
Ribosomal Protein S7 Is Both a Regulator and a Substrate of ...
Upload
independentCategory
view
0download
0
Structural aspects of RbfA action during small ribosomal subunitassembly
Partha P. Datta1,9, Daniel N. Wilson2,3,9, Masahito Kawazoe4,9, Neil K. Swami1, TatsuyaKaminishi4, Manjuli R. Sharma1, Timothy M. Booth1, Chie Takemoto4, Paola Fucini5,6,Shigeyuki Yokoyama4,7, and Rajendra K. Agrawal1,8,*
1Laboratory of Structural Pathology, Division of Molecular Medicine, Wadsworth Center, New York StateDepartment of Health, Empire State Plaza, Albany, NY, USA, 12201-0509 2Munich Center for IntergratedProtein Science CiPS, University of Munich, Germany 3Gene Center and Department of Chemistry andBiochemistry, Ludwig-Maximilians-Universitat Munchen, Feodor-Lynen-Strasse 25, D-81377 Munich,Germany 4Protein Research Group, Genomic Sciences Center, Yokohama Institute, RIKEN, 1-7-22 Suehiro-cho, Tsurumi, Yokohama, 230-0045, Japan 5Max-Planck-Institute for Molecular Genetics, Ihnestrasse 73–75, D14195 Berlin, Germany 6J. W. Goethe-Universitaet Frankfurt am Main, Institut fuer Organische Chemieund Chemische Biologie, D-60438 Frankfurt am Main, Germany 7Department of Biophysics andBiochemistry, Graduate School of Science, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo113-0033, Japan; and 8Department of Biomedical Sciences, School of Public Health, State University of NewYork at Albany, Albany, NY 12201.
SummaryRibosome binding factor A (RbfA) is a bacterial cold-shock response protein, required for an efficientprocessing of the 5′end of the 16S ribosomal RNA (rRNA) during assembly of the small (30S)ribosomal subunit. Here we present a crystal structure of Thermus thermophilus RbfA and a three-dimensional cryo-electron microscopic (EM) map of the T. thermophilus 30S·RbfA complex. RbfAbinds to the 30S subunit in a position overlapping the binding sites of the A- and P-site tRNAs, andRbfA’s functionally important C-terminus extends toward the 5′ end of the 16S rRNA. In the presenceof RbfA, a portion of the 16S rRNA encompassing helix 44, which is known to be directly involvedin mRNA decoding and tRNA binding, is displaced. These results shed light on the role played byRbfA during maturation of the 30S subunit, and also indicate how RbfA provides cells with atranslational advantage under conditions of cold shock.
IntroductionRibosomes are complex macromolecular machines, which are involved in translating anorganism’s genetic information into polypeptides (reviewed by Ramakrishnan, 2002). Allribosomes consist of two unequally sized subunits, each composed of both ribosomal RNA
*Correspondence: Rajendra K. Agrawal, e-mail: [email protected], Tel: (518) 486 5797, Fax: (518) 474 7992.9These authors contributed equallyAccession Numbers:The cryo-EM map of the 30S·RbfA complex has been deposited in the EM database (http://www.ebi.ac.uk) under ID code EMD -1413.The coordinates of the Tth RbfA crystal structure, the homology model of RbfA as fitted into the cryo-EM map, and components of 30Sribosomal subunit located in RbfA neighborhood, have been deposited in the Protein Data Bank (www.rcsb.org) under the accessioncode 2DYJ, 2R1C, and 2R1G, respectively.Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptMol Cell. Author manuscript; available in PMC 2007 December 6.
Published in final edited form as:Mol Cell. 2007 November 9; 28(3): 434–445.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(rRNA) and ribosomal protein (r-protein) molecules. The small (30S) subunit plays a directrole in decoding of the genetic message (Ogle et al., 2003); in bacteria, such as Escherichiacoli, the small subunit is composed of one 16S rRNA molecule and 21 r-proteins (designatedS1–S21) (Wittmann-Liebold, 1986). In vitro, the 30S subunit can be assembled from only itsrRNA and r-protein components (Traub and Nomura, 1968, Culver and Noller, 2000);however, the process requires non-physiological conditions, namely, high magnesium ion andsalt concentrations (Traub and Nomura, 1968). In vivo, however, maturation of rRNAs andassembly of the r-proteins into a functional ribosome appear to be highly complex processes(Culver, 2003), involving multiple accessory factors (Williamson, 2003). While theinvolvement of many protein factors in ribosomal assembly has been well characterized,including modification enzymes, such as methylases and pseudouridinylases (Decatur andFournier, 2002), RNA helicases (Iost and Dreyfus, 2006) and molecular chaperones (Alix andNierhaus, 2003; Maki et al., 2002), there appear to be a number of additional protein factors,the exact roles of which remain to be defined (reviewed by Wilson and Nierhaus, 2007). Proteinfactors implicated in 30S maturation include the highly conserved GTPase Era (Sharma et al2005), the PRC β-barrel protein RimM (Bylund et al., 1998), the ribosome-activated GTPaseRsgA (also called YjeQ; Daigle and Brown, 2004; Himeno et al., 2004), and the ribosomebinding factor A (RbfA), the focus of our study.
RbfA is a small protein (10.9 kDa in T. thermophilus) required for efficient processing of the16S rRNA (Bylund et al., 1998; Xia et al., 2003). RbfA binds to the 30S subunit, but not to thelarge (50S) subunit, nor to the 70S ribosome (Dammel and Noller, 1995; Xia et al., 2003).NMR (Huang et al., 2003; Rubin et al., 2003) and X-ray crystallographic (PDB ID 1JOS)structures of RbfA obtained from various bacterial species reveal a single-domain protein witha type-II KH-domain fold topology, characteristic of a nucleic acid-binding protein family.Consistent with a role in ribosome biogenesis, deletion of the rbfA gene (Δ rbfA) causes adecrease in the quantity of 70S ribosomes and polysomes, and a concomitant increase in 30Sand 50S ribosomal subunits (Dammel and Noller, 1995; Jones and Inouye, 1996; Bylund etal., 1998). Furthermore, ΔrbfA mutants display an accumulation of 17S rRNA, a precursor tothe 16S rRNA (Bylund et al., 1998; Inoue et al., 2003). RbfA was originally identified as amulti-copy suppressor of the cold sensitivity of a C23U mutation at the 5′-terminal helix (h1)of the 16S rRNA (Dammel and Noller, 1993, 1995). The C23U mutation is predicted tosignificantly weaken the helix, enabling formation of an alternative helix through basepairingwith nucleotides located in the upstream region of the precursor 17S rRNA (See SupplementalFigure S1A, B). Indeed, additional suppressors of the C23U mutation were identified withinthe 16S rRNA that would appear to push the equilibrium back toward formation of h1 (Dammeland Noller, 1995). The cold sensitivity of the C23U mutant and ΔrbfA strains suggests theexistence of an energy barrier to the formation of the canonical h1, which is provided by high-temperature at the permissive temperatures of these strains, and by RbfA in the case of theC23U mutant strain at cold-shock temperatures. Thus, part of the role of RbfA could be tofacilitate correct folding and maturation of h1 at the 5′ end of the 16S rRNA, which isparticularly important under cold-shock conditions.
Cold shock results in an increase in the level of non-translating ribosomes and produces atemporary cessation of bacterial growth; growth is then restored through the action of a set ofcold shock response proteins (Jones and Inouye, 1994; Graumann et al., 1996; Datta andBhadra, 2003). The role of RbfA as a cold-shock protein has been well documented. In E.coli, RbfA is encoded in an operon together with the cold-shock protein NusA (Dammel andNoller, 1995). RbfA is expressed constitutively under normal growth conditions; however, theexpression level rapidly increases upon cold shock, due to an up-regulation of the transcriptionof the rbfA mRNA (Jones and Inouye, 1996), resulting in a several-fold increase in the amountof 30S-bound RbfA (Xia et al., 2003). The elevated levels of RbfA under cold-shock conditionsare necessary to overcome the translational block at the reduced temperature, presumably by
Datta et al. Page 2
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

facilitating rapid maturation of the 30S subunits. This role is in contrast to the action of thecold-shock protein pY; the latter has been proposed to stabilize 70S ribosomes againstdissociation, and thus protect them from degradation, by binding to them (Vila-Sanjurjo et al.,2004).
Here we report a crystal structure of T. thermophilus RbfA and a cryo-EM structure of a T.thermophilus 30S·RbfA complex at resolutions of 1.84 Å and 12.5 Å, respectively. Our analysisshows that RbfA binds at the junction of the head and body i.e. at the neck region of the 30Ssubunit, with the C-terminus of RbfA approaching helix 1 located at the 5′ end of the 16SrRNA. This strategic location of RbfA on the 30S subunit, and interaction of RbfA withmultiple rRNA helices and r-proteins, is suggestive of an important role in a late step inmaturation of the 30S subunit. In addition, we find that the presence of the RbfA maintains thedecoding region of the 30S subunit in a conformation unsuitable for the subunit’s participationin protein synthesis. Specifically, RbfA appears to dramatically alter the position andconformation of helix 44, a functionally important segment of the 16S rRNA that is known tobe directly involved in mRNA decoding and in the formation of two of the intersubunit bridges,B2a and B3 (Gabashvili et al., 2000; Yusupov et al., 2001). Our results not only provide insightinto the role of RbfA during maturation of the 30S subunit, but they also suggest how RbfAconfers a translational advantage to cells under conditions of cold shock.
Results and DiscussionsCrystal Structure of the Thermus thermophilus RbfA
The crystal structure of RbfA from T. thermophilus (Tth) was determined at 1.84 Å resolutionis shown in Figure 1A, and crystallographic and refinement data are provided in Table 1. Theasymmetric unit contains two molecules (A and B), and 90 (4–94) and 89 (3–92) out of the 95residues comprising Tth RbfA, could be unambiguously modeled, respectively. The refinedmodels of the two molecules can be superimposed with a root-mean-square deviation (r.m.s.d.)of 0.53 Å for the main chain atoms. The structure shows a single KH-domain containing threeα-helices (α1 to α3) and three β-strands (β1 to β3) with an αββααβ topology (Figure 1B). Theα2 and α3 helices are arranged to form a helix-kink-helix (hkh) at the strictly conserved Ala65with a CH-π interaction between Leu62 (α2) and Phe87 (β3) (Figure 1B). It is possible that thekink serves to expand the space for an interaction and to alter the direction of basic residueson α3. The electrostatic surface potential reveals highly positively charged regions, whichencompass three loops between β1-β2, β2-α2, and α3-β3, and the surface of α2-α3 (Figure 1C).A highly conserved sequence 25DPRL28 (29DPRL32 in E. coli) forms a 310 helix both inmolecules A and B, as seen in the E. coli structure (see alignment in Supplemental Figure S2;Huang et al., 2003). While Asp25 and Arg27 form an electrostatic interaction in molecule A,the side chain direction of Arg27 is different and the interaction is not observed in moleculeB. The loop between α3-β3, where the well-conserved Arg78 (Arg88 in E. coli) is located, alsoshows a notable difference in the two molecules. These conserved regions are likely to possessfunctional importance due to the structural flexibility.
Comparison of the two Tth RbfA structures with previously known RbfA structures revealsthat, despite the low sequence conservation (20%), the overall topologies are remarkablysimilar (Figures 1D). Tth RbfA is shorter than other known RbfA proteins, being ~95 aminoacids (aa) rather than the usual 111–150 aa (with E. coli having 133 aa), and the residues appearto be lacking from the C-terminal end (Supplemental Figure S2). However, significantdifferences between the structures are clearly evident, especially when comparing the highlyflexible loop regions, as well as the N- and C-terminal ends (Figure 1D).
Datta et al. Page 3
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Cryo-EM Structure of the 30S·RbfA ComplexAfter ascertaining that Tth RbfA binds to the mature T. thermophilus 30S subunit (Figure 2A),we obtained a 3D cryo-EM map of the Tth 30S·RbfA complex at 12.5 Å resolution, accordingto the FSC criterion with a 0.5 cutoff (Bottcher, et al., 1997; Malhotra et al., 1998), or at 8.0Å according to the 3 σ cutoff (Orlova et al., 1998). The map shows all of the recognizablefeatures of the 30S subunit, namely, head with beak, body with spur, platform, and helix 44(h44) of the 16S rRNA (Figure 2B). A complex mass of extra density, with two protrudingcylindrical features on the subunit interface side was directly visible within the cryo-EM mapof the 30S·RbfA complex, located between the head and body of the 30S subunit. In order tointerpret this density, we docked the X-ray crystallographic structure of the 30S subunit(Wimberly et al., 2000; PDB1J5E) into the cryo-EM map of the 30S·RbfA complex. For anoptimum fit, the X-ray coordinates were divided into four structural domains (head, body,platform, and the 3′-minor domain encompassing h44 and h45 of the 16S rRNA), and theneach domain was fitted individually as a rigid body. Subsequently, the fitted coordinates werefiltered to match the resolution of the cryo-EM map, and then subtracted from the cryo-EMmap of the 30S·RbfA complex. This process allowed the boundaries of the extra mass of densityin the 30S·RbfA cryo-EM map to be delineated (Figure 2B).
Volumetric calculations indicate that the extra density encloses a space (~922 voxels ≈ 19,384Å3) much larger than would be expected from the volume (~512 voxels ≈ 10,764 Å3) computedfrom the Tth RbfA structure. Possible explanations for this are that either two molecules ofRbfA are binding to each 30S subunit, or binding of RbfA induces a conformational changewithin the 30S subunit.
Conformational Changes of the 30S Subunit due to RbfA BindingTwo pieces of evidence indicate that RbfA binds as a monomer to the 30S subunit, and thusmust induce a dramatic conformational change in the 16S rRNA to produce the additionaldensity over that expected for a single RbfA molecule. Firstly, analysis of the density maps ofthe 30S·RbfA complex reveals much weaker density for where h45 and the top of h44 arelocated in the 30S control. Secondly, contouring to very high threshold values leads to a lossof all but the extra bi-lobed density (See Supplemental Figure S3). At such high thresholdsonly double stranded RNA helices are expected to be visible, whereas less electron densematerial, such as protein, is lost. This observation indicates that the two protruding cylindricalfeatures of the extra density must originate from a shift in rRNA helices h44 and h45, whereasthe remainder of the extra density, which disappears at high threshold values, represents thebound RbfA protein. Closer inspection of the cryo-EM map clearly suggests that the uppersegment of h44 (nucleotides 1400–1410 and 1490–1500) is what must shift position in theRbfA-bound 30S subunit by ~25 Å, to account for the lobe of extra density that is closer to thebody of 30S (Figures 3). Since h45 is contiguous with h44, and since the loop region of h45makes intimate contacts with the minor groove at the top of h44, it seems likely that the tworegions move as a single unit. This idea is also consistent with the excellent fit of the respectiveRNA helices into the lobed densities, which allowed satisfactory overall docking of the h44–h45 region from the crystal structure into the cryo-EM map (Figure 3). In the crystal structureof the 30S subunit, the 3′ minor domain of the 16S rRNA, encompassing h44 and h45,establishes limited contacts with the r-proteins, and utilizes non-sequence specific backboneinteractions with the rest of the 30S subunit, thus indicating the potential flexibility within thisdomain (Brodersen et al., 2002). Furthermore such conformational changes are not withoutprecedent, given that shifts in h44 have been observed upon the binding of translation initiationfactor IF1 (Carter et al., 2001), as well as between different elongation states of the ribosome(VanLoock et al., 2000). Note, however, that the shifts in the two cases just mentioned werenot of the magnitude that is seen here.
Datta et al. Page 4
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Localization of RbfA on the 30S SubunitWe isolated the portion of the extra density that is directly attributable to RbfA by convertingthe fitted coordinates of h44 and h45 into electron densities and filtering them to the resolutionof the cryo-EM map. The resulting mass was then subtracted from the total extra density mass(Figure 4A), leaving a density feature that matches the structure of Tth RbfA in volume andin overall shape (Figure 4B). However, the homology model of the Tth RbfA, which includesthree additional amino acids at the C-terminus, shows a slightly better fit into the density(Figure 4C), as indicated by a cross-correlation coefficient (CCF) of 0.79. The conclusion isthat RbfA binds in the neck region, buried deep within the cleft between the head and body ofthe 30S subunit. This position of RbfA is pivotal in that it is a junction point between all fourdomains of the 30S subunit, namely the “switch” region where h1 of the 5′ domain (body) basepairs with the single-stranded region between the central (platform) and 3′ major (head)domains, as well as the linker between the 3′ major and 3′ minor (h44–45) domains located atthe base of h27 (see Supplemental Figure S1C, D). When the complex is viewed from theintersubunit side, the side of the 30S subunit that would face the 50S subunit in the 70Sribosome, RbfA is situated behind h44 and h45. The inaccessibility of the RbfA binding sitein the 30S·RbfA complex suggests that binding of RbfA to the ribosome utilizes an inducedfit-like mechanism, such that h44 and h45 shift concomitantly with RbfA’s interaction withthe neck region of the 30S subunit. One surface of the KH domain of RbfA forms a three-stranded β-sheet, which in the 30S·RbfA complex is oriented toward the neck region, whereasthe RNA-binding hkh motif, encompassing α2 and α3, faces the junction between h44 and h45(Figure 5A). The flexible C-terminus of RbfA extends from the end of β3, deep into the bodyof the 30S subunit, and approaches h1 of the 16S rRNA (Figure 6A).
Contacts of Bound RbfA with Components of the 30S SubunitThe low sequence identity between different RbfA homologues suggests that RbfA interactswith the 30S subunit utilizing general electrostatic interactions, rather than specific conservedcontacts (Supplemental Figure S4). RbfA interacts with three of the four domains in the 30Ssubunit, the head, body and 3′ minor (h44–45) domains. The most prominent interaction withthe 30S subunit involves the highly basic region on one side of RbfA, formed from the N-terminus, the hkh motif (α2-α3), and the flexible loop regions located between β1-β2 and α3-β3 (Figure 5A, B, also see Supplementary Figure S5). The positively charged residues locatedin the hkh motif are positioned so as to interact with the negatively charged phosphate-oxygenbackbone of nucleotides located in the single-stranded linker region that spans between the topof h44 and the base of h45. Similarly, the N-terminus and loop regions form a positive surfacewith which the backbone of h28, h29 and to a lesser extent h30 can interact. In contrast, it isinteresting that a large negative area, derived from the terminal end of α1 and start of β1, isoriented towards where the top of h44 is located in the native 30S subunit (Figure 5B, seeSupplemental Figure S5C). Therefore, the charge distribution supports not only our placementof RbfA and its interaction with rRNA, but also the necessity of a shift in the h44–45 region,to avoid electrostatic repulsion. In addition, the long C-terminal extensions of r-proteins S9and S13 wind their way into the P site of the 30S subunit and interact with RbfA (Figure 6B)at the periphery of the highly basic region. Deletion of either of these extensions in E. colishows that these extensions are not essential for viability; however, the deletion strains exhibitreduced growth rates, and the 30S subunits have a lower affinity for tRNA (Hoang et al.,2004). The other major site of contact between RbfA and the 30S subunit overlaps with thebinding site of IF1 (Carter et al., 2001), encompassing h18 and r-protein S12. The loop regionbetween β2-α2 approaches the backbone of nucleotides located at the top of h18 and is directlyproximal to the long flexible loop linking β1-β2 of the OB-fold of S12 (Figure 6B). This loopcontains highly conserved residues involved in mRNA-tRNA codon-anticodon discriminationat the A site; mutations within the loop affect translational fidelity (see Ogle et al., 2001).
Datta et al. Page 5
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

In the 30S·RbfA complex, rigid body docking of RbfA into the cryo-EM density places the C-terminus of RbfA in a position at the neck of the 30S subunit where it clashes with the singlestranded region linking h28 with the top of h44 and the upper region of h18 (Figure 6B).However, given the flexibility of this region in the known RbfA structures (Figure 1D), it isconceivable that the C-terminal amino acids relocate towards h18, such that the C-terminal endof the protein comes into close proximity with the loop region of h1 at the 5′ end of the 16SrRNA (Figure 6A).
Molecular Basis for the Overlapping Function between 30S Assembly FactorsThere appears to be a complicated interplay, as well as partial overlaps in function, among anumber of the 30S subunit assembly factors. It has been reported that overexpression of Era,a GTP-binding protein also involved in the 16S rRNA maturation, suppresses (at leastpartially), the cold-sensitive cell growth and defective ribosome assembly in a ΔrbfA strain(Inoue et al., 2003). Overexpression of RbfA, but not Era, can rescue a slow-growth phenotypeand assembly defects associated with deletion of the rimM gene, which encodes anotherribosome maturation factor, RimM (Bylund et al., 1998; Inoue et al., 2003). However,overexpression of RimM cannot complement the ΔrbfA strain, suggesting that some sort ofhierarchy exists during 30S assembly (Bylund et al., 1998). In light of the binding position ofRbfA determined here, we suggest an explanation for the partial functional redundancy of RbfAwith Era and RimM.
Although the principal known function of RbfA is to facilitate processing of the 5′ ends ofprecursor 17S rRNA, while that of Era is to facilitate processing of 3′ ends, our study showsthat RbfA binds in the immediate vicinity of Era’s binding position on the 30S subunit (Sharmaet al., 2005) (Figure 7A). We suggest that Era can partially compensate for RbfA functionthrough interaction with common structural elements of the ribosome, specifically h28 (Figure7B), which is directly basepaired through a pseudoknot interaction with nucleotides located inthe loop of h1 at the 5′ terminus of the 16S rRNA (Supplemental Figure S1A, B). Thus, theinteraction of Era with h28 could stabilize h1 indirectly, in a manner that RbfA normallyperforms through direct interaction.
Although the structure of RimM bound to the 30S subunit is not known, biochemical dataindicate that this maturation factor interacts directly with the head region, in the vicinity of r-proteins S13, S19, and helices 31 and 33b of the 16S rRNA, leading to the suggestion thatRimM plays a role in assembly of the head region (Lovgren et al., 2004). RbfA also participatesin multiple interactions with components of the head, including r-protein S13 and neighbouringrRNA helices. Therefore, as compared to bound Era, bound RbfA is better positioned to bringregions of the head, body, and 3′ minor domain together at a late assembly stage; in this waythe factor could compensate for the absence of RimM.
The Mechanisms of Action of RbfA during Small Subunit Assembly and Cold ShockThe binding position of RbfA on the 30S subunit supports a role for the factor during the finalsteps in the maturation of the 30S subunit (Jiang et al., 2007). In particular, the binding positionis consistent with the proposal that RbfA facilitates correct formation of h1 at the 5′ end of the16S rRNA through interaction between the C-terminus of RbfA and the loop region of h1(Supplemental Figure S1A, B; Dammel and Noller, 1995). The significantly longer C-terminalextensions present in RbfA from other bacteria, such as E. coli, rather than T. thermophilus,suggests that the interaction with h1 may be even more extensive in these species. One possibleexplanation for the shortened tail in T. thermophilus may be related to the high temperaturesat which this organism grows, by enabling efficient transition of the alternate competingconformation for h1 in the precursor 17S rRNA into the canonical h1 found in the mature 16SrRNA. It appears that, in E. coli for example, under normal growth temperatures (37 °C) RbfA
Datta et al. Page 6
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

is partially dispensable for this transition, although clearly the accumulation of 30S subunitscontaining 17S precursor observed in the ΔrbfA strain indicates that RbfA stimulates theprocess (Dammel and Noller, 1995; Bylund et al., 1998; Inoue et al., 2003). Indeed, thepresence of a C23U mutation in the stem of h1 has been proposed to weaken formation of thehelix to such an extent that under cold shock conditions, the phenotype of the mutant strain isreminiscent of the ΔrbfA strain (Dammel and Noller, 1993). The latter phenotype can be rescuedby overexpression of RbfA (Dammel and Noller, 1995). The detection of other suppressionmutations in the 16S rRNA, which appear to work through stabilization of h1, supports theidea that RbfA operates in an analogous manner (Dammel and Noller, 1995).
We believe that our complex most closely represents the post-processing of the 30S subunit,just prior to RbfA dissociation. However, one of the surprising findings that binding of RbfAinduces dramatic conformational changes in h44 and h45 of the 30S subunit, may haveimportant implications during maturation: The large shift of h44 significantly alters thelocations of two important intersubunit bridges, namely B2a and B3 (Figure 3; Gabashvili etal., 2000;Yusupov et al., 2001); such that the association of the precursor-30S·RbfA complexwith the 50S subunit would be prohibited (Dammel and Noller, 1995). Furthermore, theposition of bound RbfA on the 30S subunit overlaps that of IF1 (Carter et al., 2001), as wellas with the positions of the anticodon stem loops of A- and P-site bound tRNAs (Agrawal etal., 2000;Selmer et al., 2006; see Supplemental Figure S6). In addition, RbfA is expected toblock the path of mRNA through the 30S subunit (Yusupova et al., 2001); and due to the shiftin the 3′ end of the 16S rRNA, the anti-Shine-Dalgarno (SD) sequence is placed in anunfavourable location for base-pairing with the SD-sequence present in the 5′ UTRs of manybacterial mRNAs. Interestingly, the cold-shock protein pY binds in a location similar to thatof RbfA, and has been shown to inactivate translation by preventing binding of A- and P-sitetRNAs (Vila-Sanjurjo et al., 2004;Wilson and Nierhaus, 2004). Thus, we believe that RbfAprevents precursor 30S subunits, in which h1 has not formed and the 5′ end has not beenprocessed, from entering the translation initiation cycle. However, we do not think this is thecase for mature 30S subunits, since overexpression of RbfA in vivo does not inhibit growth(Xia et al., 2003), and addition of RbfA does not inhibit protein synthesis in vitro (M.K., C.T.and S.Y., unpublished). This may suggest that binding of RbfA to precursor 30S subunits ismuch tighter than to mature 30S subunits, and/or that RbfA may be easily released from mature,but not precursor, 30S subunits during translation initiation complex formation. Moreover,RbfA is present at very low concentrations during normal growth conditions (Xia et al.,2003), but under cold-shock conditions, when many precursor 30S subunits are trapped withan alternate h1 conformation, RbfA expression is up-regulated (Jones et al., 1996). Thus, webelieve that RbfA stimulates protein synthesis indirectly, by facilitating a more rapid supplyof active 30S subunits under cold-shock conditions, rather than by promoting translation ofspecific mRNAs.
Experimental ProceduresPurification, crystallization and structure determination of the T. thermophilus RbfA
T. thermophilus HB8 rbfA gene (TTHA0907) was cloned to expression vector pET11a(Novagen) by the RIKEN Structural Genomics Initiative (Yokoyama et al., 2000) andrecombinant RbfA was expressed in E. coli strain BL21 (DE3). The cell lysate was incubatedat 55°C and RbfA was precipitated by the addition of 70%-saturated ammonium sulfate (AS)to the supernatant. The precipitate was dissolved in 20mM Tris-HCl buffer (pH 8.0) containing1.5 M NaCl and1.2 M AS, and loaded on hydrophobic column chromatography (HiPrepButhyl). Further purification was performed by cation exchange chromatography (monoScolumn) and by size-exclusion chromatography (Superdex-75). The final buffer is 20 mM Tris-HCl buffer containing 250 mM KCl (pH 8.0). The protein yield was 1 mg per 1 g of cells.
Datta et al. Page 7
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Crystals were obtained in drops composed of 0.5 μl protein solution (2.5 mg/ml) and 0.5 μlreservoir solution [1.2 M ammonium dihydrogen phosphate, 80 mM Tris/HCl (pH 8.5), 9.6 %glycerol, 80 mM magnesium formate, 20 mM Bis-Tris propane (pH 7.0)], by the sitting dropvapor diffusion technique at 20°C. The diffraction data was collected at BL-5A beamline inPhoton Factory (Tsukuba, Japan) from a single crystal up to 1.84 Å resolution and processedusing the HKL2000 suite and SCALEPACK programs (Otwinowski and Minor, 1997).General handling of the scaled data was carried out with programs in the CCP4 suite (CCP4,1994). The phase was determined by the method of molecular replacement with programmolrep, using a homology model for T. thermophilus RbfA (SWISS MODEL http://swissmodel.expasy.org/; Schwede et al., 2003) as a search model, which was generated from1JOS (Haemophilus influenzae) with ClustalW (Higgins et al., 1994). Models were rebuilt bycombining the auto-model building results of ARP-wARP, improved using O (Jones et al.,1991) and refined with CNS (Brunger et al., 1998). The structures were refined to R-factor of19.9 % (Rfree = 24.5 %) at 1.84 Å resolution. Protein secondary structure was defined by theDSSP algorithm (Kabsch and Sander, 1983). Figures were prepared with PyMOL (DeLano;http://www.pymol.org).
Preparation of the T. thermophilus 30S subunit and the 30S·RbfA complexThe S1-depleted 30S ribosomal subunits were isolated as described earlier (Sharma et al.,2005). The 30S·RbfA complex was prepared by incubation of RbfA (a 10–40-fold excess) with30S ribosomal subunits (360 nM) for 20 min at 65 °C in a buffer A, containing 20 mM, Hepes-KOH (pH 7.8), 10 mM Mg(OAc)2, 200 mM NH4Cl, 65 mM KCl. Reactions were stopped byincubation on ice, and pre-centrifuged for 10 min at 10000 rpm, and an aliquot was removedbefore the remainder was loaded onto a 10% sucrose cushion in buffer A and centrifuged in aTLN100 rotor at 78000rpm for 30 min. Binding of RbfA to the 30S was checked by runningthe initial reaction, and the TLN100 supernatant and pellet fractions, on 20% SDS-PAGE, withCoomassie blue staining (as seen in Figure 2A).
Cryo-EM and 3D reconstructionCryo-EM grids were prepared following standard procedures (Wagenknecht et al., 1988). EMdata were collected under low-dose conditions on a Philips Tecnai F20 field emission gunelectron microscope at 200 kV, at a magnification of 50,760X. Images were recorded between0.7 and 3.5 μm under focus. A total of 131 micrographs was used. The micrographs weredigitized on a Zeiss/Imaging scanner (Z/I Imaging Corporation, Huntsville, AL) with a stepsize of 14 μm, corresponding to 2.76 Å on the object scale. The three-dimensional (3D)reconstructions were calculated using a 3D projection alignment procedure (Penczek et al.,1994). Each data set was subdivided into defocus groups and then analyzed with SPIDERsoftware (Frank et al., 1996) to generate CTF-corrected 3D cryo-EM maps (Penczek et al.,1997). Initially, 154,476 particles were manually selected. The first 3D map obtained from allof the manually selected particle images showed a fragmented mass of extra density in the neckregion of the 30S subunit, suggesting a sub-stoichiometric binding of RbfA to the 30S subunit.Therefore, the data set was subjected supervised classification (Valle et al., 2002), by use ofthe 3D map from the total image set as one of the references. The other reference was the mapof the T. thermophilus control 30S subunit (Sharma et al., 2005). Finally, 61,207 particles thatclassified with the new reference map were used to calculate the 3D reconstructions of the30S·RbfA complex. The classification helped to enrich the population of RbfA-bound 30Simages. However, the possibility that the classified population was still contaminated withimages of free (uncomplexed) 30S subunits could not be ruled out. The resolution of the final3D cryo-EM map of the 30S·RbfA complex was 12.5 Å, according to the FSC criterion witha 0.5 cutoff (Bottcher et al., 1997; Malhotra et al., 1998), or 8.9 Å, according to the 3σ cutoffcriterion (Orlova et al., 1998).
Datta et al. Page 8
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fitting of X-ray crystallographic structuresTo attain an optimum fitting of the X-ray crystallographic structure of the T. thermophilus 30Ssubunit (Wimberly et al., 2000; PDB ID 1J5E), we divided the X-ray coordinates into fourstructural domains (head, body, platform, and 16S rRNA 3′-minor domain), and then fittedeach domain individually into the cryo-EM map of the 30S·RbfA complex. Subsequently,portions of the h44 and h45 rRNA helices from the 3′-minor domain of the X-raycrystallographic structure were separately fitted into the protruding cylindrical features of theextra mass visible in the cryo-EM map of the 30S·RbfA complex. Fitted coordinates wereconverted into electron density maps with a pixel size of 2.76 Å, and the resulting maps werefiltered to match the resolution of the cryo-EM map. The density assigned to RbfA was isolatedthrough comparison of the cryo-EM map of the 30S·RbfA complex and the density mapobtained from the fitting of the 30S X-ray coordinates into the cryo-EM map of the 30S·RbfAcomplex. The cross correlation coefficient (CCF) values between the fitted coordinates for T.thermophilus RbfA and the corresponding cryo-EM density maps were determined afterconversion of the fitted coordinates into the density map, through computation of averageddensities within volume elements scale-matched to those of the cryo-EM map (i.e., with a pixelsize of 2.76 Å, and after filtration of the X-ray map to the resolution of the cryo-EM densitymap). Visualization of the fitted atomic structures and the cryo EM density maps was donewith Ribbons (Carson, 1991) and IRIS EXPLORER (Numerical Algorithms group, Inc.,Downers Grove, IL), respectively.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
Acknowledgements
We thank S. Wakatsuki for the use of BL-5A at the Photon Factory and R. Nakayama-Ushikoshi, T. Terada and M.Shirouzu for sample preparation. This work was supported by NIH R01 grant GM61576 (to R.K.A.), and also by theRIKEN Structural Genomics/Proteomics Initiative (RSGI), the National Project on Protein Structural and FunctionalAnalysis, Ministry of Education, Culture, Sports, Science and Technology of Japan (to S.Y.), and DeutscheForschungsgemeinschaft grant FU579/1–2 (to P.F.). The authors acknowledge use of the Wadsworth Center’s EMinfrastructure.
ReferencesAgrawal RK, Spahn CM, Penczek P, Grassucci RA, Nierhaus KH, Frank J. Visualization of tRNA
movements on the Escherichia coli 70S ribosome during the elongation cycle. J Cell Biol2000;150:447–460. [PubMed: 10931859]
Alix JH, Nierhaus KH. DnaK-facilitated ribosome assembly in Escherichia coli revisited. RNA2003;9:787–793. [PubMed: 12810912]
Bottcher B, Wynne SA, Crowther RA. Determination of the fold of the core protein of hepatitis B virusby electron cryomicroscopy. Nature 1997;386:88–91. [PubMed: 9052786]
Brodersen DE, Clemons WM Jr, Carter AP, Wimberly BT, Ramakrishnan V. Crystal structure of the 30S ribosomal subunit from Thermus thermophilus: structure of the proteins and their interactions with16 S RNA. J Mol Biol 2002;316:725–768. [PubMed: 11866529]
Brunger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, KuszewskiJ, Nilges M, Pannu NS, Read RJ, Rice LM, Simonson T, Warren GL. Crystallography & NMR system:A new software suite for macromolecular structure determination. Acta Crystallogr D 1998;54:905–921. [PubMed: 9757107]
Bylund GO, Wipemo LC, Lundberg LA, Wikstrom PM. RimM and RbfA are essential for efficientprocessing of 16S rRNA in Escherichia coli. J Bacteriol 1998;180:73–82. [PubMed: 9422595]
Carson M. Ribbons 2.0. J Appl Crystallogr 1991;24:103–106.
Datta et al. Page 9
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Carter AP, Clemons WM Jr, Brodersen DE, Morgan-Warren RJ, Hartsch T, Wimberly BT, RamakrishnanV. Crystal structure of an initiation factor bound to the 30S ribosomal subunit. Science 2001;291:498–501. [PubMed: 11228145]
COLLABORATIVE COMPUTATIONAL PROJECT NUMBER 4. The CCP4 Suite: Programs forProtein Crystallography. Acta Crystallogr D 1994;50:760–763. [PubMed: 15299374]
Culver GM. Assembly of the 30S ribosomal subunit. Biopolymers 2003;68:234–249. [PubMed:12548626]
Culver GM, Noller HF. In vitro reconstitution of 30S ribosomal subunits using complete set ofrecombinant proteins. Methods Enzymol 2000;318:446–460. [PubMed: 10890005]
Daigle DM, Brown ED. Studies of the interaction of Escherichia coli YjeQ with the ribosome in vitro.J Bacteriol 2004;186:1381–1387. [PubMed: 14973029]
Dammel CS, Noller HF. A cold-sensitive mutation in 16S rRNA provides evidence for helical switchingin ribosome assembly. Genes Dev 1993;7:660–670. [PubMed: 7681419]
Dammel CS, Noller HF. Suppression of a cold-sensitive mutation in 16S rRNA by overexpression of anovel ribosome-binding factor, RbfA. Genes Dev 1995;9:626–637. [PubMed: 7535280]
Datta PP, Bhadra RK. Cold shock response and major cold shock proteins of Vibrio cholerae. ApplEnviron Microbiol 2003;69:6361–6369. [PubMed: 14602587]
Decatur W, Fournier M. rRNA modifications and ribosome function. Trends Biochem Sci 2002;27:344–351. [PubMed: 12114023]
Frank J, Radermacher M, Penczek P, Zhu J, Li Y, Ladjadj M, Leith A. SPIDER and WEB: processingand visualization of images in 3D electron microscopy and related fields. J Struct Biol 1996;116:190–199. [PubMed: 8742743]
Gabashvili IS, Agrawal RK, Spahn CM, Grassucci RA, Svergun DI, Frank J, Penczek P. Solution structureof the E. coli 70S ribosome at 11.5 Å resolution. Cell 2000;100:537–549. [PubMed: 10721991]
Graumann P, Schroder K, Schmid R, Marahiel MA. Cold shock stress-induced proteins in Bacillussubtilis. J Bacteriol 1996;178:4611–4619. [PubMed: 8755892]
Higgins D, Thompson J, Gibson T, Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improvingthe sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 1994;22:4673–4680. [PubMed:7984417]
Himeno H, Hanawa-Suetsugu K, Kimura T, Takagi K, Sugiyama W, Shirata S, Mikami T, Odagiri F,Osanai Y, Watanabe D, et al. A novel GTPase activated by the small subunit of ribosome. NucleicAcids Res 2004;32:5303–5309. [PubMed: 15466596]
Hoang L, Fredrick K, Noller HF. Creating ribosomes with an all-RNA 30S subunit P site. Proc Natl AcadSci U S A 2004;101:12439–12443. [PubMed: 15308780]
Huang YJ, Swapna GV, Rajan PK, Ke H, Xia B, Shukla K, Inouye M, Montelione GT. Solution NMRstructure of ribosome-binding factor A (RbfA), a cold-shock adaptation protein from Escherichiacoli. J Mol Biol 2003;327:521–536. [PubMed: 12628255]
Inoue K, Alsina J, Chen J, Inouye M. Suppression of defective ribosome assembly in a rbfA deletionmutant by overexpression of Era, an essential GTPase in Escherichia coli. Mol Microbiol2003;48:1005–1016. [PubMed: 12753192]
Iost I, Dreyfus M. DEAD-box RNA helicases in Escherichia coli. Nucleic Acids Res 2006;34:4189–4197. [PubMed: 16935881]
Jiang M, Sullivan SM, Walker AK, Strahler JR, Andrews PC, Maddock JR. Identification of novelEscherichia coli ribosome-associated proteins using isobaric tags and multidimensional proteinidentification techniques. J Bacteriol 2007;189:3434–3444. [PubMed: 17337586]
Jones PG, Inouye M. The cold-shock response--a hot topic. Mol Microbiol 1994;11:811–818. [PubMed:8022259]
Jones PG, Inouye M. RbfA, a 30S ribosomal binding factor, is a cold-shock protein whose absence triggersthe cold-shock response. Mol Microbiol 1996;21:1207–1218. [PubMed: 8898389]
Jones PG, Mitta M, Kim Y, Jiang W, Inouye M. Cold shock induces a major ribosomal-associated proteinthat unwinds double-stranded RNA in Escherichia coli. Proc Natl Acad Sci U S A 1996;93:76–80.[PubMed: 8552679]
Datta et al. Page 10
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Jones TA, Zhou JY, Cowan SW, Kjeldgaard M. Improved methods for building protein models in electrondensity maps and the location of errors in these models. Acta Crystallogra A 1991;47:110–119.
Kabsch W, Sander C. Dictionary of protein secondary structure: pattern recognition of hydrogen-bondedand geometrical features. Biopolymers 1983;22:2577–2637. [PubMed: 6667333]
Lovgren JM, Bylund GO, Srivastava MK, Lundberg LA, Persson OP, Wingsle G, Wikstrom PM. ThePRC-barrel domain of the ribosome maturation protein RimM mediates binding to ribosomal proteinS19 in the 30S ribosomal subunits. RNA 2004;10:1798–1812. [PubMed: 15496525]
Maki JA, Schnobrich DJ, Culver GM. The DnaK chaperone system facilitates 30S ribosomal subunitassembly. Mol Cell 2002;10:129–138. [PubMed: 12150913]
Malhotra A, Penczek P, Agrawal RK, Gabashvili IS, Grassucci RA, Junemann R, Burkhardt N, NierhausKH, Frank J. Escherichia coli 70 S ribosome at 15 Å resolution by cryo-electron microscopy:localization of fMet-tRNAfMet and fitting of L1 protein. J Mol Biol 1998;280:103–116. [PubMed:9653034]
Ogle JM, Brodersen DE, Clemons WM Jr, Tarry MJ, Carter AP, Ramakrishnan V. Recognition of cognatetransfer RNA by the 30S ribosomal subunit. Science 2001;292:897–902. [PubMed: 11340196]
Ogle JM, Carter AP, Ramakrishnan V. Insights into the decoding mechanism from recent ribosomestructures. Trends Biochem Sci 2003;28:259–266. [PubMed: 12765838]
Orlova EV, Dube P, Harris JR, Beckman E, Zemlin F, Markl J, van Heel M. Structure of keyhole limpethemocyanin type 1 (KLH1) at 15 Å resolution by electron cryomicroscopy and angular reconstitution.J Mol Biol 1997;271:417–437. [PubMed: 9268669]
Otwinowski Z, Minor W. Processing of X-ray diffraction data collected in oscillation mode. MethodsEnzymol 1997;276:307–326.
Penczek PA, Grassucci RA, Frank J. The ribosome at improved resolution: new techniques for mergingand orientation refinement in 3D cryo-electron microscopy of biological particles. Ultramicrosc1994;53:251–270.
Penczek PA, Zhu J, Schroder R, Frank J. Three-dimensional reconstruction with contrast transfer functioncompensation from defocus series. Scanning Microsc 1997;11:147–154.
Ramakrishnan V. Ribosome structure and the mechanism of translation. Cell 2002;108:557–572.[PubMed: 11909526]
Rubin SM, Pelton JG, Yokota H, Kim R, Wemmer DE. Solution structure of a putative ribosome bindingprotein from Mycoplasma pneumoniae and comparison to a distant homolog. J. Struct. Funct.Genomics 2003;4:235–243.
Schwede T, Kopp J, Guex N, Peitsch MC. SWISS-MODEL: an automated protein homology-modelingserver. Nucleic Acids Res 2003;31:3381–3385. [PubMed: 12824332]
Selmer M, Dunham CM, Murphy FVt, Weixlbaumer A, Petry S, Kelley AC, Weir JR, Ramakrishnan V.Structure of the 70S ribosome complexed with mRNA and tRNA. Science 2006;313:1935–1942.[PubMed: 16959973]
Sharma MR, Barat C, Wilson DN, Booth TM, Kawazoe M, Hori-Takemoto C, Shirouzu M, YokoyamaS, Fucini P, Agrawal RK. Interaction of Era with the 30S ribosomal subunit implications for 30Ssubunit assembly. Mol Cell 2005;18:319–329. [PubMed: 15866174]
Traub P, Nomura M. Structure and function of E. coli ribosomes. V. Reconstitution of functionally active30S ribosomal particles from RNA and proteins. Proc Natl Acad Sci U S A 1968;59:777–784.[PubMed: 4868216]
Valle M, Sengupta J, Swami NK, Grassucci RA, Burkhardt N, Nierhaus KH, Agrawal RK, Frank J. Cryo-EM reveals an active role for aminoacyl-tRNA in the accommodation process. EMBO J2002;21:3557–3567. [PubMed: 12093756]
VanLoock MS, Agrawal RK, Gabashvili IS, Qi L, Frank J, Harvey SC. Movement of the decoding regionof the 16 S ribosomal RNA accompanies tRNA translocation. J Mol Biol 2000;304:507–515.[PubMed: 11099376]
Vila-Sanjurjo A, Schuwirth BS, Hau CW, Cate JHD. Structural basis for the control of translationalinitiation during stress. Nat Struct Mol Biol 2004;11:1054–1059. [PubMed: 15502846]
Wagenknecht T, Grassucci R, Frank J. Electron microscopy and computer image averaging of ice-embedded large ribosomal subunits from Escherichia coli. J Mol Biol 1988;199:137–147. [PubMed:2451023]
Datta et al. Page 11
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Williamson JR. After the ribosome structures: how are the subunits assembled? RNA 2003;9:165–167.[PubMed: 12554857]
Wilson DN, Nierhaus KH. The how and Y of cold shock. Nat Struct Mol Biol 2004;11:1026–1028.[PubMed: 15523473]
Wilson DN, Nierhaus KH. The weird and wonderful world of bacterial ribosome regulation. Crit RevBiochem Mol Biol 2007;42:187–219. [PubMed: 17562451]
Wimberly BT, Brodersen DE, Clemons WM Jr, Morgan-Warren RJ, Carter AP, Vonrhein C, Hartsch T,Ramakrishnan V. Structure of the 30S ribosomal subunit. Nature 2000;407:327–339. [PubMed:11014182]
Wittmann-Liebold, B. Ribosomal proteins: their structure and evolution. In: Hardesty, B.; Kramer, G.,editors. Structure, function and genetics of ribosomes. Springer; NY: 1986. p. 326-361.
Xia B, Ke H, Shinde U, Inouye M. The role of RbfA in 16S rRNA processing and cell growth at lowtemperature in Escherichia coli. J Mol Biol 2003;332:575–584. [PubMed: 12963368]
Yokoyama S, Hirota H, Kigawa T, Yabuki T, Shirouzu M, Terada T, Ito Y, Matsuo Y, Kuroda Y,Nishimura Y, Kyogoku Y, Miki K, Masui R, Kuramitsu S. Structural genomics projects in Japan.Nat Struct Biol 2000;7:943–945. [PubMed: 11103994]
Yusupov MM, Yusupova GZ, Baucom A, Lieberman K, Earnest TN, Cate JH, Noller HF. Crystalstructure of the ribosome at 5.5 Å resolution. Science 2001;292:883–896. [PubMed: 11283358]
Yusupova GZ, Yusupov MM, Cate JH, Noller HF. The path of messenger RNA through the ribosome.Cell 2001;106:233–241. [PubMed: 11511350]
Datta et al. Page 12
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Crystal structure of Tth RbfA and its comparison with known atomic structures of RbfAfrom other species(A) Stereo representation of the Tth RbfA is shown in cartoon (PDB code: 2DYJ). (B) Anenlarged view of the helix-kink-helix motif (cyan) with all residues in stick. Residues involvedin notable interactions, Asp25 and Arg27 on the 310 helix (magenta) and the conserved Phe87,are also shown in stick. (C) Stereo representation of Tth RbfA in the surface potential preparedby using APBS tools built in PyMOL. (D) Superposition of structures of RbfA from T.Thermophilus (molecules A and B in the asymmetric unit are shown in dark and light bluecolors, respectively), H. influenzae (green, 1JOS) and M. pneumoniae (orange, 1PA4). In allpanels, N, C, and hkh mean N-terminus, C-terminus and helix-kink-helix motif, respectively.
Datta et al. Page 13
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. Binding of RbfA to the 30S ribosomal subunit(A) SDS-PAGE results showing the binding of RbfA to the 30S subunit. T. thermophilus 30Ssubunits were incubated alone (lanes 2–4), or with increasing concentrations of RbfA (10-fold(lanes 5–7), 20-fold (lanes 8–10) and 40-fold (lanes 11–13) molar excess over 30S subunit),before being centrifuged through a 10% sucrose cushion (see Experimental Procedures). As acontrol, reactions were also performed with RbfA in the absence of 30S subunits (lanes 15–20). For each condition, aliquots of the initial (pre) reaction, supernatant (S), and pellet (P)were subjected to 20% SDS-PAGE and stained with Coomassie blue. RbfA pellets only in thepresence of the 30S subunit, and the stoichiometry of binding increases with increasing initialexcess of RbfA over 30S subunit. Lanes 1 and 14 are marker lanes, with 14, 20 and 33 kDa
Datta et al. Page 14
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

bands indicated. (B) Cryo-EM map showing extra mass of density that encompasses the bindingposition of RbfA (red) on the T. thermophilus 30S subunit (yellow).
Datta et al. Page 15
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Conformational changes in the 30S subunit upon RbfA bindingA portion of the difference map (red) shown in Figure 2B corresponds to a large positionalshift of the 3′-minor domain of the 16S rRNA involving the decoding site helix 44. Bothpositions of helix 44 (h44), as well as those of helix 45 (h45), are shown. Purple ribbons,original positions; red/orange ribbons shifted positions. B2a and B3 indicate the positions ofthe inter-subunit bridges. Arrow indicates direction of the shift. The landmarks of the 30Ssubunit are: h, head, p, platform and sp, spur.
Datta et al. Page 16
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4. Localization of RbfA on the 30S subunit, and comparison of the atomic structure of RbfAwith the cryo-EM density map(A) Interpretation of the extra mass in terms of 30S subunit conformational change, involving16S rRNA helices 44 and 45 (orange), and RbfA mass (red). The mass attributable to RbfAwas derived after subtraction of the density corresponding to shifted positions of helices 44(h44′) and 45 (h45′) from the total extra mass shown in Figure 2B. (B, C) Stereo representationsof the fittings of (B) the X-ray crystallographic structure (CCF = 0.78) and (C) and thehomology model (CCF = 0.79) of the T. thermophilus RbfA into the corresponding cryo-EMdensity. The asterisk (*) in panel B points to an unoccupied region of the cryo-EM density,due to absence of three amino acid residues and a different orientation of the tail in the X-ray
Datta et al. Page 17
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

structure. However, the same density region is nicely accounted for by the homology model(panel C). Thumbnails to the left of the panels depict the orientations of the 30S subunit, withbody (b), head (h), and platform (p) identified.
Datta et al. Page 18
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. Interactions of RbfA with 16S rRNA helices 44 and 45(A) Stereo-view presentation of the interaction between the helix-kink-helix (hkh) motif ofRbfA (red) and the linker region between the shifted positions of helices 44 (h44′) and 45(h45′). The hkh motif is shown in cyan. (B) Correlation between the electrostatic distributionof RbfA (shown with regions of positive and negative potentials, in blue and red, respectively)and interaction of RbfA with h44 and h45. Both positions of h44 and h45 are shown: lightpurple ribbons, original positions; brown ribbons, shifted positions (marked as h44′ and h45′).Panel B was made with PyMol (http://www.pymol.org). In both panels, theRbfA-30S complexis viewed from the platform side, as depicted in the thumbnail to the lower right
Datta et al. Page 19
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
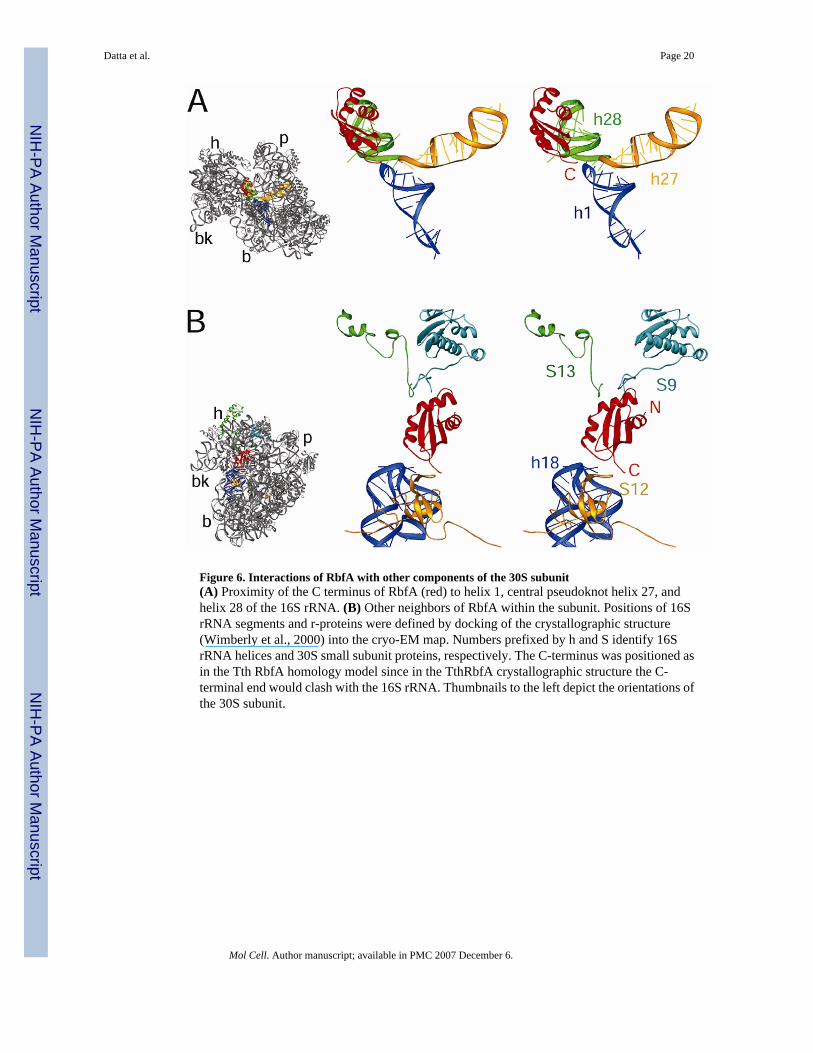
Figure 6. Interactions of RbfA with other components of the 30S subunit(A) Proximity of the C terminus of RbfA (red) to helix 1, central pseudoknot helix 27, andhelix 28 of the 16S rRNA. (B) Other neighbors of RbfA within the subunit. Positions of 16SrRNA segments and r-proteins were defined by docking of the crystallographic structure(Wimberly et al., 2000) into the cryo-EM map. Numbers prefixed by h and S identify 16SrRNA helices and 30S small subunit proteins, respectively. The C-terminus was positioned asin the Tth RbfA homology model since in the TthRbfA crystallographic structure the C-terminal end would clash with the 16S rRNA. Thumbnails to the left depict the orientations ofthe 30S subunit.
Datta et al. Page 20
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 7. Comparison of the binding positions of RbfA and Era on the 30S subunit(A) Binding position of RbfA (red) and Era (magenta; Sharma et al., 2005) on the 30S subunit.(B) RbfA (red) and Era (magenta) interact with a common structural element, h28, of the 16SrRNA (cyan). The thumbnail to the left depicts the orientation of the 30S subunit.
Datta et al. Page 21
Mol Cell. Author manuscript; available in PMC 2007 December 6.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Datta et al. Page 22
Table 1X-ray crystallographic structure determination statistics
Crystal characteristics
Space group P1 21 1Unit-cell parameters a = 28.43, b = 65.84, c = 43.17 β = 96.18Molecules/Asymmetric unit 2
Data collection
Wavelength (Å) 1.0000Resolution (Å)a 1.84 (1.96-1.84)Unique reflections 13813 (1383)Completeness (%)a 100 (100)I/σ(I)a 24.4 (3.22)Redundancy 3.7 (3.7)Rsym
a,b 0.061 (0.312)
Refinement
Resolution range (Å) 43.03-1.84Reflections 13796Rcryst/Rfree
c 0.199/0.245Rms deviation Bond lengths (Å) 0.008 Bond angles (°) 1.30 Dihedral angles (°) 21.50 Improper (°) 0.79Average B factor 25.6
aNumbers in parentheses are for the highest resolution shell.
bRsym=ΣhΣi|Ii(h)−<I(h)>|/ΣhΣI(h)
cRcryst=Σh||Fobs|−|Fcalc||/Σh|Fobs|. Rfree is Rcryst calculated with only the test set (5%) of reflections.
Mol Cell. Author manuscript; available in PMC 2007 December 6.
Copyright © 2022 FDOKUMEN




















