
High-Potential C112D/M121X (X = M, E, H, L) Pseudomonas aeruginosa Azurins
Upload
independentCategory
view
0download
0
Structural Changes and Differentially Expressed Genes inPseudomonas aeruginosa Exposed to Meropenem-CiprofloxacinCombination
Vera Lúcia Dias Siqueira,a,b Rosilene Fressatti Cardoso,b Katiany Rizzieri Caleffi-Ferracioli,b Regiane Bertin de Lima Scodro,b
Maria Aparecida Fernandez,c Adriana Fiorini,b Tania Ueda-Nakamura,a Benedito Prado Dias-Filho,a Celso Vataru Nakamuraa*
Postgraduate Program in Pharmaceutical Sciences,a Department of Clinical Analysis and Biomedicine,b and Department of Cellular Biology and Genetics,c State Universityof Maringa, Maringá, Paraná, Brazil
The effect of a meropenem-ciprofloxacin combination (MCC) on the susceptibility of multidrug-resistant (MDR) Pseudomonasaeruginosa (MRPA) clinical isolates was determined using checkerboard and time-kill curve techniques. Structural changes anddifferential gene expression that resulted from the synergistic action of the MCC against one of the P. aeruginosa isolates (1071-MRPA]) were evaluated using electron microscopy and representational difference analysis (RDA), respectively. The differen-tially expressed, SOS response-associated, and resistance-associated genes in 1071-MRPA exposed to meropenem, ciprofloxacin,and the MCC were monitored by quantitative PCR. The MCC was synergistic against 25% and 40.6% of MDR P. aeruginosa iso-lates as shown by the checkerboard and time-kill curves, respectively. The morphological and structural changes that resultedfrom the synergistic action of the MCC against 1071-MRPA were a summation of the effects observed with each antimicrobialalone. One exception included outer membrane vesicles, which were seen in a greater amount upon ciprofloxacin exposure butwere significantly inhibited upon MCC exposure. Cell wall- and DNA repair-associated genes were differentially expressed in1071-MRPA exposed to meropenem, ciprofloxacin, and the MCC. However, some of the RDA-detected, resistance-associated,and SOS response-associated genes were expressed at significantly lower levels in 1071-MRPA exposed to the MCC. The MCCmay be an alternative for the treatment of MDR P. aeruginosa. The effect of this antimicrobial combination may be not only theresult of a summation of the effects of meropenem and ciprofloxacin but also a result of differential action that likely inhibitsprotective mechanisms in the bacteria.
Pseudomonas aeruginosa is a highly successful opportunisticpathogen that displays intrinsic multidrug resistance and has a
great ability to acquire further resistance mechanisms (1, 2). Lim-ited classes of antibiotics can be used for the treatment of P. aerugi-nosa infection (3). The emergence of multidrug-resistant (MDR)P. aeruginosa (MRPA) has prompted the search for new agents oralternative therapeutics (1, 4, 5), including antimicrobial combi-nations (6–9).
Synergistic interactions between drugs have been investigatedto improve therapeutic results, decrease the potential toxicity ofantimicrobial agents, and prevent the emergence of bacterial re-sistance (6, 10, 11). Several studies have evaluated the effects of ameropenem-ciprofloxacin combination (MCC) in P. aeruginosa(12–14), including MDR isolates (8, 15). Despite some differencesin the results, likely attributable to variations in techniques andbacterial isolates, the MCC represents a promising antipseudo-monal therapeutic option (12–14). Although �-lactams and fluo-roquinolones represent classic antimicrobials with previouslyknown mechanisms of action, the molecular basis of synergismbetween these classes of drugs has not yet been elucidated.
Thus, because of high rates of resistance observed in BrazilianP. aeruginosa isolates (16) and the consequent possibility of usingantimicrobial combination therapy (17), the present study soughtto identify bacterial cell structure changes and differential tran-script results using electron microscopy (EM) and representa-tional difference analysis (RDA), respectively, in a MDR P. aerugi-nosa isolate exposed to a synergistic antimicrobial combination(MCC) selected from among MDR clinical isolates from a Brazil-ian university hospital.
MATERIALS AND METHODS
Bacterial strains. The isolates were selected after screening 75 P. aerugi-nosa isolates from several biological sources from patients admitted to auniversity hospital in northwestern Paraná, Brazil, from January 2007 toJuly 2009. A total of 32 genetically distinct MDR P. aeruginosa isolatesresistant to at least one of the two antimicrobials studied (meropenem andciprofloxacin) were tested. The resistance profiles and genetic relation-ships were previously determined (18). The selected isolates were stored intryptic soy broth (Difco Laboratories, Sparks, MD) with 15% glycerol at�80°C. P. aeruginosa ATCC 27853 was used as a control for antimicrobialactivity and the synergism tests.
Determination of MIC. The MICs of meropenem (lot no. 09405C-1;AstraZeneca, Cotia, Brazil) and ciprofloxacin (lot no. 0001396108; Sigma-Aldrich, Steinheim, Germany) were determined by microdilution usingcation-adjusted Mueller-Hinton broth (CAMHB; Difco) according to theguidelines of the Clinical and Laboratory Standards Institute (CLSI) (19),and categorical susceptibility was based on CLSI breakpoints (20). All ofthe determinations were performed in triplicate.
Received 29 November 2013 Returned for modification 17 January 2014Accepted 19 April 2014
Published ahead of print 5 May 2014
Address correspondence to Celso Vataru Nakamura, [email protected].
* Present address: Celso Vataru Nakamura, Laboratory of Technological Innovationin the Development of Drugs and Cosmetics, State University of Maringa,Maringá, Paraná, Brazil.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AAC.02584-13
July 2014 Volume 58 Number 7 Antimicrobial Agents and Chemotherapy p. 3957–3967 aac.asm.org 3957
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from

Checkerboard assay. The MCCs were assessed in microtiter platecheckerboard assays with CAMHB (21). Both drugs were tested alone andcombined at concentrations up to four times higher and lower than theMICs of the drugs tested alone. The final inoculum concentration of the P.aeruginosa isolates tested was approximately 5.0 � 105 CFU/ml, and theplates were incubated at 35°C for 18 h. The tests were performed in trip-licate. Fractional inhibitory concentrations indices (FICIs) were calcu-lated (21). Synergy was defined as FICI � 0.5, no interaction was definedas FICI � 0.5 and � 4, and antagonism was defined as FICI � 4 (22).
Time-kill curve assay. Time-kill curve assays for all of the P. aerugi-nosa isolates were performed in CAMHB. Meropenem, ciprofloxacin, andthe MCC were tested at concentrations of 0.5� and 0.25� MIC for eachisolate. All of the assays included a growth control with no antimicrobial.The final inoculum used was 105 CFU/ml. The antimicrobial and growthcontrol tubes were continuously shaken on an orbital incubator at 35°C.Aliquots (0.1 ml) were removed at 0, 3, 6, 9, 12, and 24 h and seriallydiluted in sterile saline solution to avoid antibiotic carryover. Afterward,20 �l of each dilution was immediately inoculated in tryptic soy agar(TSA; Difco) in triplicate. The plates were incubated at 35°C for 18 to 24 h,colony counts were performed, and the lowest limit of detection was 50CFU/ml. All of the time-kill assays were performed in duplicate. P. aerugi-nosa ATCC 27853 (susceptible to meropenem and ciprofloxacin) wastested as a control strain. Data from at least two independent experimentswere averaged and plotted as log10 CFU/ml versus time (h) for each timepoint over 24 h. Synergism was defined as a reduction � 102 CFU/ml withthe MCC compared with the most active single antibiotic at 24 h, with thenumber of surviving organisms in the presence of the combination being�2 log10 CFU/ml below the starting inoculum (21). Bactericidal activitywas defined as a reduction of �3 log10 CFU/ml in the colony count fromthe starting inoculum at 24 h (9).
SEM and TEM. A clinical isolate (1071-MRPA) for which the MCCshowed a synergistic action was selected for the evaluation of morpholog-ical and structural changes. The 1071-MRPA cells were exposed to 0.5�MIC of meropenem, ciprofloxacin, and the MCC for 3 and 12 h and fixedfor at least 2 h with 2.5% glutaraldehyde (Sigma)– 0.1 M cacodylate buffer(Electron Microscopy Science, Hatfield, PA) at 4°C. Subcultures on TSAwere made to ensure that no contaminating organisms were present. Priorto subculture, dilutions were performed to avoid antibiotic carryover. Theelectron microscopy experiments were performed in duplicate on differ-ent days. For scanning electron microscopy (SEM), the treated cells wereplaced on a glass support with poly-L-lysine (Sigma), dehydrated ingraded ethanol, subjected to critical-point drying in CO2, coated withgold, and observed in a Shimadzu SS-550 (Kyoto, Japan) scanning elec-tron microscope. An average of 30 to 50 microscopic fields in each samplewere selected by random scanning and photographed. For transmissionelectron microscopy (TEM), fixed cells in glutaraldehyde were postfixedfor 2 h at room temperature in 2.0% osmium tetroxide (EMS) plus 2.0%potassium ferrocyanide (Merck, Darmstadt, Germany), dehydrated in ac-etone, and embedded in EMbed resin (EMS). Ultrathin 60-nm-thick sec-tions were then prepared, stained with uranyl acetate (EMS) and leadcitrate (EMS), and observed in a JEM-1400 (JEOL, Tokyo, Japan) micro-scope that operated at 80 kV. An average of 200 fields were photographed.Two thousand cells were evaluated for the MCC and single antimicrobialtests at different times, and only cellular changes that were repeated in atleast 70% of the observed images were considered.
RNA isolation and cDNA synthesis. Overnight TSA (Difco) growthof 1071-MRPA was transferred to four vials that contained CAMHB(Difco), and the vials were incubated at 35°C to obtain growth with anoptical density at 600 nm (OD600) of 0.4. Meropenem, ciprofloxacin, andthe MCC were separately added at a final concentration of 16 �g/ml (cor-responding to 0.5� MIC). Three vials containing antimicrobial were re-ferred to as testers (tester 1 [T1], T2, and T3, respectively). A vial withoutantimicrobial was used as a control and referred to as a driver (D). Sixtyminutes after the addition of the antimicrobials, treated and control bac-terial cells were harvested at 4,700 rpm for 10 min and washed twice with
RNase-free ultrapure water. Total RNA was extracted from 1071-MRPAcells for each experimental condition (T1, T2, T3, and D) using an RNeasyminikit (Qiagen, Valencia, CA) according to the manufacturer’s instruc-tions. Quantification purity and quality assessments were performed witha Qubit2.0 fluorometer (Invitrogen, Carlsbad, CA) and on 1% agarosegels, respectively. Contaminating DNA was removed by treatment withRNase-free DNase I (Invitrogen) and subjected to conventional PCR toensure that all DNA was removed as assessed by agarose gel. First-strandcDNA synthesis was performed with Superscript III reverse transcriptase(RT) (Invitrogen) using 1 �g of DNase-treated total RNA, 0.8 �M oli-go(dT) primer (Integrated DNA Technologies, Coralville, IA), and 0.8�M Random Primer (Invitrogen). Sequentially, the second strand wassynthesized using Random Primer and Platinum Taq DNA polymeraseHigh Fidelity (Invitrogen) according to the manufacturer’s instructions.
cDNA RDA. Approximately 1 �g of T1, T2, T3, and D cDNAs wasdigested with Sau3AI (Promega, Madison, WI), and the RDAs for mero-penem (RDA1), ciprofloxacin (RDA2), and MCC (RDA3) were per-formed according to the method of Leite et al. (23), with modification, inrounds of subtractions and amplifications. Three tester-driver hybridiza-tions (1:10, 1:20, and 1:10 ratios) were performed consecutively for eachRDA condition. A reverse cDNA-RNA experiment was performed as acontrol under the same conditions as described above in which the testercultures were not exposed to the MCC and the driver cultures were ex-posed to MCC. The final RDA products were purified using a QIAquickPCR purification kit (Qiagen), cloned into pGEM-T-Easy vector (Pro-mega), and transformed into Escherichia coli DH5-� competent cells.Plasmidial DNA was extracted as previously described (24). Sequencingwas performed with universal M13 primers (Promega) and standard flu-orescence-labeling dye terminator protocols. The samples were loadedonto a MegaBACE 1000 DNA sequencer (GE Healthcare Life Science,Piscataway, NJ) for automated analysis. Expressed sequence tags (ESTs)were analyzed using the Phred, Crossmatch, and CAP3 (http://asparagin.cenargen.embrapa.br/phph/) programs. Sequences with at least 100 nu-cleotides and Phred quality � 20 were considered for further analysis. TheESTs were grouped in clusters represented by contigs and singlets. Thescreened sequences were compared to the GenBank (http://www.ncbi.nlm.nih.gov) nonredundant (nr) database from the National Center forBiotechnology Information (NCBI) using the BLASTX algorithm (25)with an E value cutoff at 10�5.
qPCR. To confirm and quantify the relative transcript levels of theselected RDA products, quantitative PCR (qPCR) was performed usingT1, T2, T3, and D cDNA and StepOnePlus real-time PCR systems (version2; Applied Biosystems, Carlsbad, CA). Transcripts detected by RDA1(meropenem exposure), RDA2 (ciprofloxacin exposure), and RDA3(MCC exposure) with redundancy above 10 were tested, including threegenes related to meropenem or ciprofloxacin resistance (mexA, ampC,and oprD) and one gene related to the SOS response (lexA). The primerswere designed using Primer Express software (version 3.0, Applied Bio-systems), with the exception of the resistance and lexA genes (Table 1).The primer concentrations were adjusted to provide amplification effi-ciencies of approximately 90% to 110% for all of the experiments. PCRwas performed using SYBR Select master mix (Applied Biosystems) with 1�l of cDNA and 400 nM each primer in a total volume of 10 �l. PCRthermal cycling was performed at 95°C for 10 min followed by 40 cycles of95°C for 15 s, 60°C for 20 s, and 72°C for 20 s. A melting curve analysis wasperformed to confirm a single PCR product. The cycle threshold (CT)values for the triplicate PCRs and duplicate biological assessments foreach cDNA sample were averaged, and then 2���CT values were calcu-lated (26). A negative control containing all of the reagents except cDNAwas also included. The rpsL gene was used to normalize all of the reactions.The P. aeruginosa antimicrobial growth-free cDNA driver (D) was used asa reference sample. The genes were considered up- or downregulatedwhen the amounts of the transcripts were at least 2 times higher or lowerthan those present in the reference sample (D) (27).
Siqueira et al.
3958 aac.asm.org Antimicrobial Agents and Chemotherapy
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from

Data analysis. Group comparisons (meropenem, ciprofloxacin, andMCC data determined by qPCR and transmission electron microscopy)were made using one-way analysis of variance (ANOVA) with SAS 9.0software (SAS OnlineDoc 9; SAS Institute, Cary, NC), followed by theTukey post hoc test. Values of P 0.01 were considered statistically sig-nificant.
RESULTSClinical isolate susceptibilities and combination assays. All ofthe P. aeruginosa isolates were MDR and collected from severalbiological sources from patients admitted in different hospitalunits (18). The MIC50 and MIC90 of meropenem and ciprofloxa-cin for P. aeruginosa were 4 and 32 �g/ml and 32 and 64 �g/ml,respectively. Seventeen of the 32 isolates (53.1%) were resistant tociprofloxacin. Five of the isolates (15.6%) were resistant to mero-penem, and 10 of the isolates (31.3%) were resistant to both anti-microbials (Table 2).
The checkerboard titrations yielded a synergistic FICI (�0.5)for eight (25.0%) of the isolates tested. For the remainder of theisolates, the antimicrobial combination displayed no interaction
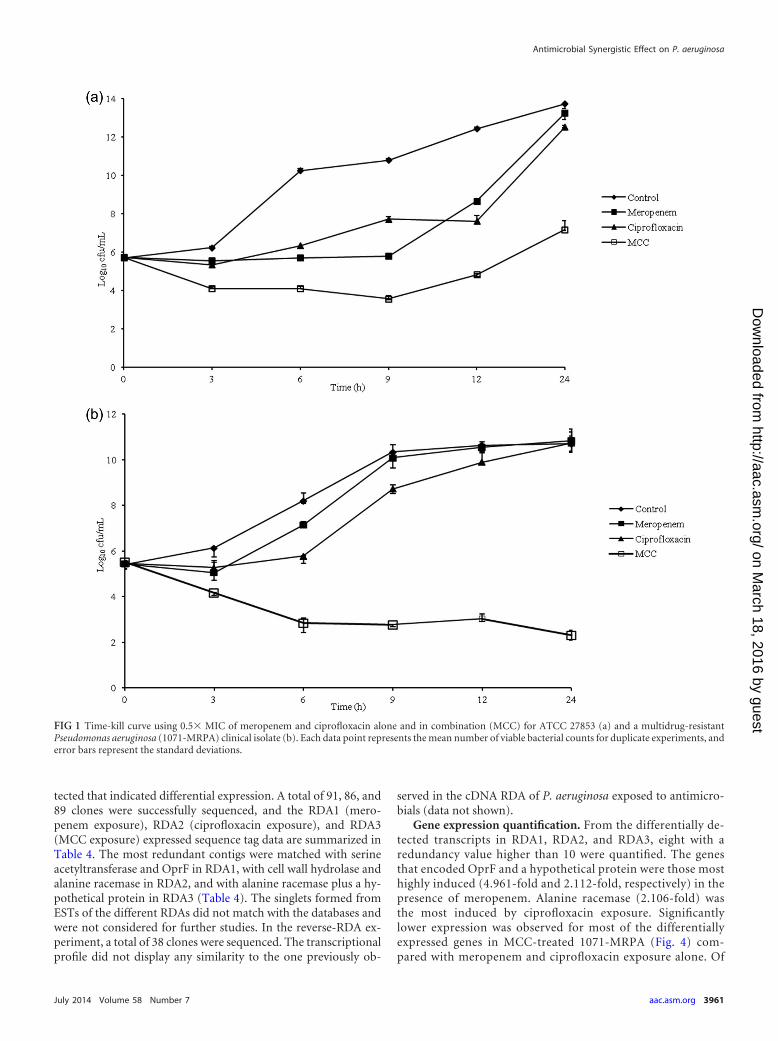
(Table 2). In all of the kill-curve assays, the viable counts in thecontrol with and without the antimicrobial agents tested at 0 hwere close to the initial inoculum count, thus ensuring that anantimicrobial carryover effect was avoided. The results of thetime-kill assays showed a �2-log10 decrease in viable counts forthe MCC at 0.5� MIC, regardless of the time of bacterial growth,in 31 (96.9%) of the isolates. This decrease was more apparentwith 12 h of exposure (87.5% of the isolates). However, synergismas previously defined (21) was observed in 13 (40.6%) of the iso-lates tested (Table 2). With combinations of 0.25� MIC plus 0.5�MIC for each antimicrobial and 0.25� MIC for both antimicro-bials, a �2-log10 reduction of viable counts was observed in 10(31.3%) of the isolates between 6 and 12 h of exposure and in 1isolate at 24 h of exposure (data not shown). Bactericidal activitywas observed in only four (12.5%) of the isolates with the MCC at0.5� MIC (Table 2). Representative time-kill curves for ATCCstrains and MDR clinical isolates using 0.5� MIC of the mero-penem, ciprofloxacin, and MCC are illustrated in Fig. 1a and b,respectively. The time-kill curve of the ATCC strain showed a
TABLE 1 Primer sequences used for real-time PCR analyses
Primer Primer sequence (5=–3=)a
GenBank accession no.or reference
OprF F—GCGGCTCGATCGGTTACTT NP_250468.1R—CGTCATGGTACTCACCGTAGGA
Serine acetyltransferase F—CCGTTCGCTCGCCACTAC ZP_15787657.1R—AAACATCTAAGTACCCTGAGGAAAAGA
Hydrolase F—CGAAATTCCTTGGTCGGGTAA ZP_10486838.1R—AGCGCCGCCATCGTTA
Alanine racemase F—CGGATCGGGTGGTTCTGTAT ZP_01680398.1R—GAGCCCCGGAGTACCTTTTATC
Uracil-DNA glycosylase F—CAGCCCCAGGATGTGATGA ZP_01682025.1R—TCATATCGACGGCGGTGTT
Hypothetical protein I F—TACCCAGAAGTGCCGAGCAT ZP_22178892.1R—CCTTTTTCACGACGGACGTT
Hypothetical protein II F—TTACGGCCAGGGCTACACA ZP_07663463.1R—CGGCTTGGCAACCCTTT
Hypothetical protein III F—GGATTAGCCCTTAAGCTTCATTGA YP_001949471.1R—CACTATGGCCGCACTTTCCA
rpsL F—GCAAGCGCATGGTCGACAAGA Dumas et al. (55)R—CGCTGTGCTCTTGCAGGTTGTGA
ampC F—CGGCTCGGTGAGCAAGACCTTC Dumas et al. (55)R—AGTCGCGGATCTGTGCCTGGTC
mexA F—CGACCAGGCCGTGAGCAAGCAGC Dumas et al. (55)R—GGAGACCTTCGCCGCGTTGTCGC
oprD F—ATCTACCGCACAAACGATGAAGG Dumas et al. (55)R—GCCGAAGCCGATATAATCAAACG
lexA F—GGATCAATCCCGCCTTCTTC Liu et al. (56)R—AAGCGTTTCACCGTGACCTC
a F, forward; R, reverse.
Antimicrobial Synergistic Effect on P. aeruginosa
July 2014 Volume 58 Number 7 aac.asm.org 3959
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from
�2-log10 reduction of viable counts between 9 and 24 h of expo-sure (Fig. 1). No reduction � 2 log10 CFU/ml below the startinginoculum was observed with 0.25� MIC against all 32 clinicalisolates (data not shown) or with 0.25� (data not shown) and0.5� MIC (Fig. 1) against the ATCC strain, indicating no syner-gism.
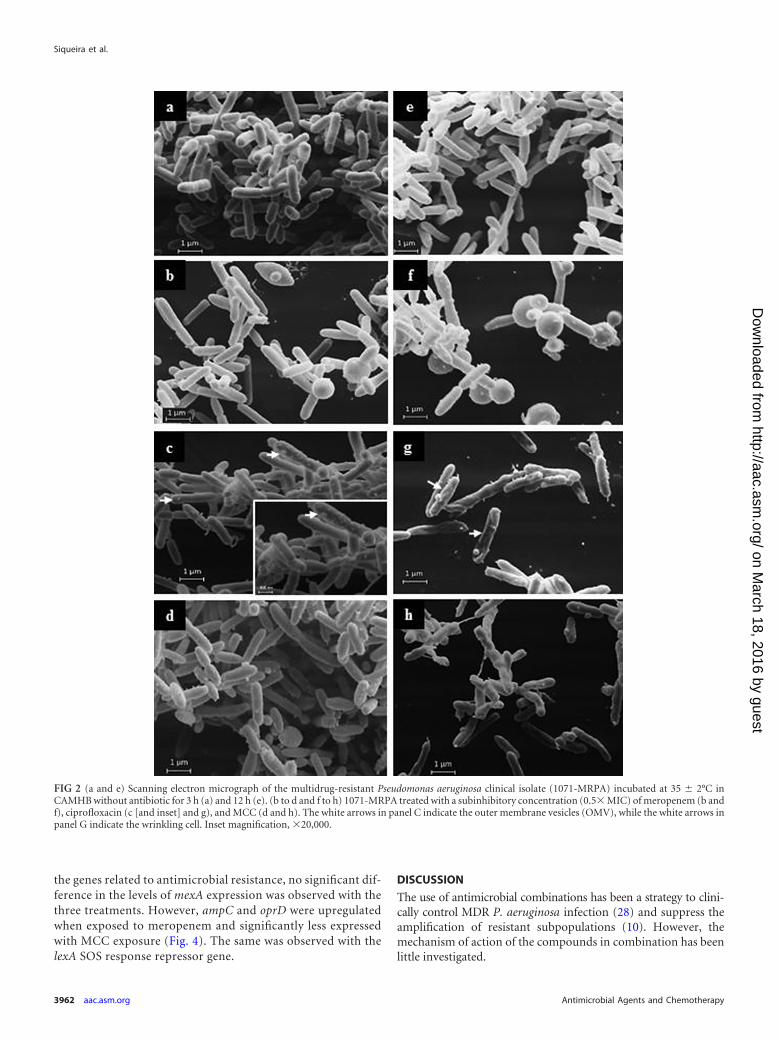
Morphological and structural changes. Because of the favor-able results from the time-kill curves and checkerboard assay,one MDR P. aeruginosa clinical isolate (1071-MRPA) was selectedfor electron microscopy assays after exposure to 0.5� MIC ofmeropenem, ciprofloxacin, and the MCC at 3 h (i.e., the start ofthe decrease in viable counts) and 12 h (i.e., the time with a greaterdecrease in viable counts). Scanning electron microscopy indi-cated that the effect of ciprofloxacin on cell morphology was lessevident than that seen with cells treated with meropenem. Round-ing and a wrinkled appearance were observed in 1071-MRPA cellsexposed to meropenem (Fig. 2b and f) and ciprofloxacin (Fig. 2cand g), respectively, mainly at 12 h of antimicrobial exposure. Thechanges caused by the MCC were observed mainly at 12 h of treat-ment and appeared to be a summation of the observed changes in1071-MRPA caused by meropenem and ciprofloxacin exposure
(i.e., rounding and cell wrinkling; Fig. 2d and h). Structures sim-ilar to those of outer membrane vesicles (OMVs) were observed inthe highest number in 1071-MRPA cells exposed to ciprofloxacinat 3 h (Fig. 2c, inset). Transmission electron microscopy was usedto observe and quantify changes inside and on the wall of 1071-MRPA cells treated with meropenem, ciprofloxacin, and theMCC. Rounding cells and cell wall disintegration were observed inmeropenem-exposed cells (Table 3 and Fig. 3b and f). A low elec-trodensity of areas without content was observed in ciprofloxacin-treated cells (Table 3 and Fig. 3c and g). Changes in MCC-treatedcells were a summation of the effects (cellular rounding and areaswithout content) observed with each antimicrobial alone (Table 3and Fig. 3d and h). Cellular changes were observed mainly at 12 hwith the different treatments. Outer membrane vesicles were ob-served in 1071-MRPA cells subjected to the three different treat-ments, but a significantly greater number of OMVs was observedin ciprofloxacin-treated cells at 3 h (Table 3 and Fig. 3c and inset).A significant decrease in the amount of OMVs was observed inMCC-exposed cells at 3 and 12 h (Table 3 and Fig. 3d and h).
Differentially expressed genes. After three rounds of subtrac-tive cDNA RDA hybridization, several DNA fragments were de-
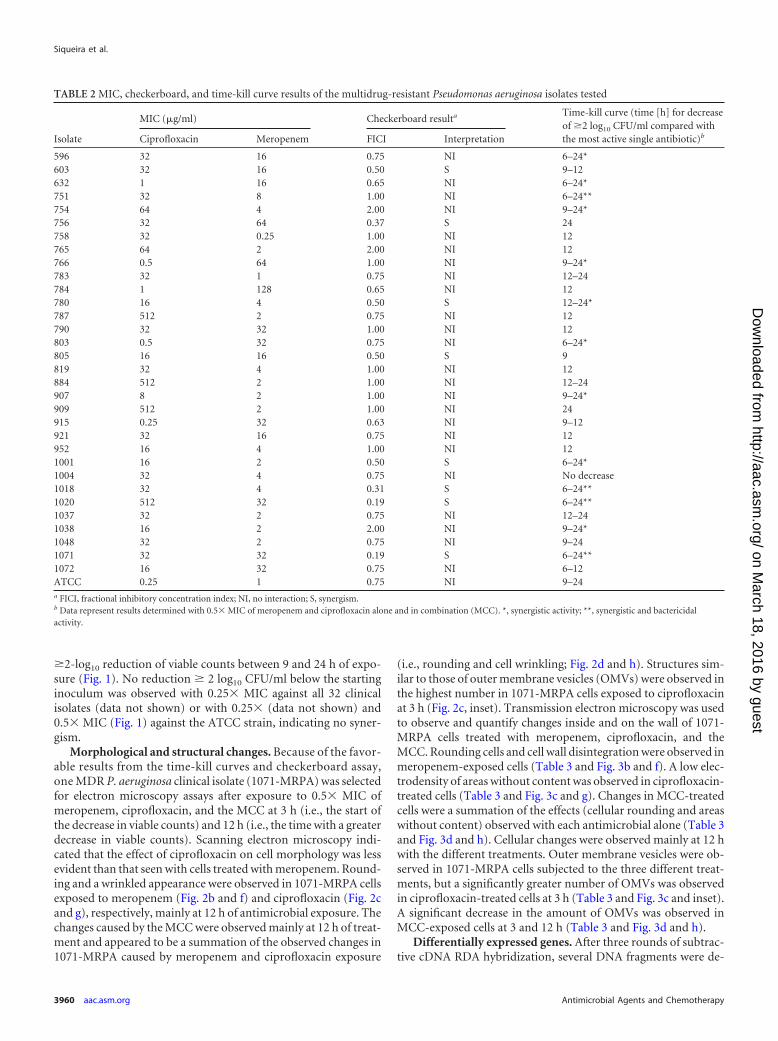
TABLE 2 MIC, checkerboard, and time-kill curve results of the multidrug-resistant Pseudomonas aeruginosa isolates tested
Isolate
MIC (�g/ml) Checkerboard resulta Time-kill curve (time [h] for decreaseof �2 log10 CFU/ml compared withthe most active single antibiotic)bCiprofloxacin Meropenem FICI Interpretation
596 32 16 0.75 NI 6–24*603 32 16 0.50 S 9–12632 1 16 0.65 NI 6–24*751 32 8 1.00 NI 6–24**754 64 4 2.00 NI 9–24*756 32 64 0.37 S 24758 32 0.25 1.00 NI 12765 64 2 2.00 NI 12766 0.5 64 1.00 NI 9–24*783 32 1 0.75 NI 12–24784 1 128 0.65 NI 12780 16 4 0.50 S 12–24*787 512 2 0.75 NI 12790 32 32 1.00 NI 12803 0.5 32 0.75 NI 6–24*805 16 16 0.50 S 9819 32 4 1.00 NI 12884 512 2 1.00 NI 12–24907 8 2 1.00 NI 9–24*909 512 2 1.00 NI 24915 0.25 32 0.63 NI 9–12921 32 16 0.75 NI 12952 16 4 1.00 NI 121001 16 2 0.50 S 6–24*1004 32 4 0.75 NI No decrease1018 32 4 0.31 S 6–24**1020 512 32 0.19 S 6–24**1037 32 2 0.75 NI 12–241038 16 2 2.00 NI 9–24*1048 32 2 0.75 NI 9–241071 32 32 0.19 S 6–24**1072 16 32 0.75 NI 6–12ATCC 0.25 1 0.75 NI 9–24a FICI, fractional inhibitory concentration index; NI, no interaction; S, synergism.b Data represent results determined with 0.5� MIC of meropenem and ciprofloxacin alone and in combination (MCC). *, synergistic activity; **, synergistic and bactericidalactivity.
Siqueira et al.
3960 aac.asm.org Antimicrobial Agents and Chemotherapy
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from
tected that indicated differential expression. A total of 91, 86, and89 clones were successfully sequenced, and the RDA1 (mero-penem exposure), RDA2 (ciprofloxacin exposure), and RDA3(MCC exposure) expressed sequence tag data are summarized inTable 4. The most redundant contigs were matched with serineacetyltransferase and OprF in RDA1, with cell wall hydrolase andalanine racemase in RDA2, and with alanine racemase plus a hy-pothetical protein in RDA3 (Table 4). The singlets formed fromESTs of the different RDAs did not match with the databases andwere not considered for further studies. In the reverse-RDA ex-periment, a total of 38 clones were sequenced. The transcriptionalprofile did not display any similarity to the one previously ob-
served in the cDNA RDA of P. aeruginosa exposed to antimicro-bials (data not shown).
Gene expression quantification. From the differentially de-tected transcripts in RDA1, RDA2, and RDA3, eight with aredundancy value higher than 10 were quantified. The genesthat encoded OprF and a hypothetical protein were those mosthighly induced (4.961-fold and 2.112-fold, respectively) in thepresence of meropenem. Alanine racemase (2.106-fold) wasthe most induced by ciprofloxacin exposure. Significantlylower expression was observed for most of the differentiallyexpressed genes in MCC-treated 1071-MRPA (Fig. 4) com-pared with meropenem and ciprofloxacin exposure alone. Of
FIG 1 Time-kill curve using 0.5� MIC of meropenem and ciprofloxacin alone and in combination (MCC) for ATCC 27853 (a) and a multidrug-resistantPseudomonas aeruginosa (1071-MRPA) clinical isolate (b). Each data point represents the mean number of viable bacterial counts for duplicate experiments, anderror bars represent the standard deviations.
Antimicrobial Synergistic Effect on P. aeruginosa
July 2014 Volume 58 Number 7 aac.asm.org 3961
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from
the genes related to antimicrobial resistance, no significant dif-ference in the levels of mexA expression was observed with thethree treatments. However, ampC and oprD were upregulatedwhen exposed to meropenem and significantly less expressedwith MCC exposure (Fig. 4). The same was observed with thelexA SOS response repressor gene.
DISCUSSION
The use of antimicrobial combinations has been a strategy to clini-cally control MDR P. aeruginosa infection (28) and suppress theamplification of resistant subpopulations (10). However, themechanism of action of the compounds in combination has beenlittle investigated.
FIG 2 (a and e) Scanning electron micrograph of the multidrug-resistant Pseudomonas aeruginosa clinical isolate (1071-MRPA) incubated at 35 2°C inCAMHB without antibiotic for 3 h (a) and 12 h (e). (b to d and f to h) 1071-MRPA treated with a subinhibitory concentration (0.5� MIC) of meropenem (b andf), ciprofloxacin (c [and inset] and g), and MCC (d and h). The white arrows in panel C indicate the outer membrane vesicles (OMV), while the white arrows inpanel G indicate the wrinkling cell. Inset magnification, �20,000.
Siqueira et al.
3962 aac.asm.org Antimicrobial Agents and Chemotherapy
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from

In the present study, important synergism of the MCC againstMDR P. aeruginosa clinical isolates was observed, mainly reflectedby the time-kill curves (40.6%). Some studies investigated thiscombination for the treatment of P. aeruginosa infection. How-ever, comparisons of these results are hampered because of meth-odological differences, with differences in the percentages of syn-ergism of 74.5% in the United States (14), 22.0% and 61.0% inTurkey (12), and 8.0% in Greece (8). Also, no synergism of theMCC was observed with American isolates (29) and Italian iso-lates (30). The checkerboard and time-kill curve assays used in thepresent study were justified because the best technique for assess-ing synergism in bacteria is unclear (31, 32). A low correlation wasfound between the checkerboard and time-kill curve results in thepresent study. The data regarding correlation between the MICand synergism results were inconclusive. The MCC time-kill curveassay showed synergism mainly in isolates resistant to ciprofloxa-cin, whereas checkerboard synergism was more evident amongisolates that were resistant to both antimicrobials.
The 1071-MRPA cell changes observed by SEM and TEM in-creased with meropenem, ciprofloxacin, and MCC exposure time.In the present study, the morphological and structural changesobserved in MCC-treated cells suggested a summation of the ef-fects observed with meropenem and ciprofloxacin treatmentalone. Nonetheless, the number of OMVs detected in MCC-treated cells was significantly lower than the number detected inmeropenem- and mainly ciprofloxacin-exposed cells. Outermembrane vesicles are constantly and naturally discharged fromthe cell wall during Gram-negative bacterial growth (33). Thesevesicles play an important role in bacterial virulence, transportingenzymes and other virulence factors for the host cell (34, 35).Apparently, these structures can increase the survival of bacteriaunder conditions of stress, such as the stress produced by antimi-crobials (36, 37). The antimicrobial ciprofloxacin targets DNAgyrase and topoisomerase IV, leading to breaks in the DNA strandand thus triggering the SOS response, stimulating bacterial outermembrane vesiculation, and contributing to cytotoxicity (36).Thus, a speculation could be made about the contribution of theMCC in reducing cytotoxicity in the host cell caused by bacterialOMVs.
Waisbren et al. (38) tested several antimicrobial combinationsother than the MCC against P. aeruginosa. They observed somecellular changes by SEM that were not observed in cells exposed tothe antimicrobials alone. According to those authors, the changesobserved involved the protective mechanisms of which the organ-
isms were deprived when exposed to synergistic combinations.Hayami et al. (39) studied MCCs using SEM and found differentcellular changes with the MCC compared with the cells exposed tothe antimicrobials alone.
In the present study, differences in the gene expression of 1071-MRPA exposed to subinhibitory concentrations of meropenem,
TABLE 3 Percentage of changes in multidrug-resistant Pseudomonasaeruginosa clinical isolate (1071-MRPA) cells, including OMV, after 3 or12 h of incubation with or without 0.5� MIC of meropenem,ciprofloxacin, and MCC by TEMa
Antimicrobial treatment
% cell changes by TEM (% OMV)
3 h 12 h
No antimicrobial (control) 6.8 (0.3) 7.4 (1.1)Meropenem 22.0b (6.9b) 68.5b (11.8b)Ciprofloxacin 23.5b (19.2b) 46.9b (14.8b)MCC 32.0d (4.2d) 92.7d (2.6c)a MCC, meropenem-ciprofloxacin combination; OMV, outer membrane vesicles; TEM,transmission electron microscopy.b P 0.01 compared to control.c P 0.01 compared to meropenem and ciprofloxacin alone.d P 0.01 compared to control and to meropenem and ciprofloxacin alone.
FIG 3 Transmission electron micrograph of multidrug-resistant Pseudomo-nas aeruginosa clinical isolate (1071-MRPA) incubated at 35 2°C in CAMHBfor 3 h (a to d) and 12 h (e to h) with a subinhibitory concentration (0.5�MIC) of meropenem (b and f), ciprofloxacin (c [and inset] and g [and inset]),and MCC (d and h). 1071-MRPA in CAMHB without antibiotic was incubatedfor 3 h (a) and 12 h (e) at 35 2°C. The arrows in panels c and g (including theinset) indicate outer membrane vesicles, while the arrows in panel f indicatecell wall disintegration. Bar, 1 �m (a to h), 200 nm (c inset), or 50 nm (g inset).
Antimicrobial Synergistic Effect on P. aeruginosa
July 2014 Volume 58 Number 7 aac.asm.org 3963
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from
ciprofloxacin, and the MCC were identified compared with 1071-MRPA not exposed to antimicrobial agents. Critical consider-ations when studying bacterial gene expression are the exposuretime and antimicrobial concentrations to be used for analysis. Theliterature on this subject has been quite variable (40–42). A longexposure time and different concentrations may be related to toxicand side effects and may not represent the primary target of theantimicrobial (23). In the present study, we used 60 min of expo-sure to 0.5� MIC (meropenem, ciprofloxacin, and the MCC),which represents the beginning of the reduction of viable cells, andantimicrobial concentrations with better synergistic action, as-sessed by time-kill assays. Considering the evidence of polyadenyl-ated mRNA in P. aeruginosa (43), the cDNA was obtained withrandom oligonucleotide hexamers plus oligo(dT) primers. Thisstrategy was used in an attempt to have the highest fidelity in theanalysis of gene expression by constructing cDNA.
By RDA analyses, transcripts related to cell wall repair or syn-thesis, DNA repair, and bacterial metabolism as well as severalhypothetical proteins of unknown function were observed. The
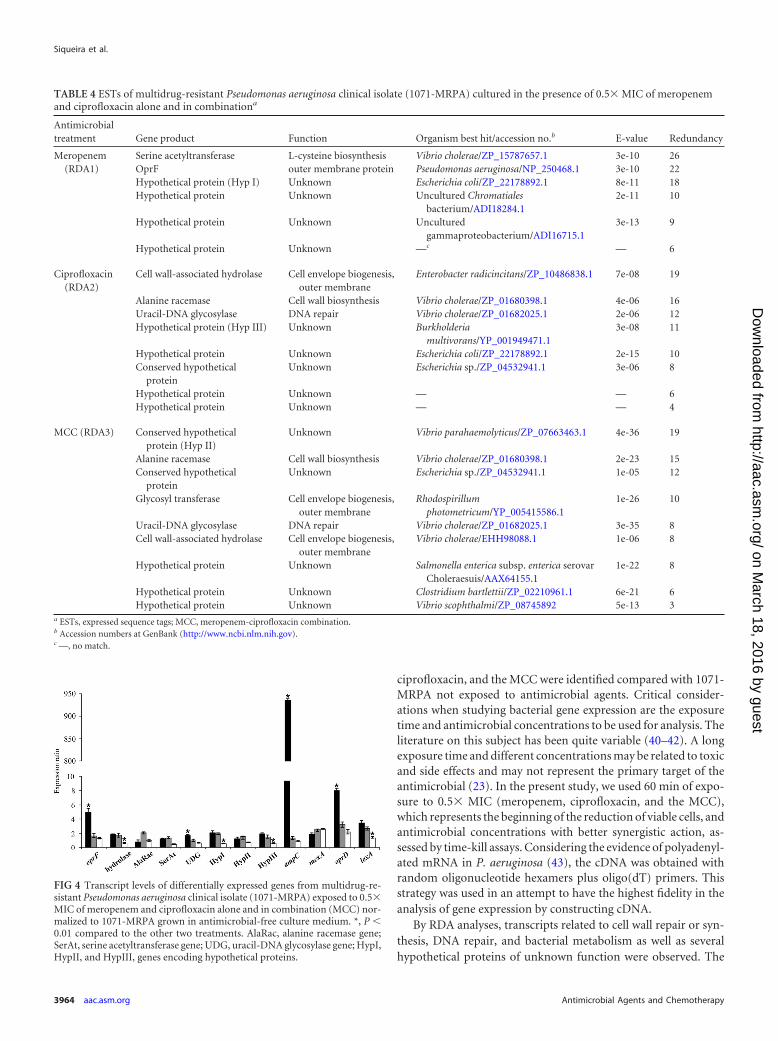
TABLE 4 ESTs of multidrug-resistant Pseudomonas aeruginosa clinical isolate (1071-MRPA) cultured in the presence of 0.5� MIC of meropenemand ciprofloxacin alone and in combinationa
Antimicrobialtreatment Gene product Function Organism best hit/accession no.b E-value Redundancy
Meropenem(RDA1)
Serine acetyltransferase L-cysteine biosynthesis Vibrio cholerae/ZP_15787657.1 3e-10 26OprF outer membrane protein Pseudomonas aeruginosa/NP_250468.1 3e-10 22Hypothetical protein (Hyp I) Unknown Escherichia coli/ZP_22178892.1 8e-11 18Hypothetical protein Unknown Uncultured Chromatiales
bacterium/ADI18284.12e-11 10
Hypothetical protein Unknown Unculturedgammaproteobacterium/ADI16715.1
3e-13 9
Hypothetical protein Unknown —c — 6
Ciprofloxacin(RDA2)
Cell wall-associated hydrolase Cell envelope biogenesis,outer membrane
Enterobacter radicincitans/ZP_10486838.1 7e-08 19
Alanine racemase Cell wall biosynthesis Vibrio cholerae/ZP_01680398.1 4e-06 16Uracil-DNA glycosylase DNA repair Vibrio cholerae/ZP_01682025.1 2e-06 12Hypothetical protein (Hyp III) Unknown Burkholderia
multivorans/YP_001949471.13e-08 11
Hypothetical protein Unknown Escherichia coli/ZP_22178892.1 2e-15 10Conserved hypothetical
proteinUnknown Escherichia sp./ZP_04532941.1 3e-06 8
Hypothetical protein Unknown — — 6Hypothetical protein Unknown — — 4
MCC (RDA3) Conserved hypotheticalprotein (Hyp II)
Unknown Vibrio parahaemolyticus/ZP_07663463.1 4e-36 19
Alanine racemase Cell wall biosynthesis Vibrio cholerae/ZP_01680398.1 2e-23 15Conserved hypothetical
proteinUnknown Escherichia sp./ZP_04532941.1 1e-05 12
Glycosyl transferase Cell envelope biogenesis,outer membrane
Rhodospirillumphotometricum/YP_005415586.1
1e-26 10
Uracil-DNA glycosylase DNA repair Vibrio cholerae/ZP_01682025.1 3e-35 8Cell wall-associated hydrolase Cell envelope biogenesis,
outer membraneVibrio cholerae/EHH98088.1 1e-06 8
Hypothetical protein Unknown Salmonella enterica subsp. enterica serovarCholeraesuis/AAX64155.1
1e-22 8
Hypothetical protein Unknown Clostridium bartlettii/ZP_02210961.1 6e-21 6Hypothetical protein Unknown Vibrio scophthalmi/ZP_08745892 5e-13 3
a ESTs, expressed sequence tags; MCC, meropenem-ciprofloxacin combination.b Accession numbers at GenBank (http://www.ncbi.nlm.nih.gov).c —, no match.
FIG 4 Transcript levels of differentially expressed genes from multidrug-re-sistant Pseudomonas aeruginosa clinical isolate (1071-MRPA) exposed to 0.5�MIC of meropenem and ciprofloxacin alone and in combination (MCC) nor-malized to 1071-MRPA grown in antimicrobial-free culture medium. *, P 0.01 compared to the other two treatments. AlaRac, alanine racemase gene;SerAt, serine acetyltransferase gene; UDG, uracil-DNA glycosylase gene; HypI,HypII, and HypIII, genes encoding hypothetical proteins.
Siqueira et al.
3964 aac.asm.org Antimicrobial Agents and Chemotherapy
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from

most redundant contig detected by the RDA in meropenem-ex-posed 1071-MRPA (RDA1) was a transcript related to serineacetyltransferase protein. Additionally, similar and reduced ex-pression levels of this protein were detected by qPCR in 1071-MRPA exposed to ciprofloxacin and the MCC (P � 0.01), respec-tively. Serine acetyltransferase catalyzes the first step of a two-stepreaction pathway for L-cysteine biosynthesis from L-serine in bac-teria, fungi, and plants (44), but the relationship between thisenzyme and the antimicrobials studied here remains to be clari-fied. The redundancy of OprF observed in RDA1 was confirmedby qPCR, which demonstrated the (4.9-fold) overexpression ofoprF. This protein is the most abundant outer membrane porin ofP. aeruginosa (45), and its role in transporting antimicrobials isnot well understood (46). OprF also plays an important role inmaintaining cell shape, particularly in adverse environments (47).The overexpression of oprF observed in 1071-MRPA cells exposedto meropenem could be an attempt to maintain cell shape, whichis quite altered by the action of meropenem, as demonstrated bySEM and TEM. Previous RDA and qPCR studies conducted in ourlaboratory with P. aeruginosa ATCC 27853 (susceptible strain)showed that OprF was also differentially overexpressed in cellsexposed to meropenem (data not shown). Recent studies suggestthat OprF is related to virulence factor production, including theformation of OMVs, through the modulation of the levels of aquorum sensing signal, the Pseudomonas quinolone signal (PQS)(48, 49). These findings reinforce the idea of a relationship be-tween OprF and meropenem. However, the relationship betweenOprF and MCC needs additional studies.
Cell wall hydrolase levels differentially detected in ciprofloxa-cin-exposed 1071-MRPA showed similar levels of expression withmeropenem exposure, too. This enzyme, which appears to be in-volved in cell wall biosynthesis and remodeling by promoting pep-tidoglycan hydrolysis (50), exhibited lower expression in MCC-exposed cells (P 0.01). A decrease in cell wall repair levels andhence in the bacterial defensive response could explain the higheractivity of the MCC against 1071-MRPA.
The gene that encodes the alanine racemase, differentially ex-pressed in 1071-MRPA under conditions of exposure to cipro-floxacin and MCC, was overexpressed under conditions of cipro-floxacin exposure. Alanine racemase is a bacterial enzyme thatcatalyzes the interconversion of L- and D-alanine, which is anamino acid that represents an important component of pepti-doglycan in most bacteria (51). Low concentrations of ciprofloxa-cin alter the structure of peptidoglycan (52). Thus, we may inferthat the bacteria also overexpressed alanine racemase to repair thepeptidoglycan as a defensive mechanism. Uracil-DNA glycosylaseproteins were also differentially expressed in 1071-MRPA exposedto ciprofloxacin and the MCC. However, they were expressedmore intensely (P 0.01) in meropenem-exposed 1071-MRPA.This enzyme plays an important role in preventing mutagenesis,eliminating uracil from DNA molecules by cleaving the N-glyco-sylic bond, and initiating the base excision repair pathway (53).Further studies are needed to better understand the relationshipbetween serine acetyltransferase, alanine racemase, and uracil-DNA glycosylase in the response of P. aeruginosa to the testedantimicrobials.
1071-MRPA is resistant to ciprofloxacin and carbapenems(i.e., imipenem and meropenem), but metallo-�-lactamase wasnot detected by genotypic and phenotypic analysis (data notshown). The loss or decreased expression of the outer membrane
porin OprD, coupled with the overexpression of the efflux pumpand AmpC, is another important mechanism of resistance in P.aeruginosa (17) and could be a mechanism involved in the resis-tance of 1071-MRPA to meropenem and ciprofloxacin. Based onthis, the ampC, mexA, and oprD genes were tested by qPCR to gainadditional knowledge about the effect of the MCC on 1071-MRPA. A significant difference in the levels of ampC and oprDexpression in 1071-MRPA was observed with meropenem, cipro-floxacin, and MCC exposure. As expected, 1071-MRPA exposedto meropenem, a �-lactam, showed ampC and oprD overexpres-sion. An intriguing observation was the significantly lower expres-sion of ampC and oprD with MCC exposure, which will requirefurther study.
The use of certain antibiotics, including ciprofloxacin, triggersthe SOS response, allowing the bacteria to adapt to the environ-ment or host, in addition to contributing to the development ofresistance (37, 54). The SOS response in P. aeruginosa involves thecontrolled derepression of 15 genes by the LexA repressor (41). Inthe present study, lexA was overexpressed with meropenem andciprofloxacin exposure, indicating that the transcription of theSOS genes was activated. These data are consistent with the SEMand TEM OMV findings, which corroborate the results of Mare-dia et al. (36). Thus, as suggested by Cirz et al. (37), an inhibitor ofLexA cleavage might have a favorable effect in P. aeruginosa ther-apy. Nonetheless, with MCC treatment, lower lexA expression andOMV production was found. Many questions remain unansweredabout LexA-regulated expression in P. aeruginosa, and furtherstudies could clarify the MCC results obtained in the presentstudy.
Although the present results need to be corroborated by addi-tional studies, they suggest that the MCC could be an alternativefor the treatment of infections caused by MDR P. aeruginosa. Theeffects of this antimicrobial combination may result not only fromthe summation of the effects of meropenem and ciprofloxacin butalso from differential actions that likely involve the protectivemechanisms of the bacteria.
ACKNOWLEDGMENTS
We thank Mychelle Vianna, Regina Cintia C. M. Velho, and Juliana Cogofor collaboration in handling of ultramicrotome and electron micro-scopes. In addition, we thank Lilian C. Baeza for many useful discussions.
This work was supported through a grant from Programa de Apoio àPesquisa Básica e Aplicada/Fundação Araucária; Coordenação de Aper-feiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacionalde Desenvolvimento Científico e Tecnológico (CNPq), Financiadora deEstudos e Projetos (FINEP), Complexo de Centrais de Apoio a Pesquisa(COMCAP), Laboratório de Ensino e Pesquisa em Análises Clínicas(LEPAC), and Programa de Pós-Graduação em Ciências Farmacêuticasda Universidade Estadual de Maringá.
REFERENCES1. Fothergill JL, Winstanley C, James CE. 2012. Novel therapeutic strate-
gies to counter Pseudomonas aeruginosa infections. Expert Rev. Anti In-fect. Ther. 10:219 –235. http://dx.doi.org/10.1586/eri.11.168.
2. Strateva T, Yordanov D. 2009. Pseudomonas aeruginosa - a phenomenonof bacterial resistance. J. Med. Microbiol. 58:1133–1148. http://dx.doi.org/10.1099/jmm.0.009142-0.
3. McGowan JE, Jr. 2006. Resistance in nonfermenting Gram-negative bac-teria: multidrug resistance to the maximum. Am. J. Med. 119(Suppl 1):S29 –S36. http://dx.doi.org/10.1016/j.amjmed.2006.03.014.
4. Mangoni ML, Maisetta G, Di Luca M, Gaddi LM, Esin S, Florio W,Brancatisano FL, Barra D, Campa M, Batoni G. 2008. Comparativeanalysis of the bactericidal activities of amphibian peptide analogues
Antimicrobial Synergistic Effect on P. aeruginosa
July 2014 Volume 58 Number 7 aac.asm.org 3965
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from

against multidrug-resistant nosocomial bacterial strains. Antimicrob.Agents Chemother. 52:85–91. http://dx.doi.org/10.1128/AAC.00796-07.
5. Mesaros N, Nordmann P, Plesiat P, Roussel-Delvallez M, Van EldereJ, Glupczynski Y, Van Laethem Y, Jacobs F, Lebecque P, Malfroot A,Tulkens PM, Van Bambeke F. 2007. Pseudomonas aeruginosa: resis-tance and therapeutic options at the turn of the new millennium. Clin.Microbiol. Infect. 13:560–578. http://dx.doi.org/10.1111/j.1469-0691.2007.01681.x.
6. El Solh AA, Alhajhusain A. 2009. Update on the treatment of Pseudomo-nas aeruginosa pneumonia. J. Antimicrob. Chemother. 64:229 –238. http://dx.doi.org/10.1093/jac/dkp201.
7. Cirioni O, Ghiselli R, Silvestri C, Kamysz W, Orlando F, MocchegianiF, Di Matteo F, Riva A, Lukasiak J, Scalise G, Saba V, Giacometti A.2007. Efficacy of tachyplesin III, colistin, and imipenem against a multi-resistant Pseudomonas aeruginosa strain. Antimicrob. Agents Chemother.51:2005–2010. http://dx.doi.org/10.1128/AAC.01576-06.
8. Kanellakopoulou K, Sarafis P, Galani I, Giamarellou H, Giamarellos-Bourboulis EJ. 2008. In vitro synergism of �-lactams with ciprofloxacinand moxifloxacin against genetically distinct multidrug-resistant isolatesof Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 32:33–39. http://dx.doi.org/10.1016/j.ijantimicag.2008.02.019.
9. Lim TP, Lee W, Tan TY, Sasikala S, Teo J, Hsu LY, Tan TT, SyahidahN, Kwa AL. 2011. Effective antibiotics in combination against extremedrug-resistant Pseudomonas aeruginosa with decreased susceptibility topolymyxin B. PLoS One 6:e28177. http://dx.doi.org/10.1371/journal.pone.0028177.
10. Louie A, Grasso C, Bahniuk N, Van Scoy B, Brown DL, Kulawy R,Drusano GL. 2010. The combination of meropenem and levofloxacin issynergistic with respect to both Pseudomonas aeruginosa kill rate and re-sistance suppression. Antimicrob. Agents Chemother. 54:2646 –2654.http://dx.doi.org/10.1128/AAC.00065-10.
11. Lister PD, Wolter DJ, Wickman PA, Reisbig MD. 2006. Levofloxacin/imipenem prevents the emergence of high-level resistance among Pseudomo-nas aeruginosa strains already lacking susceptibility to one or both drugs.J. Antimicrob. Chemother. 57:999 –1003. http://dx.doi.org/10.1093/jac/dkl063.
12. Ermertcan S, Hosgor M, Tunger O, Cosar G. 2001. Investigation ofsynergism of meropenem and ciprofloxacin against Pseudomonasaeruginosa and Acinetobacter strains isolated from intensive care unit in-fections. Scand. J. Infect. Dis. 33:818 – 821. http://dx.doi.org/10.1080/00365540110027222.
13. Solak S, Willke A, Ergonul O, Tekeli E. 2005. In vitro activity of mero-penem in combination with ciprofloxacin against clinical isolates of Pseu-domonas aeruginosa. Int. J. Antimicrob. Agents 25:181–182. http://dx.doi.org/10.1016/j.ijantimicag.2004.11.005.
14. Pankuch GA, Lin G, Seifert H, Appelbaum PC. 2008. Activity of mero-penem with and without ciprofloxacin and colistin against Pseudomonasaeruginosa and Acinetobacter baumannii. Antimicrob. Agents Chemother.52:333–336. http://dx.doi.org/10.1128/AAC.00689-07.
15. Erdem I, Kucukercan M, Ceran N. 2003. In vitro activity of combinationtherapy with cefepime, piperacillin-tazobactam, or meropenem with cip-rofloxacin against multidrug-resistant Pseudomonas aeruginosa strains.Chemotherapy 49:294 –297. http://dx.doi.org/10.1159/000074529.
16. Rossi F. 2011. The challenges of antimicrobial resistance in Brazil. Clin.Infect. Dis. 52:1138 –1143. http://dx.doi.org/10.1093/cid/cir120.
17. Zavascki AP, Carvalhaes CG, Picao RC, Gales AC. 2010. Multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii: resistancemechanisms and implications for therapy. Expert Rev. Anti Infect. Ther.8:71–93. http://dx.doi.org/10.1586/eri.09.108.
18. Siqueira VLD, Cardoso RF, Pádua RAF, Caleffi-Ferracioli KR, Helbel C,Santos ACB, Aoki EE, Nakamura CV. 2013. High genetic diversityamong Pseudomonas aeruginosa and Acinetobacter spp. isolated in a publichospital in Brazil. Braz. J. Pharm. Sci. 49:49 –56. http://dx.doi.org/10.1590/S1984-82502013000100006.
19. Clinical and Laboratory Standards Institute. 2012. Methods for dilutionantimicrobial susceptibility tests for bacteria that grow aerobically; ap-proved standard. Clinical and Laboratory Standards Institute, Wayne, PA.
20. Clinical and Laboratory Standards Institute. 2012. Performance stan-dards for antimicrobial susceptibility testing; 22nd informational supple-ment. Clinical and Laboratory Standards Institute, Wayne, PA.
21. Pillai SK, Moellering RC, Eliopoulos GM. 2005. Antimicrobial combi-nations, p 365– 440. In Lorian V (ed), Antibiotics in laboratory medicine,5th ed. Lippincott Williams & Wilkins, New York, NY.
22. Odds FC. 2003. Synergy, antagonism, and what the chequerboard putsbetween them. J. Antimicrob. Chemother. 52:1. http://dx.doi.org/10.1093/jac/dkg301.
23. Leite GGS, Baeza LC, Batista AA, Barbosa MFI, Pavan FL, Leite CQF,Silva JL, Hirata RDC, Hirata MH, Cardoso RF. 2013. Genes differen-tially expressed by Mycobacterium tuberculosis after exposure to ruthe-nium phosphinic compound and isoniazid. Intern. J. Microbiol. Res.5:356 –361. http://dx.doi.org/10.9735/0975-5276.5.1.357-362.
24. Del Sal G, Manfioletti G, Schneider C. 1989. The CTAB-DNA precipi-tation method: a common mini-scale preparation of template DNA fromphagemids, phages or plasmids suitable for sequencing. Biotechniques7:514 –520.
25. Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W,Lipman DJ. 1997. Gapped BLAST and PSI-BLAST: a new generation ofprotein database search programs. Nucleic Acids Res. 25:3389 –3402. http://dx.doi.org/10.1093/nar/25.17.3389.
26. Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression datausing real-time quantitative PCR and the 2(-Delta Delta C(T)) method.Methods 25:402– 408. http://dx.doi.org/10.1006/meth.2001.1262.
27. Ocampo-Sosa AA, Cabot G, Rodriguez C, Roman E, Tubau F, MaciaMD, Moya B, Zamorano L, Suarez C, Pena C, Dominguez MA, Mon-calian G, Oliver A, Martinez-Martinez L. 2012. Alterations of OprD incarbapenem-intermediate and -susceptible strains of Pseudomonas aerugi-nosa isolated from patients with bacteremia in a Spanish multicenterstudy. Antimicrob. Agents Chemother. 56:1703–1713. http://dx.doi.org/10.1128/AAC.05451-11.
28. Bodí M, Garnacho J. 2007. Pseudomonas aeruginosa: combined treatmentvs. monotherapy. Med. Intensiva 31:83– 87. (In Spanish.) doi:http://dx.doi.org/10.1016/S0210-5691(07)74780-0.
29. Isenberg HD, Alperstein P, France K. 1999. In vitro activity of cipro-floxacin, levofloxacin, and trovafloxacin, alone and in combination withbeta-lactams, against clinical isolates of Pseudomonas aeruginosa,Stenotrophomonas maltophilia, and Burkholderia cepacia. Diagn. Micro-biol. Infect. Dis. 33:81– 86. http://dx.doi.org/10.1016/S0732-8893(98)00126-6.
30. Campana S, Taccetti G, Farina S, Ravenni N, de Martino M. 2003.Antimicrobial susceptibility and synergistic activity of meropenemagainst Gram-negative non-fermentative bacteria isolated from cysticfibrosis patients. J. Chemother. 15:551–554. http://dx.doi.org/10.1179/joc.2003.15.6.551.
31. Cappelletty DM, Rybak MJ. 1996. Comparison of methodologies forsynergism testing of drug combinations against resistant strains of Pseu-domonas aeruginosa. Antimicrob. Agents Chemother. 40:677– 683.
32. Bonapace CR, White RL, Friedrich LV, Bosso JA. 2000. Evaluation ofantibiotic synergy against Acinetobacter baumannii: a comparison withEtest, time-kill, and checkerboard methods. Diagn. Microbiol. Infect. Dis.38:43–50. http://dx.doi.org/10.1016/S0732-8893(00)00163-2.
33. Beveridge TJ. 1999. Structures of gram-negative cell walls and their de-rived membrane vesicles. J. Bacteriol. 181:4725– 4733.
34. Bomberger JM, Maceachran DP, Coutermarsh BA, Ye S, O’Toole GA,Stanton BA. 2009. Long-distance delivery of bacterial virulence factors byPseudomonas aeruginosa outer membrane vesicles. PLoS Pathog.5:e1000382. http://dx.doi.org/10.1371/journal.ppat.1000382.
35. Tashiro Y, Uchiyama H, Nomura N. 2012. Multifunctional membranevesicles in Pseudomonas aeruginosa. Environ. Microbiol. 14:1349 –1362.http://dx.doi.org/10.1111/j.1462-2920.2011.02632.x.
36. Maredia R, Devineni N, Lentz P, Dallo SF, Yu J, Guentzel N, ChambersJ, Arulanandam B, Haskins WE, Weitao T. 2012. Vesiculation fromPseudomonas aeruginosa under SOS. ScientificWorldJournal 2012:402919. http://dx.doi.org/10.1100/2012/402919.
37. Cirz RT, O’Neill BM, Hammond JA, Head SR, Romesberg FE. 2006.Defining the Pseudomonas aeruginosa SOS response and its role in theglobal response to the antibiotic ciprofloxacin. J. Bacteriol. 188:7101–7110. http://dx.doi.org/10.1128/JB.00807-06.
38. Waisbren SJ, Hurley DJ, Waisbren BA. 1980. Morphological expressionsof antibiotic synergism against Pseudomonas aeruginosa as observed byscanning electron microscopy. Antimicrob. Agents Chemother. 18:969 –975. http://dx.doi.org/10.1128/AAC.18.6.969.
39. Hayami H, Goto T, Kawahara M, Ohi Y. 1999. Activities of beta-lactams,fluoroquinolones, amikacin and fosfomycin alone and in combinationagainst Pseudomonas aeruginosa isolated from complicated urinary tractinfections. J. Infect. Chemother. 5:130 –138. http://dx.doi.org/10.1007/s101560050022.
Siqueira et al.
3966 aac.asm.org Antimicrobial Agents and Chemotherapy
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from

40. Kolayli F, Karadenizli A, Savli H, Ergen K, Hatirnaz O, Balikci E, BudakF, Vahaboglu H. 2004. Effect of carbapenems on the transcriptional ex-pression of the oprD, oprM and oprN genes in Pseudomonas aeruginosa. J.Med. Microbiol. 53:915–920. http://dx.doi.org/10.1099/jmm.0.45692-0.
41. Juan C, Macia MD, Gutierrez O, Vidal C, Perez JL, Oliver A. 2005.Molecular mechanisms of beta-lactam resistance mediated by AmpC hy-perproduction in Pseudomonas aeruginosa clinical strains. Antimicrob.Agents Chemother. 49:4733– 4738. http://dx.doi.org/10.1128/AAC.49.11.4733-4738.2005.
42. Hay T, Fraud S, Lau CH, Gilmour C, Poole K. 2013. Antibiotic induc-ibility of the mexXY multidrug efflux operon of Pseudomonas aeruginosa:involvement of the MexZ anti-repressor ArmZ. PLoS One 8:e56858. http://dx.doi.org/10.1371/journal.pone.0056858.
43. Saravanamuthu SS, von Gotz F, Salunkhe P, Chozhavendan R, GeffersR, Buer J, Tummler B, Steinmetz I. 2004. Evidence for polyadenylatedmRNA in Pseudomonas aeruginosa. J. Bacteriol. 186:7015–7018. http://dx.doi.org/10.1128/JB.186.20.7015-7018.2004.
44. Olsen LR, Huang B, Vetting MW, Roderick SL. 2004. Structure of serineacetyltransferase in complexes with CoA and its cysteine feedback inhibi-tor. Biochemistry 43:6013– 6019. http://dx.doi.org/10.1021/bi0358521.
45. Sugawara E, Steiert M, Rouhani S, Nikaido H. 1996. Secondary structureof the outer membrane proteins OmpA of Escherichia coli and OprF ofPseudomonas aeruginosa. J. Bacteriol. 178:6067– 6069.
46. Bratu S, Landman D, Gupta J, Quale J. 2007. Role of AmpD, OprF andpenicillin-binding proteins in beta-lactam resistance in clinical isolates ofPseudomonas aeruginosa. J. Med. Microbiol. 56(Pt 6):809 – 814. http://dx.doi.org/10.1099/jmm.0.47019-0.
47. Rawling EG, Brinkman FS, Hancock RE. 1998. Roles of the carboxy-terminal half of Pseudomonas aeruginosa major outer membrane proteinOprF in cell shape, growth in low-osmolarity medium, and peptidoglycanassociation. J. Bacteriol. 180:3556 –3562.
48. Fito-Boncompte L, Chapalain A, Bouffartigues E, Chaker H, Lesouhai-
tier O, Gicquel G, Bazire A, Madi A, Connil N, Veron W, Taupin L,Toussaint B, Cornelis P, Wei Q, Shioya K, Deziel E, Feuilloley MG,Orange N, Dufour A, Chevalier S. 2011. Full virulence of Pseudomonasaeruginosa requires OprF. Infect. Immun. 79:1176 –1186. http://dx.doi.org/10.1128/IAI.00850-10.
49. Wessel AK, Liew J, Kwon T, Marcotte EM, Whiteley M. 2013. Role ofPseudomonas aeruginosa peptidoglycan-associated outer membrane pro-teins in vesicle formation. J. Bacteriol. 195:213–219. http://dx.doi.org/10.1128/JB.01253-12.
50. Johnson JW, Fisher JF, Mobashery S. 2013. Bacterial cell-wall recycling.Ann. N. Y. Acad. Sci. 1277:54 –75. http://dx.doi.org/10.1111/j.1749-6632.2012.06813.x.
51. Lambert MP, Neuhaus FC. 1972. Factors affecting the level of alanineracemase in Escherichia coli. J. Bacteriol. 109:1156 –1161.
52. Bryskier A. 1993. Fluoroquinolones: mechanisms of action and resistance.Int.J.Antimicrob.Agents2:151–183.http://dx.doi.org/10.1016/0924-8579(93)90052-7.
53. Pearl LH. 2000. Structure and function in the uracil-DNA glycosylasesuperfamily. Mutat. Res. 460:165–181. http://dx.doi.org/10.1016/S0921-8777(00)00025-2.
54. Hocquet D, Llanes C, Thouverez M, Kulasekara HD, Bertrand X,Plesiat P, Mazel D, Miller SI. 2012. Evidence for induction of integron-based antibiotic resistance by the SOS response in a clinical setting. PLoSPathog. 8:e1002778. http://dx.doi.org/10.1371/journal.ppat.1002778.
55. Dumas JL, van Delden C, Perron K, Kohler T. 2006. Analysis of antibi-otic resistance gene expression in Pseudomonas aeruginosa by quantitativereal-time-PCR. FEMS Microbiol. Lett. 254:217–225. http://dx.doi.org/10.1111/j.1574-6968.2005.00008.x.
56. Liu X, Li J, Wang Y, Li T, Zhao J, Zhang C. 2013. Green tea polyphenolsfunction as prooxidants to inhibit Pseudomonas aeruginosa and induce theexpression of oxidative stress-related genes. Folia Microbiol. (Praha) 58:211–217. http://dx.doi.org/10.1007/s12223-012-0198-2.
Antimicrobial Synergistic Effect on P. aeruginosa
July 2014 Volume 58 Number 7 aac.asm.org 3967
on March 18, 2016 by guest
http://aac.asm.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















