, Rescues the Cell Migration De-fect Induced by the Loss of Nck Proteins
(A) Lysates (15 g/lane) of the Nck1�/�
Nck2�/� MEFs (lane 1) or the Nck1�/� Nck2�/�
MEFs transfected with expression vectorsencoding FLAG-Nck-2 (lane 2) or FLAG-D257A (lane 3) or a FLAG vector lacking theNck-2 sequence (lane 4) were analyzed byWestern blotting with a mouse monoclonalantibody (clone 8.8) recognizing Nck-2. Theanti-Nck antibody also recognizes a minorband below Nck-2 in the Nck knockout aswell as the Nck-2-rescued cells. The natureof this band is not clear, but it could representa protein that shares the epitope with Nckproteins.(B–J) Migration of the (C and G) Nck1�/�
Nck2�/� MEFs, those reexpressing the (D andH) wild-type Nck-2 or the (E and I) PINCH-1binding-defective D257A Nck-2 point mutant,and the (F and J) vector control cells wasanalyzed as described in the ExperimentalProcedures. The scale bar in (C) represents0.2 mm. The bars in (B) represent means �
SD of distances traveled by the cells in threedifferent segments of the wound 18 hr afterwounding.
well documented that zyxin is functionally important for sis, can be mediated by multiple pathways, and loss ofone of the pathways may or may not affect the executionthe assembly of cell-matrix adhesion (Beckerle, 1997).
However, chronic inactivation of zyxin in mouse (derived of these processes, depending on the biological systemand the experimental approaches used.from zyxin knockout embryonic stem cells) did not show
any obvious defect in the assembly of cell-matrix adhe- In summary, we have determined the NMR structureof the ultraweak Nck2 SH3-3/PINCH LIM4 complex. Thesions (Hoffman et al., 2003). On the other hand, acute
depletion of zyxin in HeLa cells (with siRNA) demon- structure provides a molecular framework for under-standing how the two adaptors interact with each otherstrates that zyxin is required for recruiting several impor-
tant components of cell-matrix adhesions, including to mediate integrin signaling and focal adhesions. Theinterface is ultrasmall and polar (the interface is theTes, Mena, and VASP, to the adhesion sites (Garvalov
et al., 2003). Thus, mice (and certain embryonic stem smallest among any known binary protein-protein com-plexes) and represents, to our knowledge, the firstcells) may be quite resilient, whereas the ability of so-
matic cells to compensate for the loss of certain key atomic-level snapshot of ultraweak protein-proteincomplexes with KD � 10�3 M. While much is knownpathways appeared to be more limited; therefore, the
effect of acutely perturbing certain key interactions (in about the widely distributed SH3 and LIM domains, ourstructure also provides a distinct and definitive atomicour case, the Nck-2/PINCH-1 interaction) is probably
more deleterious than chronic alterations of certain view of the recognition mode for both domains wheninvolved in weak interactions. The results demonstratepathways. We believe that our data are consistent and
complementary to the mouse studies by Bladt et al. how conserved protein interaction motifs can be utilizedin different binding modes for diverse functional pro-(2003), and, together, they demonstrate that although a
Nck-1-mediated pathway can compensate for the cesses. Using the siRNA technique and cells derivedfrom the Nck-1/Nck-2 double knockout mice, we haveNck-2/PINCH-1 pathway under certain conditions (e.g.,
mice derived from embryonic stem cells in which Nck-2 further provided strong evidence that the Nck-2/PINCH-1 interaction, albeit extremely weak, is importantwas chronically inactivated), it cannot substitute the
Nck-2/PINCH-1 under other conditions (e.g., in somatic for cell shape modulation and migration. Both PINCH(Tu et al., 1998; Hobert et al., 1999; Zhang et al., 2002;human cells acutely depleted of PINCH-1 or Nck-2). The
presence of an additional pathway that can mediate a Fukuda et al., 2003; Clark et al., 2003) and Nck (Chenet al., 2000; Goicoechea et al., 2002; Bladt et al., 2003;similar process does not undermine the importance of
either pathway; in fact, many important processes, in- Rivera et al., 2004; Campellone et al., 2004) have beenindependently shown to participate in processes involv-cluding cell shape modulation, migration, and apopto-

Molecular Cell520
Figure 6. Diagram of a Complex Network In-volved in the Cell-Matrix Adhesion Regulationof Actin Cytoskeleton
The complex network that involves PINCH-1and Nck-2 provides insight into how the ul-traweak Nck-2/PINCH-1 interaction tran-siently contributes to the stability and dynam-ics of the supermolecular complex. Thetransient association of Nck-2/PINCH-1 mayfacilitate the rapid assembly/disassembly ofthe complex during cell shape modulationand movement. All proteins labeled in the dia-gram have been shown to be involved in thenetwork (see reviews by Wu, 1999, Wu andDedhar, 2001, Buday et al. 2002, Brakebuschand Fassler, 2003). The list of proteins in thenetwork is expected to continue to grow.
ing cytoskeleton and cell shape modulation (Wu, 1999; supermolecular complex network during cell shapechange and migration. Transient association of Nck-2/Wu and Dedhar, 2001; Buday et al. 2002; Brakebusch
and Fassler, 2003). However, this study has provided PINCH-1 may be an efficient means to cooperate andfacilitate the dynamic regulation of assembly/disassem-for the first time, to our knowledge, unequivocal evi-
dence that the Nck-2/PINCH-1 interaction is an impor- bly of cell-matrix adhesions for rapid cell migration.The structural and functional demonstration of thetant regulatory partner for cell shape modulation and
migration. The existence of such a weak complex re- Nck-2/PINCH-1 interaction also calls for particular at-tention to be paid to weak and ultraweak protein-proteinquires either assembly of a multicomponent complex
network in which the Nck-2/PINCH-1 pair is part of it or interactions involved in a variety of cellular processes.A vigorous molecular/atomic level investigation in thishigh local concentrations of the corresponding compo-
nents. We believe that both mechanisms occur in the area is clearly necessary to gain a thorough understand-ing of how proteins function in living cells. The feasibilityformation of the Nck-2/PINCH-1 complex. First, Figure
6 presents an interaction network involving Nck-2 and of determining structures of ultraweak PPIs by NMR, asdemonstrated in this study, also opens a new avenuePINCH-1 based on this and other published studies.
It is clear that in addition to the direct Nck-2/PINCH-1 for investigating weak PPIs at genomic and proteomicscales.interaction, the multidomain proteins Nck-2 and PINCH-1
are multivalent, bind to numerous other proteins (Wu,Experimental Procedures1999; Wu and Dedhar, 2001; Buday et al. 2002;
Brakebusch and Fassler, 2003), and form an integrin-DNA Constructs, Protein Expression, and Protein Purification
mediated cytoskeletal complex network. Most of these The preparation of DNA constructs for expressing Nck-2 SH3-3 andinteractions converge to the same points, integrin and PINCH LIM4 was described previously (Velyvis et al., 2003). Briefly,
the Nck-2 SH3-3 construct (residues 192–162) was cloned intoactin, e.g., Integrin-FAK-Nck-2-Wasp-Arp2/3-Actin andpTYB11 (New England Biolabs) with SapI and PstI restriction enzymeIntegrin-ILK-PINCH-1-parvin-Actin. The Nck-2/PINCH-1sites, and its mutant, D257A, was made by using the QuickChangepair in this supermolecular complex likely contributes tomethod (Stratagene, Inc.). The PINCH LIM4 domain (residues 188–the stability and dynamics of the total network. Second,251) was sublconed into pGEX-4T-1 (Pharmacia) vector by using
clustering of Nck (Rivera et al., 2004; Campellone et al., BamHI and XhoI sites. To prepare 15N- and/or 13C-labeled proteins,2004) has been shown to promote actin assembly that cells were grown in M9 minimal medium containing 1.1 g l�1
15N-NH4Cl and/or 3.3 g l�1 13C-glucose. For production of LIM4, welikely involves many of the components summarized inused E. coli BL21(DE3) cells, and cultures were grown at 37�C toFigure 6, and PINCH-1 is known to be clustered in actin-OD600 � 0.6 and induced for 4 hr with 1 mM IPTG. In the case ofrich cell-matrix adhesions (Zhang et al., 2002). Hence,Nck-2 SH3-3 and the mutant, E. coli ER2566 cells were grown atthe Nck-2/PINCH-1 interaction appears to occur during37�C until OD600 � 0.6 and were then induced with 0.2 mM IPTG for
the actin assembly in which Nck-2/PINCH-1 are in con- 20 hr at room temperature.tact as part of a supermolecular complex network (Fig-ure 6). On the other hand, the ultralow affinity of the Nck- Protein Purification
LIM4-expressing cells were resuspended in 40 ml phosphate-buf-2/PINCH-1 complex with the small and polar surface, asfered saline (PBS) and were lysed with 0.3 mg ml�1 lysozyme forcompared to other high-affinity complexes, e.g., the ILK/30 min at 4�C. The supernatant was applied to a 5.0 ml GSTrapPINCH-1 complex (Fukuda et al., 2003), implies that theglutathione-sepharose affinity column (Amersham-Pharmacia),
interaction is of transient nature and may be optimized equilibrated with PBS. After washing with 100 ml PBS, 100–200 Ufor rapid dissociation from interacting cytoskeletal part- thrombin (Amersham-Pharmacia) in 5 ml PBS was added to the
column and incubated for 20 hr at room temperature. Subsequentners. This dissociation triggers the disassembly of the

Structure and Function of a Weak Protein Complex521
to digestion, PINCH fragments were eluted with PBS. LIM4 was were calculated with the rmsd 0.94 A for the backbone and 1.15 Afor all heavy atoms (see Table 1 for structural statistics).further purified on anion exchange column Poros HQ20 in 25 mM
phosphate buffer (pH 6.4). Protein was eluted with a NaCl gradientup to 200 mM.
RNA Interference, Rescue, and Cell SpreadingNck-2 SH3-3 or SH3-3 (D257A) mutant-expressing cells were re-For rescue experiments, we introduced silent mutations into thesuspended in 40 ml 50 mM Tris-HCl, 1 mM EDTA, 100 mM NaClPINCH-1 siRNA-targeted region (5�-AAGGTGATGTGGTCTCTGCTC-(pH 8.3) (buffer A). Lysate was cleared by centrifugation and applied3�); the underlined T and G were replaced with C by using a Quick-to 30 ml chitin beads slurry (New England Biolabs) equilibrated inChange site-directed mutagenesis system (Stratagene). The se-buffer A. Resin was washed with 300–400 ml buffer A, and intein-quence of the resulting siRNA resistant PINCH-1 expression vectormediated cleavage at room temperature was induced by passing(pCMV-PINCH-R), which encodes a protein whose amino acid se-50 ml buffer A supplemented with 50 mM DTT. Protein was elutedquence is identical to that of PINCH-1, was confirmed by automatedafter a single 20–24 hr cleavage step. After chitin affinity, SH3-3 orDNA sequence. A similar strategy was used to generate the siRNAthe mutant (D257A) was purified further on Poros HQ20 in 25 mMresistant expression vector encoding the Nck-2 binding-defectiveTris-HCl buffer (pH 8.0).R197R198 → AA PINCH-1 mutant. HeLa cells were transfected withFour samples were made for NMR experiments in 25 mM NaH2PO4, the siRNA-resistant expression vectors encoding wild-type PINCH-1,100 mM NaCl, and 0.1–1 mM TCEP (pH 6.5) buffer: (1) 1.3 mM 15N/13C-the Nck-2 binding-defective mutant in which R197 and R198 werelabeled SH3-3; (2) 1.7 mM 15N/13C-labeled LIM4; (3) 1.0 mM 15N/13C-replaced with AA or a control vector lacking the PINCH-1 sequencelabeled LIM4 mixed with 1.5 mM unlabeled SH3-3; (4) 0.6 mM 15N/using LipofectAmine PLUS (Invitrogen). Two days after the DNA13C-labeled SH3-3 mixed with 2.5 mM unlabeled SH3-3. For SH3-3transfection, the cells were transfected with the PINCH-1 siRNAbinding studies with LIM4 and DOCK180, 0.17 mM wild-type SH3-3that specifically targets PINCH-1 mRNA or an irrelevant siRNA asand the mutant, D257A, were mixed with a slightly excess amounta control as we previously described (Fukuda et al., 2003). Theof LIM4 and DOCK180 peptide (DLLGSPTPPPPPPHQRHL). Theexpression of the wild-type and mutant form of PINCH-1 in theDOCK180 peptide was synthesized and verified as described pre-transfectants was analyzed by Western blotting.viously (Tu et al., 2001).
To suppress Nck-2 expression in HeLa cells, we transfected HeLacells with a siRNA that specifically targets Nck-2 mRNA. The tar-geted sequence that we found to effectively mediate the silencingNMR Spectroscopyof Nck-2 expression is 5�-AACTACGTGGTGGTCCTCAGT-3� (onlyAll heteronuclear NMR experiments for resonance and NOE assign-the sense sequence is shown). HeLa cells were transfected with thements of the free and bound SH3/LIM domains were performed asNck-2-specific siRNA or a 21-nucleotide irrelevant RNA as a controldescribed (Velyvis et al., 2003). The backbone and side chain 1H,by using Oligofectamine (Invitrogen, Inc.). Suppression of Nck-213C, and 15N resonances were assigned first on free SH3-3 and LIM4expression in the Nck-2 siRNA transfectants, but not in the controlbased on triple-resonance experiment spectra of HNHA, HNCO,RNA transfectants, was confirmed by Western blotting with a rabbitHNCACB, H(CCO)NH, and HCCH-TOCSY. NOE distance restraintsanti-Nck-2 antibody (Zymed, CA).for structure calculations of LIM4 and SH3-3 domains were obtained
For the spreading assay, cells were plated in Opti-MEM I serum-from 15N/13C-edited 3D NOESY spectra (mixing time 150 ms). Inter-free medium (Life Technologies) in fibronectin (10 g/ml)-coatedmolecular NOE distance restraints between SH3-3 and LIM4 do-96-well plates. The plates were incubated at 37�C under a 5% CO2/mains for the complex calculation were obtained from 15N/13C-edited95% air atmosphere, and the cell morphology was observed under15N, 13C-filtered 3D NOESY (mixing time 150 ms) (Zwahlen et al.,an Olympus IX70 fluorescence microscope and recorded with a1997). All NMR spectra were recorded on a Varian Inova 500 MHzdigital camera. Unspread cells were defined as round cells, whilespectrometer equipped with a triple-resonance probe and shieldedspread cells were defined as cells with extended processes, asz gradient unit at 25�C. Processing and analysis was done withpreviously described (Zhang et al., 2002). The percentage of cellsnmrPipe (Delaglio et al., 1995) and PIPP (Garrett et al., 1991).adopting spread morphology was quantified by analyzing at least300 cells from 4 randomly selected fields (�80 cells/field).
Structure CalculationsThe structures of bound forms of SH3-3 and LIM4 domains were Nck1/2 Knockout Cells and Rescue Experimentscalculated separately by using protocols described previously (Vel- Nck1�/� Nck2�/� MEFs were kindly provided by Dr. Tony Pawsonyvis et al., 2003) with NOE distance constraints and backbone φ,� and were cultured in DMEM containing 10% FBS as previouslyconstraints obtained from TALOS (Cornilescu et al., 1999). For both described (Bladt et al., 2003). The cells were transfected with ex-domains, 60 structures were calculated starting from the extended pression vectors encoding FLAG-tagged wild-type Nck-2 or thetemplate. LIM4 structure was first calculated without Zn. After SA, PINCH-1 binding-defective D257A Nck-2 point mutant, or a FLAGZn atoms were added and structures were extensively energy mini- vector lacking Nck-2 sequence as a control, by using Lipofectaminemized. A final set of 20 structures with the lowest NOE violation 2000 (Invitrogen). The expression of FLAG-Nck-2 and FLAG-D257Aenergies were selected as the final set. A ramachandran plot was in the corresponding transfectants was confirmed by Western blot-calculated with Procheck-NMR (Laskowski et al., 1996) to examine ting with a monoclonal anti-Nck antibody (clone 8.8) (Tu et al., 1998).the quality of the structures. Surfaces were displayed in Grasp (Ni- Cell migration was assessed by the ability of the cells to migratecholls et al., 1991). Structures were visualized and figures were into a cell-free area as described (Zhang et al., 2002; Bladt et al.,prepared with MolMol (Koradi et al., 1996). 2003). Briefly, 30 hours after transfection, the cell monolayers were
The complex structure was obtained by docking the bound SH3-3 wounded by scratching with a plastic pipette tip. After washing, theand LIM4 domains by using a rigid-body minimization followed by cells were incubated in DMEM containing 1% FBS and 0.5 g/mlconjoined rigid body/torsion angle dynamics in which the interfacial mitomycin C for 18 hr. Images of three different segments of theside chains were given full torsional degrees of freedom, while the cell-free area were recorded at 0 and 18 hr, and the distancesbackbone was free to rotate and translate (Xplor-NIH; Clore and traveled by the cells at the front in three different segments of theSchwieters, 2003). The varied interfacial side chains were based on wound were measured.chemical shift mapping (HSQC perturbation) for both domains andintermolecular NOE distance restraints, which include 199–203, 209–214, 233–236, and 249–258 for Nck-2 SH3-3 and 192, 194–195, and Acknowledgments197–200 for PINCH-1 LIM4. Rigid body minimization uses a targetfunction composed of two terms: intermolecular NOEs and intermo- We thank Dr. Tony Pawson (University of Toronto) for providing the
Nck1�/� Nck2�/� cells; F. Delaglio for nmrPipe software; D. Garrettlecular van der Waals repulsive potential. Note that instead of usingambiguous distance constraints derived from chemical shift map- for Pipp; and Asta Velyviene, Xiaolun Zhang, and Yanwu Yang for
technical assistance. The work was supported by National Institutesping (Clore and Schwieters, 2003), we used unambiguously assigned31 intermolecular NOEs, as described in the text, which were suffi- of Health grants to J.Q. (HL58758, GM62823) and C.W. (DK54639,
GM65188).cient for the convergence of the structure. A total of 50 structures

Molecular Cell522
Received: April 29, 2004 Functional organization of the yeast proteome by systematic analy-sis of protein complexes. Nature 415, 141–147.Revised: August 13, 2004
Accepted: December 29, 2004 Ghose, R., Shehtman, A., Goger, M.J., Ji, H., and Cowburn, D. (2001).Published: February 17, 2005 A novel, specific interaction involving the Csk SH3 domain and its
natural ligand. Nat. Struct. Biol. 8, 998–1004.References Goicoechea, S.M., Yizeng, T., Hua, Y., Chen, K., Shen, T.-L., Guan,
J.-L., and Wu, C. (2002). Nck-2 interacts with focal adhesion kinaseBarnett, P., Bottger, G., Klein, A.T., Tabak, H.F., and Distel, B. (2000). and modulates cell motility. Int. J. Biochem. Cell Biol. 34, 791–805.The peroxisomal membrane protein Pex13p shows a novel mode
Ho, Y., Gruhler, A., Heilbut, A., Bader, G.D., Moore, L., Adams, S.L.,of SH3 interaction. EMBO J. 19, 6382–6391.Millar, A., Taylor, P., Bennett, K., Boutilier, K., et al. (2002). System-
Beckerle, M. (1997). Zyxin: zinc fingers at sites of cell adhesion. atic identification of protein complexes in Saccharomyces cerevis-Bioessays 19, 949–957. iae by mass spectrometry. Nature 415, 180–183.Bladt, F., Aippersbach, E., Gelkop, S., Strasser, G.A., Nash, P., Tafuri, Hobert, O., Moerman, D.G., Clark, K.A., Beckerle, M.C., and Ruvkun,A., Gertler, F.B., and Pawson, T. (2003). The murine Nck SH2/SH3 G. (1999). A conserved LIM protein that affects muscular adherensadaptors are important for the development of mesoderm-derived junction integrity and mechanosensory function in Caenorhabditisembryonic structures and for regulating the cellular actin network. elegans. J. Cell Biol. 144, 45–57.Mol. Cell. Biol. 23, 4586–4597.
Hoffman, L.M., Nix, D.A., Benson, B., Boot-Hanford, R., Gustafsson,Brakebusch, C., and Fassler, R. (2003). The integrin-actin connec- E., Jamora, C., Menzies, A.S., Goh, K.L., Jensen, C.C., Gertler, F.B.,tion, an eternal love affair. EMBO J. 22, 2324–2333. et al. (2003). Targeted disruption of the murine zyxin gene. Mol. Cell.Buday, L., Wunderlich, L., and Tamas, P. (2002). The Nck family Biol. 23, 70–79.of adapter proteins: Regulators of actin cytoskeleton. Cell. Signal. Huxford, T., Mishler, D., Phelps, C.B., Huang, D.B., Sengchantha-14, 723–731. langsy, L.L., Reeves, R., Hughes, C.A., Komives, E.A., and Ghosh,Campellone, K.G., Rankin, S., Pawson, T., Kirschner, M.W., Tipper, G. (2002). Solvent exposed non-contacting amino acids play a criti-D.J., and Leong, J.M. (2004). Clustering of Nck by a 12-residue Tir cal role in NF-kappaB/IkappaBalpha complex formation. J. Mol.phosphopeptide is sufficient to trigger localized actin assembly. J. Biol. 324, 587–597.Cell Biol. 64, 407–416. Jahn, T., Seipel, P., Coutinho, S., Miething, C., Peschel, C., andCesareni, G., Panni, S., Nardelli, G., and Castagnoli, L. (2002). Can Duyster, J. (2001). Grb4/Nckbeta acts as a nuclear repressor ofwe infer peptide recognition specificity mediated by SH3 domains? v-Abl-induced transcription from c-jun/c-fos promoter elements. J.FEBS Lett. 513, 38–44. Biol. Chem. 276, 43419–43427.
Chen, M., She, H., Kim, A., Woodley, D.T., and Li, W. (2000). Nckbeta Kami, K., Takeya, R., Sumimoto, H., and Kohda, D. (2002). Diverseadapter regulates actin polymerization in NIH 3T3 fibroblasts in re- recognition of non-PxxP peptide ligands by the SH3 domain fromsponse to platelet-derived growth factor bb. Mol. Cell. Biol. 20, p67phox, Grb2 and Pex13p. EMBO J. 21, 4268–4276.7867–7880. Kang, H., Freund, C., Duke-Cohan, J.S., Musacchio, A., Wagner,Clark, K.A., McGrail, M., and Beckerle, M.C. (2003). Analysis of G., and Rudd, C.E. (2000). SH3 domain recognition of a proline-PINCH function in Drosophila demonstrates its requirement in integ- independent tyrosine-based RkxxYxY motif in immune cell adapterrin-dependent cellular processes. Development 130, 2611–2621. SKAP55. EMBO J. 19, 2889–2899.
Clore, G.M., and Schwieters, C.D. (2003). Docking of protein-protein Kang, R.S., Daniels, C.M., Francis, S.A., Shih, S.C., Salerno, W.J.,complexes on the basis of highly ambiguous intermolecular distance Hicke, L., and Radhakrishnan, I. (2003). Solution structure of a CUE-restraints derived from 1H/15N chemical shift mapping and backbone ubiquitin complex reveals a conserved mode of ubiquitin binding.15N-1H residual dipolar couplings using conjoined rigid body/torsion Cell 113, 621–630.angle dynamics. J. Am. Chem. Soc. 125, 2902–2912. Kato, M., Miyazawa, K., and Kitamura, N. (2000). A deubiquitinatingCornilescu, G., Delaglio, F., and Bax, A. (1999). Protein backbone enzyme UBPY interacts with the Src homology 3 domain of Hrs-angle restraints from searching a database for chemical shift and binding protein via a novel binding motif PX(V/I)(D/N)RXXKP. J. Biol.sequence homology. J. Biomol. NMR 13, 289–302. Chem. 275, 37481–37487.
Cowan, C.A., and Henkemeyer, M. (2001). The SH2/SH3 adaptor Koradi, R., Billeter, M., and Wuthrich, K. (1996). MOLMOL: a programGrb4 transduces B-ephrin reverse signals. Nature 413, 174–179. for display and analysis of macromolecular structures. J. Mol. Graph.
14, 51–55.Deane, J.E., Mackay, J.P., Kwan, A.H.Y., Sum, E.Y.M., Visvader,J.E., and Matthews, J.M. (2003). Structural basis for the recognition Laskowski, R.A., Rullmannn, J.A., MacArthur, M.W., Kaptein, R., andof Idb1 by the N-terminal LIM domains of LMO2 and LMO4. EMBO Thornton, J.M. (1996). AQUA and PROCHECK-NMR: programs forJ. 22, 2224–2233. checking the quality of protein structures solved by NMR. J. Biomol.
NMR 8, 477–486.Delaglio, F., Grzesiek, S., Vuister, G.W., Zhu, G., Pfeifer, J., and Bax,A. (1995). NMRPipe: a multidimensional spectral processing system Lewitzky, M., Kardinal, C., Gehring, N.H., Schmidt, E.K., Konkol, B.,based on UNIX pipes. J. Biomol. NMR 6, 277–293. Eulitz, M., Birchmeier, W., Schaeper, U., and Feller, S.M. (2001). The
C-terminal SH3 domain of the adapter protein Grb2 binds with highEspanel, X., and Sudol, M. (2001). Yes-associated protein and p53-affinity to sequences in Gab1 and SLP-76 which lack the SH3-typicalbinding protein-2 interact through their WW and SH3 domains. J.P-x-x-P core motif. Oncogene 20, 1052–1062.Biol. Chem. 276, 14514–14523.Liu, Q., Berry, D., Nash, P., Pawson, T., McGlade, C.J., and Li, S.S.Fukuda, T., Chen, K., Shi, X., and Wu, C. (2003). PINCH-1 is an(2003). Structural basis for specific binding of the Gads SH3 domainobligate partner of ILK functioning in cell shape modulation, motilityto an RxxK motif-containing SLP-76 peptide: a novel mode of pep-and survival. J. Biol. Chem. 278, 51324–51333.tide recognition. Mol. Cell 11, 471–481.
Garrett, D.S., Powers, R., Gronenborn, A.M., and Clore, G.M. (1991).Manser, E., Loo, T.H., Koh, C.G., Zhao, Z.S., Chen, X.Q., Tan, L., Tan,A common-sense approach to peak picking in two-, three-, and four-I., Leung, T., and Lim, L. (1998). PAK kinases are directly coupled todimensional spectra using automatic computer analysis of contourthe PIX family of nucleotide exchange factors. Mol. Cell 1, 183–192.diagrams. J. Magn. Reson. 95, 214–220.Mayer, B.J. (2001). SH3 domains: complexity in moderation. J. CellGarvalov, B.K., Higgins, T.E., Sutherland, J.D., Zettl, M., Scaplehorn,Sci. 114, 1253–1263.N., Kocher, T., Piddini, E., Griffiths, G., and Way, M. (2003). The
conformational state of Tes regulates its zyxin-dependent recruit- Mongiovi, A.M., Romano, P.R., Panni, S., Mendoza, M., Wong, W.T.,Musacchio, A., Cesareni, G., and Di Fiore, P.P. (1999). A novel pep-ment to focal adhesions. J. Cell Biol. 161, 33–39.tide-SH3 interaction. EMBO J. 18, 5300–5309.Gavin, A.C., Bosche, M., Krause, R., Grandi, P., Marzioch, M., Bauer,
A., Schultz, J., Rick, J.M., Michon, A.M., Cruciat, C.M., et al. (2002). Nicholls, A., Sharp, K.A., and Honig, B. (1991). Protein folding and

Structure and Function of a Weak Protein Complex523
association - insights from the interfacial and thermodynamic prop-erties of hydrocarbons. Proteins 11, 281–296.
Nooren, I.M., and Thornton, J.M. (2003). Diversity of protein-proteininteractions. EMBO J. 22, 3486–3492.
Pires, J.R., Hong, X., Brockmann, C., Volkmer-Engert, R., Schneider-Mergener, J., Oschkinat, H., and Erdmann, R. (2003). The ScPex13pSH3 domain exposes two distinct binding sites for Pex5p andPex14p. J. Mol. Biol. 326, 1427–1435.
Qin, J., Vinogradova, O., and Gronenborn, A. (2001). Protein-proteininteractions probed by NMR spectroscopy. Methods Enzymol.339, 377–389.
Rivera, G.M., Briceno, C.A., Takeshima, F., Snapper, S.B., andMayer, B.J. (2004). Inducible clustering of membrane-targeted SH3domains of the adaptor protein Nck triggers localized actin polymer-ization. Curr. Biol. 14, 11–22.
Schomburg, D., and Schomburg, I. (2001). Springer Handbook ofEnzymes, Edition 2 (New York: Springer).
Sundquist, W.I., Schubert, H.L., Kelly, B.N., Hill, G.C., Holton, J.M.,and Hill, C.P. (2004). Ubiquitin recognition by the human TSG101protein. Mol. Cell 13, 783–789.
Tu, Y., Li, F., and Wu, C. (1998). Nck-2, a novel Src homology2/3-containing adaptor protein that interacts with the LIM-only proteinPINCH and components of growth factor receptor kinase-signalingpathways. Mol. Biol. Cell 9, 3367–3382.
Tu, Y., Kucik, D.F., and Wu, C. (2001). Identification and kineticanalysis of the interaction between Nck-2 and DOCK180. FEBS Lett.491, 193–199.
van der Merwe, P.A., and Davis, S.J. (2003). Molecular interactionsmediating T cell antigen recognition. Annu. Rev. Immunol. 21,659–684.
Velyvis, A., Yang, Y., Wu, C., and Qin, J. (2001). Solution structureof the focal adhesion adaptor PINCH LIM1 domain and characteriza-tion of its interaction with the integrin-linked kinase ankyrin repeatdomain. J. Biol. Chem. 276, 4932–4939.
Velyvis, A., Vinogradova, O., Wu, C., and Qin, J. (2003). Structuraland functional insights into PINCH LIM4 domain-mediated integrinsignaling. Nat. Struct. Biol. 10, 558–564.
Wang, J.H., Meijers, R., Xiong, Y., Liu, J.H., Sakihama, T., Zhang,R., Joachimiak, A., and Reinherz, E.L. (2001). Crystal structure ofthe human CD4 N-terminal two-domain fragment complexed to aclass II MHC molecule. Proc. Natl. Acad. Sci. USA 98, 10799–10804.
Wu, C. (1999). Integrin-linked kinase and PINCH: partners in regula-tion of cell-extracellular matrix interaction and signal transduction.J. Cell Sci. 112, 4485–4489.
Wu, C., and Dedhar, S. (2001). Integrin linked kinase (ILK) and itsinteractors: a new paradigm for the coupling of extracellular matrixto actin cytoskeleton and signaling complexes. J. Cell Biol. 155,505–510.
Zhang, Y., Guo, L., Chen, K., and Wu, C. (2002). A critical role of thePINCH-integrin-linked kinase interaction in the regulation of cellshape change and migration. J. Biol. Chem. 277, 318–326.
Zwahlen, C., Legault, P., Vincent, S.J.F., Greenblatt, J., Konrat, R.,and Kay, L.E. (1997). Methods for measurement of intermolecularNOEs by multinuclear NMR spectroscopy: application to a bacterio-phage � N-peptide/boxB RNA complex. J. Am. Chem. Soc. 119,711–721.
Accession Numbers
The coordinates of the structures described here have been depos-ited in the Protein Data Bank. The entry code is 1U5S.




















