
Spatial distribution of ciguateric fish in the Republic of Kiribati
7
Spatial distribution of ciguateric fish in the Republic of Kiribati Wing Hei Chan a , Yim Ling Mak a , Jia Jun Wu a , Ling Jin a , Wai Hung Sit b , James Chung Wah Lam a , Sadovy de Mitcheson b , Leo Lai Chan a , Paul Kwan Sing Lam a , Margaret B. Murphy a,⇑ a State Key Laboratory in Marine Pollution, Department of Biology and Chemistry, Academic Building, City University of Hong Kong, Tat Chee Avenue, Kowloon, Hong Kong SAR b School of Biological Sciences, University of Hong Kong, Pokfulam Road, Hong Kong SAR article info Article history: Received 20 October 2010 Received in revised form 2 February 2011 Accepted 15 February 2011 Available online xxxx Keywords: Ciguatera Coral reef fish Moray eel Mouse neuroblastoma assay Spatial distribution Republic of Kiribati abstract Ciguatera is food poisoning caused by human consumption of reef fish contaminated with ciguatoxins (CTXs). The expanding international trade of tropical fish species from ciguatera-endemic regions has resulted in increased global incidence of ciguatera, and more than 50 000 people are estimated to suffer from ciguatera each year worldwide. The Republic of Kiribati is located in the Pacific Ocean; two of its islands, Marakei and Tarawa, have been suggested as high-risk areas for ciguatera. The toxicities of coral reef fish collected from these islands, including herbivorous, omnivorous and carnivorous fish (24% [n = 41], 8% [n = 13] and 68% [n = 117], respectively), were analyzed using the mouse neuroblastoma assay (MNA) after CTX extraction. The MNA results indicated that 156 fish specimens, or 91% of the fish samples, were ciguatoxic (CTX levels >0.01 ng g À1 ). Groupers and moray eels were generally more toxic by an order of magnitude than other fish species. All of the collected individuals of eight species (n = 3– 19) were toxic. Toxicity varied within species and among locations by up to 10 000-fold. Cephalapholis argus and Gymnothorax spp. collected from Tarawa Island were significantly less toxic than those from Marakei Island, although all individuals were toxic based on the 0.01 ng g À1 threshold. CTX concentra- tions in the livers of individuals of two moray eel species (Gymnothorax spp., n = 6) were nine times greater than those in muscle, and toxicity in liver and muscle showed a strong positive correlation with body weight. The present study provides quantitative information on the ciguatoxicity and distribution of toxicity in fish for use in fisheries management and public health. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Ciguatera is food poisoning caused by human consumption of a variety of reef fish containing natural marine toxins called cigua- toxins (CTXs). Ciguatera is endemic to tropical regions, but its inci- dence is increasing in other parts of the world because of the expanding international trade in tropical fish species (Dickey and Plakas, 2010) and the lack of practical and accurate routine testing methods for CTX surveillance. Outbreaks of ciguatera have been re- ported in tropical or sub-tropical regions around the world, includ- ing Hong Kong (Wong et al., 2008), Okinawa, Japan (Oshiro et al., 2010), French Polynesia (Chinain et al., 2010), Australia (Dickey and Plakas, 2010), and Texas and Southern California in the US (CDC, 2006). More than 50 000 people are estimated to suffer from ciguatera each year globally (Van Dolah, 2000). The average inci- dence rate in the endemic US territories is estimated to be 5–70 cases per 10 000 (CDC, 2009), while the highest global incidence rate, 464 cases per 10 000 people, was recorded in Raivavae Island in the South Pacific in 2002 (Caillaud et al., 2010). In 2008, about 4% of the fishes imported into the US originated from ciguatera- endemic regions (NMFS, 2009). Symptoms of ciguatera include abdominal pain, vomiting and diarrhea, as well as reversal of hot and cold sensations, numbness and tingling. Cardiovascular symptoms such as irregular pulse, decreased blood pressure, bradycardia and dizziness have also been reported (Lehane and Lewis, 2000; Dickey and Plakas, 2010; Roeder et al., 2010). Symptoms can be exacerbated with repeated exposure to CTXs (Tester et al., 2010). Death may also occur in ex- treme cases when the internal organs of ciguateric fish are con- sumed (Hamilton et al., 2010). The mechanism of action of CTX was first proposed by Rayner (1972), who stated that toxicity was related to voltage-sensitive sodium ion (Na + ) channels (VSSCs) in the membranes of nerve cells. CTX binding to site 5 of VSSCs results in an excessive influx of Na + , leading to an increase in cellular excitability and altering the recovery of resting membrane potential. This nerve hyperexcit- ability then causes gastrointestinal, neurological and cardiovascu- lar disorders; the symptoms can occurs as quickly as 4 h after ingestion and last for months or years (Lehane and Lewis, 2000; Empey Campora et al., 2008). Structurally, CTXs are heat-stable lipid-soluble cyclic polyether compounds containing 13–14 rings (Lehane and Lewis, 2000) and 0045-6535/$ - see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.chemosphere.2011.02.036 ⇑ Corresponding author. Tel.: +852 3442 6858; fax: +852 3442 0522. E-mail address: [email protected] (M.B. Murphy). Chemosphere xxx (2011) xxx–xxx Contents lists available at ScienceDirect Chemosphere journal homepage: www.elsevier.com/locate/chemosphere Please cite this article in press as: Chan, W.H., et al. Spatial distribution of ciguateric fish in the Republic of Kiribati. Chemosphere (2011), doi:10.1016/ j.chemosphere.2011.02.036
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Spatial distribution of ciguateric fish in the Republic of Kiribati

Chemosphere xxx (2011) xxx–xxx
Contents lists available at ScienceDirect
Chemosphere
journal homepage: www.elsevier .com/locate /chemosphere
Spatial distribution of ciguateric fish in the Republic of Kiribati
Wing Hei Chan a, Yim Ling Mak a, Jia Jun Wu a, Ling Jin a, Wai Hung Sit b, James Chung Wah Lam a,Sadovy de Mitcheson b, Leo Lai Chan a, Paul Kwan Sing Lam a, Margaret B. Murphy a,⇑a State Key Laboratory in Marine Pollution, Department of Biology and Chemistry, Academic Building, City University of Hong Kong, Tat Chee Avenue, Kowloon, Hong Kong SARb School of Biological Sciences, University of Hong Kong, Pokfulam Road, Hong Kong SAR
a r t i c l e i n f o
Article history:Received 20 October 2010Received in revised form 2 February 2011Accepted 15 February 2011Available online xxxx
Keywords:CiguateraCoral reef fishMoray eelMouse neuroblastoma assaySpatial distributionRepublic of Kiribati
0045-6535/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.chemosphere.2011.02.036
⇑ Corresponding author. Tel.: +852 3442 6858; fax:E-mail address: [email protected] (M.B. Mu
Please cite this article in press as: Chan, W.H.,j.chemosphere.2011.02.036
a b s t r a c t
Ciguatera is food poisoning caused by human consumption of reef fish contaminated with ciguatoxins(CTXs). The expanding international trade of tropical fish species from ciguatera-endemic regions hasresulted in increased global incidence of ciguatera, and more than 50 000 people are estimated to sufferfrom ciguatera each year worldwide. The Republic of Kiribati is located in the Pacific Ocean; two of itsislands, Marakei and Tarawa, have been suggested as high-risk areas for ciguatera. The toxicities of coralreef fish collected from these islands, including herbivorous, omnivorous and carnivorous fish (24%[n = 41], 8% [n = 13] and 68% [n = 117], respectively), were analyzed using the mouse neuroblastomaassay (MNA) after CTX extraction. The MNA results indicated that 156 fish specimens, or 91% of the fishsamples, were ciguatoxic (CTX levels >0.01 ng g�1). Groupers and moray eels were generally more toxicby an order of magnitude than other fish species. All of the collected individuals of eight species (n = 3–19) were toxic. Toxicity varied within species and among locations by up to 10 000-fold. Cephalapholisargus and Gymnothorax spp. collected from Tarawa Island were significantly less toxic than those fromMarakei Island, although all individuals were toxic based on the 0.01 ng g�1 threshold. CTX concentra-tions in the livers of individuals of two moray eel species (Gymnothorax spp., n = 6) were nine timesgreater than those in muscle, and toxicity in liver and muscle showed a strong positive correlation withbody weight. The present study provides quantitative information on the ciguatoxicity and distribution oftoxicity in fish for use in fisheries management and public health.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Ciguatera is food poisoning caused by human consumption of avariety of reef fish containing natural marine toxins called cigua-toxins (CTXs). Ciguatera is endemic to tropical regions, but its inci-dence is increasing in other parts of the world because of theexpanding international trade in tropical fish species (Dickey andPlakas, 2010) and the lack of practical and accurate routine testingmethods for CTX surveillance. Outbreaks of ciguatera have been re-ported in tropical or sub-tropical regions around the world, includ-ing Hong Kong (Wong et al., 2008), Okinawa, Japan (Oshiro et al.,2010), French Polynesia (Chinain et al., 2010), Australia (Dickeyand Plakas, 2010), and Texas and Southern California in the US(CDC, 2006). More than 50 000 people are estimated to suffer fromciguatera each year globally (Van Dolah, 2000). The average inci-dence rate in the endemic US territories is estimated to be 5–70cases per 10 000 (CDC, 2009), while the highest global incidencerate, 464 cases per 10 000 people, was recorded in Raivavae Islandin the South Pacific in 2002 (Caillaud et al., 2010). In 2008, about
ll rights reserved.
+852 3442 0522.rphy).
et al. Spatial distribution of cig
4% of the fishes imported into the US originated from ciguatera-endemic regions (NMFS, 2009).
Symptoms of ciguatera include abdominal pain, vomiting anddiarrhea, as well as reversal of hot and cold sensations, numbnessand tingling. Cardiovascular symptoms such as irregular pulse,decreased blood pressure, bradycardia and dizziness have alsobeen reported (Lehane and Lewis, 2000; Dickey and Plakas, 2010;Roeder et al., 2010). Symptoms can be exacerbated with repeatedexposure to CTXs (Tester et al., 2010). Death may also occur in ex-treme cases when the internal organs of ciguateric fish are con-sumed (Hamilton et al., 2010).
The mechanism of action of CTX was first proposed by Rayner(1972), who stated that toxicity was related to voltage-sensitivesodium ion (Na+) channels (VSSCs) in the membranes of nervecells. CTX binding to site 5 of VSSCs results in an excessive influxof Na+, leading to an increase in cellular excitability and alteringthe recovery of resting membrane potential. This nerve hyperexcit-ability then causes gastrointestinal, neurological and cardiovascu-lar disorders; the symptoms can occurs as quickly as 4 h afteringestion and last for months or years (Lehane and Lewis, 2000;Empey Campora et al., 2008).
Structurally, CTXs are heat-stable lipid-soluble cyclic polyethercompounds containing 13–14 rings (Lehane and Lewis, 2000) and
uateric fish in the Republic of Kiribati. Chemosphere (2011), doi:10.1016/
2 W.H. Chan et al. / Chemosphere xxx (2011) xxx–xxx
consisting of more than 20 congeners, of which 12 have been struc-turally characterized and classified into three groups according totheir geographical distribution: the Caribbean CTXs (C-CTX), PacificCTXs (P-CTX) and Indian Ocean CTXs (I-CTX) (Hamilton et al.,2002). The most potent congener, P-CTX-1, is approximately 16times more toxic than the least potent P-CTX-4B (Lehane and Le-wis, 2000). The safe level of exposure to P-CTX-1 equivalents in fishhas been proposed as 0.01 ng g�1 of fish flesh based on results fromthe mouse bioassay toxicity test and including an uncertainty fac-tor of 10 (Lehane and Lewis, 2000). Studies of fish remnantsresponsible for ciguatera in human patients suggested that toxicityhigher than 0.31 ng g�1 P-CTX-3C equivalents is likely to causesymptoms of intoxication (Chinain et al., 2010).
More than 400 fish species, including groupers, snappers, jacks,mackerels, triggerfish and surgeonfish, are known to be associatedwith a high risk of ciguatera (Tester et al., 2010). They are generallydistributed between 35�N and 35�S, and within this region theiroccurrence is discrete, heterogeneous and site-specific. The distri-bution of ciguatoxic fishes has been found to be consistent withthat of toxic dinoflagellate communities (Dickey and Plakas,2010) because CTXs are metabolic products of toxins producedby epiphytic, benthic dinoflagellates in the genus Gambierdiscus,especially Gambierdiscus toxicus (Roeder et al., 2010) that are trans-formed and accumulated along the marine food chain. Herbivorousfishes accumulate and concentrate precursor P-CTX-4A/B or gam-biertoxins (GTXs) when grazing on Gambierdiscus-associated sea-
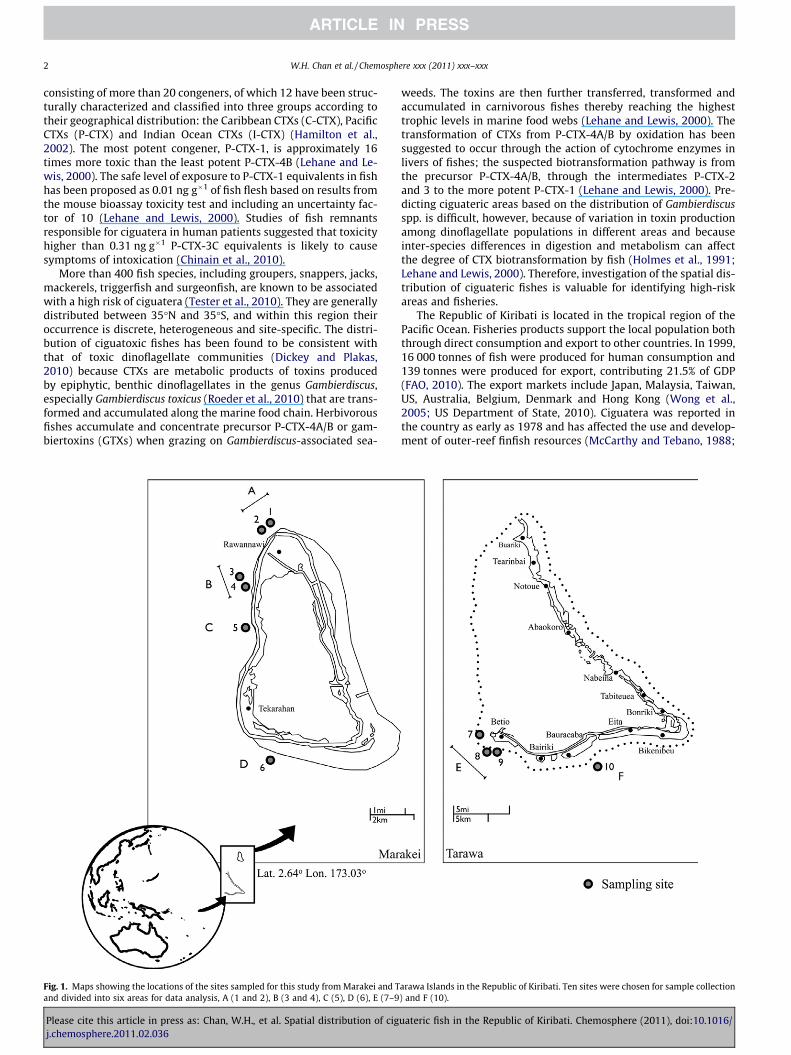
Fig. 1. Maps showing the locations of the sites sampled for this study from Marakei and Tand divided into six areas for data analysis, A (1 and 2), B (3 and 4), C (5), D (6), E (7–9
Please cite this article in press as: Chan, W.H., et al. Spatial distribution of cigj.chemosphere.2011.02.036
weeds. The toxins are then further transferred, transformed andaccumulated in carnivorous fishes thereby reaching the highesttrophic levels in marine food webs (Lehane and Lewis, 2000). Thetransformation of CTXs from P-CTX-4A/B by oxidation has beensuggested to occur through the action of cytochrome enzymes inlivers of fishes; the suspected biotransformation pathway is fromthe precursor P-CTX-4A/B, through the intermediates P-CTX-2and 3 to the more potent P-CTX-1 (Lehane and Lewis, 2000). Pre-dicting ciguateric areas based on the distribution of Gambierdiscusspp. is difficult, however, because of variation in toxin productionamong dinoflagellate populations in different areas and becauseinter-species differences in digestion and metabolism can affectthe degree of CTX biotransformation by fish (Holmes et al., 1991;Lehane and Lewis, 2000). Therefore, investigation of the spatial dis-tribution of ciguateric fishes is valuable for identifying high-riskareas and fisheries.
The Republic of Kiribati is located in the tropical region of thePacific Ocean. Fisheries products support the local population boththrough direct consumption and export to other countries. In 1999,16 000 tonnes of fish were produced for human consumption and139 tonnes were produced for export, contributing 21.5% of GDP(FAO, 2010). The export markets include Japan, Malaysia, Taiwan,US, Australia, Belgium, Denmark and Hong Kong (Wong et al.,2005; US Department of State, 2010). Ciguatera was reported inthe country as early as 1978 and has affected the use and develop-ment of outer-reef finfish resources (McCarthy and Tebano, 1988;
arawa Islands in the Republic of Kiribati. Ten sites were chosen for sample collection) and F (10).
uateric fish in the Republic of Kiribati. Chemosphere (2011), doi:10.1016/
W.H. Chan et al. / Chemosphere xxx (2011) xxx–xxx 3
cited by Beets (2001)). A survey of local fishermen conducted in1983 reported that ciguatera cases and toxic fish species wereassociated with two islands, Marakei and Tarawa, both of whichhave recently been reported as having a high risk of ciguatera(Tikai, 1988; Lewis, 2006). Quantitative information on the toxicityand distribution of ciguatoxic fishes in these islands, however, isstill limited. The present study therefore aims to investigate theciguatoxicity and distribution of toxicity in fish collected from thisarea to provide further information for fisheries management andpublic health.
In this study, the toxicities of wild-caught coral reef fishes andthe spatial distribution of ciguatoxic fishes from the Republic ofKiribati were studied using mammalian cell-based viability assays,which have been widely used for the identification of marine tox-ins. The mouse neuroblastoma assay (MNA) is one of the most sta-ble methods used for studying VSSC-dependent neurotoxinsincluding CTXs (Cañete and Diogène, 2008). Coupling mouse neu-ronal carcinoma Neuro-2a cells and the MTT [3-(4,5-dimethylthia-zol-2-yl)-2,5-diphenyltetrazolium] cell proliferation assay with thedrugs veratridine, a sodium channel activator, and ouabian, whichcan maximize the effect of veratridine by inhibiting the Na+/K+
pump, allows the assay to be more specific to CTXs with sensitivityas high as the pg g�1 level (Ledreux et al., 2009).
2. Materials and methods
2.1. Fish samples
All fish samples were collected from the Republic of Kiribati,which is composed of 32 islands. Some areas of the Republic ofKiribati have been described as high-risk areas for ciguatoxicfishes, such as the southern reef of Tarawa and the western reef
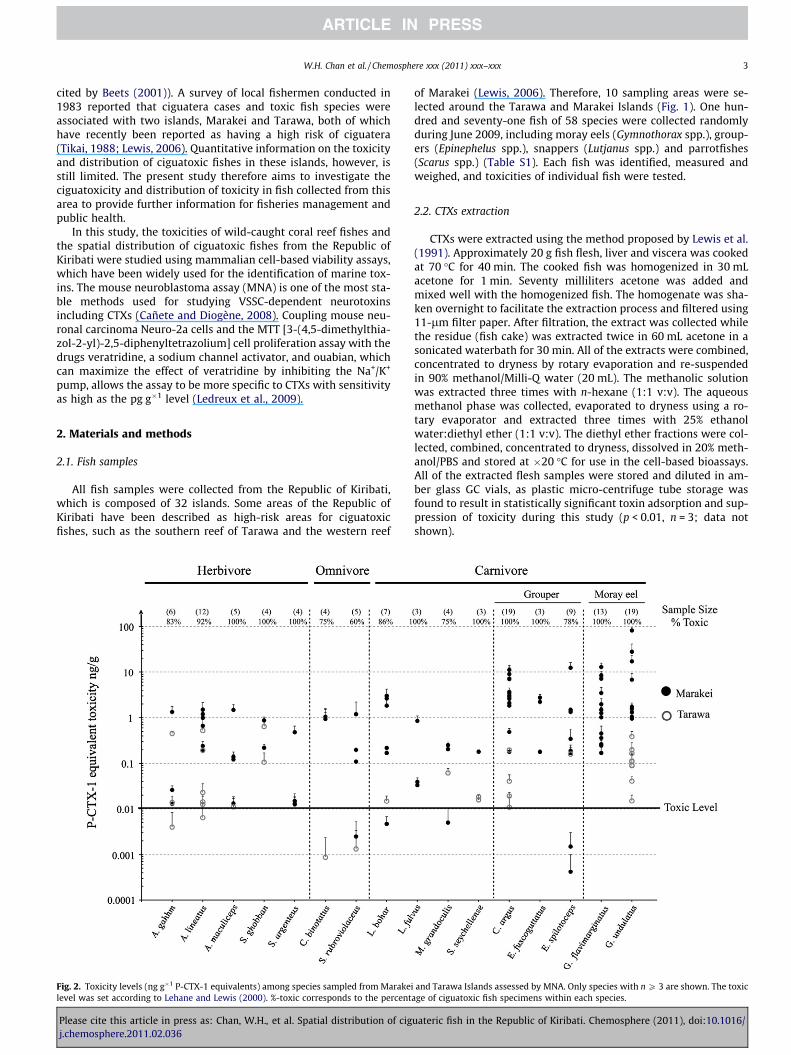
Fig. 2. Toxicity levels (ng g�1 P-CTX-1 equivalents) among species sampled from Marakeilevel was set according to Lehane and Lewis (2000). %-toxic corresponds to the percent
Please cite this article in press as: Chan, W.H., et al. Spatial distribution of cigj.chemosphere.2011.02.036
of Marakei (Lewis, 2006). Therefore, 10 sampling areas were se-lected around the Tarawa and Marakei Islands (Fig. 1). One hun-dred and seventy-one fish of 58 species were collected randomlyduring June 2009, including moray eels (Gymnothorax spp.), group-ers (Epinephelus spp.), snappers (Lutjanus spp.) and parrotfishes(Scarus spp.) (Table S1). Each fish was identified, measured andweighed, and toxicities of individual fish were tested.
2.2. CTXs extraction
CTXs were extracted using the method proposed by Lewis et al.(1991). Approximately 20 g fish flesh, liver and viscera was cookedat 70 �C for 40 min. The cooked fish was homogenized in 30 mLacetone for 1 min. Seventy milliliters acetone was added andmixed well with the homogenized fish. The homogenate was sha-ken overnight to facilitate the extraction process and filtered using11-lm filter paper. After filtration, the extract was collected whilethe residue (fish cake) was extracted twice in 60 mL acetone in asonicated waterbath for 30 min. All of the extracts were combined,concentrated to dryness by rotary evaporation and re-suspendedin 90% methanol/Milli-Q water (20 mL). The methanolic solutionwas extracted three times with n-hexane (1:1 v:v). The aqueousmethanol phase was collected, evaporated to dryness using a ro-tary evaporator and extracted three times with 25% ethanolwater:diethyl ether (1:1 v:v). The diethyl ether fractions were col-lected, combined, concentrated to dryness, dissolved in 20% meth-anol/PBS and stored at �20 �C for use in the cell-based bioassays.All of the extracted flesh samples were stored and diluted in am-ber glass GC vials, as plastic micro-centrifuge tube storage wasfound to result in statistically significant toxin adsorption and sup-pression of toxicity during this study (p < 0.01, n = 3; data notshown).
and Tarawa Islands assessed by MNA. Only species with n P 3 are shown. The toxicage of ciguatoxic fish specimens within each species.
uateric fish in the Republic of Kiribati. Chemosphere (2011), doi:10.1016/

4 W.H. Chan et al. / Chemosphere xxx (2011) xxx–xxx
2.3. Neuro-2a cell culture
Neuro-2a neuroblastoma cells (ATCC, CCL131; ATCC, Manassas,VA) were cultured in Roswell Park Memorial Institute (RPMI)-1640medium (Gibco, Life Technologies, Carlsbad, CA) supplementedwith 10% heat inactivated fetal bovine serum (BD Biosciences,San Jose, CA), 2 g L�1 Na2CO3 and antibiotic solution (50 unitsmL�1
penicillin, 50 lg mL�1 streptomycin and 2.5 lg mL�1 Fungizone�
(Gibco Life Technologies, Carlsbad, CA)) at 37 �C in 5% CO2. Cellsubcultures were conducted three times per week (dilution 1:3)in 75 cm2 tissue culture flasks (Nunc, Rockilde, Denmark).
2.4. Cell viability assays
Neuro-2a cells were seeded into 96-well flat-bottom plates at aconcentration of 2.5 � 105 cells mL�1 in 200 lL of medium per well.To avoid edge effects, peripheral wells received 300 lL PBS. After24 h, the medium was renewed with complete RPMI 1640 contain-ing 0.1 mM ouabain and 0.01 mM veratridine. Cells were dosed with10 lL well�1 P-CTX-1 standard (Dr. R. Lewis, University of Queens-land) at seven concentrations ranging from 1.9 pg mL�1 to125 pg mL�1 in five replicates; sample extracts were tested intriplicate. All wells received the same final volume of 200 lL. Cellviability was assessed by adding 50 lL, 0.5 mg mL�1 MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (Sigma–Aldrich, St.Louis, MO). After 90 min incubation at 37 �C, 50 lL isopropanol with0.1 N HCl was added and mixed well. Absorbance was measuredusing a microplate reader (Molecular Devices Spectra Max M2) at595 nm with a reference wavelength of 655 nm. The optical densityacquired for each well was then normalized by the MTT blank.
2.5. Data analysis
P-CTX-1 standard curves were plotted using non-linear regres-sion against the log of the toxin concentration. Toxicity values for
Fig. 3. Spatial distribution and toxicities of ciguatoxic C. argus and Gymnothorax spp. fromhaving a toxicity of 17.17 ng g�1 was also collected from Marakei Island, but the exact s
Please cite this article in press as: Chan, W.H., et al. Spatial distribution of cigj.chemosphere.2011.02.036
all fish samples were determined from the standard curves and themethod limit of quantification (LOQ) was 1.6 pg g�1. The assayswere conducted twice and the toxicity results are reported as meanP-CTX-1 equivalents ± SEM of all replicates between the twoassays. The inter-plate relative standard deviation was 18.4%, andinter-assay relative standard deviation was 24.7%. Twelve Cepha-lapholis argus muscle samples were also analyzed using a novelinstrumental method (manuscript in preparation), and the resultsfrom the MNA were found to overestimate P-CTX-1 concentrationsby 1.5–2.7-fold; this overestimation is due to the fact that the MNAcan also detect the activity of brevetoxins and other CTX analogues.Statistical analysis was conducted using Student’s t-tests if datapassed normality and equal variance tests; otherwise nonparamet-ric Mann–Whitney U tests were used (SigmaStat 3.5, Aspire Soft-ware International, Washburn, VA). Linear regression was used toanalyse relationships between body weight and toxin concentra-tion. Statistical significance was accepted at p < 0.05.
3. Results and discussion
3.1. Fish flesh toxicity
The collected fish species were categorized according to theirfeeding habits into three groups: herbivore, omnivore and carni-vore (Table S1). Herbivores and omnivores represented 24%(n = 41) and 8% (n = 13) of the specimens, respectively, and theremaining 68% (n = 117) samples were carnivorous fishes. Withinthe carnivorous fishes, 37% (n = 43) and 28% (n = 33) were groupersand moray eels, respectively.
Based on the MNA results of flesh extract, 111 fish specimensaccounting for 65% of the fish samples collected from the two is-lands in the Republic of Kiribati contained ciguatoxicityP0.1 ng g�1 P-CTX-1 equivalents, the minimum ciguatoxicity esti-mated in muscle of fish involved in a ciguatera outbreak in thePacific (Lehane and Lewis, 2000). To address uncertainties due to
Marakei and Tarawa Islands in the Republic of Kiribati. One additional G. undulatusampling location is unknown, and therefore it is not shown on the figure.
uateric fish in the Republic of Kiribati. Chemosphere (2011), doi:10.1016/

W.H. Chan et al. / Chemosphere xxx (2011) xxx–xxx 5
the quantity of fish consumed, individual variation among fishes,inter-species variation and use of the mouse bioassay, a safety fac-tor of 10 was applied (Lehane and Lewis, 2000) and fish containing60.01 ng g�1 P-CTX-1 are considered to be safe for human con-sumption. According to this more conservative value, 156 fishspecimens (91%) were ciguatoxic. Only 10%, 23% and 12.4% of her-bivorous, omnivorous and carnivorous fishes, respectively, col-lected from the two islands were safe for human consumption.Due to the relatively small sample size of the omnivores, the per-centage estimated of safe for consumption may be overestimated.Among the carnivorous fishes, most of the groupers (91%) and all ofthe moray eels were ciguatoxic.
The P-CTX-1 equivalent toxicities of each species having a sam-ple size larger than or equal to three are shown in Fig. 2. The threemost toxic fishes were all moray eels (G. undulatus); their P-CTX-1equivalent toxicities were 81.84, 27.88 and 17.17 ng g�1, respec-tively. All of the collected individuals of Acanthurus maculiceps(n = 5), Scarus ghobban (n = 4), Siganus argenteus (n = 4), Lutjanusfulvus (n = 3), Sargocentron seychellense (n = 3), Epinephelus fusco-guttatus (n = 3) and two moray eel species Gymothorax flavimargin-atus (n = 13) and G. undulatus (n = 19) were found to be toxic; fiveof these species are carnivorous. It is noteworthy that toxicity var-ied within species among locations by up to 10 000-fold, reflectingthe importance of understanding site-specific factors that deter-mine whether a fish becomes ciguatoxic.
Groupers and moray eels collected from Marakei and Tarawa Is-lands generally were more toxic by an order of magnitude than theother fish species sampled. The average toxicities of groupers andmoray eels were 2.02 ng g�1 P-CTX-1 equivalents (range:4.0 pg g�1–12.43 ng g�1 P-CTX-1 equivalents) and 5.65 ng g�1 P-CTX-1 equivalents (range: 0.02–81.84 ng g�1 P-CTX-1 equivalents),respectively. Herbivorous, omnivorous and other carnivorous fishcontained relatively similar average toxicities of 0.36 ng g�1,0.69 ng g�1 and 0.39 ng g�1 P-CTX-1 equivalents, respectively.These results support those reported in previous reviews suggest-ing that ciguatera was most often associated with ingestion ofgroupers and that moray eels are generally the most ciguatoxicfishes (Lehane and Lewis, 2000). This pattern is likely due to the
Fig. 4. P-CTX-1 equivalent toxicities of muscle and liver tissues from moray eels (Gymnotmeasured in liver tissue and muscle tissue are shown above each column. The insetGymnothorax spp. liver and muscle.
Please cite this article in press as: Chan, W.H., et al. Spatial distribution of cigj.chemosphere.2011.02.036
high trophic level occupied by these two fishes; however, a studyin the Red Sea found that groupers and eels engaged in cooperativehunting behavior (Bshary et al., 2006), and therefore it is possiblethat consumption of a common food source contributes to the tox-icity observed in these species. Herbivorous fish are often found tobe toxic in the South Pacific, but they are not implicated in cigua-tera in other parts of the world such as the Caribbean Sea (Pottieret al., 2001). The groupers C. argus and Epinephelus spilotoceps werefound to contain P-CTX-1 equivalents of up to 10 ng g�1, a levelwhich is 1000-fold higher than the safety level. Consumption ofthese species fished from this area in the Republic of Kiribatishould therefore be avoided. Moray eels were generally an orderof magnitude more toxic than groupers, while groupers were twoorders of magnitude more toxic than the other species collected.
CTXs bioaccumulate in tropical and sub-tropical fishes (Litakeret al., 2010), and the toxicity of individual fish depends upon thebioaccumulation time and the amount of toxin present in foodsources (Llewellyn, 2010). Highest-trophic-level consumers withinthe food web are therefore expected to accumulate the highest CTXlevels, though the distribution of CTXs at lower trophic levels is notwell understood, and the relationship between body size andciguatoxin content is not clear for many species. Moray eels occupythe highest trophic level within the area sampled in the presentstudy, prey on other carnivorous fishes as well as scavenging theremains of larger fish, and are able to grow to more than 10 kg inbody weight. Thus moray eels are considered to be high-risk spe-cies for ciguatera.
3.2. Ciguateric fish distribution
Fish samples collected from Tarawa Island were significantlyless toxic than those from Marakei (p < 0.01, Mann–Whitney Utest; Fig. 2). The P-CTX-1 equivalent toxicities of the samples col-lected from Tarawa were all less than 1 ng g�1, though the majoritycould still be considered ciguatoxic based on the 0.01 ng g�1 P-CTX-1 equivalents threshold. Overall, 87%, 96%, 86% and 100% offish were rated as ciguateric within areas A, B, C and D of MarakeiIsland, respectively, while, 95% and 66% of fish were found to be
horax spp.) sampled in the Republic of Kiribati. Fold-differences between the toxicityshows the relationship between body weight and P-CTX-1-equivalent toxicity in
uateric fish in the Republic of Kiribati. Chemosphere (2011), doi:10.1016/

6 W.H. Chan et al. / Chemosphere xxx (2011) xxx–xxx
ciguateric, respectively, at sites E and F of Tarawa Island; the occur-rence of ciguateric fish from the two islands was generally similar.However, toxicity higher than 1 ng g�1 P-CTX-1 equivalents wasonly found at sites A (62%), B (65%) and C (46%).
Due to limited sample sizes and different capabilities for CTXaccumulation among species, the spatial distribution of ciguatoxicfish in Marakei and Tarawa Islands are shown for C. argus and Gym-nothorax spp. only (Fig. 3). C. argus and Gymnothorax spp. collectedfrom Tarawa Island (0.01–0.20 ng g�1 P-CTX-1 equivalents and0.02–0.40 ng g�1 P-CTX-1 equivalents, respectively) were signifi-cantly less toxic than those from Marakei Island (0.18–11.17 ng g�1
P-CTX-1 equivalents and 0.17–81.84 ng g�1 P-CTX-1 equivalents,respectively) (p < 0.01, Mann–Whitney U test). Among the speci-mens collected from Marakei Island, the average toxicities of fisheswere in the order of site B (5.82 ng g�1 P-CTX-1 equivalents) > siteA (4.08 ng g�1 P-CTX-1 equivalents) > site C (2.45 ng g�1 P-CTX-1equivalents) > site D (0.18 ng g�1 P-CTX-1 equivalents) for C. argusand site B (29.46 ng g�1 P-CTX-1 equivalents) > site C (3.14 ng g�1
P-CTX-1 equivalents) > site A (1.34 ng g�1 P-CTX-1 equiva-lents) > site D (0.3 ng g�1 P-CTX-1 equivalents) for Gymnothoraxspp. Samples from site B were relatively toxic, whereas those fromsite D were generally less toxic for both fish species. The distribu-tion pattern of ciguatoxic fish was similar to the results of a localsurvey showing that dinoflagellates were usually located in thenorthwestern region of Marakei Island: densities of G. toxicus werefound to be threefold higher at site B compared with site D(MRAMU, 1999). Another previous study found that the highestG. toxicus densities near sites E and F of Tarawa Island were similarto those near site D of Marakei (2–3 cells g�1; Tebano, 1984), a pat-tern which is reflected in the toxicity of the collected fish. Popula-tion studies of G. toxicus and risk assessments in ciguatera-proneregions often rely on quantifying dinoflagellates on microalgae.However, G. toxicus density and toxicity can vary greatly withchanges in season, seawater temperature and salinity (Llewellyn,2010), and many algal hosts of G. toxicus possess a variety of de-fences against being grazed by herbivores. Therefore the measure-ment of G. toxicus densities to identify ciguatera-endemic areas isnot always accurate, and should be supported with analysis of fishtoxicity. The results of the present work indicate that P-CTX-1equivalent toxicity is species- and location-dependent; for exam-ple, individual E. spilotoceps of similar size collected from the samearea of Marakei Island varied in their toxicity by up to three ordersof magnitude, and the smallest individual C. argus collected fromMarakei Island was found to exceed the toxicity threshold of0.01 ng g�1 P-CTX-1 by 900-fold. Therefore, some very commonand highly toxic Pacific species, such as C. argus and E. spilotoceps,have the potential to be marker species for identifying ciguatera-endemic areas for risk assessment purposes.
3.3. Tissue distribution in moray eel
The ciguatoxicities of the muscle and liver of moray eels areshown in Fig. 4. The samples analysed in this case were stored inplastic micro-centrifuge tubes, as the samples were extractedand stored before the discovery of the adsorption effect from theplastic tubes. Although the data are therefore not perfect, generalpatterns of the distribution of P-CTX-1 equivalent toxicity betweenthe liver and muscle can be estimated. Liver was an average of ninetimes more toxic than muscle (ranging from 4.09- to 15.43-fold) insix Gymnothorax spp. of different sizes collected from MarakeiIsland. The toxicity measured in liver from the heaviest morayeel (14 kg) was 539 ng g�1, 50 000 times higher than the safetylevel. The P-CTX-1 equivalent toxicity in liver and muscle of morayeel showed a strong positive relationship with the body weight(R2 = 0.994 and 0.955; Fig. 4, inset), reflecting strong bioaccumula-tion of CTXs, particularly in the liver. The ability of moray eels to
Please cite this article in press as: Chan, W.H., et al. Spatial distribution of cigj.chemosphere.2011.02.036
accumulate such high toxin levels without effect suggests that aresistance mechanism is present in this species, though this mech-anism is not currently known.
4. Conclusions
The results of the present study demonstrated that:
(1) Most of the fish samples collected from the two islands ofthe Republic of Kiribati were found to be ciguateric usingthe MNA, providing quantitative support to a previous state-ment that Marakei and Tarawa Islands are high-risk areas forciguatera (Lewis, 2006).
(2) The P-CTX-1 equivalent toxicity was species- and location-dependent, and within Gymnothorax spp. a strong positiverelationship between the body weight and P-CTX-1 equiva-lent toxicity in liver and muscle was observed.
(3) The occurrence of ciguateric fish was similar between thetwo sampled islands, while the toxicities of individual fishfrom Marakei were significantly greater than those fromTarawa (p < 0.01).
(4) Some carnivorous species such as groupers and moray eelscontained higher P-CTX-1 equivalent toxicities than herbi-vores and omnivores.
Further studies are warranted to determine the fluxes of CTXs inmarine ecosystems using stable isotope and fatty acid analyses tounderstand how these toxins accumulate in food webs.
Acknowledgments
We thank Mr. Being Yeeting (Senior Fisheries Scientist, Secre-tariat of the Pacific Community, New Caledonia) and the FisheriesDivision (Ministry of Fisheries and Marine Resource Development,Republic of Kiribati) for facilitating the sample collection. Wethank Ms. Rachel P.P. Wong for identification of fish species andMr. Albert C.C. Au, Mr. Xiwen Jiang, Mr. Minghua Wang and Mr.Yue Gao (School of Biological Science, The University of HongKong) for technical assistance. The work described in this paperwas supported by grants from the Research Grants Council of theHong Kong Special Administrative Region, China (City U3/CRF/08), State Key Laboratory of Marine Environmental Science(Xiamen University) cooperative Project (MEL0607) and a ScienceFaculty Collaborative Seed Grant from the University of Hong Kong.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.chemosphere.2011.02.036.
References
Beets, J., 2001. Declines in finfish resources in Tarawa lagoon, Kiribati, emphasizethe need for increased conservation effort. Atoll Res. Bull. 490, 1–14.
Bshary, R., Hohner, A., Ait-el-Djoudi, K., Fricke, H., 2006. Interspecificcommunicative and coordinated hunting between groupers and giant morayeels in the Red Sea. PLoS Biol. 4, e431.
Caillaud, A., De la Iglesia, P., Darius, H.T., Pauillac, S., Aligizaki, K., Fraga, S., Chinain,M., Diogène, J., 2010. Update on methodologies available for ciguatoxindetermination: perspectives to confront the onset of ciguatera fish poisoningin Europe. Mar. Drugs 8, 1838–1907.
Cañete, E., Diogène, J., 2008. Comparative study of the use of neuroblastoma cells(Neuro-2a) and neuroblastomaxglioma hybrid cells (NG108-15) for the toxiceffect quantification of marine toxins. Toxicon 52, 541–550.
Centers for Disease Control and Prevention (CDC), 2006. Ciguatera Fish Poisoning –Texas, 1998, and South Carolina, 2004. MMWR Morb. Mortal. Wkly. Rep. 55, pp.935–937.
uateric fish in the Republic of Kiribati. Chemosphere (2011), doi:10.1016/

W.H. Chan et al. / Chemosphere xxx (2011) xxx–xxx 7
Centers for Disease Control and Prevention (CDC), 2009. Cluster of Ciguatera FishPoisoning – North Carolina, 2007. MMWR Morb. Mortal. Wkly. Rep. 58, pp. 283–235.
Chinain, M., Darius, H.T., Ung, A., Fouc, M.T., Revel, T., Cruchet, T., Pauillac, S.,Laurent, D., 2010. Ciguatera risk management in French Polynesia: the casestudy of Raivavae Island (Australes Archipelago). Toxicon 56, 674–690.
Dickey, R.W., Plakas, S.M., 2010. Ciguatera: a public health perspective. Toxicon 56,123–136.
Empey Campora, C., Dierking, J., Tamaru, C.S., Hokama, Y., Vincent, D., 2008.Detection of ciguatoxin in fish tissue using sandwich ELISA and neuroblastomacell bioassay. J. Clin. Lab. Anal. 22, 246–253.
FAO, 2010. Kiribati-profiles Home. Fishery and Aquaculture Country Profiles. FAOFisheries and Aquaculture Department. The United Nation <http://www.fao.org/fishery/countrysector/FI-CP_KI/en>.
Hamilton, B., Hurbungs, M., Vernoux, J.P., Jones, A., Lewis, R.J., 2002. Isolation andcharacterisation of Indian Ocean ciguatoxin. Toxicon 40, 685–693.
Hamilton, B., Whittle, N., Shaw, G., Eaglesham, G., Moore, M.R., Lewis, R.J., 2010.Human fatality associated with Pacific ciguatoxin contaminated fish. Toxicon56, 668–673.
Holmes, M.J., Lewis, R.J., Poli, M.A., Gillespie, N.C., 1991. Strain dependentproduction of ciguatoxin precursors (gambiertoxins) by Gambierdiscus toxicus(Dinophyceae) in culture. Toxicon 29, 761–775.
Ledreux, A., Krys, S., Bernard, C., 2009. Suitability of the Neuro-2a cell line for thedetection of palytoxin and analogues (neurotoxic phycotoxins). Toxicon 53,300–308.
Lehane, L., Lewis, R.J., 2000. Ciguatera: recent advances but the risk remains. Int. J.Food Microbiol. 61, 91–125.
Lewis, R.J., 2006. Ciguatera: Australian perspectives on a global problem. Toxicon48, 799–809.
Lewis, R.J., Sellin, M., Poli, M.A., Norton, R.S., MacLeod, J.K., Shell, M.M., 1991.Purification and characterization of ciguatoxins from moray eel (Lycodontisjavanicus, Muraenidae). Toxicon 29, 1115–1127.
Litaker, R.W., Vandersea, M.W., Faust, M.A., Kibler, S.R., 2010. Global distribution ofciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon 56, 711–730.
Llewellyn, L.E., 2010. Revisiting the association between sea surface temperatureand the epidemiology of fish poisoning in the South Pacific: reassessing the linkbetween ciguatera and climate change. Toxicon 56, 691–697.
Please cite this article in press as: Chan, W.H., et al. Spatial distribution of cigj.chemosphere.2011.02.036
Marine Resource Assessment and Monitoring Unit, 1999. Live Ref Fish andCiguatoxic (Fish Poisoning) Reef Areas Surveys Conducted on the Island ofMarakei. Marine Resource Assessment and Monitoring Unit Report: 1999.Republic of Kiribati.
McCarthy, D., Tebano, T., 1988. Ciguatoxic Fish Poisoning and the CausativeOrganism in the Gilbert Islands, Kiribati. Atoll Research and Development Unit,University of South Pacific and University of Hawaii 125.
NMFS. Fisheries Statistics of the United States, 2009. Current Fisheries Statistics No.2008, vol. 118. Washington, D.C., pp. 1–15.
Oshiro, N., Yogi, K., Asato, S., Sasaki, T., Tamanaha, K., Hirama, M., Yasumoto, T.,Inafuku, Y., 2010. Ciguatera incidence and fish toxicity in Okinawa, Japan.Toxicon 56, 656–661.
Pottier, I., Vernoux, J.P., Lewis, R.J., 2001. Ciguatera fish poisoning in theCaribbean islands and Western Atlantic. Rev. Environ. Contam. Toxicol. 168,99–141.
Rayner, M.D., 1972. Mode of action of ciguatoxin. Fed. Proc. 31, 1139–1145.Roeder, K., Erler, K., Kibler, S., Tester, P., The, H.V., Nguyen-Ngoc, L., Gerdts, G.,
Luckas, B., 2010. Characteristic profiles of Ciguatera toxins in different strains ofGambierdiscus spp. Toxicon 56, 731–738.
Tebano, T., 1984. Population Density Study on a Toxic Dinoflagellate Responsible forCiguateria Fish Poisoning on South Tarawa Island, Republic of Kiribati. IslandResearch and Development Unit, Tanaea, Tarawa, Republic of Kiribati.
Tester, P.A., Feldman, R.L., Nau, A.W., Kibler, S.R., Litaker, W.R., 2010. Ciguatera fishpoisoning and sea surface temperatures in the Caribbean Sea and the WestIndies. Toxicon 56, 698–710.
Tikai, T., 1988. Ciguatoxic Fish Poisoning in Kiribati. Twentieth Regional TechnicalMeeting on Fisheries. South Pacific Commission, Noumea, New Caledonia,August 1–5 1988. SPC/Fisheries 20/WP.25.
US Department of State, 2010. Background Notes-Kiribati. Bureau of Public Affairsand Pacific Affairs, US Department of State, Washington <http://www.state.gov/r/pa/ei/bgn/1836.htm>.
Van Dolah, F.M., 2000. Marine algal toxins: origins, health effects, and theirincreased occurrence. Environ. Health Perspect. 108, 133–141.
Wong, C.K.K., Hung, P., Lee, K.L., Kam, K.M.M., 2005. Study of an outbreak ofciguatera fish poisoning in Hong Kong. Toxicon 46, 563–571.
Wong, C.K.K., Hung, P., Lee, K.L., Mok, T., Chung, T., Kam, K.M.M., 2008. Feature ofciguatera fish poisoning cases in Hong Kong 2004–2007. Biomed. Environ. Sci.21, 521–527.
uateric fish in the Republic of Kiribati. Chemosphere (2011), doi:10.1016/




















