



Structural and functional characterization of tree proteins involved in redox regulation: a new...
26
Structural and functional characterization of Rv2966c reveals an RsmD-like methyltransferase from M. tuberculosis and the role of its N-terminal domain in target recognition # Atul Kumar 1 , Kashyap Saigal 2 , Ketan Malhotra 2 , Krishna Murari Sinha 2 *, Bhupesh Taneja 1 * 1 Institute of Genomics and Integrative Biology (CSIR), Mall Road, Delhi University Campus, Delhi- 110007; 2 Institute of Molecular Medicine, 254, Okhla Industrial Estate, Phase III, New Delhi-110020. *Address for Correspondence: Krishna Murari Sinha, IMM, 254, Okhla Industrial Estate, Phase III, New Delhi-110020. INDIA. Fax: +91 11 41028709, E-mail: [email protected] or Bhupesh Taneja, IGIB (CSIR), Mall Road, Delhi University Campus, Delhi-110007. Fax: +91-11-27667471. E-mail: [email protected] Running title: Rv2966c is an ortholog of RsmD Abstract: 9 out of 10 methylated nucleotides of E. coli 16S rRNA are conserved in Mycobacterium tuberculosis. All the 10 different methyltransferases are known in E. coli whereas only TlyA and GidB have been identified in mycobacteria. Here we have identified Rv2966c of Mycobacterium tuberculosis as an ortholog of RsmD protein of E. coli. We have shown that rv2966c can complement rsmD-deleted E. coli cells. Recombinant Rv2966c can use 30S ribosomes purified from rsmD-deleted E. coli as substrate and methylate G966 of 16S rRNA in vitro. Structure determination of the protein shows the protein to be a two-domain structure with a short hairpin domain at the N-terminus and a C- terminal domain with the SAM-MT fold. We show that the N-terminal hairpin is a minimalist functional domain that helps Rv2966c in target recognition. Deletion of the N-terminal domain prevents binding to nucleic acid substrates and the truncated protein fails to carry out the m2G966 methylation on 16S rRNA. The N-terminal domain also binds DNA efficiently, a property that may be utilized under specific conditions of cellular growth. Keywords: RNA methyltransferase/ RsmD/ SAM-MT fold/ SAM binding/ Rv2966c Introduction: Mycobacterium tuberculosis (M. tb) is one of the most infectious pathogens worldwide. It causes about 2 million deaths and 10 million cases of new infections every year (1,2). The major hurdle in the treatment of the disease is the emergence of drug resistant M. tb strains. Multiple drug resistant (MDR) and extremely drug resistant strains (XDR) of mycobacterium are becoming more and more prevalent (3,4). Understanding the molecular mechanism of drug resistance is an area of intense research now. Ribosome is one of the most common targets of antibiotics in the cell. Mutations in different components of ribosome are responsible for drug resistance. Several mutations in 23S and 16S rRNA genes, rRNA methyltransferases and ribosomal proteins in different mycobacterial strains have been shown to be responsible for resistance to different drugs. An A1400G mutation in 16S rRNA (rrs) gene leads to resistance to amikacin-kanamycin in M. tb strain ATCC 35827 and 13 other clinical isolates (5). Substitutions of C1401A/T or G1483T led to kanamycin resistance (6). Mycobacteria belonging to M. tb complex have intrinsic resistance to macrolides (7,8). The resistance has been attributed to erm genes which are 23S rRNA methyltransferases (9). Inactivation of tlyA gene which encodes a 2’-O- methyltransferase and methylates nucleotide C1409 of 16S rRNA and C1920 of 23S rRNA of M. tb makes it resistant to capreomycin and http://www.jbc.org/cgi/doi/10.1074/jbc.M110.200428 The latest version is at JBC Papers in Press. Published on April 7, 2011 as Manuscript M110.200428 Copyright 2011 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on May 25, 2016 http://www.jbc.org/ Downloaded from by guest on May 25, 2016 http://www.jbc.org/ Downloaded from by guest on May 25, 2016 http://www.jbc.org/ Downloaded from by guest on May 25, 2016 http://www.jbc.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Structural and functional characterization of tree proteins involved in redox regulation: a new...

Structural and functional characterization of Rv2966c reveals an RsmD-like methyltransferase from M. tuberculosis and the role of its N-terminal domain in target recognition#
Atul Kumar1, Kashyap Saigal2, Ketan Malhotra2, Krishna Murari Sinha2*, Bhupesh Taneja1* 1Institute of Genomics and Integrative Biology (CSIR), Mall Road, Delhi University Campus, Delhi-110007; 2Institute of Molecular Medicine, 254, Okhla Industrial Estate, Phase III, New Delhi-110020.
*Address for Correspondence: Krishna Murari Sinha, IMM, 254, Okhla Industrial Estate, Phase III, New Delhi-110020. INDIA. Fax: +91 11 41028709, E-mail: [email protected] or Bhupesh Taneja, IGIB (CSIR), Mall Road, Delhi University Campus, Delhi-110007. Fax: +91-11-27667471. E-mail: [email protected]
Running title: Rv2966c is an ortholog of RsmD
Abstract:
9 out of 10 methylated nucleotides of E. coli 16S rRNA are conserved in Mycobacterium tuberculosis. All the 10 different methyltransferases are known in E. coli whereas only TlyA and GidB have been identified in mycobacteria. Here we have identified Rv2966c of Mycobacterium tuberculosis as an ortholog of RsmD protein of E. coli. We have shown that rv2966c can complement rsmD-deleted E. coli cells. Recombinant Rv2966c can use 30S ribosomes purified from rsmD-deleted E. coli as substrate and methylate G966 of 16S rRNA in vitro. Structure determination of the protein shows the protein to be a two-domain structure with a short hairpin domain at the N-terminus and a C-terminal domain with the SAM-MT fold. We show that the N-terminal hairpin is a minimalist functional domain that helps Rv2966c in target recognition. Deletion of the N-terminal domain prevents binding to nucleic acid substrates and the truncated protein fails to carry out the m2G966 methylation on 16S rRNA. The N-terminal domain also binds DNA efficiently, a property that may be utilized under specific conditions of cellular growth.
Keywords: RNA methyltransferase/ RsmD/ SAM-MT fold/ SAM binding/ Rv2966c
Introduction:
Mycobacterium tuberculosis (M. tb) is one of the most infectious pathogens worldwide. It causes about 2 million deaths and 10 million cases of new infections every year (1,2). The major hurdle in the treatment of the disease is the emergence of drug resistant M. tb strains. Multiple drug resistant (MDR) and extremely drug resistant strains (XDR) of mycobacterium are becoming more and more prevalent (3,4). Understanding the molecular mechanism of drug resistance is an area of intense research now.
Ribosome is one of the most common targets of antibiotics in the cell. Mutations in different components of ribosome are responsible for drug resistance. Several mutations in 23S and 16S rRNA genes, rRNA methyltransferases and ribosomal proteins in different mycobacterial strains have been shown to be responsible for resistance to different drugs. An A1400G mutation in 16S rRNA (rrs) gene leads to resistance to amikacin-kanamycin in M. tb strain ATCC 35827 and 13 other clinical isolates (5). Substitutions of C1401A/T or G1483T led to kanamycin resistance (6). Mycobacteria belonging to M. tb complex have intrinsic resistance to macrolides (7,8). The resistance has been attributed to erm genes which are 23S rRNA methyltransferases (9). Inactivation of tlyA gene which encodes a 2’-O-methyltransferase and methylates nucleotide C1409 of 16S rRNA and C1920 of 23S rRNA of M. tb makes it resistant to capreomycin and
http://www.jbc.org/cgi/doi/10.1074/jbc.M110.200428The latest version is at JBC Papers in Press. Published on April 7, 2011 as Manuscript M110.200428
Copyright 2011 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

2
viomycin (10). Loss of GidB which is a 7-methylguanosine methyltransferase specific for 16S rRNA confers streptomycin resistance to the bacteria (11).
rRNA methyltransferases appear to be very strong candidates for drug targeting against mycobacteria but surprisingly there is very little information available about them. Erm, TlyA and GidB are the only methyltransferases which have been studied in mycobacteria (9-12). In E. coli, 10 methylated nucleotides in 16S rRNA and 12 in 23S rRNA are known with more than 20 rRNA methyltransferases responsible for these modifications (13). Alignment of 16S rRNA sequences of E. coli and M. tb show very high conservation with as many as 9 out of 10 methylated nucleotides being conserved. The very fact that each nucleotide in E. coli is methylated by a specific methyltransferase necessitates identification of the different methyltransferases in mycobacteria. Here, we have identified and characterized Rv2966c as an RsmD-like methyltransferase from M. tb by genome database search, structural and biochemical analysis. RsmD has m2G966 methyltransferase activity for 16S rRNA in E. coli (14). We have shown that rv2966c can complement rsmD-deleted E. coli cells. We have also determined its crystal structure to 1.9 angstrom resolution which reveals that Rv2966c is a structural homolog of RsmD. We also show for the first time, the role of a minimalist N-terminal domain in recruiting Rv2966c to ribosome to carry out its methyltransferase function.
Experimental Procedures:
Recombinant Rv2966c constructs: cloning, expression and purification: Open reading frame encoding Rv2966c was PCR-amplified from H37Rv genomic DNA using forward and reverse primers. Forward primer introduced a BamHI site at the start codon while the reverse primer introduced an XhoI site 3’ to the stop codon. The PCR product was digested with BamHI and XhoI and cloned in similarly digested pET28-His10-Smt3 vector to give the expression plasmid pMTase1. pMTase1 encodes full length Rv2966c polypeptide fused to an N-terminal His10-Smt3 tag. The tagged protein was expressed by growing
BL21 (DE3) cells transformed with pMTase1 in presence 50 μg/ml kanamycin. Protein expression was induced by growing the cells at 37°C to A600 ≈ 0.5, chilling them on ice for an hour followed by addition of ethanol to 2% and 0.2 mM IPTG. Cells were further grown with constant shaking at 17°C for 18 hours, harvested by centrifugation and stored at -80°C. All the subsequent steps were performed at 4°C. Cell pellet was thawed on ice and resuspended in buffer A (10 mM HEPES, pH 7.0, 10% glycerol, 0.1% β-mercaptoethanol and 300 mM NaCl). The cells were lysed by sonication, centrifuged to remove insoluble material and the supernatant was applied on a Ni-NTA agarose (Qiagen) column pre-equilibrated with buffer A. The column was then washed with 20 column volumes of buffer B (10 mM Potassium phosphate buffer, pH 7.4, 10% (v/v) glycerol, 0.1% (v/v) β-mercaptoethanol, 300 mM NaCl, 40 mM imidazole) and bound proteins were eluted with 3 column volumes of buffer B containing 250 mM imidazole. The eluted protein was mixed with Smt3-specific protease Ulp1 (Ulp1: protein ratio was 1: 500) and incubated at 4°C overnight to cleave His10-Smt3 tag. Tag-free protein was recovered in flow-through by passage of the mixture over Ni-NTA agarose column. Recombinant wild type Rv2966c (WT) thus purified was dialyzed against buffer C containing 10 mM Potassium phosphate buffer, pH 7.4, 5 mM EDTA, 100 mM NaC1, 10% (v/v) glycerol, 0.1% (v/v) β-mercaptoethanol. The dialysate was applied onto an ion-exchange column to remove nucleic acids. Purified protein was pooled, dialyzed against buffer D [10 mM Potassium phosphate buffer, pH 7.4, 5 mM EDTA, 10% (v/v) glycerol, 0.1% v/v β-mercaptoethanol, 50mM NaCl] and concentrated using Amicon ultra 3KDa molecular cutoff filter units and stored at 4°C until further use.
N-terminal truncated Rv2966c was expressed by PCR amplifying the truncated reading frames from pMTase1. Forward primers were designed to amplify the reading frame from the 19th or 24th amino acid residues of Rv2966c reading frame. The primers introduced a BamHI site at initiation of the reading frames. Reverse primer used was the same as used to amplify full length Rv2966c. PCR products were digested with BamHI and
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

3
XhoI and inserted between BamHI and XhoI sites of pET28-His10-Smt3 to give the expression plasmid pMTase4 and pMTase5. The proteins were expressed and purified from E. coli BL21 (DE3) following the same procedure as described above for the full length Rv2966c to result in truncated proteins Δ18RM and Δ23RM, lacking residues 1-18 or 1-23, respectively.
Construct for determining in vivo activity of full length and truncated Rv2966c: In vivo activity of full length, and truncated proteins were determined by expressing them under arabinose promoter in the pBAD24 vector containing ampicillin selection marker (15). Plasmids pMTase1, pMTase4 and pMTase5 were digested with NcoI and XhoI and the fragments containing the genes along with the fused N-terminal His10-Smt3 tag were cloned between NcoI and SalI sites of pBAD24 giving the expression constructs pMTase3, pMTase6 and pMTase7, respectively. A control plasmid pBSmt2 was generated by digesting pET28-His10-Smt3 vector with NcoI and XhoI and ligating the resulting 405 bp fragment between the NcoI and SalI sites of pBAD24. The control plasmid hence expresses His10-Smt3 protein only without any additional fusion partner.
A construct of Rv2966c lacking positively charged residues in the NTD was made by mutating Arg-3, Arg-12 and Arg-13 in the NTD to Ala in pMTase3. Mutagenesis was carried out using Quikchange site-directed mutagenesis kit (Stratagene). In the first cycle both Arg-12 and Arg-13 were mutated and the resultant construct was used as a template to make the third mutation, R3A to give the plasmid pMTase8. Presence of the desired mutations was confirmed by DNA sequencing. pMTase8 was used to monitor the in vivo activity of triple Arg-mutant of Rv2966c, 3RM. pMTase8 was digested with NcoI and HindIII and the fragment containing rv2966c (triple Arg-mutant) gene along with the fused N-terminal His10-Smt3 tag was subcloned between NcoI and HindIII sites of pET28-His10-Smt3 giving pMTase3RM. pMTase3RM hence expresses 3RM, which was purified as wild type Rv2966c.
Complementation of rsmD-deleted E. coli cells: m2G966 methyltransferase activity of full-length
Rv2966c, truncated and mutant proteins was determined by transforming rsmD-deleted E. coli KL16 cells (KL16ΔrsmD) (16) with pMTase3, pMTase6, pMTase7, pMTase8 or pBsmt2. Cells were selected against 100 μg/ml ampicillin and 50 μg/ml kanamycin. Protein was expressed by growing the cells at 37°C to A600 ≈ 0.5 and inducing with 0.2% arabinose for 3 hours. Cells were harvested and stored at -80°C.
Total RNA was isolated from the cells (RNeasy kit, Qiagen) and annealed to 32P-5’-end labeled primer P1 complementary to sequence 980-996 of E. coli 16S rRNA. Primer extension was carried out using AMV reverse transcriptase (Fermentas) as per the manufacturer’s protocol. The reaction was terminated with formamide/EDTA and products were analyzed on 15% denaturing polyacrylamide/7M urea gel and visualized by phosphoimaging. Oligonucleotides were also synthesized complementary to sequence 965-996, 966-996, 967-996 and 968-996 (labeled ME1, ME2, ME3 and ME4, respectively) and used as markers to analyze the length of product of primer extension. The product size of primer extension would be 30 nucleotides long (equal to ME3: 967-996) if G966 is methylated.
Circular Dichroism spectroscopy measurements: Far-UV CD spectra were collected on a Jasco J815 CD spectrometer in a quartz cuvette with a path length of 0.1 cm at room temperature. Ellipticity data was collected in the range of 250-195 nm. Concentration of the proteins (Rv2966c, Δ18RM or Δ23RM) for CD measurements was kept at 0.5 mg/ml in 5 mM HEPES, pH 7.4 and 50 mM KCl. Each spectrum was recorded as an average of four scans. In all experiments, contributions of the buffer to the spectra were subtracted and mean residue ellipticities were determined before plotting the spectra.
Electrophoretic mobility shift assay (EMSA): EMSA with DNA was carried out with pUC18 DNA (linearized with HindIII). Varying concentrations of Rv2966c, Δ18RM, Δ23RM or 3RM as indicated in figure legends were incubated with DNA in a buffer containing 50 mM Tris-HCl, pH 7.9; 250 mM NaCl; 10% glycerol; 1 mM EDTA; 0.5% Triton-X-100 for 60 min at 30°C.
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

4
For EMSA with RNA, RNA purified as above (RNeasy kit, Qiagen) was similarly incubated with Rv2966c, Δ18RM, Δ23RM or 3RM in an identical buffer as for DNA binding experiments. The protein-nucleic acid complex was analyzed on 0.8% agarose gel by ethidium bromide staining.
Western blot analysis: rsmD-deleted E. coli KL16 cells (16) transformed with pMTase3, pMTase6, pMTase7 or pMTase8 were grown at 37°C to A600 ≈ 0.5 and induced with 0.2% arabinose for 1 hour at 37°C. Total cell extract was prepared from each of the induced cells and fractionated on 12% SDS-PAGE. The proteins were transfered onto nitrocellulose membrane and probed with anti-His (Qiagen) and anti-DnaK (Stressgen) at 1:5000 dilution for both the antibodies.
Ribosome purification and in vitro methylation assay: Ribosome was isolated from both KL16ΔrsmD as well as KL16ΔrsmD complemented with pMTase3 as has been described before (17). In brief, the cells were grown in 500 ml of LB medium in presence of suitable antibiotic(s) at 37°C to A600≈ 0.6 and then induced with 0.2% arabinose. Cells were grown further for 3 hours with constant shaking at the same temperature, centrifuged, washed with Tris buffer saline (TBS) containing 10 mM Mg(OAc)2 and stored at -80°C. Pellets were suspended in 2 ml of resuspension buffer (20 mM HEPES, pH 7.5, 60 mM NH4Cl, 10.5 mM Mg (OAc)2, 0.5 mM EDTA and 10 mM β-mercaptoethanol) for each gram of wet cell pellet and lysed by sonication. Insoluble fraction was removed by centrifugation at 15,000 rpm for 40 min. The supernatant was treated with DNase I on ice for 30 min and centrifuged further at 31,000 rpm in a Beckman SW32 rotor for 4 h to pellet ribosomes. Ribosome pellet was suspended in 1 ml of extraction buffer (same as resuspension buffer but with 1M NH4Cl) and centrifuged again at 44,000 rpm for 12 h in a Beckman SW28 rotor on an 18% sucrose cushion. Ribosome pellets thus obtained were resuspended in the extraction buffer containing 2 mM Mg(OAc)2 to disassemble the ribosomal subunits. The subunits were separated by centrifuging through a 10-30% sucrose gradient for 12 hours at 31,500 rpm in a Beckman SW28 rotor. Fractions containing 30S ribosomes were collected,
concentrated through Amicon Ultra50K, dialyzed against buffer containing 20mM Hepes pH 7.4, 120mM NH4Cl and 10% glycerol and stored at -80°C until further use.
In vitro methylation: In vitro methyltransferase assay was done using a non-radioactive colorimetric assay kit (G-biosciences) with slight modification of the protocol as suggested by the supplier. Methyltransferase reaction was first carried out in a 20 μl reaction mixture containing 100 ng of enzyme (Rv2966c, Δ18RM or Δ23RM), 4 μg of purified 30S ribosome subunit, 100 μM S-adenosylmethionine (SAM) and incubated at 37°C for 1h. 10 μl of the reaction mixture was added to 90 μl of ‘master mix’ without SAM as suggested by the supplier. This reaction was further incubated at 37°C. The methyltransferase reaction results in an increase in S-adenosyl homocysteine due to transfer of methyl group from SAM to substrate (16S rRNA). S-adenosyl-homocysteine is rapidly converted to urate and hydrogen peroxide by coupled enzymatic reactions. The rate of production of hydrogen peroxide is monitored colorimetrically with 3,5 dichloro-2-hydroxy-benzene sulfonic acid at 510 nm to measure the methyltransferase reaction. A parallel control reaction to subtract background was done where methyltransferase assay was done in absence of SAM.
Protein crystallization, data collection and structure refinement: Rv2966c was crystallized by hanging drop vapor diffusion method by mixing 4 μl of 17 mg/ml Rv2966c (in 10 mM Potassium phosphate buffer, pH 7.4, 5 mM EDTA, 10% (v/v) glycerol, 0.1% v/v β-mercaptoethanol, 50mM NaCl, 100 μM SAM) and 2 μl of reservoir solution (100 mM Tris-HCl, pH7.4, 3.0 M potassium acetate) at 24°C. Long rod-shaped crystals were obtained in two-three days. The crystals were cryo-protected in a solution containing 25% ethylene glycol in addition to the crystal reservoir solution and flash-frozen in liquid nitrogen before data collection.
Diffraction data were collected at BM14 beam line at ESRF. The crystals belong to the space group P3221 and contain one molecule in the crystallographic asymmetric unit. Data were processed and integrated using MOSFLM (18) and
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

5
scaled using SCALA (19). Data collection and refinement statistics are summarized in Table 1.
To solve the structure of Rv2966c by molecular replacement, polyalanine model of several bacterial putative methyltransferases (PDB ID: 2FPO, 2FHP, 2IFT) were used as templates in Phaser (19). The best solution was obtained with the polyalanine model of putative methyltransferase of Enterococcus faecalis (PDBID: 2FHP) as the starting model. The initial model was further improved by iterative cycles of model building in COOT (20) and refinement using Refmac5 (21). Structure comparisons were performed with the help of DALI server (22). A final R-factor of 17.8% and Rfree of 22.1% were obtained (Table 1). The final coordinates have been deposited with PDB with the accession code 3P9N.
Results:
Identification and sequence analysis of Rv2966c of Mycobacterium tuberculosis:
Search of H37Rv genome database (http://www.tigr.org) for nucleic acid methyltransferases identified Rv2966c as one of the putative methyltransferases. Rv2966c consists of an open reading frame of 188 amino acid residues. A BLAST search of Rv2966c against non-redundant protein sequences shows a strong sequence homology with several putative uncharacterized methyltransferases of Actinobacteria.
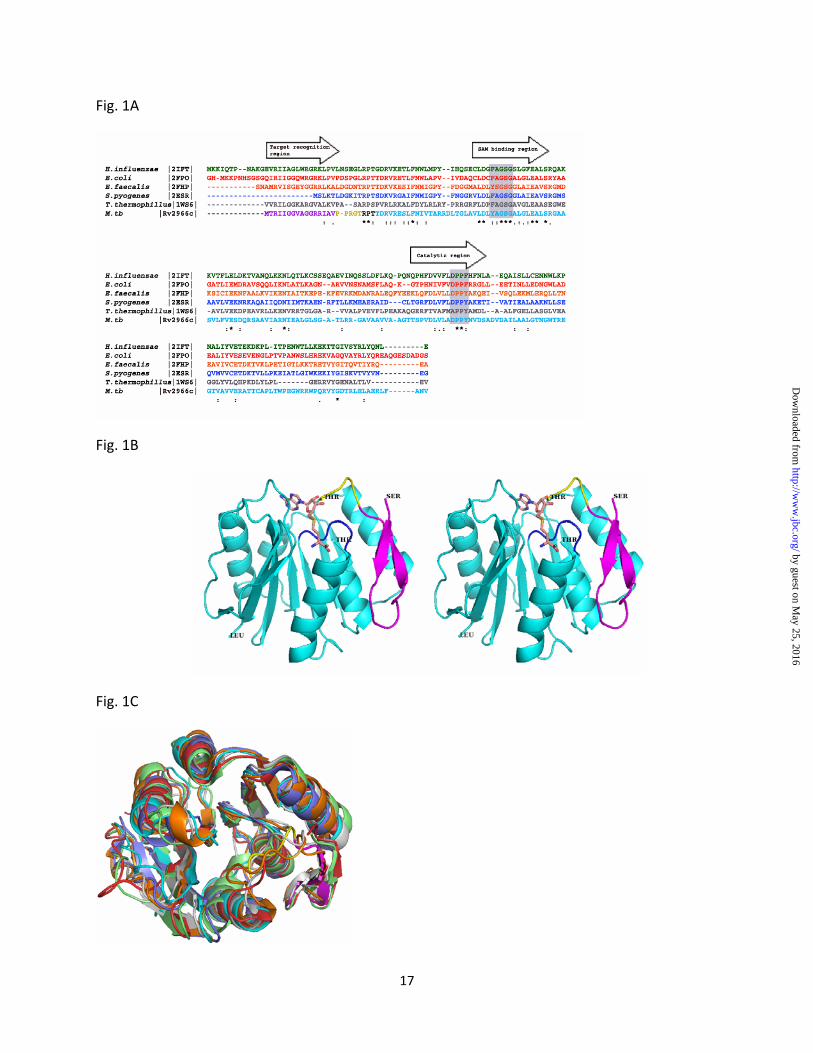
A search for presence of conserved domains in the protein identified residues 17-182 to contain domain belonging to Class I S-adenosylmethionine (SAM)-dependent methyltransferase superfamily. The protein contains a SAM-binding motif 51YAGSG55 (equivalent to conserved motif I, Y/FXGXG of DNA methyltransferases) and a 119DPPY123 motif (equivalent to conserved motif IV, D/N/SPPY/F of DNA methyltransferases) (Fig. 1A) (23-25). The 51YAGSGALG58 sequence of Rv2966c represents a general nucleotide binding motif in the SAM-binding pocket while the ‘DPPY’ motif is present at the active site in several DNA
methyltransferases and N5-glutamine methyltransferases (26-28).
The stretch of first 16 amino acid residues at N-terminus of Rv2966c did not show the presence of any known conserved domain. This stretch appears to be variable in length in different organisms although several residues in this region appear to be conserved (Fig. 1A). As described later in the text, the N-terminal region of Rv2966c appears to play a role in target recognition. Rv2966c hence belongs to a distinct family of nucleic acid methyltransferase resembling the ζ-class of DNA methyltransferases consisting of three distinct regions in the following sequential order: a target recognition region, SAM-binding region and lastly the catalytic region (23).
The closest characterized match to Rv2966c was found to be RsmD from E. coli (14) (e-value of 10-
18) with which it has 67 identical and 92 similar amino acid residues with 16 gaps in the sequence alignment. Other structural homologs, for instance Haemophilus influenzae putative methyltransferase (PDBID 2IFT, unpublished work, e-value 10-20), E. faecalis putative methyltransferase (PDBID 2FHP, unpublished work, e-value 10-14) among several others could be identified, although their biochemical functions have still not been characterized. RsmD is a 16S rRNA specific methyltransferase that methylates G966 at N2 position (14). Rv2966c hence may be a protein with similar role specific for modification of nucleotide equivalent to m2G966 in 16S rRNA in mycobacteria.
Crystallization and structural determination of Rv2966c:
Rv2966c was expressed by transforming plasmid pMTase1 in E. coli BL21 (DE3) cells. The protein was purified to homogeneity from soluble extract as His10-Smt3 fusion at N-terminus over Ni-NTA and ion exchange columns, followed by cleavage of the tag by Ulp1 protease as described in the methods section (Suppl Fig. S1). The purified protein was set-up for crystallizations. The structure of Rv2966c was determined to 1.9 Å resolution by molecular replacement using the coordinates of E. faecalis putative
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

6
methyltransferase as the search model and refined to a final R-work of 17.8% and Rfree of 22.1%. The final model of Rv2966c consists of 182 residues of the entire molecule including an additional Ser at the N-terminus obtained as a cloning artifact. Four residues at the C-terminus and three residues in the linker region were disordered and could not be included in the final model. The overall structure of Rv2966c consists of two distinct domains, a short N-terminal domain (NTD) comprising residues 1-16 and a larger C-terminal domain (CTD) comprising residues 25 to 188 resembling the SAM-MT fold present in several SAM-dependent methyltransferases (29) (Fig. 1B). The two domains are linked to each other by a short linker region (residues 17 to 24), which is partially disordered in the present structure.
Several methyltransferases have a domain in addition to the SAM-MT fold that defines its substrate binding ability. In Rv2966c, only a short NTD in addition to the C-terminal SAM domain is present. The NTD is a beta-hairpin comprising a small two-stranded beta-sheet separated by a long loop of nine residues. The NTD appears to be the only region present in the protein structure that is likely to interact with substrate molecules. The NTD is connected to the CTD via a flexible linker region (Fig. 1B). The linker region may enable large movements of the entire NTD to interact with nucleic acid substrates. This small stretch of residues, however, is too short to search for structural homologs in DALI.
In contrast to the variable substrate binding domain, the CTD is a highly conserved structure consisting of the classical SAM-MT fold found in several SAM-dependent methyltransferases (29). It consists of a central eight-stranded β-sheet flanked by α-helices on both sides. The first five strands of the β-sheet are parallel to each other and encompass the SAM binding site towards their C- terminus end (Fig. 1B), while the remaining strands of the β-sheet are antiparallel.
A search for structural homologs of Rv2966c was carried out in DALI using the final coordinates of Rv2966c. Rv2966c shows similarity to methyltransferases belonging to different classes. The top structural matches were found with E. coli RsmD (PDBID 2FPO) and several putative
methyltransferases with structural similarities encompassing both the domains of Rv2966c (Fig. 1C, Suppl Table, ST1). Structures of these homologs have been determined as part of structural biology consortium efforts but the function of these proteins is unknown. Interestingly, the linker region of Rv2966c was found to be disordered in all the homologous structures, except in one subunit of E. faecalis and T. thermophilus structures. An analysis of the coordinate files for each structure also indicated the absence of a bound SAM in any of these structures. Other methyltransferases showing structural similarities with that of Rv2966c belonged to diverse families of methyltransferases. e.g. Precorrin-6Y C5,15- methyltransferase (Z-score 18.8 and r.s.m.d of 2.2 Å for 159 residues), 23S rRNA m5C1962 methyltransferase (Z-score 18.9 and r.s.m.d of 2.6 Å for 170 residues), catechol-O-methyltransferase (Z-score 18.2 and r.s.m.d of 2.6 Å for 164 residues). However, the structural similarities with other methyltransferases were limited to the core SAM-domain but not the target recognition domain and hence are not discussed further.
S-adenosyl methionine (SAM) binding site
Superposition of Rv2966c structure with other SAM dependent methyltransferases helped identify SAM binding pocket. Residues in “YAGSGALG” nucleotide binding motif are part of the SAM binding site on one side while the linker region is present at the other end of the SAM binding pocket. Although SAM was included in the crystallization conditions of Rv2966c, an unambiguous density for SAM in this pocket could not be identified. Absence of SAM in the binding region appears to have resulted in a partially disordered binding pocket (missing density for linker region). However, SAM could be modeled very well into this binding pocket replacing a network of water molecules present in the SAM binding pocket (Fig 2). As shown in the figure, residues of the SAM-binding motif as well as the catalytic motif are well positioned to interact with the SAM molecule.
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

7
In vivo activity: Rv2966c complements E. coli RsmD
To determine the in vivo activity of Rv2966c, pMTase3 was transformed into rsmD-deleted E. coli KL16 strain (16) and the protein was induced with 0.2% arabinose at 37°C for 3 hours. Total RNA was isolated from the induced cells and used for primer extension analysis as described in materials and methods. Primer extension by reverse transcriptase stops 3’ to a base methylated at positions involved in Watson-Crick base pairing like m2G or m2A. Hence if G966 is methylated at N2, primer extension will stop at C967. As can be seen from Fig. 3, extension of primer by reverse transcriptase stops at nucleotide C967 when the cells are transformed with pMTase3 (lane 4) but not when it is transformed with vector alone (lane 2). rv2966c is hence able to complement E. coli KL16ΔrsmD strain by methylating G966 at N2 position and inhibiting extension of primer at C967. An identical result is obtained in wild type strain (Fig. 3, lane 5) whereas there is read through in the rsmD-deleted strain (Fig 3, lane 1). Primer extension reaction carried out on total RNA purified from E. coli KL16ΔrsmD and methylated in vitro with recombinant Rv2966c resulted in read through at the G966 position (Fig3, lane3), suggesting that Rv2966c failed to methylate G966 at N2 in this reaction.
In vitro activity of Rv2966c
E. coli RsmD as well as other nucleotide-specific RNA methyltransferases, viz. RsmC and RsmE that modify m2G1207 and m3U1498 of 16S rRNA respectively (30,31) lack in vitro methyltransferase activity in the absence of ribosomal subunits. In order to monitor the in vitro activity of Rv2966c, Rv2966c was incubated with 30S ribosomal subunits in presence of SAM and the methyltransferase activity was monitored colorimetrically. As shown in Fig. 4, an increase in absorbance at 510 nm, indicating the methyltransferase reaction, occurs when Rv2966c is incubated with purified 30S ribosome from the rsmD-deleted E. coli KL16 strain. No in vitro methyltransferase activity was observed when Rv2966c was incubated with 30S ribosomal
subunit of E. coli KL16ΔrsmD complemented with pMTase3 indicating that RNA was already methylated in vivo.
Although the necessity of ribosome for in vitro methylase activity is unclear, it is likely that the small N-terminal domain of Rv2966c is incapable to binding efficiently to RNA and requires additional factors for stable binding in order to carry out the methylase reaction. Another possibility is that Rv2966c has a preference for the short stem and loop structure containing G966 nucleotide position in 16S rRNA. Whether the base recognition is sequence, RNA secondary-structure or ribosomal-position specific, remains to be ascertained.
N-terminus of Rv2966c is the target recognition region that helps in binding to nucleic acids: Rv2966c has a conserved stretch of about 16 amino acid residues at its N-terminus which is not part of the known ‘methyltransferase’ domain (Fig. 1). We surmised that this conserved stretch of amino acid residues might have an important role to play which was not determined yet. We addressed this by making two truncations of 18 and 23 amino residues from N-terminus and purifying the recombinant truncated proteins Δ18RM and Δ23RM from their respective expression vectors, pMTase4 and pMTase5 as described in the methods section.
Quaternary structures of the full length (WT) and truncated proteins were determined by doing size exclusion chromatography (data not shown). All the three proteins eluted from the column as monomers. The overall stability and correct folded state of the proteins was confirmed by Circular Dichroism (Fig. 5A).
In order to explore the role of NTD in nucleic acid binding, nucleic acid binding ability of Rv2966c was determined by EMSA (Fig 5B). Both DNA and RNA were used for binding assay and the protein-nucleic acid complex was analyzed on agarose gel by ethidium bromide staining (Fig. 5B). Full length Rv2966c (WT) binds to RNA as can be seen by the gradual slowing of the migration of rRNA band on the gel (Fig. 5B, upper panel, lanes 2-4) and the binding is concentration
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

8
dependent. Surprising result is the formation of complex between DNA and Rv2966c (Fig. 5B, lower panel, lanes 2-4). Deletion of the N-terminal hairpin prevents binding to DNA or RNA (Fig 5B, lanes 5-10, upper and lower panels).
Binding of protein to DNA is much stronger than it is to RNA as is evident from the much slower migration of DNA substrate on gel even with less amount of protein (Fig 5B). Ability of Rv2966c to bind to DNA with a higher apparent affinity than RNA is surprising since no methyltransferase activity onto substrate DNA molecules by Rv2966c has been identified so far.
N-terminal truncated proteins lack in vivo and in vitro m2G966 methyltransferase activity: As the truncated proteins did not bind to RNA, we wanted to determine their m2G966 methyltransferase activity. To monitor the in vivo activity of Δ18RM and Δ23RM, plasmids pMTase6 and pMTase7 were transformed into E. coli KL16ΔrsmD. Activity of the truncated proteins was determined by doing a primer extension assay on total RNA isolated from the transformed strain as described before. The truncated proteins did not show any m2G966 methyltransferase activity as seen in Fig 5C (lanes 2, 4) as there is no inhibition of primer extension by reverse transcriptase at C967 in these lanes whereas there is clear termination at the same position with wild type (full length) protein (Fig. 5C, lane 3), as expected. A western blot was done with the total cell extract of E. coli KL16ΔrsmD cells expressing Δ18RM and Δ23RM (or 3RM) to find out whether the lack of complementation was because of low level of expression of the truncated (or mutated) proteins. As seen in Fig. 5D, expression level of all proteins was found to be similar.
Incubation of the 30S ribosomes from E. coli KL16ΔrsmD with Δ18RM or Δ23RM indicated that the truncated Rv2996c lacked in vitro methyltransferase activity as well (Fig. 4).
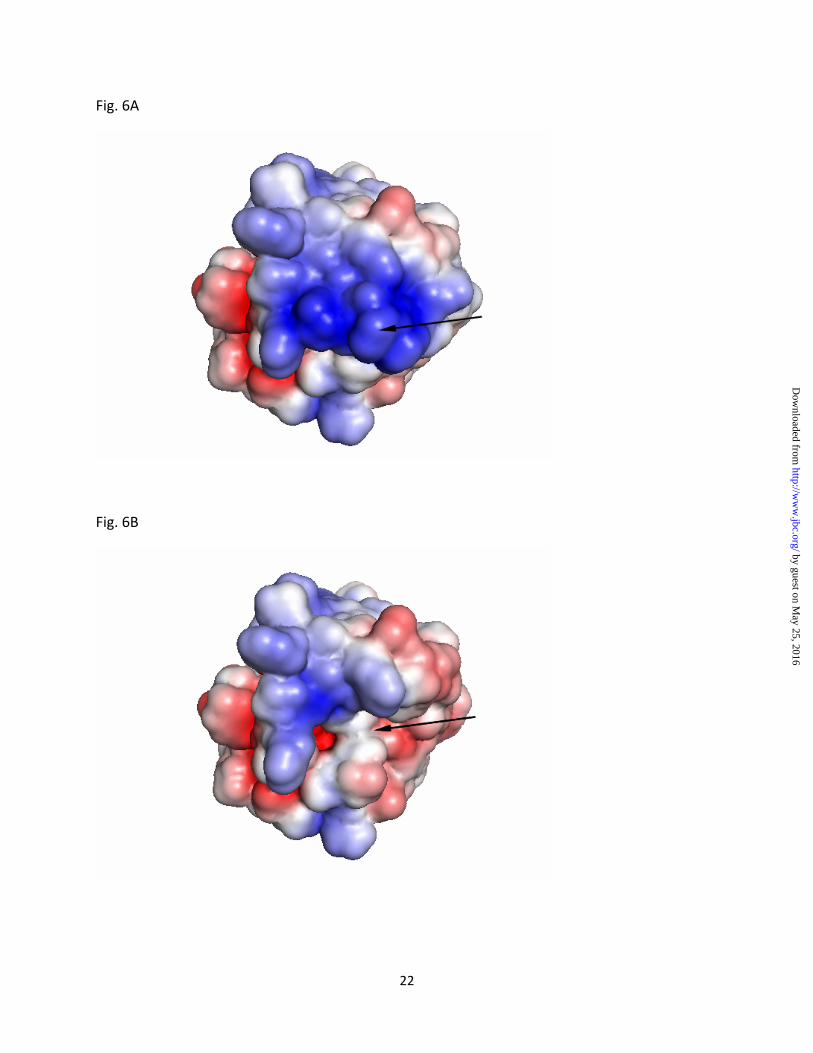
Mutant protein 3RM does not have in vitro DNA binding activity: Electrostatic surface representation of nucleic acid binding proteins is a
good indicator of the binding region on the protein. An electrostatic surface representation of Rv2966c shows a region of positive potential for the N-terminal domain that may interact with negatively charged backbone of nucleic acid substrates (Fig. 6A). An electrostatic surface generated for the N-terminal truncated protein (Fig. 6B), on the other hand, shows a marked reduction in the positively charged region, which may lead to loss of interaction with 16S rRNA. To probe the role of positively charged residues of the NTD in nucleic acid binding, a triple Arg-mutant of Rv2966c, 3RM, was made mutating conserved arginine residues at positions 3, 12 and 13 to alanine. The mutated protein 3RM did not bind to both DNA and RNA in vitro (Fig. 7A) and had no m2G966 activity in vivo (Fig 7B).
Discussion:
Ribosomal RNAs play very important roles in the structure and function of ribosomes. Both 23S and 16S rRNAs contain several highly modified nucleotides (32). Tertiary structure of ribosomes has revealed that the modified nucleotides are located at important functional sites underlying their importance in ribosome biochemistry (33). Here we have identified Rv2966c from Mycobacterium tuberculosis as an ortholog of RsmD of E. coli. We show that Rv2966c can complement RsmD in a mutant E. coli strain. Rv2966c can also use 30S ribosomes of E. coli as substrate and methylates N2-G966 of 16S rRNA. Ability of Rv2966c to use E. coli ribosomes as substrate shows conservation of not only the tertiary structure of ribosomes and the proteins but also the nature of interactions between RsmD and the components of ribosomes between the two organisms. There is not much conservation of the primary structures between the two proteins as has also been seen with other Rsm homologs (24) but the structure and the amino acid residues at the conserved motifs are highly similar. Open reading frame of Rv2966c consists of 188 amino acid residues with a conserved typeI Rossman fold SAM-binding domain from 17-182 amino acid residues. This domain contains SAM-binding motifs 51YAGSG55 and a conserved active site 119DPPY123, present in nucleic acid
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

9
methyltransferases (23). Homologs of Rv2966c are present in all the mycobacterial species including M. leprae, a strain of mycobacterium which has lost 33% of its genome. It shows that the protein might play an important role in mycobacterial physiology. rsmD-deleted E. coli cells do not show any significant phenotype except modest decrease in growth rate (14). This could be because of the presence of modifications in other nucleotides in the same loop region of 16S rRNA at the P-site binding pocket of the ribosome. As these modified bases are positioned together at important sites in ribosomes, absence of one modification does not make any significant difference in cellular growth under normal condition. These modifications might give growth advantage to the bacteria under stress. M. tuberculosis and several other mycobacterial species spend much of their life cycle within human (host) macrophages and Rv2966c (or its homologs) might have a role in bacterial survival at that time.
Rv2966c shows no detectable activity on RNA in absence of ribosomes or when ribosomes are isolated from wild type E. coli or rsmD-complemented cells where the RNA is already methylated. Substrate specificity of RsmD has been attributed to the interaction between the protein and ribosomes (14). The NTD seems necessary and sufficient to enable this interaction with ribosomes as seen by the loss of methyltransferase activity both in vivo and in vitro in both the truncated and mutant proteins. The conserved arginines play an important role in binding to the negatively charged nucleic acid substrates. Role of arginines in the NTD was confirmed as the mutant protein 3RM did not bind to nucleic acids in vitro and also lacked any in vivo m2G966 activity.
Rv2966c, like RsmD, has a very short N-terminal extension in the form of a short beta-hairpin comprising a small two-stranded beta-sheet separated by a long loop. Other rRNA methyltransferases like RlmG and RlmL have longer N-terminal extensions which have been shown to be involved in RNA substrate binding (34,35). Beta-hairpins play an important role in interactions with several nucleic acids. Interestingly, Rv2118c, an m1A58 tRNA
methyltransferase contains three-tandem beta-hairpins as the only target recognition region in addition to the SAM-MT fold (36,37) although the nature of interactions of the hairpin with tRNA is unclear. Flexible beta-hairpins of architectural proteins, HU and IHF also take part in stabilizing interactions with unusual bent DNA (38,39). The flexible linker region of Rv2966c is likely to allow large conformational movements of the N-terminal β-hairpin domain to enable specific interactions with RNA near the P-site of the ribosome.
The NTD of Rv2966c is important for binding not only RNA but also DNA substrates. Somewhat surprisingly, Rv2966c shows stronger binding to DNA than RNA under given conditions. Truncated constructs of Rv2966c lacking the N-terminal 18 or 23 amino acid residues and the mutant protein 3RM did not show any significant binding to either DNA or RNA (Fig. 4B, Fig. 7A). The proteins did not show any m2G966 methyltransferase activity confirming an important role of N-terminus region in target recognition and nucleic acid binding. It is worth noting that the stretch of amino acid residues at N-terminus are fairly conserved in RsmD homologs in different organisms including mycobacterium (although annotated reading frames of many of these homologs including MSMEG_2413 and ML1664 need to be corrected by including additional amino acid residues at N-terminus) (Suppl Fig. S2). The NTD of Rv2966c represents the smallest functional domain to the best our knowledge.
It is provocative to think that Rv2966c and its orthologs are also involved in methylating DNA. Rv2966c has a strong binding to DNA and the complex is stable even at 65°C (data not shown) and also contains the conserved ‘DPPY’ motif at its active site which is present in many other DNA methyltransferases (26,27). Moreover, it seems unlikely for cells to maintain Rv2966c or its homologs at a high metabolic cost just for one function, which has already been shown to be dispensable without significant phenotypic changes in E. coli. A single methyltransferase methylating more than one substrate is not without precedence. Eukaryotic Dnmt2 has been shown to methylate both tRNA and DNA (40). Methylation of specific nucleotides on tRNA in Drosophila protects them from stress-induced cleavage (41).
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

10
Even the bacterial exocyclic DNA methyltransferases specific for adenine or cytosine methylates the other purine nucleotide (42,43).
m2G966 is located at P-site of ribosomes and binds to the tip of tRNA anticodon (44). P-site in bacteria is target of many antibiotics and modifications of nucleotides at the site has been
shown to play a role in drug sensitivity of the organism. We have started to learn more about their roles with the solution of tertiary structure of ribosomes. It will be interesting to further explore the role of methylation at G958 (equivalent position to m2G966 of E. coli 16S rRNA) and that of Rv2966c in M. tuberculosis drug resistance.
References
1. Maher D, and Raviglione, M. Clin. Chest Med (2005) 26: 167-182.
2. World Health Organization. Report WHO/HTM/TB/2006.361.: World Health Organization, Geneva, Switzerland., 2006.
3. Dye C, Espinal, MA, Watt, CJ, Mbiaga, C, and Williams, BG. Journal of Infectious Diseases. (2002) 185:1197-1202.
4. Shah NS, Wright A, Bai GH, Barrera L, Boulahbal F, Martín-Casabona N, Drobniewski F, Gilpin C, Havelková M, Lepe R, Lumb R, Metchock B, Portaels F, Rodrigues MF, Rüsch-Gerdes S, Van Deun A, Vincent V, Laserson K, Wells C, and Cegielski JP. Emerg. Infect. Dis. (2007) 13:380-387.
5. Alangaden GJ, Kreiswirth, BN, Aouad, A, Khetarpal, M, Igno, FR, Moghazeh, SL, Manavathu, EK, and Lerner, SA. Antimicrob. Agents Chemother. (1998) 42: 1295-1297.
6. Suzuki Y, Katsukawa C, Tamaru A, Abe C, Makino M, Mizuguchi Y, and Taniguchi, H. J. Clin. Microbiol. (1998) 36: 1220-1225.
7. Rastogi N, Goh, KS, Berchel, M, and Bryskier, A. Antimicrob Agents Chemother (2000) 44:2848-2852.
8. Falzari K, Zhu, Z, Pan, D, Liu, H, Hongmanee, P, and Franzblau, SG. Antimicrob Agents Chemother (2005) 49: 1447-1454.
9. Buriánková K, Doucet-Populaire, F, Dorson, O, Gondran, A, Ghnassia, JC, Weiser, J, and Pernodet, JL. Antimicro. Agents Chemother (2004) 48: 143-150.
10. Johansen, SK, Maus, CE, Plikaytis, BB, and Douthwaite, S. Mol Cell (2006) 23:173-182.
11. Okamoto, S, Tamaru, A, Nakajima, C, Nishimura, K, Tanaka, Y, Tokuyama, S, Suzuki, Y, and Ochi, K. Mol Microbiol (2007) 63:1096-1106.
12. Jugheli, L, Bzekalava, N, de Rijk, P, Fissette, K, Portaels, F, and Rigouts L. Antimicrob Agents Chemother (2009) 53: 5064-5068.
13. Andersen, NM, and Douthwaite, S. J Mol Biol (2006) 359: 777-786.
14. Lesnyak, DV, Osipiuk, J, Skarina, T, Sergiev, PV, Bogdanov, AA, Edwards, A, Savchenko A, Joachimiak, A, andDontsova OA. J Biol Chem (2007) 282: 5880-5887.
15. Guzman, LM, Belin, D, Carson, MJ, and Beckwith, J. J Bacteriol (1995) 177: 4121-4130.
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

11
16. Das, G, Thotala, DK, Kapoor, S, Karunanithi, S, Thakur, SS, Singh, NS, and Varshney, U. Embo J (2008) 27: 840-851.
17. Gutgsell, N, Deutscher, MP, and Ofengand, J. RNA (2005) 11:1141-1152.
18. Leslie, AGW. Joint CCP4 + ESF EAMCB Newsletter on Protein Crystallography. (1992) 26.
19. Collaborative Computational Project 4. Acta Crystallogr (1994) D50: 760-763.
20. Emsley, P, and Cowtan, K. Acta Crystallogr (2004) D60: 2126-2132.
21. Murshudov, GN, Vagin, AA, and Dodson, EJ. Acta Crystallogr (1997) D53: 240-255.
22. Holm L, and Sander, C. J Mol Biol (1993) 233:123-38.
23. Malone, T, Blumenthal, RM, and Cheng, X. J Mol Biol (1995) 253:618-32.
24. Schubert HL, Blumenthal, RM, and Cheng, X. Trends Biochem Sci (2003a) 28:329-335.
25. Bheemanaik S, Reddy YV, Rao DN. Biochem J (2006) 399:177-90.
26. Goedecke, K , Pignot, M, Goody, RS, Scheidig, AJ, and Weinhold, E. Nat Struct Biol (2001) 8: 121-125.
27. Gong, W, O'Gara, M, Blumenthal, RM, and Cheng, X. Nucleic Acids Res (1997) 25:2702–2715.
28. Schubert, HL, Phillips, JD, and Hill CP. Biochemistry (2003b) 42: 5592-5599.
29. Martin, JL, and McMillan, FM. Curr Opin Struct Biol (2002) 12:783-93.
30. Tscherne, JS, Nurse, K, Popienick, P, and Ofengand, J. J Biol Chem (1999) 274:924-9.
31. Basturea, GN, and Deutscher, MP. RNA (2007) 13:1969-76.
32. Brimacombe, R, Mitchell, P, Osswald, M, Stade, K, and Bochkariov D. FASEB J, (1993) 7:161-7.
33. Decatur, WA, and Fournier, MJ. Trends Biochem Sci (2002) 27: 344–351.
34. Sergiev, PV, Lesnyak, DV, Bogdanov, AA, and Dontsova OA. J Mol Biol (2006) 364:26-31.
35. Lesnyak, DV, Sergiev, PV, Bogdanov, AA, and Dontsova, OA. J Mol Biol (2006) 364: 20-25.
36. Gupta, A, Kumar, PH, Dineshkumar, TK, and Varshney, U, Subramanya, HS. J Mol Biol (2001) 312: 381-91.
37. Varshney, U, Ramesh, V, Madabushi, A, Gaur, R, Subramanya, HS, RajBhandary, UL. Nucleic Acids Res (2004) 32: 1018-27.
38. Swinger, KK, Lemberg, KM, Zhang, Y, Rice, PA. EMBO J (2003) 14: 3749-60.
39. Rice, PA, Yang, S, Mizuuchi, K, Nash HA. Cell (1996) 87:1295-306.
40. Schaefer, M, and Lyko, F. Chromosoma (2010) 119: 35-40.
41. Schaefer, M, Pollex, T, Hanna, K, Tuorto, F, Meusburger, M, Helm, M, Lyko, F. Genes Dev (2010) 24: 1590-1595.
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

12
42. Jeltsch, A, Christ, F, Fatemi, M, and Roth M. J Biol Chem (1999) 274: 19538-19544.
43. Jeltsch A. Biol Chem. (2001) 382: 707-710.
44. Selmer, M, Dunham, CM, Murphy, FV 4th, Weixlbaumer, A, Petry, S, Kelley, AC, Weir, JR, and Ramakrishnan, V. Science (2006) 313:1935-1942.
45. Baker, NA, Sept, D, Joseph, S, Holst, MJ, and McCammon, JA. Proc Natl Acad Sci, USA. (2001) 98: 10037-10041.
46. Diederichs, K, and Karplus, PA. Nat. Struct. Biol. (1997) 4: 269-275.
Footnotes:
#We would like to thank Dr. S. Ramachandran and Dr. Rakesh Sharma for critical evaluation of the manuscript. We thank Dr. Rajesh Gokhale and Prof. NK Ganguly for their help, suggestions and support for the project, colleagues at IMM and IGIB for kind help with reagents, Santosh Kumar for help with CD experiments, Prof. Umesh Varshney for his generous gift of rsmD knockout strains of E. coli, Prof. Stewart Shuman for pET28-His10-Smt3 plasmid and the wonderful team at BM14, ESRF for data collection. Institutional start-up fund from IMM to KMS is gratefully acknowledged. This work was supported by FAC-02 project grant of CSIR to BT.
The abbreviations used are S-Adenosyl Methionine, SAM, M. tuberculosis, M. tb; NTD, N-terminal domain; CTD, C-terminal domain
Figure Legends:
Figure 1: Rv2966c: overall structure and comparisons
(A) Sequence alignment of Rv2966c homologs: Sequence alignment of Rv2966c and its structural homologs as identified from DALI. Sequences are shown in same color as their structural counterparts in (C). The conserved F/YXGXG motif in the SAM-binding region and DPPY/F in the catalytic region are marked in gray boxes. The sequential arrangement of Target recognition region, SAM-binding region and Catalytic region classifies the protein to be similar to the ζ-class of DNA methyltransferases (Malone et al, 1995).
(B) Stereo diagram of the overall structure of Rv2966c showing the NTD (magenta), linker region (yellow) and the CTD containing the SAM-MT fold (cyan). Rv2966c is a SAM-dependent methyltransferase. The conserved YAGSG motif in the SAM-binding region is shown in black. A SAM molecule, shown in sticks has been modeled in the putative SAM-binding pocket formed towards the C-terminus of the parallel β-strands. Residues at N-terminus (Ser), C-terminus (Leu) and at the break in linker region (Thr) are labeled.
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

13
(C) Structure superposition of Rv2966c homologs. Superposition of Rv2966c with H. influenzae methyltransferase, PDBID: 2IFT (green), E. coli RsmD, PDBID: 2FPO (red), E. faecalis methyltransferase, PDBID: 2FHP (orange), S. pyogenes methyltransferase, PDBID: 2ESR (purple), T. thermophilus methyltransferase, PDBID: 1WS6 (gray) shows that all the structures have a same overall fold. Rv2966c is shown in same color notations as Fig. 1B.
Figure 2: Stereo view of SAM binding pocket of Rv2966c.
(A) A network of waters, marked W, occupies the SAM-binding pocket. (B) SAM can be modeled into this pocket into positions occupied by waters in the current structure. The putative H-bonding interactions of the modeled SAM are shown as dashed lines. An acetate molecule, ACT is well placed to interact with the 6-amino group of the adenosine moiety of SAM. Tyr-51 of the Y/FXGXG motif stacks against the adenosine of SAM molecule. Residues, Asp-119, Pro-120 and Pro-121 of the DPPY/F motif are present close to the SAM binding pocket. The side chain of Tyr-122 of the DPPY/F motif is disordered in the present structure.
Figure 3: Primer extension analysis for in vivo activity of Rv2966c.
Primer extension reaction was carried out using AMV reverse transcriptase to monitor the in vivo methylation carried out by Rv2996c. Products were analyzed by running a denaturing polyacrylamide gel and visualized by phosphoimaging. Primer extension products in the different lanes correspond to reactions carried out on total RNA isolated from E. coli KL16ΔrsmD (lane 1), E. coli KL16ΔrsmD containing pBSmt2 expressing only the His10-Smt3 tag (lane 2), in vitro reaction of total RNA from E. coli KL16ΔrsmD incubated with purified Rv2966c (lane 3), E. coli KL16ΔrsmD complemented with pMTase3 (lane 4), parent E. coli KL16 strain (lane 5). Lanes 6-9 contain contain 32P-end labeled oligonucleotide markers ME1, ME2, ME3 and ME4 corresponding to sequence 965-996, 966-996, 967-996 and 968-996 of lengths 32, 31, 30 and 29 nucleotides, respectively, to ascertain the size of extension products (see text for more details). Position and size of ME1 and ME4 are indicated.
Figure 4: In vitro activity of Rv2966c
In vitro methyltransferase activity of Rv2966c was monitored colorimetrically as described in the Methods section. A methyltransferase reaction was carried out by incubating Rv2966c (filled diamonds), Δ18RM (filled square) or Δ23RM (filled triangles) with purified 30S ribosome subunit from E. coli KL16ΔrsmD and 100 μM SAM at 37°C for 1h. No in vitro methyltransferase activity was observed when Rv2966c was incubated with 30S ribosomal subunit of E. coli KL16ΔrsmD complemented with pMTase3 (open squares).
Figure 5: Role of NTD in binding and activity.
(A) CD analysis: CD of Rv2966c, Δ18RM and Δ23RM (top, middle and bottom panels, respectively) indicates secondary structures which is suggestive of folded protein.
(B) EMSA for nucleic acid binding
Binding of Rv2966c and truncated proteins to DNA and RNA was analyzed by running the reaction mixture on agarose gel and visualizing by ethidium bromide staining. Full length Rv2966c (WT) binds
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

14
RNA (upper panel) and DNA (lower panel) (lanes 2-4) in a concentration dependent manner. Truncated proteins lacking the N-terminal hairpin, Δ18RM (lanes 5-7) and Δ23RM (lanes 8-10) fail to bind the nucleic acids. A control with no protein is indicated in lane 1 in both the panels. In the reaction mix, 100, 200 or 400 ng of full length (Rv2966c) or truncated proteins (Δ18RM and Δ23RM) were incubated with RNA at 30°C for 60 min (see Methods section for more details). For DNA binding, a similar reaction was carried out with 50, 100 or 200 ng of full length (Rv2966c) or truncated proteins (Δ18RM and Δ23RM) to monitor gel shift. Position and sizes of linear DNA markers (in kilobases) is indicated in lane M.
(C) In vivo activity of truncated proteins
In vivo activity of Δ18RM or Δ23RM was monitored by primer extension analysis using AMV reverse transcriptase and analyzing the reaction products on denaturing polyacrylamide gel. Gel was visualized by phosphoimaging. Reactions were carried out with RNA purified from E. coli KL16ΔrsmD (lane 1), E. coli KL16ΔrsmD transformed with pMTase6 expressing Δ18RM (lane 2), E. coli KL16ΔrsmD transformed with pMTase3 expressing Rv2966c (full length) (lane 3), E. coli KL16ΔrsmD transformed with pMTase7 expressing Δ23RM (lane 4). Lanes 5-8: contain 32P-end labeled oligonucleotide markers ME1, ME2, ME3 and ME4 corresponding to sequence 965-996, 966-996, 967-996 and 968-996 and lengths 32, 31, 30 and 29 nucleotides, respectively to determine the size of extension products. Position and size of ME1 and ME4 are indicated.
(D) Expression level of wild type, deleted and mutant Rv2966c are similar:
Upper panel: Western blot of total cell extract of E. coli KL16DrsmD cells expressing full length Rv2966c (WT), lane 1, Δ18RM (lane 2), Δ23RM (lane 3) and 3RM (lane 4) shows an equivalent level of expression for all proteins. The blot was probed with anti-His. Lower panel: The same blot was probed with anti-DnaK which was used as loading control.
Figure 6: Molecular surface representation of Rv2966c
(A) The electrostatic surface potential of Rv2966c shows the NTD (marked by arrow) to have a positively charged surface. (B) An electrostatic surface generated for Rv2966c lacking the NTD (deleted region marked by arrow) shows a reduction in positive surface on the protein. The electrostatic potential was calculated with the program APBS (Baker et al, 2001). The surface is colored with a blue to red gradient from +3.5 to -3.5 KbT/e.
Figure 7: Role of conserved arginines in binding and activity
(A) EMSA for nucleic acid binding.
Binding of Rv2966c and 3RM to DNA and RNA was analyzed by running the reaction mixture on agarose gel and visualizing by ethidium bromide staining. 3RM fails to bind RNA (upper panel, lanes 5-7) or DNA (lower panel, lanes 5-7). A control with no protein is indicated in lane 1 in both the panels, while concentration dependent binding of nucleic acids with Rv2966c (WT) is seen in lanes 2-4. In the reaction mix, 100, 200 or 400 ng of Rv2966c (WT) or mutant protein, 3RM were incubated with RNA at 30°C for 60 min (see Methods section for more details). For DNA binding, a similar reaction was carried out with 50, 100 or 200 ng of Rv2966c (WT) or mutant protein (3RM) to monitor gel shift. 3RM, like Δ18RM and Δ23RM (Fig. 5) fails to bind both DNA and RNA. A minor difference in appearance of bands from figure 5 is due to gel running artefact. Position and sizes of linear DNA markers (in kilobases) is indicated.
(B) In vivo activity of mutant protein
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

15
In vivo activity of 3RM was monitored by primer extension analysis using AMV reverse transcriptase and analyzing the reaction products on denaturing polyacrylamide gel as described earlier. Reactions were carried out with RNA purified from E. coli KL16ΔrsmD (lane 1), E. coli KL16ΔrsmD transformed with pMTase3 expressing full length Rv2966c (WT) (lane 2), E. coli KL16ΔrsmD transformed with pMTase8 expressing 3RM (lane 3). Position and size of markers ME1 and ME4, corresponding to sizes 32 and 29 nucleotides are indicated.
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

16
Table 1: Data collection and model refinement statistics
Crystal and Data Collection
Space group P32 2 1 Cell dimensions A=b=54.78 Å, c=100.04
Å, α=β=90°, γ=120°
Resolution (Å)a 27.39 – 1.9 (2.0 – 1.9)
Number of unique reflections 14232 (955)
Completeness (%) 99.94 (100.0) Multiplicity 10.7 (10.8) Rmeas (%)
b 9.3 (33.6)
<<I>/σ(<I>)>c 4.9 (2.3) Refinement
Resolution (Å) 27.39 -1.9 (1.95 – 1.9)
Number of reflections working/test
13511 / 712
R-work (%)d 17.8 (18.2) Rfree(%)e 22.1 (19.8) Protein residues/atomsf 182/1374 Alternate conformations 4 Water molecules 181 Other atoms 2 acetates Wilson B-factor (Å2) 17.1 B-factor (Å2)
Protein atoms (chain) 15.5 Water molecules 27.3
r.m.s. deviations bond lengths (Å) 0.015 bond angles (°) 1.53 Ramachandran plot Most Favored regions (%) 96.7 Disallowed regions (%) 0.0
aNumbers in paranthesis correspond to highest resolution shell. bRmeas as described in Diederichs and Karplus (46). c<<I>/σ(<I>)> = Mean Ih over the standard deviation of the mean Ih averaged over all reflections in a resolution shell. dRwork= ∑ ||Fo| – |Fc||/ ∑|Fo|, where |Fo| is the observed structure factor amplitude and |Fc| the calculated structure factor amplitude. eRfree: Rfactor based on 5% of the data excluded from refinement. fTotal number of protein atoms, including those in alternate conformations.
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from
17
Fig. 1A
Fig. 1B
Fig. 1C
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

21
Fig. 5A Fig. 5B
Fig. 5C Fig. 5D
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from
22
Fig. 6A
Fig. 6B
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

23
Fig. 7A
Fig. 7B
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from

1
Structural and functional characterization of Rv2966c reveals an RsmD-like methyltransferase from M. tuberculosis and the role of its N-terminal domain in target recognition
Kumar, A et al (2010)
Supplementary Data:
Supplementary Figure S1: Analysis of aliquots (3μg each) of Rv2966c (WT) (lane 1), Δ23RM (lane 2), Δ18RM (lane 3) and 3RM (lane 4) by SDS−PAGE. The Coomassie blue-stained gel is shown. Lane 5 contains a molecular weight marker; the sizes (in kDa) of marker proteins are indicated on the right.
Supplementary Figure S2: Sequence comparison of N-terminal region (residues 1-23) of Rv2966c of M. tuberculosis (M. tb) and its orthologs from M. smegmatis, MSMEG_2413 and M. leprae, ML1664 show strong conservation of the N-terminal target recognition region. The respective annotated reading frames of both orthologs had to be corrected to include the additional residues in this region.

2
Supplementary Table ST1: Structure comparisons of Rv2966c with DALI
A search for structural homologs of Rv2966c with DALI yielded hits with several methyltransferases. The top hits among these were with RsmD and its putative homologs. Pariwise r.m.s deviations (in Å) obtained upon superposing structurally equivalent Cα atoms are given below. Other methyltransferases with similarities with the core C-terminal SAM-domain were not further analyzed.
aR.m.s.d. is the root mean square deviation obtained upon superposition of respective methyltransferase structures against that of Rv2966c.
PDB ID 2FHP 2FPO 1WS6 2IFT 2ESR
Organism E. faecalis E. coli T. thermophillus H. influenzae S. pyogenes
Z-score 26.7 26.6 24.5 23.7 21.4
R.m.s.da 1.5 1.7 1.6 1.9 1.6
Residues aligned
174 177 167 170 151
% identity 30 33 31 28 29

Atul Kumar, Kashyap Saigal, Ketan Malhotra, Krishna Murari Sinha and Bhupesh Tanejarecognition
targetmethyltransferase from M. tuberculosis and the role of its N-terminal domain in Structural and functional characterization of Rv2966c reveals an RsmD-like
published online April 7, 2011J. Biol. Chem.
10.1074/jbc.M110.200428Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2011/04/07/M110.200428.DC1.html
http://www.jbc.org/content/early/2011/04/07/jbc.M110.200428.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 25, 2016
http://ww
w.jbc.org/
Dow
nloaded from




















