
Structural insights from GRP78-NF-κB binding interactions: A computational approach to understand a...
9
Structural insights from GRP78–NF-κB binding interactions: A computational approach to understand a possible neuroprotective pathway in brain injuries Marco Fidel Ávila a , Daniel Torrente a , Ricardo Cabezas a , Ludis Morales a , Luis Miguel García-Segura b , Janneth Gonzalez a,n , George E. Barreto a,n a Departamento de Nutrición y Bioquímica, Facultad de Ciencias, Pontificia Universidad Javeriana, Bogotá D.C., Colombia b Instituto Cajal, CSIC, Avenida Doctor Arce 37, 28002 Madrid, Spain HIGHLIGHTS GRP78–NF-κB interactions may explain a significant regulation of cell survival and inflammation. Amino acids GLU 215 and GLY 403 in GRP78 are involved in the interaction with NF- κB. IkB alpha as a possible subunit regulated by GRP78. N- and C-terminal domains of GRP78 are potential interacting sites. article info Article history: Received 19 March 2013 Received in revised form 23 September 2013 Accepted 9 December 2013 Available online 18 December 2013 Keywords: Protein interaction NF-κB regulation Brain damage Neuroprotection abstract GRP78 participates in multiple functions in the cell during normal and pathological conditions, controlling calcium homeostasis, protein folding and Unfolded Protein Response. GRP78 is located in the endoplasmic reticulum, but it can change its location under stress, hypoxic and apoptotic conditions. NF-κB represents the keystone of the inflammatory process and regulates the transcription of several genes related with apoptosis, differentiation, and cell growth. The possible relationship between GRP78–NF-κB could support and explain several mechanisms that may regulate a variety of cell functions, especially following brain injuries. Although several reports show interactions between NF-κB and Heat Shock Proteins family members, there is a lack of information on how GRP78 may be interacting with NF-κB, and possibly regulating its downstream activation. Therefore, we assessed the computational predictions of the GRP78 (Chain A) and NF-κB complex (IkB alpha and p65) protein–protein interactions. The interaction interface of the docking model showed that the amino acids ASN 47, GLU 215, GLY 403 of GRP78 and THR 54, ASN 182 and HIS 184 of NF-κB are key residues involved in the docking. The electrostatic field between GRP78–NF-κB interfaces and Molecular Dynamic simulations support the possible interaction between the proteins. In conclusion, this work shed some light in the possible GRP78–NF-κB complex indicating key residues in this crosstalk, which may be used as an input for better drug design strategy targeting NF-κB downstream signaling as a new therapeutic approach following brain injuries. & 2013 Elsevier Ltd. All rights reserved. 1. Introduction The molecular chaperone GRP78 belongs to the highly con- served family of proteins Hsp (Heat shock proteins). It participates in multiple cellular functions, including facilitation of nascent protein folding inside Endoplasmic reticulum (ER), modulation of the Unfolded Protein Response (UPR), regulation the efflux of endoplasmic reticulum Ca 2 þ (Coe and Michalak, 2009) and regulation of cell death related with apoptotic mechanisms (Wang et al., 2010). GRP78 interacts and binds to caspases 7 and 9 interfering with the cleavage of these proteins (Dong et al., 2011; Zhang and Zhang, 2010). Previous studies suggest a role of GRP78 in cell death by regulating apoptotic mechanisms in purkinje cell in the cerebellum (Wang et al., 2010) and SHSY5Y neuroblastoma cells treated with prostaglandin under rotenone insult (Xiaoxia et al., 2002), cerebral ischemia (Ouyang et al., 2012a), and signals as a membrane receptor (Burikhanov et al., 2013; Liu et al., 2010; Nakajima et al., 2011) transducing intracellular ligands and extra- cellular signals associated to cancer, cell survival, cell death program, proliferation and metastasis (Gonzalez-Gronow et al., 2009; Kim et al., 2006; Lee et al., 2006; Misra and Pizzo, 2010; Misra et al., 2011; Rao et al., 2002). Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/yjtbi Journal of Theoretical Biology 0022-5193/$ - see front matter & 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.jtbi.2013.12.010 n Corresponding authors. Tel.: þ57 1 320 8320x4096. E-mail addresses: [email protected] (J. Gonzalez), [email protected] (G.E. Barreto). Journal of Theoretical Biology 345 (2014) 43–51
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Structural insights from GRP78-NF-κB binding interactions: A computational approach to understand a...

Structural insights from GRP78–NF-κB binding interactions:A computational approach to understand a possibleneuroprotective pathway in brain injuries
Marco Fidel Ávila a, Daniel Torrente a, Ricardo Cabezas a, Ludis Morales a, LuisMiguel García-Segura b, Janneth Gonzalez a,n, George E. Barreto a,n
a Departamento de Nutrición y Bioquímica, Facultad de Ciencias, Pontificia Universidad Javeriana, Bogotá D.C., Colombiab Instituto Cajal, CSIC, Avenida Doctor Arce 37, 28002 Madrid, Spain
H I G H L I G H T S
� GRP78–NF-κB interactions may explain a significant regulation of cell survival and inflammation.� Amino acids GLU 215 and GLY 403 in GRP78 are involved in the interaction with NF- κB.� IkB alpha as a possible subunit regulated by GRP78.� N- and C-terminal domains of GRP78 are potential interacting sites.
a r t i c l e i n f o
Article history:Received 19 March 2013Received in revised form23 September 2013Accepted 9 December 2013Available online 18 December 2013
Keywords:Protein interactionNF-κB regulationBrain damageNeuroprotection
a b s t r a c t
GRP78 participates in multiple functions in the cell during normal and pathological conditions, controllingcalcium homeostasis, protein folding and Unfolded Protein Response. GRP78 is located in the endoplasmicreticulum, but it can change its location under stress, hypoxic and apoptotic conditions. NF-κB represents thekeystone of the inflammatory process and regulates the transcription of several genes related with apoptosis,differentiation, and cell growth. The possible relationship between GRP78–NF-κB could support and explainseveral mechanisms that may regulate a variety of cell functions, especially following brain injuries. Althoughseveral reports show interactions between NF-κB and Heat Shock Proteins family members, there is a lack ofinformation on how GRP78 may be interacting with NF-κB, and possibly regulating its downstreamactivation. Therefore, we assessed the computational predictions of the GRP78 (Chain A) and NF-κB complex(IkB alpha and p65) protein–protein interactions. The interaction interface of the docking model showed thatthe amino acids ASN 47, GLU 215, GLY 403 of GRP78 and THR 54, ASN 182 and HIS 184 of NF-κB are keyresidues involved in the docking. The electrostatic field between GRP78–NF-κB interfaces and MolecularDynamic simulations support the possible interaction between the proteins. In conclusion, this work shedsome light in the possible GRP78–NF-κB complex indicating key residues in this crosstalk, which may beused as an input for better drug design strategy targeting NF-κB downstream signaling as a new therapeuticapproach following brain injuries.
& 2013 Elsevier Ltd. All rights reserved.
1. Introduction
The molecular chaperone GRP78 belongs to the highly con-served family of proteins Hsp (Heat shock proteins). It participatesin multiple cellular functions, including facilitation of nascentprotein folding inside Endoplasmic reticulum (ER), modulation ofthe Unfolded Protein Response (UPR), regulation the efflux ofendoplasmic reticulum Ca2þ (Coe and Michalak, 2009) andregulation of cell death related with apoptotic mechanisms
(Wang et al., 2010). GRP78 interacts and binds to caspases 7 and9 interfering with the cleavage of these proteins (Dong et al., 2011;Zhang and Zhang, 2010). Previous studies suggest a role of GRP78in cell death by regulating apoptotic mechanisms in purkinje cellin the cerebellum (Wang et al., 2010) and SHSY5Y neuroblastomacells treated with prostaglandin under rotenone insult (Xiaoxiaet al., 2002), cerebral ischemia (Ouyang et al., 2012a), and signalsas a membrane receptor (Burikhanov et al., 2013; Liu et al., 2010;Nakajima et al., 2011) transducing intracellular ligands and extra-cellular signals associated to cancer, cell survival, cell deathprogram, proliferation and metastasis (Gonzalez-Gronow et al.,2009; Kim et al., 2006; Lee et al., 2006; Misra and Pizzo, 2010;Misra et al., 2011; Rao et al., 2002).
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/yjtbi
Journal of Theoretical Biology
0022-5193/$ - see front matter & 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.jtbi.2013.12.010
n Corresponding authors. Tel.: þ57 1 320 8320x4096.E-mail addresses: [email protected] (J. Gonzalez),
[email protected] (G.E. Barreto).
Journal of Theoretical Biology 345 (2014) 43–51

GRP78 interacts with several proteins due to its multiple functions,but the mechanisms of some interactions are not completely clear. Online databases, as STRING and EBI-EMBL IntAct, report interactionswith transcription and translation factors and other chaperones(Szklarczyk et al., 2011). For example, GRP78 interacts with three ofthe principal regulators of the UPR, IRE1, ATF6 and PERK, thusmodulating the activation of cell response to ER stress (Chen andGao, 2002; Wang et al., 2009). Two GRP78 domains had beenpreviously identified: (i) the 44-kDa NH2-terminal domain, which isresponsible for its ATPase activity (Stetler et al., 2010); and (ii) a 20-kDa COOH-terminal polypeptide binding domain (Delie et al., 2012).In addition, it contains a highly helical and variable 10-kDa COOH-terminal tail of unknown function, but, as an Hsp member, it could actas a lid for the substrate binding domain (Wang et al., 2009). Prositedatabase reports four different GRP78 signature patterns, IDLGTTYS,VFDLGGGTFDVSLL, IVLVGGSTRIPKIQQ and KDEL, demonstrating thatthese signature patterns are related with possible interacting sites andthe ER location signal (Sigrist et al., 2013).
Previous studies report that many chaperone family membersare located in different cellular compartments with multiple andvaried functions as signaling and homeostasis regulation(Nakajima et al., 2011; Quinones et al., 2008; Stetler et al., 2010),and that GRP78 may have extracellular function as a possibleinflammatory resolution molecule (Shields et al., 2012). However,the mechanisms by which GRP78 translocate to another cell placesare poorly understood. It is known that small pool of GRP78translocate to membrane in association with MTJ-1 (Chevalieret al., 2000), acting as a co-receptor to viruses, major histocompat-ibility complex Class I and alpha-2 macroglobulin (Misra et al.,2011). In fact, the activation of this receptor promotes proliferativeand anti-apoptotic functions in the cell (Misra and Pizzo, 2010)with a crosstalk between AKT, UPR and NF-κB (Misra et al., 2006;Misra and Pizzo, 2010). Independently, this pathway is alsoactivated via Subtilase AB, a protease which selectively degradesGRP78 and induces the activation of NF-κB (Yamazaki et al., 2009).It is known that Hsp proteins may regulate apoptosis, thereforecomputational studies have been directed to design compoundsthat might interact with the hATPase domain of Hsp70 to mod-ulate the apoptotic response (Lee et al., 2008). Besides thesecellular functions driven or regulated by GRP78, this co-chaperone may also regulate transcriptions factors implicated inthe inflammatory process following brain damage, possibly byreducing neuronal damage and oxidative stress withdrawal(Ouyang et al., 2012b; Xie et al., 2012; Zhang and Kaufman,2008), being NF-κB as the main transcription factor implicatedin mediating downstream pro-inflammatory signaling.
Nuclear factor-κB (NF-κB) is a pleiotropic transcription factor thatregulates genes involved at multiple stages of immune responses,including innate immune cell activation, inflammation, dendritic cellmaturation and lymphocyte activation, differentiation, cell growth,tumorigenesis and apoptosis (Hayden and Ghosh, 2008). NF-κBactivation is tightly controlled by canonical and atypical pathwaysthat regulate proteolysis of IκBα (an inhibitor of NF-κB) and IκBrelated proteins (Beinke and Ley, 2004; Hayden and Ghosh, 2008).Regulation of canonical NF-κB signaling by ubiquitination anddeubiquitination plays a critical role in NF-κB activation (Karin andBen-Neriah, 2000; Swinney et al., 2002).
NF-κB is a homo- or heterodimeric complex formed by the Rel-like domain-containing proteins RELA/p65, RELB, NF-κB1/p105,NF-κB1/p50, REL and NF-κB2/p52 and the heterodimeric p65-p50complex (Hayden and Ghosh, 2008). The common upstreamregulatory step in both canonical and non-canonical pathways isactivation of an IκBα kinase (IKK) complex, which consists ofcatalytic kinase subunits (IKKα and/or IKKβ) and a scaffold sensingprotein, NF-κB essential modulator (NEMO), also named IKKγ(Hayden and Ghosh, 2008). As such, activation of NF-κB dimers is
the result of phosphorylation-induced degradation of IκBα inhi-bitor by IKK, which enables NF-κB dimers to enter the nucleus andactivate specific target gene expression (Gilmore, 1990).
Possible NF-κB activation is happen to occur through calciumrelease (Pahl and Baeuerle, 1996; Petranka et al., 2001), reactiveoxygen species production (Chandel et al., 2000), IRE1, and PERKsignal pathway during ER stress (Pahl and Baeuerle, 1997). On theother hand, two previous reports indicate the expression of GRP78and NF-κB under oxidative stress and ER stress conditions(Bernstein et al., 1999; Crowley-Weber et al., 2003). It is possiblethe existence of GRP78–NF-κB family members protein interac-tions, as there are several reports showing interactions andregulation of NF-κB with Hsp members (Guzhova et al., 1997;Hinz et al., 2007; Kammanadiminti and Chadee, 2006; Lim et al.,1990; Park et al., 2003), the same protein family where GRP78belongs. For example, Hsp70 interacts with IκBα and p65 subunits(Guzhova et al., 1997), while Hsp27 and Hsp90 regulates TNF alphathrough IKK (Dodd et al., 2009). Additionally, it is suggested thatHsp0s are key regulators of NF-κB activity (Chen and Gao, 2002;Kim et al., 2013; Tam et al., 2012; Xiaoxia et al., 2002). Althoughprevious studies report binary interactions between GRP78 andNF-κB pathway members (Bouwmeester et al., 2004; Rual et al.,2005; Tieri et al., 2012; Fig. S3), the significance and role of theseinteractions remain unclear. A better characterization of GRP78–NF-κB crosstalk could support and explain several mechanismsthat may regulate homeostasis, apoptosis, inflammation and pro-liferation in the cells of brain pathologic conditions (Lin et al.,2012; Pereira., 2013). Experimental evidence points to a possibleintegration of GRP78 with regulation of the inflammatory process.For example, GRP78 physically binds to Tissue Factor (TF;Bhattacharjee et al., 2005) and negatively regulates inflammatoryand procoagulant activity. Additionally, TF mediates inflammation(Chu, 2005) and is highly expressed in astrocytes under patholo-gical conditions (Eddleston et al., 1993; Gonzalez-Dunia et al.,1996). A current research workflow tries to propose and explainthe ER stress and inflammation crosstalk in both physiological andpathological conditions (Hotamisligil, 2010; Kolattukudy and Niu,2012; Tam et al., 2012; Zhang, 2010; Zhang and Kaufman, 2008).Therefore, in the present study, we assessed an in-depth computa-tional study on how GRP78 may be binding to NF-κB, thussuggesting that this type of interaction has implications in thepathological mechanisms of neurological disorders.
2. Materials and methods
2.1. Atomic coordinates and structures
The atomic coordinates of GRP78 (PDB: 3LDL) (Macias et al.,2011) were downloaded from the Protein Data Bank. Several PDBentries to GRP78 are available, but the PDB entry 3LDL wasselected due to its resolution (2.3 Å) (Macias et al., 2011), organism(human) and presence of interacting sites (Nucleotide bindingdomain) (Finn et al., 2010). The chain A of the homodimer GRP78was selected to the interaction and docking assessment due to thefact that Hsp proteins are active in monomer conditions (Aprileet al., 2013). As for NF-κB (PDB: 1IKN) the atomic coordinates werealso downloaded and selected due to its resolution and thepresence of all the subunits of the protein, the chains D and A ofthe heterodimer NF-κB that corresponds to IκBα and p65 sub-units, respectively (Huxford et al., 1998). Although the chain A of1IKN belongs to mus-musculus, no significant difference with thehuman protein at structural and sequence level was observed.Water molecules and associated ligands were removed in bothGRP78 and NF-κB proteins by using Mollegro Virtual Dockerversion 5.5 and Pymol version 1.5. Subsequently, the edited
M.F. Ávila et al. / Journal of Theoretical Biology 345 (2014) 43–5144

molecules were energy minimized (GROMOS96 Forcefield) beforeperforming the docking simulations to avoid significant changes inthe surface of the molecules and the minimum of energy.
2.2. Protein domain description and protein interactions
The description of most relevant domains found in the GRP78 andNF-κB was assessed by servers PFAM, CDD-Blast, PP interactionprediction, and STRING. Pfam is a collection of multiple protein-sequence alignments and HMMs, and provides a good repository ofmodels for identifying protein families, domains and repeats. Thereare two parts to the Pfam database: Pfam A, a set of manually curatedand annotated models; and PfamB, which has higher coverage, but isfully automated (with no manual curation). Pfam B HMMs arecreated from alignments generated by ProDom in the automaticclustering of the protein sequences in SWISS-PROT and TrEMBL.
2.2.1. CDD-blastCD-Search is NCBI0s interface to searching the Conserved
Domain Database with protein query sequences. It uses RPS-BLAST,a variant of PSI-BLAST, to quickly scan a set of pre-calculatedposition-specific scoring matrices (PSSMs) with a protein query(Marchler-Bauer et al., 2013).
2.2.2. Protein–protein interactions prediction (STRING)A search was carried out using the protein sequence of GRP78 and
NF-κB in the Search Tool for the Retrieval of Interacting Genes/Proteins(STRING 9.0). STRING 9.0 database allowed the identification ofpredicted and known protein interactions, encompassing physicaland functional associations (Szklarczyk et al., 2011).
2.2.3. Prediction of interacting sitesThe interacting sites were assessed using Consurf server to
predict the conserved regions of GRP78 chain A and NF-κB chainsD and A. Subsequently, the selection of possible interacting siteswas evaluated with the server tool Predict-Protein, assessingsolvent accessible surface, sequence based protein–protein bind-ing, and with the Detect Cavities subroutine of Mollegro virtualdocker version 5.5. Interacting sites were assessed using Q-sitefinder, a server that uses an energy-based method for the predic-tion of protein–ligand binding sites. It works by binding hydro-phobic (CH3) probes to the protein, and finding clusters of probeswith the most favorable binding energy (Laurie and Jackson,2005). The advantage of this is that putative binding sites areidentified as closely as possible to the actual binding site. It usesthe interaction energy between the protein and a simple Van derWaals probe to locate energetically binding sites. Q-site finderranks 10 possibilities of the most energetically favorable bindingsites, where those ranked at the top are the most favorable ones(Laurie and Jackson, 2005).
2.3. Docking (ClusPro)
The docking algorithms of ClusPro evaluate billion of putativecomplexes, retaining a preset number with favorable surfacecomplementarities (Comeau et al., 2004a, 2004b; Kozakov et al.,2006, 2010). A filtering method is then applied to this set ofstructures, selecting those with good electrostatic and desolvationfree energies for further clustering. The algorithm rotates the firstprotein with 70.000 rotations. For each rotation, it translates theligand in x, y, z relative to the second protein on a grid. Subse-quently, the algorithm chooses the translation with the best scorefrom each rotation (Comeau et al., 2004a, 2004b; Kozakov et al.,2006, 2010). ClusPro implements an improved method in thealgorithm, which includes a novel Fast Fourier Transform (FFT)
docking method with pairwise potentials implemented in PIPER(Kozakov et al., 2006). PIPER is a major improvement relative toZDOCK that also includes a novel method of generating referencestates for molecular recognition potentials (Chuang et al., 2008)(Decoys as the Reference State or DARS), a clustering technique forthe detection of near native conformations (Kozakov et al., 2005),a method of eliminating some of the non-native clusters byanalyzing the stability of funnels in the free energy landscape(Kozakov et al., 2008), and a new medium range optimizationmethod using Semi-Definite programming based Underestimation(SDU) (Shen et al., 2008) with a subsequent and final minimizationstep. As a strategy to determine possible interactions betweenGRP78–NF-κB, directed dockings were performed between theGRP78 chain A–NF-κB chain A (GRP78 (A)–NF-κB (A)) and GRP78chain A–NF-κB chain D (GRP78 (A)–NF-κB (D)).
2.4. Docking validation by molecular dynamics simulation
The best structural and minimized model for the complexesGRP78 (A)–NF-κB (A) and GRP78 (A)–NF-κB (D) obtained from thedocking analysis was subjected to Molecular dynamics (MD) simula-tion to refine and validate the protein interface. MD simulations wereperformed using the GROMACS package v. 4.5.5 (Hess et al., 2008).The two complexes were inserted into SPC solvated cubic box, andthe system was neutralized by adding negatively charged Cl�
counter ions at random positions. All of the simulations were per-formed using the AMBER99 force field. The systems were submittedto 5000 steps of energy minimization using the steepest descentmethod with GROMACS program.
MD simulations were performed in the Gibbs ensemble at300 K and 1 atm using the Berendsen algorithm. The simulationswere conducted in two steps: equilibration and trajectory collec-tion. For the equilibration stage, 100 ps were performed with allatomic protein positions restrained. The second step was asimulation without restraints, performed for 20 ns. In both stagesof the simulation, a 2-fs time step was applied. The final con-formation obtained at the end of the MD simulation was used foridentifying specific interactions at the interface, computing inter-residue distances and other calculations.
2.5. Docking interface analysis and hot-spots
The interface of interactions was analyzed with LigPlotþ v1.4(Laskowski and Swindells, 2011), using the DIMPLOT subroutine. Thisprogram shows hydrophobic and hydrogen bonds interactionsbetween interacting residues. Electrostatic potential surface, interac-tions of the complexes were calculated using Pymol v1.5, additionallythe Accessible solvent Area change (ΔASA) was calculated using theGet_area subroutine of Pymol v1.5 (DeLano, 2012). Interface interac-tion was assessed with Proface server (Saha et al., 2006), and hot-spot determination was evaluated with the KFC Server, which usesthe Fast Atomic Density Evaluator (FADE) and the K-FADE classifica-tion (KFC2-A and KFC2-B; Zhu and Mitchell, 2011).
3. Results
3.1. Domain description and protein interactions
Pfam server and CDD-BLAST classified GRP78 as a Hsp70 familymember, and described 3 principal domains: (i) N-terminal ATPasedomain (binds ATP and hydrolyzes it to ADP, and drives conforma-tional changes in the two other domains); (ii) Substrate bindingdomain, which contains a groove with an affinity for neutral,hydrophobic amino acid residues, and interacts with peptides upto seven amino acids in length; and (iii) C-terminal domain – rich
M.F. Ávila et al. / Journal of Theoretical Biology 345 (2014) 43–51 45
in alpha helical structure, and regulates the substrate bindingdomain). Pfam also reported outer membrane protein (OmpH-like)domain, but the e-value was not significant. On the other hand,Pfam and CDD-BLAST retrieved a Rel homology domain (RHD) inthe NF-κB chain A-this domain is found in eukaryotic transcriptionfactors-, and the Ankyrin repeat domain in the D chain, whichcan mediate protein–protein interactions (Mosavi et al., 2004).In respect to STRING 9.0, the interacting sites were made usingQ-site finder, and results showed no physical and functionalinteractions between GRP78 and NF-κB based on experimentaland co-expression data.
3.2. The prediction of the interacting sites was consequent with theconserved domains
The resulting prediction of the interacting sites of the GRP78chain A and NF-κB chains A and D was consequent with conservedand functional regions. As a result, interacting amino acids werechosen to direct the docking between the GRP78–NF-κB chains.Table 1 shows the resulting interacting residues, while Table 2 themotifs and domains that were possibly participating in theseinteracting sites. Q-site finder server displayed 10 ligand bindinginteracting sites to each protein (GRP78 and NF-κB). For GRP78,most ligand binding sites were related with the nucleotide bindingdomain of the protein. The third-ranked binding interacting sitewas near to the place where GRP78 possibly interacts with NF-κB(in the chains A and D). In addition, for NF-κB, Q-site finder alsodisplayed 10 ligand binding sites, where the first three of rankedsites we found amino acids that are possibly interacting withGRP78, as for example ASN 212 and ARG 158.
3.3. Docking simulation by ClusPro reveals the potential models ofthe interactions
Selected chains of GRP78 and NF-κB were uploaded to ClusProserver for docking simulations. The server retrieved 29 models in
each case (GRP78 (A)–NF-κB (A) and GRP78 (A)–NF-κB (D)) of thedirected docking with the selected residues. The most favorablecomplex was chosen to analyze the electrostatic, hydrophobic andbalanced favored scores (Tables 3 and 4) and to carry out the MDsimulations. Other criteria to select models were given by Profaceserver that included area of surface contact, scores of the interac-tion sites and stability.
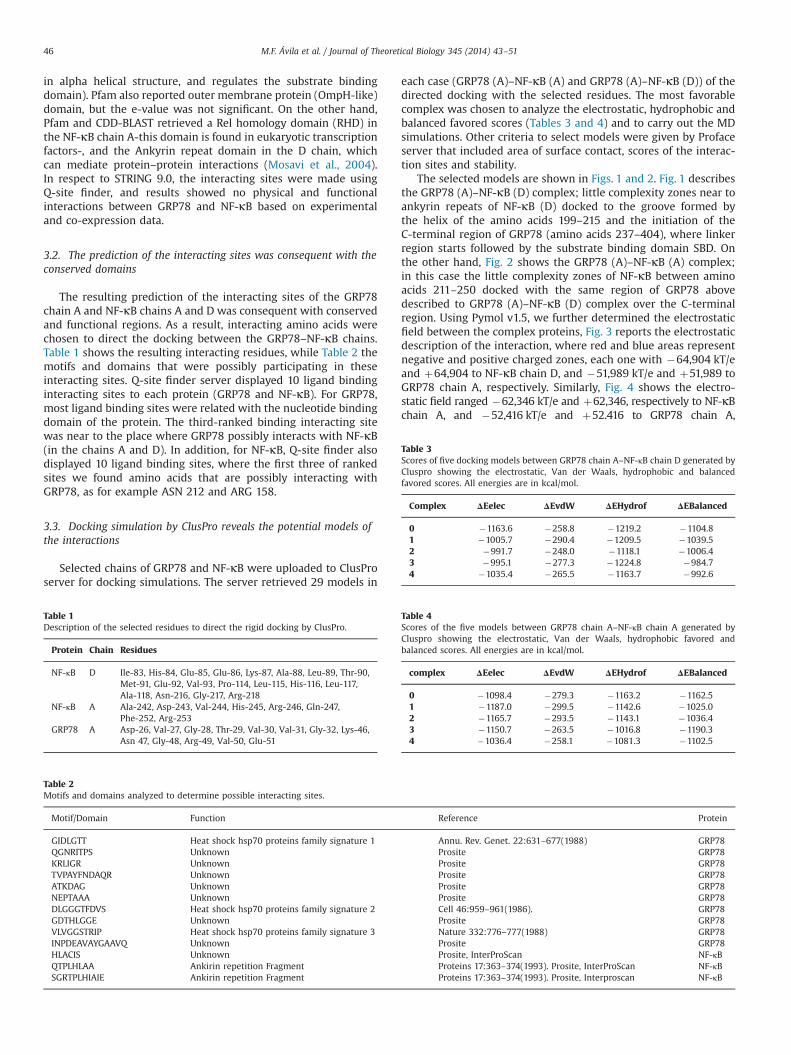
The selected models are shown in Figs. 1 and 2. Fig. 1 describesthe GRP78 (A)–NF-κB (D) complex; little complexity zones near toankyrin repeats of NF-κB (D) docked to the groove formed bythe helix of the amino acids 199–215 and the initiation of theC-terminal region of GRP78 (amino acids 237–404), where linkerregion starts followed by the substrate binding domain SBD. Onthe other hand, Fig. 2 shows the GRP78 (A)–NF-κB (A) complex;in this case the little complexity zones of NF-κB between aminoacids 211–250 docked with the same region of GRP78 abovedescribed to GRP78 (A)–NF-κB (D) complex over the C-terminalregion. Using Pymol v1.5, we further determined the electrostaticfield between the complex proteins, Fig. 3 reports the electrostaticdescription of the interaction, where red and blue areas representnegative and positive charged zones, each one with �64,904 kT/eand þ64,904 to NF-κB chain D, and �51,989 kT/e and þ51,989 toGRP78 chain A, respectively. Similarly, Fig. 4 shows the electro-static field ranged �62,346 kT/e and þ62,346, respectively to NF-κBchain A, and �52,416 kT/e and þ52.416 to GRP78 chain A,
Table 1Description of the selected residues to direct the rigid docking by ClusPro.
Protein Chain Residues
NF-κB D Ile-83, His-84, Glu-85, Glu-86, Lys-87, Ala-88, Leu-89, Thr-90,Met-91, Glu-92, Val-93, Pro-114, Leu-115, His-116, Leu-117,Ala-118, Asn-216, Gly-217, Arg-218
NF-κB A Ala-242, Asp-243, Val-244, His-245, Arg-246, Gln-247,Phe-252, Arg-253
GRP78 A Asp-26, Val-27, Gly-28, Thr-29, Val-30, Val-31, Gly-32, Lys-46,Asn 47, Gly-48, Arg-49, Val-50, Glu-51
Table 2Motifs and domains analyzed to determine possible interacting sites.
Motif/Domain Function Reference Protein
GIDLGTT Heat shock hsp70 proteins family signature 1 Annu. Rev. Genet. 22:631–677(1988) GRP78QGNRITPS Unknown Prosite GRP78KRLIGR Unknown Prosite GRP78TVPAYFNDAQR Unknown Prosite GRP78ATKDAG Unknown Prosite GRP78NEPTAAA Unknown Prosite GRP78DLGGGTFDVS Heat shock hsp70 proteins family signature 2 Cell 46:959–961(1986). GRP78GDTHLGGE Unknown Prosite GRP78VLVGGSTRIP Heat shock hsp70 proteins family signature 3 Nature 332:776–777(1988) GRP78INPDEAVAYGAAVQ Unknown Prosite GRP78HLACIS Unknown Prosite, InterProScan NF-κBQTPLHLAA Ankirin repetition Fragment Proteins 17:363–374(1993). Prosite, InterProScan NF-κBSGRTPLHIAIE Ankirin repetition Fragment Proteins 17:363–374(1993). Prosite, Interproscan NF-κB
Table 4Scores of the five models between GRP78 chain A–NF-κB chain A generated byCluspro showing the electrostatic, Van der Waals, hydrophobic favored andbalanced scores. All energies are in kcal/mol.
complex ΔEelec ΔEvdW ΔEHydrof ΔEBalanced
0 �1098.4 �279.3 �1163.2 �1162.51 �1187.0 �299.5 �1142.6 �1025.02 �1165.7 �293.5 �1143.1 �1036.43 �1150.7 �263.5 �1016.8 �1190.34 �1036.4 �258.1 �1081.3 �1102.5
Table 3Scores of five docking models between GRP78 chain A–NF-κB chain D generated byCluspro showing the electrostatic, Van der Waals, hydrophobic and balancedfavored scores. All energies are in kcal/mol.
Complex ΔEelec ΔEvdW ΔEHydrof ΔEBalanced
0 �1163.6 �258.8 �1219.2 �1104.81 �1005.7 �290.4 �1209.5 �1039.52 �991.7 �248.0 �1118.1 �1006.43 �995.1 �277.3 �1224.8 �984.74 �1035.4 �265.5 �1163.7 �992.6
M.F. Ávila et al. / Journal of Theoretical Biology 345 (2014) 43–5146
respectively, when in complex. These data demonstrated an electro-static favored complementarity in each of one complex.
3.4. Description of the interaction interface GRP78–NF-κB
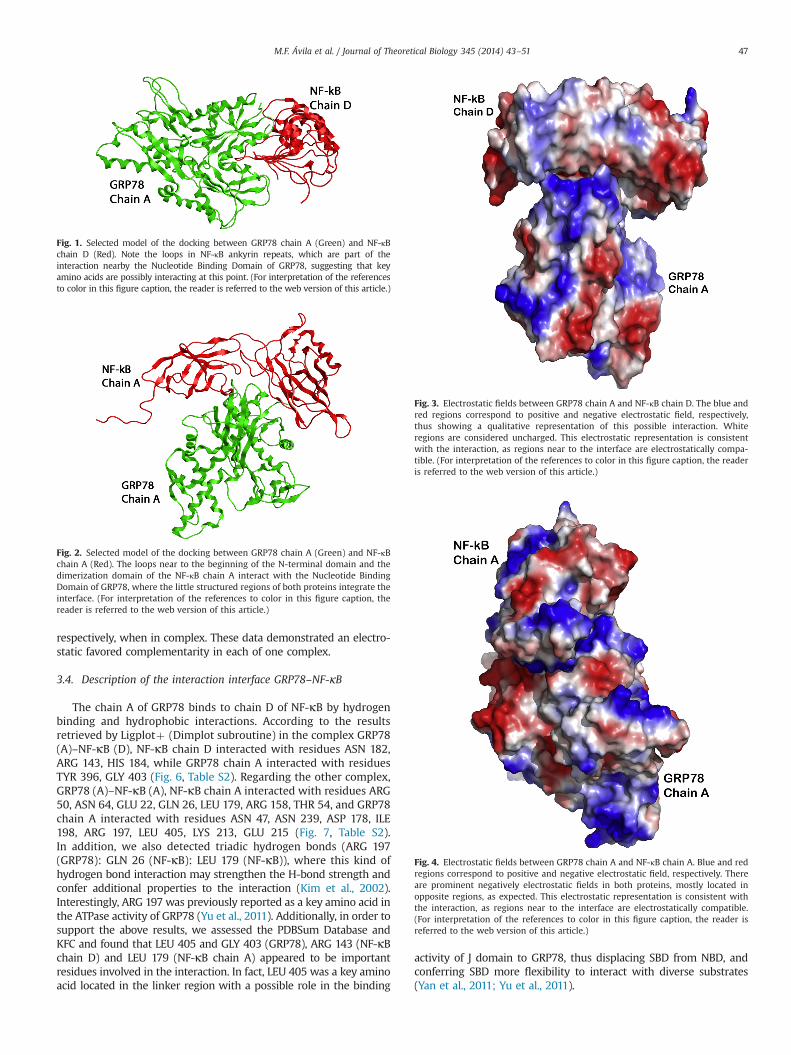
The chain A of GRP78 binds to chain D of NF-κB by hydrogenbinding and hydrophobic interactions. According to the resultsretrieved by Ligplotþ (Dimplot subroutine) in the complex GRP78(A)–NF-κB (D), NF-κB chain D interacted with residues ASN 182,ARG 143, HIS 184, while GRP78 chain A interacted with residuesTYR 396, GLY 403 (Fig. 6, Table S2). Regarding the other complex,GRP78 (A)–NF-κB (A), NF-κB chain A interacted with residues ARG50, ASN 64, GLU 22, GLN 26, LEU 179, ARG 158, THR 54, and GRP78chain A interacted with residues ASN 47, ASN 239, ASP 178, ILE198, ARG 197, LEU 405, LYS 213, GLU 215 (Fig. 7, Table S2).In addition, we also detected triadic hydrogen bonds (ARG 197(GRP78): GLN 26 (NF-κB): LEU 179 (NF-κB)), where this kind ofhydrogen bond interaction may strengthen the H-bond strength andconfer additional properties to the interaction (Kim et al., 2002).Interestingly, ARG 197 was previously reported as a key amino acid inthe ATPase activity of GRP78 (Yu et al., 2011). Additionally, in order tosupport the above results, we assessed the PDBSum Database andKFC and found that LEU 405 and GLY 403 (GRP78), ARG 143 (NF-κBchain D) and LEU 179 (NF-κB chain A) appeared to be importantresidues involved in the interaction. In fact, LEU 405 was a key aminoacid located in the linker region with a possible role in the binding
activity of J domain to GRP78, thus displacing SBD from NBD, andconferring SBD more flexibility to interact with diverse substrates(Yan et al., 2011; Yu et al., 2011).
Fig. 1. Selected model of the docking between GRP78 chain A (Green) and NF-κBchain D (Red). Note the loops in NF-κB ankyrin repeats, which are part of theinteraction nearby the Nucleotide Binding Domain of GRP78, suggesting that keyamino acids are possibly interacting at this point. (For interpretation of the referencesto color in this figure caption, the reader is referred to the web version of this article.)
Fig. 2. Selected model of the docking between GRP78 chain A (Green) and NF-κBchain A (Red). The loops near to the beginning of the N-terminal domain and thedimerization domain of the NF-κB chain A interact with the Nucleotide BindingDomain of GRP78, where the little structured regions of both proteins integrate theinterface. (For interpretation of the references to color in this figure caption, thereader is referred to the web version of this article.)
Fig. 3. Electrostatic fields between GRP78 chain A and NF-κB chain D. The blue andred regions correspond to positive and negative electrostatic field, respectively,thus showing a qualitative representation of this possible interaction. Whiteregions are considered uncharged. This electrostatic representation is consistentwith the interaction, as regions near to the interface are electrostatically compa-tible. (For interpretation of the references to color in this figure caption, the readeris referred to the web version of this article.)
Fig. 4. Electrostatic fields between GRP78 chain A and NF-κB chain A. Blue and redregions correspond to positive and negative electrostatic field, respectively. Thereare prominent negatively electrostatic fields in both proteins, mostly located inopposite regions, as expected. This electrostatic representation is consistent withthe interaction, as regions near to the interface are electrostatically compatible.(For interpretation of the references to color in this figure caption, the reader isreferred to the web version of this article.)
M.F. Ávila et al. / Journal of Theoretical Biology 345 (2014) 43–51 47
3.5. Molecular dynamics validation
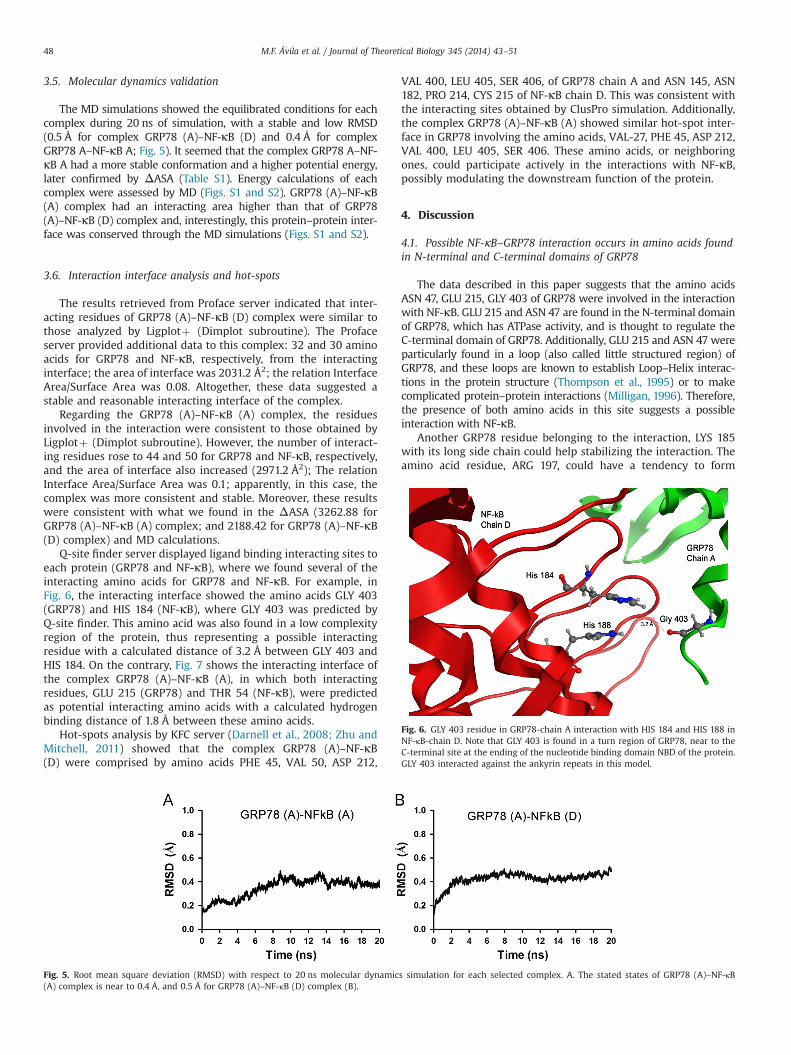
The MD simulations showed the equilibrated conditions for eachcomplex during 20 ns of simulation, with a stable and low RMSD(0.5 Å for complex GRP78 (A)–NF-κB (D) and 0.4 Å for complexGRP78 A–NF-κB A; Fig. 5). It seemed that the complex GRP78 A–NF-κB A had a more stable conformation and a higher potential energy,later confirmed by ΔASA (Table S1). Energy calculations of eachcomplex were assessed by MD (Figs. S1 and S2). GRP78 (A)–NF-κB(A) complex had an interacting area higher than that of GRP78(A)–NF-κB (D) complex and, interestingly, this protein–protein inter-face was conserved through the MD simulations (Figs. S1 and S2).
3.6. Interaction interface analysis and hot-spots
The results retrieved from Proface server indicated that inter-acting residues of GRP78 (A)–NF-κB (D) complex were similar tothose analyzed by Ligplotþ (Dimplot subroutine). The Profaceserver provided additional data to this complex: 32 and 30 aminoacids for GRP78 and NF-κB, respectively, from the interactinginterface; the area of interface was 2031.2 Å2; the relation InterfaceArea/Surface Area was 0.08. Altogether, these data suggested astable and reasonable interacting interface of the complex.
Regarding the GRP78 (A)–NF-κB (A) complex, the residuesinvolved in the interaction were consistent to those obtained byLigplotþ (Dimplot subroutine). However, the number of interact-ing residues rose to 44 and 50 for GRP78 and NF-κB, respectively,and the area of interface also increased (2971.2 Å2); The relationInterface Area/Surface Area was 0.1; apparently, in this case, thecomplex was more consistent and stable. Moreover, these resultswere consistent with what we found in the ΔASA (3262.88 forGRP78 (A)–NF-κB (A) complex; and 2188.42 for GRP78 (A)–NF-κB(D) complex) and MD calculations.
Q-site finder server displayed ligand binding interacting sites toeach protein (GRP78 and NF-κB), where we found several of theinteracting amino acids for GRP78 and NF-κB. For example, inFig. 6, the interacting interface showed the amino acids GLY 403(GRP78) and HIS 184 (NF-κB), where GLY 403 was predicted byQ-site finder. This amino acid was also found in a low complexityregion of the protein, thus representing a possible interactingresidue with a calculated distance of 3.2 Å between GLY 403 andHIS 184. On the contrary, Fig. 7 shows the interacting interface ofthe complex GRP78 (A)–NF-κB (A), in which both interactingresidues, GLU 215 (GRP78) and THR 54 (NF-κB), were predictedas potential interacting amino acids with a calculated hydrogenbinding distance of 1.8 Å between these amino acids.
Hot-spots analysis by KFC server (Darnell et al., 2008; Zhu andMitchell, 2011) showed that the complex GRP78 (A)–NF-κB(D) were comprised by amino acids PHE 45, VAL 50, ASP 212,
VAL 400, LEU 405, SER 406, of GRP78 chain A and ASN 145, ASN182, PRO 214, CYS 215 of NF-κB chain D. This was consistent withthe interacting sites obtained by ClusPro simulation. Additionally,the complex GRP78 (A)–NF-κB (A) showed similar hot-spot inter-face in GRP78 involving the amino acids, VAL-27, PHE 45, ASP 212,VAL 400, LEU 405, SER 406. These amino acids, or neighboringones, could participate actively in the interactions with NF-κB,possibly modulating the downstream function of the protein.
4. Discussion
4.1. Possible NF-κB–GRP78 interaction occurs in amino acids foundin N-terminal and C-terminal domains of GRP78
The data described in this paper suggests that the amino acidsASN 47, GLU 215, GLY 403 of GRP78 were involved in the interactionwith NF-κB. GLU 215 and ASN 47 are found in the N-terminal domainof GRP78, which has ATPase activity, and is thought to regulate theC-terminal domain of GRP78. Additionally, GLU 215 and ASN 47 wereparticularly found in a loop (also called little structured region) ofGRP78, and these loops are known to establish Loop–Helix interac-tions in the protein structure (Thompson et al., 1995) or to makecomplicated protein–protein interactions (Milligan, 1996). Therefore,the presence of both amino acids in this site suggests a possibleinteraction with NF-κB.
Another GRP78 residue belonging to the interaction, LYS 185with its long side chain could help stabilizing the interaction. Theamino acid residue, ARG 197, could have a tendency to form
Fig. 5. Root mean square deviation (RMSD) with respect to 20 ns molecular dynamics simulation for each selected complex. A. The stated states of GRP78 (A)–NF-κB(A) complex is near to 0.4 Å, and 0.5 Å for GRP78 (A)–NF-κB (D) complex (B).
Fig. 6. GLY 403 residue in GRP78-chain A interaction with HIS 184 and HIS 188 inNF-κB-chain D. Note that GLY 403 is found in a turn region of GRP78, near to theC-terminal site at the ending of the nucleotide binding domain NBD of the protein.GLY 403 interacted against the ankyrin repeats in this model.
M.F. Ávila et al. / Journal of Theoretical Biology 345 (2014) 43–5148

clusters with head to tail hydrogen bonding due to the presence ofthe three charged groups (Shukla and Trout, 2010). Lysine andArginine are usually found exposed at the protein surface to makeproteins soluble and interact with the nucleic acids negativelycharged phosphate backbone (Shukla and Trout, 2010). The aspartatein the positions 178 and 212 is involved in the protein–proteininteraction, and is commonly found on protein surface, or buried inthe hydrophobic core, stabilizing the protein structure if ion pairs areformed, mostly with Arginine (Sokalingam et al., 2012). Other aminoacids integrate the Nucleotide Binding Domain (Thr37, Thr38, Asp391,Ser300, Ile368, Lys296) of GRP78 and are highly conserved through-out Hsp70 protein family (Macias et al., 2011). These amino acids fillthe positions near to the ligand (ATP), and are subjected to modula-tion (Macias et al., 2011); however, they do not make part of theinteraction interface, despite that many of them are near to thepossible interaction site, denoting possible biological function theymight be carrying out. These observations are in accordance with ourproposed model of interacting residues in GRP78–NF-κB crosstalk.On the other hand, it is possible that the amino acids GLY 403, VAL404 and LEU 405, found in the C-terminal domain of GRP78 alsointervene in the interacting complex and participate in the NBD-SBDdynamics in complex with NF-κB. It is known that asparaginestabilizes hydrogen bond interactions (Stapley and Doig, 1997) inthe protein and also in glycolisation (O0Connor and Imperiali, 1996).Here we found that GRP78 amino acids GLU 215 and GLY 403interacted with the A (THR 54) and D (HIS 184) chains of NF-κB.Interestingly, these physical interactions may modulate the activity ofboth GRP78 and NF-κB with its subsequent downstream functions.Other amino acids also participated in the complex, as describedearlier, and should be considered as important for further studies.
4.2. GRP78–NF-κB interactions represent a significant cellularfunctions modulation
NF-κB is a promiscuous interacting protein that regulatesmultiple key functions in the cell (Hayden and Ghosh, 2008;Pahl and Baeuerle, 1996). Currently, the knowledge of its interac-tions has led to the development of inhibitors that block theinitiation of inflammation and downstream cascades involvedwith apoptosis and autoimmune diseases and several cancer types(Ivanenkov et al., 2011). Overexpression of GRP78 indicates stressconditions in the cell and has been related as a response ofsurvival (Wang et al., 2009; Zhang and Zhang, 2010). In fact, theresistance to some chemotherapeutic schemes has been associatedto high levels of GRP78, therefore NF-κB and GRP78 (GRP78 is
considered a potent resolution molecule in inflammation process)constitute potential therapeutic molecules (Ivanenkov et al., 2011;Panayi et al., 2004; Shields et al., 2011).
NF-κB–GRP78 physical interactions are not completely eluci-dated, but the inference and analysis of interactions with proteinsthat belong to the same family can provide key information thatmay explain the results reported here. For example, Hsp70, whichhas more than 95% of homology with GRP78 as assessed by usingthe blastp tool, a close parent of GRP78 – also considered a Hsp70isoform located predominantly in the endoplasmic reticulum(Macias et al., 2011), interacts with NF-κB in the Rel/p65 regula-tory complex, as previously demonstrated by inmmunoprecipita-tion assays, though the domains involved in this interaction werenot described. In another study, the chaperone Hsp27 regulatesNF-κB activity, as demonstrated by experimental studies of Hsp27-IKK, evaluated by directed mutations in Ser32 of IκBα (down-stream target of IKK) (Guo et al., 2009). Additionally, Cheng andcols demonstrated that Hsp90 interacts with the IKK complex andregulates TNF alfa signaling (Chen et al., 2002), supporting the ideathat Hsp proteins have a key role in the regulation and inhibitionof NF-κB via the interaction with its subunits (Xiaoxia et al., 2002).
4.3. IkB alpha as a possible subunit regulated by GRP78
Previous reports suggest that the interacting site in IκBα withproteins, as p53, is happen to occur through the alpha domain-containing C-terminus (between amino acids 244–314; Chang,2002). Interestingly, these interactions are mediated by conditionsthat overexpress GRP78, like hypoxia and apoptotic stress (Chang,2002). Conversely, other study reports that the N-terminus regionof IκBα interacts with ANT in the region comprised betweenamino acids 2–72, later confirmed using the two-hybrid system inyeast (Bottero et al., 2001). The interacting site of IκBα varies fromthe C-terminus and N-terminus in the protein, and depends on thedownstream signaling of NF-κB. In the present paper, we showthat models of NF-κB–GRP78 complex contained some aminoacids that belonged to the reported interactions sites of IκBα,supporting the idea of one possible plausible interaction.
Other studies showed that damage and stress inductors, likenicotine, dioxycholate, hypoxia, stimulate the expression of GRP78,HSP70, NF-κB and others proteins simultaneously in a dosis depen-dent rate (Bernstein et al., 1999; Crowley-Weber et al., 2003). This isimportant because signal translation gives rise to proteins that caninteract and modulate cell response, and shall guide cell to theresolution of an inflammatory process or apoptosis. In this context,our results may contribute to elucidate the mechanism of interactionthat occurs in cell under stress and pathological conditions, thusmaking a connection-interaction between several vital functions ofthe cell and a possible and already proposed crosstalk between ERstress and inflammation pathways (Hotamisligil, 2010; Kim et al.,2013; Kolattukudy and Niu, 2012; Misra et al., 2006; Tam et al., 2012;Yamazaki et al., 2009; Zhang, 2010; Zhang and Kaufman, 2008).For example, NF-κB drives the synthesis of pro-inflammatory pro-teins, and that GRP78 may contribute to inflammatory resolution anddecrease ER stress. Although the models of GRP78–NF-κB presentedhere represent a great advance in our understanding of how GRP78may regulate NF-κB, it is necessary to address other molecularinteractions of NF-κB with IKK (an upstream activator, for example),to broaden up our knowledge into these different cellular paradigms.
Acknowledgments
This work was supported in part by grants PUJ IDs 4327, 5024and 4367 to GEB.
Fig. 7. GLU 215 residue in GRP78-chain A interaction with THR 54 in NF-κB-chainA. In this interaction, amino acids were found in turn regions of GRP78 and NF-κB.The calculated distance of the hydrophobic interaction was 1.8 Å.
M.F. Ávila et al. / Journal of Theoretical Biology 345 (2014) 43–51 49

Appendix A. Supplementary material
Supplementary data associated with this article can be found inthe online version at http://dx.doi.org/10.1016/j.jtbi.2013.12.010.
References
Aprile, F., Dhulesia, A., Stengel, F., Roodveldt, C., Benesch, J., Tortora, P., Robinson, C.,Salvatella, X., Dobson, C., Cremades, N., 2013. Hsp70 oligomerization is mediatedby an interaction between the interdomain linker and the substrate-bindingdomain. PloS One 8, http://dx.doi.org/10.1371/journal.pone.0067961.
Beinke, S., Ley, S.C., 2004. Functions of NF-kappaB1 and NF-kappaB2 in immune cellbiology. Biochem. J. 382, 393–409, http://dx.doi.org/10.1042/BJ20040544.
Bernstein, H., Payne, C.M., Bernstein, C., Schneider, J., Beard, S.E., Crowley, C.L., 1999.Activation of the promoters of genes associated with DNA damage, oxidativestress, ER stress and protein malfolding by the bile salt, deoxycholate. Toxicol.Lett. 108, 37–46.
Bhattacharjee, G., Ahamed, J., Pedersen, B., El-Sheikh, A., Mackman, N., Ruf, W., Liu,C., Edgington, T., 2005. Regulation of tissue factor-mediated initiation of thecoagulation cascade by cell surface grp78. Arterioscler. Thromb. Vasc. Biol. 25,1737–1743, http://dx.doi.org/10.1161/01.atv.0000173419.31242.56.
Bottero, V., Rossi, F., Samson, M., Mari, M., Hofman, P., Peyron, J.F., 2001. Ikappab-alpha, the NF-kappa B inhibitory subunit, interacts with ANT, the mitochon-drial ATP/ADP translocator. J. Biol. Chem. 276, 21317–21324, http://dx.doi.org/10.1074/jbc.M005850200.
Bouwmeester, T., Bauch, A., Ruffner, H., Angrand, P.O., Bergamini, G., Croughton, K.,Cruciat, C., Eberhard, D., Gagneur, J., Ghidelli, S., Hopf, C., Huhse, B., Mangano,R., Michon, A.M., Schirle, M., Schlegl, J., Schwab, M., Stein, M.A., Bauer, A., Casari,G., Drewes, G., Gavin, A.C., Jackson, D.B., Joberty, G., Neubauer, G., Rick, J.,Kuster, B., Superti-Furga, G., 2004. A physical and functional map of the humanTNF-alpha/NF-kappa B signal transduction pathway. Nat. Cell Biol. 6, 97–105,http://dx.doi.org/10.1038/ncb1086.
Burikhanov, R., Shrestha-Bhattarai, T., Qiu, S., Shukla, N., Hebbar, N., Lele, S.,Horbinski, C., Rangnekar, V., 2013. Novel mechanism of apoptosis resistancein cancer mediated by extracellular PAR-4. Cancer Res. 73, 1011–1019, http://dx.doi.org/10.1158/0008-5472.can-12-3212.
Chandel, N.S., Trzyna, W.C., McClintock, D.S., Schumacker, P.T., 2000. Role ofoxidants in NF-kappa B activation and TNF-alpha gene transcription inducedby hypoxia and endotoxin. J. Immunol. 165, 1013–1021.
Chang, N.S., 2002. The non-ankyrin C terminus of Ikappa Balpha physically interactswith p53 in vivo and dissociates in response to apoptotic stress, hypoxia, DNAdamage, and transforming growth factor-beta 1-mediated growth suppression.J. Biol. Chem. 277, 10323–10331, http://dx.doi.org/10.1074/jbc.M106607200.
Chen, G., Cao, P., Goeddel, D.V., 2002. TNF-induced recruitment and activation ofthe IKK complex require Cdc37 and Hsp90. Mol. Cell 9, 401–410.
Chen, L., Gao, X., 2002. Neuronal apoptosis induced by endoplasmic reticulumstress. Neurochem. Res. 27, 891–898.
Chevalier, M., Rhee, H., Elguindi, E., Blond, S., 2000. Interaction of murine BiP/GRP78 with the DnaJ homologue MTJ1. J.f Biol. Chem. 275, 19620–19627, http://dx.doi.org/10.1074/jbc.M001333200.
Chu, A.J., 2005. Tissue factor mediates inflammation. Arch. Biochem. Biophys. 440,123–132, http://dx.doi.org/10.1016/j.abb.2005.06.005.
Chuang, G.Y., Kozakov, D., Brenke, R., Comeau, S.R., Vajda, S., 2008. DARS (decoys asthe reference state) potentials for protein–protein docking. Biophys. J. 95,4217–4227, http://dx.doi.org/10.1529/biophysj.108.135814.
Coe, H., Michalak, M., 2009. Calcium binding chaperones of the endoplasmicreticulum. Gen. Physiol. Biophys., F96–F103 (28 Spec No Focus).
Comeau, S.R., Gatchell, D.W., Vajda, S., Camacho, C.J., 2004a. ClusPro: an automateddocking and discrimination method for the prediction of protein complexes.Bioinformatics 20, 45–50.
Comeau, S.R., Gatchell, D.W., Vajda, S., Camacho, C.J., 2004b. ClusPro: a fullyautomated algorithm for protein-protein docking. Nucl. Acids Res. 32, W96–-W99, http://dx.doi.org/10.1093/nar/gkh354.
Crowley-Weber, C.L., Dvorakova, K., Crowley, C., Bernstein, H., Bernstein, C.,Garewal, H., Payne, C.M., 2003. Nicotine increases oxidative stress, activatesNF-kappaB and GRP78, induces apoptosis and sensitizes cells to genotoxic/xenobiotic stresses by a multiple stress inducer, deoxycholate: relevance tocolon carcinogenesis. Chem. Biol. Interact. 145, 53–66.
Darnell, S.J., LeGault, L., Mitchell, J.C., 2008. KFC Server: interactive forecasting ofprotein interaction hot spots. Nucl. Acids Res. 36, W265–W269, http://dx.doi.org/10.1093/nar/gkn346.
DeLano, W.L., 2012. The PyMOL Molecular Graphics System. DeLano Scientific LLC,San Carlos, CA, USA.
Delie, F., Petignat, P., Cohen, M., 2012. GRP78 protein expression in ovarian cancerpatients and perspectives for a drug-targeting approach. J. Oncol. 2012, 468615,http://dx.doi.org/10.1155/2012/468615.
Dodd, S.L., Hain, B., Senf, S.M., Judge, A.R., 2009. Hsp27 inhibits IKKbeta-inducedNF-kappaB activity and skeletal muscle atrophy. FASEB J. 23, 3415–3423, http://dx.doi.org/10.1096/fj.08-124602.
Dong, D., Stapleton, C., Luo, B., Xiong, S., Ye, W., Zhang, Y., Jhaveri, N., Zhu, G., Ye, R.,Liu, Z., Bruhn, K.W., Craft, N., Groshen, S., Hofman, F.M., Lee, A.S., 2011. A criticalrole for GRP78/BiP in the tumor microenvironment for neovascularization
during tumor growth and metastasis. Cancer Res. 71, 2848–2857, http://dx.doi.org/10.1158/0008-5472.CAN-10-3151.
Eddleston, M., de la Torre, J.C., Oldstone, M.B., Loskutoff, D.J., Edgington, T.S.,Mackman, N., 1993. Astrocytes are the primary source of tissue factor in themurine central nervous system. A role for astrocytes in cerebral hemostasis. J.Clin. Invest. 92, 349–358, http://dx.doi.org/10.1172/JCI116573.
Finn, R.D., Mistry, J., Tate, J., Coggill, P., Heger, A., Pollington, J.E., Gavin, O.L.,Gunasekaran, P., Ceric, G., Forslund, K., Holm, L., Sonnhammer, E.L., Eddy, S.R.,Bateman, A., 2010. The Pfam protein families database. Nucl. Acids Res. 38,D211–D222, http://dx.doi.org/10.1093/nar/gkp985.
Gilmore, T.D., 1990. NF-kappa B, KBF1, dorsal, and related matters. Cell 62, 841–843.Gonzalez-Dunia, D., Eddleston, M., Mackman, N., Carbone, K., de la Torre, J.C., 1996.
Expression of tissue factor is increased in astrocytes within the central nervoussystem during persistent infection with borna disease virus. J. Virol. 70,5812–5820.
Gonzalez-Gronow, M., Selim, M.A., Papalas, J., Pizzo, S.V., 2009. GRP78: a multi-functional receptor on the cell surface. Antioxid. Redox Signal 11, 2299–2306,http://dx.doi.org/10.1089/ARS.2009.2568.
Guo, K., Kang, N.X., Li, Y., Sun, L., Gan, L., Cui, F.J., Gao, M.D., Liu, K.Y., 2009.Regulation of HSP27 on NF-kappaB pathway activation may be involved inmetastatic hepatocellular carcinoma cells apoptosis. BMC Cancer 9, 100, http://dx.doi.org/10.1186/1471-2407-9-100.
Guzhova, I.V., Darieva, Z.A., Melo, A.R., Margulis, B.A., 1997. Major stress proteinHsp70 interacts with NF-kB regulatory complex in human T-lymphoma cells.Cell Stress Chaperones 2, 132–139.
Hayden, M.S., Ghosh, S., 2008. Shared principles in NF-kappaB signaling. Cell 132,344–362, http://dx.doi.org/10.1016/j.cell.2008.01.020.
Hess, B., Kutzner, C., Spoel, D., Lindahl, E., 2008. GROMACS 4: algorithms for highlyefficient, load-balanced, and scalable molecular simulation. J. Chem. TheoryComput. 4, 435–447.
Hinz, M., Broemer, M., Arslan, S.C., Otto, A., Mueller, E.C., Dettmer, R., Scheidereit, C.,2007. Signal responsiveness of IkappaB kinases is determined by Cdc37-assisted transient interaction with Hsp90. J. Biol. Chem. 282, 32311–32319,http://dx.doi.org/10.1074/jbc.M705785200.
Hotamisligil, G., 2010. Endoplasmic reticulum stress and the inflammatory basis ofmetabolic disease. Cell 140, 900–917, http://dx.doi.org/10.1016/j.cell.2010.02.034.
Huxford, T., Huang, D., Malek, S., Ghosh, G., 1998. The crystal structure of theIkappaBalpha/NF-kappaB complex reveals mechanisms of NF-kappaB inactiva-tion. Cell 95, 759–770.
Ivanenkov, Y.A., Balakin, K.V., Lavrovsky, Y., 2011. Small molecule inhibitors of NF-kBand JAK/STAT signal transduction pathways as promising anti-inflammatorytherapeutics. Mini. Rev. Med. Chem. 11, 55–78.
Kammanadiminti, S.J., Chadee, K., 2006. Suppression of NF-kappaB activation byEntamoeba histolytica in intestinal epithelial cells is mediated by heat shockprotein 27. J. Biol. Chem. 281, 26112–26120, http://dx.doi.org/10.1074/jbc.M601988200.
Karin, M., Ben-Neriah, Y., 2000. Phosphorylation meets ubiquitination: the controlof NF-[kappa]B activity. Annu. Rev. Immunol. 18, 621–663, http://dx.doi.org/10.1146/annurev.immunol.18.1.621.
Kim, H., Jeong, J., Kim, S., Park, S., Chae, H., Lee, Y., 2013. Inhibition of endoplasmicreticulum stress alleviates lipopolysaccharide-induced lung inflammationthrough modulation of NF-κB/HIF-1α signaling pathway. Sci. Rep. 3, 1142,http://dx.doi.org/10.1038/srep01142.
Kim, K.S., Kim, D., Lee, J.Y., Tarakeshwar, P., Oh, K.S., 2002. Catalytic mechanism ofenzymes: preorganization, short strong hydrogen bond, and charge buffering.Biochemistry 41, 5300–5306.
Kim, R., Emi, M., Tanabe, K., Murakami, S., 2006. Role of the unfolded proteinresponse in cell death. Apoptosis: Int. J. Program. Cell Death 11, 5–13, http://dx.doi.org/10.1007/s10495-005-3088-0.
Kolattukudy, P., Niu, J., 2012. Inflammation, endoplasmic reticulum stress, autop-hagy, and the monocyte chemoattractant protein-1/CCR2 pathway. Circ. Res.110, 174–189, http://dx.doi.org/10.1161/circresaha.111.243212.
Kozakov, D., Schueler-Furman, O., Vajda, S., 2008. Discrimination of near-nativestructures in protein–protein docking by testing the stability of local minima.Proteins 72, 993–1004, http://dx.doi.org/10.1002/prot.21997.
Kozakov, D., Clodfelter, K.H., Vajda, S., Camacho, C.J., 2005. Optimal clustering fordetecting near-native conformations in protein docking. Biophys. J. 89, 867–-875, http://dx.doi.org/10.1529/biophysj.104.058768.
Kozakov, D., Brenke, R., Comeau, S.R., Vajda, S., 2006. PIPER: an FFT-based proteindocking program with pairwise potentials. Proteins 65, 392–406, http://dx.doi.org/10.1002/prot.21117.
Kozakov, D., Hall, D.R., Beglov, D., Brenke, R., Comeau, S.R., Shen, Y., Li, K., Zheng, J.,Vakili, P., Paschalidis, I., Vajda, S., 2010. Achieving reliability and high accuracy inautomated protein docking: ClusPro, PIPER, SDU, and stability analysis in CAPRIrounds 13-19. Proteins 78, 3124–3130, http://dx.doi.org/10.1002/prot.22835.
Laskowski, R.A., Swindells, M.B., 2011. LigPlotþ: multiple ligand–protein interac-tion diagrams for drug discovery. J. Chem. Inf. Model 51, 2778–2786, http://dx.doi.org/10.1021/ci200227u.
Laurie, A.T., Jackson, R.M., 2005. Q-SiteFinder: an energy-based method for theprediction of protein-ligand binding sites. Bioinformatics 21, 1908–1916, http://dx.doi.org/10.1093/bioinformatics/bti315.
Lee, E., Nichols, P., Spicer, D., Groshen, S., Yu, M., Lee, A., 2006. GRP78 as a novelpredictor of responsiveness to chemotherapy in breast cancer. Cancer Res. 66,7849–7853, http://dx.doi.org/10.1158/0008-5472.can-06-1660.
M.F. Ávila et al. / Journal of Theoretical Biology 345 (2014) 43–5150

Lee, J.-Y., Jung, K.-W., Kim, Y., 2008. Inhibitor design for human heat shock protein70 ATPase domain by pharmacophore-based in silico screening. Bull. KoreanChem. Soc. 29, 1717.
Lim, M.Y., Davis, N., Zhang, J.Y., Bose Jr., H.R., 1990. The v-rel oncogene product iscomplexed with cellular proteins including its proto-oncogene product andheat shock protein 70. Virology 175, 149–160.
Lin, W.-C., Chuang, Y.-C., Chang, Y.-S., Lai, M.-D., Teng, Y.-N., Su, I.-J., Wang, C.C., Lee, K.-H.,Hung, J.-H., 2012. Endoplasmic reticulum stress stimulates p53 expression throughNF-κB activation. PloS One 7, http://dx.doi.org/10.1371/journal.pone.0039120.
Liu, M., Spellberg, B., Phan, Q.T., Fu, Y., Lee, A.S., Edwards Jr., J.E., Filler, S.G., Ibrahim,A.S., 2010. The endothelial cell receptor GRP78 is required for mucormycosispathogenesis in diabetic mice. J. Clin. Invest. 120, 1914–1924, http://dx.doi.org/10.1172/JCI42164.
Macias, A.T., Williamson, D.S., Allen, N., Borgognoni, J., Clay, A., Daniels, Z., Dokurno,P., Drysdale, M.J., Francis, G.L., Graham, C.J., Howes, R., Matassova, N., Murray, J.B., Parsons, R., Shaw, T., Surgenor, A.E., Terry, L., Wang, Y., Wood, M., Massey, A.J., 2011. Adenosine-derived inhibitors of 78 kDa glucose regulated protein(Grp78) ATPase: insights into isoform selectivity. J. Med. Chem. 54, 4034–4041,http://dx.doi.org/10.1021/jm101625x.
Marchler-Bauer, A., Zheng, C., Chitsaz, F., Derbyshire, M.K., Geer, L.Y., Geer, R.C.,Gonzales, N.R., Gwadz, M., Hurwitz, D.I., Lanczycki, C.J., Lu, F., Lu, S., Marchler, G.H., Song, J.S., Thanki, N., Yamashita, R.A., Zhang, D., Bryant, S.H., 2013. CDD:conserved domains and protein three-dimensional structure. Nucl. Acids Res.41, D348–D352, http://dx.doi.org/10.1093/nar/gks1243.
Milligan, R.A., 1996. Protein–protein interactions in the rigor actomyosin complex.Proc. Natl. Acad. Sci. U.S.A. 93, 21–26.
Misra, U., Deedwania, R., Pizzo, S., 2006. Activation and cross-talk between Akt, NF-kappaB, and unfolded protein response signaling in 1-LN prostate cancer cellsconsequent to ligation of cell surface-associated GRP78. J. Biol. Chem. 281,13694–13707, http://dx.doi.org/10.1074/jbc.M511694200.
Misra, U.K., Pizzo, S.V., 2010. Ligation of cell surface GRP78 with antibody directedagainst the COOH-terminal domain of GRP78 suppresses Ras/MAPK and PI3-kinase/AKT signaling while promoting caspase activation in human prostatecancer cells. Cancer Biol. Ther. 9, 142–152.
Misra, U.K., Payne, S., Pizzo, S.V., 2011. Ligation of prostate cancer cell surface GRP78activates a proproliferative and antiapoptotic feedback loop: a role for secretedprostate-specific antigen. J. Biol. Chem. 286, 1248–1259, http://dx.doi.org/10.1074/jbc.M110.129767.
Mosavi, L.K., Cammett, T.J., Desrosiers, D.C., Peng, Z.Y., 2004. The ankyrin repeat asmolecular architecture for protein recognition. Protein Sci. 13, 1435–1448, http://dx.doi.org/10.1110/ps.03554604.
Nakajima, S., Hiramatsu, N., Hayakawa, K., Saito, Y., Kato, H., Huang, T., Yao, J., Paton,A., Paton, J., Kitamura, M., 2011. Selective abrogation of BiP/GRP78 bluntsactivation of NF-κB through the ATF6 branch of the UPR: involvement of C/EBPβ and mTOR-dependent dephosphorylation of Akt. Mol. Cell. Biol. 31,1710–1718, http://dx.doi.org/10.1128/mcb.00939-10.
O0Connor, S.E., Imperiali, B., 1996. Modulation of protein structure and function byasparagine-linked glycosylation. Chem. Biol. 3, 803–812.
Ouyang, Y.B., Lu, Y., Yue, S., Giffard, R.G., 2012a. miR-181 targets multiple Bcl-2family members and influences apoptosis and mitochondrial function in astro-cytes. Mitochondrion 12, 213–219, http://dx.doi.org/10.1016/j.mito.2011.09.001.
Ouyang, Y.B., Lu, Y., Yue, S., Xu, L.J., Xiong, X.X., White, R.E., Sun, X., Giffard, R.G.,2012b. miR-181 regulates GRP78 and influences outcome from cerebralischemia in vitro and in vivo. Neurobiol. Dis. 45, 555–563, http://dx.doi.org/10.1016/j.nbd.2011.09.012.
Pahl, H.L., Baeuerle, P.A., 1996. Activation of NF-kappa B by ER stress requires bothCa2þ and reactive oxygen intermediates as messengers. FEBS Lett. 392, 129–136.
Pahl, H.L., Baeuerle, P.A., 1997. The ER-overload response: activation of NF-kappa B.Trends Biochem. Sci. 22, 63–67.
Panayi, G.S., Corrigall, V.M., Henderson, B., 2004. Stress cytokines: pivotal proteinsin immune regulatory networks; opinion. Curr. Opin. Immunol. 16, 531–534,http://dx.doi.org/10.1016/j.coi.2004.05.017.
Park, K.J., Gaynor, R.B., Kwak, Y.T., 2003. Heat shock protein 27 association with the Ikappa B kinase complex regulates tumor necrosis factor alpha-induced NF-kappa B activation. J. Biol. Chem. 278, 35272–35278, http://dx.doi.org/10.1074/jbc.M305095200.
Pereira, C., 2013. Crosstalk between endoplasmic reticulum stress and proteinmisfolding in neurodegenerative diseases. ISRN Cell Biol., 2013, http://dx.doi.org/10.1155/2013/256404.
Petranka, J., Wright, G., Forbes, R.A., Murphy, E., 2001. Elevated calcium inpreneoplastic cells activates NF-kappa B and confers resistance to apoptosis.J. Biol. Chem. 276, 37102–37108, http://dx.doi.org/10.1074/jbc.M008448200.
Quinones, Q., de Ridder, G., Pizzo, S., 2008. GRP78: a chaperone with diverse rolesbeyond the endoplasmic reticulum. Histol. Histopathol. 23, 1409–1416.
Rao, R., Peel, A., Logvinova, A., del Rio, G., Hermel, E., Yokota, T., Goldsmith, P., Ellerby,L., Ellerby, H., Bredesen, D., 2002. Coupling endoplasmic reticulum stress to the celldeath program: role of the ER chaperone GRP78. FEBS Lett. 514, 122–128.
Rual, J.F., Venkatesan, K., Hao, T., Hirozane-Kishikawa, T., Dricot, A., Li, N., Berriz, G.F., Gibbons, F.D., Dreze, M., Ayivi-Guedehoussou, N., Klitgord, N., Simon, C.,Boxem, M., Milstein, S., Rosenberg, J., Goldberg, D.S., Zhang, L.V., Wong, S.L.,Franklin, G., Li, S., Albala, J.S., Lim, J., Fraughton, C., Llamosas, E., Cevik, S., Bex, C.,Lamesch, P., Sikorski, R.S., Vandenhaute, J., Zoghbi, H.Y., Smolyar, A., Bosak, S.,
Sequerra, M., Doucette-Stamm, L., Cusick, M.E., Hill, D.E., Roth, F.P., Vidal, M.,2005. Towards a proteome-scale map of the human protein-protein interactionnetwork. Nature 437, 1173–1178, http://dx.doi.org/10.1038/nature04209.
Saha, R.P., Bahadur, R.P., Pal, A., Mandal, S., Chakrabarti, P., 2006. ProFace: a serverfor the analysis of the physicochemical features of protein–protein interfaces.BMC Struct. Biol. 6, 11, http://dx.doi.org/10.1186/1472-6807-6-11.
Shen, Y., Paschalidis, I., Vakili, P., Vajda, S., 2008. Protein docking by the under-estimation of free energy funnels in the space of encounter complexes. PLoSComput. Biol. 4, e1000191, http://dx.doi.org/10.1371/journal.pcbi.1000191.
Shields, A.M., Panayi, G.S., Corrigall, V.M., 2011. Resolution-associated molecularpatterns (RAMP): RAMParts defending immunological homeostasis? Clin. Exp.Immunol. 165, 292–300, http://dx.doi.org/10.1111/j.1365-2249.2011.04433.x.
Shields, A.M., Panayi, G.S., Corrigall, V.M., 2012. A new-age for biologic therapies:long-term drug-free therapy with BiP? Front. Immunol. 3, 17, http://dx.doi.org/10.3389/fimmu.2012.00017.
Shukla, D., Trout, B.L., 2010. Interaction of arginine with proteins and the mechan-ism by which it inhibits aggregation. J. Phys. Chem. B 114, 13426–13438, http://dx.doi.org/10.1021/jp108399g.
Sigrist, C.J., de Castro, E., Cerutti, L., Cuche, B.A., Hulo, N., Bridge, A., Bougueleret, L.,Xenarios, I., 2013. New and continuing developments at PROSITE. Nucl. AcidsRes. 41, D344–D347, http://dx.doi.org/10.1093/nar/gks1067.
Sokalingam, S., Raghunathan, G., Soundrarajan, N., Lee, S.G., 2012. A study on theeffect of surface lysine to arginine mutagenesis on protein stability andstructure using green fluorescent protein. PLoS One 7, e40410, http://dx.doi.org/10.1371/journal.pone.0040410.
Stapley, B.J., Doig, A.J., 1997. Hydrogen bonding interactions between glutamineand asparagine in alpha-helical peptides. J. Mol. Biol. 272, 465–473, http://dx.doi.org/10.1006/jmbi.1997.1262.
Stetler, R.A., Gan, Y., Zhang, W., Liou, A.K., Gao, Y., Cao, G., Chen, J., 2010. Heat shockproteins: cellular and molecular mechanisms in the central nervous system. Prog.Neurobiol. 92, 184–211, http://dx.doi.org/10.1016/j.pneurobio.2010.05.002.
Swinney, D.C., Xu, Y.Z., Scarafia, L.E., Lee, I., Mak, A.Y., Gan, Q.F., Ramesha, C.S.,Mulkins, M.A., Dunn, J., So, O.Y., Biegel, T., Dinh, M., Volkel, P., Barnett, J.,Dalrymple, S.A., Lee, S., Huber, M., 2002. A small molecule ubiquitinationinhibitor blocks NF-kappa B-dependent cytokine expression in cells and rats. J.Biol. Chem. 277, 23573–23581, http://dx.doi.org/10.1074/jbc.M200842200.
Szklarczyk, D., Franceschini, A., Kuhn, M., Simonovic, M., Roth, A., Minguez, P., Doerks,T., Stark, M., Muller, J., Bork, P., Jensen, L.J., Mering, C.v., 2011. The STRING databasein 2011: functional interaction networks of proteins, globally integrated andscored. Nucl. Acids Res. 39, D561–D568, http://dx.doi.org/10.1093/nar/gkq973.
Tam, A., Mercado, E., Hoffmann, A., Niwa, M., 2012. ER stress activates NF-κB byintegrating functions of basal IKK activity, IRE1 and PERK. PloS One 7, http://dx.doi.org/10.1371/journal.pone.0045078.
Thompson, T.B., Chou, K.C., Zheng, C., 1995. Analysis of the loop–helix interaction inbundle motif protein structures. J. Protein Chem. 14, 559–566.
Tieri, P., Termanini, A., Bellavista, E., Salvioli, S., Capri, M., Franceschi, C., 2012.Charting the NF-κB pathway interactome map. PloS One 7, http://dx.doi.org/10.1371/journal.pone.0032678.
Wang, M., Wey, S., Zhang, Y., Ye, R., Lee, A.S., 2009. Role of the unfolded proteinresponse regulator GRP78/BiP in development, cancer, and neurological disorders.Antioxid. Redox Signal 11, 2307–2316, http://dx.doi.org/10.1089/ARS.2009.2485.
Wang, M., Ye, R., Barron, E., Baumeister, P., Mao, C., Luo, S., Fu, Y., Luo, B., Dubeau, L.,Hinton, D.R., Lee, A.S., 2010. Essential role of the unfolded protein responseregulator GRP78/BiP in protection from neuronal apoptosis. Cell Death Differ.17, 488–498, http://dx.doi.org/10.1038/cdd.2009.144.
Xiaoxia, W., Zheng-Hong, Q., Yan, L., Yumei, W., Xiannu, J., Thomas, N.C., Bennett, M.C., 2002. Prostaglandin A1 inhibits rotenone-induced apoptosis in SH-SY5Ycells. J. Neurochem., 83, http://dx.doi.org/10.1046/j.1471-4159.2002.01224.x.
Xie, L., Tiong, C.X., Bian, J.S., 2012. Hydrogen sulfide protects SH-SY5Y cells against6-hydroxydopamine-induced endoplasmic reticulum stress. Am. J. Physiol. Cell.Physiol. 303, C81–C91, http://dx.doi.org/10.1152/ajpcell.00281.2011.
Yamazaki, H., Hiramatsu, N., Hayakawa, K., Tagawa, Y., Okamura, M., Ogata, R.,Huang, T., Nakajima, S., Yao, J., Paton, A., Paton, J., Kitamura, M., 2009. Activationof the Akt-NF-kappaB pathway by subtilase cytotoxin through the ATF6 branchof the unfolded protein response. J. Immunol. 183, 1480–1487, http://dx.doi.org/10.4049/jimmunol.0900017 (Baltimore, Md.: 1950).
Yan, M., Li, J., Sha, B., 2011. Structural analysis of the Sil1-Bip complex reveals themechanism for Sil1 to function as a nucleotide-exchange factor. Biochem. J.438, 447–455, http://dx.doi.org/10.1042/bj20110500.
Yu, H., Li, S., Yang, C., Wei, M., Song, C., Zheng, Z., Gu, Y., Du, H., Zhang, J., Xia, N., 2011.Homology model and potential virus-capsid binding site of a putative HEV receptorGrp78. J. Mol. Model. 17, 987–995, http://dx.doi.org/10.1007/s00894-010-0794-5.
Zhang, K., 2010. Integration of ER stress, oxidative stress and the inflammatoryresponse in health and disease. Int. J. Clin. Exp. Med. 3, 33–40.
Zhang, K., Kaufman, R., 2008. From endoplasmic-reticulum stress to the inflamma-tory response. Nature 454, 455–462, http://dx.doi.org/10.1038/nature07203.
Zhang, L.H., Zhang, X., 2010. Roles of GRP78 in physiology and cancer. J. CellBiochem. 110, 1299–1305, http://dx.doi.org/10.1002/jcb.22679.
Zhu, X., Mitchell, J.C., 2011. KFC2: a knowledge-based hot spot prediction methodbased on interface solvation, atomic density, and plasticity features. Proteins79, 2671–2683, http://dx.doi.org/10.1002/prot.23094.
M.F. Ávila et al. / Journal of Theoretical Biology 345 (2014) 43–51 51




















