
Cystic fibrosis: a review of pulmonary and nutritional therapies

This article is protected by copyright. All rights reserved. 1
Received Date: 26-Nov-2013 1
Accepted Date: 05-Jun-2014 2
Spatial distribution of microbial communities in the shallow submarine alkaline 3
hydrothermal field of the Prony Bay, New Caledonia1 4
5
Marianne Quéméneur1*
, Méline Bes1, Anne Postec
1, Nan Mei
1, Jérôme Hamelin
2, Christophe 6
Monnin3
,
Valérie Chavagnac3,
Claude Payri4, Bernard Pelletier
4, Linda Guentas-7
Dombrowsky4,5
, Martine Gérard6, Céline Pisapia
6, Emmanuelle Gérard
6, Bénédicte Ménez
6, 8
Bernard Ollivier1, Gaël Erauso
1. 9
10
1 Aix Marseille Université, CNRS, Université de Toulon, IRD, MIO UM 110, 13288, 11
Marseille, France 12
2 INRA, UR0050, Laboratoire de Biotechnologie de l’Environnement, 11100 Narbonne, 13
France 14
3 GET UMR5563 (CNRS/UPS/IRD/CNES), Observatoire Midi-Pyrénées, Université de 15
Toulouse, CNRS, IRD, 14 Avenue Edouard Belin, 31400 Toulouse, France 16
4 Institut pour la Recherche et le Développement (IRD) Centre de Nouméa, MIO UM 110, 17
promenade Laroque, 98848 Nouméa, Nouvelle Calédonie 18
5 Université de Toulon, Laboratoire des Matériaux Polymères Interfaces Environnement 19
Marin (MAPIEM), ISITV- Avenue Georges Pompidou BP56 - 83162 La Valette-du-Var 20
Cedex, France 21
6 Institut de Physique du Globe de Paris, Sorbonne Paris Cité, Université de Paris Diderot, 22
UMR CNRS 7154, 75005 Paris, France 23
24
* Corresponding author: Marianne Quéméneur 25
Address: Aix Marseille Université, CNRS/INSU, IRD, Mediterranean Institute of 26
Oceanography (MIO), UM 110, 13288 Marseille, France 27
E-mail: [email protected]; Phone: +0033(0)491828577; Fax: +0033(0)491828570 28
This article has been accepted for publication and undergone full peer review but has not been
through the copyediting, typesetting, pagination and proofreading process, which may lead to
differences between this version and the Version of Record. Please cite this article as doi:
10.1111/1758-2229.12184
Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 2
29
Running title: Microbial communities of Prony hydrothermal field 30
Keywords: Microbial diversity, serpentinization, hydrothermal system, alkaliphilic, Prony bay 31
32
Abstract 33
34
The shallow submarine hydrothermal field of the Prony Bay (New Caledonia) discharges 35
hydrogen- and methane-rich fluids with low salinity, temperature (<40°C) and high pH (11) 36
produced by the serpentinization reactions of the ultramafic basement into the lagoon 37
seawater. They are responsible for the formation of carbonate chimneys at the lagoon 38
seafloor. CE-SSCP fingerprinting, quantitative PCR and sequence analysis of 16S rRNA 39
genes revealed changes in microbial community structure, abundance and diversity depending 40
on the location, water depth and structure of the carbonate chimneys. The low archaeal 41
diversity was dominated by few uncultured Methanosarcinales similar to those found in other 42
serpentinization-driven submarine and subterrestrial ecosystems (e.g. Lost City, The Cedars). 43
The most abundant and diverse bacterial communities were mainly composed of Chloroflexi, 44
Deinococcus-Thermus, Firmicutes, and Proteobacteria. Functional gene analysis revealed 45
similar abundance and diversity of both Methanosarcinales methanoarchaea and 46
Desulfovibrionales- and Desulfobacterales sulfate-reducers in the studied sites. Molecular 47
studies suggest that redox reactions involving hydrogen, methane and sulfur compounds (e.g. 48
sulfate) are the energy driving forces of the microbial communities inhabiting the Prony 49
hydrothermal system. 50
51
Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 3
Introduction 52
53
A wide variety of submarine hydrothermal systems has been investigated worldwide since 54
their first discovery in the late 1970s (Corliss et al., 1979). Depending on the basement rock 55
hosting the hydrothermal system, different energy sources are available and thus 56
preferentially used as electron donors by the indigenous microorganisms. In ultramafic-hosted 57
oceanic hydrothermal systems, hydrogen (H2) and abiotic organic compounds dissolved in the 58
hydrothermal fluids are the dominant energy sources generated by serpentinization processes 59
(i.e. the metamorphic hydration of ultramafic rocks/minerals into serpentine) (Charlou et al., 60
2002; Kelley et al., 2005; Ludwig et al., 2006; Proskurowski et al., 2008). 61
The Lost City Hydrothermal Field (LCHF) (30°07′N, 42°07′W) is a well-known serpentinite-62
hosted submarine hydrothermal system (at water depth of ~800 meters below sea level, mbsl) 63
(Kelley et al., 2005). LCHF hosts carbonate chimneys up to 60 meters high above the 64
seafloor, and generate fluids with moderate temperatures (40-80°C at chimney vents, ca. 90-65
110°C in the subsurface), high pH (9-10.8), high dissolved H2 (1-15 mM), methane (1-2 mM), 66
and sulfate (5.9-12.9 mM) concentrations (Lang et al., 2010; Ludwig et al., 2006; 67
Proskurowski et al., 2008). Molecular surveys of the LCHF microbial diversity provided 68
evidence for a limited number of dominant phylogenetic prokaryotic lineages (Brazelton et 69
al., 2013; Kelley et al., 2005; Schrenk et al., 2004). The LCHF archaeal communities 70
consisted mainly of Lost City Methanosarcinales (LCMS), and ANaerobic MEthanotrophic 71
archaea (ANME-1 group), while bacterial communities included sulfide/methane-oxidizing 72
Gammaproteobacteria, H2/sulfide-oxidizing Epsilonproteobacteria, and sulfate-reducing and 73
H2-producing Firmicutes (Schrenck et al., 2004; Brazelton et al., 2006; Brazelton et al., 74
2013). In this respect, H2, CH4 and sulfur compounds are considered as the main components 75
sustaining the growth of LCHF microbial communities (Schrenk et al., 2013). 76 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 4
Shallow submarine vents support complex biological communities, characterized by the 77
coexistence of both chemosynthetic and photosynthetic organisms (Canet and Prol-Ledesma, 78
2007; Maugeri et al., 2009). To our knowledge, the Prony Hydrothermal Field (PHF) is the 79
only example of a shallow ultramafic-hosted submarine hydrothermal systems so far (at water 80
depth <50 mbsl). The PHF is located in the Prony bay on the New Caledonia ophiolitic nappe 81
(southwest Pacific Ocean) (Launay and Fontes, 1985; Quesnel et al., 2013). PHF comprises 82
large carbonate chimneys where H2- and CH4-rich and high pH (up to pH 11.3) fluids 83
discharge at low temperature (<40°C) into the lagoon (Launay and Fontes, 1985; Monnin et 84
al., submitted). In contrast to LCHF, but similarly to subterrestrial serpentinite-hosted 85
hydrothermal systems, such as those found in ophiolite massifs (e.g., Oman, Liguria) (Boulart 86
et al., 2013; Chavagnac et al., 2013; Monnin et al., 2011), the highly alkaline PHF fluids have 87
a low salinity inherited from their meteoric origin (Launay and Fontes, 1985; Monnin et al., 88
submitted), which may variably impact the PHF-associated microbial communities, 89
depending on the complexity of hydrothermal circulation from land to sea and thus the 90
location of the vents. Easy accessibility of PHF for sampling, as compared to deep-sea 91
hydrothermal systems (e.g. Lost City), makes PHF an ideal natural observatory to study 92
biogeochemical processes linked to serpentinization in submarine hydrothermal systems. 93
This study aimed to determine the abundance, the structure and the diversity of both archaeal 94
and bacterial communities using both functional- and 16S rRNA gene-based PCR methods in 95
several shallow and active carbonate chimney samples from four distinct sites located at 96
different water depths in the Prony bay. The microbial community of the shallow submarine 97
PHF and their potential in H2, CH4 or sulfur cycling was then compared with those of other 98
serpentinization-driven sites studied so far. 99
100 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 5
Results and discussion 101
102
Sampling sites in PHF and geochemistry 103
Samples of carbonate concretions and fluids were collected by scuba divers in four active 104
springs at different water depths in the Prony Bay (Fig. 1A): (i) at the Bain des Japonais 105
(BdJ), a spring in shallow water uncovered at low tide in the Carenage Bay, (ii) at the Needle 106
of Prony (ST07) at 16 mbsl in the North Bay, (iii) at Site ST08 at 47 mbsl in the Central Bay, 107
and (iv) at Site ST12 at 38 mbsl in the West Bay (Supplementary Experimental Procedures). 108
The PHF carbonate structures have different characteristics, depending on their size, age and 109
the associated level of discharged fluids (Fig. 1B): (i) at the BdJ, numerous fluid outlets build 110
a flat carbonate plateau on top of which grow small needle-like brittle structures with thin 111
walls (a few tens of cm high and a few centimeters in diameter) (Fig. S1), (ii) the concretion 112
sampled at ST07 (up to 30 cm in diameter) was part of a large-sized and competent 113
carbonated edifice (Fig. S2). The presence of encrusted marine shells inside the highly-114
consolidated chimney along with coral coating likely attest for a marine fingerprint in the 115
edification along with a long lasting existence and a reduced permeability, (iii) the ST08 116
hydrothermal vent (10 to 15 cm in diameter) was fairly active, with white tips at its top 117
demonstrating an active fluid discharge (Fig. S3), and (iv) ST12 massive chimney (~25 cm in 118
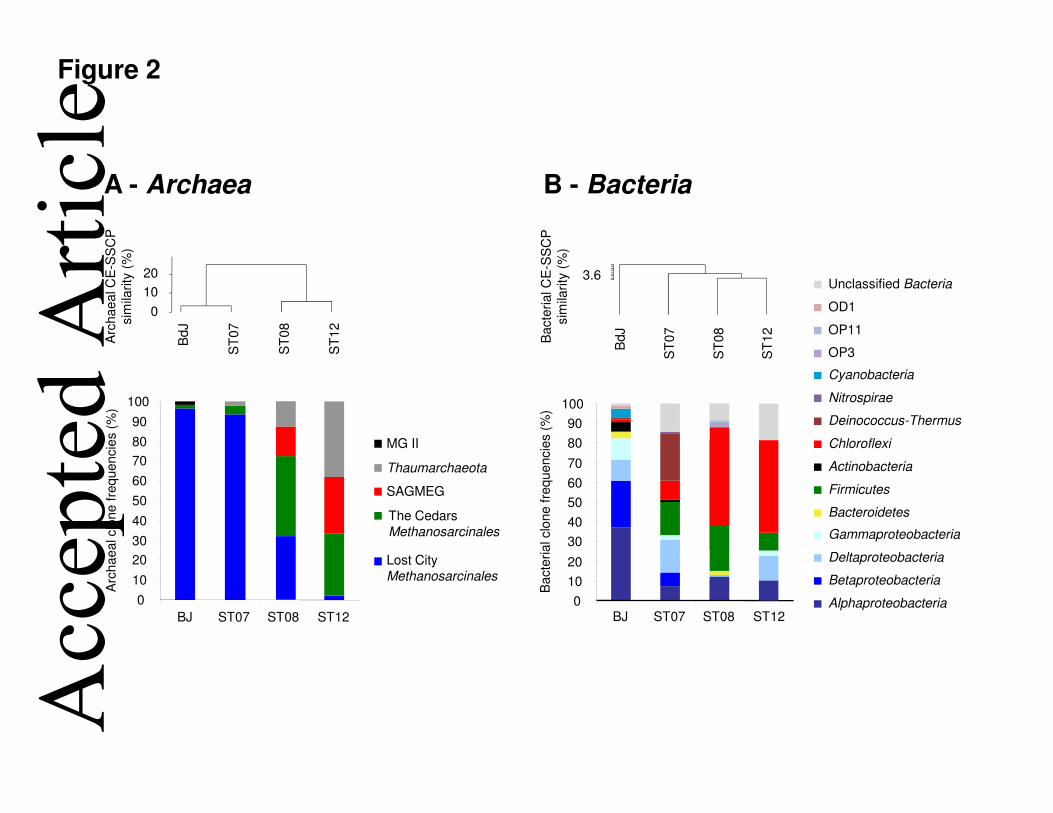
diameter) had several fluid diffusion centers and black spots of bitumen that correspond to 119
degraded organic matter (Fig. S4). Submarine chimney surfaces were colonized by various 120
organisms (e.g. ascidia, sponges, corals), which probably further contribute to reduce 121
permeability with the surrounding seawater of the site. PHF carbonate chimneys were mainly 122
composed of aragonite (CaCO3), brucite [Mg(OH)2], calcite (CaCO3), and magnesium 123
carbonates (MgCO3) (Pisapia et al., 2013), as observed at LCHF (Ludwig et al., 2006). 124 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 6
PHF fluids had high pH values ranging from 10.1 (ST07) to 11.1 (BdJ) (Table S1). The 125
highest concentrations of Cl- (163.9 mM), SO4
2- (7.5 mM), and major cations (e.g. 97.3 mM 126
Na+), measured in ST07, were 3 to 4 times lower than those of Prony ambient seawater. 127
Although these values are probably overestimated due to the likely contamination of the 128
hydrothermal fluid by the surrounding seawater, they indicated the low salinity level of PHF 129
fluids. The temperature of the Prony alkaline fluids ranged from 30°C to 37°C (BdJ). Gas 130
bubbles escaping at BdJ were composed of N2 (66.7%vol), CH4 (17.8%vol) and H2 131
(15.0%vol) (CH4/H2~1-2; Monnin et al., submitted) which is similar to those reported for 132
gases emitted at The Cedars, California, hyperalkaline springs (pH 11.5) (Morrill et al., 2013; 133
Suzuki et al., 2013). The CH4 concentration (8.0 mM) was 4 times higher than that of Lost 134
City fluids (1-2 mM), whereas the H2 concentration (6.4 mM) was two times lower than the 135
highest LCHF values (Schrenk et al., 2013). The variable CH4/H2 ratios were interpreted as 136
reflecting the different tectono-metamorphic evolution of the ultrabasic substratum (Boulart et 137
al., 2013). CO2 and O2 levels in gas bubbles were below the detection limit, as found in other 138
serpentinization-derived fluids, such as alkaline (pH 11.4) groundwater from Cabeço de Vide 139
Aquifer (CVA) in Portugal (Tiago and Verissimo, 2013; Morill et al., 2013). 140
141
Microbial community abundance and structure in PHF 142
The 16S rRNA gene was used as target gene for qPCR experiments to assess the archaeal and 143
bacterial abundance in the four studied chimney samples, while the dsrB and mcrA genes 144
were used to quantify the sulfate-reducers and methanoarchaea (methanogens and/or 145
anaerobic methanotrophs), respectively. The abundance of bacterial 16S rRNA genes was 146
more than one order of magnitude higher than that of archaeal 16S rRNA genes in all samples 147
(Table S2). According to the average 16S rRNA copy number in archaeal and bacterial 148
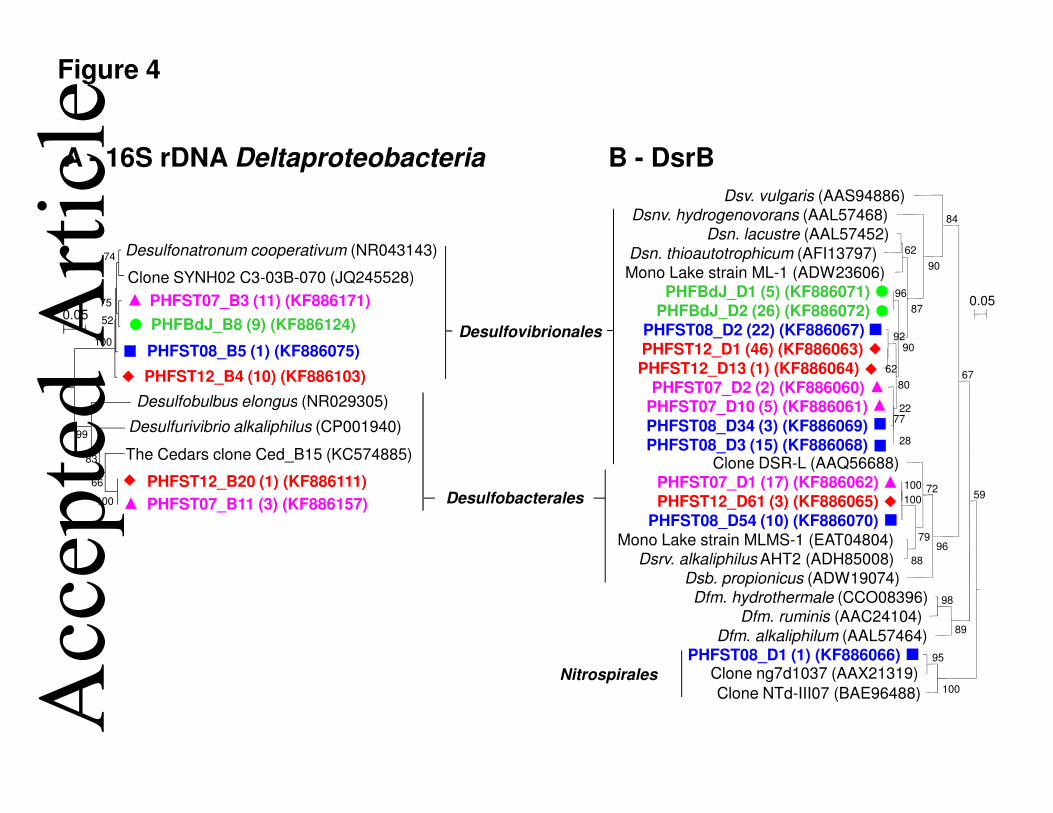
genomes (1.62 and 3.82, respectively) (Sun et al., 2013), Archaea accounted for 15.8% 149 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 7
(ST08) to 24.9% (ST07) of the total prokaryotic cells in PHF. Similar total cell numbers 150
(ranging from 0.02 to 8.4 x 108 cells per g of chimneys) and Archaea percentages were 151
obtained from LCHF samples using fluorescence in situ hybridization (Kelley et al., 2005). 152
Assuming that archaeal and bacterial cells contain, respectively, only one copy of mcrA and 153
dsrB genes, methanoarchaea accounted for 18.2% (BdJ) to 77.9% (ST12) of the archaeal 154
community, whereas the percentage of sulfate-reducers only represented from 2.2% (ST12) to 155
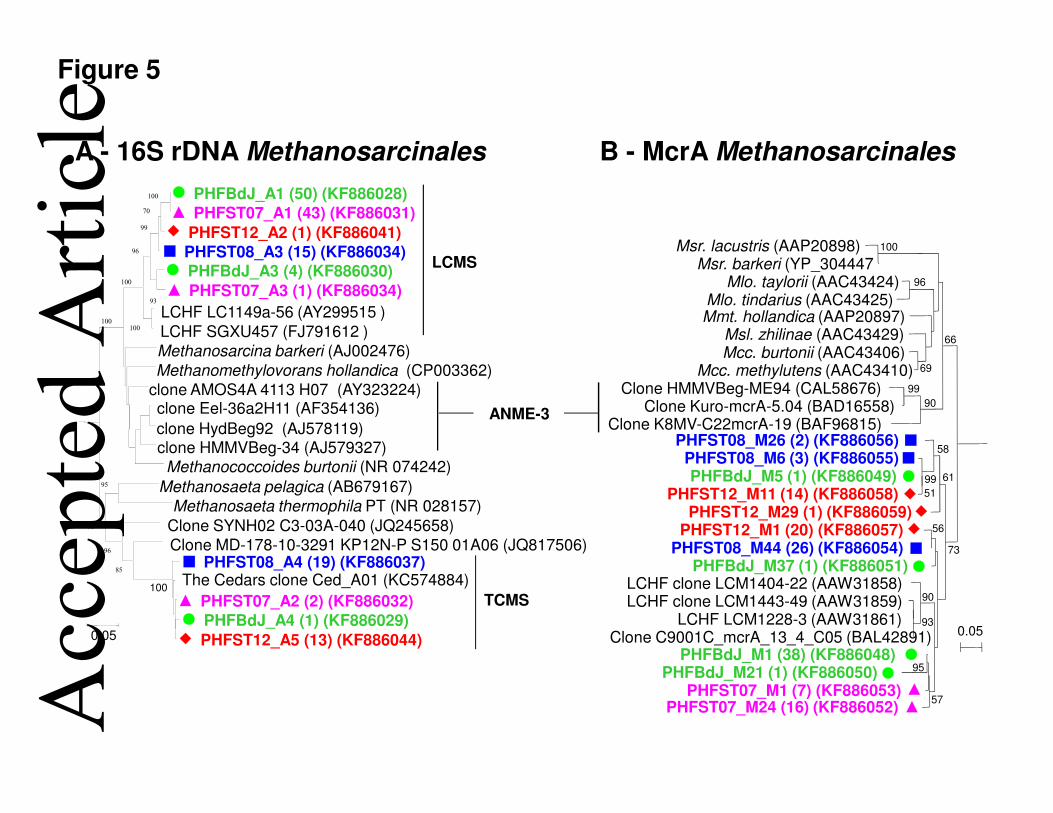
8.1% (ST07) of the bacterial community. 156
The CE-SSCP profiles of both bacterial and archaeal 16S rRNA genes allowed comparing the 157
PHF microbial community structures (Fig. S5), and were used to construct the dendrograms 158
shown in Fig. 2. The cluster analysis revealed two groups of archaeal CE-SSCP patterns: 159
BdJ/ST07 and ST08/ST12. Compared to BdJ and ST07, which shared a large dominant peak, 160
the two other encrusted chimneys (ST08 and ST12) collected at the greater depths showed a 161
more diverse archaeal community. The bacterial CE-SSCP profiles displayed two times the 162
number of peaks than the archaeal CE-SSCP profiles on average (Fig. S5). Cluster analysis 163
demonstrated a higher similarity among the three submarine samples ST07, ST08 and ST12, 164
with a slight difference for BdJ, the only site uncovered at low tide. 165
The dominant members of the microbial community of the four Prony chimney samples were 166
identified by analyzing clone libraries of bacterial and archaeal 16S rRNA gene sequences, as 167
well as, functional dsrB and mcrA sequences. Table S2 lists the number of clones, OTUs, 168
species richness, coverage and diversity indices calculated for each clone libraries. These 169
coverage values, supported by the Shannon index of diversity (H’) and the Chao1 estimator of 170
total species richness, demonstrated that PHF bacterial communities were more diverse than 171
archaeal communities, as already reported in other serpentinization-related ecosystems 172
(Brazelton et al., 2010; Brazelton et al., 2006; Suzuki et al., 2013; Tiago and Verissimo, 173
2013). Pyrosequencing 16S rRNA gene data obtained from ST07 sample was comparable 174 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 8
with that derived from clones (Fig. S6), and confirmed the low archaeal diversity and 175
abundance (20-25% of prokaryotic community). The extreme conditions and energy 176
limitations inside the porous carbonate chimneys may contribute to the establishment in it of 177
low and specific archaeal diversity (Brazelton et al., 2011; Valentine, 2007; Schrenk et al., 178
2004). Low diverse Archaea could live in direct contact with hyperalkaline and reduced fluids 179
and should be considered as primary producers using CH4 and possibly H2 in these alkaline 180
ecosystem. The more diverse bacteria could colonize different niches within the chimney both 181
in direct and indirect contacts with hydrothermal fluids. 182
183
Phylogenetic composition of microbial communities in different PHF sites 184
All sites combined, the PHF microbial communities consisted mainly of: Alpha- (16%), Beta- 185
(8%), Delta- (10%), Gammaproteobacteria (4%), Chloroflexi (28%), Firmicutes (12%) and 186
Deinococcus-Thermus (6%) within the Bacteria domain, and of Euryarchaeota (88%) and 187
Thaumarchaeota (12%) within the Archaea domain (Fig. 2). Both PHF archaeal and bacterial 188
community compositions changed depending on chimney sites and structure. 189
Sample BdJ and ST07 displayed the highest occurrence and diversity of Proteobacteria (Fig. 190
2). Alphaproteobacteria were mainly represented in BdJ by phototrophic anaerobic sulfur-191
oxidizing Rhodobacterales (Fig. 7). Betaproteobacteria, detected in BdJ (24%) and ST07 192
(7%), were related to H2-oxidizing chemolithoautotrophic Hydrogenophaga members 193
(Willems et al., 1989) (Fig. 3A). ST07 displayed the highest occurrence of 194
Deltaproteobacteria sequences (17%), which were divided in two groups in both 16S rRNA- 195
and dsrB-based phylogenetic trees (Fig. 4). The first and most abundant deltaproteobacterial 196
group was closely affiliated with the alkaliphilic Desulfonatronum cooperativum (97-98% 197
identity) (Zhilina et al., 2005) (Fig. 4). The second deltaproteobacterial group, only retrieved 198
in ST07 and ST12, was related to the alkaliphilic chemolithoautotrophic Desulfurivibrio 199 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 9
alkaliphilus (91% identity) (Sorokin et al., 2008). Both H2-oxidizing Desulfonatronum and 200
Desulfurivibrio genera, isolated from soda lakes, can use sulfate, thiosulfate, elemental sulfur 201
and/or polysulfide as terminal electron acceptors (Pikuta et al., 1998; Sorokin et al., 2008; 202
Zhilina et al., 2005). Deinococcus-Thermus sequences constituted 24% of the ST07 library 203
and closely clustered with the CVA sequences (Fig. S8A). Deinococci are known to be 204
chemoorganotrophic and resistant to extreme conditions (i.e., alkaline, moderately saline, 205
high temperature, ionizing radiations) (Albuquerque et al., 2005). The majority of BdJ and 206
ST07 archaeal sequences (94-96%) were closely affiliated with the uncultured Lost City 207
Methanosarcinales (LCMS) (Schrenk et al., 2004) (96-97% identity) (Fig. 5), as related to 208
ANME-3 and more distantly related to the Methanomethylovorans hollandica (91% identity), 209
which can use methylamines for methanogenesis (Lomans et al., 1999). 210
ST08 and ST12 samples were mainly represented by Chloroflexi sequences (>47%) belonging 211
to Dehalococcoidia class (87-96% identity) (Hug et al., 2012) (Fig. 3B), the function of which 212
is still unknown in the natural environment (Wasmund et al., 2013). The majority of 213
Firmicutes sequences detected in ST08 sample (23%) clustered with Clostridiales sequences 214
from CVA, The Cedars, and the hydrothermal alkaline groundwater of a deep South African 215
gold mine (pH 9.2; depth of 1290m) (96-97% identity) (Lin et al., 2006; Suzuki et al., 2013; 216
Tiago and Verissimo, 2013) (Fig. S9). PHFST08_B19 OTU was related to the alkaliphilic H2-217
oxidizing Dethiobacter alkaliphilus isolated from Mongolian soda lakes (Sorokin et al., 218
2008). PHFST12_B13 was affiliated with subsurface Clostridiales from LCHF carbonate 219
chimneys (91% identity) (Brazelton et al., 2006). Contrary to samples BdJ and ST07, the 220
ST08 and ST12 Euryarchaeota sequences were mainly (31-40%) related to The Cedars 221
Methanosarcinales sequences (98-99% identity) (Suzuki et al., 2013) (Fig. 5). South African 222
Gold Mine Euryarchaeotic Group (SAGMEG) sequences were exclusively found in samples 223
ST08 and ST12 (15% and 29%, respectively). The metabolic role of SAGMEG, in both 224 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 10
marine and terrestrial subsurface environments, still remains unknown. Thaumarchaeota 225
sequences were also exclusively found in samples ST08 (13%) and ST12 (38%) (Fig. S10), 226
and were affiliated with uncultured symbionts of marine sponges, which covered the surface 227
of these chimneys. PHFST08_A6 OTU clustered with Nitrosopumilus maritimus, the first 228
cultivated ammonia-oxidizing archaeon isolated from the rocky bottoms of a marine aquarium 229
(Konneke et al., 2005). 230
231
Comparison of PHF microbial community with other serpentinizing systems 232
PHF microbial diversity, as well as chemistry and mineralogy, are comparable to other 233
continental and submarine serpentinized sites (Schrenk et al., 2013; Suzuki et al., 2013). 234
Regarding Bacteria, the members of the H2-oxidizing Hydrogenophaga detected in BdJ and 235
ST07 chimneys were already found in many continental hyperalkaline springs (Brazelton et 236
al., 2013; Suzuki et al., 2013; Tiago and Verissimo, 2013). Such Hydrogenophaga sequences 237
were detected neither in the LCHF, nor in both encrusted deepest ST08 and ST12 PHF 238
chimneys. In the Tablelands ultrabasic springs (pH 10.5-12.6), these Hydrogenophaga 239
members were specifically associated with oxic-anoxic transition zones where H2-rich 240
subsurface fluids mix with oxygenated surface water (Brazelton et al., 2013). Both alpha- and 241
gammaproteobacterial sequences related to Methylosinus and Methylomicrobium genera were 242
detected at BdJ, suggesting potential aerobic methane oxidation in porous BdJ structures. In 243
contrast, Firmicutes and Chloroflexi were mainly recovered from submarine PHF chimneys, 244
which could be considered as more anoxic. Similar patterns have recently been observed in 245
The Cedars serpentinizing sites, where Chloroflexi and Firmicutes dominated the spring fed 246
solely by deep groundwater, while Beta- and Gammaproteobacteria dominated the shallow 247
groundwater-fed springs (Suzuki et al., 2013). These findings suggest the key role of 248
Chloroflexi members in anoxic zones of the PHF carbonate chimneys, while Hydrogenophaga 249 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 11
members seem to be important in oxic/anoxic transition zones. Both 16S rRNA- and dsrB 250
gene library analyses showed that the alkaliphilic H2-oxidizing and sulfate-reducing 251
Desulfonatronum members were common in PHF (Fig. 4), but they were retrieved neither 252
from LCHF, nor from the continental serpentinizing sites. Desulfurivibrio members were 253
detected only in the PHF submarine chimneys and in The Cedars (Suzuki et al., 2013), but not 254
in the LCHF. In contrast to Lost City (Brazelton and Baross, 2010), sulfur compounds could 255
be oxidized at PHF by gammaproteobacterial sulfur/sulfide-oxidizing 256
Thioalkalivibrio/Thiohalophilus genera (vs. Thiomicrospira genus at LCHF) (Fig. S8B). 257
Regarding Archaea, the most abundant Euryarchaeota phylotypes of both 16S rRNA gene 258
and mcrA clone libraries were associated with the Methanosarcinales (Fig. 5). In the PHF 259
system, Methanosarcinales similar to those found at The Cedars surprisingly dominated the 260
concretion samples collected at the greatest depths (40% at ST08), while the LCMS were 261
predominant in the shallower ones (96% at BdJ) (Fig. 2). Methanosarcinales members have 262
been isolated from diverse environments and are known to produce methane from different 263
substrates (e.g. H2, acetate, and methylated compounds) in a wide range of temperature and 264
pH conditions (Kendall and Boone, 2006). Moreover, some Methanosarcinales sequences had 265
those of LCMS, which are able to produce and oxidize methane (Brazelton et al., 2011), as its 266
closest phylogenetic relatives. Interestingly, the former co-occurred with those of sulfate-267
reducing bacteria belonging to Desulfobacterales (e.g. Desulfobulbus spp.) in some 268
submarine chimneys (e.g. ST08, ST12). Thus, a potential Anaerobic Methane Oxidation 269
(AMO) could take place in the inner parts of the PHF carbonate chimney in direct contact 270
with the H2 and CH4-rich fluids (anoxic zones), as previously demonstrated in LCHF 271
(Brazelton et al., 2011; Schrenk et al., 2004). Thaumarchaeota were detected only in 272
submarine PHF chimneys and have also been detected at LCHF, but it is unclear whether they 273 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 12
are autotrophic nitrifiers or representatives of a hydrothermal ecotype with a different 274
physiology (Lincoln et al., 2013). 275
276
Conclusion 277
PHF is an alkaline hydrothermal system hosted in an ultrabasic formation and is composed of 278
several springs (emerging at low tide or completely under water) discharging into the lagoon 279
seawater anoxic, H2- and CH4-rich, high pH fluids. Its shallow submarine location (at less 280
than 50 mbsl) and the low salinity of hydrothermal fluids originating from meteoric waters are 281
unique features for serpentinization-associated environments studied so far. Microbial 282
diversity and structure changed in the four PHF carbonate concretions collected at different 283
water depths in the Prony Bay. Factors that could affect PHF microbial diversity include age, 284
porosity, and colonization by various organisms of the chimney which may have impact on its 285
permeability and availability of nutrients. It is possible that distance to the recharge area and 286
presence of a fault close to the deepest sites with involvement in deep fluids circulation, might 287
also influence the PHF microbial structure. A large-scale study integrating numerous 288
chimneys and fluid samples from other geographically distant sites in the Prony Bay should 289
help in demonstrating the impact of various physico-chemical environmental factors (Eh, 290
organic carbon content, dissolved H2, CH4 and O2 levels, nutrient concentrations, age of the 291
edifice and permeability) on the existing microbial communities. In view of their abundance, 292
diversity and broad distribution in PHF carbonate chimneys, prokaryotes are likely to play a 293
key role in the carbon and sulphur budgets in this shallow submarine serpentinite-hosted 294
environment, similarly to what is observed in deep submarine hyperalkaline settings. 295
296
Acknowledgments 297 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 13
This project was financially supported by IRD and by the French national program EC2CO-298
Biohefect/Ecodyn/Dril/MicrobiEn (MicroProny) CNRS/INSU. We are grateful to the divers’ 299
team of IRD Nouméa (Eric Folcher, John Butcher, and Bertrand Bourgeois) for collecting 300
samples, and to Captain Jean-François Barazer and his crew on board the R/V Alis. 301
302
Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 14
References 303
304
Albuquerque, L., Simões, C., Nobre, M.F., Pino, N.M., Battista, J.R., Silva, M.T., Rainey, 305
F.A., and da Costa, M.S. (2005) Truepera radiovictrix gen. nov., sp. nov., a new radiation 306
resistant species and the proposal of Trueperaceae fam. nov. FEMS Microbiol Lett 247(2): 307
161-169. 308
Brazelton, W.J. and Baross, J.A. (2010) Metagenomic comparison of two Thiomicrospira 309
lineages inhabiting contrasting deep-sea hydrothermal environments. PloS one, 5(10): 310
e13530. 311
Brazelton, W.J., Ludwig, K.A., Sogin, M.L., Andreishcheva, E.N., Kelley, D.S., Shen, C.C., 312
Edwards, R.L., and Baross, J.A. (2010) Archaea and bacteria with surprising microdiversity 313
show shifts in dominance over 1,000-year time scales in hydrothermal chimneys. Proc Natl 314
Acad Sci 107(4): 1612-1617. 315
Brazelton, W.J., Mehta, M.P., Kelley, D.S., and Baross, J.A. (2011) Physiological 316
differentiation within a single-species biofilm fueled by serpentinization. MBio, 2(4). 317
Brazelton, W.J., Morrill, P.L., Szponar, N. and Schrenk, M.O. (2013) Bacterial communities 318
associated with subsurface geochemical processes in continental serpentinite springs. Appl 319
Environ Microbiol 79(13): 3906-3916. 320
Brazelton, W.J., Nelson, B., and Schrenk, M.O. (2012) Metagenomic evidence for H2 321
oxidation and H2 production by serpentinite-hosted subsurface microbial communities. Front 322
Microbiol, 2. 323
Brazelton, W.J., Schrenk, M.O., Kelley, D.S., and Baross, J.A. (2006) Methane-and sulfur-324
metabolizing microbial communities dominate the Lost City hydrothermal field ecosystem. 325
Appl Environ Microbiol 72(9): 6257-6270. 326 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 15
Boulart, C., Chavagnac, V., Monnin, C., Delacour, A., Ceuleneer, G., and Hoareau, G (2013) 327
Differences in gas venting from ultramafic-hosted warm spings: the example of oman and 328
voltri ophiolites. Ofioliti 38(2), 142-156. 329
Canet, C., and Prol-Ledesma, R.M. (2007) Mineralizing processes at shallow submarine 330
hydrothermal vents: Examples from Mexico. Geol Soc Am. Special papers 422: 359. 331
Charlou, J.L., Donval, J.P., Fouquet, Y., Jean-Baptiste, P., and Holm, N. (2002) Geochemistry 332
of high H2 and CH4 vent fluids issuing from ultramafic rocks at the Rainbow hydrothermal 333
field (36°14′N, MAR). Chem Geol 191(4): 345-359. 334
Chavagnac, V., Monnin, C., Ceuleneer, G., Boulart, C., and Hoareau, G. (2013) 335
Characterization of hyperalkaline fluids produced by low-temperature serpentinization of 336
mantle peridotites in the Oman and Ligurian ophiolites. Geochem Geophy Geosy 14(7): 337
2496-2522. 338
Corliss, J.B., Dymond, J., Gordon, L.I., Edmond, J.M., von Herzen, R.P., Ballard, R.D., 339
Green, K., Williams, D., et al. (1979) Submarine thermal springs on the Galapagos Rift. 340
Science, 203(4385): 1073-1083. 341
Hug, L., Beiko, R., Rowe, A., Richardson, R., and Edwards, E. (2012) Comparative 342
metagenomics of three Dehalococcoides-containing enrichment cultures: the role of the non-343
dechlorinating community. BMC genomics 13(1): 327. 344
Kelley, D.S., Karson, J.A., Früh-Green, G.L., Yoerger, D.R., Shank, T.M., Butterfield, D.A., 345
Hayes, J.M., Schrenk, M.O., et al. (2005) A serpentinite-hosted ecosystem: the Lost City 346
hydrothermal field. Science 307(5714): 1428-1434. 347
Kendall, M.M., and Boone, D.R. (2006) The order Methanosarcinales. The prokaryotes, 3: 348
244-256. 349
Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 16
Könneke, M, Bernhard, A.E., de la Torre, J.R., Walker, C.B., Waterbury, J.B., Stahl D.A. 350
(2005) Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 351
437(7058):543-6 352
Lang, S.Q., Butterfield, D.A., Schulte, M., Kelley, D.S., and Lilley, M.D. (2010) Elevated 353
concentrations of formate, acetate and dissolved organic carbon found at the Lost City 354
hydrothermal field. Geochim Cosmochim Acta 74(3): 941-952. 355
Launay, J. and Fontes, J.C. (1985) Les sources thermales de Prony (Nouvelle-Calédonie) et 356
leurs precipites chimiques. Exemple de formation de brucite primaire. Geologie de la France 357
1: 83-100. 358
Lin, L.-H., Hall, J., Onstott, T.C., Gihring, T., Lollar, B.S., Boice, E., Pratt, L., Lippmann-359
Pipke, J., and Bellamy, R.E.S. (2006) Planktonic microbial communities associated with 360
fracture-derived groundwater in a deep gold mine of South Africa. Geomicrobiol J 23(6): 361
475-497. 362
Lincoln, S.A., Bradley, A.S., Newman, S.A., and Summons, R.E. (2013) Archaeal and 363
bacterial glycerol dialkyl glycerol tetraether lipids in chimneys of the Lost City Hydrothermal 364
Field. Organic Geochem 60: 45-53. 365
Lomans, B.P., Maas, R., Luderer, R., Op den Camp, H.J., Pol, A., van der Drift, C., and 366
Vogels, G.D. (1999) Isolation and characterization of Methanomethylovorans hollandica gen. 367
nov., sp. nov., isolated from freshwater sediment, a methylotrophic methanogen able to grow 368
on dimethyl sulfide and methanethiol. Appl Environ Microbiol 65(8): 3641-3650. 369
Ludwig, K.A., Kelley, D.S., Butterfield, D.A., Nelson, B.K. and Fruh-Green, G. (2006) 370
Formation and evolution of carbonate chimneys at the Lost City Hydrothermal Field. 371
Geochim Cosmochim Acta 70(14): 3625-3645. 372
Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 17
Maugeri, T.L., Lentini, V., Gugliandolo, C., Italiano, F., Cousin, S., and Stackebrandt, E. 373
(2009) Bacterial and archaeal populations at two shallow hydrothermal vents off Panarea 374
Island (Eolian Islands, Italy). Extremophiles, 13(1): 199-212. 375
Maurizot, P. and Vendé-Leclerc, M. (2009) Carte géologique de la Nouvelle-Calédonie au 376
1/500 000. Direction de l’Industrie, des Mines et de l’Energie, Service de la Géologie de 377
Nouvelle-Calédonie, Bureau de Recherches Géologiques et Minières, Nouméa. 378
Monnin, C., Chavagnac, V., Boulart, C., Payri, C., Ménez, B., Gérard, M., Gérard, E., 379
Quéméneur, M., Erauso, G., Postec, A. , Dombrowski, L., and
Pelletier, B. The low 380
temperature hyperalkaline hydrothermal system of the Prony Bay (New Caledonia). 381
Biogeosciences. Submitted. 382
Monnin, C., Chavagnac, V., Ceuleneer, G., Boulart, C. and Hoareau, G. (2011) 383
Characterization of hyperalkaline fluids produced by serpentinization of mantle peridotites in 384
Oman and in Liguria (Northern Italy). Mineralog. Mag 75: 1490. 385
Morrill, P.L., Kuenen, J.G., Johnson, O.J., Suzuki, S., Rietze, A., Sessions, A.L., Fogel, M.L., 386
and Nealson, K.H. (2013) Geochemistry and geobiology of a present-day serpentinization site 387
in California: The Cedars. Geochim Cosmochim Acta 109:222-240 388
Pikuta, E.V., Zhilina, T.N., Zavarzin, G.A., Kostrikina, N.A., Osipov, G.A., and Rainey, F.A. 389
(1998) Desulfonatronum lacustre gen. nov., sp. nov.: a new alkaliphilic sulfate-reducing 390
bacterium utilizing ethanol. Microbiology, 67(1): 105-113. 391
Pisapia, C., Gerard, E., Gerard, M., and Menez, B., 2013. Organomineralization drives early 392
chimney edification at the hyperalkaline hydrothermal field of the Prony Bay (New 393
Caledonia). Mineralog Mag 77(5): 1974. 394
Proskurowski, G., Lilley, M.D., Seewald, J.S., Früh-Green, G.L., Olson, E.J., Lupton, J.E., 395
Sylva, S.P., and Kelley D.S. (2008) Abiogenic hydrocarbon production at Lost City 396
hydrothermal field. Science, 319(5863): 604-607. 397 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 18
Quesnel, B., Gautier, P., Boulvais, P., Cathelineau, M., Maurizot, P., Cluzel, D., Ulrich M., 398
Guillot, S., et al. (2013) Syn-tectonic, meteoric water-derived carbonation of the New 399
Caledonia peridotite nappe. Geology. doi: 10.1130/G34531.1 400
Schrenk, M.O., Brazelton, W.J., and Lang, S.Q. (2013) Serpentinization, carbon, and deep 401
life. Rev Mineral Geochem 75: 575-606. 402
Schrenk, M.O., Kelley, D.S., Bolton, S.A., and Baross, J.A. (2004) Low archaeal diversity 403
linked to subseafloor geochemical processes at the Lost City Hydrothermal Field, Mid-404
Atlantic Ridge. Environ Microbiol, 6(10): 1086-1095. 405
Sorokin, D.Y., Tourova, T.P., Mussmann, M., and Muyzer, G. (2008) Dethiobacter 406
alkaliphilus gen. nov. sp. nov., and Desulfurivibrio alkaliphilus gen. nov. sp. nov.: two novel 407
representatives of reductive sulfur cycle from soda lakes. Extremophiles, 12(3): 431-439. 408
Sun, D.-L., Jiang, X., Wu, Q.L., and Zhou, N.-Y. (2013) Intragenomic Heterogeneity of 16S 409
rRNA Genes Causes Overestimation of Prokaryotic Diversity. Appl Environ Microbiol 410
79(19): 5962-5969. 411
Suzuki, S., Ishii, S., Wu, A., Cheung, A., Tenney, A., Wanger, G., Kuenen, J.G., and Nealson, 412
K.H (2013) Microbial diversity in The Cedars, an ultrabasic, ultrareducing, and low salinity 413
serpentinizing ecosystem. P Natl Acad Sci 110(38): 15336-15341. 414
Tiago, I., and Verissimo, A. (2013) Microbial and functional diversity of a subterrestrial high 415
pH groundwater associated to serpentinization. Environ microbiol 15(6):1687-706. 416
Valentine, D. L. (2007) Adaptations to energy stress dictate the ecology and evolution of the 417
Archaea. Nat Rev Microbiol 5(4), 316-323. 418
Wasmund, K., Schreiber, L., Lloyd, K.G., Petersen, DG., Schramm, A., Stepanauskas, R., 419
Jørgensen, B.B., and Adrian, L (2013) Genome sequencing of a single cell of the widely 420
distributed marine subsurface Dehalococcoidia, phylum Chloroflexi. ISME J 8(2), 383-397. 421 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 19
Willems, A., Busse, J., Goor, M., Pot, B., Falsen, E., Jantzen, E., Hoste, B., Gillis, M., et al. 422
(1989) Hydrogenophaga, a new genus of hydrogen-oxidizing bacteria that includes 423
Hydrogenophaga flava comb. nov. (formerly Pseudomonas flava), Hydrogenophaga 424
palleronii (formerly Pseudomonas palleronii), Hydrogenophaga pseudoflava (formerly 425
Pseudomonas pseudoflava and "Pseudomonas carboxydoflava"), and Hydrogenophaga 426
taeniospiralis (formerly Pseudomonas taeniospiralis). Int J Syst Bacteriol 39(3): 319-333. 427
Zhilina, T.N., Zavarzina, D.G., Kuever, J., Lysenko, A.M., and Zavarzin, G.A. (2005) 428
Desulfonatronum cooperativum sp. nov., a novel hydrogenotrophic, alkaliphilic, sulfate-429
reducing bacterium, from a syntrophic culture growing on acetate. Int J Syst Evol Micr 55(3): 430
1001-1006. 431
432
Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 20
Figure legends 433
434
Figure 1. Sampling sites in the Prony bay. Simplified geological map of the Prony bay 435
(Maurizot and Vendé-Leclerc, 2009) displaying the location of the four sampling sites: Bain 436
des Japonais (BdJ), the Needle of Prony (ST07), ST08 and ST12 (A). Photographs of the 437
chimneys (B) collected at BdJ, ST07, ST08 and ST12. Chimney and fluid samples were 438
collected in 2011 by scuba divers, who appear in picture ST07. The top of chimneys 439
(approximately 30 cm high) were collected at ST07, ST08 and ST12 and the inner parts were 440
sampled for DNA extraction. BdJ chimney samples corresponded to the crushed small 441
chimney shown on picture BdJ. 442
443
Figure 2. Structures of archaeal (A) and bacterial (B) communities at the phylum and 444
subphylum level in the four chimney samples collected in the Prony bay. The clustering 445
analyses were based on euclidean distance and Ward's linkage of CE-SSCP profiles of 446
archaeal and bacterial 16S rRNA gene fragments retrieved from chimneys of the four sites: 447
BdJ, ST07, ST08 and ST12. The diagram of relative phylogenetic abundance was based on 448
frequencies of 16S rRNA gene sequences affiliated with major phylogenetic groups in the 449
clone libraries from the four chimneys of the Prony Bay. MGII and SAGMEG mean Marine 450
Group II and South Africa Gold Mine Euryarchaeotic Group, respectively. 451
452
Figure 3. Neighbor-joining phylogenetic tree of the Betaproteobacteria (A) and the 453
Chloroflexi (B) 16S rRNA gene sequences from the PHF chimneys. Each OTU is 454
represented by one sequence with ≥97% similarity grouping. Numbers between brackets 455
indicate the number of clones obtained for each sequence type. PHF sequences are marked in 456
bold and their origin is indicated by different font colors and by the following markers: BdJ 457 Acc
epte
d A
rticl
e

This article is protected by copyright. All rights reserved. 21
(, green font), ST07 (, pink font), ST08 (, blue font), and ST12 (, red font). Bootstrap 458
values <50% are not shown. 459
460
Figure 4. Neighbor-joining phylogenetic trees showing the relationship between 461
Deltaproteobacteria 16S rRNA gene sequences (A) and DsrB protein sequences from four 462
PHF chimneys. Each OTU is represented by one sequence with ≥97% similarity grouping. 463
Numbers between brackets indicate the number of clones obtained for each sequence type. 464
PHF sequences are marked in bold and their origin is indicated by different font colors and by 465
the following markers: BdJ (, green font), ST07 (, pink font), ST08 (, blue font), and 466
ST12 (, red font). Bootstrap values <50% are not shown. Genus name have been 467
abbreviated in the DsrB tree as follows: Dsv., Desulfovibrio; Dsnv., Desulfonatronovibrio; 468
Dsn., Desulfonatronum; Dsrv., Desulfurivibrio; Dsb., Desulfobulbus; Dfm., 469
Desufotomaculum. 470
471
Figure 5. Neighbor-joining phylogenetic trees showing the relationship between 472
Methanosarcinales 16S rRNA gene sequences (A) and McrA protein sequences from 473
four PHF chimneys. Each OTU is represented by one sequence with ≥97% similarity 474
grouping. Numbers between brackets indicate the number of clones obtained for each 475
sequence type. The PHF Methanosarcinales sequences are marked in bold and their origin is 476
indicated by different font colors and by following markers: BdJ (, green font), ST07 (, 477
pink font), ST08 (, blue font), and ST12 (, red font). Bootstrap values <50% are not 478
shown. Genus name have been abbreviated in the McrA tree as follows: Msr., 479
Methanosarcina; Mlo., Methanolobus; Mmt., Methanomethylovorans; Msl., Methanosalsum; 480
Mcc., Methanococcoides. Environmental (uncultured) Methanosarcinales clusters have been 481
abbreviated as follows: LCMS, Lost City Methanosarcinales; ANME-3, Anaerobic 482
Methanotrophs of group 3; TCMS, The Cedars Methanosarcinales. 483
Acc
epte
d A
rticl
e

Figure 1
ST12
BdJ
ST08
BdJ ST07
A B
Acc
epte
d A
rticl
e
100 100
Unclassified Bacteria
OD1
OP11
OP3
Cyanobacteria
Nitrospirae
BdJ
ST
07
ST
08
ST
12
0
10
20
Arc
haeal C
E-S
SC
P
sim
ilarity
(%
)
3.6
BdJ
ST
07
ST
08
ST
12
Bacte
rial C
E-S
SC
P
sim
ilarity
(%
)
A - Archaea B - Bacteria
Figure 2
0
10
20
30
40
50
60
70
80
90
100
BJ ST12
Arc
haeal clo
ne f
requencie
s (
%)
MG II
Thaumarchaeota
SAGMEG
The Cedars
Methanosarcinales
Lost City
Methanosarcinales
0
10
20
30
40
50
60
70
80
90
100
Bacte
rial clo
ne f
requencie
s (
%)
Deinococcus-Thermus
Chloroflexi
Actinobacteria
Firmicutes
Bacteroidetes
Gammaproteobacteria
Deltaproteobacteria
Betaproteobacteria
AlphaproteobacteriaST07 ST08 BJ ST12 ST07 ST08
Acc
epte
d A
rticl
e

Burkholderiales
100
94
93
75
75
73
61
57
100
91
0.05
PHFBdJ_B4 (2) (KF886116)Ralstonia pickettii (NR 102967)
Hydrogenophaga intermedia (FJ009392) Clone KSB18 (JX047095)
Hydrogenophaga atypica (AJ585992)PHFBdJ_B6 (18) (KF886128)
The Cedars clone Ced_B12 (KC574882)Malikia spinosa (AB681972)
The Cedars clone Ced_B11 (KC574880)CVA clone CVCloAm3Ph105 (AM778002)
PHFST07_B1 (6) (KF886164)
Clone LC1524B-50 (DQ270636)
A - Betaproteobacteria
Figure 3
PHFST08_B10 (2) (KF886088)PHFST12_B5 (3) (KF886100)
Abyssal marine sediment clone SPG12 461 471 B7 (FJ746255)Subseafloor sediment clone 1H3M40 (JN230084)
PHFST12_B10 (4) (KF886095)Dehalogenimonas sp. SBP1 (JQ994267)
Dehalococcoides ethenogenes 195 (NR 074116)Dehalococcoides sp. GT strain G (NR 074288)
Holocene sediments clone KM35B-155 (AB300104)Siliciclastic sediment clone CK 2C4 105 (EU488394)
PHFST08_B7 (8) (KF886076)PHFST12_B1 (17) (KF886112)
Mud volcano clone AMSMV-5-B75 (HQ588482)Hypersaline microbial mat clone SBZO 4493 (JN533673)
The Cedars clone Ced B01 (KC574890)PHFST07_B6 (7) (KF886162)PHFST08_B3 (33) (KF886074)PHFST12_B3 (16) (KF886101)
Dehalococcoidia
99
86
100
100
100
61
100
99
100
9971
67
10098
87
85
0.05
B - Chloroflexi
Acc
epte
d A
rticl
e
22
62
92
80
96
90
62
87
90
84
67
0.05
PHFST07_D10 (5) (KF886061)PHFST07_D2 (2) (KF886060)
PHFST12_D13 (1) (KF886064)
PHFST12_D1 (46) (KF886063)
PHFST08_D2 (22) (KF886067)
PHFBdJ_D2 (26) (KF886072)
PHFBdJ_D1 (5) (KF886071)
Mono Lake strain ML-1 (ADW23606)
Dsn. thioautotrophicum (AFI13797)
Dsn. lacustre (AAL57452)
Dsnv. hydrogenovorans (AAL57468)
Dsv. vulgaris (AAS94886)
Desulfovibrionales
PHFST12_B4 (10) (KF886103)
PHFST07_B3 (11) (KF886171)
PHFBdJ_B8 (9) (KF886124)
PHFST08_B5 (1) (KF886075)
Desulfonatronum cooperativum (NR043143)
Clone SYNH02 C3-03B-070 (JQ245528)
Desulfobulbus elongus (NR029305)
0.05
A - 16S rDNA Deltaproteobacteria B - DsrB
74
100
75
52
Figure 4
95
98
100
88
100
79
72
22
28
77
96
89
100
59
Clone NTd-III07 (BAE96488)
Clone ng7d1037 (AAX21319) PHFST08_D1 (1) (KF886066)
Dfm. alkaliphilum (AAL57464)
Dfm. ruminis (AAC24104)
Dfm. hydrothermale (CCO08396)
Dsb. propionicus (ADW19074)
Dsrv. alkaliphilus AHT2 (ADH85008)
Mono Lake strain MLMS-1 (EAT04804)
PHFST08_D54 (10) (KF886070)
PHFST12_D61 (3) (KF886065)
PHFST07_D1 (17) (KF886062)
Clone DSR-L (AAQ56688)PHFST08_D3 (15) (KF886068)
PHFST08_D34 (3) (KF886069)
PHFST07_D10 (5) (KF886061)
PHFST12_B20 (1) (KF886111)
PHFST07_B11 (3) (KF886157)
Desulfobulbus elongus (NR029305)
Desulfurivibrio alkaliphilus (CP001940)
The Cedars clone Ced_B15 (KC574885)
Desulfobacterales
Nitrospirales
99
83
66
100
Acc
epte
d A
rticl
e
PHFBdJ_A1 (50) (KF886028)
PHFST07_A1 (43) (KF886031)
PHFST12_A2 (1) (KF886041)
PHFST08_A3 (15) (KF886034)
PHFBdJ_A3 (4) (KF886030)
PHFST07_A3 (1) (KF886034)
LCHF LC1149a-56 (AY299515 )
LCHF SGXU457 (FJ791612 )
Methanosarcina barkeri (AJ002476)
Methanomethylovorans hollandica (CP003362)
clone AMOS4A 4113 H07 (AY323224)
100
70
99
93
100
96
100
100
clone Eel-36a2H11 (AF354136) Clone Kuro-mcrA-5.04 (BAD16558)Clone HMMVBeg-ME94 (CAL58676)
Mcc. methylutens (AAC43410)Mcc. burtonii (AAC43406)Msl. zhilinae (AAC43429)
Mmt. hollandica (AAP20897)Mlo. tindarius (AAC43425)
Mlo. taylorii (AAC43424)Msr. barkeri (YP_304447
Msr. lacustris (AAP20898)
99
69
96
100
90
66
LCMS
ANME-3
A - 16S rDNA Methanosarcinales B - McrA Methanosarcinales
Figure 5
clone HMMVBeg-34 (AJ579327) clone HydBeg92 (AJ578119)
Methanosaeta pelagica (AB679167)
Methanosaeta thermophila PT (NR 028157)
Clone SYNH02 C3-03A-040 (JQ245658)
Clone MD-178-10-3291 KP12N-P S150 01A06 (JQ817506) PHFST08_A4 (19) (KF886037)
The Cedars clone Ced_A01 (KC574884)
PHFST07_A2 (2) (KF886032)
PHFBdJ_A4 (1) (KF886029)
PHFST12_A5 (13) (KF886044)
100
85
96
95
70
clone Eel-36a2H11 (AF354136)
PHFST07_M24 (16) (KF886052)PHFST07_M1 (7) (KF886053)
PHFBdJ_M21 (1) (KF886050)PHFBdJ_M1 (38) (KF886048)
Clone C9001C_mcrA_13_4_C05 (BAL42891)LCHF LCM1228-3 (AAW31861)
LCHF clone LCM1443-49 (AAW31859)LCHF clone LCM1404-22 (AAW31858)
PHFBdJ_M37 (1) (KF886051) PHFST08_M44 (26) (KF886054)PHFST12_M1 (20) (KF886057)PHFST12_M29 (1) (KF886059)
PHFST12_M11 (14) (KF886058)PHFBdJ_M5 (1) (KF886049)
PHFST08_M6 (3) (KF886055) PHFST08_M26 (2) (KF886056)
Clone K8MV-C22mcrA-19 (BAF96815)Clone Kuro-mcrA-5.04 (BAD16558)
95
93
57
90
56
51
99
58
61
90
73
0.05
Methanococcoides burtonii (NR 074242)
ANME-3
0.05
TCMS
Acc
epte
d A
rticl
e
Copyright © 2022 FDOKUMEN




















