
Reproductive factors affecting survivorship in captive callitrichidae

Zoo Biology 13:167-176 (1994)
Spatial Selection in Captive Adult Female Chimpanzees Tammie Bettinger, Janette Wallis, and Tracy Carter
Tulsa Zoological Park, Tulsa, Oklahoma (T.B.); Department of Zoology, Oklahoma State University, Stillwater, Oklahoma (T. B., T. C.); Department of Obstetrics and Gynecology, University of Oklahoma College of Medicine, Oklahoma City, Oklahoma (J. W.)
Spatial selection refers to an animal’s frequent use of certain areas of its enclosure or range. Through differential use of habitat, an individual can limit the amount of time spent in close proximity to other group members. The four adult female chimpanzees at the Tulsa Zoo were studied to determine if they used their enclo- sure differently from one another, if each had selected areas it occupied most frequently, and if an individual’s preferred area was used by other female group members. Pairwise comparisons showed that the females used their island habitat differently from one another with the exception of one pair. Although these two females overlapped in their use of the enclosure, they separated their use tempo- rally, thereby reducing the amount of time spent in close proximity to one another. Each female had a selected site that she occupied more frequently than did the other females. However, there was variation in the extent to which the females occupied each other’s selected sites. Time of day and temperature had no effect on use of the habitat, however, one female showed a seasonal variation in her use of the enclosure. The spatial selection exhibited by these females may be com- parable to the use of core areas by wild female chimpanzees. 0 1994 Wiley-Liss, Inc.
Key words: core area, enclosure, habitat, spacing behavior, spatial selection
INTRODUCTION
The behavior of female chimpanzees (Pun troglodytes) has received less atten- tion from researchers than that of male chimpanzees. Nishida [ 19891 attributes this to an emphasis by researchers on the cooperative behavior of males and the fact that females do not interact frequently and spend much of their time alone or with their
Received for publication April 26, 1993; revision accepted October 28, 1993.
Address reprint requests to Tammie Bettinger, Tulsa Zoological Park, 5701 E. 36th St. No., Tulsa, OK 74115.
0 1994 Wiley-Liss, Inc.

168 Bettinger et al.
offspring. However, recent studies reflect an increased interest in the behavior of female chimpanzees.
Nishida [ 19791 described female association patterns at Mahale as “passive aggregations, ” meaning that associations between females occurred only inadvert- ently when females where accompanying adult males. With the exception of the Bossou population [Sugiyama and Koman, 1979; Sugiyama, 19881, female chim- panzees tend to be less gregarious than males [Goodall, 1986; Boesch and Boesch, 1989; Nishida, 1989; Wrangham et al., 19921. At Gombe, anestrous adult female chimpanzees spent 65-70% of their time alone or with their dependent offspring [Halperin, 1979; Wrangham and Smuts, 19801. When females do interact, their relationships have been characterized as being neutral or slightly aggressive [Wrang- ham et al., 19921.
In addition to being less sociable, females range shorter distances than males [Reynolds and Reynolds, 1965; Wrangham and Smuts, 1980; Goodall, 1986; Hase- gawa, 19901. Anestrous females at Gombe spent 80% of their time in “core areas” about one-half the size of the males’ home range [Wrangham and Smuts, 19801. A core area is defined as an area of the home range that is used disproportionately more than the rest of the range [Jennrich and Turner, 19691. The core areas of female chimpanzees in the wild are not exclusive and may overlap extensively [Wrangham, 19791. However, the relatively solitary nature of females suggests that they may separate themselves temporally in overlapping areas.
Wrangham [1979] suggested that the use of core areas by female chimpanzees may be in response to feeding competition. Indeed, Nishida [1989] found that pro- tection of offspring and competition over food were the two main sources of conflict for the female chimpanzees at Mahale. Thus, females may benefit by having a small core area in which to range. The smaller core area may enable females to identify food sources more easily, and therefore conserve energy by traveling shorter distances. Nishida [ 19891 proposed that female reproductive success may depend on acquiring a core area near the center of the group’s territory. He suggested that females with peripheral ranges may incur a greater risk of aggression from other female group members, as well as from neighboring males who may attack their offspring.
In zoos, the typical chimpanzee group is composed of several adult females and only one or two adult males. Therefore, captive group composition may necessitate frequent interaction between females; a situation which is opposite that found in the wild. Indeed, this is what has been found for the female chimpanzees at the Arnhem Zoo. de Waal [ 19861 reported that the females in his study frequently associated with one another, even forming strong coalitions. However, this finding may be a result of the periodic unstable dominance hierarchy among the male chimpanzees during the study period.
Comparing spacing behavior between captive and wild chimpanzees is further complicated by the fact that captive chimpanzees cannot leave their group. Given adequate space in captivity, however, female chimpanzees could select certain areas of their habitat for more frequent use. By occupying different areas of the enclosure, they could reduce the amount of time they spend in close proximity to one another.
The nine chimpanzees in this study were housed in a large complex outdoor facility, although they could not leave, this facility allowed them the opportunity to separate themselves visually or physically from other group members. Although food was provided and there were no potential dangers such as those experienced by wild
Spatial Selection in Captive Chimpanzees 169
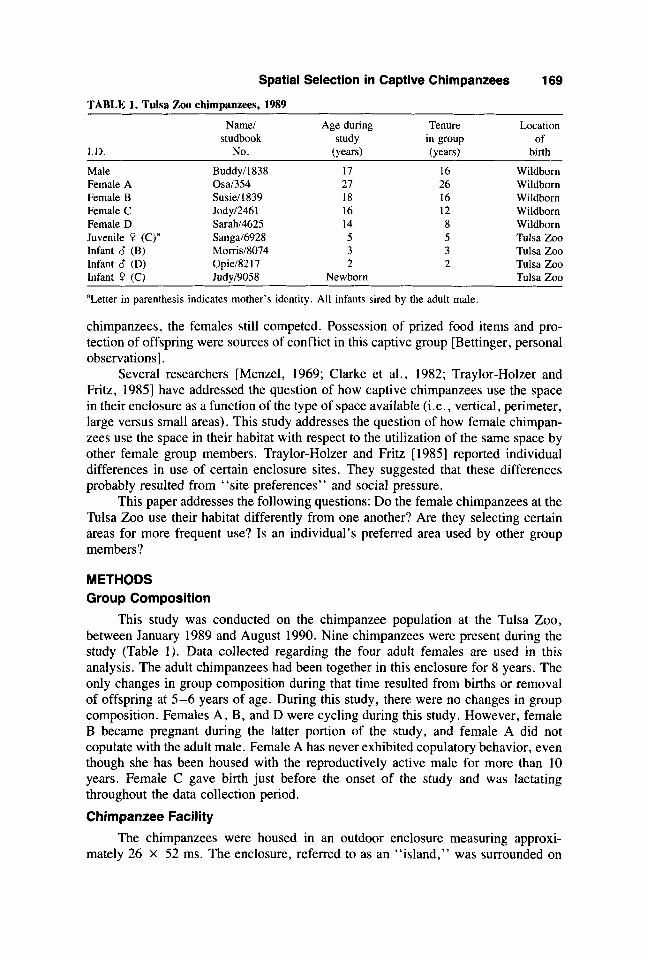
TABLE 1. Tulsa Zoo chimDanzees, 1989 ~ ~~ ~
Name1 Age during Tenure Location studbook study in group of
- I.D. No. (years) (years) birth
Male Buddy11838 17 16 Wildborn Female A Osal354 27 26 Wildborn Female B Susiel1839 18 16 Wildborn Female C Jodyl2461 16 12 Wildborn Female D Sarahl4625 14 8 Wildborn Juvenile P (C)” Sangal6928 5 5 Tulsa Zoo Infant d (B) Moms18074 3 3 Tulsa Zoo Infant 6 (D) Opiel82 1 I 2 2 Tulsa Zoo Infant 0 (C) Judy19058 Newborn Tulsa Zoo
aLetter in parenthesis indicates mother’s identity. All infants sired by the adult male.
chimpanzees, the females still competed. Possession of prized food items and pro- tection of offspring were sources of conflict in this captive group [Bettinger, personal observations].
Several researchers [Menzel, 1969; Clarke et al., 1982; Traylor-Holzer and Fritz, 198.51 have addressed the question of how captive chimpanzees use the space in their enclosure as a function of the type of space available (i.e., vertical, perimeter, large versus small areas). This study addresses the question of how female chimpan- zees use the space in their habitat with respect to the utilization of the same space by other female group members. Traylor-Holzer and Fritz [ 19851 reported individual differences in use of certain enclosure sites. They suggested that these differences probably resulted from ‘‘site preferences” and social pressure.
This paper addresses the following questions: Do the female chimpanzees at the Tulsa Zoo use their habitat differently from one another? Are they selecting certain areas for more frequent use? Is an individual’s preferred area used by other group members?
METHODS Group Composition
This study was conducted on the chimpanzee population at the Tulsa Zoo, between January 1989 and August 1990. Nine chimpanzees were present during the study (Table 1). Data collected regarding the four adult females are used in this analysis. The adult chimpanzees had been together in this enclosure for 8 years. The only changes in group composition during that time resulted from births or removal of offspring at 5-6 years of age. During this study, there were no changes in group composition. Females A, B, and D were cycling during this study. However, female B became pregnant during the latter portion of the study, and female A did not copulate with the adult male. Female A has never exhibited copulatory behavior, even though she has been housed with the reproductively active male for more than 10 years. Female C gave birth just before the onset of the study and was lactating throughout the data collection period.
Chimpanzee Facility
The chimpanzees were housed in an outdoor enclosure measuring approxi- mately 26 X 52 ms. The enclosure, referred to as an “island,” was surrounded on

170 Bettinger et al.
CHIMP ISLAND AT THE TULSA ZOO
Moat I t I I I I I I I
I I I I I I
Den
! I I I I
Moat
Fig. 1 . Diagram of the chimpanzee enclosure at the Tulsa Zoo showing the ground-level grid used for this study. This diagram shows only solid man-made vertical structures. The island was covered in thick natural vegetation and had numerous ropes not shown.
three sides by a water moat 6 m wide and on the fourth side by the night den wall. Included on the island were numerous vertical structures such as ropes, poles, a platform, termite mound, and cargo nets, as well as rocks and natural vegetation. Additionally, four concrete drainage pipes covered with dirt, referred to in this study as “tunnels,” were also in the enclosure. The dense vegetation, tunnels and hilly terrain enabled the chimpanzees to visually isolate themselves from each other and the public. The chimpanzees were on exhibit 6-10 Nday, weather permitting; they had access to an inside den at night and during cold weather (usually below 4.4”C).
Method of Observation and Analysis
Data were collected by instantaneous sampling on a focal animal [Lehner, 19791. Each female was observed for 10 min with data recorded at 30-sec intervals. Each 30-sec sample recorded location, behavior, and group member within 5 ft of the focal animal or in the same tunnel as the focal animal. One hundred hours of data were collected for this study. Order of observation was established by randomization schedule. Females were observed equally during each observation session. Due to weather and maintenance constraints, the chimpanzees did not have access to the island equally throughout the year. Therefore, samples could not be evenly distrib- uted by season (fall = 15%, spring = 2496, summer = 53%, winter = 8%).
For analysis, the enclosure was divided into 7 sections of approximately equal size (Fig. 1). Each section was further subdivided according to characteristic of the space: horizontal space (on the ground), vertical space (cargo nets, poles, ropes or vegetation that support the individual off the ground), tunnels, platform, and termite mound. This gave a possibility of 19 locations that individuals could occupy.
Data were analyzed using chi-square tests to determine how each subject’s use of the enclosure compared to that of other female group members. Since the data set

Spatial Selection in Captive Chimpanzees 171
TABLE 2. Pairwise chi-square tests of space use in each female's most frequently occupied area (df = 1)
Female Area A B C D _____ ~
A 1T chi-square
B 3T chi-square
C 1P chi -square
D 5T chi-square
P*
P*
P*
P*
555.96 <o.o001 28.86
<O.OOol 67.79 <O.OOol
304.18 <O.OOol
147.26 <O.OOol 238.57 <O.OOol
358.52 <o.o001 532.41 <O.OOoI
19.75 <0.0001
315.36 <O.oOOl 527.22 <o.o001 12.93
= O.OOo3
*Significant probability levels remain significant after sequential Bonferonni adjustment for overall alpha level set a priori at P = 0.05.
for each female was compared to that of other females, chi-square probability results were adjusted using the sequential Bonferonni adjustment for multiple related tests [Rice, 19891. Chi-square tests were also used to determine whether individuals al- tered their use of the enclosure in response to diurnal, thermal, or seasonal changes. Time of day was divided into three categories: (1) before 10:00, (2) 1O:Ol to 14:00, and (3) 14:Ol to 17:OO. Temperature was divided into three categories: (1) below 16"C, (2) 16.1" to 29.9"C, and (3) above 30°C. Seasons were divided into four categories: (1) winter = Dec-Feb, (2) spring = Mar-May, (3) summer = Jun-Aug, (4) fall = SepNov.
RESULTS Use of the Enclosure
Figure 2 shows the percentage of observations each female spent in the 19 possible locations. Each subject had an area of the enclosure that she used signifi- cantly more often than did other female group members (Table 2).
Five of the six comparisons between the subjects' overall use of the island showed that each female used the space differently from the other females with the exception of the comparison between females C and D (Table 3).
The overlap in habitat use by females C and D included areas they occupied as well as areas they did not use. Reviewing the two areas most frequently occupied by these females reveals that they are the inverse of one another (Fig. 2). Although these females frequented the same sites, they tended temporally to separate their use of overlapping areas, thus reducing the amount of time they simultaneously occupied the same area (Table 4). Females C and D seldom occupied areas frequented by female A or female B.
None of the females altered their use of the enclosure in response to variation in temperature or differing time of day. Female C was the only individual to show a seasonal variation in her use of the enclosure (df = 51, chi-square = 69.03, P = 0.047).
Use of Another Individual's Preferred Area Figure 3 shows a comparison of each female's most frequently occupied area
and the extent to which the other females used this same space. The areas most often

172 Bettinger et al.
Area 1H 1T tv 1M 1P 2H 2T 2v
6V BH
7H BV
a- e
I
8
M
BH BV 7H
ezza I
- B
3v 4H 4v 6H 6T
Area 1H 1T 1v 1M 1P 2H 2T 2v 3H
4H 4 v 6H 6T 6V
0 Animat C Anlmal 0
Fig. 2. Percentage of observations each female chimpanzee spent in the 19 locations in the enclosure.
occupied by female A and B, respectively, were seldom used by other group mem- bers. This was true whether or not the female that frequently occupied the area was present (Table 4, female present; Fig. 2, overall use). Locations most frequently used by female C and female D were also used by female A, but seldom by female B.
DISCUSSION
The disproportionate use of certain areas of the enclosure by the female chim- panzees in this study may reflect a behavior comparable to the use of core areas by wild female chimpanzees. The female chimpanzees in this study separated themselves from other group members through differential use of their island habitat. Spatial separation was achieved by two strategies: (1) use of different areas of the exhibit, and (2) temporal separation in overlapping areas.

Spatial Selection in Captive Chimpanzees 173
TABLE 3. Results of chi-square tests used to analyze differences between each female's overall use of the enclosure
Female Chi-square df Probability"
A X B 116.749 18 <0.0001 A X C 86.264 18 <0.0001 A X D 83.198 18 <0.0001 B X C 113.501 18 <0.0001 B X D 108.059 18 <0.0001 C x D 26.185 18 = 0.0956
aSignificant probability levels remain significant after sequential Bon- feronni adjustment for an overall a level set a priori at P = 0.05.
Percent time
50 r 30
20
10
0 1T
Female A
3T 1P Section of the island
Female B 0 Female C
*
5T
Female D
Fig. 3. habitat. *This subject used this location significantly more often than did other subjects.
Comparison of overall subject use of the four most frequently occupied areas of the island
The most common strategy was for the females to occupy different areas of the enclosure (five of six comparisons). However, one comparison, female C and D, showed a significant overlap in the use of the island. This overlap was partially offset by the females' tendency to occupy these areas at different times. Additionally, 5T, the area most often used simultaneously by these females, differed from other areas in the study. This tunnel was T shaped and three times larger than the other tunnels. It was the only tunnel in which females could be simultaneously occupying, yet be visually isolated from one another. Therefore, even when both females were in this tunnel, they were not necessarily in close proximity to one another. Data on associ- ation patterns of females C and D showed that they spent 8-9% of the observations in close proximity to one another [Bettinger, 19901. Overlap in the use of some areas cannot be attributed to lack of available space in the enclosure; there were several areas which were used infrequently, including a tunnel.

174 Bettinger et al.
TABLE 4. Percentage of observations during which another female simultaneously occupied the same area as the focal animal
Females
A B C D (%I (%) (%) (%)
Focal/ Area" N1T - 0 0 1 Bl3T 0 0 0
20 C/IP 9 2 D/5T 21 1 47
aArea is the most frequently used location by this female
- -
-
Three of the females most frequently occupied a tunnel. Tunnels provide pro- tection from the elements, but they also provide visual separation from other group members and the zoo public. Visual separation may be an important mechanism for reducing tension and limiting aggression between captive females. Reducing aggres- sion in captive groups is especially important, since the chimpanzees cannot leave the enclosure and may not be able to escape their aggressor adequately. Female aggres- sion in this study was recorded only 8 times in 100 hr of observation [Bettinger,
de Waal (1982) reported that aggressive incidents among the Arnhem Zoo chimpanzees were nearly twice as high in the winter when the chimpanzees were restricted to their inside quarters, which lack the complexity and size provided in the outdoor enclosure. Additionally, Nieuwenhuijsen and de Waal [ 19821 reported that by the end of the winter period, most of the chimpanzees had bald patches. Hair plucking and overgrooming may be indicative of boredom or it may be a result of tension caused by forced close proximity to others.
The female chimpanzees in this study consistently occupied their selected areas. Neither variation in temperature nor differences in time of day had an effect on the females' use of the habitat. Only female C showed a seasonal variation in use of the enclosure. A factor that may have contributed to female C's variation in enclosure use was the type of space she occupied. While the other females most often occupied a tunnel, female C most frequently used an elevated platform that provided no protec- tion from the elements. Regardless of season, female C frequently occupied the platform; however, during fall, summer and winter, she also used more protected areas.
This study shows that the female chimpanzees differentially used their island habitat by occupying different areas or using overlapping areas at different times. The overall consistency with which they occupied their selected space suggests that these females were actively regulating proximity to one another. Indirect evidence suggests that the subjects associated a particular area with the female who most often occupied it. For example, females with less tenure seldom used areas frequently occupied by others with longer tenure. This occurred despite no observation of a female taking overt action to exclude another from her area. It is possible that as a result of the long-term stable composition of this group, disputes over space were settled long ago.
The results of this study raise several additional questions. What factors influ- ence spatial use? Is the differential use of the island an extension of dominance
19911.

Spatial Selection in Captive Chimpanzees 175
interactions among females? Although the dominance hierarchy of these females had not been formally assessed at the time of this study, we suspect that it correlated with tenure. However, variables such as age of offspring, interactions among offspring, individual differences in sociality, and avoidance of, or attraction to, other group members may also affect spatial use. Answers to these questions are beyond the scope of this study. However, future studies could easily address these topics. Data from such studies can supply important information on exhibit use in addition to answering theoretical questions regarding female chimpanzee behavior.
CONCLUSIONS
1. Pairwise comparisons showed that the female chimpanzees used their island habitat differently from one another, with the exception of one comparison.
2. The females maintained spatial separation by using different areas of the enclosure or by separating themselves temporally in overlapping areas.
3. There was no diurnal or thermal variation in the females’ use of the enclo- sure; however, one female showed a seasonal variation.
4. The availability of areas in which chimpanzees can isolate themselves from other group members may be beneficial in reducing stress caused by close proximity to other chimpanzees.
ACKNOWLEDGMENTS
We thank the personnel of the Tulsa Zoo for allowing this research to be conducted on their chimpanzees and Tulsa Zoo Friends for providing funding for this project. We sincerely appreciate the cooperation and support of primate keepers A. Morris and P. Murphy. Dr. William Warde’s guidance with the statistical analysis is very much appreciated.
REFERENCES
Bettinger, T. Spatial selection, association patterns and behavior of captive chimpanzees (Pan trogl- odytes) at the Tulsa Zoological Park. Master’s thesis, Oklahoma State University, Stillwater, Oklahoma, 1990.
Bettinger, T. Behavior exhibited by captive adult chimpanzees. Pp. 27-36 in CHIMPANZOO PROCEEDINGS, April 1991. Oakland, Calif., Jane Goodall Institute, 1991.
Boesch, C.; Boesch, H. Hunting behavior of wild chimpanzees in the Tai National Park. AMERI- CAN JOURNAL OF PHYSICAL ANTHRO-
Clarke, A.; Juno, C.; Maple, T. Behavioral effects of a change in the physical environment: A pilot study of captive chimpanzees. ZOO BIOLOGY
Goodall, J. THE CHIMPANZEES OF GOMBE STREAM: PATTERNS OF BEHAVIOR. Cam- bridge, Belknap Press, 1986.
Halperin, S. Temporary association patterns in free
POLOGY 78547-573, 1989.
11371-380, 1982.
ranging chimpanzees: an assessment of individ- ual grouping preferences. Pp. 491-499 in THE GREAT APES. D. Hamburg, E. McCown, eds. Menlo Park, BenjamidCummings, 1979.
Hasegawa, T. Sex differences in ranging patterns. Pp. 99-1 14 in THE CHIMPANZEES OF THE MAHALE MOUNTAINS: SEXUAL AND LIFE HISTORY STRATEGIES. T. Nishida ed. Japan, University of Tokyo Press, 1990.
Jennrich, R.; Turner, F. Measurement of non-cir- cular home range. JOURNAL OF THEORETI-
Lehner, P. HANDBOOK OF ETHOLOGICAL METHODS. New York, Garland STPM Press, 1979.
Menzel, E. Chimpanzee utilization of space and responsiveness to objects. Pp. 72-80 in PRO-
TIONAL CONGRESS OF PRIMATOLOGY, Vol. 1. BEHAVIOR. C. Carpenter, ed. Basel, S. Karger, 1969.
CAL BIOLOGY 22:227-237, 1969.
CEEDINGS OF THE SECOND INTERNA-

176 Bettinger et al. Nieuwenhuijsen, K.; de Waal, F.B.M. Effects of
spatial crowding on social behavior in a chim- panzee colony. ZOO BIOLOGY 1:5-28, 1982.
Nishida, T. The social structure of the chimpan- zees of the Mahale Mountains. Pp. 73-122 in THE GREAT APES. D. Hamburg, E. McCown, eds. Menlo Park, BenjamidCummings, 1979.
Nishida, T. Social interactions between resident and immigrant female chimpanzees at Mahale. Pp. 68-89 in UNDERSTANDING CHIMPAN- ZEES. P. Heltne, L. Marquardt, eds. Cam- bridge, Harvard University Press, 1989.
Reynolds, V.; Reynolds, F. Chimpanzees of the Budongo Forest. Pp. 368-424 in PRIMATE BE- HAVIOR: FIELD STUDIES OF MONKEYS AND APES. I. DeVore, ed. New York, Holt, Rinehart & Winston, 1965.
Rice, W. Analyzing tables of statistical tests. EV-
Sugiyama, Y. Grooming interactions among adult chimpanzees at Bossou, Guinea, with special ref- erence to social structure. INTERNATIONAL JOURNAL OF PRIMATOLOGY 9:393-407, 1988.
Sugiyama, Y.; Koman, J. Social structure and dy- namics of wild chimpanzees at Bossou, Guinea.
OLUTION 43:223-225, 1989.
PRIMATES 20:323-339. 1979.
Traylor-Holzer, K.; Fritz, P. Utilization of space by adult and juvenile groups of captive chimpan- zees (Pan troglodytes). ZOO BIOLOGY 4:115- 127, 1985.
de Waal, F. CHIMPANZEE POLITICS: POWER AND SEX AMONG APES. New York, Harper & Row, 1982.
de Waal, F. The integration of dominance and so- cial bonding in primates. THE QUARTERLY REVIEW OF BIOLOGY 61:459-479, 1986.
Wrangham, R . Sex differences in chimpanzee dis- persion. Pp. 481-489 in THE GREAT APES. D. Hamburg, E. McCown, eds. Menlo Park, Ben- jamidCummings, 1979.
Wrangham, R.; Smuts, B. Sex differences in the hehavioural ecology of chimpanzees in the Go- mbe National Park, Tanzania. JOURNAL OF REPRODUCTIVE FERTILITY, SUPPLEMEN- TAL 28113-31, 1980.
Wrangham, R.; Clark, A,; Isabirye-Basuta, G. Fe- male social relationships and social organization of Kibale Forest Chimpanzees, Pp. 81-98 in TOPICS IN PRIMATOLOGY, Vol. I HUMAN ORIGINS. T. Nishida, W. McGrew, P. Marler, M. Pickford and F. de Waal, eds. Tokyo, Uni- versity of Tokyo Press, 1992.
Copyright © 2022 FDOKUMEN




















