Confronting Drought in Africa's Drylands - Open Knowledge ...
Upload
khangminh22Category
view
0download
0
University of Texas at El PasoDigitalCommons@UTEP
Open Access Theses & Dissertations
2016-01-01
Spatiotemporal Variability Of Plant Phenology InDrylands: A Case Study From The NorthernChihuahuan DesertNaomi Robin LunaUniversity of Texas at El Paso, [email protected]
Follow this and additional works at: https://digitalcommons.utep.edu/open_etdPart of the Ecology and Evolutionary Biology Commons, and the Environmental Sciences
Commons
This is brought to you for free and open access by DigitalCommons@UTEP. It has been accepted for inclusion in Open Access Theses & Dissertationsby an authorized administrator of DigitalCommons@UTEP. For more information, please contact [email protected].
Recommended CitationLuna, Naomi Robin, "Spatiotemporal Variability Of Plant Phenology In Drylands: A Case Study From The Northern ChihuahuanDesert" (2016). Open Access Theses & Dissertations. 684.https://digitalcommons.utep.edu/open_etd/684

SPATIOTEMPORAL VARIABILITY OF PLANT PHENOLOGY IN
DRYLANDS: A CASE STUDY FROM THE NORTHERN CHIHUAHUAN
DESERT
Naomi Robin Luna, B.Sc.
Master’s Program in Environmental Science
APPROVED:
__________________________________________
Craig E. Tweedie, Ph.D.
__________________________________________
Dawn Browning, Ph.D.
__________________________________________
Jennie McLaren, Ph.D.
_______________________________________
Charles Ambler, Ph.D.
Dean of the Graduate School

Copyright ©
by
Naomi Robin Luna
2016

SPATIOTEMPORAL VARIABILITY OF PLANT PHENOLOGY IN DRYLANDS: A CASE STUDY
FROM THE NORTHERN CHIHUAHUAN DESERT
By
NAOMI ROBIN LUNA, B.Sc.
THESIS
Presented to the Faculty of the Graduate School of
The University of Texas at El Paso
in Partial Fulfillment
of the Requirements
for the Degree of
MASTER OF SCIENCE
Environmental Science Program
THE UNIVERSITY OF TEXAS AT EL PASO
December 2016

iv
Acknowledgments
I would like to thank my advisors Dr. Craig E. Tweedie, Dr. Dawn Browning, and Dr. Jennie
McLaren for their encouragement and help in developing and understanding my project. There
are too many others that I would like to thank who helped me so much throughout my stint as a
graduate student, but I would like to thank all of the students and staff (Dr. Geovany Ramirez,
Gesuri Ramirez, Dr. Christine Laney, Dr. Sandra Villarreal, and Dr. Xia Song) from the Systems
Ecology Laboratory at UTEP for their encouragement and guidance throughout my project. I am
especially thankful for Dr. Geovany Ramirez and his development of the Phenology Analyzer
Software, which played a critical role in the processing of data for this project which made a
world of a difference from alternate software currently available. I am also very grateful for
having had the chance to collaborate with Dr. Dawn Browning, your guidance and enthusiasm
throughout this project was much appreciated. I would also like to thank the Jornada
Experimental Range LTER program which provided the perfect platform needed to complete my
project. Last, but certainly not least, I would like to thank my family and friends for their
support, guidance, and patience throughout my challenges. More importantly I am thankful for
my son, Dominic, whom I dedicate this thesis to. I hope you realize one day that all the hard
work, dedication, and sacrifices I make in life are for you, I love you beyond words.
This material is supported by the National Science Foundation (NSF) - CREST - 1242122. All
the ideas, outcomes, and conclusions given in this work are those of the author(s) and do not
necessarily reflect the views of NSF.
NOTE: This thesis was submitted to my Supervising Committee on December 13, 2016.

v
Abstract
With global change, which includes climate change, there is a sense of urgency to understand
how shifts in climate will affect ecosystems. Although several studies have improved
understanding of how and why some ecosystems respond, most studies have not explored
simultaneous responses of different land cover types throughout a given region. Dryland
ecosystems, such as the Chihuahuan Desert, appear to respond to climate variability and
currently make up about 40% of global land surface area. It is expected that drylands will expand
to cover 60% of land surface area on earth by mid-century making this ecosystem more critical
to global land-atmosphere interactions than previously thought.
The goal of the proposed study is to determine how plant phenology in multiple desert land cover
types responds to seasonal and inter annual climate variability over five years. Phenology is the
timing of major growth stages in plants and animals which has been shown to provide important
insights into the environmental state heavily influenced by climate change. Dryland plant
phenology is relatively understudied.
Time series imagery acquired by static digital cameras in five land cover types on the USDA
Jornada Experimental Range in Southern New Mexico between 2010 and 2015 were analyzed
with alternative remote sensing techniques at the landscape level phenology. At each site, both
the phenology of the landscape and replicates of key species were analyzed using custom
phenology analysis software developed within the Systems Ecology Lab at the University of
Texas at El Paso. This study is expected to expand the current knowledge of the effects of
climate variability and change in dryland ecosystems by understanding which land cover types

vi
and species are more/less sensitive to change. The study is also novel in that it will explore
image processing methods that have yet to be fully explored by ecosystem scientists.
Grasses displayed greater seasonal fluctuation in greening thought to be tied closely to rainfall
events, where shrubs displayed a more consistent inter-annual growth pattern. This is
hypothesized to be attributed to accessibility to deeper water storage attainable by the more
extensive root systems commonly found in shrubs. Exploration into the use of alternate spectral
signatures from images to capture timing of key growth stages proved to be useful in the patchy
land cover. More extensive research needs to be done, but this study has hinted to advantages for
using alternate color models for processing images within these extreme and complex
ecosystems. These results may provide strong implications to predicting future ecosystem states
of the northern Chihuahuan Desert region including ecosystem properties and processes such as
biodiversity and land-atmosphere carbon fluxes.

vii
Table of Contents
Acknowledgments ......................................................................................................................... iiv
Abstract.. ..........................................................................................................................................v
Table of Contents ........................................................................................................................... vii
List of Tables ................................................................................................................................ viii
List of Figures ................................................................................................................................ iix
1. Introduction ...................................................................................................................... 1
1.1. Background and Rationale ............................................................................................... 1
1.2. Goals and objectives ...................................................................................................... 16
1.3. Study Area ..................................................................................................................... 17
2. Inter-comparison of plant and landscape phenology in different land cover types on
the Jornada Experimental Range ................................................................................... 33
2.1 Introduction .................................................................................................................... 33
2.2 Methods.......................................................................................................................... 33
2.3 Results ............................................................................................................................ 40
2.4 Discussion ...................................................................................................................... 48
3. Use of alternate color space to derive phenological trends in a Chihuahuan Desert
shrubland ........................................................................................................................ 50
3.1 Introduction .................................................................................................................... 50
3.2 Methods.......................................................................................................................... 51
3.3 Results ............................................................................................................................ 61
3.1 Discussion ...................................................................................................................... 71
4. General Conclusions ...................................................................................................... 73
5. Suggestions for future work ........................................................................................... 74
6. References ...................................................................................................................... 76
7. Curriculum Vitae ........................................................................................................... 90

viii
List of Tables
Table 1.1: Focal plant species monitored in this study.. ........................................................................... 22
Table 2.1: Cameras used to address objectives one and two. .................................................................. 36
Table 3.1: Number of individuals for each key plant species observed at site and transect. .................... 57
Table 3.2: Difference of means of proportion of plants for creosote bush (3 clusters) captured
by landscape spectral indices................................................................................................... 64
Table 3.3: Difference of means of proportion of plants for creosote bush (4 clusters) captured
by species spectral indices. ...................................................................................................... 65
Table 3.4: Difference of means of proportion of plants for creosote bush (4 clusters) captured
by landscape spectral indices................................................................................................... 65
Table 3.5: Difference of means of proportion of plants for honey mesquite (3 clusters) captured
by species spectral indices ....................................................................................................... 67
Table 3.6: Difference of means of proportion of plants for honey mesquite (3 clusters) captured
by landscape spectral indices................................................................................................... 68
Table 3.7: Difference of means of proportion of plants for honey mesquite (4 clusters) captured
by species spectral indices ....................................................................................................... 69
Table 3.8: Difference of means of proportion of plants for honey mesquite (4 clusters) captured
by landscape spectral indices................................................................................................... 70

ix
List of Figures
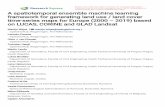
Figure 1.1: Tripod with a fixed camera mounted on top to capture images of the landscape .................. 8
Figure 1.2: RGB color model ................................................................................................................... 9
Figure 1.3: HSV color model ................................................................................................................. 12
Figure 1.4: L*a*b* color model ............................................................................................................. 13
Figure 1.5: Map of the USDA-ARS JER near Las Cruces, New Mexico. ............................................. 18
Figure 1.6: Phenocam locations within the JER. .................................................................................... 19
Figure 1.7: U.S. distribution map of the shrub honey mesquite ............................................................. 29
Figure 1.8: U.S. distribution map of the evergreen creosote bush. ......................................................... 29
Figure 1.9: U.S. distribution map of the grass bush muhly.. .................................................................. 30
Figure 1.10: U.S. distribution map of the deciduous tarbush ................................................................... 30
Figure 1.11: U.S. distribution map of the grass tobosa grass ................................................................... 31
Figure 1.12: U.S. distribution map of the grass mesa dropseed. .............................................................. 31
Figure 1.13: U.S. distribution map of the grass black grama. .................................................................. 32
Figure 1.14: U.S. distribution map of the grass fluff grass ....................................................................... 32
Figure 2.1: Gradient Compensation Module. ......................................................................................... 37
Figure 2.2: Color gradient displayed in image ....................................................................................... 37
Figure 2.3: Gradient Compensation Module – image correction sample. .............................................. 38
Figure 2.4: Phenology Analyzer Software (PAS) interface. ................................................................... 38
Figure 2.5: Regions of interest (ROI) for landscape and species specific phenology across
all land cover types. ............................................................................................................. 39
Figure 2.6: Daily and cumulative rainfall (mm) for 2012-2015.. ........................................................... 42
Figure 2.7: Landscape phenology trends across all land cover types from 2012-2015.. ........................ 42
Figure 2.8: Phenology trends (GCC) for honey mesquite and graminoids observed in all LCTs .......... 44
Figure 2.9: IBPE1 landscape and species 28-day GCC trend lines ........................................................ 45

x
Figure 2.10: PAS9 landscape and species 28-day GCC trend lines ......................................................... 46
Figure 2.11: SCAN1 landscape and species 28-day GCC trend lines ...................................................... 46
Figure 2.12: SCAN2 landscape and species 28-day GCC trend lines ...................................................... 46
Figure 2.13: TROM landscape and species 28-day GCC trend lines ....................................................... 47
Figure 2.14: TWEE landscape and species 28-day GCC trend lines ........................................................ 47
Figure 3.1: UTEP-Systems Ecology Lab (SEL). Phenology transects ................................................... 54
Figure 3.2: UTEP-SEL’s site with the relative field of view for cam2 .................................................. 54
Figure 3.3: Flow chart of methods for Ch. 3 .......................................................................................... 55
Figure 3.4: Field of View for cam2 within PAS ..................................................................................... 56
Figure 3.5: Key phenophases for Larrea tridentata (creosote bush) ....................................................... 58
Figure 3.6: Key phenophases for Prosopis glandulosa (honey mesquite) .............................................. 59
Figure 3.7: Key phenophases for Prosopis glandulosa (honey mesquite) .............................................. 60
Figure 3.8: Proportion of plants for creosote bush with 3 cluster groups. .............................................. 62
Figure 3.9: Proportion of plants for creosote bush with 4 cluster groups. .............................................. 63
Figure 3.10: Proportion of plants for honey mesquite with 3 cluster groups ........................................... 63
Figure 3.11: Proportion of plants for honey mesquite with 4 cluster groups... ........................................ 63

1
1. Introduction
1.1. Background and Rationale
1.1.1. Definition and importance of plant phenology
Phenology is the study of the timing of growth and reproductive patterns for a particular
organism. For plants, this includes events such as the timing, rate of development, and duration
of initial leaf growth, production of flower buds, opening of flowers, release of seeds, and onset
of senescence. Plant phenology is sensitive to climate, and the alteration of a range of ecosystem
properties and processes (Cook et al., 2012; Crimmins et al., 2010; Menzel et al., 2006; Zhang &
Friedl, 2004). The study of plant phenology and phenological change has been undertaken for
centuries, but the fervor for phenological studies has increased dramatically over the past few
decades due in part to the urgency for understanding how climate change will impact plant
phenology both within and between ecosystems (Ibáñez et al., 2010; Menzel et al., 2006;
Richardson et al., 2009). As early as the eighteenth century, plant phenology had already been
linked to climatic conditions. Such evidence suggests that climate change will likely alter spatial
and temporal phenological patterns causing changes in ecosystem properties and processes
(Bowers & Dimmitt, 1994; Huenneke et al., 2002; Joiner et al., 2014; Munson et al., 2013).
Research has linked shifts in spatial and temporal ecosystem properties and processes to changes
in plant phenology (Cook et al., 2012; Joiner et al., 2014). Plant-animal interactions can be
particularly sensitive to the timing and duration of specific plant growth stages (Bascompte et al.,
2016; Elzinga et al., 2007; Hegland et al., 2009). In a study that examined plant-animal
relationships for selected small rodent species in the Chihuahuan Desert, for example, a strong
relationship between small mammal population size and the timing of plant growth development

2
was noted (Ernest et al., 2000). Similarly, the timing of plant production relies heavily on the
timing and duration of climatic conditions, such as, precipitation and temperature which can shift
due to climate change (Ernest et al., 2000).
There are various motivations and benefits for monitoring plant phenology both in managed (e.g.
agriculture, grazing) and unmanaged settings (e.g. conservation, invasive species).
Advancement of plant phenology research has been especially important in agriculture because
research findings have helped to improve yield, the efficiency of agricultural production, and
economic gain (Bowers & Dimmitt, 1994; Loomis & Connor, 1992). In most modern
agricultural operations, the timing of phenophases such as flowering and fruiting provides insight
to the likely timing and success of proceeding phenophases such as seed set, which can be used
to predict the timing of harvests and the logistics of transportation of products to market
(Ahrends et al., 2008; Campillo et al., 2008; Morisette et al., 2009).
Although agriculture might be a more obvious and relatable sector that relies on plant phenology
studies, there are other sectors that rely on these studies/observations also. Japanese festivities,
for example, celebrate the production of blossoms from the Japanese cherry trees which is of
critical economic and social importance (Allen et al., 2014). It was determined that with
increasing temperatures flowering of Japanese cherry trees might occur one month earlier by
mid-century (Allen et al., 2014), which could mean having to adjust the timing of festivities to
ensure festivities coincide with peak flowering. Furthermore, the economic profitability of honey
production and the associated links to pollination success can be strongly dependent on the
interaction, or lack of interaction between honeybees and flowering phenology (Bagella et al.,
2013). More commonly, phenophases are used as a metric of ecosystem productivity where

3
shifts or differences in climate and other ecosystem properties and processes are recognized
constraints and controls (Kramer et al., 2000).
Monitoring plant phenology has been used to show the impacts of climate change on plant
species and ecosystem dynamics (Arora & Boer, 2005; Cleland et al., 2006; Richardson et al.,
2007). Research in colder ecosystems provide evidence that decreasing snow depth and earlier
snowmelt as a result of climate change has already had and will likely continue to play a
significant role in plant responses (Cooper et al., 2011; Van Wijk et al., 2003; Wipf et al., 2009).
The growing season for most areas in the Arctic and subarctic is limited to the summer months
(May-August/September) (Euskirchen et al., 2006; Tucker et al., 2001). This short growing
season is only exacerbated when the timing of snowmelt occurs later each successive year
coupled with decreasing snow depth which makes it more difficult for plants to establish strong
roots (Cooper et al., 2011; Van Wijk et al., 2003). “Green-up” of plants within a given
ecosystem marks the beginning of a new growing season and ultimately controls other
phenological stages (e.g. flowering, fruiting, seed set) both for individual plants and whole
ecosystems (Cong et al., 2013; Pettorelli et al., 2005; Schwartz & Karl, 1990). Global studies
have noted a variable but generally earlier onset of green-up across biomes, further providing
evidence that climate change is impacting ecosystems (Badeck et al., 2004; Cook et al., 2012;
Keenan et al., 2014; Peñuelas et al., 2009). Furthermore, certain plant species may succumb to
critical thresholds that can threaten plant diversity and distribution. For some species, such
thresholds could determine their listing on endangered species lists in the future (Kelly &
Goulden, 2008; Thomas et al., 2004; Thuiller et al., 2005).

4
Capturing spatial and temporal canopy state patterns can give insight to energy, water, and land-
atmosphere exchange of carbon within the ecosystem (Cleland et al., 2007; Geesing et al., 2000;
Joiner et al., 2014; Kurc & Benton, 2010; Richardson et al., 2013). Carbon-cycling is closely
linked to the timing of greening in most ecosystems(Joiner et al., 2014; Mizunuma et al., 2013).
In drylands, phenology and C-cycling are strongly linked (Knapp et al., 2008; Kurc & Benton,
2010). Most modern studies reporting on such activities, have utilized the analysis of time-lapse
photography from stationary digital cameras using digital image analysis techniques (Brown et
al., 2016). As mentioned above, agricultural practices rely heavily on phenological observations
and the analysis of digital imagery from stationary cameras has been shown to provide unbiased
and non-subjective observations for determining more accurate harvest times (Ahrends et al.,
2008; Campillo et al., 2008; Crimmins & Crimmins, 2008; Kurc & Benton, 2010; Przeszlowska
et al., 2006; Richardson et al., 2007; Vanamburg et al., 2006). Other disciplines that have
increasingly used static digital camera stations include, but are not limited to, wildlife/habitat
monitoring (Bater et al., 2011; Brawata et al., 2013; Deacy et al., 2016), coastal erosion
monitoring (Jones et al., 2009; Walker, 2014; Whitehead et al., 2010), and dust monitoring
(Lorenz, 2009; Skiles et al., 2015). Several of these studies have demonstrated direct links to
improved habitat management. Many different areas of study utilize time lapse images for data
collection due to the ease by which high temporal resolution observations can be made,
decreased field time, improved degree of quantifiable results, and generally higher quality data;
all of which have stimulated the digital image revolution in the environmental sciences (Julitta et
al., 2014; Keenan et al., 2014; Melaas et al., 2016; Nagai et al., 2014; Toomey et al., 2015;
Young et al., 2015).

5
1.1.2. Biophysical controls and relationships of plant phenology
Abiotic factors beyond climate change can influence the phenological patterns of plants.
Biophysical controls such as spatial distribution, light availability and strength, and soil moisture
conditions and geographic variability can alter plant phenological patterns (Forrest et al., 2010;
Parmesan & Yohe, 2003; Root et al., 2003; Rosenzweig et al., 2008). The magnitude of impact
from any given controlling factor can depend on the stresses from multiple co-occurring
biophysical factors including competition among other plant species for space, light, and water
(Harris, 1977; Myneni & Williams, 1994; White et al., 1997). Light availability and light
intensity vary with canopy structure (Cheng et al., 2006), which can also be influenced by
atmospheric conditions (e.g. clouds, dust) (Kurc & Benton, 2010; Loomis & Connor, 1992;
Rathcke & Lacey, 1985). Some plant species, including several perennial grasses in dryland
ecosystems, grow under the canopy of shrubs, which can limit light availability for these grass
species (Archibald & Scholes, 2007; Tucker et al., 2001; Van Auken, 2009) and lead to delayed
growth (Archibald & Scholes, 2007; Thomson & Siddique, 1997). Soil moisture can also
influence plant phenology by either accelerating or delaying plant growth when optimal
conditions do not apply (Kidron & Gutschick, 2013; Kurc & Benton, 2010).
Plant phenology also influences the biological structure and function of ecosystems in multiple
ways. These include but are not limited to: the timing of flower emergence that may directly
affect key pollinator’s cycle (Chew & Whitford, 1992; Forrest et al., 2010; Lightfoot et al.,
1989) grain production which is an important energy source for some small mammals (Brown,
et al., 1979); roosting environments for various winged animals (Matuzak & Brightsmith, 2007;
Ober et al., 2005; Scott, 2004); and nesting sites for various species that rely on specific plant

6
growth stages for rearing offspring (Hingrat et al., 2007; Sedinger & Raveling, 1986; Wagner,
1997). Litter fall is an important phenological stage that may control nutrient cycling (Barlow et
al., 2007; Campanella & Bertiller, 2008; Facelli & Pickett, 1991). As the importance of plant
phenology to organism and ecosystem structure and function has been increasingly recognized, a
plethora of methodologies have developed to heighten the intricacies of phenological research
and that has permitted scaling of studies across broader scales of space and time (Joiner et al.,
2014; Moore et al., 2016; Nelson & Papuga, 2009; Piao et al., 2007).
1.1.3. Approaches to measuring plant phenology
Various methods exist to measure plant phenology, but some are more efficient and less
cumbersome than others. Some of the more commonly practiced methods include (1) human
observations, which require a researcher to physically go out to the field, monitor, and record
plant phenological stages (phenophases). Although in-situ observations are still commonly
practiced, (2) remote sensing techniques are not subjective, are less expensive, can generally
cover a much larger area, and can record at higher frequency making this a highly preferred
method in most modern phenological and multi-scale phenological studies. The analysis of
satellite imagery has be used to monitor green-up and senescence at the landscape or regional
scale for several decades (Archibald & Scholes, 2007; Cong et al., 2013; White & Nemani,
2006) and allows researchers to more easily interpret phenological change at regional or global
scales (Allen et al., 2014).
Satellite imagery can be robust and expensive if high quality data is needed, but alternate remote
sensing techniques exist that capture imagery over smaller areas at higher resolution (e.g.
ground-level images), yet still permit insightful ecosystem studies (Browning et al., 2015; Ma et

7
al., 2013; Pettorelli et al., 2007; Tucker et al., 2001). While satellite imagery captures regional
scale ecosystem productivity there are smaller scale techniques that are optimal for capturing
landscape and plant level phenological trends. Hard-mounted digital cameras that capture repeat
imagery of a fixed field of view (FOV) within RGB (red, green, and blue) color space
(phenocams, Figure 1.1) have become popular and have arguably transformed phenological
research over the past decade.
Phenocam images are made up of pixels that each contains a ratio of red, green, and blue
channels(Barsky, n.d.; Connolly & Fliess, 1997; Saitoh et al., 2012; Sonnentag et al., 2012). The
quality of such images depends on the capacity of the camera itself. The more pixels the camera
can capture for each images, the more detail will be represented. The quality needed in a camera
depends on the goals of the project. The specifications of the cameras used for this project will
be addressed below. Digital time lapse photography and associated image analysis is now a
popular data collection method that can be used to cheaply and consistently record plant
phenological trends in most ecosystems (Ansley et al., 2001; Richardson et al., 2007). For plant
phenological observations, a greenness index (derived from the RGB color model) within a given
region of interest (ROI) in the image is typically what most researchers focus on (Benton et al.,
2008; Hufkens et al., 2004; Toomey et al., 2015). This approach is generally sufficient to capture
plant to landscape phenological change (Bater et al., 2011; Mizunuma et al., 2013; Toomey et
al., 2015). There are, however, alternate color models that remain poorly explored in the
ecosystem sciences that may be equally useful adept as the aforementioned RGB color model. In
order to fully understand how each color model is represented and applied to image analysis
techniques, the following section will expand on these concepts and their applications to
phenological studies.

8
Figure 1.1: Tripod with a hard-mounted digital camera programmed to acquire repeat images of a fixed
field of view (Image: Robin Luna).

9
RGB color model:
RGB (red, green, blue) is an additive color model, which means the color perceived in an image
depends on how many red, green, and blue rays are added to each pixel and collectively will
represent the image being captured (Figure 1.2) (Connolly & Fliess, 1997). RGB closely matches
the human perceptions of color (Connolly & Fliess, 1997; Ford & Roberts, 1998). RGB is the
standard color model used for plant phenological analysis because of its adequacy in
representing greenness within a given ROI (Bater et al., 2011; Proulx & Parrott, 2008) and is
what affordable off-the-shelf cameras offer. An equation (Equation 1) is readily used that
calculates this greenness within the image, which is critical for calculating the amount of green
captured within a given ROI (Hufkens et al., 2004; Richardson et al., 2007; Vezhnevets, n.d.).
This equation, coined the green excess index (GEI), calculates greenness in vegetation based on
the absolute channel brightness (DN) for green with respect to red and blue channels for a given
ROI (Bater et al., 2011; Graham et al., 2009; Mizunuma et al., 2013; Seager et al., 2007). Other
commonly used equations derived from the RGB color model include the normalized difference
Figure 1.2: RGB color model. R=red channel, G=green channel, and B=blue channel (Barsky, n.d.).

10
vegetation index (NDVI; Equation 2) (Gamon et al., 1995; Gamon et al., 2013; Richardson et
al., 2007), and more recently, the green chromatic coordinate (GCC; Equation 3) (Ahrends et al.,
2008; Klosterman et al., 2014; Sonnentag et al., 2012; Toomey et al., 2015). These color models
have all been tested rigorously in different studies for overall efficacy and thus, have been
adopted as a few of the more reliable indices for phenocam-based based plant phenological
studies (Benton, 2009; Richardson et al., 2007; Toomey et al., 2015).
Richardson’s (etal.) 2007 paper provided the building blocks for future plant phenological
studies, which included the use of the green excess index (GEI) as the basic equation rooted from
the RGB color model that allows researchers to quantify the temporal “greenness” of a landscape
and growth trends that can be linked to climate variability and change (Migliavacca et al., 2011;
Mizunuma et al., 2013; Richardson et al., 2007, 2009). Along with GEI, the indices NDVI and
GCC have proven to be both reliable and informative for documenting changes in green plant
productivity in a given ecosystem (Gamon et al., 2013; Huemmrich et al., 2010; Migliavacca et
al., 2011). Although RGB color space is regularly used in ecological research there are several
important caveats; it is device dependent, there is a high correlation between color channels, and
there is mixing of chrominance and luminance data, which can limit the use of this color model
for some analyses (Vezhnevets, n.d.).
(1)
where DN represents the digital number (channel brightness) for red, green, and blue channels
(Richardson et al., 2007) .

11
(2)
where NIR is the reflectance in near infrared wavelength and RED is the reflectance in red
wavelength (Gamon et al., 2006).
(3)
where DN represents the digital numbers for red (R), green (G), and blue (B) channels (Toomey
et al., 2015).

12
HSV color model:
Although the HSV color model has been rarely explored in phenological studies, there are
several studies that demonstrate potential utility beyond the capacity of the RGB color space.
HSV (hue, saturation, and value – a transformation of the RGB color model) will be used in this
study to determine its capacity for capturing specific plant and landscape phenological trends
(see Chapter 3). HSV is based on a different concept to the RGB color model and represents
colors in their purest form that can be most effectively visualized as an inverse cone (Figure 1.3)
(Proulx & Parrott, 2008). Hue (H) is the specific color that is being displayed that can be seen
along the perimeter (circumference) of the cone in (Figure 1.3) (Crimmins & Crimmins, 2008;
Smith, 1978). Once the specific color is identified, saturation (S) is calculated and represents
how rich the color is. In other words, the higher the saturation value the more washed out the
color will appear. High S values can be seen closest to the central axis of the cone visualizing
HSV color space in Figure 1.3 and low S values can be seen closest to the perimeter of the cone
Figure 1.3: HSV color model, H=hue, S=saturation, and V=value (Barsky, n.d.).

13
where colors are the most ‘pure’ (Figure 1.3) (Crimmins & Crimmins, 2008; Smith, 1978). Value
(V) represents the richness of the color and can range from pure black (the peak of the cone ~
low richness) to high richness at the plane of the cone where color is most ‘pure’ (Crimmins &
Crimmins, 2008; Smith, 1978). Although the previously mentioned RGB indices (GEI, NDVI,
and GCC) are frequently used for phenological studies, HSV has also been explored and proven
in some cases to be more reliable than RGB color space under certain circumstances (Benton,
2009; Crimmins & Crimmins, 2008; Pekel et al., 2014). Benton (2009) found that hue was
optimal for the detection of creosote bush flowering. Hue sometimes overestimated the number
of flowers present, but was more accurate than RGB color space options investigated (Benton
2009).
L*a*b* color model:
Figure 1.4: L*a*b* color model. L* corresponds to the lightness or darkness in the image. Channels a*
and b* represent the opponent color theory for red and green & blue and yellow, respectively (image:
http://dba.med.sc.edu/price/irf/Adobe_tg/models/cielab.html).

14
L*a*b* (luminance, channel a, and channel b, respectively; Figure 1.4) will be used to determine
the efficiency in capturing specific growth patterns in plant phenology (see Chapter 3 below).
This color model is based on opponent-color theory which reflects more closely the human
perception of colors. L* measures the reflectance properties of a pixel - how light or dark the
image is; channel a* represents the color gained from the red and green opponent colors, and
channel b* represents the resultant color from the opponent colors yellow and blue (Connolly &
Fliess, 1997; Hill et al., 1997). There appears to be no published research that has explored the
utility of L*a*b* the color model in phenological studies, suggesting that the study completed in
Chapter 3 below may be pioneering this research.
1.1.4. Review and challenges of phenological studies in drylands
One of the historical and still current data collection methods include human observations, which
requires individuals to observe and record growth stages of given plant species with datasheets
developed by entities such as the National Phenology Network (NPN), the National Ecological
Observatory Network (NEON) and ProjectBudBurst. The terminologies and definitions
developed by the NPN has created a level of consistency evident across data collectors and has
helped make data collection and dispersion possible to a wide range of interest groups (Denny,
2012; Richardson et al., 2007). Since accuracy and consistency are critical to recording plant
phenophases, data collection methods expanded to remotely sensed methods, such as time lapse
images acquired on phenocams are helping to capture phenological variance in a range of
ecosystems (Benton, 2009; Richardson et al., 2007; Toomey et al., 2015). Various remote
sensing techniques (e.g. NDVI from satellite imagery) have been used in many plant
phenological studies, and have shown a strong relationship between the timing of key growth

15
stages and climate variability at the regional and global scale (Justice et al., 1985; Keenan et al.,
2014; Migliavacca et al., 2011).
Dryland plant phenology studies have been conducted across the globe, and generally support
forecasts of dryland ecosystem expansion (Archer et al., 2000; Naito & Cairns, 2011; Van
Auken, 2009). In addition to the heterogeneous spatial distribution of plants in drylands, which
often creates for phenological studies (Browning et al., 2012; Walker et al., 2014), other research
hurdles include: a low abundance of historical and present dryland research relative to
ecosystems such as rainforest or temperate forests; and the strong dependence of dryland
phenophase development on precipitation that is uncommon and spatially heterogeneous in
drylands making some pheonphase shifts difficult to determine (Gibbens, 1991; Reynolds et al.,
2004). Often, it is difficult for satellite based imagery to detect subtle phenophase shifts due to
the high occurrence bare ground in drylands (Walker et al., 2012), which can be overcome
through regular ground-based measurements (i.e. human observations) to calibrate/verify more
broad-scale remotely sensed data (satellite images) (Badeck et al., 2004; Vilhar et al., 2013).
Although human observations are susceptible to high levels of subjectivity, observations of this
kind have proven useful for calibrating other methods in order to useful remote sensing
techniques that can be applied at a broader scales and across biomes (Graham et al., 2009; Mbow
et al., 2013).
Readily available historical dryland plant phenology research does not compare to that of other
ecosystems such as, temperate or rainforest biomes (Richardson et al., 2007; Yanoff &
Muldavin, 2008). When using key words to search for relevant studies within the Science Direct
database, the number of papers located for “temperate plant phenology” or “rainforest plant

16
phenology” resulted in over 4,000 studies; whereas, the number of papers relevant to “dryland
plant phenology” only results in about 700 papers (Science Direct). Another possible
contribution to the lack of attention drylands have received may be that dryland phenophase
transitions rely heavily on certain climatic events, such as rainfall, which can be sporadic and
uncommon in drylands making detection of key phenophase shifts difficult (Munson et al., 2015;
Walker, 2014; Yanoff & Muldavin, 2008). In other biomes, phenophase shifts appear to be
controlled by temperature and photoperiod, which are more readily measured and consistent
from year to year (Kramer et al., 2000; Melaas et al., 2013; Piao et al., 2007; Richardson et al.,
2007).
1.2. Goals and objectives
The overarching goal of this study is to determine the spatial and temporal dynamics of plant
phenology in different land cover types of a northern Chihuahuan Desert landscape. To achieve
this goal the study links: (1) conventional phenocam-derived analysis of species and landscape
plant phenology in different land cover types with, (2) a novel analysis of phenocam imagery
that utilizes alternate color space. Specifically, this study will address the following questions in
two primary chapters (Questions 1 and 2 ~ Chapter 2, Question 3 ~ Chapter 3):
1. Is landscape plant phenology spatially variable across a Chihuahuan Desert landscape?
2. Do different plant species drive greening trends in different land cover types?
3. Does phenological analysis with alternate color space offer advantages over customary
approaches that utilize only RGB color space?

17
1.3. Study Area
This study was performed on the United States Department of Agriculture-Agricultural Research
Services (USDA-ARS) Jornada Experimental Range (JER; Figure 1.5) that hosts the Jornada
Basin Long Term Ecological Research (LTER) site and encompasses an approximate area of
200,000 ha in Doña Ana County, southern New Mexico, USA in the northern Chihuahuan
Desert, the largest desert in North America (Figure 1.5) (Campbell, 1929; Herrick et al., 2006;
Peters et al., 2013). Although the JER can be considered a shrubland as a whole, with
Creosotebush (Larrea tridentata) and Mesquite (Prosopis glandulosa) as the two dominant plant
species, there are still a variety of land cover types (LCT) present. In Chapter two, this study
focuses on four different plant communities (land cover types) commonly found throughout the
Chihuahuan Desert: shrubland, shrubland-sandy ridge, grassland-tobosa playa, and grassland
(Figure 1.6) (Peters et al., 2006). Subtle differences in elevation, soil composition, plant
population, soil permeability, and other variables can change the LCT for a given area (Peters
and Gibbens, 2006; Peters et al., 2013). The advantage of examining phenological trends in these
different land cover types is that research findings can be extrapolated regionally using land
cover maps.

18
Figure 1.5: The USDA-ARS JER near Las Cruces, New Mexico. This JER research range is managed by
the New Mexico State University (NMSU). Map courtesy of (jornada-www.nmsu.edu).

19
Figure 1.6: Phenocam locations within the JER. Starting from the left: [1] IBPE1 - grassland, [2] PAS9 -
grassland, [3] SCAN1 - grassland-tobosa playa, [4] SCAN2 - shrubland-sandy ridge, [5] TWEE -
shrubland, and [6] TROM - shrubland.

20
1.3.1. Climate
The Climatic record for the JER dates back to 1915, making this an ideal location for long-term
research. The Chihuahuan desert is the largest desert in North America and is located in the rain
shadow of the Sierra Madre Occidental and the Sierra Madre Oriental, which flank the western
and eastern edges of the desert, respectively (Shmida et al., 1985; Sowell, 2001). Based on the
Koppen classification the JER is a mid-latitude (cold) desert (Havstad et al., 2006) due to its high
solar radiation, low relative humidity, wide ranges in diurnal temperatures, variability in annual
precipitation, and high rates of evaporation, which surpasses precipitation allowing a moisture
deficit to persist. The JER has a mean annual air temperature of 15°C and a mean annual
precipitation of approximately 250 mm, most of which falls as rainfall in late summer monsoons
that are common between July and September (Bestelmeyer et al., 2013; Havstad et al., 2006).
1.3.2. Landscape
The JER (783km2) is located in Southern New Mexico within the Chihuahuan Desert (Havstad et
al., 2000). Landforms include, but are not limited to, rocky mountain slopes, stony bajadas, and
silty basin floors (Monger, 2006). Soil development since the Quaternary has been influenced by
fluctuations in climate, parent material and topographical position (Gile et al., 1981; Monger,
2006). The JER has a slightly sloping surface that is modified by winds and creates the coppice
dunes seen throughout the range. As classified by the USDA ARS JER habitats of the JER are
regionally representative of the northern Chihuahuan Desert (Havstad et al., 2006).

21
1.3.3. Vegetation
Grasslands flourished throughout the American south-west over 150 years ago (Ares et al., 1974;
Bhark & Small, 2003; Dick-peddie, 1975); however, with extensive land settlement and grazing
between the late 1800’s and mid 1900’s (Ares et al., 1974) and persistent drought during the
1950’s, the abundance of grass cover has decreased in the region (Gibbens & Beck, 1988).
Generally, the plant communities in the region are classified as desert-grassland transition
(Peters and Gibbens, 2006). The JER, specifically, has been classified as having five major land
cover types (Peters and Gibbens, 2006) which include: [1] grasslands dominated by Black Grama
(Bouteloua eriopoda), [2] playa grasslands, shrublands dominated by [3] Tarbush (Flourensia
cernua), or [4] Creosotebush (Larrea tridentata), or [5] Honey Mesquite (Prosopis glandulosa).
For this study, eight key plant species that are common throughout the Chihuahuan Desert were
selected for observation. The species are as follows: Honey Mesquite (Prosopis glandulosa),
Creosotebush (Larrea tridentata), Tarbush (Flourensia cernua), Bush Muhly (Muhlenbergia
porteri), Mesa Dropseed (Sporobolous flexuosus), Tobosa Grass (Pleuraphis mutica), Fluff
Grass (Dasyochloa pulchella), and Black Grama (Bouteloua eriopoda). The following sections
briefly outline the distinguishing biological characteristics for each of these species (Table 1.1).

22
Table 1.1: Focal plant species monitored in this study. Sites can be seen in Figure 2.3, above. Taxonomy
is derived from the USDA Plant Database (http://plants.usda.gov/java/).

23
Honey Mesquite (Prosopis glandulosa)
Knowledge of the physical appearance and temporal emergence of key phenophases of any plant
species is critical for accurate observation and record keeping, which will be briefly covered for
each of the following species. Prosopis glandulosa (honey mesquite) is a C3 deciduous shrub and
lives for approximately 200 years (Figure 1.7) (Peters and Gibbens, 2006). Honey mesquite is a
member of the Fabaceae (legume) family, and can occupy about 30-55% of the plant land cover
for plant communities in which it dominates (honey mesquite shrublands or shrublands) (Peters
and Gibbens, 2006). Honey mesquite has very extensive and deep roots (Gibbens & Lenz,
2001a) and is present throughout the JER but is most common on gravelly/sandy soils (Havstad
et al., 2006). Honey mesquite can grow to heights of about 6m tall, but has been documented to
have a greater total biomass below-ground then above-ground (Gibbens & Lenz, 2001a). When
not managed in grazed rangelands, honey mesquite can be an aggressively invasive that displaces
other native species that are not as tolerant to disturbance (Moran et al., 1993). This feature in
part explains the extensive expansion of this and other grazing-tolerant shrub species in
southwest rangelands over past century (“Desert Plants…,” n.d.). Honey mesquite has the
capacity to form symbiotic relationships with N-fixing bacteria (Geesing et al., 2000).
One or two spines can be seen at each node and leaves are alternate and bipinnate. There is one
paired division per leaf and about 6 to 15 leaflets (15 to 62 mm long) per pinna (“Welcome
to…," n.d.). Leaves bud in late spring/early summer. Commonly, leaves reach full size 45 to 60
days after bud break. Flowers typically bloom from spring into summer and resemble yellowish
tiny frothy-like clusters of flowers called catkins ("Plants &…," n.d.). Seedpods are 7 to 20 cm
long with a reddish-brown tint and inside are the brown seeds that reach 6 to 7 mm in length

24
(Havstad et al., 2006). Honey mesquite seeds are vital energy sources for some small and large
mammals such as, jack rabbits and cattle. When milled, humans can also use these seeds as meal
or flour (“Plants &…,” n.d.). Numerous bird species are known to nest in the canopy of honey
mesquite and are benefited from the height advantage and protection these plants offer (“Desert
Plants…,” n.d.). Honey mesquite is expansive and highly tolerant of dryland conditions, making
it an important plant species in southwest dryland ecosystem studies.
Creosotebush (Larrea tridentata)
Larrea tridentata (creosote bush) is an evergreen long-lived C3 perennial shrub (Figure 1.8)
(Miller & Huenneke, 2000) that is common throughout the U.S. Southwest drylands (Reynolds,
1986), especially on well drained slopes and plains where caliche is present. It can occupy about
28-45% of the land cover within a given LCT (Peters and Gibbens, 2006). Creosote bush also
prefers more porous soils unlike honey mesquite (Hamerlynck et al., 2000). Its adaptation to
these water limited environments mostly come from physiological adaptations (Waide et al.,
1999). creosote bush also has a deep, wide spread, and non-overlapping root system
(Hamerlynck et al., 2000). Even when under water stress, creosote bush can maintain relatively
high photosynthetic rates year-round (Franco et al., 1994; Odening et al., 1974) making this
species a hardy in dryland environments. creosote bush is prominently featured in Whitford’s ( et
al., 1997) theory of ‘islands of fertility’ which proposes that due to a combination of canopy
architecture, nutrient uptake, litter fall, and shrub interspace soil erosion, water accumulates
underneath the canopies of creosote bush promoting nutrient uptake and enhanced plant growth
(Whitford et al., 1997).

25
Creosote bush has a large number of medium to large sized flexible branches extending from the
base of the shrub and reaching a height of 1.2 m although it rarely grows up to 3.6 m (Federal
Forest Services). Leaves on creosote bush are usually about 0.6 to 1.2 cm in length and can bud
at various times throughout the year, although they do stop budding and/or drop some of their
leaves in extreme conditions (drought or frost) (Federal Forest Services). Yellow flowers bloom
from February-August when ideal growing conditions prevail (Federal Forest Services). From
the center of the flowers a small greenish fuzzy ball shaped fruit with five sections emerges.
When fruits mature and turn a brown color, they dehisce, and drop five individual ripe fruits
(Federal Forest Services).
Tarbush (Flourensia cernua)
Flourensia cernua (tarbush) is a C3 perennial shrub. It is typically found in arroyos where there
is a higher prevalence of soil water (Figure 1.10), but can also be common near playas on
clay/silt soils (Peters et al., 2006) and areas with predominantly bare ground with dispersed
shrubs and grasses. Sometimes tarbush can be known to smell like tar as a result of out-gassing
of secondary compounds found in the leaves (Estell et al., 1998). It also has an elaborate root
system that allows it to acquire water both deep in the soil and near the soil surface (Gibbens &
Lenz, 2001b). It can tolerate flooding but only for a short amount of time (Dick-Peddie, 1993).
Instead of a trunk, tarbush has branches that extend obliquely from the base and can grow up to 2
m tall. Tarbush has smooth, dark green leaves that are alternate and range from 1.7 to 2.5 cm in
length and 1 cm in width (“Plants &…,” n.d.). Flowers on tarbush are yellow, small, single, and
often difficult to see and occur in late spring. Each flower head has up to 20 flowers and the
seeds are flattened and hairy achenes.

26
Bush Muhly (Muhlenbergia porteri)
Muhlenbergia porteri (bush muhly) is a C4 perennial grass and is most common between
boulders, cliffs, in between shrubs, near dry arroyos, and in some grassland (Figure 1.9). It is
grazed on by cattle (Miller & Donart, 1981; “Plants &…,” n.d.; “Welcome to…,” n.d.) and can
be heavily grazed during the winter when the availability of other grasses is sparse (Miller &
Donart, 1981). It can be seen most commonly underneath honey mesquite or creosote bush
(Chew, 1982; Miller & Donart, 1981) as described by the ‘islands of fertility’ hypothesis
(Whitford et al., 1997) discussed above. It can reach heights from 25 to 100 cm tall. Initial leaf
growth in bush muhly can be seen in early spring to late summer and flowering can be seen in
early spring and summer (Kemp, 1983; Livingston et al., 1995). Leaf blades are usually flat or
folded, 0.5 to 2 mm wide, and usually rough in texture (“Plants &…,” n.d.). Flowers are fine,
many-branched, purplish, and when it is in full bloom the entire plant can have a cobwebby
appearance.
Tobosa Grass (Pleuraphis mutica)
Pleuraphis mutica (tobosa grass) is a perennial grass (Figure 1.11). Tobosa grass is also heavily
grazed on by cattle and horses (“Welcome to…,” n.d.), but can recover from grazing relatively
quickly (“Plants &…,” n.d.). This grass grows best on clay-like soils and slopes. Roots for
tobosa grass are shallow and can extend from 0.6 to 1.8 m in depth (“Plants &…,” n.d.). This is
an erect grass that can to grow to approximately 0.6 m at maturity (“Welcome to…,” n.d.). Its
active growth period falls between spring and summer and can produce light yellow flowers,
with ripe brown seed color that follows flowering. Its seed production begins mid-late summer
and ends in the fall (“Welcome to…,” n.d.).

27
Mesa Dropseed (Sporobolus flexuosus)
Sporobolus flexuosus (mesa dropseed) is a perennial grass (Figure 1.12) and is most common on
sandy and/or loamy soils with a preference for well-drained soils (“Welcome to…,” n.d.). Mesa
dropseed are short-lived (4-5 years) perennials and greens up during the spring, and if the
conditions are ideal, again in late fall. During extreme drought periods, mesa dropseed goes into
dormancy. Culms can grow up to 2 m tall. The growing season for this grass is from March
through November. Mesa dropseed produces hermaphrodite inflorescences in late fall
(September to November) (Gibbens, 1991). Seeds are open, oblong panicles that can grow to 10-
30 cm long (“Plants &…,” n.d.). Mesa dropseed has one floret that produces one small seed with
a hard coat.
Fluff Grass (Dasyochloa pulchella)
Dasyochloa pulchella (fluff grass) is also a C4 perennial grass (Figure 1.14). This grass is a
colonizing species with a short-lived life cycle typically found on gravelly soils in ecosystems
with a high percentage of bare ground (Pezzani et al., 2006). It can form open mats and is
weakly rooted (“Plants &…,” n.d.). In its initial growth the leaves are light green, and once they
have matured turn into a green whitish color. The clusters usually bend over to the ground and
root themselves (“Plants &…,” n.d.). This grass displays an erect growth pattern that forms
culms 4 to 10 cm in height. Their “fluffy” appearance develops at the maturity of their growth
stage and is caused by fascicled spikelets with white hairs (Powell, 1998). It has 1-5 spikelets
which are attached to the rachis and contain 5-10 florets. Flowering occurs mid-July through
mid-September.

28
Black Grama (Bouteloua eriopoda)
Bouteloua eriopoda (black grama) is a perennial grass (Figure 1.13) that is widely regarded as a
major source of fodder for livestock in the southwest U.S. where it is sometimes cut for hay
(USDA-plants). Black grama mostly grows in gravelly soils, sandy dunes, and is seldom seen in
clay soils (USDA-plants), in contrast to the other grasses examined in this study. Black grama
forms a weak sod and roots at the nodes of the stems and is an ideal species to prevent soil
erosion (Federal Forest Services). The leaf blade can grow from 25 to 71 cm in height and can
roll inwards during drier periods (USDA-plants). The stem is solid and the seed-head has 3 to 8
spikes per head and 18 to 20 spikelets per spike (USDA-plants). In order to have successful
asexual reproduction by stoloniferous growth, the plant generally requires two successive
favorable growing seasons (USDA-plants).

29
Figure 1.7: A-B. A. U.S. distribution map of the shrub Prosopis glandulosa - honey mesquite (USDA
Plants Database), B. honey mesquite on the JER displaying full leaf extension (photo taken: August
2013).
Figure 1.8: A-B. A. U.S. distribution map of the evergreen Larrea tridentata - creosote bush (USDA
Plants Database), B. creosote bush at the JER displaying young unfolded leaves (photo taken: June 2013).

30
Figure 1.9: A-B. A. U.S. distribution map of the deciduous shrub Flourensia cernua - tarbush (USDA
Plants Database), B. tarbush at the JER displaying flower buds (photo taken: September 2013).
Figure 1.10: A-B. A. U.S. distribution map of the grass Muhlenbergia porteri - bush muhly (USDA
Plants Database), B. Bush muhly at the JER displaying ripe grains (photo taken: August 2014).

31
Figure 1.11: A-B. A. U.S. distribution map of the grass Pleuraphis mutica - tobosa grass (USDA Plants
Database), B. tobosa grass at the JER displaying initial leaf growth and tall shoots of grass blades (photo
taken: August 2014).
Figure 1.12: A-B. A. U.S. distribution map of the grass Sporobolus flexuosus - mesa dropseed (USDA
Plants Database), B. Mesa dropseed at the JER displaying ripe grains (photo taken: August 2014).

32
Figure 1.13: A-B. A. U.S. distribution map of the grass Dasyochloa pulchella - fluff grass (USDA Plants
Database), B. fluff grass at the JER displaying flower heads (photo taken: August 2015).
Figure 1.14: A-B. A. U.S. distribution map of the grass Bouteloua eriopoda - black grama (USDA Plants
Database), B. Black grama at the JER displaying flower heads (photo from: museum2.utep.edu).

33
2. Inter-comparison of plant and landscape phenology in different land cover
types on the Jornada Experimental Range
2.1 Introduction
Monitoring and understanding phenology trends in drylands are critical for predicting the type of
impact this ecosystem will have on regional and global scale atmospheric carbon cycling (Gao &
Reynolds, 2003; Migliavacca et al., 2011). When compared to other systems, dryland plant
phenological studies and its contribution to these land atmosphere interactions have not been as
heavily explored (Kurc & Benton, 2010). Since drylands cover about 40% of the Earth’s land
surface, and are predicted to significantly expand over the next few decades (Peters et al., 2006;
Poulter et al., 2014; Reynolds et al., 2007) improving our understanding of the spatiotemporal
dynamics and the controls of phenology are paramount for improving land-management,
modeling future ecosystem states, and understanding how change in drylands may impact the
Earth System. This chapter uses remote sensing techniques commonly practiced for studies
similar to this; specifically, time lapse images analysis was used to capture greening trends from
2012 to 2015. Greening trends at the species level were compared to landscape level greening
with the aim of capturing species drivers and contributors of overall landscape greening, in other
words, this comparison will give insight to suggest which plant species have more influence on
landscape level greening.
2.2 Methods
This study spanned four land cover types (LCTs) on the Jornada Experimental Range (JER;
Figure 1.6), which were chosen to represent the variety of LCTs seen more broadly throughout
the Chihuahuan Desert. Phenocams were used to capture images at regular intervals at each site

34
(Table 2.1). Images captured at solar noon were used in analyses to minimize the potential of
shading in the field of view and both limit complications associated with differences in sun angle
in analyses, and more accurately represent phenological trends across LCTs. Images from the
Wingscape cameras have shown to display degradation in the quality of the images over time
with a shadow gradient that required pre-processing before any analysis could be performed
(Figure 2.1).
Collaboration with the Department of Computer Science and UTEP’s Cyber-ShARE Center for
Excellence has assisted the study develop a program within MatLab that can calibrate, correct,
and process the images. Dr. Geovany Ramirez, a former doctoral graduate in computer science,
collaboratively developed this software (hereafter Phenology Analyzer Software or PAS). In
order to calibrate the images, we captured images of a near-perfect white board for each camera
to quantify the shadow gradient. A model of this gradient was generated to visualize the error,
and in order to correct this gradient, a gradient with opposite color values was laid on top of the
first model Figure 2.2. After calibration, the software can automatically correct all the images in
a specified directory (Figure 2.3) and save the new images into a different directory to avoid
overwriting original images. Image analysis was performed with the PAS that allows users to
apply multiple regions of interest (ROIs) of different shapes and sizes, and to save ROIs for
future use and reference (Figure 2.4 a & b). PAS also allows users to view images with multiple
color spaces such as the ones previously mentioned: RGB, HSV, and L*a*b*. These alternate
color spaces will be explored in the next chapter. Once the all images have been calibrated,
corrected, and processed with PAS (Figure 2.5 a-f) the values are saved as a csv file where more
statistical analysis can be performed.

35
This section of the study aims (1) to determine if there is any spatial variability in landscape
plant phenology across LCTs, and (2) to determine if there are specific plant species that seem to
drive plant phenology within each LCT. Specific landscape and individual plant species ROIs
were chosen for Objective 1 (Figure 2.5 a-f) and processed with PAS. Landscape ROIs were
chosen to include the top half of the landscape within the field of view (excluding the sky),
which captures vegetation representative of each LCT and omits most bare ground captured in
the lower half of the field of view (Fig. 2.5 a-f). Species specific ROIs were chosen based on
which plants within the field of view were most visible and had the least amount of overlap from
other plants in order to best represent each species (Fig. 2.5 a-f). After the images were
processed for all hours, a filter was used to include only images captured at noon (solar zenith
angle; solar noon) to minimize shadowing and challenges in image analysis associated with
shifts in sun angle (Keenan et al., 2014; Peters et al., 2014). The same process was used for both
the landscape and species specific comparisons. The green chromatic coordinate (GCC) index
(Ahrends et al., 2008; Richardson et al., 2007; Sonnentag et al., 2012; Toomey et al., 2015) was
used to determine phenological changes across four years of phenocam images acquired as
described above, phenological trends will be referenced as ‘GCC’ from this point forward.
This formula - derived from the standard red, green, and blue (RGB) color model has been used
in multiple studies, including dryland studies to show seasonal change in vegetation phenology
and thus represents a proven metric suitable for inter-comparison between ecosystems and
studies. GCC was used to plot phenological trends at the landscape and species level for each of
the LCTs as shown below (Fig. 2.7- 2.14). Scatter plots were created for each of the ROIs (Fig.
2.5a-f) with a 28-day running average plotted on top to help visualize trends. Figure 2.7 displays
the landscape level phenological trends captured with time lapse images along with daily

36
precipitation, which has been shown to be an important driver of greening trends in dryland
ecosystems (Lesica & Kittelson, 2010; Ogle & Reynolds, 2004).
Table 2.1: Cameras used to address objectives one and two. Only photos captured at solar noon were
used for this project to reduce shading and other complications associated with the analysis of imagery
acquired at different sun angles.

37
Figure 2.1: Image depicting the color gradient (area enclosed in red rectangle) for the TWEE site. All
cameras displayed a similar error and were corrected independently.
Figure 2.2: A model of the calibration process with the Phenology Analyzer Software, which identified
and corrected the shadow gradient (enclosed in the red rectangle) displayed in the TWEE image (Fig.
2.1). [1] ‘Calibration Image’ captured a pure white panel by the camera - red rectangle is the same area as
displayed in Fig. 2.1 which depicts the gradient issue; [2] The ‘Illumination Model’ is the visual model
generated from the white panel image; [3] The ‘Compensation Image’ is the same gradient from
‘Illumination Model’, but with opposite values.

38
Figure 2.3: A model of the Phenology Analyzer Software correcting the TWEE images from [1]
specified directory; [2] Original image (gradient enclosed in red rectangle) and the [3] corrected image
(fixed gradient enclosed in red rectangle); There is also the option to correct the images in a [4] ‘Raw’ or
‘Normalized’ process which can then be [5] stored in a new directory.
Figure 2.4 a&b: A screen shot of the Phenology Analyzer Software processing images for the TWEE
site. Images were used from the directory occupied with corrected images (step 5 in Fig. 2.3). [1]
‘Directory’ which the PAS will use to load all images to be processed; [2] Multiple color spaces included
in the analysis; [3] An ROI can either be a rectangle, ellipse, polygonal shape, or a free-hand shape. Each
can be saved and stored for future use; [4] ‘Live View’ shows the real time values for each color option;
[5] ‘Plots’ generates a quick and dirty plot for any of the options selected; [6] ‘Save Data’ will save the
values generated from the ROIs into a .csv file; [7] A rectangular ROI was chosen to represent the
landscape phenological phases; and [8] free-hand ROIs were carefully chosen and outlined to represent
the phenological phases for Honey Mesquite within this LCT.

39
Figure 2.5 a-f: Regions of interest (ROI) used for landscape and species specific phenology. White =
landscape; green = honey mesquite; orange = mesa dropseed; pink = black grama; pink (dashed) = mix of
black grama and mesa dropseed; purple = tobosa grass; blue = tarbush; red = creosote bush; red (dashed)
= mix of creosote bush and tarbush; yellow = bush muhly. (a) IBPE1 ROIs – grassland; (b) PAS9 ROIs –
grassland; (c) SCAN1 ROIs – grassland-tobosa playa; (d) SCAN2 ROIs – shrubland-sandy ridge; (e)
TROM ROIs – shrubland; and (f) TWEE ROIs – shrubland.

40
2.3 Results
TROM is a creosotebush-honey mesquite shrubland site with bush muhly growing in shrub
interspaces (Figure 2.5 e). Similar to all LCTs, except SCAN2, TROM displayed a generally
broad peak during the growing season of 2012 that consisted of three separate spikes in GCC
between July and September, which was not evident in 2013 or 2014 for this LCT (Figure 2.7).
Each of these spikes in GCC followed three rain events that occurred about 1-2 weeks prior to
peaks in GCC. In 2013, there was an early GCC peak (mid-June) that preceded the first rainfall
events for the year. After the rainfall events occurred between July through August, TROM
peaked in GCC from September through October. In 2014, another early spike in GCC was
captured mid-June which did not seem to follow any precipitation events. This was the last peak
in GCC captured for TROM before the camera no longer functioned (Figure 2.7).
TWEE is also a creosote-honey mesquite shrubland site with bush muhly intertwined beneath
the canopies of shrubs with bare ground exposed between shrubs (Figure 2.5 f). TWEE also
displayed three peaks in GCC that like TROM followed three separate rainfall events. In 2013,
an early peak in GCC was captured that did not seem to follow any significant precipitation
events. The second peak in 2013 was also seen in September/October (as in TROM) after heavy
rainfall events in the preceding months. In 2014, there was another early peak in GCC at TWEE,
which occurred in mid-June and did (similar to 2013) not follow any heavy precipitation (Figure
2.7). Towards the end of October there was another peak in GCC for TWEE, but this peak
(unlike the first) did follow a precipitation event (Figure 2.7). Similar patterns were also seen in
2015, until the camera stopped working in mid-August.

41
SCAN1 - grassland-tobosa playa - displays almost no bare ground and is dominated by tobosa
grass with scattered honey mesquite (Figure 2.5 c). SCAN2 - shrubland-sandy ridge – has more
bare ground exposure than SCAN1, but is still dominated by tobosa grass with scattered
establishments of honey mesquite, tarbush, and creosotebush (Figure 2.5 d). SCAN1 and SCAN2
displayed similar trends in generalized landscape GCC greening and browning trends across all
years (Figure 2.7). SCAN1 displays one clear peak in GCC in 2012, which occurs around the
same time as that for TWEE, PAS9, and IBPE1; however, it does not display a prominent second
peak in GCC that year like other LCTs (Figure 2.7). During 2013 - 2015, peaks in GCC occurred
slightly ahead of those for other LCTs (mid-late August). Unlike GCC trends in other sites, there
was a negligible seasonal shift in GCC for 2012 (Figure 2.7). During 2013 and 2014, peaks in
GCC aligned well with trends documented for the other LCTs (Figure 2.7). The camera ceased
functioning in spring 2015 for SCAN2.
PAS9 and IBPE1 display a low amount of bare ground and include the same dominant plant
species - black grama, mesa dropseed, and honey mesquite (Figure 2.5 b & a, respectively).
These grassland sites displayed similar patterns in GCC, but the camera at PAS9 did not function
after mid-2014. The two peaks in GCC (June and August) were evident for both grassland sites
in both 2012 and 2013 (Figure 2.7). IBPE1 continued recording through 2015 and resulted in
GCC peaks around mid-late August for both 2014 and 2015 (Figure 2.7).
Precipitation is a limiting factor for plant life in drylands which can directly affect phenology
which may result in shifts throughout trophic levels. 2012 and 2013 experienced very few rain
events (drought years) which led to a low amount of annual cumulative precipitation especially
when compared to the proceeding years (Figure 2.6). 2013 experienced a few more intense

42
rainfall events with the highest input of 42 mm in September. 2014 and 2015 both received
noticeably more rainfall events than 2012 and 2013, making these years ‘abnormally wet years’
(Figure 2.6). The highest rainfall event in 2014 was about 45 mm in late September. Although
2015 did not receive as many intense precipitation events as 2014, rainfall input was more
consistent throughout the year (Figure 2.6).
Figure 2.6: Daily and cumulative rainfall (mm) for 2012-2015. MOY = month of the year.
Figure 2.7: Landscape phenology trends across all land cover types (LCTs) from 2012-2015. Each LCT
scatter plot has a 28-day moving average trend line set on top. TROM = black; TWEE = red; SCAN1 =
green; SCAN2 = purple; PAS9 = yellow; IBPE1 = blue.

43
Since honey mesquite (C3 shrub) and warm season C4 graminoid species were present at all six
sites, this allowed the study to explore these trends across all sites for similarities or
dissimilarities (Figure 2.8 a&b). For the most part, GCC trends for honey mesquite and
graminoids were consistent across all LCTs (Figure 2.8 a), meaning that the timing of peaks in
maximum greening captured by the cameras were similar across sites for each species. There
were at least two separate and distinct peaks in 2012 for both honey mesquite and graminoid
GCC values (mid-July and mid-September) with the exception of TROM, which captured three
peaks (July-September). SCAN2 did not see any peaks in GCC for 2012 and retained a
comparatively flat line all year (Figure 2.8 a). There was another early peak in GCC for honey
mesquite seen across all LCTs in 2013 around mid-June. The three peaks in greening can also be
seen for graminoid species in 2012, but only for the TROM site. All other LCTs seemed to stay
flat until the next peak in productivity in mid-September (Figure 2.8 b), which was also captured
for honey mesquite (Figure 2.8 a).
2014 honey mesquite productivity seemed to repeat the same pattern observed in 2013,
displaying an early peak in GCC (Figure 2.8 a) with the exception of SCAN2, which remained
flat until the occurrence of precipitation events during September (Figure 2.8 b). Both the TROM
and PAS9 cameras ceased working mid-August. In 2014, GCC for honey mesquite at TWEE,
IBPE1, and SCAN1, no second peak in greenness was observed. Although there was no early
peak in GCC for graminoid species, there was a peak in GCC after the rainfall events that
occurred between July and September 2014. Rainfall events in 2015 were followed by peaks in
GCC for honey mesquite and graminoid species at the TWEE, SCAN2, and IBPE1 sites (Figure
2.8 a & b).

44
Figure 2.8 a&b: Phenology trends (GCC) for honey mesquite and graminoids observed in all LCTs. (a)
Shrub - honey mesquite. (b) GCC values for all graminoids (TROM & TWEE- bush muhly; SCAN1 &
SCAN2-tobosa grass; PAS9- mesa dropseed; and IBPE1- black grama / mesa dropseed mix) across all
four LCTs. TROM = black; TWEE = red; SCAN2 = purple; SCAN1 = green; PAS9 = yellow; IBPE1 =
blue.

45
In order to determine which plant species appears to drive GCC in each LCT, we plotted
landscape GCC (black trend line included across all the LCTs) against GCC values generated for
each species. For this section of the results, we will focus on the species specific GCC trends and
assess how closely the match landscape trends. At IBPE1, landscape GCC was most strongly
aligned with graminoid species GCC - green trend line (Figure 2.9). At PAS9 landscape trends in
GCC closely matched GCC trends for mesa dropseed (Figure 2.10). For SCAN1, landscape GCC
appeared to be most strongly aligned with trends in tobosa grass (Figure 2.11). At SCAN2
(Figure 2.12) landscape GCC and species specific GCC trend aligned well with all species
(tobosa grass, honey mesquite, creosote bush, and tarbush)suggesting that there was no dominant
species driving landscape patterns in phenology at this site. TROM landscape GCC aligned with
almost all species trends with the exception of creosote bush (Figure 2.13). Some LCTs
displayed clear similarities in landscape and species specific GCC trends while others did not. At
TWEE, landscape GCC appeared to be controlled by honey mesquite and creosote bush (Figure
2.14).
Figure 2.9: Landscape and species 28-day GCC trend lines plotted for IBPE1 (grassland) from 2012-
2015. Black = landscape; green = honey mesquite; and pink (dashed) = black grama & mesa dropseed.
Blue dashed line is cumulative precipitation for each individual year.

46
Figure 2.10: Landscape and species 28-day GCC trend lines plotted for PAS9 (grassland) from 2012-
2015. Black = landscape; green = honey mesquite; pink = black grama; and orange = mesa dropseed.
Blue dashed line is cumulative precipitation for each individual year.
Figure 2.11: Landscape and species 28-day GCC trend lines plotted for SCAN1 (grassland-tobosa playa)
from 2012-2015. Black = landscape; green = honey mesquite; and purple = tobosa grass. Blue dashed
line is cumulative precipitation for each individual year.
Figure 2.12: Landscape and species 28-day GCC trend lines plotted for SCAN2 (shrubland sandy-ridge)
from 2012-2015. Black = landscape; green = honey mesquite; purple = tobosa grass; red = creosote
bush; and blue = tarbush. Blue dashed line is cumulative precipitation for each individual year.

47
Figure 2.13: Landscape and species 28-day GCC trend lines plotted for TROM (shrubland) from 2012-
2015. Black = landscape; green = honey mesquite; red = creosote bush; blue = tarbush; red (dashed) =
mix of creosote bush and tarbush; and yellow = bush muhly. Blue dashed line is cumulative precipitation
for each individual year.
Figure 2.14: Landscape and species 28-day GCC trend lines plotted for TWEE (shrubland) from 2012-
2015. Black = landscape; green = honey mesquite; red = creosote bush; and yellow = bush muhly. Blue
dashed line is cumulative precipitation for each individual year.

48
2.4 Discussion
The main goal for this chapter was to document landscape and species-specific greening (GCC)
trends across different LCTs commonly seen throughout the JER and northern Chihuahuan
Desert (Peters and Gibbens, 2006). More specifically, the aim was to identify species-specific
drivers and/or contributors to landscape greening trends within each LCT. When compared to
other LCTs, the earlier landscape greening (early June) captured by cameras in the shrubland
LCTs was most likely driven by shrub level greening trends which dominate this LCT
(Huenneke et al., 2002; Maynard et al., 2016). Since 2012-2013 received little to no rain input in
the earlier part of the year, the early peak in greening from shrubs is evidence in its likely ability
to access water at deeper soil horizons (coupled with ideal climatic conditions, i.e. temperature)
from which initial growth is stimulated (Browning et al., 2012; Gibbens & Lenz, 2001a). This
may also be evidence that the productivity of shrubs is also tied to winter water supply and
possibly the lower evapotranspiration at this of the year, thus allowing deeper water infiltration
and transport by deep rooted plants (Gibbens & Lenz, 2001a; Munson et al., 2013).
During the non-drought years (2014-2015) there is a clear shift in the response time for the first
spring green-up events, which can be seen across most non-shrub dominated LCTs (Lesica &
Kittelson, 2010; Reynolds et al., 1999). Earlier precipitation events for 2014-2015 resulted in
quick responses for overall landscape greening trends which might be attributed to the quick
responses commonly seen in grasses (Munson et al., 2013; Walker et al., 2014). Grasses are
known for quickly reacting to precipitation events that reach their shallow root systems (Gibbens
& Lenz, 2001a; Peters et al., 2010) resulting in greater seasonal fluctuation tied to rainfall events
(Peters et al., 2014; Peters et al., 2013). There seemed to be strong ties between earlier greening

49
events during non-drought years and later greening events during drought years (Munson et al.,
2013, 2015).
The grassland-tobosa playa was the only LCT that captured a clear species driver for overall
landscape phenology trends. This site is dominated by tobosa grass with few mesquite plants
scattered throughout (Peters & Gibbens, 2006); therefore, the cameras were likely able to capture
the greening signatures from the greater species coverage of tobosa grass over shrubs, resulting
in similar greening trend lines at the species and landscape level. Due to the higher rate of
seasonal fluctuation in grasses, long-term phenological monitoring is critical when examining
drought effects on the properties and dynamics of a dryland region especially in an area where
cattle grazing is a critical contributor to plant stress (Havstad et al., 2006; Peters & Gibbens,
2006). Ranchers and land manager outside the JER could incorporate the low-cost, high data
quality, and quick installation of phenocams to adjust cattle grazing patterns according to
precipitation input and grass phenology patterns (Browning et al., 2015). This applied method
would allow ranchers to practice a more sustainable land management strategy, resulting in a
more productive and responsive ecosystem (Frank et al., 2013). This application might not only
continue to provide a critical food source for cattle, but it may also allow the ecosystem to
improve carbon storage (Frank et al., 2013; Han et al., 2016).

50
3. Use of alternate color space to derive phenological trends in a Chihuahuan
Desert shrubland
3.1 Introduction
Since its inception in 1912, the JER has played a key role in the development of new and/or
more efficient methods of data collection and analysis. Currently, phenophase observations are
primarily conducted by human observers periodically recording changes on tagged plants over
time (Denny, 2012). This method appears to offer high fidelity information but has
disadvantages including low cost efficiency due to the physical requirement of researchers to
make these observations; a risk of data inconsistency due to differences in subjectivity between
observers; and low frequency of data collection due to the presence required by an observer
(Booth et al., 2006; Kurc & Benton, 2010; Menzel, 2002). Although this method is tied to some
challenges, it is still necessary to serve as ground control for testing newer more efficient data
collection methods (Crimmins & Crimmins, 2008; Richardson et al., 2007).
One of the newer, highly preferred, and reliable methods of data collection for phenological
studies is the use of time lapse photography from stationary digital cameras – or ‘phenocams’ as
they are now more commonly referred (Crimmins & Crimmins, 2008; Keenan et al., 2014;
Richardson et al., 2007; Toomey et al., 2015). This method (as opposed to the former method
mentioned) is relatively cost efficient due to the minimal field presence required of human
observers (Benton, 2009; Seefeldt & Booth, 2006). It also eliminates the issue of subjectivity and
increases the frequency of data collection, allowing researchers to capture phenophase changes at
finer temporal scales and from plant to landscape spatial scales (Ahrends et al., 2008; Benton,
2009; Neeser et al., 2000; Richardson et al., 2007). The introduction of phenocams has been
challenged by the need for processing and analyzing large quantities of phenocam images, and

51
although there are established and trusted methods already set in place, there are still a myriad of
digital image analysis techniques that have yet to be explored for vegetation phenological
research.
Time lapse images for phenological studies are most commonly analyzed with imagery software
where regions of interest (ROIs) are chosen by the user and the red, green, blue (RGB) color
channels are used to assign a quantitative values to the ROIs chosen (Benton, 2009; Kercher et
al., 2003; Neeser et al., 2000). In this study, we apply this same technique of analyzing the
digital spectral signature within ROIs but we explore the potential of alternate color space for
advancing this field of research. Specifically, we focused on camera 2 at the UTEP-SEL site at
the JER (Figure 3.2), where we extracted landscape and species (creosote bush and honey
mesquite) ROIs from 2011-2015 with the PAS. We derived different color channel values for
these ROIs and conducted an inter-comparison of these values with detailed phenophase
measurements (Figure 3.5 - 3.7).
3.2 Methods
This study utilized a phenocam (Figure 3.2; Table 2.1) that was situated in a study area (Latitude:
32.581956, Longitude: -106.635025) with an elevation of 1188m and slope of ~2 degrees (Figure
3.1). This study site is maintained and operated by the University of Texas at El Paso’s Systems
Ecology Lab (UTEP-SEL) and is located within a shrubland LCT (Fig. 3.1 ).Time lapse images
were captured hourly from 700 to 1900 spanning 2011-2015, but only images at solar noon were
used in this study to avoid complications in digital image analysis caused by sun angle (Figure
3.3 a) (Gonzalez-Alonso, 2011; Kurc & Benton, 2010). PAS was used to process images for
landscape and species specific ROIs (Figure 3.4 a-c) that included only honey mesquite and

52
creosote bush plants since these were the only two species that could be clearly distinguished
within the field of view (Figure 3.3 b). Images were filtered to include only those with DN
values >39, which helped eliminate images that were overcast, or marred by other uncommon
phenomenon that could impact results (Figure 3.3 c) (following the method of Toomey et al.,
2015).
Images were processed with the color space options: RGB, HSV and L*a*b*. Most plant
phenological studies - focused on northern hardwood forests and rarely expand into the dryland
ecosystems - apply RGB channels and typically analyze data derived for spectral indices such as
GEI or GCC (Equation 1 and 3), which have proven to be reliable and emphasize on the green
channel from RGB which is especially important for plant phenological studies(Benton, 2009;
Kurc & Benton, 2010; Toomey et al., 2015). However, this project explores alternate color
spaces and compares them to physical phenophase observations (Figure 3.3 - 3.5) made weekly
on plants at the site. A goal of this study is to determine if less investigated color channels (such
as HSV and L*a*b*) in the analysis of phenocams imagery offers a greater potential for
documenting phenological trends in dryland shrublands.
Weekly phenological observations (Figure 3.5 – 3.7) were made at the site from 2011-2015,
which included the shrubs creosote bush and honey mesquite. Phenological observations were
made using standardized status protocols reported in Denny (et al., 2014), which are based on
recording the presence/absence of pre-defined criteria that is indicative of a specific phenophase,
meaning if a phenophase was present on a tagged plant, it was given a “1” for that day, and if it
was absent it was given a “0.” In this study, phenophase measurements from sites along two
transects adjacent to the phenocam’s field of view (Table 3.1; Figure 3.1) were totaled per

53
transect then averaged between the two in order to establish a dataset representative of the
landscape at the study site (Figure 3.3 d). The phenocam data was then aligned for comparison
(by date) with the averaged phenophase observations (Figure 3.3 e).
To enhance inter-comparison of the phenophase and phenocam datasets, cluster analysis of the
phenophase data was conducted with the Hierarchical Clustering option in JMP 4. Cluster
analyses were conducted separately on observations collected for creosote bush and honey
mesquite (Figure 3.3 f). Cut off points were chosen to better represent any major shifts in
phenophases (i.e. seasons). Specifically, clusters of three and four were chosen to determine
which was best suited for capturing major shifts in phenophases (Figure 3.3 g). Following cluster
analysis, a one-way ANOVA in JMP 4 was used to determine which color channel derived from
the landscape and species specific ROIs could best capture the phenological state represented by
each cluster (Figure 3.3 h). A Tukey HSD test in JMP 4 was then run (only on significant results
from the ANOVA test) to determine which clusters were significantly different from each other,
thus identifying the color channels that might best capture each shift in phenology (Figure 3.3 i).

54
Figure 3.1: The USDA-ARS JER on the left with a zoomed in view of UTEP-Systems Ecology Lab
(SEL) site within the eastern JER. Red dots are phenology transects monitored at the site, and the two
transects circled in green are included in this study.
Figure 3.2: UTEP-SEL’s site with the relative field of view for the camera used to address research
objective 3.

55
Figure 3.3 a-i: Flow chart of methods for Ch. 3. (a) phenocam image dataset; (b) process images with
PAS; (c) filter images for any noise; (d) phenology dataset; (e) align image and phenology datasets by
date; (f) run hierarchical cluster analysis on phenology data; (g ) cluster groups into seasons; (h) one-way
ANOVA on clusters and spectral indices from images; (i) Tukey test to determine which clusters best
described by spectral indices. Steps in blue re: phenocam images only; steps in green re: phenology
dataset only; and steps in orange re: both datasets together.

56
Figure 3.4 a-c: Field of View for cam2 within PAS. (a) The rectangle overlaid on the image is the
landscape ROI; (b) the five free-hand ROIs encircle individual creosote bush plants; (c) and five free-
hand ROIs encircle individual honey mesquite plants.

57
Table 3.1: The number of individuals for each key plant species observed at site and transect.

58
Figure 3.5 a-f: Key phenophases for Larrea tridentata (creosote bush): (A) breaking leaf buds; (B) young
unfolded leaves; (C) open flowers; (D) full flowering; (E) ripe fruits; (F) flower buds; and (G) fruits
(Denny et al., 2012). (Photographs by: Libia Gonzalez)

59
Figure 3.6 a-f: Key phenophases for Prosopis glandulosa (honey mesquite): (A) breaking leaf buds; (B)
young unfolded leaves; (C) >25% of full leaf size; (D) >75% of full leaf size; (E) 50% leaves fallen; (F)
all leaves fallen (Denny et al., 2012). (Photographs by: Libia Gonzalez) Additional phenophases
monitored for honey mesquite are given in Figure 3.5.

60
Figure 3.7 g-l: Key phenophases for Prosopis glandulosa (honey mesquite): (G) open flowers; (H) full
flowering; (I) fruits; (J) ripe fruits; (K) recent fruit drop; and (L) flower buds (Denny et al., 2012).
(Photographs by: Libia Gonzalez)

61
3.3 Results
The hierarchical cluster analysis from JMP captured temporal shifts in the timing of key
phenophases with moderate overlap in the occurrence of some phenophases. For the three cluster
analysis, creosote bush was grouped by: (1) max greening, (2) flowering, and (3) fruiting (Figure
3.8). For the four cluster analysis for creosote bush, the shifts captured were: (1) green-up, (2)
max greening, (3) flowering, and (4) fruiting (Figure 3.9) where green-up was only captured in
2011. For the three cluster analysis for honey mesquite the shifts captured were: (1) dormant, (2)
green-up/max greening, and (3) flowering/fruiting (Figure 3.10) where fruiting/flowering was
only captured in 2013-2015. Finally, the four shifts captured in the four cluster analysis for
honey mesquite were: (1) senescence/dormant, (2) green-up/max greening, (3) flowering, and (4)
fruiting, where 3 and 4 were only captured in 2013-2014, as shown in Figure 3.11.
There were no significant relationships from the one-way ANOVA for creosote bush spectral
indices at the species level for the three-cluster classification. There were, however, strong
relationships between phenocam spectral properties for creosote bush at the landscape level and
the three clusters (i.e. phenophase shifts; Table 3.2). The Tukey test showed there was no color
model that could strongly capture the max greening phenophase. There were, however, a few
color channels that captured flowering and fruiting phenophases, but the most significant (for
both phenophase shifts) was for ‘Hue’ (from the HSV color model) with a p-value of 0.0055
(Table 3.2). GEI and GCC were also able to capture flowering and fruiting with p-values of
0.0162 and 0.0246, respectively.

62
The four-level classification for creosote bush had various color channels capable of capturing
the four phenophase shifts, supported by the Tukey HSD test (Table 3.3). ‘BlueDN’ and
‘totalRGB’ were able to capture ~75% of all phenophases - early green-up, greening, and fruiting
- with the strongest p-value of <0.0001. As for the flowering phenophase, only ‘Hue’ was able to
capture flowering with a p-value of 0.0279 (Table 3.3.). For this same classification, ANOVA’s
at the landscape level showed that early-green up and flowering were captured by ‘Hue’ with a
strong p-value of 0.005 (Table 3.4). ‘GEI’ was able to capture greening with a p-value of 0.0108
and fruiting was captured best (p-value 0.0122) by ‘BlueDN’, which was the same for the
species level values mentioned above (Table 3.4).
Figure 3.8: Key creosote bush phenophase presence (%) with cluster groups (3) plotted in the
background. Black line represents the clusters established from the Cluster Analysis.

63
Figure 3.9: Key creosote bush phenophase presence (%) with cluster groups (4) plotted in the
background. Black line represents the clusters established from the Cluster Analysis.
Figure 3.10: Key honey mesquite phenophase presence (%) with cluster groups (3) plotted in the
background. Black line represents the clusters established from the Cluster Analysis.
Figure 3.11: Key honey mesquite phenophase presence (%) with cluster groups (4) plotted in the
background. Black line represents the clusters established from the Cluster Analysis.

64
Table 3.2: Difference of means for phenological observations (3 clusters) of creosote bush captured by
landscape ROI color channels. Rows in grey signify non- significant results from the one-way ANOVA.
Numbers in grey represent no difference in means from the other phenophases. Numbers in red (p-value)
are colored in order of significance - deep and bold red are highly significant.

65
Table 3.3: Difference of means for phenological observations (4 clusters) of creosote bush captured by
species ROI color channels. Rows in grey signify non- significant results from the one-way ANOVA.
Numbers in grey represent no difference in means from the other phenophases. Numbers in red (p-value)
are colored in order of significance - deep and bold red are highly significant.
Table 3.4: Difference of means for phenological observations (4 clusters) of creosote bush captured by
landscape ROI color channels. Rows in grey signify non- significant results from the one-way ANOVA.
Numbers in grey represent no difference in means from the other phenophases. Numbers in red (p-value)
are colored in order of significance - deep and bold red are highly significant.

66
Results for the species level honey mesquite analyses (including 3 phenophases) showed that
differences in mean ‘BlueDN’ could be used to most effectively distinguish the three phenophase
clusters (Table 3.5). At the landscape level comparison, no one color channel could distinguish
all phenophase clusters; rather, different color channels proved to be more effective for a given
phenophase cluster. For the dormant phenophase (cluster 1), there were a few color channels that
captured this cluster effectively including ‘redDN’, ‘GEI’, ‘GCC’, ‘totalRGB’, ‘Hue’, ‘Value’,
‘a*’, and ‘test’ (all had p <0.0001, Table 3.6). ‘L*’ proved to be the best channel to capture
green-up/peak productivity with a p-value of 0.0002. Flowering/fruiting was the best captured
with ‘pctR’ (p-value 0.0226; Table 3.6).
Honey mesquite clusters that included 4 groups at the species level did not have a prominent
color channel that captured all phenophase clusters separately (Table 3.7). Color channels that
best captured senescence/dormant, green-up/growing season, and flowering were: ‘greenDN’,
‘blueDN’, ‘pctR’, ‘totalRGB’, and ‘L*’ all with a p-value of <0.0001. The only channel that
captured fruiting was ‘Value’ with a p-value of <0.0001 (Table 3.7). As for the landscape level
color channel values, senescence/dormant, green-up/growing season, and flowering were
captured by ‘redDN’, ‘GCC’, ‘nNDVI’, and ‘Value’ which had p-values of 0.0001 and <0.0001
(Table 3.8). None of the color channels were able to capture fruiting at the landscape level (Table
3.8).

67
Table 3.5: Difference of means for phenological observations (3 clusters) of honey mesquite captured by
species ROI color channels. Rows in grey signify non- significant results from the one-way ANOVA.
Numbers in grey represent no difference in means from the other phenophases. Numbers in red (p-value)
are colored in order of significance - deep and bold red are highly significant.

68
Table 3.6: Difference of means for phenological observations (3 clusters) of honey mesquite captured by
landscape ROI color channels. Rows in grey signify non- significant results from the one-way ANOVA.
Numbers in grey represent no difference in means from the other phenophases. Numbers in red (p-value)
are colored in order of significance - deep and bold red are highly significant.

69
Table 3.7: Difference of means for phenological observations (4 clusters) of honey mesquite captured by
species ROI color channels. Rows in grey signify non- significant results from the one-way ANOVA.
Rows in grey signify these did not have significant results from the one-way ANOVA. Numbers in grey
represent no difference in means from the other phenophases. Numbers in red (p-value) are colored in
order of significance - deep and bold red are highly significant.

70
Table 3.8: Difference of means for phenological observations (4 clusters) of honey mesquite captured by
species ROI color channels. Rows in grey signify non- significant results from the one-way ANOVA.
Numbers in grey represent no difference in means from the other phenophases. Numbers in red (p-value)
are colored in order of significance - deep and bold red are highly significant.

71
3.1 Discussion
This chapter included an exploratory analysis that assessed the utility of alternate spectral indices
from time lapse images to capture key phenological states in a dryland ecosystem. No single
channel or spectral index studied across RGB, HSV, or L*a*b* color space was capable of
capturing all phenological states derived from the cluster analysis of phenophases. Between both
shrub species included, only ‘BlueDN’ for honey mesquite was able to capture all phenological
stages. Interestingly, ’BlueDN’ is not the color channel typically used to capture plant
phenological development in the more commonly studied biomes such as temperate forests
(Richardson et al., 2007; Toomey et al., 2015). Heterogeneity and low foliar cover, characteristic
of a shrubland land cover type, might be factors affecting these more accepted methods of image
analysis for plant phenology in dryland ecosystems (Huenneke et al., 2002; Kurc & Benton,
2010). As such, this study appears to have further illustrated the need for additional exploratory
analyses in alternate color space for phenological studies.
There might also be an underlying issue of distance between the camera lens and the point of
interest (plant) when processing the images. Considering the distance between the camera lens
and plants was about 15 to 20m (Gonzalez-Alonso, 2011), and given that changes in greenness
are subtle for creosote bush (Denny, 2012; Ogle & Reynolds, 2002), these factors may have
made it difficult for cameras to capture those changes over time. ‘Hue’ was able to capture the
timing of flowering and fruiting seen in creosote bush, which may be due to the ability of ‘Hue’
to capture more subtle changes in color than the standard RGB color model as noted in previous
studies (Benton, 2009; Laliberte et al., 2007). Results from this study suggest that new
approaches to dryland phenological studies employing time-lapse image processing should

72
consider alternate approaches to those developed for temperate deciduous and boreal ecosystems,
which typically display higher foliar coverage and more regular shorter-duration annual plant
phenological cycles (Broich et al., 2014).
Based on the results from this chapter, future plant phenological studies in drylands might
consider correlating alternate spectral indices from time lapse images with ecosystem gross
primary productivity (Kurc & Benton, 2010; Moore et al., 2016). This type of data analysis may
give insight as to how dryland plant phenology contributes to land-atmosphere interactions and
specifically carbon and water fluxes (Cong et al., 2013; Poulter et al., 2014; Zhang et al., 2006).
Water is a limiting factor in drylands and there may be correlations between soil moisture depth
and plant phenological responses during long drought periods and/or wetter years (Kurc &
Benton, 2010). Dryland phenology has also shown to play a critical role in regional atmospheric
carbon cycling, but monitoring these greening events has proven to be challenging with the
heterogeneous landscape characteristic of dryland systems (Browning et al., 2015; Huenneke et
al., 2002).

73
4. General Conclusions
In this study, we examined the spatiotemporal variability of plant phenology across different land
cover types that are common on the JER and throughout the northern Chihuahuan Desert.
Grasses seem to drive landscape phenological trends, but only in grassland LCTs; whereas, in
shrubland LCTs grasses act more as contributors to landscape trends. Species cover appears to
play a major role in determining which species will act as the major drivers of landscape plant
phenology for any given LCT. Earlier peaks in greenness for shrubs, relative to grasses, provides
evidence that winter groundwater storage and availability to deeper rooted shrubs likely plays a
critical control in determining the difference in the timing of greening captured by cameras.
Although grass responses are sensitive to irregular precipitation events they still act as key
contributors to overall landscape phenology trends as well as a main source of energy for large
mammals (cattle) and income for ranchers whom own these cattle. Time lapse cameras could be
easily implemented into cattle grazing methods in order to lower the detrimental effects of
overgrazing previously noted across the JER.
This study also explored the capability of capturing key plant phenophase shifts with the use of
alternate spectral indices derived from time lapse image analysis. While there are many studies
readily available from different ecosystems, phenological studies in dryland ecosystems remain
scant. The patchy heterogeneous landscape of drylands cannot compare to that of the lush boreal
or dense rainforest ecosystems where phenocam analysis methods have been developed and
applied. This study has shown that non-conventional approaches using alternate color space
might offer a greater efficacy for capturing phenological events that are not easily discerned in
RGB color space.

74
5. Suggestions for future work
This study sets the tone for many studies that can build from the results found here. One idea is
determining which factors most heavily influence (drive) spatiotemporal variability in dryland
plant phenology. Precipitation is a limiting factor in these environments, but there are other
factors such as temperature, soil moisture, and carbon fluxes that surely play a role in
phenological observations. Correlations can be made between the timing of major shifts in
change (i.e. onset greening, maximal greening, and onset senescence) and the aforementioned
drivers. While there are strong ties to precipitation and grass phenology, there may been another
that more heavily drives shrub phenology. This may elucidate just how dynamic and complex
these dryland plants are and what resources they rely on most for growth.
Scaling is an issue in all ecosystems, but fine-scale studies are not relatively studied in dryland
environments. It is known that plant phenology in drylands contributes significantly to regional
and global scale carbon cycling through photosynthesis. Linking leaf-level physiological
measurements to plant level phenology can help elucidate how these two different scale
measurements are related and how that can be interpreted in future studies through time lapse
image analysis methods. This can also be applied to even broader scale measurements such as
satellite imagery. This may prove helpful by improving data analysis methods in pre-existing
cross scale analysis issues in patchy low plant cover environments that is typical of a dryland
ecosystem.
Another interesting study might be to redo Ch.3, but with images at a smaller scale. The distance
from the camera lens to the point of interest (individual plants in the field of view) may have
played a role in capturing subtle changes in greenness common for dryland shrubs. Maybe a

75
closer range approach will provide allow color signatures from images to better capture
phenological measurements. This, again, is another example of a scaling issue and can elucidate
the strong need to address cross scale analysis especially in heterogeneous landscapes which
makes capturing plant phenology with images even more difficult.

76
6. References
Ahrends, H. E., Brügger, R., Stöckli, R., Schenk, J., Michna, P., Jeanneret, F., … Eugster, W. (2008).
Quantitative phenological observations of a mixed beech forest in northern Switzerland with digital
photography. Journal of Geophysical Research, 113(G4), G04004.
http://doi.org/10.1029/2007JG000650
Allen, J. M., Terres, M. A., Katsuki, T., Iwamoto, K., Kobori, H., Higuchi, H., … Silander, J. A. (2014).
Modeling daily flowering probabilities: Expected impact of climate change on Japanese cherry
phenology. Global Change Biology, 20(4), 1251–1263. http://doi.org/10.1111/gcb.12364
Ansley, R., Ben Wu, X., & Kramp, B. (2001). Observation: Long-term increases in mesquite canopy
cover in a North Texas savanna. Journal of Range Management, 54(2), 171–176.
http://doi.org/10.2307/4003179
Archer, S., Boutton, T. W., Hibbard, K. A., & In, D. (2000). TREES IN GRASSLANDS :
BIOGEOCHEMICAL CONSEQUENCES OF WOODY PLANT EXPANSION Global
Biogeochemical Cycles in the Climate System ( E-D Schulze , SP Harrison , M Heimann , EA
Holland , J Lloyd , IC Prentice , D Schimel , eds .). Academic Press , San I . Intro.
Archibald, S., & Scholes, R. J. (2007). Leaf green-up in a semi-arid African savanna –separating tree and
grass responses to environmental cues. Journal of Vegetation Science, 18(4), 583.
http://doi.org/10.1111/j.1654-1103.2007.tb02572.x
Ares, Fred N. , Campbell, R. S. (1974). The Jornada Experimental Range - An epoch in the era of
southwestern range management. Denver, Colorado: Belke Pringitng Co.
Arora, V. K., & Boer, G. J. (2005). A parameterization of leaf phenology for the terrestrial ecosystem
component of climate models. Global Change Biology, 11(1), 39–59. http://doi.org/10.1111/j.1365-
2486.2004.00890.x
Badeck, F. W., Bondeau, A., Böttcher, K., Doktor, D., Lucht, W., Schaber, J., & Sitch, S. (2004).
Responses of spring phenology to climate change. New Phytologist, 162(2), 295–309.
http://doi.org/10.1111/j.1469-8137.2004.01059.x
Bagella, S., Satta, A., Floris, I., Caria, M. C., Rossetti, I., & Podani, J. (2013). Effects of plant community
composition and flowering phenology on honeybee foraging in Mediterranean sylvo-pastoral
systems. Applied Vegetation Science, 16(4), 689–697. http://doi.org/10.1111/avsc.12023
Barlow, J., Gardner, T. A., Ferreira, L. V., & Peres, C. A. (2007). Litter fall and decomposition in
primary, secondary and plantation forests in the Brazilian Amazon. Forest Ecology and
Management, 247(1), 91–97. http://doi.org/10.1016/j.foreco.2007.04.017
Barsky, B. A. (n.d.). and Color Space Theory.
Bascompte, J., Jordano, P., Bascompte, J., & Jordano, P. (2016). Plant-Animal Mutualistic Networks :
The Architecture of Biodiversity Linked references are available on JSTOR for this article : Plant-
Animal Mutualistic Networks : The Architecture of Biodiversitv, 38(2007), 567–593.
Bater, C. W., Coops, N. C., Wulder, M. a, Hilker, T., Nielsen, S. E., McDermid, G., & Stenhouse, G. B.
(2011). Using digital time-lapse cameras to monitor species-specific understorey and overstorey
phenology in support of wildlife habitat assessment. Environmental Monitoring and Assessment,

77
180(1–4), 1–13. http://doi.org/10.1007/s10661-010-1768-x
Benton, L. M. L. (2009). Automated repeat digital photography for continuous phenological monitoring:
An analysis of flowering in a semiarid shrubland. Thesis, 77.
Benton, L., Weltzin, J., Richardson, A. D., & Losleben, M. V. (2008). PS 78-72: Towards a nation-wide
phenology camera monitoring network. The 93rd ESA Annual …, 3824. Retrieved from
https://eco.confex.com/eco/2008/techprogram/P14746.HTM
Bestelmeyer, B. T., Duniway, M. C., James, D. K., Burkett, L. M., & Havstad, K. M. (2013). A test of
critical thresholds and their indicators in a desertification-prone ecosystem: more resilience than we
thought. Ecology Letters, 16(3), 339–45. http://doi.org/10.1111/ele.12045
Bhark, E. W., & Small, E. E. (2003). Association between Plant Canopies and the Spatial Patterns of
Infiltration in Shrubland and Grassland of the Chihuahuan Desert, New Mexico. Ecosystems, 6(2),
185–196. http://doi.org/10.1007/s10021-002-0210-9
Booth, D. T., Cox, S. E., & Berryman, R. D. (2006). Point sampling digital imagery with “SamplePoint”.
Environmental Monitoring and Assessment, 123(1–3), 97–108. http://doi.org/10.1007/s10661-005-
9164-7
Bowers, J., & Dimmitt, M. (1994). Flowering phenology of six woody plants in the northern Sonoran
Desert. Bulletin of the Torrey Botanical Club, 121(3), 215–229. Retrieved from
http://www.jstor.org/stable/2997177
Brawata, R. L., Raupach, T. H., & Neeman, T. (2013). Techniques for Monitoring Carnivore Behavior
Using Automatic Thermal Video. Wildlife Society Bulletin, 37(4), 862–871.
http://doi.org/10.1002/wsb.315
Broich, M., Huete, A., Tulbure, M. G., Ma, X., Xin, Q., Paget, M., … Held, A. (2014). Land surface
phenological response to decadal climate variability across Australia using satellite remote sensing.
Biogeosciences, 11(18), 5181–5198. http://doi.org/10.5194/bg-11-5181-2014
Brown, James H.; Reichman, O. J.; Davidson, D. W. (1979). Ecosystems, Granivory in Desert. Annual
Review of Ecology and Systematics, 10, 201–227.
Brown, T. B., Hultine, K. R., Steltzer, H., Denny, E. G., Denslow, M. W., Granados, J., … Richardson,
A. D. (2016). Using phenocams to monitor our changing Earth: toward a global phenocam network.
Frontiers in Ecology and the Environment, 14(2), 84–93. http://doi.org/10.1002/fee.1222
Browning, D. M., Duniway, M. C., Laliberte, A. S., & Rango, A. (2012). Hierarchical analysis of
vegetation dynamics over 71 years: soil-rainfall interactions in a Chihuahuan Desert ecosystem.
Ecological Applications : A Publication of the Ecological Society of America, 22(3), 909–26.
Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/22645820
Browning, D. M., Rango, A., Karl, J. W., Laney, C. M., Vivoni, E. R., & Tweedie, C. E. (2015).
Emerging technological and cultural shifts advancing drylands research and management. Frontiers
in Ecology and the Environment, 13(1), 52–60. http://doi.org/10.1890/140161
Campanella, M. V., & Bertiller, M. B. (2008). Plant phenology, leaf traits and leaf litterfall of contrasting
life forms in the arid Patagonian Monte, Argentina. Journal of Vegetation Science, 19(1), 75–85.
http://doi.org/10.3170/2007-8-18333
Campbell, R. S. (1929). Vegetative Succession in the Prosopis Sand Dunes of Southern New Mexico.

78
Ecology, 10(4), 392–398.
Campillo, C., Prieto, M. H., Daza, C., Moñino, M. J., & García, M. I. (2008). Using digital images to
characterize canopy coverage and light interception in a processing tomato crop. HortScience, 43(6),
1780–1786.
Cheng, Y., Gamon, J., Fuentes, D., Mao, Z., Sims, D., Qiu, H., … Rahman, a. (2006). A multi-scale
analysis of dynamic optical signals in a Southern California chaparral ecosystem: A comparison of
field, AVIRIS and MODIS data. Remote Sensing of Environment, 103(3), 369–378.
http://doi.org/10.1016/j.rse.2005.06.013
Chew, R. M. (1982). Changes in Herbaceous and Suffrutescent Perennials in Grazed and Ungrazed
Desertified Grassland in Southeastern Arizona , 1958-1978, 108(1), 159–169.
Chew, R. W., & Whitford, W. G. (1992). A long-term positive effect of kangaroo rats (Dipodomys
spectabilis) on creosotebushes (Larrea tridentata). Journal of Arid Environments, (22(4)), 375–386.
Cleland, E. E., Chiariello, N. R., Loarie, S. R., Mooney, H. a, & Field, C. B. (2006). Diverse responses of
phenology to global changes in a grassland ecosystem. Proceedings of the National Academy of
Sciences of the United States of America, 103(37), 13740–4.
http://doi.org/10.1073/pnas.0600815103
Cleland, E. E., Chuine, I., Menzel, A., Mooney, H. a, & Schwartz, M. D. (2007). Shifting plant phenology
in response to global change. Trends in Ecology & Evolution, 22(7), 357–65.
http://doi.org/10.1016/j.tree.2007.04.003
Cong, N., Wang, T., Nan, H., Ma, Y., Wang, X., Myneni, R. B., & Piao, S. (2013). Changes in satellite-
derived spring vegetation green-up date and its linkage to climate in China from 1982 to 2010: A
multimethod analysis. Global Change Biology, 19(3), 881–891. http://doi.org/10.1111/gcb.12077
Connolly, C., & Fliess, T. (1997). A Study of Efficiency and Accuracy in the Transformation from RGB
to CIELAB Color Space, 6(7), 1046–1048.
Cook, B. I., Wolkovich, E. M., Davies, T. J., Ault, T. R., Betancourt, J. L., Allen, J. M., … Travers, S. E.
(2012). Sensitivity of Spring Phenology to Warming Across Temporal and Spatial Climate
Gradients in Two Independent Databases. Ecosystems, 15(8), 1283–1294.
http://doi.org/10.1007/s10021-012-9584-5
Cooper, E. J., Dullinger, S., & Semenchuk, P. (2011). Late snowmelt delays plant development and
results in lower reproductive success in the High Arctic. Plant Science, 180(1), 157–167.
http://doi.org/10.1016/j.plantsci.2010.09.005
Crimmins, M. a, & Crimmins, T. M. (2008). Monitoring plant phenology using digital repeat
photography. Environmental Management, 41(6), 949–58. http://doi.org/10.1007/s00267-008-9086-
6
Crimmins, T. M., Crimmins, M. A., & David Bertelsen, C. (2010). Complex responses to climate drivers
in onset of spring flowering across a semi-arid elevation gradient. Journal of Ecology, 98(5), 1042–
1051. http://doi.org/10.1111/j.1365-2745.2010.01696.x
Deacy, W. W., Leacock, W. B., Eby, L. A., & Stanford, J. A. (2016). A time-lapse photography method
for monitoring salmon ( Oncorhynchus spp. ) passage and abundance in streams. PeerJ, 4, e2120.
http://doi.org/10.7717/peerj.2120

79
Denny, E. G. (2012). USA-NPN Technical Report Series 2012 004 USA National Phenology Network
USA NPN Plant and Animal Phenophase Definitions.
Desert Plants - Cactus - Wildflowers - Flora - DesertUSA. (n.d.). Retrieved December 13, 2016, from
http://www.desertusa.com/flora.html
Dick-peddie, W. A. (1975). Vegetation of southern New Mexico, 81–84.
Dick-Peddie, W. A. (1993). New Mexico vegetation: past, present, and future.
Elzinga, J. A., Atlan, A., Biere, A., Gigord, L., Weis, A. E., & Bernasconi, G. (2007). Time after time:
flowering phenology and biotic interactions. Trends in Ecology and Evolution, 22(8), 432–439.
http://doi.org/10.1016/j.tree.2007.05.006
Ernest, S. K. M., Brown, J. H., & Parmenter, R. R. (2000). Rodents, plants, and precipitation: spatial and
temporal dynamics of consumers and resources. Oikos, 88, 470–482. http://doi.org/10.1034/j.1600-
0706.2000.880302.x
Estell, R. E., Fredrickson, E. L., Anderson, D. M., Havstad, K. M., & Remmenga, M. D. (1998).
Relationship of tarbush leaf surface terpene profile with livestock herbivory. Journal of Chemical
Ecology, 24(1), 1–12. http://doi.org/10.1023/A:1022399426352
Euskirchen, E. S., McGuire, A. D., Kicklighter, D. W., Zhuang, Q., Clein, J. S., Dargaville, R. J., …
Smith, N. V. (2006). Importance of recent shifts in soil thermal dynamics on growing season length,
productivity, and carbon sequestration in terrestrial high-latitude ecosystems. Global Change
Biology, 12(4), 731–750. http://doi.org/10.1111/j.1365-2486.2006.01113.x
Facelli, J. M., & Pickett, S. T. A. (1991). Plant litter: Its dynamics and effects on plant community
structure. The Botanical Review, 57(1), 1–32. http://doi.org/10.1007/BF02858763
Ford, A., Roberts, A., & Roberts@rd, A. (1998). Colour Space Conversions.
Forrest, J., Inouye, D. W., & Thomson, J. D. (2010). Flowering phenology in subalpine meadows: Does
climate variation influence community co-flowering patterns? Ecology, 91(2), 431–440.
http://doi.org/10.1890/09-0099.1
Franco, a. C., de Soyza, a. G., Virginia, R. a., Reynolds, J. F., & Whitford, W. G. (1994). Effects of
plant size and water relations on gas exchange and growth of the desert shrub Larrea tridentata.
Oecologia, 97(2), 171–178. http://doi.org/10.1007/BF00323146
Frank, A. S. K., Dickman, C. R., Wardle, G. M., Greenville, A. C., Asner, G., Elmore, A., … Read, J.
(2013). Interactions of Grazing History, Cattle Removal and Time since Rain Drive Divergent
Short-Term Responses by Desert Biota. PLoS ONE, 8(7), e68466.
http://doi.org/10.1371/journal.pone.0068466
Gamon, J. a., Huemmrich, K. F., Stone, R. S., & Tweedie, C. E. (2013). Spatial and temporal variation in
primary productivity (NDVI) of coastal Alaskan tundra: Decreased vegetation growth following
earlier snowmelt. Remote Sensing of Environment, 129, 144–153.
http://doi.org/10.1016/j.rse.2012.10.030
Gamon, J., Field, C., Goulden, M., Griffin, K. L., Hartley, A. E., Joel, G., … Valentini, R. (1995).
Relationships between NDVI, canopy structure, and photosynthesis in three Californian vegetation
types. Ecological …, 5(1), 28–41. Retrieved from http://www.creaf.uab.es/Global-
Ecology/Pdfs_UEG/EcolAppl1995.pdf

80
Gao, Q., & Reynolds, J. (2003). Historical shrub–grass transitions in the northern Chihuahuan Desert:
modeling the effects of shifting rainfall seasonality and event size over a landscape gradient. Global
Change Biology, 1475–1493. Retrieved from http://onlinelibrary.wiley.com/doi/10.1046/j.1365-
2486.2003.00676.x/full
Geesing, D., Felker, P., & Bingham, R. L. (2000). Influence of mesquite (Prosopis glandulosa) on soil
nitrogen and carbon development: Implications for global carbon sequestration. Journal of Arid
Environments, 46(2), 157–180. http://doi.org/10.1006/jare.2000.0661
Gibbens, R. P. (1991). Some effects of precipitation patterns on mesa dropseed phenology. Journal of
Range Management, 44(1), 86–90.
Gibbens, R. P., & Beck, R. F. (1988). Chnages in grass basal area and forb densities over a 64-year period
on grassland types of the Jornada Experimental Range. Plant Ecology, 41(3), 186–192.
http://doi.org/10.1093/infdis/jit783
Gibbens, R. P., & Lenz, J. M. (2001a). Root systems of some Chihuahuan Desert plants. Journal of Arid
Environments, 49(2), 221–263. http://doi.org/10.1006/jare.2000.0784
Gibbens, R. P., & Lenz, J. M. (2001b). Root systems of some Chihuahuan Desert plants. Journal of Arid
Environments, 49(2), 221–263. http://doi.org/10.1006/jare.2000.0784
Gile, L. H., Hawley, J. W., & Grossman, R. B. (1981). Soils and geomorphology in the Basin and Range
area of southern New Mexico: Guidebook to the Desert Project. New Mexico Bureau of Mines &
Mineral Resources., 39.
Gonzalez-Alonso, L. (n.d.). Develpoment of low-cost network of webcams for monitoring plant phenology
in a Chihuahuan Desert shrubland.
Gonzalez-Alonso, L. (2011). Development of a low-cost network of webcams for monitoring plant
phenology in a Chihuahuan desert shrubland. The University of Texas at El Paso.
Graham, E. a., Yuen, E. M., Robertson, G. F., Kaiser, W. J., Hamilton, M. P., & Rundel, P. W. (2009).
Budburst and leaf area expansion measured with a novel mobile camera system and simple color
thresholding. Environmental and Experimental Botany, 65(2–3), 238–244.
http://doi.org/10.1016/j.envexpbot.2008.09.013
Hamerlynck, E. P., Mcauliffe, J. R., & Smith, S. D. (2000). Effects of surface and sub-surface soil
horizons on the seasonal performance of Larrea tridentata (creosotebush). Functional Ecology,
14(5), 596–606. http://doi.org/10.1046/j.1365-2435.2000.00469.x
Han, J., Li, L., Chu, H., Miao, Y., & Chen, S. (2016). The effects of grazing and watering on ecosystem
CO2 fluxes vary by community phenology. Environmental Research, 144, 64–71.
http://doi.org/10.1016/j.envres.2015.09.002
Harris, G. a. (1977). Among Phenology Grass as a Factor of Competition Seedlings. Journal of Range
Management, 30(3), 172–177.
Havstad, K. M., Huenneke, L. F., & Schlesinger, W. H. (2006). Structure and function of a Chihuahuan
desert ecosystem: the Jornada Basin long-term ecological research site.
Havstad, Kris M. , Fredrickson, Ed L. , Huenneke, L. F. (2006). Grazing Lifestock Management in an
Arid Ecosystem. In Structure and function of a Chihuahuan Desert ecosystem the Jornada Basin
long-term ecological research site (pp. 266–277). Oxford, New York: Oxford University Press.

81
Havstad, K., Kustas, W., & Rango, A. (2000). Jornada Experimental Range: A unique arid land location
for experiments to validate satellite systems. Remote Sensing of …, (74), 13–25. Retrieved from
http://www.sciencedirect.com/science/article/pii/S0034425700001188
Hegland, S. J., Nielsen, A., Lázaro, A., Bjerknes, A. L., & Totland, Ø. (2009). How does climate
warming affect plant-pollinator interactions? Ecology Letters, 12(2), 184–195.
http://doi.org/10.1111/j.1461-0248.2008.01269.x
Herrick, J. E., Havstad, K. M., & Rango, A. (2006). Remediation research in the Jornada Basin: past and
future. Structure and function of a Chihuahuan Desert Ecosystem.
Hill, B., Roger, T., & Vorhagen, F. W. (1997). Comparative analysis of the quantization of color spaces
on the basis of the CIELAB color-difference formula. ACM Transactions on Graphics, 16(2), 109–
154. http://doi.org/10.1145/248210.248212
Hingrat, Y., Ysnel, F., Jalme, M. Saint, Le Cuziat, J., Béranger, P.-M., & Lacroix, F. (2007). Assessing
habitat and resource availability for an endangered desert bird species in eastern Morocco: the
Houbara Bustard. Biodiversity and Conservation, 16(3), 597–620. http://doi.org/10.1007/s10531-
005-0603-0
Huemmrich, K. F., Gamon, J. a., Tweedie, C. E., Oberbauer, S. F., Kinoshita, G., Houston, S., … Oechel,
W. C. (2010). Remote sensing of tundra gross ecosystem productivity and light use efficiency under
varying temperature and moisture conditions. Remote Sensing of Environment, 114(3), 481–489.
http://doi.org/10.1016/j.rse.2009.10.003
Huenneke, L. F., Anderson, J. P., Remmenga, M., & Schlesinger, W. H. (2002). Desertification alters
patterns of aboveground net primary production in Chihuahuan ecosystems. Global Change Biology,
8, 247–264. http://doi.org/10.1046/j.1365-2486.2002.00473.x
Hufkens, K., Friedl, M. A., Richardson, A., Milliman, T., & Migliavacca, M. (2004). Vegetation
Phenology from MODIS / AVHRR / PhenoCam : scaling and validation possibilities, 2004.
Ibáñez, I., Primack, R. B., Miller-Rushing, A. J., Ellwood, E., Higuchi, H., Lee, S. D., … Silander, J. a.
(2010). Forecasting phenology under global warming. Philosophical Transactions of the Royal
Society of London. Series B, Biological Sciences, 365(1555), 3247–3260.
http://doi.org/10.1098/rstb.2010.0120
Joiner, J., Yoshida, Y., Vasilkov, a. P., Schaefer, K., Jung, M., Guanter, L., … Belelli Marchesini, L.
(2014). The seasonal cycle of satellite chlorophyll fluorescence observations and its relationship to
vegetation phenology and ecosystem atmosphere carbon exchange. Remote Sensing of Environment,
152, 375–391. http://doi.org/10.1016/j.rse.2014.06.022
Jones, B. M., Arp, C. D., Beck, R. A., Grosse, G., Webster, J. M., & Urban, F. E. (2009). Erosional
history of Cape Halkett and contemporary monitoring of bluff retreat, Beaufort Sea coast, Alaska.
Polar Geography, 32(3–4), 129–142. http://doi.org/10.1080/10889370903486449
Julitta, T., Cremonese, E., Migliavacca, M., Colombo, R., Galvagno, M., Siniscalco, C., … Menzel, A.
(2014). Using digital camera images to analyse snowmelt and phenology of a subalpine grassland.
Agricultural and Forest Meteorology, 198, 116–125. http://doi.org/10.1016/j.agrformet.2014.08.007
Justice, C. O., Townshend, J. R. G., Holben, B. N., & Tucker, C. J. (1985). Analysis of the phenology of
global vegetation using meteorological satellite data. International Journal of Remote Sensing, 6(8),
1271–1318. http://doi.org/10.1080/01431168508948281

82
Keenan, T. F., Darby, B., Felts, E., Sonnentag, O., Friedl, M. A., Hufkens, K., & Keefe, J. O. (2014).
Tracking forest phenology and seasonal physiology using digital repeat photography : a critical
assessment, 24(6), 1478–1489.
Kelly, A. E., & Goulden, M. L. (2008). Rapid shifts in plant distribution with recent climate change.
Proceedings of the National Academy of Sciences of the United States of America, 105(33), 11823–
6. http://doi.org/10.1073/pnas.0802891105
Kemp, P. R. (1983). Phenological patterns of Chihuahuan Desert plants in relations to the timing of water
availability. Journal of Ecology, 71(2), 427–436.
Kercher, S. M., Frieswyk, C. B., & Zedler, J. B. (2003). Effects of sampling teams and estimation
methods on the assessment of plant cover. Journal of Vegetation Science, 14(6), 899–906.
http://doi.org/10.1111/j.1654-1103.2003.tb02223.x
Kidron, G. J., & Gutschick, V. P. (2013). Soil moisture correlates with shrub-grass association in the
chihuahuan desert. Catena, 107, 71–79. http://doi.org/10.1016/j.catena.2013.02.001
Klosterman, S. T., Hufkens, K., Gray, J. M., Melaas, E., Sonnentag, O., Lavine, I., … Richardson, a. D.
(2014). Evaluating remote sensing of deciduous forest phenology at multiple spatial scales using
PhenoCam imagery. Biogeosciences Discussions, 11(2), 2305–2342. http://doi.org/10.5194/bgd-11-
2305-2014
Knapp, A. K., Briggs, J. M., Collins, S. L., Archer, S. R., Bret-Harte, M. S., Ewers, B. E., … Cleary, M.
B. (2008). Shrub encroachment in North American grasslands: Shifts in growth form dominance
rapidly alters control of ecosystem carbon inputs. Global Change Biology, 14, 615–623.
http://doi.org/10.1111/j.1365-2486.2007.01512.x
Kramer, K., Leinonen, I., & Loustau, D. (2000). The importance of phenology for the evaluation of
impact of climate change on growth of boreal, temperate and Mediterranean forests ecosystems: an
overview. International Journal of Biometeorology, 44(2), 67–75. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/10993560
Kurc, S. a., & Benton, L. M. (2010). Digital image-derived greenness links deep soil moisture to carbon
uptake in a creosotebush-dominated shrubland. Journal of Arid Environments, 74(5), 585–594.
http://doi.org/10.1016/j.jaridenv.2009.10.003
Laliberte, a. S., Rango, a., Herrick, J. E., Fredrickson, E. L., & Burkett, L. (2007). An object-based
image analysis approach for determining fractional cover of senescent and green vegetation with
digital plot photography. Journal of Arid Environments, 69(1), 1–14.
http://doi.org/10.1016/j.jaridenv.2006.08.016
Lesica, P., & Kittelson, P. M. (2010). Precipitation and temperature are associated with advanced
flowering phenology in a semi-arid grassland. Journal of Arid Environments, 74(9), 1013–1017.
http://doi.org/10.1016/j.jaridenv.2010.02.002
Lightfoot, D. C., Whitford, W. G., & Cruces, L. (1989). Oecologia and canopy arthropods. Differences,
166–175.
Livingston, M., Roundy, B. A., & Smith, S. E. (1995). Association of native grasses and overstory species
in southern Arizona.
Loomis, R. S., & Connor, D. J. (1992). Crop Ecology: Productivity and Management in Agricultural

83
Systems - R. S. Loomis, D. J. Connor - Google Books. Retrieved December 12, 2016, from
https://books.google.com/books?id=0e5vKY5XDmoC&printsec=frontcover#v=onepage&q&f=false
Lorenz, R. D. (2009). Power law of dust devil diameters on Mars and Earth. Icarus (Vol. 203).
Ma, X., Huete, A., Yu, Q., Coupe, N. R., Davies, K., Broich, M., … Eamus, D. (2013). Spatial patterns
and temporal dynamics in savanna vegetation phenology across the north australian tropical transect.
Remote Sensing of Environment, 139, 97–115. http://doi.org/10.1016/j.rse.2013.07.030
Matuzak, G. D., & Brightsmith, D. J. (2007). Roosting of Yellow-naped Parrots in Costa Rica: estimating
the size and recruitment of threatened populations. Journal of Field Ornithology, 78(2), 159–169.
http://doi.org/10.1111/j.1557-9263.2007.00099.x
Maynard, J. J., Karl, J. W., & Browning, D. M. (2016). Effect of spatial image support in detecting long-
term vegetation change from satellite time-series. Landscape Ecology, 31(9), 2045–2062.
http://doi.org/10.1007/s10980-016-0381-y
Mbow, C., Fensholt, R., Rasmussen, K., & Diop, D. (2013). Can vegetation productivity be derived from
greenness in a semi-arid environment? Evidence from ground-based measurements. Journal of Arid
Environments, 97, 56–65. http://doi.org/10.1016/j.jaridenv.2013.05.011
Melaas, E. K., Friedl, M. A., & Richardson, A. D. (2016). Multiscale modeling of spring phenology
across Deciduous Forests in the Eastern United States. Global Change Biology, 22(2), 792–805.
http://doi.org/10.1111/gcb.13122
Menzel, A. (2002). Phenology: its importance to the global change community, 54(4), 379–385.
Menzel, A., Sparks, T. H., Estrella, N., Koch, E., Aaasa, A., Ahas, R., … Zust, A. (2006). European
phenological response to climate change matches the warming pattern. Global Change Biology,
12(10), 1969–1976. http://doi.org/10.1111/j.1365-2486.2006.01193.x
Migliavacca, M., Galvagno, M., Cremonese, E., Rossini, M., Meroni, M., Sonnentag, O., … Richardson,
A. D. (2011). Using digital repeat photography and eddy covariance data to model grassland
phenology and photosynthetic CO2 uptake. Agricultural and Forest Meteorology, 151(10), 1325–
1337. http://doi.org/10.1016/j.agrformet.2011.05.012
Miller, R. E., & Huenneke, L. F. (2000). Demographic variation in a desert shrub, Larrea tridentate, in
response to a thinning treatment. Journal of Arid Environments, 45(4), 315–323.
http://doi.org/10.1006/jare.2000.0649
Miller, R. F., & Donart, G. B. (1981). Response of Muhlenbergia porteri Scribn. To Season of
Defoliation. Journal of Range Management, 34(2), 91. http://doi.org/10.2307/3898119
Mizunuma, T., Wilkinson, M., L. Eaton, E., Mencuccini, M., I. L. Morison, J., & Grace, J. (2013). The
relationship between carbon dioxide uptake and canopy colour from two camera systems in a
deciduous forest in southern England. Functional Ecology, 27(1), 196–207.
http://doi.org/10.1111/1365-2435.12026
Monger, H. C. (2006). Soil Development in the Jornada Basin. In Structure and function of a Chihuahuan
Desert ecosystem - the Jornada Basin long-term ecological research site (pp. 81–106). Oxford, New
York: Oxford University Press.
Moore, Caitlin E; Keenan, Trevor F; Duursma, Remko A; Albert I J M van Dijk; Hutley, L. B. (2016).
Reviews and syntheses: Australian vegetation phenology: new insights from satellite remote sensing

84
and digital repeat photography, 5085–5102.
Moran, V. C., Hoffmann, J. H., & Zimmermann, H. G. (1993). Objectives, Constraints, and Tactics in the
Biological Control of Mesquite Weeds (Prosopis) in South Africa. Biological Control, 3(1), 80–83.
http://doi.org/10.1006/bcon.1993.1013
Morisette, J. T., Richardson, A. D., Knapp, A. K., Fisher, J. I., Graham, E. A., Abatzoglou, J., … Liang,
L. (2009). Tracking the rhythm of the seasons in the face of global change: phenological research in
the 21st century. Frontiers in Ecology and the Environment, 7(5), 253–260.
http://doi.org/10.1890/070217
Munson, S. M., Muldavin, E. H., Belnap, J., Peters, D. P. C., Anderson, J. P., Reiser, M. H., …
Christiansen, T. a. (2013). Regional signatures of plant response to drought and elevated
temperature across a desert ecosystem. Ecology, 94(9), 2030–41. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/24279274
Munson, S. M., Webb, R. H., Housman, D. C., Veblen, K. E., Nussear, K. E., Beever, E. a., … Tallent, N.
G. (2015). Long-term plant responses to climate are moderated by biophysical attributes in a North
American desert. Journal of Ecology, n/a-n/a. http://doi.org/10.1111/1365-2745.12381
Myneni, R. B., & Williams, D. L. (1994). On the relationship between FAPAR and NDVI. Remote
Sensing of Environment, 49(3), 200–211. http://doi.org/10.1016/0034-4257(94)90016-7
Nagai, S., Yoshitake, S., Inoue, T., Suzuki, R., Muraoka, H., Nasahara, K. N., & Saitoh, T. M. (2014).
Year-to-year blooming phenology observation using time-lapse digital camera images. Journal of
Agricultural Meteorology, 70(3), 163–170. http://doi.org/10.2480/agrmet.D-13-00021
Naito, a. T., & Cairns, D. M. (2011). Patterns and processes of global shrub expansion. Progress in
Physical Geography, 35, 423–442. http://doi.org/10.1177/0309133311403538
NEESER, C., MARTIN, A. R., JUROSZEK, P., & MORTENSEN, D. A. (2000). A Comparison of
Visual and Photographic Estimates of Weed Biomass and Weed Control 1. Weed Technology, 14(3),
586–590. http://doi.org/10.1614/0890-037X(2000)014[0586:ACOVAP]2.0.CO;2
Nelson, K., & Papuga, S. K. (2009). Continuous Monitoring of Dynamic Pulse-Driven Phenological
Phases in a Semiarid Shrubland, 152(2), 2007.
Ober, H. K., STEIDL, R. J., & DALTON, V. M. (2005). RESOURCE AND SPATIAL-USE PATTERNS
OF AN ENDANGERED VERTEBRATE POLLINATOR, THE LESSER LONG-NOSED BAT.
Journal of Wildlife Management, 69(4), 1615–1622. http://doi.org/10.2193/0022-
541X(2005)69[1615:RASPOA]2.0.CO;2
Odening, W. R., Strain, B. R., & Oechel, W. C. (1974). The Effect of Decreasing Water Potential on Net
CO2 Exchange of Intact Desert Shrubs. Ecology, 55(5), 1086–1095. http://doi.org/10.2307/1940358
Ogle, K., & Reynolds, J. F. (2002). Desert dogma revisited: Coupling of stomatal conductance and
photosynthesis in the desert shrub, Larrea tridentata. Plant, Cell and Environment, 25(7), 909–921.
http://doi.org/10.1046/j.1365-3040.2002.00876.x
Ogle, K., & Reynolds, J. F. (2004). Plant responses to precipitation in desert ecosystems: integrating
functional types, pulses, thresholds, and delays. Oecologia, 141(2), 282–94.
http://doi.org/10.1007/s00442-004-1507-5
Parmesan, C., & Yohe, G. (2003). A globally coherent fingerprint of climate change impacts across

85
natural systems. Nature, 421(6918), 37–42. http://doi.org/10.1038/nature01286
Pekel, J. F., Vancutsem, C., Bastin, L., Clerici, M., Vanbogaert, E., Bartholomé, E., & Defourny, P.
(2014). A near real-time water surface detection method based on HSV transformation of MODIS
multi-Spectral time series data. Remote Sensing of Environment, 140, 704–716.
http://doi.org/10.1016/j.rse.2013.10.008
Peñuelas, J., Rutishauser, T., & Filella, I. (2009). Phenology Feedbacks on Climate Change. Science,
324(5929).
Peters, Debra P.C. , Gibbens, R. P. (2006). Plant Communitites in the Jornada Basin: The Dynamic
Landscape. In Structure and function of a Chihuahuan Desert ecosystem - the Jornada Basin long-
term ecological research site (pp. 211–231). Oxford, New York: Oxford University Press.
Peters, A. J., Eve, M. D., Holt, E. H., Whitford, W. G., Whitfordi, W. G., Eve, D., & Howard, E. (2014).
with patterns growth plant community of desert Analysis spectra satellite resolution high temporal,
34(2), 418–432.
Peters, D. P. C., Bestelmeyer, B. T., Herrick, J. E., Fredrickson, E. L., Monger, H. C., & Havstad, K. M.
(2006). Disentangling Complex Landscapes: New Insights into Arid and Semiarid System
Dynamics. BioScience, 56(6), 491. http://doi.org/10.1641/0006-
3568(2006)56[491:DCLNII]2.0.CO;2
Peters, D. P. C., Herrick, J. E., Monger, H. C., & Huang, H. (2010). Soil-vegetation-climate interactions
in arid landscapes: Effects of the North American monsoon on grass recruitment. Journal of Arid
Environments, 74(5), 618–623. http://doi.org/10.1016/j.jaridenv.2009.09.015
Peters, D. P. C., Yao, J., Browning, D., & Rango, A. (2013). Mechanisms of grass response in grasslands
and shrublands during dry or wet periods. Oecologia. http://doi.org/10.1007/s00442-013-2837-y
Pettorelli, N., Pelletier, F., Von Hardenberg, A., Festa-Bianchet, M., & Côté, S. D. (2007). Early onset of
vegetation growth vs. rapid green-up: Impacts on juvenile mountain ungulates. Ecology, 88(2), 381–
390. http://doi.org/10.1890/06-0875
Pettorelli, N., Weladji, R. B., Holand, Ø., Mysterud, A., Breie, H., & Stenseth, N. C. (2005). The relative
role of winter and spring conditions : linking climate and phenology to alpine reindeer body mass,
(September 2004), 24–26. http://doi.org/10.1098/rsbl.2004.0262
Pezzani, F., Montaña, C., & Guevara, R. (2006). Associations between arbuscular mycorrhizal fungi and
grasses in the successional context of a two-phase mosaic in the Chihuahuan Desert. Mycorrhiza,
16(4), 285–295. http://doi.org/10.1007/s00572-006-0044-y
Piao, S., Friedlingstein, P., Ciais, P., Viovy, N., & Demarty, J. (2007). Growing season extension and its
impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Global
Biogeochemical Cycles, 21(3), 1–11. http://doi.org/10.1029/2006GB002888
Plants & Animals | US Forest Service. (n.d.). Retrieved December 13, 2016, from
http://www.fs.fed.us/learn/plants-animals
Poulter, B., Frank, D., Ciais, P., Myneni, R. B., Andela, N., Bi, J., … van der Werf, G. R. (2014).
Contribution of semi-arid ecosystems to interannual variability of the global carbon cycle. Nature,
509(7502), 600–603. http://doi.org/10.1038/nature13376
Powell, A. M. (1998). Trees & shrubs of the trans-pecos and adjacent areas. University of Texas Press.

86
Proulx, R., & Parrott, L. (2008). Measures of structural complexity in digital images for monitoring the
ecological signature of an old-growth forest ecosystem. Ecological Indicators, 8(3), 270–284.
http://doi.org/10.1016/j.ecolind.2007.02.005
Przeszlowska, A., Trlica, M. J., & Weltz, M. A. (2006). Near-Ground Remote Sensing of Green Area
Index on the Shortgrass Prairie. Rangeland Ecology & Management, 59(4), 422–430.
http://doi.org/10.2111/05-059R1.1
Rathcke, B., & Lacey, E. P. (1985). Phenological Patterns of Terrestrial Plants. Source: Annual Review of
Ecology and Systematics Ann. Rev. Ecol. Syst, 16(16). Retrieved from
http://www.jstor.org/stable/2097047
Reynolds, J. F. (1986). Adaptative strategies of desert shrubs with special reference to the creosotebush
(Larrea tridentata).
Reynolds, J. F., Kemp, P. R., Ogle, K., & Fernández, R. J. (2004). Modifying the “pulse-reserve”
paradigm for deserts of North America: precipitation pulses, soil water, and plant responses.
Oecologia, 141(2), 194–210. http://doi.org/10.1007/s00442-004-1524-4
Reynolds, J. F., Smith, D. M. S., Lambin, E. F., Ii, B. L. T., Mortimore, M., Batterbury, S. P. J., …
Walker, B. (2007). Global Desertification : Building a Science for Dryland Development, (MAY),
847–851.
Reynolds, J. F., Virginia, R. A., Kemp, P. R., de Soyza, A. G., & Tremmel, D. C. (1999). IMPACT OF
DROUGHT ON DESERT SHRUBS: EFFECTS OF SEASONALITY AND DEGREE OF
RESOURCE ISLAND DEVELOPMENT. Ecological Monographs, 69(1), 69–106.
http://doi.org/10.1890/0012-9615(1999)069[0069:IODODS]2.0.CO;2
Richardson, A. D., Braswell, B. H., Hollinger, D. Y., Jenkins, J. P., & Ollinger, S. V. (2009). Near-
surface remote sensing of spatial and temporal variation in canopy phenology. Ecological
Applications : A Publication of the Ecological Society of America, 19(6), 1417–28. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/19769091
Richardson, A. D., Jenkins, J. P., Braswell, B. H., Hollinger, D. Y., Ollinger, S. V, & Smith, M.-L.
(2007). Use of digital webcam images to track spring green-up in a deciduous broadleaf forest.
Oecologia, 152(2), 323–34. http://doi.org/10.1007/s00442-006-0657-z
Richardson, A. D., Keenan, T. F., Migliavacca, M., Ryu, Y., Sonnentag, O., & Toomey, M. (2013).
Climate change, phenology, and phenological control of vegetation feedbacks to the climate system.
Agricultural and Forest Meteorology, 169, 156–173. http://doi.org/10.1016/j.agrformet.2012.09.012
Root, T. L., Price, J. T., Hall, K. R., Schneider, S. H., Rosenzweig, C., & Pounds, J. A. (2003).
Fingerprints of global warming on wild animals and plants. Nature, 421(6918), 57–60.
http://doi.org/10.1038/nature01333
Rosenzweig, C., Karoly, D., Vicarelli, M., Neofotis, P., Wu, Q., Casassa, G., … Imeson, A. (2008).
Attributing physical and biological impacts to anthropogenic climate change. Nature, 453(7193),
353–357. http://doi.org/10.1038/nature06937
Saitoh, T. M., Nagai, S., Saigusa, N., Kobayashi, H., Suzuki, R., Nasahara, K. N., & Muraoka, H. (2012).
Assessing the use of camera-based indices for characterizing canopy phenology in relation to gross
primary production in a deciduous broad-leaved and an evergreen coniferous forest in Japan.
Ecological Informatics, 11, 45–54. http://doi.org/10.1016/j.ecoinf.2012.05.001

87
Schwartz, M. D., & Karl, T. R. (1990). Spring Phenology: Nature’s Experiment to Detect the Effect of
“Green-Up” on Surface Maximum Temperatures. Monthly Weather Review.
http://doi.org/10.1175/1520-0493(1990)118<0883:SPNETD>2.0.CO;2
Scott, P. E. (2004). TIMING OF AGAVE PALMERI FLOWERING AND NECTAR-FEEDING BAT
VISITATION IN THE PELONCILLOS AND CHIRICAHUA MOUNTAINS. The Southwestern
Naturalist, 49(4), 425–434. http://doi.org/10.1894/0038-4909(2004)049<0425:TOAPFA>2.0.CO;2
Seager, R., Ting, M., Held, I., Kushnir, Y., Lu, J., Vecchi, G., … Naik, N. (2007). Model projections of
an imminent transition to a more arid climate in southwestern North America. Science (New York,
N.Y.), 316(5828), 1181–4. http://doi.org/10.1126/science.1139601
Sedinger, J. S., & Raveling, D. G. (1986). Timing of Nesting by Canada Geese in Relation to the
Phenology and Availability of Their Food Plants. The Journal of Animal Ecology, 55(3), 1083.
http://doi.org/10.2307/4435
Seefeldt, S. S., & Booth, D. T. (2006). Measuring Plant Cover in Sagebrush Steppe Rangelands: A
Comparison of Methods. Environmental Management, 37(5), 703–711.
http://doi.org/10.1007/s00267-005-0016-6
Shmida, A., Noy, I., Deserts, H., & Shrublands, A. (1985). Biogeography of the desert floras of the world,
12, 23–77.
Skiles, S. M., Painter, T. H., Belnap, J., Holland, L., Reynolds, R. L., Goldstein, H. L., & Lin, J. (2015).
Regional variability in dust-on-snow processes and impacts in the Upper Colorado River Basin.
Hydrological Processes, 29(26), 5397–5413. http://doi.org/10.1002/hyp.10569
Smith, A. (1978). Color gamut transform pairs. ACM Siggraph Computer Graphics, (2), 12–19. Retrieved
from http://dl.acm.org/citation.cfm?id=807361
Sonnentag, O., Hufkens, K., Teshera-Sterne, C., Young, A. M., Friedl, M., Braswell, B. H., …
Richardson, A. D. (2012). Digital repeat photography for phenological research in forest
ecosystems. Agricultural and Forest Meteorology, 152, 159–177.
http://doi.org/10.1016/j.agrformet.2011.09.009
Sowell, J. (2001). Desert Ecology - an introduction to life in the arid southwest. Salt Lake City: The
University of Utah Press.
Thomas, C. D., Thomas, C. D., Cameron, A., Cameron, A., Green, R. E., Green, R. E., … Williams, S. E.
(2004). Extinction risk from climate change. Nature, 427(6970), 145–8.
http://doi.org/10.1038/nature02121
Thomson, B. D., & Siddique, K. H. M. (1997). Grain legume species in low rainfall Mediterranean-type
environments II. Canopy development, radiation interception, and dry-matter production. Field
Crops Research, 54(2), 189–199. http://doi.org/10.1016/S0378-4290(97)00048-8
Thuiller, W., Lavorel, S., Araujo, M. B., Sykes, M. T., & Prentice, I. C. (2005). Climate change threats to
plant diversity in Europe. Proceedings of the National Academy of Sciences, 102(23), 8245–8250.
http://doi.org/10.1073/pnas.0409902102
Toomey, M., Friedl, M. a, Frolking, S., Hufkens, K., Klosterman, S., Sonnentag, O., … Richardson, A. D.
(2015). Greenness indices from digital cameras predict the timing and seasonal dynamics of canopy-
scale photosynthesis. Ecological Applications, 25(1), 99–115. http://doi.org/10.1890/14-0005.1

88
Tucker, C. J., Slayback, D. A., Pinzon, J. E., Los, S. O., Myneni, R. B., & Taylor, M. G. (2001). Higher
northern latitude normalized difference vegetation index and growing season trends from 1982 to
1999. International Journal of Biometeorology, 45(4), 184–190. http://doi.org/10.1007/s00484-001-
0109-8
Van Auken, O. W. (2009). Causes and consequences of woody plant encroachment into western North
American grasslands. Journal of Environmental Management, 90(10), 2931–42.
http://doi.org/10.1016/j.jenvman.2009.04.023
Vanamburg, L. K., Trlica, M. J., Hoffer, R. M., & Weltz, M. A. (2006). Ground based digital imagery for
grassland biomass estimation. International Journal of Remote Sensing, 27(5), 939–950.
http://doi.org/10.1080/01431160500114789
Van Wijk, M. T., Williams, M., Laundre, J. A., & Shaver, G. R. (2003). Interannual variability of plant
phenology in tussock tundra: Modelling interactions of plant productivity, plant phenology,
snowmelt and soil thaw. Global Change Biology, 9(5), 743–758. http://doi.org/10.1046/j.1365-
2486.2003.00625.x
Vezhnevets, V. (n.d.). A Survey on Pixel-Based Skin Color Detection Techniques.
Vilhar, U., Beuker, E., Mizunuma, T., Skudnik, M., Lebourgeois, F., Soudani, K., & Wilkinson, M.
(2013). Tree Phenology. Developments in Environmental Science, 12, 169–182.
http://doi.org/10.1016/B978-0-08-098222-9.00009-1
Wagner, D. (1997). The Influence of Ant Nests on Acacia Seed Production, Herbivory and Soil Nutrients.
The Journal of Ecology, 85(1), 83. http://doi.org/10.2307/2960629
Waide, R. B., Willig, M. R., Steiner, C. F., Mittelbach, G., Gough, L., Dodson, S. I. & Parmenter, R.
(1999). The relationship between productivity and species richness. Annual Review of Ecology and
Systematics, 257–300.
Walker, B. J. (2014). The Interaction of Aeolian and Fluvial Processes in Dry Washes on the Colorado
Plateau , USA.
Walker, J. J., de Beurs, K. M., & Wynne, R. H. (2014). Dryland vegetation phenology across an elevation
gradient in Arizona, USA, investigated with fused MODIS and landsat data. Remote Sensing of
Environment, 144, 85–97. http://doi.org/10.1016/j.rse.2014.01.007
Walker, J. J., De Beurs, K. M., Wynne, R. H., & Gao, F. (2012). Evaluation of Landsat and MODIS data
fusion products for analysis of dryland forest phenology. Remote Sensing of Environment, 117, 381–
393. http://doi.org/10.1016/j.rse.2011.10.014
Welcome to the PLANTS Database | USDA PLANTS. (n.d.). Retrieved December 13, 2016, from
http://plants.usda.gov/java/
White, M. A., & Nemani, R. R. (2006). Real-time monitoring and short-term forecasting of land surface
phenology. Remote Sensing of Environment, 104(1), 43–49. http://doi.org/10.1016/j.rse.2006.04.014
White, M. a., Thornton, P. E., & Running, S. W. (1997). A continental phenology model for monitoring
vegetation responses to interannual climatic variability. Global Biogeochemical Cycles, 11(2), 217–
234. http://doi.org/10.1029/97GB00330
Whitehead, K., Moorman, B., Wainstein, P., & Habib, a. (2010). Monitoring rapidly evolving Landscape
Features using Ground-Based Time-Lapse Photography: A Case Study for Proglacial Icing. 2010

89
Canadian Geomatics Conderence and Symposium of Commison I, International Society for
Photogrammetry and Remote Sensing, (Moorman 2005), 6.
Whitford, W. G., Anderson, J., & Rice, P. M. (1997). Stemflow contribution to the “fertile island” effect
in creosotebush,Larrea tridentata. Journal of Arid Environments, 35(3), 451–457.
http://doi.org/10.1006/jare.1996.0164
Wipf, S., Stoeckli, V., & Bebi, P. (2009). Winter climate change in alpine tundra: Plant responses to
changes in snow depth and snowmelt timing. Climatic Change, 94(1–2), 105–121.
http://doi.org/10.1007/s10584-009-9546-x
Yanoff, S., & Muldavin, E. (2008). Grassland-shrubland transformation and grazing: A century-scale
view of a northern Chihuahuan Desert grassland. Journal of Arid Environments, 72(9), 1594–1605.
http://doi.org/10.1016/j.jaridenv.2008.03.012
Young, D. S., Hart, J. K., & Martinez, K. (2015). Image analysis techniques to estimate river discharge
using time-lapse cameras in remote locations. Computers and Geosciences, 76, 1–10.
http://doi.org/10.1016/j.cageo.2014.11.008
Zhang, X., & Friedl, M. (2004). Climate controls on vegetation phenological patterns in northern mid‐and
high latitudes inferred from MODIS data. Global Change …, 1133–1145.
http://doi.org/10.1111/j.1365-2486.2004.00784.x
Zhang, X., Friedl, M. a., & Schaaf, C. B. (2006). Global vegetation phenology from Moderate Resolution
Imaging Spectroradiometer (MODIS): Evaluation of global patterns and comparison with in situ
measurements. Journal of Geophysical Research, 111(G4), G04017.
http://doi.org/10.1029/2006JG000217

90
7. Curriculum Vitae
Naomi Robin Luna completed her Bachelor’s Degree at The University of Texas at El Paso in
2013. During her last two years she worked as a Research Assistant at UTEP where she
collaborated in several projects related to dryland ecosystems and image analysis for monitoring
plant productivity in these harsh environments. In 2013, she was awarded the EPA Air Quality
Internship at UTEP. While working in the lab, Robin found her passion for research in dryland
ecosystems and decided to enter the Master’s program in Environmental Science at UTEP under
the direction of Dr. Craig E. Tweedie. Her thesis entitled “Spatiotemporal variability in plant
phenology in drylands: a case study from the northern Chihuahuan Desert” investigates the
variability of plant productivity between different plant communities across a dryland ecosystem
via spectral signatures from time lapse imagery; along with this it explores new methods for
analyzing these time lapse images.
During her studies, Robin worked as a research assistant position, as well as one semester as a
teaching assistant giving her the skills to communicate science to other students. Robin
participated in a program called TIERA, working as a mentor, helping students explore the
possibilities beyond academia through research. She has presented at several national
conferences many of them awarded with travel scholarship. Ms. Luna’s research interests focus
on the effects of climate change on plant productivity and its effects on this harsh environment.
Copyright © 2022 FDOKUMEN