
Strict regulation of c-Raf kinase levels is required for early organogenesis of the vertebrate inner...
9
Strict regulation of c-Raf kinase levels is required for early organogenesis of the vertebrate inner ear Carmen Sanz 1 , Yolanda Leo´n 1 , Jakob Troppmair 2 , Ulf R Rapp 2 and Isabel Varela-Nieto 1 1 Instituto de Investigaciones Biome´dicas, Consejo Superior de Investigaciones Cientı´ficas (CSIC), Arturo Duperier 4, 28029 Madrid, Spain; 2 Institut fu ¨r Medizinische Strahlenkunde und Zellforschung (MSZ), Bayerische Julius-Maximillians-Universita ¨t, Versbacher Str. 5, 97078 Wu ¨rzburg, Germany Regulation of organogenesis involves a dynamic balance of the mechanisms regulating cell division, dierentiation and death. Here we have investigated the pattern of expression of c-Raf kinase in the inner ear during early developmental stages and the consequences of manipulat- ing c-Raf levels by misexpression of c-raf viral vectors in organotypic cultures of otic vesicle explants. We found that otic vesicles expressed c-Raf and its level remained constant during embryonic days 2 and 3 (E2-E3). c-Raf activity was increased in response to insulin like growth factor-I (IGF-I) and the activation by IGF-I of the c-Raf kinase pathway was a requirement to turn on cell proliferation in the otic vesicle. Overexpression of c-raf in E2.5 explants increased the proliferative response to low serum and IGF-I and blocked dierentiation induced by retinoic acid. The increase in c-Raf levels also prevented nerve growth factor (NGF)-dependent induc- tion of programmed cell death. Consistent with these results, the expression of a dominant negative c-Raf mutant potentiated retinoic acid action and decreased the rate of cell proliferation. We conclude that a strict control of c-Raf levels is essential for the co-ordination of the biological processes that operate simultaneously during early inner ear development. Keywords: insulin-like growth factor-I; mitogen-acti- vated protein kinase; NGF; organotypic otic vesicle cultures; RCAS vectors; retinoic acid Introduction c-Raf (c-Raf-1) is a cytoplasmic protein serine/ threonine kinase involved in signal transduction from the plasma membrane to the nucleus (Mark and Rapp, 1984; Daum et al., 1994; Naumann et al., 1997). c-Raf has a key role in the response to many growth factors and stimulators of cellular proliferation (Magnuson et al., 1994; Alessi et al., 1994; Marshall, 1994) and in cell transformation by oncogenes (Troppmair et al., 1994; Cacace et al., 1996). c-Raf regulates the mitogen- activated protein (MAP) kinase pathway by phosphor- ylating and activating MAP kinase kinase, which then phosphorylates and activates MAP kinase (Kyriakis et al., 1992; Marshall, 1994). There are three known active members of the Raf family of kinases: c-Raf, A- Raf and B-Raf. A-Raf is expressed in urogenital and kidney tissues, with low levels in the brain. The expression of the B-Raf isoform is highest in the nervous system and testis whereas c-Raf is ubiquitously expressed, with high levels found in fetal brain (Storm et al., 1990). c-Raf is a member of a small family of proteins that has been highly conserved during evolution from Drosophila to mammals. In avian cells there is an homologue of c-raf termed c-mil (Jansen et al., 1984; Koenen et al., 1988). Raf proteins are necessary for vulval development in Caenorhabditis elegans (Georgi et al., 1990; Han et al., 1993), R7 photoreceptor formation in Drosophila melanogaster (Perrimon and Desplan, 1994; Brunner et al., 1994; Wassarman et al., 1996) and mesoderm induction in Xenopus laevis (MacNicol et al., 1995; Xu et al., 1996). Transgenic chimeric c-Raf-deficient mice show growth retardation (Naumann et al., 1997) and B-Raf-deficient mice die in utero and display defects in endothelial cell dierentiation and survival (Wojnowski et al., 1997). Hence, it has been proposed that Raf kinases are essential for growth and development (Daum et al., 1994; Naumann et al., 1997), but the detailed functional role of c-Raf kinase in early organogenesis remains to be investigated. The vertebrate inner ear develops from the embryonic otic vesicle. At early stages of develop- ment, a patch of the head ectoderm thickens and forms the otic placode. The otic placode then invaginates and deepens to form a pear-shaped structure, the otic vesicle or otocyst. This is a transient structure that undergoes a distinct period of intense cell proliferation (stages 18 – 20, Hamburger and Hamilton, 1951) which precedes the dierentiation of the various cell types and compartments that will conform the adult inner ear (Bissonnette and Fekete, 1996). The neurons of the cochleovestibular ganglion (CVG) originate at the ventral aspect of the otic vesicle where they organize, become postmitotic and then dierentiate (Hemond and Morest, 1991). The early development of the chicken inner ear can be reproduced in vitro and it is a well characterised model for studying early organogenesis in the sensory organs and nervous system (Bissonnette and Fekete, 1996; Torres and Giraldez, 1998). The basic molecules responsible for normal ontogenesis of the inner ear have started to be unravelled by complementary in vivo and in vitro studies. Organotypic cultures of otic vesicle explants have been the preferred approach to dissect which growth factors and intracellular mediators play physiological roles during inner ear development. IGF- I, NGF and retinoic acid are among the best characterized signals. NGF induces apoptosis in specific areas of the otocyst epithelium and in the Correspondence: I Varela-Nieto Received 4 May 1998; revised 22 July 1998; accepted 23 July 1998 Oncogene (1999) 18, 429 – 437 ª 1999 Stockton Press All rights reserved 0950 – 9232/99 $12.00 http://www.stockton-press.co.uk/onc
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Strict regulation of c-Raf kinase levels is required for early organogenesis of the vertebrate inner...

Strict regulation of c-Raf kinase levels is required for early organogenesisof the vertebrate inner ear
Carmen Sanz1, Yolanda Leo n1, Jakob Troppmair2, Ulf R Rapp2 and Isabel Varela-Nieto1
1Instituto de Investigaciones BiomeÂdicas, Consejo Superior de Investigaciones Cientõ®cas (CSIC), Arturo Duperier 4, 28029Madrid, Spain; 2Institut fuÈr Medizinische Strahlenkunde und Zellforschung (MSZ), Bayerische Julius-Maximillians-UniversitaÈt,Versbacher Str. 5, 97078 WuÈrzburg, Germany
Regulation of organogenesis involves a dynamic balanceof the mechanisms regulating cell division, di�erentiationand death. Here we have investigated the pattern ofexpression of c-Raf kinase in the inner ear during earlydevelopmental stages and the consequences of manipulat-ing c-Raf levels by misexpression of c-raf viral vectors inorganotypic cultures of otic vesicle explants. We foundthat otic vesicles expressed c-Raf and its level remainedconstant during embryonic days 2 and 3 (E2-E3). c-Rafactivity was increased in response to insulin like growthfactor-I (IGF-I) and the activation by IGF-I of the c-Rafkinase pathway was a requirement to turn on cellproliferation in the otic vesicle. Overexpression of c-rafin E2.5 explants increased the proliferative response tolow serum and IGF-I and blocked di�erentiation inducedby retinoic acid. The increase in c-Raf levels alsoprevented nerve growth factor (NGF)-dependent induc-tion of programmed cell death. Consistent with theseresults, the expression of a dominant negative c-Rafmutant potentiated retinoic acid action and decreased therate of cell proliferation. We conclude that a strictcontrol of c-Raf levels is essential for the co-ordinationof the biological processes that operate simultaneouslyduring early inner ear development.
Keywords: insulin-like growth factor-I; mitogen-acti-vated protein kinase; NGF; organotypic otic vesiclecultures; RCAS vectors; retinoic acid
Introduction
c-Raf (c-Raf-1) is a cytoplasmic protein serine/threonine kinase involved in signal transduction fromthe plasma membrane to the nucleus (Mark and Rapp,1984; Daum et al., 1994; Naumann et al., 1997). c-Rafhas a key role in the response to many growth factorsand stimulators of cellular proliferation (Magnuson etal., 1994; Alessi et al., 1994; Marshall, 1994) and in celltransformation by oncogenes (Troppmair et al., 1994;Cacace et al., 1996). c-Raf regulates the mitogen-activated protein (MAP) kinase pathway by phosphor-ylating and activating MAP kinase kinase, which thenphosphorylates and activates MAP kinase (Kyriakis etal., 1992; Marshall, 1994). There are three knownactive members of the Raf family of kinases: c-Raf, A-Raf and B-Raf. A-Raf is expressed in urogenital and
kidney tissues, with low levels in the brain. Theexpression of the B-Raf isoform is highest in thenervous system and testis whereas c-Raf is ubiquitouslyexpressed, with high levels found in fetal brain (Stormet al., 1990). c-Raf is a member of a small family ofproteins that has been highly conserved duringevolution from Drosophila to mammals. In avian cellsthere is an homologue of c-raf termed c-mil (Jansen etal., 1984; Koenen et al., 1988). Raf proteins arenecessary for vulval development in Caenorhabditiselegans (Georgi et al., 1990; Han et al., 1993), R7photoreceptor formation in Drosophila melanogaster(Perrimon and Desplan, 1994; Brunner et al., 1994;Wassarman et al., 1996) and mesoderm induction inXenopus laevis (MacNicol et al., 1995; Xu et al., 1996).Transgenic chimeric c-Raf-de®cient mice show growthretardation (Naumann et al., 1997) and B-Raf-de®cientmice die in utero and display defects in endothelial celldi�erentiation and survival (Wojnowski et al., 1997).Hence, it has been proposed that Raf kinases areessential for growth and development (Daum et al.,1994; Naumann et al., 1997), but the detailedfunctional role of c-Raf kinase in early organogenesisremains to be investigated.The vertebrate inner ear develops from the
embryonic otic vesicle. At early stages of develop-ment, a patch of the head ectoderm thickens and formsthe otic placode. The otic placode then invaginates anddeepens to form a pear-shaped structure, the oticvesicle or otocyst. This is a transient structure thatundergoes a distinct period of intense cell proliferation(stages 18 ± 20, Hamburger and Hamilton, 1951) whichprecedes the di�erentiation of the various cell typesand compartments that will conform the adult innerear (Bissonnette and Fekete, 1996). The neurons of thecochleovestibular ganglion (CVG) originate at theventral aspect of the otic vesicle where they organize,become postmitotic and then di�erentiate (Hemondand Morest, 1991).The early development of the chicken inner ear can
be reproduced in vitro and it is a well characterisedmodel for studying early organogenesis in the sensoryorgans and nervous system (Bissonnette and Fekete,1996; Torres and Giraldez, 1998). The basic moleculesresponsible for normal ontogenesis of the inner earhave started to be unravelled by complementary in vivoand in vitro studies. Organotypic cultures of otic vesicleexplants have been the preferred approach to dissectwhich growth factors and intracellular mediators playphysiological roles during inner ear development. IGF-I, NGF and retinoic acid are among the bestcharacterized signals. NGF induces apoptosis inspeci®c areas of the otocyst epithelium and in the
Correspondence: I Varela-NietoReceived 4 May 1998; revised 22 July 1998; accepted 23 July 1998
Oncogene (1999) 18, 429 ± 437ã 1999 Stockton Press All rights reserved 0950 ± 9232/99 $12.00
http://www.stockton-press.co.uk/onc

CVG (Frago et al., 1998) through its low a�nity p75receptor which is expressed in the inner ear (vonBartheld et al., 1991). Retinoic acid causes growtharrest and cell di�erentiation (Leo n et al., 1995a; Leeand Cotanche, 1996) and IGF-I is a potent growthpromoting and survival factor for the otic vesicle (Leo net al., 1995b; Frago et al., 1998). IGF-I and its higha�nity tyrosine kinase receptor are expressed duringthe proliferative period of otocyst development (Leo net al., 1995b). Although the cellular actions of IGF-Iand NGF in the otic vesicle are beginning to beunderstood, the signalling pathways they trigger stillremain largely unknown. In this context, viral vectorscan be used to deliver both wild type and dominantnegative mutant genes to analyse the functionalconsequences of overexpression (Hughes et al., 1987;Federspiel and Hughes, 1997). Therefore, ectopicexpression of developmentally important genes bymeans of retroviral vectors can provide uniqueinformation on their functional roles during inner eardevelopment (Federspiel and Hughes, 1997; Fekete etal., 1997).Herein we show that the levels of c-Raf kinase are
maintained during the early proliferative stages of theotocyst formation and that IGF-I modulated the Raf/MAP kinase pathway in organotypic cultures. c-Rafkinase activation was essential for otic vesicleproliferation. We further explored the role andimportance of c-Raf kinase by misexpressing both c-
raf and a dominant negative c-raf mutant (Raf-C4)cDNA by means of RCAS retroviral vectors. To thisend, we set up the procedure to misexpress genes inorganotypic cultures of otic vesicles. This has allowedus to show that an increase in c-Raf levels enhancedthe proliferative response to low-doses of mitogens,limited di�erentiation induced by retinoic acid andblocked NGF induction of apoptosis. Conversely,blockade of endogenous c-Raf kinase activity byoverexpression of Raf-C4 potentiated retinoic acide�ects and counteracted IGF-I action. We proposethat a strict control of the levels of c-Raf kinase isessential for the normal progression of inner eardevelopment, thus allowing the system to balance themain biological processes of cell growth, morphogen-esis and apoptosis.
Results
c-Raf kinase pattern of expression and modulation ofactivity
We studied c-Raf protein levels in otic vesicles betweenembryonic stages 17 and 20 (E2 and E3), theproliferative period of the otic vesicle. Figure 1ashows that there was an endogenous expression of c-Raf with no signi®cant changes in c-Raf levels duringthis period. The presence of c-raf transcripts in the
Figure 1 c-Raf expression and activity in the otic vesicle. (a) Immunoblot analysis of c-Raf kinase in otic vesicles. Otic vesicles weredissected at the Hamburger and Hamilton stages indicated. Lysates were analysed by SDS±PAGE and Western blotting asdescribed in Materials and methods. The p74 c-Raf band was quanti®ed by densitometry. Average densitrometrical values areplotted on bars. Results are expressed as percentage of c-Raf ± protein content in stage 19 otic vesicles and they are given asmean+s.e.m. of at least three independent experiments. A representative blot of c-Raf levels at the di�erent stages is shown in theupper row (left) and compared with that of extracts of NIH3T3 control ®broblasts and ®broblasts stably transfected with c-raf(right). (b) Expression of c-raf mRNA in the otic vesicle. Photomicrographs from 60 mm sections of stage 18 chick embryos showingthe otic vesicle and CVG. The samples were processed for in situ hybridization with a digoxigenin labelled c-raf RNA antisenseprobe (right) or a c-raf sense probe (left.) as control. Vibratome sections were obtained in whole mount preparations as described inthe Materials and methods section. Bar, 100 mm. A, anterior; D, dorsal. Photomicrographs are representative of four independentexperiments each involving at least ®ve explants per condition. (c) Immunoblot analysis of phosphorylated MAP kinase (MAPK-P)in explanted otic vesicles. For Western blotting of phosphorylated MAP kinase, otic vesicles were dissected at stage 18 and culturedwithout additives (0S) or in the presence of IGF-I (1 nM) for 15 min, either with or without PD098059 (2 mM) or geldanamycin(0.5 mM). Explants were pre-treated for 5 h with the di�erent inhibitors
Misexpression of c-Raf kinase in the otic vesicleC Sanz et al
430
epithelium of the otocyst was studied by in situhybridization. Figure 1b shows that c-raf mRNA wasdistributed homogeneously in the otic vesicle at stage18 (E2.5). The lack of variation in Raf levels was inaccordance with the observations that Raf is regulatedat the kinase activity level in most cell contexts(Morrison, 1995). MAP kinase is downstream Rafkinase activation, both kinases function in a cascadethat plays a critical role in the regulation of cell growthand di�erentiation (Marshall, 1994). In this regard, thedegree of phosphorylation of MAP kinase is consid-ered an index of Raf activity. We explored MAPkinase phosphorylation by Western blot analysis(Figure 1c). IGF-I increased MAP kinase phosphoryla-tion threefold over control explants, when explantsfrom stage 18 were incubated with 1 nM IGF-I. Theobservation that MAP kinase was activated did notnecessarily prove the functional role of MAP kinase inthat process. To further con®rm that activation of thec-Raf kinase pathway was involved in the cellularresponse to IGF-I, we used the inhibitors PD098059and geldanamycin. PD098059 blocks the c-Rafactivation of MAP kinase (Coolican et al., 1997), andgeldanamycin selectively promotes c-Raf proteindegradation (Schulte et al., 1996). Both PD098059(2 mM) and geldanamycin (0.5 mM) blocked IGF-Ie�ects on MAP kinase activation (Figure 1c) and cellproliferation (not shown). PD098059 reduced phos-phorylated MAP kinase levels to basal and geldana-mycin had a similar e�ect. The e�ect of geldanamycintreatment and that of IGF-I on c-Raf protein levelswere checked. Geldanamycin treatment decreased c-Raf protein levels whereas IGF-I per se did not induceany change in c-Raf expression (not shown). Theseresults con®rm that the activation of the c-Raf/MAPkinase pathway is a necessary step in the cellularresponse to IGF-I.
Generation and characterization of RCAS raf viruses
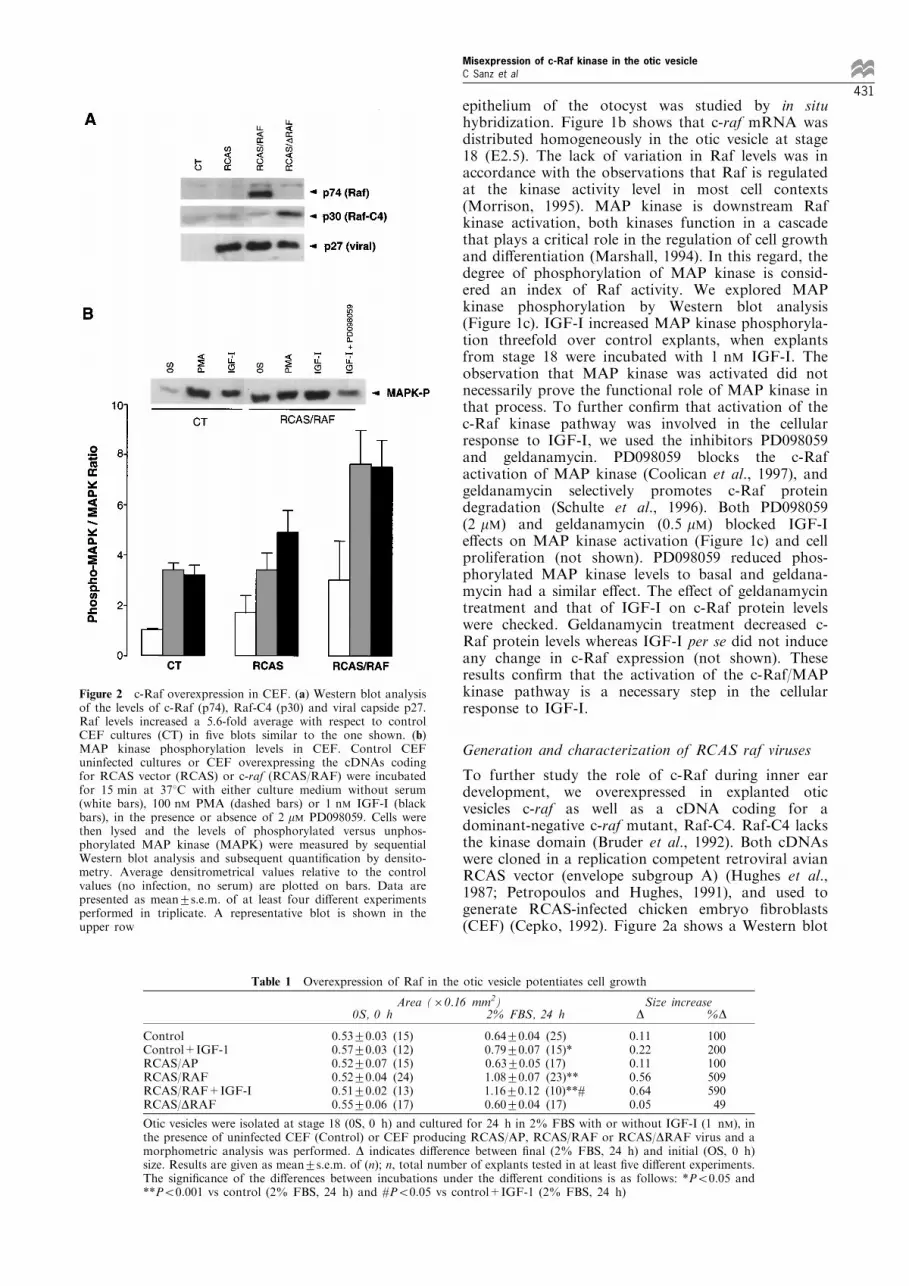
To further study the role of c-Raf during inner eardevelopment, we overexpressed in explanted oticvesicles c-raf as well as a cDNA coding for adominant-negative c-raf mutant, Raf-C4. Raf-C4 lacksthe kinase domain (Bruder et al., 1992). Both cDNAswere cloned in a replication competent retroviral avianRCAS vector (envelope subgroup A) (Hughes et al.,1987; Petropoulos and Hughes, 1991), and used togenerate RCAS-infected chicken embryo ®broblasts(CEF) (Cepko, 1992). Figure 2a shows a Western blot
Figure 2 c-Raf overexpression in CEF. (a) Western blot analysisof the levels of c-Raf (p74), Raf-C4 (p30) and viral capside p27.Raf levels increased a 5.6-fold average with respect to controlCEF cultures (CT) in ®ve blots similar to the one shown. (b)MAP kinase phosphorylation levels in CEF. Control CEFuninfected cultures or CEF overexpressing the cDNAs codingfor RCAS vector (RCAS) or c-raf (RCAS/RAF) were incubatedfor 15 min at 378C with either culture medium without serum(white bars), 100 nM PMA (dashed bars) or 1 nM IGF-I (blackbars), in the presence or absence of 2 mM PD098059. Cells werethen lysed and the levels of phosphorylated versus unphos-phorylated MAP kinase (MAPK) were measured by sequentialWestern blot analysis and subsequent quanti®cation by densito-metry. Average densitrometrical values relative to the controlvalues (no infection, no serum) are plotted on bars. Data arepresented as mean+s.e.m. of at least four di�erent experimentsperformed in triplicate. A representative blot is shown in theupper row
Table 1 Overexpression of Raf in the otic vesicle potentiates cell growth
Area (60.16 mm2) Size increase0S, 0 h 2% FBS, 24 h D %D
ControlControl+IGF-1RCAS/APRCAS/RAFRCAS/RAF+IGF-IRCAS/DRAF
0.53+0.03 (15)0.57+0.03 (12)0.52+0.07 (15)0.52+0.04 (24)0.51+0.02 (13)0.55+0.06 (17)
0.64+0.04 (25)0.79+0.07 (15)*0.63+0.05 (17)1.08+0.07 (23)**1.16+0.12 (10)**#0.60+0.04 (17)
0.110.220.110.560.640.05
10020010050959049
Otic vesicles were isolated at stage 18 (0S, 0 h) and cultured for 24 h in 2% FBS with or without IGF-I (1 nM), inthe presence of uninfected CEF (Control) or CEF producing RCAS/AP, RCAS/RAF or RCAS/DRAF virus and amorphometric analysis was performed. D indicates di�erence between ®nal (2% FBS, 24 h) and initial (OS, 0 h)size. Results are given as mean+s.e.m. of (n); n, total number of explants tested in at least ®ve di�erent experiments.The signi®cance of the di�erences between incubations under the di�erent conditions is as follows: *P50.05 and**P50.001 vs control (2% FBS, 24 h) and #P50.05 vs control+IGF-1 (2% FBS, 24 h)
Misexpression of c-Raf kinase in the otic vesicleC Sanz et al
431

analysis of proteins p74 c-Raf, p30 Raf-C4 and theviral capsid p27 (Stewart et al., 1990). The increaseobserved in c-Raf levels varied from 4 ± 7-fold withrespect to uninfected cells or CEF infected with RCAS(Figure 2b). The next step was to study theconsequences of the overexpression of these genes inCEF. IGF-I and phorbol esters are two well knowne�ectors of c-Raf kinase in a variety of cell systems(Bruder et al., 1992; Troppmair et al., 1994; Chao etal., 1994). Figure 2b shows that CEF cells thatoverexpress c-raf exhibited increased levels of MAPkinase phosphorylation in response to IGF-I orphorbol ester PMA. Incubation of overexpressed c-Raf-CEF with the MAP kinase inhibitor PD098059blocked IGF-I-induced phosphorylation of MAPkinase by 70% (Figure 2b). These results indicatethat CEF cells overexpress a functional c-Raf kinaseand that the c-Raf levels in CEF are limiting for thetransduction of the extracellular stimuli.
Misexpression of Raf in the otic vesicle
To achieve misexpression of c-Raf in explanted oticvesicles we developed the experimental approachillustrated in Figure 3a. CEF were transfected withthe various cDNAs and after the ®rst cycle of infectionhad occurred, the infected cells started to export viralparticles to the cell culture medium. Culture super-natants were collected, concentrated by centrifugation,resuspended in a small volume and used for the nextstep. Otic vesicle explants were then co-cultured withCEF producing viruses seeded in inserts in 2% foetalbovine serum (FBS) containing 10 ml of the concen-trated virus stock. This allowed rapid viral infection of
the explants without direct contact with CEF. After24 h explanted otic vesicles were already infected andthe inserts were removed. The explants were thenincubated with the di�erent factors for the timesindicated. Infection was monitored by Westernblotting (Figure 3b) or immunostaining for theretroviral p27 protein (not shown). Under theseconditions, the average increase of c-Raf levels in theotic vesicles was threefold in 23 explants out of 32, asmeasured by Western blotting (Figure 3c).The biological assays used to investigate c-Raf
misexpression in organotypic cultures of otic vesiclesincluded measurement of cell growth rate by followingexpression of the proliferating cell nuclear antigen(PCNA) and morphometrical analysis as described byLeo n et al., (1998). Otic vesicles were dissected fromstage 18 chicken embryos and cultured under di�erentconditions for 24 h. In the presence of 2% FBS, theotic vesicles which misexpressed c-raf increased in sizewhen compared to control explants kept in the sameculture conditions with uninfected CEF (Figure 4 andTable 1). Similar results were obtained regarding thelevels of PCNA in the explants (Figure 4, lower row).FBS is rich in IGF-I binding proteins that can trapIGF-I and mask the expected e�ects (Spagnoli andRosenfeld, 1997). To avoid this, we used a modi®edfactor, a long-IGF-I that has low a�nity for thebinding proteins. Incubation with 1 nM long-IGF-Iinduced a further increase in the otocyst size (Table 1).The otic vesicles that overexpressed the dominantnegative c-Raf mutant (RCAS/DRAF) did not changein size in response to the stimuli and maintained asimilar aspect to that of control otic vesicles (0S, 0 h)(Figure 4 and Table 1). Neither the co-culture with
Figure 3 c-Raf misexpression in the otic vesicle. (a) A scheme of the experimental approach used to misexpress genes in explantedotic vesicles. (b) Representative Western blotting of the viral protein p27 that indicates the degree of infection of explants co-cultured with uninfected CEF or CEF producing di�erent RCAS viruses. (c) Western blot analysis of the levels of c-Raf (p74) inexplanted otic vesicles co-cultured with CEF in the presence of 2% FBS plus 10 ml of viral stock (left, OV) and in the CEFproducing virus inserts (right, CEF). c-Raf levels increased an average threefold with respect to control explants
Misexpression of c-Raf kinase in the otic vesicleC Sanz et al
432

uninfected CEF, nor infection with RCAS, nor thecontrol with virus bearing the human alkalinephosphatase cDNA (RCAS/AP) had any apparente�ects on cell growth (Figure 4 and Table 1).The increase in size observed in c-Raf-overexpressing
otic vesicles (RCAS/RAF) was, however, not accom-panied by the typical morphological changes observedin explants incubated with either serum or IGF-I (Leo net al., 1995b). We decided to test whether misexpres-sion of c-Raf was a�ecting morphogenesis and celldi�erentiation induced by retinoic acid. After incuba-tion of explanted otic vesicles with retinoic acid, PCNAlevels decreased (Figure 5a). PCNA is a marker for
proliferating cells whose expression is developmentallyregulated in vivo and induced by IGF-I in vitro (Leo net al., 1998; Frago et al., 1998). Retinoic acid promotescell di�erentiation and downregulates Fos levels in theotic vesicle (Represa et al., 1990; Leo n et al., 1995a).Here we show that retinoic acid also a�ects cell cycleprogression by decreasing PCNA levels. Overexpres-sion of c-Raf blocked retinoic acid-induced morpholo-gical changes (not shown) and downregulation ofPCNA expression (Figure 5b, dashed bars). Incuba-tion of explanted otic vesicles with 50 nM retinoic acidin the presence of 2% FBS induced 47% decrease inPCNA levels, whereas in otic vesicles overexpressing c-Raf it was only reduced 22%, with respect to thecorresponding maximum PCNA levels. On the con-trary, overexpression of the dominant negative Rafmutant potentiated the retinoic acid e�ect, thusdecreasing 56% the PCNA levels (Figure 5b, blackbars). These results indicate that c-Raf is involved inthe cellular response to retinoic acid and suggest that c-
Figure 4 c-Raf overexpression in the otic vesicle potentiates cellgrowth. Otic vesicles were isolated at stage 18 (0S) and incubatedfor 24 h in 2% FBS culture medium in the presence of insertsseeded with control CEF cells (CT), CEF producing RCAS/AP,RCAS/RAF or RCAS/DRAF virus, before performing eithermorphometric analysis (see Table 1) or quanti®cation of PCNAlevels. Calibration bar, 130 mm. Photographs are representative ofat least 10 independent experiments performed in triplicate
Figure 5 Retinoic acid e�ects are modulated by c-Raf levels. (a) E�ects of retinoic acid on PCNA levels in explanted otic vesicles.Otic vesicles were explanted from stage 18 chick embryos and co-cultured for 24 h with control CEF cells in 2% FBS. Then, CEFwere removed and the explants incubated for 16 h with 2% FBS in the presence of increasing doses of retinoic acid. Lysates wereanalysed by SDS±PAGE and Western blotting. Speci®c PCNA bands were quanti®ed by densitometry. (b) Otic vesicles wereexplanted and co-cultured with control CEF cells (white bars) or CEF producing either RCAS/RAF (dashed bars) or RCAS/DRAFvirus (black bars) for 24 h. CEF-containing inserts were then removed and the explants cultured for a further period of 16 h in thepresence of the retinoic acid doses indicated. PCNA levels were measured as above. Results in a and b are expressed relative to the2% FBS (100) value and are given as mean+s.e.m. of at least four di�erent experiments performed in triplicate
Table 2 Overexpression of Raf abolishes NGF-induced cell death
± RCAS RCAS/RAF RCAS/DRAF
0SIGF-INGF
10080+3 (13)169+10 (13)
10076+5 (12)142+6 (8)
10052+5 (12)*91+5 (12)**
10076+8 (9)146+8 (6)
Explanted otic vesicles were cultured in 2% FBS for 24 h in theabsence (7) or presence of CEF-producing RCAS, RCAS/RAF orRCAS/DRAF viruses. CEF inserts were then removed and theexplants were incubated with culture medium without serum (0S),IGF-I (1 nM) or NGF (4 nM) for 16 h. Soluble nucleosomes werequanti®ed in single otic vesicle explants. For each experimentalcondition the absorbance obtained with otic vesicles cultured in theabsence of additives was given a value of 100. Results are given asmean+s.e.m. of (n); n, total number of explants tested in at leastthree di�erent experiments. Statistical signi®cance is as follows:*P50.05 and **P50.001 vs RCAS
Misexpression of c-Raf kinase in the otic vesicleC Sanz et al
433

Raf levels may contribute in the otocyst to cellcommitment to enter the cell cycle and proliferate, orelse to exit and di�erentiate.IGF-I is also a cell survival factor for the otic vesicle
and the CVG. On the other hand NGF inducesapoptosis mediated by the p75 neurotrophin receptorin speci®c areas of the otocyst (Frago et al., 1998).After incubation with NGF, only cells expressing p75receptors enter apoptosis. A key biochemical event inapoptosis is nuclear DNA fragmentation, which can bedetected by ELISA immunoassay. Using this approach,we tested the consequences of overexpressing c-Raf andRaf-C4 in the explanted otic vesicles. The results aresummarized in Table 2. Otic vesicles were cultured orco-cultured with RCAS virus producing-CEF. Com-pared with control otic vesicles, co-culture in thepresence of low serum produced a small basal e�ect.In otic vesicles overexpressing c-Raf, IGF-I survivale�ects were higher and NGF-induced cell death wascompletely abolished. Explants expressing the domi-nant negative Raf-C4 exhibited cellular responsessimilar to those seen in control cultures. Hence,competing c-Raf kinase is not su�cient to triggerapoptosis in the population of cells that do not expressp75 low a�nity NGF receptors.
Discussion
There are three main processes which are fundamentalfor organogenesis: cell proliferation, cell di�erentiationand programmed cell death. Cell division andapoptosis are responsible for generating the appro-priate number of cells that form an organ and celldi�erentiation accounts for the specialisation of thesurviving cells. As such, these pathways must be co-ordinately regulated for normal development to occur.Proto-oncogenes that lie upstream the MAP kinasepathway are important regulators of cell growth anddi�erentiation in a variety of cell types. c-Raf kinaseparticipates in this signal transduction network thattransmits proliferative messages from the cell mem-brane to the nucleus (Magnuson et al., 1994; Daum etal., 1994; Marshall, 1994). Increased c-Raf kinaseactivity is associated with an increase in the phosphor-ylation degree of MAP kinase during the cellularresponse to various mitogenic and survival agents,including IGF-I (Alessi et al., 1994; Chao et al., 1994;Marshall, 1994). In the present work, we show thatduring the early organogenesis of the inner ear c-Rafkinase expression is maintained, suggesting that c-Rafis required for the intense mitotic activity reportedduring this period. During this stage and also later indevelopment, IGF-I is a key factor for the regulationof cell proliferation and survival in the otic vesicle(Leo n et al., 1995b, 1998; Oesterle et al., 1997; Zhenget al., 1997; Frago et al., 1998). Furthermore, a partialdeletion in both copies of the Igf-I gene is associatedwith sensorineural deafness in humans (Woods et al.,1996). Our data indicate that the c-Raf/MAP kinasepathway is activated during the proliferative responseto IGF-I, for which response c-Raf is essential.To further analyse the importance of keeping c-Raf
levels tightly regulated during this period, we developedmethods to alter the protein dosage in organotypiccultures of otic vesicles. RCAS vectors have been used
in the past to misexpress genes in the inner ear(Kiernan and Fekete, 1997; Fekete et al., 1997) butthe procedures described did not allow strong infectionor the study of responses to extracellular stimuli suchas IGF-I, NGF or retinoic acid. In contrast, our co-cultures of infected CEF cells with otic vesicles didallow a detailed analysis of events in a short time.Overexpression of c-Raf caused an increased cellularproliferation in response to low serum and IGF-I. Onthe contrary, overexpression of a dominant negative c-Raf mutant protein impaired these e�ects. These datasuggest that c-Raf kinase is a rate limiting step in theMAP kinase cascade following growth factor estimula-tion of the otic vesicle. Typically, modulation of c-Rafkinase is achieved by controlling its degree ofphosphorylation (Chao et al., 1994; Morrison, 1995;Cacace et al., 1996). However, our data indicate that c-Raf levels may be limiting and that precise modulationof c-Raf kinase levels contribute to control cellularresponses. The abundance of c-Raf protein is alsotightly regulated during early Xenopus development(MacNicol et al., 1995). Taken together, these resultssuggest that this might be a general mechanismoperating during development to control the extent ofcellular proliferation. Moreover, treatment of oticvesicles with retinoic acid induces di�erentiation ofthe otic epithelia, concomitantly with a reduction ingrowth rate (Leo n et al., 1995a). c-Raf overexpressionwas accompanied by a remarkable decrease in theresponse to retinoic acid which was evaluated as theincrease in PCNA levels in parallel with themorphological analysis. By competing c-Raf kinaseactivity with a dominant negative c-Raf mutant, wehave observed that c-Raf potentiates retinoic acidaction by shifting the pro®le of the dose responsecurve. The otic ephitelium is morphologically homo-geneous but has regional and cell fate speci®ty which isdetermined by the asymmetric expression of severalgenes (reviewed by Fekete, 1996; Torres and Giraldez,1998). Our data indicate that the dosage of a givengene may introduce another level of regulation.A potential role for the c-Raf/MAP kinase pathway
in the regulation of apoptosis has been proposed onthe basis of data obtained in a number of cell types aswell mice lacking c-raf or B-raf genes (reviewed byPritchard and McMahon, 1997). Programmed celldeath at early developmental stages in the nervoussystem is essential for tissue morphogenesis andneurogenesis (Ra� et al., 1993; Weil et al., 1997).NGF can have both pro- and anti-apoptotic actions,depending on the set of receptors expressed in thetarget cell (reviewed in Bergeron and Yuan, 1998). Inthe inner ear, NGF-induced apoptosis has beenproposed to play a role in maintaining the number ofneuronal precursors and in the formation of theendolymphatic duct (Frago et al., 1998). Overexpres-sion of c-Raf causes a profound alteration in theresponse to NGF. An increase in c-Raf levelssuppresses NGF induction of apoptosis and potenti-ates IGF-I actions as a survival factor. These resultscon®rm the importance of maintaining c-Raf levels inthe otic vesicle during E2-3 since their unbalance wouldalter early morphogenesis by increasing the total cellnumber. Furthermore, the data presented here con®rmsa role for c-Raf in the suppression of apoptosis andwould suggest that modulation of c-Raf levels
Misexpression of c-Raf kinase in the otic vesicleC Sanz et al
434

contributes to gain speci®city in the response to a givengrowth factor or to the local combination ofextracellular stimuli that encounter a developing cell.In conclusion, our results indicate that avian
retroviruses are a useful tool to dissect the role ofkey signalling proteins in the response to extracellularfactors during embryonic development, and that c-Rafkinase has a critical role in the integration ofmechanisms that regulate cellular proliferation, differ-entiation and apoptosis during the organogenesis of theinner ear.
Materials and methods
Construction of viral vectors and production of viral stocks
RCAS is a replication-competent retroviral vector derivedof Rous sarcoma virus (Hughes et al., 1987). It contains aClaI site for cloning of exogenous genes. RCAS envelopesubgroup A vector and RCAS containing the humanphosphatase alkaline cDNA (RCAS/AP) were the gener-ous gift of Dr S Hughes (National Cancer Institute-Frederick, MD, USA). In order to clone c-raf and Raf-C4 cDNAs into RCAS, c-raf and Raf-C4 were subclonedinto the PAJ10 adapter plasmid to obtain ClaI ends. PAJ10is a Bluescript plasmid containing a multicloning adapter¯anked by ClaI sites that was a kind gift of Dr D MartinZanca (CSIC, Salamanca, Spain), c-raf was cleaved fromElneo c-raf (Heidecker et al., 1990) by EcoRI-ApaIdigestion and inserted into PAJ10 (PAJ10/RAF). Then c-raf was removed from PAJ10 by digestion with ClaI andinserted into RCAS at the ClaI site (RCAS/RAF). RCASclones containing the raf inserts in the sense orientationwere identi®ed and isolated. RSV-C4 encodes a dominantnegative Raf protein (Raf-C4) (Bruder et al., 1992). TheXhoI ±XbaI fragment was isolated from this vector,subcloned into PAJ10 (PAJ10/DRAF) and cloned intoRCAS as described above (RCAS/DRAF).
CEF were transfected with 10 mg of each plasmidcontaining the proviruses by calcium phosphate co-precipita-tion (Petropoulus et al., 1989). Transfected cells were thenpassaged and expanded for 10 days in CEF culture medium;at the end of this period all cells were infected, as estimatedby immunohistochemistry with anti-p27 antibody. Viralsupernatants were collected and concentrated 100-fold bycentrifugation (3 h 28 000 r.p.m.). The titer of the viral stockwas determined as described by (Fekete and Cepko, 1993).Titers were typically in the range 2 ± 36108 cfu/ml. Five viralpreparations were used for these studies with similar results.
Otic vesicle isolation and cell culture
Chicken embryos were obtained from fertilized eggs(Granja RodrõÂ guez Serrano, Salamanca. Spain) that wereincubated at 388C in humid atmosphere. The embryos werestaged according to Hamburger and Hamilton criteria,(1951). Embryos were removed from eggs and placed in 35-mm Petri dishes (NUNC, Roskilde, Denmark) containingphosphate bu�ered saline. Otic vesicles were dissected fromembryos corresponding to stage 18 using a dissectionstereomicroscope and transferred into four-well cultureplates (NUNC). The standard culture medium consisted ofserum-free M199 medium with Earle's salts (Biowhitaker,Walkersville, MD, USA) supplemented with 2 mM gluta-mine (Biowhitaker), and antibiotics (penicillin, 50 IU/mland streptomycin, 50 mg/ml) (Biochrom, Berlin, Germany).FBS was purchased from Biowhitaker. Incubations werecarried out at 378C in a water-saturated atmospherecontaining 5% CO2. Estimation of otocyst size was carriedout by measuring ratios of surface area as described (Miner
et al., 1988; Leo n et al., 1995a, 1998). Computer-generatedphotomicrographs were produced by scanning the obtainedphotos with a Studio ScanII (Agfa) using the programsAdobe Photoshop 2.5 and MacDraw Pro on Macintoshcomputers.
Line 0 CEF were prepared from 10 day chicken embryos(kind gift from Dr J Martin, IIB-CSIC, Madrid) and culturedas described (Hunter, 1979). After infection, CEF werecultured with various mitogens as described in the text orused for the production of virus. When indicated, otic vesicleswere dissected from chick embryos corresponding to stage 18and cultured as described above. Explants and CEF cellsproducing virus were co-cultured at 378C and 5% CO2 forthe times indicated in the presence of 2% FBS plus 10 ml ofconcentrated viral stock. The degree of infection wasestimated by Western blotting or immunocytochemistry witha polyclonal anti-p27 antibody (generous gift from Dr VolkerM Vogt, Cornell University, NY, USA). The followingcontrols were included for each block of experiments: co-culture with uninfected CEF or with CEF producing RCASor RCAS/AP virus. No clear e�ects were observed in any ofthe cases and therefore the control conditions are not shownin every experiment.
In some experiments NIH3T3 ®broblasts, either wild typeor overexpressing c-raf, were used to prepare control proteinextracts. NIH3T3 were subcultured as described (Heideckeret al., 1990).
In situ hybridization
In situ hybridization was performed as described (Leo n etal., 1998), using a digoxigenin-labelled RNA probe for c-raf. A single-stranded c-raf RNA probe was prepared bythe transcription with T7 RNA polymerase of thelinearized plasmid PAJ10/RAF with SalI (Promega,Southampton, UK). Control sense probes were preparedby linearising the plasmid with HindIII and transcribingwith T3 RNA polymerase (Promega). For histologicalexamination the embryos were hybridized and post-®xed in4% paraformaldehyde, embedded in gelatine-albumin, andsectioned (60 mm) with a vibratome (Leica VT 1000 M,Heidelberg, Germany).
Immunoblotting and MAP kinase assay
For Western blotting, otic vesicle explants were homo-genized in SDS ± PAGE sample bu�er with 1 mM
phenylmethylsulfonyl-¯uoride and then frozen immedi-ately. Gels were loaded with equivalent amounts ofproteins. Otic vesicle proteins were resolved using 10%SDS ± PAGE before being electrophoretically transferredonto PVDF membranes (Dupont-NEN, Boston, MA,USA). Filters were blocked with Tris-bu�ered salinecontaining 5% (wt/vol) non-fat dried milk and incubatedwith the primary speci®c antibody. Filters were subse-quently washed and incubated with the correspondingsecondary antibody conjugated with peroxidase. Boundperoxidase activity was visualized by chemiluminiscence(Dupont-NEN) and quanti®ed by analysing scannedimages of the Western blots using the program NIHImage 1.59 on Macintosh computers. The relativeintensities of the corresponding bands were then com-pared. When indicated, cellular extracts of CEF wereprepared using RIPA bu�er (10 mM Tris-HCl pH 8.0,150 mM NaCl, 1% Triton X-100, 1% aprotinin, 250 mMphenylmethylsulfonyl ¯uoride, 1 mM sodium ¯uoride and100 mM sodium orthovanadate) and analysed as above.Protein concentration was estimated by using the BCAprotein assay (Pierce).
Rabbit polyclonal antibody to human c-Raf was fromUpstate Biotechnology Incorporated (UBI, New York,USA). This antibody recognizes the highly conserved
Misexpression of c-Raf kinase in the otic vesicleC Sanz et al
435

sequence corresponding to residues 380 ± 396 of the sub-domain III of the 74 kDa human c-Raf kinase (Pelech et al.,1990). Rabbit polyclonal antibody to Raf-C4 (anti-BRaf#301) recognizes 17 residues in the carboxyl region ofB-Raf (Bruder et al., 1992). Mouse monoclonal to MAPkinase and rabbit polyclonal anti phosphospeci®c MAPkinase antibodies were from Zymed (San Francisco, CA,USA) and New England Biolabs (Beverly, MA, USA),respectively. These antibodies recognize the carboxylterminus of MAP kinase and the tyrosine phosphorylatedresidues 196 ± 209 of human p44 MAP kinase, respectively.The proliferative cell nuclear antigen (PCNA) is a nuclearprotein of 36 kDa associated with the cell cycle (Bravo andMacDonald-Bravo, 1987). Levels of PCNA were taken as anindex of cell proliferation. Mouse monoclonal anti-PCNAwas from ATOM (ATOM, Barcelona, Spain). Secondaryantibodies conjugated with peroxidase were purchased fromBio-Rad (Richmond, CA, USA).
For the MAP kinase assay, otic vesicles were stimulatedfor 15 min by addition of 1 nM IGF-I to the culture medium.Explants were immediately lysed in RIPA bu�er for Westernblot processing. Levels of phosphorylated MAP kinase weremeasured and normalized with respect to total MAP kinaselevels under each condition. PD098059 (2'-amino-3'-methoxy-¯avone), a selective inhibitor of MAP kinase kinase(Coolican et al., 1997), was purchased from Calbiochem(San Diego, CA, USA). Geldanamycin treatment selectivelydepletes c-Raf cellular contents, disrupting the c-Raf/MAPkinase signalling pathway (Schulte et al., 1996). Geldanamy-cin (kind gift of Dr J Renart, IIB-CSIC, Madrid) was fromthe Developmental Therapeutics Program, National CancerInstitute, Bethesda, MD, USA. Retinoic acid was from Sigma(St. Louis, MS, USA), 2.5S mouse NGF was from UBI,recombinant human IGF-I from Boehringer Mannheim(Mannheim, Germany), and long-IGF-I from GroPep(Adelaide, Australia).
Cell death determination by immunoassay
Quanti®cation of cell death was performed by using theCell Death Detection ELISA from Boehringer Mannheimbased on the detection of histone-associated DNAfragments (mono- and oligo-nucleosomes) in the cellscytoplasm. Explanted otic vesicles were co-cultured withCEF infected with the di�erent RCAS vectors for 24 h.The inserts were removed and the explants cultured for afurther period of 16 h in the presence or absence of 1 nMIGF-I or 4 nM (100 ng/ml) NGF. Otic vesicles werehomogenized and the cell extracts subjected to ELISAdetermination as indicated by the manufacturer anddescribed (Frago et al., 1998).
AbbreviationsChicken embrionary ®broblasts (CEF); cochleovestibularganglion (CVG); embryonic day (E); insulin-like growthfactor-I (IGF-I); mitogen activated protein kinase (MAPkinase); nerve growth factor (NGF).
AcknowledgementsYL held a CSIC research contract and CS was supportedby Ministerio de Education y Cultura. We wish to thankDrs Stephen Hughes, Volker M Vogt and Luis Alvarez fortheir generous help and the continuos technical adviceprovided along this work, and to Drs Luis Alvarez,Fernando Giraldez, Flora de Pablo, Jaime Renart, DavidJones and Carmen G Vallejo, for critical reading of themanuscript. We also thank Antonio Ferna ndez and JavierPe rez for photographic work and illustrations respectively.The research was supported by grants from Direccio nGeneral Investigacio n Ciencia y Tecnologõ a (PM96-0075)and Europharma (Boehringer Ingelheim Inc.) to I V-N andDeutsche Forschungsgemeinschaft (DFG) Sonder-forschungsbereiche (SFB) 465 and 172 to URR.
References
Alessi D, Saito Y, Campbell D, Cohen P, Sithanandam G,Rapp U, Ashworth A, Marshall C and Cowley S. (1994).EMBO J., 13, 1610 ± 1619.
Bergeron L and Yuan J. (1998). Currr. Opin. Neurol., 8, 55 ±63.
Bissonnette JP and Fekete DM. (1996). J. Comp. Neurol,174, 1 ± 11.
Bravo R and MacDonald-Bravo H. (1987). J. Cell. Biol.,105, 1549 ± 1554.
Bruder JT, Heidecker G and Rapp UR. (1992). GenesDevelop., 6, 545 ± 556.
Brunner D, Oellers N, Szabad J, Biggs WH, Zipursky SL andHafen E. (1994). Cell, 76, 875 ± 888.
Cacace A, Ue�ng M, Philipp A, Han EK, Kolch W andWeistein IB. (1996). Oncogene, 13, 2517 ± 2526.
Cepko CL. (1992). Current Protocols in Molecular Biology.Ausubel FM, Brent R, Kingston RE, Moore DD, SeidmanJG, Smith JA and Struhl K (eds). Greene and Wiley-Interscience: New York, pp. 9.10.1 ± 9.13.5.
Chao T-SO, Foster DA, Rapp UR and Rosner MR. (1994).J. Biol. Chem., 269, 7337 ± 7341.
Coolican S, Samuel D, Ewton D, McWade F and Florini J.(1997). J. Biol. Chem., 272, 6653 ± 6662.
Daum G, Eisenmann-Tappe I, Fries H-W, Troppmair J andRapp UR. (1994). Trends Biochem. Sci., 19, 474 ± 479.
Federspiel M and Hughes S. (1997). Methods Cell Biol., 52,179 ± 214.
Fekete DM. (1996). Curr. Opin. Neurol., 6, 533 ± 541.Fekete DM and Cepko C. (1993).Mol. Cell. Biol., 13, 2604 ±2613.
Fekete D, Homburger S, Waring M, Riedl A and Garcia L.(1997). Development, 124, 2451 ± 2461.
Frago L, Leo n Y, De la Rosa E, Go mez-MunÄ oz A andVarela-Nieto I. (1998). J. Cell Science, 111, 549 ± 556.
Georgi L, Albert P and Riddle D. (1990). Cell, 61, 635 ± 645.Hamburger V and Hamilton HL. (1951). J. Morphol., 88,49 ± 92.
HanM, Golden A, Han Y and Sternberg PW. (1993).Nature,363, 133 ± 140.
Heidecker G, Huleihel M, Cleveland JL, Kolch W, Beck TW,Lloyd P, Pawson T and Rapp UR. (1990).Mol. Cell. Biol.,10, 2503 ± 2512.
Hemond SG and Morest DK. (1991). Anat. Embryol., 184,1 ± 13.
Hughes SH, Greenhouse JJ, Petropoulos CJ and Sutrave P.(1987). J. Virol., 61, 3004 ± 3012.
Hunter E. (1979). Methods in Enzymology. Jakoby WB andPastan IH (eds). Academic Press, Inc: San Diego, pp.379 ± 393.
Jansen HW, Lurz R and Bister K. (1984). Nature, 307, 281 ±284.
Kiernan A and Fekete D. (1997). Audiol. Neurootol., 2, 12 ±24.
Koenen M, Sippel AE, Trachmann C and Bister K. (1988).Oncogene, 2, 179 ± 185.
Kyriakis JM, App H, Zhang X, Banerjee P, Brautigan DL,Rapp UR and Avruch J. (1992). Nature, 358, 417 ± 421.
Lee KH and Cotanche DA. (1996). Hear. Res., 94, 1 ± 13.Leo n Y, Sa nchez J, Miner C, Ariza-McNaughton L, RepresaJ and Giraldez F. (1995a). Dev. Biol., 167, 75 ± 86.
Misexpression of c-Raf kinase in the otic vesicleC Sanz et al
436

Leo n Y, Sanz C, Giraldez F and Varela-Nieto I. (1998). J.Comp. Neurol., 398, 323 ± 332.
Leo n Y, Va zquez E, Sanz C, Vega J, Mato JM, Giraldez F,Represa J and Varela-Nieto I. (1995b). Endocrinology,136, 3494 ± 3503.
MacNicol A, Muslin A, Howard E, Kikuchi A, MacNicol Mand Williams L. (1995). Mol. Cell. Biol., 15, 6686 ± 6693.
Magnuson N, Beck T, Vahidi H, Hahn H, Smola U andRapp UR. (1994). Semin. Cancer Biol., 5, 247 ± 253.
Mark G and Rapp U. (1984). Science, 224, 285 ± 289.Marshall CJ. (1994). Curr. Opin. Genet. Dev., 4, 82 ± 89.Miner C, Represa JJ, Barbosa E and Giraldez F. (1988).
Roux's Arch. Dev. Biol., 197, 294 ± 297.Morrison DK. (1995). Mol. Reprod. Dev., 42, 507 ± 514.Naumann U, Eisenmann-Tape I and Rapp UR. (1997).
Recent Results Cancer Res., 143, 237 ± 244.Oesterle EC, Tsue T and Rubel EW. (1997). J. Comp.
Neurol., 380, 262 ± 274.Pelech SL, Sanghera JS and Daya-Makin M. (1990).
Biochem. Cell Biol., 68, 1297 ± 1330.Perrimon N and Desplan C. (1994). Trends Biochem. Sci., 19,509 ± 513.
Petropoulus CJ, Rosenberg MP, Jenkins NA, Copeland NGand Hughes SH. (1989). Mol. Cell. Biol., 9, 3785 ± 3792.
Petropoulus CJ and Hughes SH. (1991). J. Virol., 65, 3728 ±3737.
Pritchard C and McMahon M. (1997). Nat. Genet., 16, 214 ±215.
Ra� MC, Barres B, Burne JF, Coles HS, Ishizaki Y andJacobson MD. (1993). Science, 262, 695 ± 700.
Represa J, Sa nchez A, Miner C, Lewis J and Giraldez F.(1990). Development, 110, 1081 ± 1090.
Schulte TW, Blagosklonny M, Romanova L, Mushinski JF,Monia BP, Johnston JF, Nguyen P, Trepel J and NeckersLM. (1996). Mol. Cell. Biol., 16, 5839 ± 5845.
Spagnoli A and Rosenfeld RG. (1997). Curr. Opin.Endocrinol. Diabetes, 4, 1 ± 9.
Stewart L, Schatz G and Vogt VM. (1990). J. Virol., 64,5076 ± 5092.
Storm SM, Cleveland JL and Rapp UR. (1990). Oncogene, 5,345 ± 351.
Torres M and Giraldez F. (1998). Mech. Dev., 71, 5 ± 21.Troppmair J, Bruder JT, Munoz H, Lloyd PA, Kyriakis J,Banerjee P, Avruch J and Rapp UR. (1994). J. Biol.Chem., 269, 7030 ± 7035.
Von Bartheld CS, Patterson SL, Heuer JG, Wheeler EF,Bothwell M and Rubel EW. (1991). Development, 113,455 ± 470.
Wassarman DA, Solomon NM, Chang HC, Karim FD,Therrien M and Rubin GM. (1996). Genes Dev., 10, 272 ±278.
Weil M, Jacobson MD and Ra� MC. (1997). Curr. Biol., 7,281 ± 284.
Woods KA, Camacho-Hubner C, Savage MO and Clark AJ.(1996). N. Engl. J. Med., 335, 1363 ± 1367.
Wojnowski L, Zimmer AM, Beck TW, Hahn H, Bernal R,Rapp UR and Zimmer A. (1997). Nat. Genet., 16, 293 ±297.
Xu RH, Dong Z, Maeno M, Kim J, Suzuki A, Ueno N,Sredni D, Colburn NH and Kung HF. (1996). Proc. Natl.Acad. Sci. USA, 93, 834 ± 838.
Zheng JL, Helbig C and Gao WQ. (1997). J. Neurosci., 17,216 ± 226.
Misexpression of c-Raf kinase in the otic vesicleC Sanz et al
437




















