
Spatial distribution and trophic diversity of nematode populations under Acacia raddiana along a...
17
www.elsevier.com/locate/jnlabr/yjare Journal of Arid Environments Journal of Arid Environments 56 (2004) 339–355 Spatial distribution and trophic diversity of nematode populations under Acacia raddiana along a temperature gradient in the Negev Desert ecosystem Stanislav Pen-Mouratov a , Xueli He a,b , Yosef Steinberger a, * a Faculty of Life Sciences, Bar-Ilan University, Ramat Gan 52900, Israel b Faculty of Life Sciences, Northwest Science and Technology University of Agriculture and Forestry, Shaanxi 712100, China Received 13 September 2002; received in revised form 6 March 2003; accepted 2 April 2003 Abstract The aim of this study was to determine the spatial distribution and trophic diversity of free- living soil nematode populations in the most extreme xeric period, along a temperature gradient from the northern Negev Desert toward its southern edge, with mean annual temperature of 20–23 C and mean annual rainfall of 150–42 mm, respectively. Seventeen nematode taxa were identified, with bacterial feeding found to be dominant, with 52%, 24% and 12% at three stations, respectively. We found that when environmental temperatures reached high values, the influence of soil moisture and organic matter on the nematode population minimized, temperature being one of the factors determining community composition and structure. r 2003 Elsevier Science Ltd. All rights reserved. Keywords: Nematode community; Trophic groups; Acacia raddiana; Ecological indices; Desert ecosystem 1. Introduction Acacia is the dominant tree and one of the most important biological resources in the Negev Desert and the Arava Valley, Israel (Zohary, 1972). Most trees in these areas are either Acacia raddiana Savi, A. tortilis (Forssk.) Hayne or A. negevensis (Halevy and Orshan, 1972; Danin, 1983) and they provide food and shelter for many ARTICLE IN PRESS *Corresponding author. Fax: +972-3-5351824. E-mail address: [email protected] (Y. Steinberger). 0140-1963/03/$ - see front matter r 2003 Elsevier Science Ltd. All rights reserved. doi:10.1016/S0140-1963(03)00058-2
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Spatial distribution and trophic diversity of nematode populations under Acacia raddiana along a...

www.elsevier.com/locate/jnlabr/yjare
Journal of AridEnvironments
Journal of Arid Environments 56 (2004) 339–355
Spatial distribution and trophic diversity ofnematode populations under Acacia raddianaalong a temperature gradient in the Negev
Desert ecosystem
Stanislav Pen-Mouratova, Xueli Hea,b, Yosef Steinbergera,*aFaculty of Life Sciences, Bar-Ilan University, Ramat Gan 52900, Israel
bFaculty of Life Sciences, Northwest Science and Technology University of Agriculture and Forestry,
Shaanxi 712100, China
Received 13 September 2002; received in revised form 6 March 2003; accepted 2 April 2003
Abstract
The aim of this study was to determine the spatial distribution and trophic diversity of free-
living soil nematode populations in the most extreme xeric period, along a temperature
gradient from the northern Negev Desert toward its southern edge, with mean annual
temperature of 20–23�C and mean annual rainfall of 150–42mm, respectively. Seventeen
nematode taxa were identified, with bacterial feeding found to be dominant, with 52%, 24%
and 12% at three stations, respectively. We found that when environmental temperatures
reached high values, the influence of soil moisture and organic matter on the nematode
population minimized, temperature being one of the factors determining community
composition and structure.
r 2003 Elsevier Science Ltd. All rights reserved.
Keywords: Nematode community; Trophic groups; Acacia raddiana; Ecological indices; Desert ecosystem
1. Introduction
Acacia is the dominant tree and one of the most important biological resources inthe Negev Desert and the Arava Valley, Israel (Zohary, 1972). Most trees in theseareas are either Acacia raddiana Savi, A. tortilis (Forssk.) Hayne or A. negevensis
(Halevy and Orshan, 1972; Danin, 1983) and they provide food and shelter for many
ARTICLE IN PRESS
*Corresponding author. Fax: +972-3-5351824.
E-mail address: [email protected] (Y. Steinberger).
0140-1963/03/$ - see front matter r 2003 Elsevier Science Ltd. All rights reserved.
doi:10.1016/S0140-1963(03)00058-2

desert animals (Ernst et al., 1989; Ruess and Halter, 1990; Milton and Dean, 1995;Oba, 1998; Rohner and Ward, 1999). The biological diversity at these sites isdetermined to a large extent by the structure and long-term dynamics of the Acacia
trees (Wiegand et al., 1998).The African Acacia species in the Negev Desert, A. raddiana, is currently believed
to be endangered (Ward and Rohner, 1997). Many current investigations haveconcentrated on the evaluation of the conservation status of these species and theestimation of the mortality of mature Acacia trees (Ward and Rohner, 1997;Perevolotsky et al., 1998; Rohner and Ward, 1999). The results of these studies haveprovided important insights into the interaction between plants and environment.However, the study of the interaction of Acacia trees and its nearest environmentseems to be far from complete (Wiegand et al., 1999; Lahav-Ginott et al., 2001). Oneway of understanding the adaptation and contribution of A. raddiana in the Israelidesert is by studying the interaction between trees and soil biota, where the latterreflects their food source and provides information on the functioning of the soilfood web (Moore and Ruiter, 1991). One of the important soil biota groups whichplay a leading role as regulators of energy flow are the nematode population (Chew,1974). The soil free-living nematodes are one of the most numerous groups amongthe multicellular animals (Bongers and Ferris, 1999), participating in fundamentalecological processes in soil, such as decomposition and nutrient cycling (Freckman,1988; De Goede and Bongers, 1994). However, since they are sensitive to ecosystemdisturbances (Wardle et al., 1995; Freckman and Virginia, 1997; Yeates et al., 1999),they can be used as stable indicators for understanding processes in the soils ofdifferent ecosystems, including the semi-arid and arid zones of deserts (Manlay et al.,2000; Liang et al., 2001).
The main factors determining the activity and diversity of soil biota in aridecosystems are soil temperature, soil water potential (Steinberger and Sarig, 1993;Whitford, 1996), soil organic matter and vegetation (Freckman and Mankau, 1986;Duponnois et al., 1999). Soil temperature and soil moisture content were found to beone of the most important factors which directly or indirectly affect the seasonalpopulation dynamics of nematodes in the soil by altering metabolic activity andquality of their food source (Yeates and Coleman, 1982). These assertions are typicalfor hot desert ecosystems (Alon and Steinberger, 1999). The climate models ofWatson et al. (1998) indicate that the mean annual global surface temperature willrise by 1–3.5�C by the year 2100. This predominant factor will obviously determinechanges not only in ecosystems but in the nematode community as well.
The main aims of the present study were to define the trophic and spatialstructures of soil nematode communities living under A. raddiana along a climaticgradient in the Negev Desert.
Based on previous investigations, we assume that the trophic structure of thenematode population is spatially dependent on the location of the Acacia and variesin different parts of the desert that has peculiar climatological conditions.Furthermore, under the most unfavorable conditions in the desert ecosystem, e.g.,during the dry period, temperature can function as the main ecological factor,determining the changes in the structure of the nematode community.
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355340

The study of the nematode community in the Acacia rhizosphere will greatlycontribute to the understanding of the rhizosphere’s environmental contribution asmediator under extreme xeric environments and its affect on soil biota community.
2. Materials and methods
2.1. Location and geo-climatological conditions
Soil samples were collected from three sites along a climatological gradientrunning from the northern Negev Desert 25 km east of Dimona (ST I) (30058�N;35020�E), with a mean annual temperature of 20�C and a mean annual rainfall of150mm, toward south of the Dead Sea, Ein Ofarin in the Arava Valley (ST II)(30045�N; 35018�E) with a mean yearly temperature of 23�C and mean rainfall of42mm, and southwest towards the Faran Desert (ST III) (30012�N; 34055�E) with amean annual temperature of 21�C and a mean annual rainfall of below 42mm. Allthree sites exhibit arid conditions with a common feature of low and unpredictablerainfall and desert climate, i.e., mild, rainy winters and hot, dry summers. Radiationmay reach 3.14� 104 kJm�2 d�1. Many locations in this area are dotted bynumerous green Acacia trees such as A. spirocarpa, A. negevensis and A. raddiana,contrasting sharply with the brown-yellow landscape (Evenari et al., 1982). The soilsare brown, shallow, rocky, desert soils (brown lithosols), and loessial and gray desertsoils (loessial sierozems) (Dan et al., 1972).
2.2. Sampling
Samples were taken from under A. raddiana during the driest part of the summer(August 2000), from the 0–50 cm soil layer. A total of 143 soil samples were collectedfrom each 10-cm layer using a 7-cm diameter auger.
Sampling points were located in the following order: near the stems (herewithcalled ‘‘under’’), at the edge of the canopy of the Acacia trees (‘‘radius’’), and tworadii from the stem (‘‘control’’). Soil samples were placed in individual plastic bagsand transported to the laboratory in an insulated container. The soil samples werekept in cold storage at 4�C and before biological and chemical analysis they weresieved through a 2-mm mesh sieve.
2.3. Laboratory analysis
Soil moisture was determined gravimetrically by drying at 105�C to a constantweight. Organic matter content was calculated from the percentage of organiccarbon estimated by oxidization with dichromate in the presence of H2SO4, withoutapplication of external heat (Rowell, 1994).
Nematodes were extracted from 100 g soil samples using the Baermann funnelprocedure (Cairns, 1960) and nematode populations were expressed per 100 g drymass soil. The recovered organisms were counted and preserved in formalin
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355 341

(Steinberger and Sarig, 1993). The total nematodes extracted were used foridentification, mainly to genus level, using a compound microscope.
2.4. Ecological indices and statistical analysis
The characteristics of the nematode communities were described by means of thefollowing indices: (1) total number of nematodes (100 g�1 dry soil); (2)trophic structure: bacterial-feeding (BF), fungal-feeding (FF), plant-parasitic(PP) and omnivores-predators (OP) (Steinberger and Loboda, 1991; Steinbergerand Sarig, 1993; Liang et al., 2000); (3) ecological indices: F/B=FF/BF(Twinn, 1974); WI=(FF+BF)/PP (Wasilewska, 1994); trophic diversity,T ¼ 1=SP2
i ; where Pi is the proportion of the ith trophic group (Heip et al., 1988);Shannon index, H ¼ �SPi (lnPi), where p is the proportion of individuals inthe ith taxon (Shannon and Weaver, 1949); maturity index SMi ¼ Sniri where ni isthe c � p value assigned by Bongers (1990) of the ith genus in the nematode and ri
the proportion of the genus in the nematode community. The c � p values describethe nematode life strategies, and range from 1 (for colonizers, tolerant todisturbance) to 5 (for persisters, sensitive to disturbance); maturity indexmodification (SMI), includes both PP and free-living nematodes (Yeates, 1994);specific richness, SR ¼ ðS � 1Þ=lnðNÞ; where S is the number of taxa and N is thenumber of individuals identified (Yeates and King, 1997); evenness index, J 0 ¼H 0=lnðSÞ; where H 0 is the Shannon–Weiner index, ln is the log base (e) and S is equalto number of taxa (Pielou, 1975).
All obtained data were examined by analysis of variance (ANOVA) usingnematode species, nematode trophic groups, soil moisture, organic matter, location,points of different distances from stem and depth as location values at the threestations along the temperature gradient. Differences with Po0:05 were consideredsignificant.
3. Results
3.1. Soil moisture and organic matter
No significant differences (p > 0:05) were found in soil moisture and organicmatter contents (n ¼ 143) between the different stations. Soil moisture contentincreased from 0–10 to 20–30 cm soil depths with no significant changes toward thedeeper layers of soil. Organic matter content decreased toward the deeperlayers from 0–10 to 40–50 cm near the stems (base) and at the edge of the treecanopies. In contrast, organic matter of the control soil samples increasedtoward the deeper layers from 0–10 to 40–50 cm. The soil organic matterunder A. raddiana at all depths ranged between 7.0% and 13.9%. The values ofsoil organic matter in the control soil samples were found to vary between 5.9%and 7.2%.
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355342

3.2. Nematode community structure
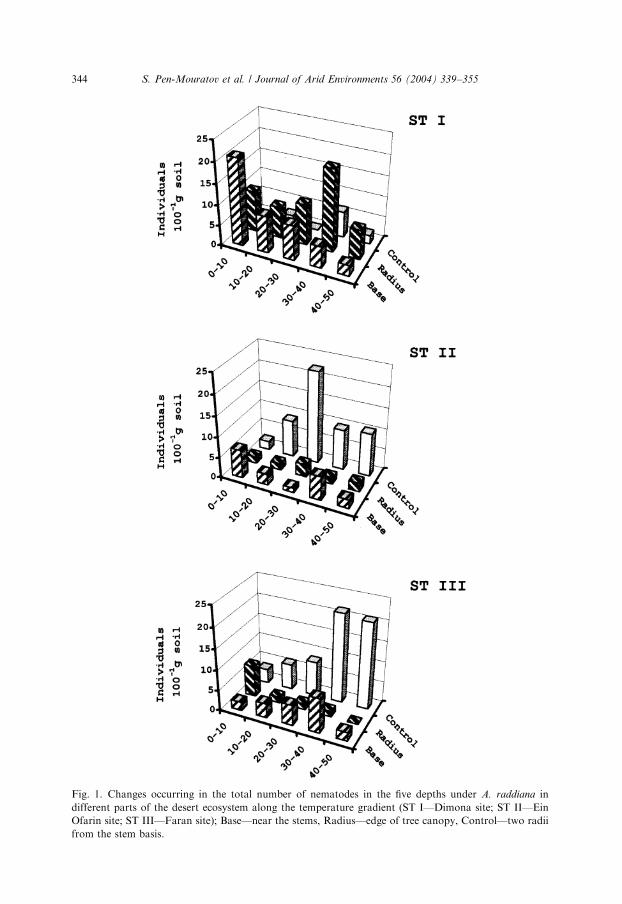
Seventeen nematode taxa were identified in the present investigation. The meanvalues of nematode found in the soil samples ranged from 0 to 22 individuals per100-g dry soil. The total number of nematodes extracted at the ST I—Dimona siteunder trees was found to be significantly higher in comparison to the other twolocations (po0:05), the ST II—Ein Ofarin and ST III—Faran sites (Fig. 1).Moreover, three different trends were obtained, each representing one of the threestations. Furthermore, at each station the ‘‘under’’, ‘‘radius’’ and ‘‘control’’performed a different trend (Table 1).
Under Acacia at the ST I site (Fig. 1A), a gradual decrease in total number ofnematodes from 21 to 3 individuals 100 g�1 soil from 0–10 to 40–50 cm depth wasobserved. At the canopy edge at the ST I site, the total number of nematodes rangedbetween 7 and 20 individuals per 100 g�1 soil, reaching maximum in the 30–40 cmsoil layer, while in the control soil samples, the total number of nematodes was foundto be significantly lower (po0:05) in comparison to the other two samplinglocations, ranging between 0 to a maximum of 6 individuals 100 g�1 soil (Fig. 1).
At the ST II and ST III sites, the mean maximal value of soil free-living nematodepopulation found in the control samples at 20–30 cm depth (ST II) and at the 30–40and 40–50 cm layers (ST III) was 22, 21 and 21 individuals g�1, respectively (Fig. 1—ST IIB and ST III). At the other layers of the Ein Ofarin and Faran sites, the totalnumber of nematodes ranged between 0 and 10 individuals 100 g�1 dry soil (Fig. 1—ST II and ST III).
The total number of nematodes under tree ‘‘base’’ and ‘‘radius’’ decreased fromthe STI toward the STII and STIII sites and amounted to 58%, 23%, 20% and 74%,14%, 12%, respectively. But these data did not demonstrate differences between thetotal number of nematodes next to the tree stems (‘‘base’’) and the edge of thecanopy of the Acacia trees (‘‘radius’’), where the relative abundance of soil free-livingnematodes was found to be higher in the soil samples taken at the STI and increasedtoward the STIII site. The significantly different patterns in total nematode numbersbetween sites, depths and the three sampling locations, visually elucidate the transectof abiotic effect.
3.2.1. Trophic groups
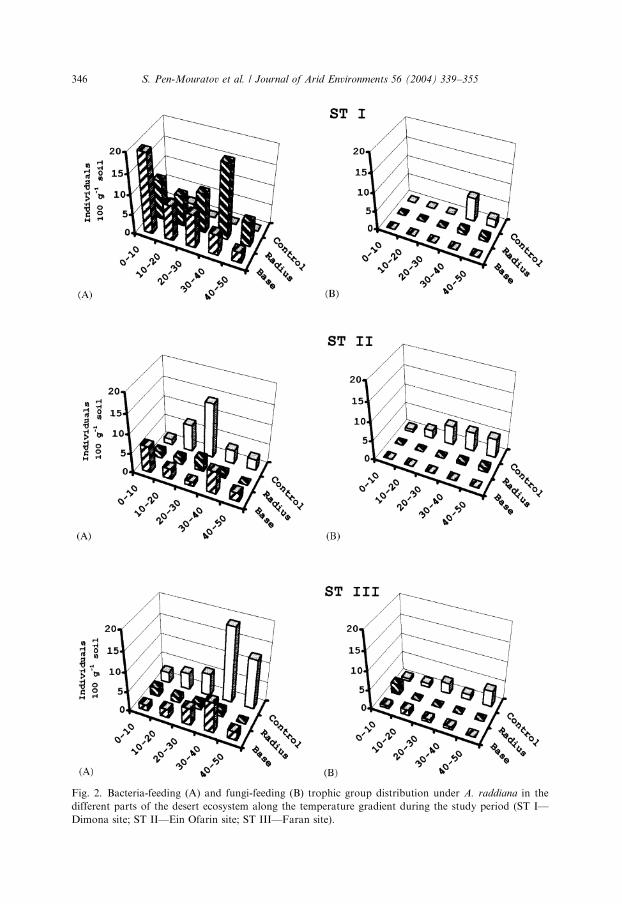
BF nematodes were found to be the most dominant trophic group at all stations(Table 1) following, in general, the trend described for total number of nematodepopulation decreasing from ST I toward the ST II and ST III sites under tree ‘‘base’’and ‘‘radius’’ and amounting to 19, 8, 6 and 22, 3, 2 individuals 100 g�1 soil,respectively (Fig. 2 ST IA; ST IIA; ST IIIA). Data obtained showed that at the twoupper layers (0–20 cm) at the Dimona site (STI), the mean values were higher than atthe Ein Ofarin (STII) and Faran (STIII) sites. However, in the control samples at thedifferent sites, the value at the STIII showed the highest values at the deeper layers(30–50 cm) reaching maximum values of 18.8 individuals 100 g�1 soil. Densities ofBF trophic groups were found to be significantly different between the observed sites(p ¼ 0:01; n ¼ 143) (Fig. 2 ST IA; ST IIA; ST IIIA; Table 2).
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355 343
ARTICLE IN PRESS
Fig. 1. Changes occurring in the total number of nematodes in the five depths under A. raddiana in
different parts of the desert ecosystem along the temperature gradient (ST I—Dimona site; ST II—Ein
Ofarin site; ST III—Faran site); Base—near the stems, Radius—edge of tree canopy, Control—two radii
from the stem basis.
S. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355344
ARTIC
LEIN
PRES
STable 1
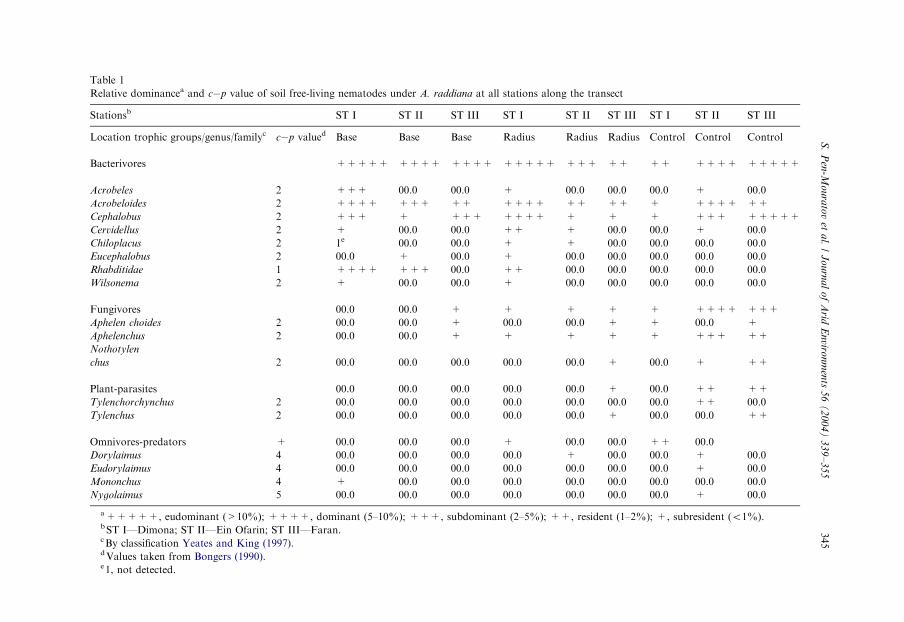
Relative dominancea and c�p value of soil free-living nematodes under A. raddiana at all stations along the transect
Stationsb ST I ST II ST III ST I ST II ST III ST I ST II ST III
Location trophic groups/genus/familyc c�p valued Base Base Base Radius Radius Radius Control Control Control
Bacterivores +++++ ++++ ++++ +++++ +++ ++ ++ ++++ +++++
Acrobeles 2 +++ 00.0 00.0 + 00.0 00.0 00.0 + 00.0
Acrobeloides 2 ++++ +++ ++ ++++ ++ ++ + ++++ ++
Cephalobus 2 +++ + +++ ++++ + + + +++ +++++
Cervidellus 2 + 00.0 00.0 ++ + 00.0 00.0 + 00.0
Chiloplacus 2 1e 00.0 00.0 + + 00.0 00.0 00.0 00.0
Eucephalobus 2 00.0 + 00.0 + 00.0 00.0 00.0 00.0 00.0
Rhabditidae 1 ++++ +++ 00.0 ++ 00.0 00.0 00.0 00.0 00.0
Wilsonema 2 + 00.0 00.0 + 00.0 00.0 00.0 00.0 00.0
Fungivores 00.0 00.0 + + + + + ++++ +++
Aphelen choides 2 00.0 00.0 + 00.0 00.0 + + 00.0 +
Aphelenchus 2 00.0 00.0 + + + + + +++ ++
Nothotylen
chus 2 00.0 00.0 00.0 00.0 00.0 + 00.0 + ++
Plant-parasites 00.0 00.0 00.0 00.0 00.0 + 00.0 ++ ++
Tylenchorchynchus 2 00.0 00.0 00.0 00.0 00.0 00.0 00.0 ++ 00.0
Tylenchus 2 00.0 00.0 00.0 00.0 00.0 + 00.0 00.0 ++
Omnivores-predators + 00.0 00.0 00.0 + 00.0 00.0 ++ 00.0
Dorylaimus 4 00.0 00.0 00.0 00.0 + 00.0 00.0 + 00.0
Eudorylaimus 4 00.0 00.0 00.0 00.0 00.0 00.0 00.0 + 00.0
Mononchus 4 + 00.0 00.0 00.0 00.0 00.0 00.0 00.0 00.0
Nygolaimus 5 00.0 00.0 00.0 00.0 00.0 00.0 00.0 + 00.0
a+++++, eudominant (>10%); ++++, dominant (5–10%); +++, subdominant (2–5%); ++, resident (1–2%); +, subresident (o1%).bST I—Dimona; ST II—Ein Ofarin; ST III—Faran.cBy classification Yeates and King (1997).dValues taken from Bongers (1990).e1, not detected.
S.
Pen
-Mo
ura
tov
eta
l./
Jo
urn
al
of
Arid
En
viron
men
ts5
6(
20
04
)3
39
–3
55
345
ARTICLE IN PRESS
Fig. 2. Bacteria-feeding (A) and fungi-feeding (B) trophic group distribution under A. raddiana in the
different parts of the desert ecosystem along the temperature gradient during the study period (ST I—
Dimona site; ST II—Ein Ofarin site; ST III—Faran site).
S. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355346
The changes in FF and PP trophic groups of the nematode population arepresented in Fig. 2 ST IB; ST IIB; ST IIIB and Fig. 3 ST IA; ST IIA; ST IIIA). Oneof the most peculiar findings is the significantly higher (po0:05) population outsidethe rhizosphere and radius effect region in the control soil samples, reachingmaximum mean values of 6.3 and 4.2 individuals 100 g�1 soil for FF and plantparasitic trophic groups, respectively. The densities of the OP trophic group, like thedensities of fungi-feeding trophic groups, were low at the observed stations and wererelated to locations and sites (Fig. 3B). Similar to the FF, the PP trophic groupexhibited a significant difference between locations, but not between sampling sites(Table 2).
The omnivore-predator trophic groups were found in relatively low numbers, withhigher presence in the open spaces (control), and maximum mean value (2individuals 100 g�1 soil) at the STIII.
3.3. Nematode species diversity
A total of 17 genera were found with eight bacterivores, three fungivores, twoplant-parasites and four OP (Table 1). A significant decrease in bacterial-feeders
ARTICLE IN PRESS
Table 2
Univariate statistics by ‘‘ANOVA’’ for the nematode communities under A. raddiana
Variable Stations p value Location p value Locat. & station p value Depth p value
(1) Total number of nematodes
0.13 0.11 0.03 0.67
(2) Trophic structure
BF 0.01 0.71 0.03 0.47
FF 0.19 0.0001 0.53 0.48
PP 0.09 0.02 0.27 0.82
OP 0.07 0.09 0.04 0.3
(3) Ecological indices
F/B 0.29 0.0004 0.42 0.43
WI 0.29 0.01 0.02 0.59
T 0.006 0.19 0.0001 0.62
H0 0.09 0.03 0.0001 0.57
MI 0.01 0.07 0.0032 0.78
SMI 0.02 0.04 0.004 0.79
SR 0.09 0.05 0.0001 0.78
J0 0.13 0.16 0.001 0.34
Indices are: (3) total abundance: individuals 100 g�1 dry soil; (4) trophic structure—BF, bacterial-feeding;
FF, fungal-feeding; PP, plant parasitic; OP, omnivores-predatores. (5) Ecological indices—WI, ratio of
bacterial-feeding and fungal-feeding to plant-parasitic; F/B, fungal-feeding/bacterial-feeding ratio; T,
trophic diversity; H0, Shannon index; MI, maturity index; SMI; modified maturity index; SR, richness; J0,
evenness index.
S. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355 347

ARTICLE IN PRESS
Fig. 3. Plant parasite (A) and omnivore-predator (B) trophic group distribution under A. raddiana in the
different parts of the desert ecosystem along the temperature gradient during the study period (ST I—
Dimona site; ST II—Ein Ofarin site; ST III—Faran site).
S. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355348

presence was observed from the relatively mild sites to the extreme site where a totalof eight dominant taxa decreased 50% toward the second (four taxa) and third (twotaxa) sites. However, an opposite trend was obtained for the fungal-feeder nematodetaxa, which increased from the relatively mild toward the harsh environment.Regarding plant-porites, representative taxa were present at the ST I site, while OPwere not found in the ST III samples. Cephalobus was found at the STI and STIII(next to the tree stem—‘‘base’’), at the ST I site (the edge of the canopy of trees—‘‘radius’’), and at the ST II and ST III sites (control soil samples—‘‘control’’) in thecapacity of eudominant, dominant and subdominant, respectively. Rhabditides werefound at the ST I and ST II (base) sites as dominant and subdominant, respectively.Acrobeles (base) and Aphelenchus (control) were found as subdominants at the ST Iand ST II sites, respectively.
The species dispersal of other nematode species were restricted by location. Forexample, Tylenchorchynchus and Tylenchus were found as subresidents in controlsamples at the ST II and ST III sites, respectively. Dorylaimus was found at the edgeof the canopy of the Acacia trees and in the control soil samples at the ST II assubresident. Eudorylaimus and Nygolaimus were found in the control samples at theST II site as subresidents. Mononchus was found next to tree stems at the ST I site assubresident (Table 1). In general, nematode abundance in the different trophicgroups was spatially dependent on tree location, therefore, species nematodeabundance of BF trophic group under trees is almost double the amount of that inthe control soil samples. The proportion of species abundance of BF trophic groupunder trees (‘‘base’’ and ‘‘radius’’) and in control samples amounted to 12%, 14%,7%, respectively. The proportion of species nematode abundance of FF trophicgroups increased from base to control and amounted to 2%, 5%, 7%, respectively.
The plant parasitic trophic group was absent at all soil sample stations next to treestems (‘‘base’’), present in the ‘‘radius’’ soil samples only at station 3 (0.56nematodes, in the mean 100 g�1 dry soil samples) and in the control soil samples atstations 2 and 3 (0.67 and 1.34 nematodes 100 g�1 of dry soil, respectively) (po0:05;n ¼ 143). Proportion of omnivore-predator nematodes at the ST I, ST II and ST IIIsites amounted to 1%, 1%, 3% of the total population, respectively. Differences inthe plant-parasitic group were found between sampling location and omnivore-predator groups were found between location-site interaction (Table 2).
3.4. Ecological indices
The ratio of FF to BF (F/B) at the different sampling sites and locations below theAcacia tree was found to be less than one (Table 2). F/B index at stations 1 and 2next to tree stems and at station 1 in the control samples could not be calculatedbecause one or more of the two trophic groups was not detected. Mean value of F/Bindex in the ‘‘base’’ soil samples at station 3 was equal to 0.16. The value for the‘‘radius’’ soil samples ranged between 0.05 and 0.13. F/B index in control samplesvaried from 0.11 to 0.74. However, significant effects were observed only between thelocations (po0:0004), and significant effects were found between the stations orbetween the depths (po0:05) (Table 2).
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355 349

Values of ratio of BF and FF to PP (WI) index were found to be very low at allstations at the base and tree canopy sampling locations. However, an increaseappeared in control soil samples at stations 2 and 3, with mean values rangingbetween 1.47 and 1.36, elucidating significant location effect (po0:01).
At all the observed sites, the mean values of trophic diversity (T) changed, rangingfrom 0.42 to 9.03 for the control samples and from 1.25 to 10.65 for the soil samplesunder trees. Mean values of trophic diversity were maximal under trees at station 1,with no difference at the other two stations. In the control soil samples, T meanvalue was maximal at stations 2 and 3 and minimal at station 1. At all the observedsites, mean trophic diversity changed, ranging from 0.45 to 1.60. Changes in meanvalues of MI and its modification were similar to each other at the observed station.They were maximal at station 1 and minimal at station 2. Mean values of Shannonindex (H0) were maximal in the control soil samples (0.44), minimal for the ‘‘base’’soil samples (0.29), and significantly affected by sampling location (po0:03) and bythe interaction between the site and sampling location (Table 2).
The c�p values reflecting the nematode life strategies and their sensitivity toenvironmental disturbance ranged from 1 (for colonizers, tolerant to disturbance) to5 (for persisters, sensitive to disturbance) (Table 2). Total number of nematodeshaving c�p values 1 and 2, decreased from ST I toward the ST II and ST III sitesunder tree ‘‘base’’ and ‘‘radius’’ and amounted to 19%, 8%, 7% and 22%, 4%, 4%,respectively. In contrast, total number of nematodes in control soil samples increasedfrom ST I toward the ST II and ST III sites and amounted to 1%, 16%, 18%,respectively. Total number of nematodes having c�p value of 4 and 5, were low at allobserved stations and ranged from 0% to 1.3% of total number of nematodes of allsites. In general, nematodes belonging to c�p values of 1 and 2 and 4 and 5composed 98% and 2%, of the total population, respectively.
MI values were not different under trees and in control samples. In contrast to MI,the values of MI modification were significantly different under trees and in controlsamples (Table 2). At all the observed sites, MI and MI modification ranged from 0to 3.
Species richness was maximal in the control samples (0.38) and minimal in the‘‘base’’ soil samples (0.26). Significant differences were found (po0:05) as a result oflocation and the interaction between site and location, while no site effect was found.
No significant difference in evenness index (J0) was observed between the sites,locations and depths, however, the importance of site location interaction was foundto be significant.
4. Discussion
Activity, density and species diversity in the desert ecosystem are connectedtraditionally with soil moisture, soil organic matter and temperature of environment.These factors lead to mosaic distribution of soil organisms in the different groundareas. One of the factors influencing soil organism location, especially in the desert
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355350

ecosystem, is plant presence. Plants, essentially, are the local ecological factorcreating niches which moderate the environmental surroundings.
Our data indicated that there were no significant differences in soil moisture andorganic matter content between the different sites during the study period. In thisenvironment, long-term temperature effect could function as a major ecologicalfactor in determining changes in the structure of the nematode community, while theactivity of other dominant environmental factors (soil moisture and organic matter)was minimized. No significant correlation was found between trophic groups andorganic matter, and not between trophic groups and soil moisture. These data agreewith data reported by Verschoor et al. (2001), who detected a low correlationbetween nematode density and environmental factors, including soil temperature,soil moisture, precipitation and root biomass. However, they (Verschoor et al., 2001)emphasized the importance of environmental factors as a more spatial influence onthe species character than on the total number of nematodes.
The nematode mean under trees was maximal at the ST I site, decreasing towardST II and ST III, however, statistical analysis did not reveal any difference of totalnematode density in general, between the observed stations. Moreover, total numberof nematodes at each location (base, radius and control) exhibited significantdifferences at the observed stations (po0:05). Total number of nematodes under tree‘‘base’’ and ‘‘radius’’ decreased from ST I toward the ST II and ST III sites andamounted to 58%, 23%, 20% and 74%, 14%, 12%, respectively, while total numberof nematodes in control soil samples increased from ST I toward the ST II and ST IIIsites and amounted to 3%, 46% and 51%, respectively.
These data indicate that nematode populations exhibit sensitivity to temperaturechanges by active migration. In addition, in the northern part of the desert during agiven season of year, nematodes preferred to exist in the upper layers of the treerhizosphere and, probably in the deeper soil layers outside of the tree rhizosphere’sinfluence.
In contrast, in the southern part of the desert during a given season of year,nematodes preferred to exist in the upper soil layers of area without trees, and,probably in the deeper soil layers under trees.
The most numerous nematode species found at the investigated sites wereCephalobus (34%), Acrobeloides (28%), Rhabditis (12%) and Aphelenchus (8%).
The BF trophic group was dominant at all stations. The BF group, like totalnumber of nematodes, decreased from ST I toward the ST II and ST III sites undertree ‘‘base’’ and ‘‘radius’’ and amounted to 19%, 8%, 6% and 22%, 3%, 2%,respectively. In contrast, BF group in control soil samples increased from ST Itoward the ST II and ST III sites and amounted to 0.2%, 9.0% and 14.0%,respectively. The BF group exhibited the highest resistance to the high temperaturesof the desert compared to the other trophic groups.
Both total number of nematodes and number of separate trophic groups of the‘‘base’’ were not different from the total number of nematode and number ofseparate trophic groups of the ‘‘radius’’. Densities and species diversity of FF and PPtrophic groups were significantly higher toward the extreme sites in the control thanunder the trees. This might mean that under predominantly high temperature values
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355 351

of desert environment, decomposition occupies a more leading role in soil processes;processes are mainly fungi community-controlled. During the time of ourinvestigation, the control soil samples also demonstrated more species diversity inomnivore-predator trophic groups.
In observed research, ecological indices such as F/B, WI, H0, SR and J0 showedsignificant differences only on locations but not between different sites. The trophicdiversity index describing functional groups within the nematode communitiesexhibited different values at the different investigation stations along theclimatological gradient. The T value obtained by us was significantly higher at theST I site than at the other two sites. These data demonstrate that the conditions ofthe nematode community reached a more steady state at the ‘‘northern’’ station thanat the more southern station. The mean trophic diversity value in the present study(T ¼ 0:84) was lower than the value (T ¼ 1:98) obtained by Steinberger et al. (2001)along the climate gradient in the west-eastern part of the Judean Desert in Israelduring a longer period of investigation.
Comparing total number of nematodes of two groups belonging to different life-strategies showed that all nematodes, both those belonging to r-life-strategy andthose belonging to K-life-strategy, have spatial dependence. Total number ofnematodes belonging to r-strategy (tolerant to disturbance) decreased from ST Itoward the ST II and ST III sites under tree ‘‘base’’ and ‘‘radius’’ and amounted to19%, 8%, 7% and 22%, 4%, 4%, of the total nematode abundance, respectively. Incontrast, total number of nematodes in control soil samples increased from ST Itoward the ST II and ST III sites and amounted to 1%, 16% and 18%, respectively.Total number of nematodes belonging to K-strategy (sensitive to disturbance) waslow at all the observed stations and ranged from 0% to 1.3% total nematodes at allsites. It is necessary to indicate that nematodes belonging to ‘‘temperate’’ life groups,i.e., having c�p value equal to 3, were absent along the temperature gradient. Thisprobably means that nematodes located in the mid-area of the c�p life strategy scalehave less chance to survive in the harsh desert environment than nematodes locatedat the two ends of the above-mentioned scale.
The MI and its modification (SMI) were sensitive to changes along theclimatological gradient. The values of MI and SMI related to the ‘‘free-living’’component of the nematode community varied at the observed stations and were1.32 and 1.34, respectively. The maturity indices in the present investigation werelower than the data (1.80–3.54) obtained in other sites around the world(Wasilewska, 1994; Yeates, 1994; Porazinka et al., 1997; Liang et al., 2001). Themeans of the maturity indices were higher at station 1 than at the other two stations.This finding is in agreement with the data of other investigators (Bakonyi and Nagy,2000) who determined that MI is an index sensitive to temperature changes. The MIand its modification, and the trophic diversity value indicated that nematodecommunities are more stable in the northern part of the examined desert than in thesouthern part.
This study demonstrated that under equally low soil moisture and organic mattercontent which can directly and indirectly influence the food source, long-termtemperature effect seems to be the main factor for restricting nematode activity,
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355352

leading to changes in the nematode community structure. Of all the observed trophicgroups, BF was found to be the trophic group with the highest resistance to deserttemperatures.
The bacteria-feeding trophic group, trophic diversity, MI and its modificationwere found to be indicators for studying changes in the desert habitat along aclimatological gradient. Moreover, the modification of MI, including PP nematodes,proved to be more sensitive at the different locations under trees and in the controlsoil samples than the MI (Table 3). This result was the opposite of that obtained byBongers et al. (1997), who believe that the inclusion of plant parasitic into the MIleads to an index less sensitive to changes in the environment than the original MI. Inconclusion, in analyzing the obtained data, we arrived at the conclusion that underdesert conditions, when environmental temperatures reach high values, the influenceof soil moisture and organic matter on the nematode population minimizes,elucidating the temperature as being one of the factors determining communitycomposition and structure.
References
Alon, A., Steinberger, Y., 1999. Response of the soil microbial biomass and nematode population to a
wetting event in nitrogen-amended Negev desert plots. Biology and Fertility of Soils 30, 147–152.
Bakonyi, G., Nagy, P., 2000. Temperature- and moisture-induced changes in the structure of the nematode
fauna of a semiarid grassland—patterns and mechanisms. Global Change Biology 6, 697–707.
Bongers, T., 1990. The maturity index: an ecological measure of environmental disturbance based on
nematode species composition. Oecologia 83, 14–19.
Bongers, T., Ferris, H., 1999. Nematode community structure as a bioindicator in environmental
monitoring. Trends in Ecology & Evolution 14, 224–228.
Bongers, T.H., Van der Muelen, T.H., Korthals, G., 1997. Inverse relationship between the nematode
maturity index and plant parasite index under enriched nutrient conditions. Applied Soil Zoology 6,
195–199.
Cairns, E.J., 1960. Methods in nematology. In: Sasser, J.N., Jenkins, W.R. (Eds.), Nematology,
Fundamentals and Recent Advances with Emphasis on Plant Parasitic and Soil Forms. University of
North Carolina Press, Chapel Hill, pp. 33–84.
Chew, R.M., 1974. Consumers as regulators of ecosystems; an alternative to energetics. Ohio Journals of
Science 72, 359–370.
Dan, J., Yaalon, D.H., Royumdjisky, H., Raz, Z., 1972. The soil association map of Israel. Israel Journal
of Earth Sciences 2, 29–49.
Danin, A., 1983. Desert Vegetation of Israel and Sinai. Cana Publications, Jerusalem, p. 148.
De Goede, R.G.M., Bongers, T., 1994. Nematode community structure in relation to soil and vegetation
characteristics. Applied Soil Ecology 1, 29–44.
Duponnois, R., Neyra, M., Senghor, K., Ba, A.M., 1999. Effects of the root-knot nematode Meloidogyne
javanica on the symbiotic relationships between different strains of Rhizobia and Acacia holosericea
(A Cunn. ex G. Don.). European Journal of Soil Biology 35, 99–105.
Ernst, W.H.O., Tolsma, D.J., Decelle, J.E., 1989. Predation of seeds of Acacia seedlings tortilis by insects.
Oikos 54, 294–300.
Evenari, M.E., Shanan, L., Tadmore, W., 1982. The Negev: The challenge of a desert. Harvard University
Press, Cambridge, MA.
Freckman, D.W., 1988. Bacterivorous nematodes and organic matter decomposition. Agriculture
Ecosystems & Environment 24, 196–217.
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355 353

Freckman, D.W., Mankau, R., 1986. Abundance, distribution, biomass and energetics of soil nematodes
in a northern Mojave Desert ecosystem. Pedobiologia 29, 129–142.
Freckman, D.W., Virginia, R.A., 1997. Low-diversity Antarctic soil nematode communities: distribution
and response to disturbance. Ecology 78, 363–369.
Halevy, G., Orshan, G., 1972. Ecological studies on Acacia species in the Negev and Sinai I. Distribution
of Acacia Raddiana, A. tortilis and A. gerrardi ssp. negevensis as related to environmental factors.
Israel Journal of Botany 21, 197–208.
Heip, C., Herman, P.M.J., Soetaert, K., 1988. Data processing, evaluation and analysis. In: Higgins, R.P.,
Thiel, H. (Eds.), Introduction to the Study of Meiofauna. Smithsonian Institute Press, Washington
DC, pp. 197–231.
Lahav-Ginott, S., Kadmon, R., Gersani, M., 2001. Evaluating the viability of acacia populations in the
Negev Desert: a remote sensing approach. Biological Conservation 98, 127–137.
Liang, W., Pinhasi-Adiv, Y., Shtultz, H., Steinberger, Y., 2000. Nematode population dynamics under the
canopy of desert halophytes. Arid Soil Research and Rehabilitation 14, 183–192.
Liang, W., Lavian, I., Steinberger, Y., 2001. Effect of agricultural management on nematode communities
in a Mediterranean agroecosystem. Journal of Nematology 33, 208–215.
Manlay, R.J., Cadet, P., Thioulouse, J., Chotte, J.L., 2000. Relationships between abiotic and biotic soil
properties during fallow periods in the sudanian zone of Senegal. Applied Soil Ecology 14, 89–101.
Milton, S.J., Dean, W.R.J., 1995. How useful is the keystone species concept, and can it be applied to
Acacia erioloba in the Kalahari Desert? Zeitschrift fuer Okologie Naturschutz 4, 147–156.
Moore, J.C., Ruiter, P.C., 1991. Temporal and spatial heterogeneity of trophic interactions within below-
ground food webs. Agriculture Ecosystems & Environment 34, 371–397.
Oba, G., 1998. Effects of excluding goat herbivory on Acacia tortilis woodland around pastoralist
settlements in northwest Kenya. Acta Oecologica-International Journal of Ecology 19, 395–404.
Perevolotsky, A., Lotan, R., Chamo, Y., 1998. Factors affecting Acacia regeneration in the Northern
Arava. Report to The Nature Reserves Authority, Negev, Israel.
Pielou, E.C., 1975. Ecological Diversity. Wiley, New York, p. 165.
Porazinka, D.L., McSorley, R., Duncan, L.W., Graham, J.H., Wheaton, T.A., Rohner, C., Ward, D.,
1997. Chemical and mechanical defense against herbivory in two sympatric species of desert Acacia.
Journal of Vegetation Science 8, 717–726.
Rohner, C., Ward, D., 1999. Large mammalian herbivores and the conservation of arid Acacia stands in
the Middle East. Conservation Biology 13, 1162–1171.
Rowell, D.L., 1994. Soil Science: Methods and Applications. Longman Group UK Ltd., London, p. 48.
Ruess, R.W., Halter, F.L., 1990. The impact of large herbivores on the Seronera woodlands, Serengeti
National Park, Tanzania. African Journal of Ecology 28, 259–275.
Shannon, C.E., Weaver, W., 1949. The Mathematical Theory of Communication. University of Illinois
Press, Urbana, IL.
Steinberger, Y., Loboda, I., 1991. Nematode population dynamics and trophic structure in a soil profile
under the canopy of the desert shrub Zygophyllum dumosum. Pedobiologia 35, 191–197.
Steinberger, Y., Sarig, S., 1993. Response by soil nematode populations in the soil microbial biomass to a
rain episode in the hot, dry Negev Desert. Biology and Fertility of Soils 16, 188–192.
Steinberger, Y., Liang, W., Savkina, E., Mesh, T., Barness, G., 2001. Nematode community composition
and diversity associated with a topoclimatic transect in a rain shadow desert. European Journal of Soil
Biology 37, 315–320.
Twinn, D.C., 1974. Nematodes. In: Dickinson, C.H., Pugh, G.J.F. (Eds.), Biology of Plant Litter
Decomposition. Academic Press, London, pp. 421–465.
Verschoor, B.C., De Goede, R.G.M., de Hoop, J.W., de Vries, F.W., 2001. Seasonal dynamics and vertical
distribution of plant-feeding nematode communities in grasslands. Pedobiologia 45, 213–233.
Ward, D., Rohner, C., 1997. Anthropogenic causes of high mortality and low recruitment in three Acacia
tree taxa in the Negev desert, Israel. Biodiversity and Conservation 6, 877–893.
Wardle, D.D., Yeates, G.W., Watson, R.N., Nicholson, K.S., 1995. The detritus food-web and the
diversity of soil fauna as indicators of disturbance regimes in agro-ecosystems. Plant and Soil 170,
35–43.
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355354

Wasilewska, L., 1994. The effect of age of meadows on succession and diversity in soil nematode
communities. Pedobiologia 38, 1–11.
Watson, R.T., Zinyowera, M.C., Moss, R.H., 1998. The Regional Impacts of Climate Change: An
Assessment of Vulnerability. A Special Report by IPCC Working Group II. Cambridge University
Press, Cambridge.
Whitford, W.G., 1996. The importance of the biodiversity of soil biota in arid ecosystems. Biodiversity
and Conservation 5, 185–195.
Wiegand, K., Jeltsch, F., Ward, D., Rohner, C., 1998. Decline of the Negev’s Acacias—a spatially explicit
simulation model as an aid for sustainable management. In: Uso, J.L., Brebbia, C.A., Power, H. (Eds.),
Ecosystems and Sustainable Development. Computational Mechanics Publications, Southhampton,
pp. 63–72.
Wiegand, K., Jeltsch, F., Ward, D., 1999. Analysis of the population dynamics of acacia trees in the Negev
desert, Israel with a spatially explicit computer simulation model. Ecological Modelling 117, 203–224.
Yeates, G.W., 1994. Modification and qualification of the nematode maturity index. Pedobiologia 38,
97–101.
Yeates, G.W., Coleman, D.C., 1982. Role of nematodes in decomposition. In: Freckman, D.W. (Ed.),
Nematodes in Soil Ecosystems. University of Texas, Austin, TX, pp. 55–80.
Yeates, G.W., King, K.L., 1997. Soil nematodes as indicators of the effect of management on grasslands in
the New England Tablelands (NSW): comparison of native and improved grasslands. Pedobiologia 41,
526–536.
Yeates, G.W., Newton, P.C.D., Ross, D.J., 1999. Response of soil nematode fauna to naturally elevated
CO2 levels influenced by soil pattern. Nematology 1, 285–293.
Zohary, M., 1972. Flora Palestina Israel. Academy of Sciences and Humanities, Jerusalem.
ARTICLE IN PRESSS. Pen-Mouratov et al. / Journal of Arid Environments 56 (2004) 339–355 355




















