
Staining method for determination of the penetration of immobilized enzyme into a porous support
11
Staining Method for Determination of the Penetration of Immobilized Enzyme into a Porous Support S. W. CARLEYSMITH,* M. B. L. EAMES,t and M. D. LILLY, Department of Chemical and Biochemical Engineering, University College London, Torrington Place, London, England Summary The penetration of bovine serum albumin and penicillin acylase into Amberlite XAD7 beads was determined by staining split beads. The rates of penetration were measured and correlated with a theoretically derived equation. INTRODUCTION Enzymes have been immobilized onto a wide range of porous supports, either particulate or membranous. Mathematical models used to predict the behavior of these immobilized enzymes normally assume that the enzyme is evenly distributed throughout the sup- port. This may not be the reality, although Lasch et a1.l showed the homogeneous distribution of immobilized protein in Sepharose 6B beads by using fluorescein-tagged protein and observing sections of beads under ultraviolet light. We have been investigating the im- mobilization of penicillin acylase onto porous spherical particles of the resin, Amberlite XAD-7. In the absence of added buffer, the hydrolysis of benzylpenicillin by penicillin acylase produces phen- ylacetic acid, which is dissociated at the pH values that are normally used for the reaction. Also, both reaction products cause inhibition of the reaction, so that the concentrations of hydrogen ions, reac- tant, and products within the porous beads have complex profiles. Because of acid production and inhibition by products, the enzyme in the center of the beads is unlikely to contribute significantly to the hydrolytic reaction. Thus for optimum catalyst design it is im- portant to determine the location of the enzyme within the bead, * Present address: Beecharn Pharmaceuticals (UK Division) Worthing, England. t Present address: BP Research, Sunbury-on-Thames, England. Biotechnology and Bioengineering, Vol. XXII, Pp. 957-967 (1980) @ 1980 John Wiley & Sons, Inc. OOO6-3592/80/0022-0957$0 1 .OO
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Staining method for determination of the penetration of immobilized enzyme into a porous support

Staining Method for Determination of the Penetration of Immobilized Enzyme into a
Porous Support
S. W. CARLEYSMITH,* M. B. L. EAMES,t and M. D. LILLY, Department of Chemical and Biochemical Engineering, University
College London, Torrington Place, London, England
Summary
The penetration of bovine serum albumin and penicillin acylase into Amberlite XAD7 beads was determined by staining split beads. The rates of penetration were measured and correlated with a theoretically derived equation.
INTRODUCTION
Enzymes have been immobilized onto a wide range of porous supports, either particulate or membranous. Mathematical models used to predict the behavior of these immobilized enzymes normally assume that the enzyme is evenly distributed throughout the sup- port. This may not be the reality, although Lasch et a1.l showed the homogeneous distribution of immobilized protein in Sepharose 6B beads by using fluorescein-tagged protein and observing sections of beads under ultraviolet light. We have been investigating the im- mobilization of penicillin acylase onto porous spherical particles of the resin, Amberlite XAD-7. In the absence of added buffer, the hydrolysis of benzylpenicillin by penicillin acylase produces phen- ylacetic acid, which is dissociated at the pH values that are normally used for the reaction. Also, both reaction products cause inhibition of the reaction, so that the concentrations of hydrogen ions, reac- tant, and products within the porous beads have complex profiles. Because of acid production and inhibition by products, the enzyme in the center of the beads is unlikely to contribute significantly to the hydrolytic reaction. Thus for optimum catalyst design it is im- portant to determine the location of the enzyme within the bead,
* Present address: Beecharn Pharmaceuticals (UK Division) Worthing, England. t Present address: BP Research, Sunbury-on-Thames, England.
Biotechnology and Bioengineering, Vol. XXII, Pp. 957-967 (1980) @ 1980 John Wiley & Sons, Inc. OOO6-3592/80/0022-0957$0 1 .OO

958 CARLEYSMITH, EAMES, AND LILLY
especially as it is possible to immobilize the enzyme under condi- tions that result in the protein being fixed only at or near the bead surface.
Direct observation of the protein distribution was found to be possible by using a dye that bound irreversibly to the protein. Protein staining (widely used for detecting protein in analytical systems involving gel diffusion) has also been used to detect protein immobilized in layers in gel membranes.2
THEORY
Penetration of Protein into a Porous Activated Support
Assume a porous spherical particulate support (Fig. 1) and: 1) The rate of protein adsorption is much higher than the rate of
diffusion and therefore adsorption is at equilibrium and the system is diffusion controlled.
2) Once adsorbed to the activated support, protein is irreversibly bound.
3) The diffusivity of the soluble protein in the saturated zone is constant.
4) The protein adsorption capacity of the support is much greater than the amount of protein dissolved in the void volume of the support, i.e., p %- 4,.
5 ) The equilibrium concentration ( p ) of adsorbed protein can be regarded as constant and independent of the corresponding protein concentration ( C , ) in free solution over the range of protein solu- tions examined, i.e., the specific volumetric capacity of the support for protein is constant.
It follows from these assumptions that all protein entering the particle is adsorbed at the boundary between the protein saturated and unsaturated zones, and that the concentration profile of protein
Un7tuuted
\ \ - - Protein
boundary
Particle surface
Fig. 1. Diagram of partially saturated bead particle.

PENETRATION OF IMMOBILIZED ENZYMES 959
through the saturated zone is rapidly established compared to the rate of movement of the boundary.
The rate of adsorption of protein (a) is given by
dr b a = 4.nrb2p - dt
kglsec
The protein flux (J,) at any radius r is given by Fick's Law
dc eff dr
J , = - D - kg/m2.sec
where Deff is the effective diffusivity of the soluble protein in the saturated zone. The negative sign indicates that material enters the particle. Thus the total transport of protein into the particlehec (a) is given by
dc a = -4.nr2Deff-
dr kglsec
Integrating across the saturated zone,
a = -4 7rDeff Co[rbR/ (R - r b ) ] kg/sec (3)
(4)
t c = PR2/6DeffCo (54
p = rb/R (5b)
7 = t / t c (5c)
Combining eqs. (1) and (3):
-D, ,Codt /p = ( R - rb) rbdrb/R
To put this into a dimensionless form, define
So eq. (4) becomes
-dT = 6(1 - p ) p dp
Integrating with respect to time,
7 = I - 3p2 + 2 p 3
1 - p is plotted against 7 in Figure 2.
then p = 0, so from eq. (6), T = 1 . For the special case of penetration to the center of the particle,
so t / t c = T = 1
t = pR2/6DeffCo = 1 ,

%O CARLEYSMITH, EAMES, AND LILLY
0 1 2 7-
Fig. 2. Theoretical curves for the immobilization of protein to porous activated spherical particles, where 7 is the dimensionless reaction time, p is the proportion of the particle that would be saturated with protein, and ( 1 - p ) is the fractional radial penetration of protein into the particle. (-) Case where the external protein concentration remains constant and (- - -) case where the initial amount of protein available equals the maximum amount that can be immobilized to the particle.
Thus t , is the time taken for the protein to reach the center of the particle when the external coupling protein concentration remains unchanged at C, . It can be compared with
t = p Ax2/2DeffC0
derived by Goldman et a1.2 for the penetration of protein to a depth Ax into a membrane.
Proportion of Support Saturated with Protein
Of more practical significance than the movement of the boundary is the amount of protein immobilized, expressed as a fraction of total saturation of the support. At time t , r = i b and the total protein adsorbed (P,) is given by
4 3 (7) P , = - T ( R 3 - rb3)p kg
This is made dimensionless by defining
p = P t ( 3 / 4 r R 3 p )
p is the proportion of support saturated with protein. Combining eqs. (7), (S), and (5b):
p = (1 -p )1’3 (9)

PENETRATION OF IMMOBILIZED ENZYMES %I
So from eqs. (6) and (9):
7 = 3 - 3 ( 1 - - 2 p
p is plotted as a function of7 in Figure 2.
Cuse of Changing External Protein Concentration
The foregoing treatment was where there was a constant C, , i.e., a sufficiently large excess of coupling protein so that the con- centration does not change significantly during the immobilization period. A more useful result is obtained for the case when the amount of coupling protein is limited. As protein is adsorbed, C o falls to C , at time t , so that
c, = c, - P , / V
c, = c, - (4T p / 3 V ) ( R 3 - rb3)
Substituting eq. (7),
( 1 1 )
The adsorption of proteidsec (a ) is now given by
a = -4.rrD,&,[rbR/(R - r b ) ] (12)
[cf. eq. (3)]. Substituting eq. ( 1 1 ) into eq. (12) and combining eq. ( 1 ) :
This may be made dimensionless by defining
+ = 3 V c o / 4 ~ R 3 p
$ is the ratio of total challenge protein at the start ( t = 0) to the total capacity of the support.
Substituting for dimensionless groups in eq. (13):
When + = 1, the challenge protein at the start equals the capacity of the support. For this special case.

962 CARLEYSMITH, EAMES, AND LILLY
Upon integration,
7 = 6(l /p + In p - 1) (17)
Of more interest is the fractional saturation (p) of the support.
(18)
1 - p is plotted against 7 in Figure 2 .
From eqs. (9) and (17):
7 = 6 (1 - @ ) - * I 3 + 21n (1 - @) - 6
p is plotted against T in Figure 2.
Immobilization of Enzyme beneath a Layer of Inactive Protein in a Spherical Porous Particle
Equation (7) gives the amount of enzyme protein immobilized in a layer from the particle surface inwards and may be written as follows:
4 p = - 3 .rrpR3 (1 - p3) (19)
It is also possible to immobilize an enzyme as a layer beneath a layer of inactive protein by exposing the particle first to a solution of the inactive protein, washing, and then exposing the particle to the enzyme solution. Assuming that p is the same for both the active and inert proteins, the amount of enzyme coupled will be given by,
(20) P = - 7rpR3 ( p I 3 - pZ3)
where p1 and pz are the respective values for the inner boundaries of the inert protein and enzyme layers. Assuming as before that p is constant with particle depth, the depth of the enzyme inner layer required to give an activity equivalent (in the absence of diffusional limitation of the reaction rate) to a layer adjacent to the surface may be found by combining eqs. ( 19) and (20) to give
I - p3 = p: - pZ3
4 3
or rearranging
pz” = p3 + p: - 1 (21)
Having selected a value for p l , p2 can be calculated, and hence from eq. (6) the time T~ to produce the inert layer. Similarly, the total time required to achieve a penetration pz, T ~ , can be calculated.

PENETRATION OF IMMOBILIZED ENZYMES 963
The difference, T~ - T ~ , is the time required for adsorption of the second enzyme layer.
MATERIALS AND METHODS
Amberlite XAD-7 beads, 20-50 mesh [Rohm and Haas (UK) Ltd., Croydon, England] were sieved to give a size fraction of 420-850 pm. The beads were activated as described e l ~ e w h e r e , ~ and then suspended in a solution of penicillin acylase or bovine serum al- bumin (BSA) at 20°C for various times. Unless stated otherwise, l g damp activated beads were presented with I g protein. The beads were then separated by filtration, washed six times with 50 ml lots of deionized water, and then stored at 0°C in deionized water until required.
Beads were split using a scalpel blade and stained for 30 min with a 1.5 mgfliter solution of Ponceau S CI27195 (molecular weight of 761) in 3% (wiv) aqueous trichloroacetic acid. A sample of whole beads, stained and then split, gave the same results. Removal of the unbound dye was accomplished by repeated rinsing of the split beads with 2% ( v h ) aqueous solution of glacial acetic acid for 30 min. After this treatment only a barely detectable color remained in the beads that had been activated but to which protein had not been bound. The split beads were dried in an oven at 45°C for 10 min and then mounted for photography on a thin film of Araldite Rapid Adhesive (Ciba-Geigy Co.) on a microscope slide. The beads were photographed under a tungsten-halogen light. As the magni- fications of the microscope and the enlarger head used to print the photographs were known, the diameters of the beads and the depths of penetration of protein could be obtained from measurements made on the photographic prints.
Immobilized penicillin acylase was assayed with a pH titrator as described p r e v i ~ u s l y . ~ Batch reactions were done in a 50 ml reactor containing 40 ml 0.2M benzylpenicillin at 37°C maintained at pH 8 by addition of 5N NaOH.
RESULTS AND DISCUSSION
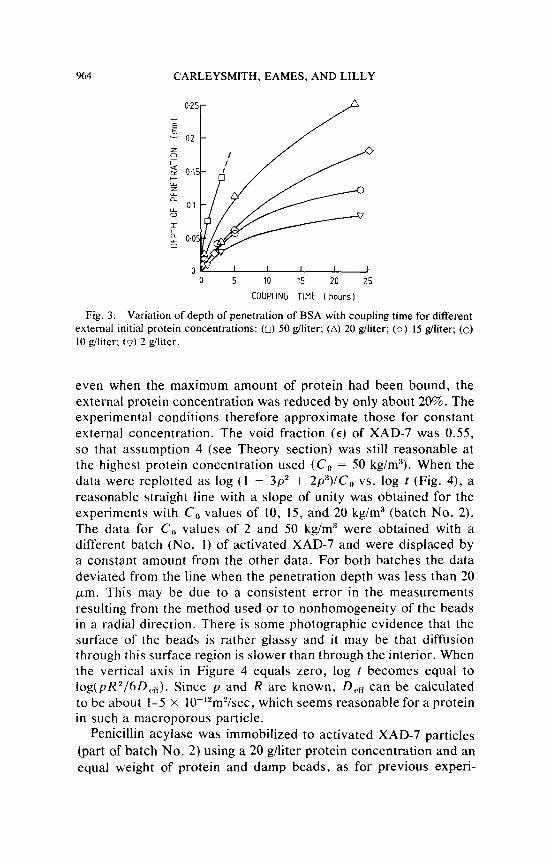
The depths of penetration of BSA into the 600-700 p m diam beads with coupling time for different initial external protein con- centrations are shown in Figure 3. The total binding capacity ( p ) for BSA or penicillin acylase of the activated XAD-7 was about 190 kg/m3, assuming a density for the damp beads of 11 10 kg/m3. Thus
%4 CARLEYSMITH, EAMES, AND LILLY
0 5 10 15 20 25
COUPLING TIME I hours)
Variation of depth of penetration of BSA with coupling time for different external initial protein concentrations: (0) 50 giliter; (A) 20 giliter; (0) IS giliter; (0) 10 giliter; (v) 2 giliter.
Fig. 3.
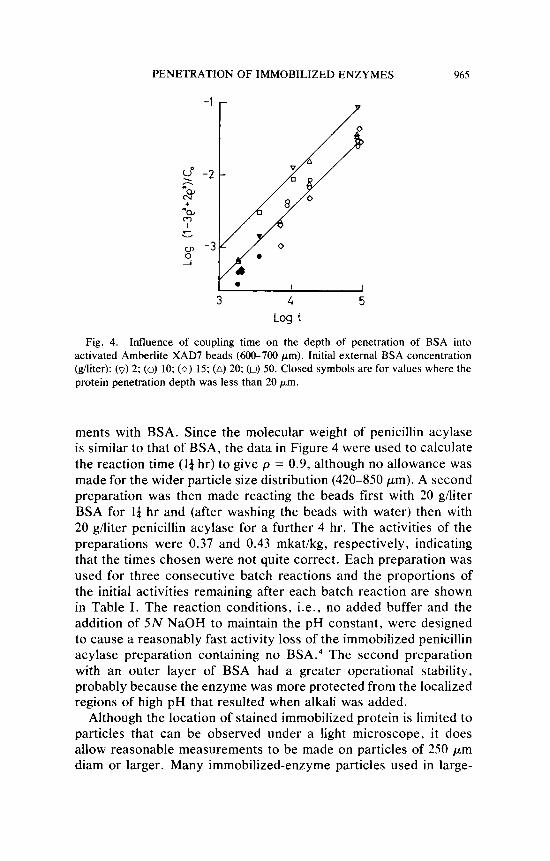
even when the maximum amount of protein had been bound, the external protein concentration was reduced by only about 20%. The experimental conditions therefore approximate those for constant external concentration. The void fraction ( E ) of XAD-7 was 0.55, so that assumption 4 (see Theory section) was still reasonable at the highest protein concentration used ( C , = 50 kg/m3). When the data were replotted as log (1 - 3p2 + 2p3) /C, vs. log r (Fig. 4), a reasonable straight line with a slope of unity was obtained for the experiments with C , values of 10, 15, and 20 kg/m3 (batch No. 2). The data for C , values of 2 and 50 kg/m3 were obtained with a different batch (No. I ) of activated XAD-7 and were displaced by a constant amount from the other data. For both batches the data deviated from the line when the penetration depth was less than 20 pm. This may be due to a consistent error in the measurements resulting from the method used or to nonhomogeneity of the beads in a radial direction. There is some photographic evidence that the surface of the beads is rather glassy and i t may be that diffusion through this surface region is slower than through the interior. When the vertical axis in Figure 4 equals zero, log r becomes equal to log(pR2/6D,,,,). Since p and R are known, D,, can be calculated to be about 1-5 x 10-'*m2/sec, which seems reasonable for a protein in such a macroporous particle.
Penicillin acylase was immobilized to activated XAD-7 particles (part of batch No. 2) using a 20 glliter protein concentration and an equal weight of protein and damp beads, as for previous experi-
PENETRATION OF IMMOBILIZED ENZYMES 965
I 1
3 4 5 Log t
Fig. 4. Influence of coupling time on the depth of penetration of BSA into activated Amberlite XAD7 beads (600-700 pm). Initial external BSA concentration (giliter): (0) 2; (0) 10; ( 0 ) 15; (A) 20; (0) 50. Closed symbols are for values where the protein penetration depth was less than 20 pm.
ments with BSA. Since the molecular weight of penicillin acylase is similar to that of BSA, the data in Figure 4 were used to calculate the reaction time ( l a hr) to give p = 0.9, although no allowance was made for the wider particle size distribution (420-850 pm). A second preparation was then made reacting the beads first with 20 g/liter BSA for Id hr and (after washing the beads with water) then with 20 g/liter penicillin acylase for a further 4 hr. The activities of the preparations were 0.37 and 0.43 mkat/kg, respectively, indicating that the times chosen were not quite correct. Each preparation was used for three consecutive batch reactions and the proportions of the initial activities remaining after each batch reaction are shown in Table I. The reaction conditions, i.e., no added buffer and the addition of 5N NaOH to maintain the pH constant, were designed to cause a reasonably fast activity loss of the immobilized penicillin acylase preparation containing no BSA.4 The second preparation with an outer layer of BSA had a greater operational stability, probably because the enzyme was more protected from the localized regions of high pH that resulted when alkali was added.
Although the location of stained immobilized protein is limited to particles that can be observed under a light microscope, it does allow reasonable measurements to be made on particles of 250 pm diam or larger. Many immobilized-enzyme particles used in large-

966 CARLEYSMITH, EAMES, AND LILLY
TABLE I Stability of Immobilized Penicillin Acylase
after Reuse in a Batch Reactor
Activity remaining (%) after batch No.
Immobilized protein 1 2 3
Penicillin acylase 85 75 68 BSA followed by
penicillin acylase 97 95 93
scale reactors are of this size.5 Thus this technique may be useful for the optimum design of immobilized-enzyme particles in reactions other than the deacylation of benzylpenicillin.
Nomenclature rate of adsorption of protein per particle (kg/sec) initial protein concentration in challenge solution (kg/m3) protein concentration in challenge solution at time t (kg/m3) diffusivity of protein in saturated zone of particle (m*/sec) flux of protein into particle at radius r (kg/m* sec) specific protein capacity of support (kg/m3) protein adsorbed per particle at time t (kg) radial position in particle (m) radial position of boundary of the protein saturated zone (m) radius of spherical particle (m) time (sec) time for protein to reach center (i.e., fully saturated) of the particle at constant challenge protein concentration C o (sec) volume of challenge solution (m3) depth of protein penetration into membrane (m) proportion of support saturated with protein (= 3 P1/47-rR3p) support void fraction (m3/m3) dimensionless radial position of boundary of protein saturated zone (= rb lR) dimensionless time of coupling reaction (= t/t,)
ratio of total challenge protein at start to capacity of the support (= 3 V C o / 4 n R 3 p )
References 1. J . Lasch, M. Iwig, and H. Hanson, Eur. J. Biochern., 27, 431 (1972). 2. R. Goldman, 0. Kedem, I. H. Silman, S. R. Caplan, and E. Katachalski,
3 . S. W. Carleysmith, P. Dunnill, and M. D. Lilly, Biotechnol. Bioeng., in press. Biochemistry. 7 , 486 (1968).

PENETRATION OF IMMOBILIZED ENZYMES 967
4. S . W. Carleysrnith and M. D. Lilly, Biotechnol. Bioeng., 21, 1057 (1979). 5. M. D. Lilly, Proceedings of First European Congress of Biotechnology, Inter-
laken, September 1978 (Dechema Monographs, Berlin, 1978), Vol. 82, pp. 165-180.
Accepted for Publication August 20, 1979




















