
Soft-walled, monothalamous benthic foraminiferans in the Pacific, Indian and Atlantic Oceans:...
21
Deep-Sea Research I 51 (2004) 33–53 Soft-walled, monothalamous benthic foraminiferans in the Pacific, Indian and Atlantic Oceans: aspects of biodiversity and biogeography Andrew J. Gooday a, *, Saori Hori b , Yuko Todo b , Takuji Okamoto b , Hiroshi Kitazato b,c , Anna Sabbatini d a Southampton Oceanography Centre, DEEPSEAS Benthic Biology Group, Empress Dock, European Way, Southampton SO14 3ZH, United Kingdom b Department of Life and Earth Sciences, Shizuoka University, Ohya 836, Shizuoka 422-8529, Japan c Institute for Frontier Research on Earth Evolution (IFREE), Japan Marine Science and Technology Center (JAMSTEC), 2-15 Natsushimacho, Yokosuka 237-0061, Japan d Istituto di Scienze del Mare, Facolt " a di Scienze, Universit " a degli Studi di Ancona, Via Brecce Bianche, 60131 Ancona, Italy Received 23 August 2002; received in revised form 16 April 2003; accepted 14 July 2003 Abstract Delicate foraminiferans with test walls that are either agglutinated (Order Astrorhizida, Families Saccamminidae and Psammosphaeridae) or composed of organic material (Order Allogromiida) are often an important element of deep-sea benthic assemblages. Many species have soft, flexible walls and, except for a few allogromiids, are monothalamous (single-chambered). We examined quantitative multicorer samples from abyssal (4263–5570 m) water depths in the North and western Equatorial Pacific and a qualitative sample from a hadal site (7800 m water depth) in the Atacama Trench, SE Pacific. Soft-walled foraminiferans made up 29–42% of all complete, ‘live’ (rose Bengal stained) foraminiferans in the >32 mm fractions (0–1 cm sediment layer) of the North Pacific samples and 63% in the western Equatorial Pacific sample. They included the following morphotypes: (i) organic-walled allogromiids with one or occasionally two apertures; (ii) Nodellum- and Resigella-like forms in which the organic wall is generally brownish in colour and the test sometimes divided into more or less well-defined chambers by constrictions or partitions; (iii) flask- like agglutinated saccamminids with one aperture; (iv) oval saccamminids with two apertures; (v) agglutinated spheres which lack obvious apertures (psammosphaerids). In addition, numerous isolated spherical chambers, believed to be fragments derived from much larger komokiacean foraminiferans, were present in some samples and easily confused with saccamminids. At each Pacific station, we recognised 26–55 morphologically distinct forms (morphospecies) in addition to numerous ‘indeterminate’ specimens that could not be categorised on the basis of morphological criteria. A comparison of the Pacific assemblages with existing data on monothalamous assemblages from abyssal and bathyal depths in the Atlantic and Indian Oceans (>63 mm fraction) suggests that some morphospecies have wide geographical ARTICLE IN PRESS *Corresponding author. Tel.: +44-02380-596353; fax: +44-02380-596247. E-mail address: [email protected] (A.J. Gooday). 0967-0637/$ - see front matter r 2003 Elsevier Ltd. All rights reserved. doi:10.1016/j.dsr.2003.07.002
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Soft-walled, monothalamous benthic foraminiferans in the Pacific, Indian and Atlantic Oceans:...

ARTICLE IN PRESS
Deep-Sea Research I 51 (2004) 33–53
*Correspondin
E-mail addres
0967-0637/$ - see
doi:10.1016/j.dsr.
Soft-walled, monothalamous benthic foraminiferans in thePacific, Indian and Atlantic Oceans: aspects of biodiversity
and biogeography
Andrew J. Goodaya,*, Saori Horib, Yuko Todob, Takuji Okamotob,Hiroshi Kitazatob,c, Anna Sabbatinid
aSouthampton Oceanography Centre, DEEPSEAS Benthic Biology Group, Empress Dock, European Way,
Southampton SO14 3ZH, United KingdombDepartment of Life and Earth Sciences, Shizuoka University, Ohya 836, Shizuoka 422-8529, Japan
c Institute for Frontier Research on Earth Evolution (IFREE), Japan Marine Science and Technology Center (JAMSTEC),
2-15 Natsushimacho, Yokosuka 237-0061, Japand Istituto di Scienze del Mare, Facolt "a di Scienze, Universit "a degli Studi di Ancona, Via Brecce Bianche, 60131 Ancona, Italy
Received 23 August 2002; received in revised form 16 April 2003; accepted 14 July 2003
Abstract
Delicate foraminiferans with test walls that are either agglutinated (Order Astrorhizida, Families Saccamminidae and
Psammosphaeridae) or composed of organic material (Order Allogromiida) are often an important element of deep-sea
benthic assemblages. Many species have soft, flexible walls and, except for a few allogromiids, are monothalamous
(single-chambered). We examined quantitative multicorer samples from abyssal (4263–5570m) water depths in the
North and western Equatorial Pacific and a qualitative sample from a hadal site (7800m water depth) in the Atacama
Trench, SE Pacific. Soft-walled foraminiferans made up 29–42% of all complete, ‘live’ (rose Bengal stained)
foraminiferans in the >32 mm fractions (0–1 cm sediment layer) of the North Pacific samples and 63% in the western
Equatorial Pacific sample. They included the following morphotypes: (i) organic-walled allogromiids with one or
occasionally two apertures; (ii) Nodellum- and Resigella-like forms in which the organic wall is generally brownish in
colour and the test sometimes divided into more or less well-defined chambers by constrictions or partitions; (iii) flask-
like agglutinated saccamminids with one aperture; (iv) oval saccamminids with two apertures; (v) agglutinated spheres
which lack obvious apertures (psammosphaerids). In addition, numerous isolated spherical chambers, believed to be
fragments derived from much larger komokiacean foraminiferans, were present in some samples and easily confused
with saccamminids. At each Pacific station, we recognised 26–55 morphologically distinct forms (morphospecies) in
addition to numerous ‘indeterminate’ specimens that could not be categorised on the basis of morphological criteria. A
comparison of the Pacific assemblages with existing data on monothalamous assemblages from abyssal and bathyal
depths in the Atlantic and Indian Oceans (>63mm fraction) suggests that some morphospecies have wide geographical
g author. Tel.: +44-02380-596353; fax: +44-02380-596247.
s: [email protected] (A.J. Gooday).
front matter r 2003 Elsevier Ltd. All rights reserved.
2003.07.002

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–5334
and bathymetric distributions. These wide-ranging morphospecies could provide good targets for future molecular-level
studies addressing species differentiation among soft-shelled, monothalamous deep-sea foraminiferans.
r 2003 Elsevier Ltd. All rights reserved.
Keywords: Abyssal; Allogromiids; North and Equatorial Pacific; Northeast Atlantic; Arabian Sea
1. Introduction
Local species diversity (a diversity) in the deepsea is high for many macro- and meiofaunal taxa(Grassle and Maciolek, 1992; Etter and Mulli-neaux, 2000; Snelgrove and Smith, 2002). Amongthe meiofauna, the foraminiferans are particularlyrich in species (Gooday et al., 1998). Hard-shelled,calcareous foraminiferans are fairly well documen-ted (Douglas and Woodruff, 1981; Murray, 1991)but much less is known about the soft-walled,predominantly monothalamous (single-cham-bered) taxa. These comprise forms in which thetest is composed of either agglutinated particles(e.g. ‘soft-walled saccamminids’ sensu Gooday,1986a and later papers) or organic material(‘allogromiids’ in the traditional sense). Studiesconducted during the last 15 years at bathyal andabyssal sites in the North Atlantic Ocean (Gooday,1986a, b, 1996; Gooday et al., 1998) and IndianOcean (Gooday et al., 2000) have revealed thatmonothalamous foraminiferans often form a sub-stantial component of meiofaunal communities.These delicate organisms are difficult to study
and represent a largely undocumented source ofbiodiversity on the ocean floor. Many deep-seaspecies have small tests that are destroyed orbecome unrecognisable when dried and providetaxonomists with relatively few morphologicalcharacters on which to base species definitions.Recognising the same species in samples fromdifferent regions is often problematic and hence weknow little about their biogeography. Informationabout species ranges is important in order toestablish the extent of diversity at regional scales(Levin et al., 2001a). The lack of knowledge aboutbiodiversity at larger spatial scales in the deep sea(Gray, 2002) make this an urgent task. With theseissues in mind, the aims of the present paper are to(1) give new information on the biodiversity of
soft-walled foraminiferans in the North andEquatorial Pacific and (2) establish the distribu-tion of some easily recognisable morphospecies(species based on morphological characteristics) inthese two areas of the Pacific, at a hadal site in theSE Pacific, and at previously studied sites in theNorth Atlantic and Indian Oceans. A moregeneral aim is to identify morphotypes suitablefor targeting in future studies addressing speciesdiversity at the molecular level. Our results arebased on material that has been processed andanalysed in a consistent way by the same workinggroup.Previous studies of foraminiferans in the abyssal
North Pacific include those of Brady (1884),Cushman (1910, 1911, 1913, 1914, 1915, 1917),Bernstein et al. (1978), Schr .oder et al. (1988),Saidova (1975, 1981, 2000), Tendal and Hessler(1977) and Smith (1973). With the exception ofTendal and Hessler’s (1977) introduction to theKomokiacea, none of these authors treated soft-shelled foraminiferans. An earlier publication(Gooday et al., 2001) provided preliminary de-scriptions of assemblages from one of our NorthPacific sites (Station 2/3).
2. Materials and methods
2.1. Study sites
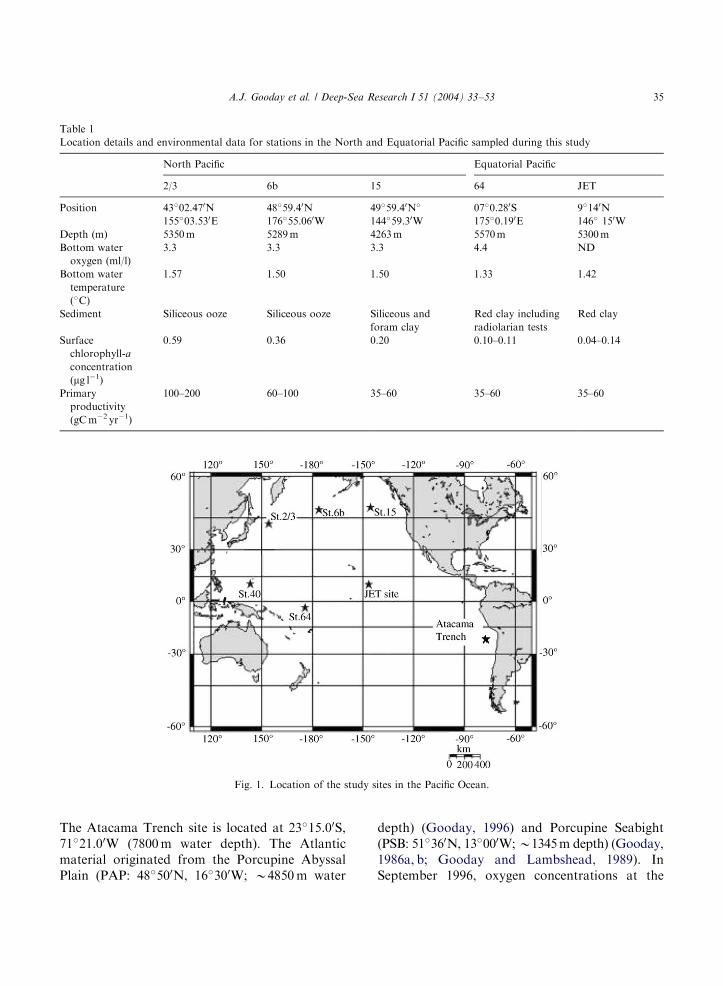
Environmental data for our five Pacific sites aresummarised in Table 1 and their locations shownin Fig. 1. All sites are overlain by oxic bottomwater (O2: 3.3–4.4ml l
�1=148–198 mmol l�1) and,except for Station 15, are situated below thecarbonate compensation depth (CCD). Primaryproductivity and surface chlorophyll-a concentra-tions decrease from west to east in the NorthPacific and are lowest at the central Pacific sites.
ARTICLE IN PRESS
Table 1
Location details and environmental data for stations in the North and Equatorial Pacific sampled during this study
North Pacific Equatorial Pacific
2/3 6b 15 64 JET
Position 43�02.470N 48�59.40N 49�59.40N� 07�0.280S 9�140N
155�03.530E 176�55.060W 144�59.30W 175�0.190E 146� 150W
Depth (m) 5350m 5289m 4263m 5570m 5300m
Bottom water
oxygen (ml/l)
3.3 3.3 3.3 4.4 ND
Bottom water
temperature
(�C)
1.57 1.50 1.50 1.33 1.42
Sediment Siliceous ooze Siliceous ooze Siliceous and
foram clay
Red clay including
radiolarian tests
Red clay
Surface
chlorophyll-a
concentration
(mg l�1)
0.59 0.36 0.20 0.10–0.11 0.04–0.14
Primary
productivity
(gCm�2 yr�1)
100–200 60–100 35–60 35–60 35–60
Fig. 1. Location of the study sites in the Pacific Ocean.
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–53 35
The Atacama Trench site is located at 23�15.00S,71�21.00W (7800m water depth). The Atlanticmaterial originated from the Porcupine AbyssalPlain (PAP: 48�500N, 16�300W; B4850m water
depth) (Gooday, 1996) and Porcupine Seabight(PSB: 51�360N, 13�000W;B1345m depth) (Gooday,1986a, b; Gooday and Lambshead, 1989). InSeptember 1996, oxygen concentrations at the

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–5336
sediment surface at the PAP were B4.4–5.1ml l�1
(=200–230 mmol l�1) (Witbaard et al., 2000). ThePorcupine Seabight site is overlain by bottomwater with an oxygen concentration ofB4.0ml l�1
(=B180 mmol l�1) (Gooday, 2002). Both Atlanticsites are situated above the CCD. The IndianOcean material was collected off the Oman margin(18�59.330N, 58�59.090E; B3400m depth) at a siteabove the CCD and beneath the well-developedoxygen minimum zone where oxygen values arearound 3ml l�1 (=135 mmol l�1) (Gooday et al.,2000). Sediments at the Atlantic and Indian Oceanlocalities are carbonate oozes.
2.2. Sampling and laboratory methods
All samples were obtained with a hydraulicallydampened multiple corer. The North and CentralPacific material was collected during R.V. Haku-
ho-maru Cruises KH-95-2 (July 7–September 27,1995; Station 64) and KH-97-2 (July 9–September8, 1997; Stations 2/3, 6b, 15) (Kawaguchi, 1998;Hori, 2001). Two samples (64A, 64B) fromseparate deployments of the corer were collectedat station 64. The samples from the JET (JapanDeep-Sea Mining Impact Experiment) stationwere collected during JET Cruise I (July 1995).Kajitani (1997) provides an overview of the JETstudy. Samples were collected in the NorthAtlantic between 1989 and 1997 during variouscruises of R.R.S. Discovery, and from the hadalSE Pacific during the Atacama Trench Interna-tional Expedition (September 1997; R.V. Vidal
Gomez). Surface chlorophyll-a measurementswere made at sea during the R.V. Hakuho-maru
cruises (summer 1995, 1997) as follows: 200ml ofseawater were filtered on a glass filter (WhatmanGF/F), decanted for 24 h with 6ml DMF andmeasured with a Turner Design FlourescentPhotometer.The JET station core was subsampled with a
cut-off syringe (diameter 28mm and cross-sec-tional area 6.2 cm2) and the top 1 cm layersliced into the following layers: 0–0.25 cm, 0.25–0.50 cm, 0.50–0.75 cm and 0.75–1.0 cm. At otherPacific stations (2/3, 6a, 15, 64), the entire core(surface area 52.8 cm2) was used and the top 3 cmof the sediment was subdivided into 0.5 cm-
thick layers. Each layer was fixed and stored in4% formaldehyde solution (10% formalin), buf-fered with hexamethylene tetramine. The upper1 cm layer of sediment was examined for thisstudy.In the laboratory, wet sediment layers were split
separately with a plankton splitter into 1/4 splits.One of these splits was then sieved into thefollowing size fractions: >500, 250–500, 125–250,63–125, 32–63 mm (Station 64); >300, 32–300 mm(JET Station), >63, 32–63 mm (Stations 2/3, 6b,15); >500, 150–500, 125–150, 63–125 mm (NorthAtlantic stations); >300, 125–300, 63–125 mm(Indian Ocean). The Atacama Trench samplewas treated with Ludox and sieved on a 20 mmscreen (Sabbatini et al., 2002). Sieve fractionswere stained with rose Bengal and hand-sorted inwater under a binocular microscope for stainedbenthic foraminiferans. Individuals were stored inglycerol in a glass cavity slide and examined indetail under a compound microscope. All photo-graphs were taken on Ilford Delta 100ASAfilm using an Olympus BH-2 photomicroscope.The JET station samples were sorted at theMarine Biological Research Institute of JapanCo Ltd. and the specimens provided to us inglass vials. Since we could not re-examine theoriginal sample residues in order to check thethoroughness with which soft-shelled foraminifer-ans (unfamiliar to non-specialists) had beensorted, this material was used only to establishthe presence of species, not for quantitative countsof specimen numbers.The tests of most of the calcareous foraminifer-
ans (rotaliids and miliolids) were dissolved,leaving only the cell body. The formalin used topreserve the samples was buffered and so dissolu-tion was probably caused by carbonate under-saturation on the seafloor. This made theseparation of species difficult. However, we wereable to discriminate between some species on thebasis of the configuration of the cell body, whichreflects the number, shape and arrangement of thechambers.Diversity indices (Shannon-Wiener, Fisher a)
were calculated with the ‘BioDiversity Pro’ soft-ware (r Natural History Museum, London,Scottish Association of Marine Sciences).
ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–53 37
2.3. Terminology
Recent molecular studies indicate that theOrder Allogromiida (sensu Sen Gupta, 1999)is a polyphyletic group and cannot be separatedfrom monothalamous agglutinated foramini-ferans in the Order Astrorhizida (Pawlowski
Table 2
Abundance, species richness and species diversity (Fisher a index, Sh
North Pacific
2/3 6b 15
Indiv./fraction (1/4)
Total forams 549 738 337
Total monothalamous 202 235 138
Allogromiids 85 95 37
Saccamminids 60 88 52
Psammosphaerids 57 52 49
Indiv./10 cm2
Total forams 416 559 255
Total monothalamous 152 178 104
Allogromiids 64 72 28
Saccamminids 45 67 39
Psammosphaerids 43 39 37
Species numbers
Total forams 85 121 86
Total monothalamous 31 55 33
Allogromiids 14 20 14
Saccamminids 9 22 9
Psammosphaerids 8 13 10
Fisher index
Total forams 30.9 40.2 44
Total monothalamous 11.9 25.2 15
Allogromiids 5.58 8.77 10
Saccamminids 3.24 9.86 3
Psammosphaerids 3.20 6.83 4
Shannon-Wiener (loge)
Total forams 3.91 3.97 3
Total monothalamous 2.93 3.44 2
Allogromiids 2.20 2.60 2
Saccamminids 1.61 2.30 0
Psammosphaerids 1.60 2.17 1
Abundance is given in terms of individual sample fractions (1/4 s
normalised to 10 cm2 surface area. A and B refer to separate sam
allogromiids, saccamminids and psammosphaerids are given sep
polythalamous organic-walled allogromiids.
et al., 2002). In this paper we use ‘allogromiid’in the traditional sense, but only as a convenientlabel for a heterogeneous assemblage oforganic-walled foraminiferans which are mainlymonothalamous but include some polythal-amous taxa (Nodellum, Placopsilinella andResigella).
annon-Wiener) of stained foraminifera in Pacific samples
W. Equatorial Pacific: Stn 64
A B
Complete Fragments
469 337 271
268 212
106 60
64 35
98 117
355 255 205
202 161
80 45
48 27
74 89
56 82 35
26 53
14 22
7 17
5 14
.2 18.5 44.1
.7 7.59 38.1
.6 4.47 13.6
.31 2.18 13.0
.50 1.20 13.1
.98 3.66 4.02
.74 2.78 3.78
.37 2.22 2.89
.99 1.71 2.67
.97 1.06 2.47
plit of 0–1 cm layer of core, 52.8 cm2 total surface area) and
ples from Station 64. Data for total monothalamous taxa,
arately. Note that ‘monothalmous’ taxa also include the
ARTICLE IN PRESS
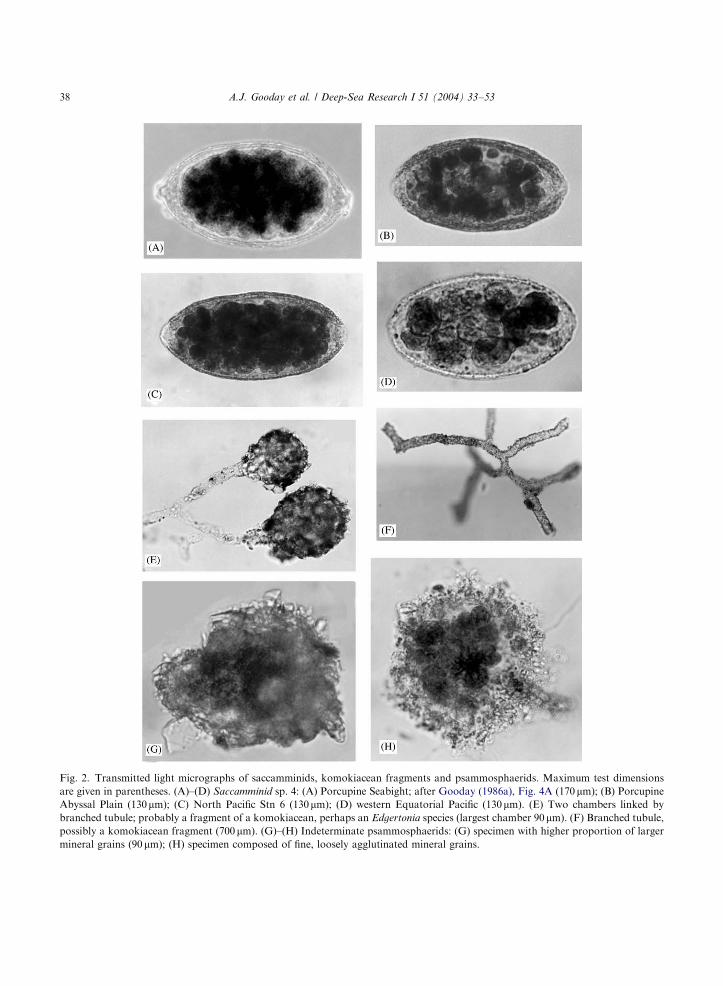
Fig. 2. Transmitted light micrographs of saccamminids, komokiacean fragments and psammosphaerids. Maximum test dimensions
are given in parentheses. (A)–(D) Saccamminid sp. 4: (A) Porcupine Seabight; after Gooday (1986a), Fig. 4A (170 mm); (B) PorcupineAbyssal Plain (130mm); (C) North Pacific Stn 6 (130mm); (D) western Equatorial Pacific (130mm). (E) Two chambers linked by
branched tubule; probably a fragment of a komokiacean, perhaps an Edgertonia species (largest chamber 90mm). (F) Branched tubule,possibly a komokiacean fragment (700 mm). (G)–(H) Indeterminate psammosphaerids: (G) specimen with higher proportion of larger
mineral grains (90mm); (H) specimen composed of fine, loosely agglutinated mineral grains.
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–5338

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–53 39
3. Results
3.1. Population density, taxonomic composition,
and diversity in Pacific samples
The numbers of complete, rose Bengal stainedindividuals (>32 mm sieve fraction; 0–1 cm layer)from the North Pacific stations (Stations 2/3, 6b,15) ranged from 255 to 559 per 10 cm2 (see Table 2for original counts from 1/4 splits). Sample A fromStation 64 in the western Equatorial Pacific yielded355 complete specimens per 10 cm2. Sample Bfrom the same station (>32 mm fraction; 0–0.5 cmlayer only) yielded 255 individuals and 205fragments per 10 cm2. Many of the fragments wereeither tubular (Fig. 2F) or isolated sphericalchambers with tubular extensions. Occasionally,these extensions developed into a branched tubule;in one case, this gave rise to two chambers (Fig. 2E),
Table 3
Gross taxonomic composition of stained foraminiferal assemblages (>
North Pacific
Group Stn 2/3 Stn 6b
Total Indet Total Indet
Allogromiida 15.5 25.9 12.9 18.9
Saccamminidae 10.9 16.7 11.9 6.8
Psammosphaeridae 10.3 38.6 7.0 25.0
Lagenammina 4.6 4.0 5.8 2.3
Tubular forms 5.5 0 0.7 0
Hyperamminacea 0.2 0 1.5 0
Ammodiscacea 0.5 0 2.3 0
Hormosinacea 19.7 10.2 14.6 0
Trochamminacea 19.1 6.7 16.7 0
Other MAFa 8.3 2.2 11.8 0
Other agglutinated taxab 0.4 0 0.3 0
Miliolina 0.7 0 1.4 0
Rotaliina 4.2 34.8 13.1 3.1
Total number 549 82 738 41
All data are from the 0–1.0 cm layer. Under each station, the figures
grouping (taxonomic rank varies) as a percentage of the total sta
indeterminate specimens (i.e. those that could not be assigned to a wor
each taxon or grouping. The large number of indeterminate rotali
calcareous tests were dissolved, making species discrimination proble
final row.aMAF=Multilocular Agglutinated Foraminifera: includes families
and Pseudobolivinidae.bVanhoeffenella spp., Tholosina sp., Astrammina sp.
suggesting that the chambers are fragments of akomokiacean, possibly an Edgertonia species.The general taxonomic composition was
broadly similar at all stations. Soft-shelled taxa(i.e. the predominantly monothalamous allogro-miids, saccamminids, psammosphaerids) ac-counted for 32–41% of individuals at NorthPacific sites and 58–63% at Stn 64 (Table 3).Other numerically important groups were Lagen-
ammina spp., hormosinaceans (Reophax and Hor-
mosina spp.), trochamminaceans and othermultilocular agglutinated taxa. Rotaliids werecommon in some samples. All assemblages werehighly diverse (Table 2), yielding 85–121 morphos-pecies of which 26–55 (36–65%) were soft-shelledmonothalamous forms. Among these, the allogro-miids were usually the most species rich as well asthe most diverse (Fisher and Shannon-Wienerdiversity indices).
32mm) at North and equatorial Pacific sites
W. Central Pacific: Stn 64
Stn 15 Sample A Sample B
Total Indet Total Indet Total Indet
11.0 21.1 22.8 7.55 17.8 8.3
15.4 9.6 13.6 18.7 10.4 0
14.5 24.5 21.0 22.4 34.6 23.9
6.5 13.6 12.0 14.3 13.6 0
3.9 0 0.9 0 6.8 0
1.2 0 0.4 0 0 0
1.5 0 0 0 0.3 0
5.3 16.7 15.3 7.0 5.9 0
10.7 5.6 5.2 0 2.7 11.1
13.6 17.4 1.7 0 2.7 0
0.3 0 2.1 0 0.9 0
0 0 1.3 66.7 0 0
16.0 51.9 3.4 12.5 4.4 6.7
337 69 469 61 337 35
in the left-hand column give the percentage of each taxon or
ined assemblage; the right-hand column gives the number of
king species) as a percentage of the total number of specimens in
ids and miliolids in some samples reflects the fact that many
matic. The total number of stained foraminifera is given in the
Spiroplectamminidae, Lituolidae, Discamminidae, Eggerellidae

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–5340
3.2. Soft-shelled morphotypes and species in Pacific
samples
Most of the soft-shelled forms in our Pacificsamples were monothalamous (single chambered),although organic-walled tests consisting of anumber of more or less well-defined chambersalso occurred. We assigned these forms to sevenmorphological categories (morphotypes), eachcontaining one or more morphospecies. In orderto learn about species ranges, the North andEquatorial Pacific morphospecies were comparedwith those present in samples from the AtacamaTrench (7800m water depth) in the SE Pacific(Sabbatini et al., 2002), the NW Arabian Sea(3400m water depth) (Gooday et al., 2000), thePorcupine Abyssal Plain (4850m depth) andPorcupine Seabight (1345m depth) in the NEAtlantic (Gooday, 1986a, b, 1996). Many of thePacific morphospecies were restricted to individualsamples or areas, but twelve distinctive typesoccurred in more than one ocean (Table 4). Theseprobably represent either single species or groupsof closely related species. They are briefly char-
Table 4
Occurrence of selected soft-shelled foraminiferal morphotypes and m
North Atlantic and Indian Oceans
Morphospecies North Pacific W. Equat. Pacific
2/3 6 15 JET 64
5350m 5289m 4263m 5300m 5570m
Allogromiid sp. 1 a a a a a
?Chitinosiphon sp. — a a a a
Nodellum-like 1 a a a a a
Nodellum-like 2 a a a a a
Nodellum-like 3 — a — a a
Resigella moniliforme a a a a a
Resigella-like 1 a a a a a
Resigella-like 2 — — — — a
Placopsilinella sp. a a a a a
Saccamminid 2 — a — a —
Saccamminid 4 a a — a a
Saccamminid 5 a a a a a
Saccamminid 6 — a — — —
Psammosphaerid sp. 5 a — — — a
Vanhoeffenella spp. a a — a a
Water depth is given for each station. Twelve of the 15 morphospeci
AT=Atacama Trench; PAP=Porcupine Abyssal Plain; PSB=PorcuaPresence.
acterised below and illustrated in Figs. 2–6. Wealso consider the distribution of Vanhoeffenella, ataxon presently classified, on the basis of testmorphology, in the Order Astrorhizida, FamilyAstrorhizidae (Loeblich and Tappan, 1987).
3.2.1. Nodellum-like allogromiids
Disregarding komokiaceans, the majority ofdeep-water, soft-shelled foraminiferans that havebeen described formally are large species withrelatively thick, brownish organic walls. Nodellum
membranacea (Brady, 1879) was reported from theSouth Atlantic and subsequently from the NorthAtlantic Ocean (Brady, 1884; Gooday, 1986a;Schr .oder, 1986) and Pacific Ocean (Saidova,1975; Zheng and Fu, 2001). Saidova (1970) erecteda new genus and species, Xenothekella elongata,for a Nodellum-like form from hadal depths (9170–9335m) in the Kurile-Kamchatka Trench, westernPacific Ocean. This is the deepest-known organic-walled foraminiferan and differs from Nodellum inhaving an unconstricted tubular test. Resig (1982)described Nodellum (=Resigella) moniliforme fromthe abyssal eastern Equatorial Pacific. This species
orphospecies at stations in the North and Equatorial Pacific,
SE Pacific NE Atlantic Indian Ocean Illustration
AT PAP PSB
7800m 4850m 1340m 3400m
— a a — Figs. 5E–Fa a a — Figs. 3I–La a a a Figs. 3A–E
— a a a Figs. 3F–Ha a a a Figs. 4F–J
— — — — Fig. 5C
— a — — Figs. 4A–Ea — — — Fig. 5D
— a a a Figs. 5A–B
— a — — Figs. 6A–Ca a — a Figs. 2A–D
— a a a Figs. 6D–G
— a — — Figs. 6I–J
— — — — Fig. 6Ha a — — Figs. 5H–J
es occur in more than one ocean.
pine Seabight.
ARTICLE IN PRESS
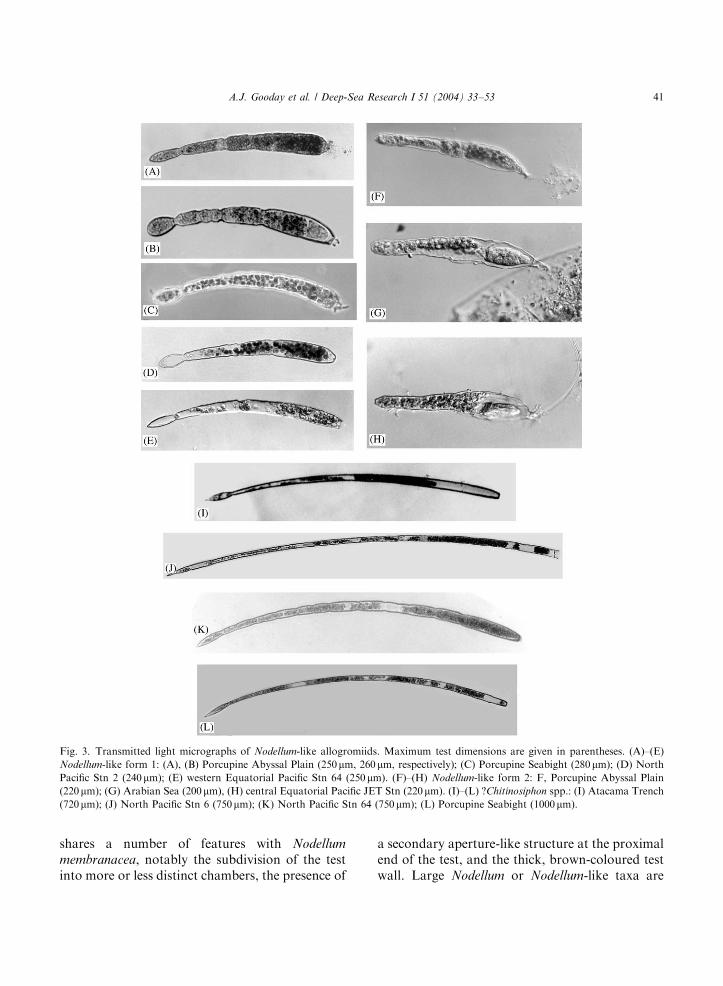
Fig. 3. Transmitted light micrographs of Nodellum-like allogromiids. Maximum test dimensions are given in parentheses. (A)–(E)
Nodellum-like form 1: (A), (B) Porcupine Abyssal Plain (250mm, 260mm, respectively); (C) Porcupine Seabight (280mm); (D) NorthPacific Stn 2 (240mm); (E) western Equatorial Pacific Stn 64 (250mm). (F)–(H) Nodellum-like form 2: F, Porcupine Abyssal Plain
(220mm); (G) Arabian Sea (200mm), (H) central Equatorial Pacific JET Stn (220mm). (I)–(L) ?Chitinosiphon spp.: (I) Atacama Trench
(720mm); (J) North Pacific Stn 6 (750mm); (K) North Pacific Stn 64 (750mm); (L) Porcupine Seabight (1000mm).
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–53 41
shares a number of features with Nodellum
membranacea, notably the subdivision of the testinto more or less distinct chambers, the presence of
a secondary aperture-like structure at the proximalend of the test, and the thick, brown-coloured testwall. Large Nodellum or Nodellum-like taxa are
ARTICLE IN PRESS
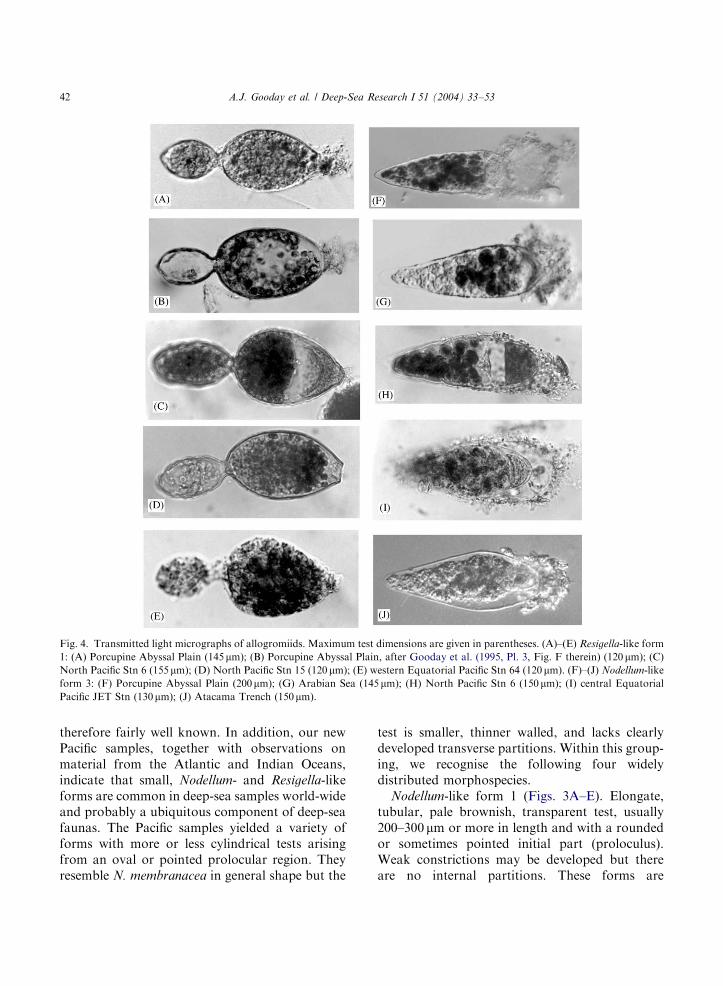
Fig. 4. Transmitted light micrographs of allogromiids. Maximum test dimensions are given in parentheses. (A)–(E) Resigella-like form
1: (A) Porcupine Abyssal Plain (145 mm); (B) Porcupine Abyssal Plain, after Gooday et al. (1995, Pl. 3, Fig. F therein) (120 mm); (C)North Pacific Stn 6 (155mm); (D) North Pacific Stn 15 (120 mm); (E) western Equatorial Pacific Stn 64 (120 mm). (F)–(J) Nodellum-like
form 3: (F) Porcupine Abyssal Plain (200mm); (G) Arabian Sea (145 mm); (H) North Pacific Stn 6 (150 mm); (I) central EquatorialPacific JET Stn (130mm); (J) Atacama Trench (150mm).
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–5342
therefore fairly well known. In addition, our newPacific samples, together with observations onmaterial from the Atlantic and Indian Oceans,indicate that small, Nodellum- and Resigella-likeforms are common in deep-sea samples world-wideand probably a ubiquitous component of deep-seafaunas. The Pacific samples yielded a variety offorms with more or less cylindrical tests arisingfrom an oval or pointed prolocular region. Theyresemble N. membranacea in general shape but the
test is smaller, thinner walled, and lacks clearlydeveloped transverse partitions. Within this group-ing, we recognise the following four widelydistributed morphospecies.
Nodellum-like form 1 (Figs. 3A–E). Elongate,tubular, pale brownish, transparent test, usually200–300 mm or more in length and with a roundedor sometimes pointed initial part (proloculus).Weak constrictions may be developed but thereare no internal partitions. These forms are

ARTICLE IN PRESS
Fig. 5. Transmitted light micrographs of allogromiids and an astrorhiziid. Maximum test dimensions are given in parentheses. (A)–(B)
Placopsilinella sp.: (A) Porcupine Abyssal Plain (150 mm); (B) western Equatorial Pacific Stn 64 (115 mm). (C) R. moniliforme (Resig)
from central Equatorial Pacific JET Stn (330m). (D) Resigella-like form 2 from Atacama Trench (150mm). (E)–(F) Allogromiid sp. 1:
(E) central Equatorial Pacific JET Stn (120mm); (F) Porcupine Seabight, after Gooday (1986a, Fig. 3b therein) (187mm). (G)Allogromiid with two apertures at opposite ends of the test (70mm). (H)-(J) Family Astrorhizidae, Vanhoffenella spp. (H) Porcupine
Abyssal Plain (240 mm); (I) North Pacific, Stn 2/3 (200mm); (J) Atacama Trench (260 mm)
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–53 43
morphologically variable, particularly withregard to the relative length of the test and theshape of the proloculus, and possibly include more
than one species. They occur in all samplesexamined and seem to be common and widelydistributed.
ARTICLE IN PRESS
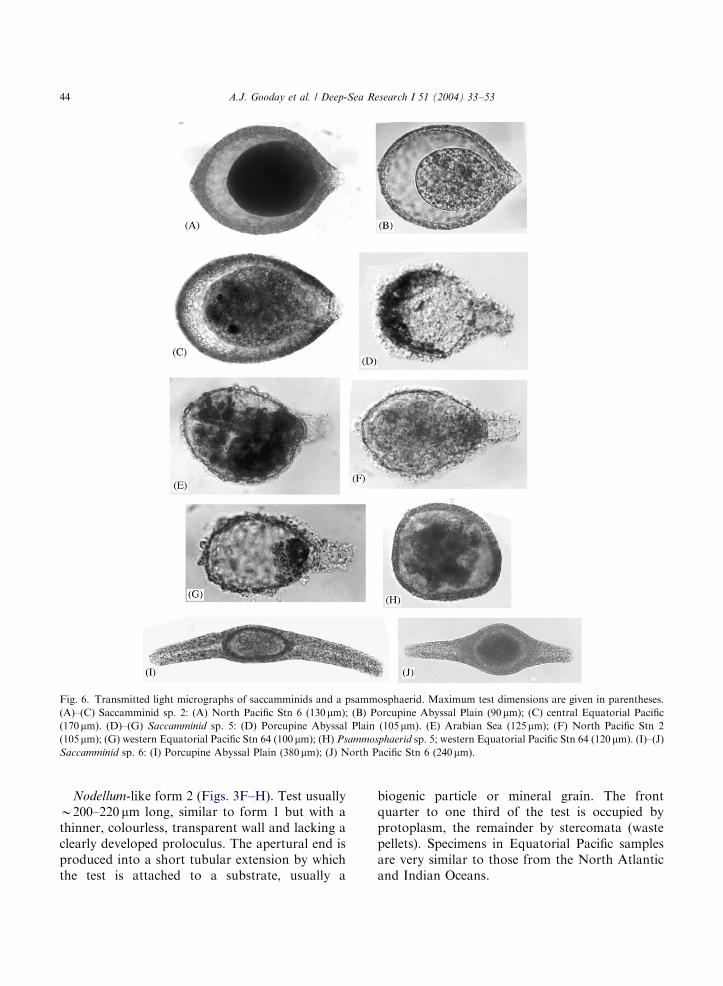
Fig. 6. Transmitted light micrographs of saccamminids and a psammosphaerid. Maximum test dimensions are given in parentheses.
(A)–(C) Saccamminid sp. 2: (A) North Pacific Stn 6 (130mm); (B) Porcupine Abyssal Plain (90mm); (C) central Equatorial Pacific(170mm). (D)–(G) Saccamminid sp. 5: (D) Porcupine Abyssal Plain (105 mm). (E) Arabian Sea (125mm); (F) North Pacific Stn 2
(105mm); (G) western Equatorial Pacific Stn 64 (100 mm); (H) Psammosphaerid sp. 5; western Equatorial Pacific Stn 64 (120mm). (I)–(J)Saccamminid sp. 6: (I) Porcupine Abyssal Plain (380mm); (J) North Pacific Stn 6 (240mm).
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–5344
Nodellum-like form 2 (Figs. 3F–H). Test usuallyB200–220 mm long, similar to form 1 but with athinner, colourless, transparent wall and lacking aclearly developed proloculus. The apertural end isproduced into a short tubular extension by whichthe test is attached to a substrate, usually a
biogenic particle or mineral grain. The frontquarter to one third of the test is occupied byprotoplasm, the remainder by stercomata (wastepellets). Specimens in Equatorial Pacific samplesare very similar to those from the North Atlanticand Indian Oceans.

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–53 45
Nodellum-like form 3 (Figs. 4F–J). Test short,transparent, tubular, 120–200 mm long, widest infront of the mid point and tapering towards theproximal end. The front end is rounded andproduced into a short tubular extension endingin an aperture. Immediately behind this is a small,concentrated mass of cytoplasm with a crescenticor semicircular outline. The remainder of thetest interior is occupied by stercomata. The fronthalf (sometimes the entire test) is covered in a thinlayer of loosely agglutinated material. Thisfeature, which may be some kind of feeding cyst,forms a tent-like canopy over the terminaltube. Specimens from the North and centralPacific (Stns 6, 64, JET), the Arabian Sea andthe abyssal North Atlantic (PAP site) are almostidentical (Figs. 4F–I). A similar form, but with aslightly different shape and therefore possibly adistinct species, occurs in the Atacama Trench(Fig. 4J).?Chitinosiphon spp. (Figs. 3I–L). A very elongate
form, up to 1000 mm or more in length, with apointed proloculus giving rise to a narrow, usuallycurved, needle-shaped tube. The test wall istransparent and typically brownish in colour. Itis fairly common in the Porcupine Seabight(Gooday, 1986a, Fig. 1E therein) and PorcupineAbyssal Plain (NE Atlantic) and occurs occasion-ally in samples from the Equatorial and NorthPacific and the Atacama Trench (7800m)(Sabbatini et al., 2002). Slight differences inthe shape of the proloculus and the relativewidth of the tubular part of the test suggestthat more than one species is represented.Thalmann and Bermudez (1954) establishedChitinosiphon rufescens for a Nodellum-likespecies from Caribbean waters off Cuba (waterdepth 522–3294m), which they compare to theagglutinated genus Bathysiphon. Loeblich andTappan (1964) regarded C. rufescens as asynonym of N. membranacea but the pointedproloculus and tubular rather than subdividedtest suggest that the establishment of a separategenus is justified. Our specimens are much smallerthan C. rufescens (maximum length B1mmcompared to >3mm), lack irregular constrictionsand are therefore placed in Chitinosiphon onlytentatively.
3.2.2. Multilocular allogromiids
Our Pacific samples yielded several chamberedallogromiid species that share certain features withthe Nodellum-like forms described above. Inparticular, the organic test wall is fairly thick andusually brownish in colour, although this is notalways evident in small forms. The Pacific materialcontains three distinct morphospecies, two ofwhich occur in other oceans.
Resigella moniliforme (Resig, 1982) (Fig. 5C).The test is >300 mm in length, has a fairly thick,brown, organic wall and consists of a series ofrounded chambers separated by waist-like con-strictions (Fig. 4D). This species is presentlyunknown outside the Pacific Ocean.
Resigella-like form 1 (Figs. 4A–E). The test(length 120–160 mm) consists of two, occasionallythree, globular chambers joined by a narrow waist-like constriction. The second chamber taperstowards the aperture. As in the case of someNodellum-like forms, the test may be attached by ashort tubular extension to a sediment particle andthere is a small mass of protoplasm just inside theaperture. The remainder of the test is occupied bystercomata. This form occurs in the abyssal NorthAtlantic and in the North and Equatorial Pacific.A similar species (illustrated here as Fig. 5D) iscommon in the Atacama Trench sample (Sabba-tini et al., 2002).
Placopsilinella sp. (Figs. 5A and B). The test islength 140–200 mm long and has a relatively thick,brownish, organic wall and consists of a series ofrounded chambers joined by short stolons. Thechambers are arranged in a more or less linearpattern or in a variety of more irregular patterns.Specimens from the Equatorial Pacific and theAtlantic PAP site are closely similar. Gooday(1986a, Fig. 2 therein) illustrates an attachedspecimen of Placopsilinella from the bathyal NEAtlantic (Porcupine Seabight).
3.2.3. Organic-walled allogromiids with a single
aperture
Allogromiids with a single aperture are commonin deep-sea samples and include a variety ofspherical, oval and elongate forms (Gooday,1986a, 2002). In some cases, the test contains onlyprotoplasm, while in others, stercomata are also

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–5346
present. Our Pacific material yielded at least 12morphospecies of this type. One has been recog-nised in other oceans.
Allogromiid sp. 1 (Figs. 5E–F). A morphologi-cally simple form with an oval, transparent test,120–200 mm in length and a single apertureassociated with a nipple-like protrusion. The testinterior usually contains a mixture of stercomataand stained cytoplasm. This morphospecies is wellknown from the bathyal North Atlantic (Gooday,1986a, Fig. 3B therein) and the abyssal PorcupineAbyssal Plain (Gooday, 1996) and occurs at all ofthe Pacific sites.
3.2.4. Organic-walled allogromiids with two
terminal apertures
These forms are generally oval or sausageshaped and lack stercomata. One deep-waterspecies, Tinogullmia riemanni, is common in theNE Atlantic where it is often associated withphytodetrital deposits (Gooday, 1990). They arerelatively uncommon in the Pacific samples andinclude only three morphospecies, one of which isillustrated in Fig. 4G. None of the Pacific formshas been recognised in other oceans.
3.2.5. Soft-walled saccamminids with one terminal
aperture
Flask-shaped saccamminids are common inboth shallow-water and deep-sea settings. At least15 morphospecies are present in the Pacificsamples, including two that are widely distributed.
Saccamminid sp. 2 (Figs. 6A–C). Test rounded,oval in shape, 100–180 mm long with a clearlydeveloped aperture. The wall is relatively thick andcomposed of flat-lying, plate-like grains thatimpart a silvery highlight when the exterior surfaceis viewed in reflected light. The protoplasmic bodytypically occupies only part of the test interior.This morphospecies is fairly common in the NorthAtlantic and also occurs in the North Pacific (Stn6) and central Pacific (JET Stn). Slight differencesin the shape of the test suggest that more than onespecies may be present.
Saccamminid sp. 5 (Figs. 6D–G). Test flask-shaped, 100–150 mm long, with a clearly developedapertural neck. The wall consists of an organiclayer with a loosely agglutinated outer coating of
variable thickness. The organic layer is continuouswith a septum that appears to form a completeinternal partition across the base of the neck. Apatch of material that stains strongly with roseBengal is usually located immediately inside thisfeature (Fig. 6G). The rest of the test cavitycontains a mixture of stercomata and cytoplasm.This morphospecies occurs at all of our Pacificstations except the Atacama Trench and iscommon in the North Atlantic (illustrated byGooday, 1994, Fig. 3f therein) and Indian Oceans.It seems identical to Hippocrepina sp., reported byHeinz (1999, Pl. 5, Fig. 1 therein) from the bathyal(898–919m) NW Mediterranean (Gulf of Lions,Gulf of Taranto). A similar species from theabyssal Indian Ocean was assigned to Hippocrepi-
na indivisa by Kurbjeweit et al. (2000).
3.2.6. Soft-walled saccamminids with two terminal
apertures
Oval saccamminids with two terminal aperturesare a consistent feature of deep-sea foraminiferalassemblages and may be particularly important intrench samples (Sabbatini et al., 2002). Our Pacificmaterial includes at least ten forms belonging tothis distinctive morphotype. In some cases, the testcontains stercomata in addition to cytoplasmwhile in others only cytoplasm is present. Thefollowing two morphospecies also occur in otheroceans.
Saccamminid sp. 4 (Figs. 2A–D). The test issymmetrical, oval, 100–180 mm long, and taperstowards the rounded or bluntly pointed ends, eachof which is provided with a small, often indistinctaperture. The surface of the test often has a slightspeckly sheen. The interior contains stercomata aswell as cytoplasm. This morphospecies occurs atall the Pacific stations, except Stn 15, and at thePAP and PSB sites in the North Atlantic.
Saccamminid sp. 6 (Figs. 6I–J). This uncommonbut very distinctive form has a central oval capsulefrom either end of which arise two slightly curvedtubular extensions. These two components of thetest are composed of distinctly different kinds ofagglutinated particles. The overall length is 200–400 mm and the central capsule 100–150 mm long.Very similar individuals occur in the North Pacific

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–53 47
(Station 6a) and Atlantic (Porcupine AbyssalPlain) samples.
3.2.7. Psammosphaerids
Small agglutinated spheres without obviousapertures are often an important component ofdeep-sea foraminiferal assemblages. We recog-nised 13 morphospecies in our North and Equa-torial Pacific material. A few are easily recognisedand occur in more than one sample; for example, amorphospecies with a thick, fine-grained wall andheterogeneous contents (Fig. 6H) occurs at Sta-tions 2/3 and 64 (sample B). Many deep-seapsammosphaerids, however, are rather featureless,lack distinctive characters that could be used as abasis for separating species, and are thereforeparticularly problematic. Pacific specimens areoften very small (80–120 mm). The test wall usuallyconsists of a mixture of fine particles and largerquartz grains, but the relative proportion ofdifferent-sized particles varies considerably (Figs.2G–H), making it difficult to define consistentlyrecognisable forms. These indeterminate indivi-duals represent a quarter to a third (23–39%) of allpsammosphaerids in the North and equatorialPacific samples (Table 3).
3.2.8. Vanhoeffenellids
Vanhoeffenella spp. (Figs. 4H–J). Deep-seaspecimens of this well-known and widely distrib-uted genus are generally assigned to a singlespecies, V. gaussi Rhumbler. In our material thetest is 200–260 mm long, very flat with anagglutinated rim and transparent upper and lowermembranes through which the central cell body isvisible. Specimens from the North and EquatorialPacific, the Atacama Trench and the AtlanticOcean are generally small with a leaf-like outlineand two opposing apertures, although somedifferences in shape are evident between thosefrom the Atlantic and Pacific Oceans. Earland(1933) illustrated examples of V. gaussi fromaround South Georgia, in some of which theagglutinated rim is much wider than in Atlanticand Pacific individuals. Wide-rimmed vanhoeffe-nellids were observed recently in samples frombathyal and abyssal depths in the Weddell Sea(Pawlowski and Gooday, unpublished). The mor-
phological variability of Vanhoeffenella suggeststhat several distinct species exist in the deep sea.Earland (1933) already established one species, V.
oculus, based on a single specimen from the SouthGeorgia area that is oval in shape with two small,delicate, organic tubular extensions.
4. Discussion
4.1. Problems in estimating local diversity among
soft-shelled foraminiferans
The study of soft-shelled foraminifera at thespecies level involves substantial difficulties. Asignificant proportion (often >10%) of saccam-minids, psammosphaerids and allogromiids in ourPacific samples could not be separated easily intomorphospecies and therefore were considered tobe ‘indeterminate’ (Table 3). The number ofmorphospecies recognised, and the proportion ofindeterminate specimens, is somewhat subjective,depending partly on the persistence of theresearcher in attempting to organise these ratherfeatureless specimens into consistently recognisa-ble types. These problems are illustrated by sampleB from Station 64 which was sorted very carefullyfor all stained foraminiferans. Of the 608 speci-mens extracted, 337 (55%) were complete indivi-duals and the remainder were considered to befragments. More than a third (1 1 7) of thecomplete tests were agglutinated spheres, of which28 (29%) were regarded as completely indetemi-nate; i.e. we could not place them in any of ourworking morphospecies. Fifty-three individuals(45%) were sorted into four categories on thebasis of the thickness and relative abundance oflarge and small grains in the test wall (Figs. 2Gand H). As explained above, however, each type isvariable and we cannot be certain that theyrepresent distinct species. Only 25 psammosphaer-id specimens (i.e. 21% of all complete agglutinatedspheres) could be confidently assigned to 14working morphospecies.Fragmentation leads to additional problems,
particularly in the case of komokiaceans andtubular taxa (Figs. 2E and F). It is particularlyimportant to distinguish between saccamminids

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–5348
(complete individuals) and what appear to beisolated komokiacean chambers (fragments). Sixtysaccamminid-like chambers with an organic wall,overlain by an agglutinated layer and drawn outon one side into a tubular extension, were presentin sample B from Station 64. The fact that,occasionally, the tubular extensions branchedand linked two chambers together (Fig. 2E),suggests that they were derived from a komokia-cean, probably a species of Edgertonia.
4.2. Abundance and diversity
Despite the difficulties discussed above, we wereable to discriminate between numerous forms inour Pacific samples on the basis of features such astest morphology, the number and characteristicsof the apertures, and the thickness and composi-tion of the test wall. Our results confirm that, inrelative terms, soft-shelled agglutinated and or-ganic-walled foraminiferans, the majority of themmonothalamous, are at least as abundant atabyssal sites in the Pacific Ocean (Table 2) as theyare at similar depths in other oceans (data fromGooday, 1996, 2002; Gooday et al., 1998, 2000;Gooday, unpublished). Our Pacific samplesyielded 28–80 organic-walled allogromiids per10 cm2 (=11–23% of all stained foraminifera)compared to 20–64 per 10 cm2 (=7–18%) at theAtlantic PAP site, 18–29 per 10 cm2 (=8–18%) atother abyssal NE Atlantic sites and 142 per 10 cm2
(=23%) at 3400m depth in the Arabian Sea.Saccamminids and psammosphaerids togethernumbered 76–116 stained individuals per 10 cm2
(19–45% of all stained foraminifera) at the Pacificsites compared to 34–49 per 10 cm2 (=8–15%) atthe PAP, 19–42 per 10 cm2 (=9–21%) at otherabyssal Atlantic sites, and 216 per 10 cm2 (=30%)in the Arabian Sea. Their contribution to totalspecies richness followed a similar pattern. Allo-gromiids were represented by 14–22 species (=16–27% of all foraminiferal species) in the Pacificsamples, 17–24 species (=13–19%) at the PAP,14–24 (=9–16%) at other abyssal NE Atlanticsites, and 37 (=17%) at the 3400m-deep ArabianSea site. The corresponding data for saccamminidsand psammosphaerids combined were 12–35species (=20–38%) at the Pacific sites, 21–34
(=17–26%) at the PAP, 29–37 (=25–30%) atother NE Atlantic sites, and 45 (=21%) at 3400mdepth in the Arabian Sea.These similarities are surprising in view of the
environmental contrasts between different parts ofthe Pacific (Table 1) and particularly between thePacific and Atlantic Oceans (Levin and Gooday,2003). Atlantic and Pacific bottom watermassesare very different. The northeast Atlantic Oceanfloor is bathed by young, well-oxygenated water(6–7ml l�1=270–310 mmol l�1) derived from theGreenland-Norwegian Sea, whereas the NorthPacific lies at the end of the thermohaline conveyerbelt and is therefore occupied by old bottom waterwith an oxygen content of 3–4ml l�1 (135–185 mmol l�1). There are also important differencesin sediment types. The Atlantic sites lie above theCCD and the sediments are calcareous oozes.Three of the four Pacific sites lie below the CCDand the sediments are therefore devoid of carbo-nate. They also contain a higher proportion offine-grained material than the Atlantic sedimentsand a substantial siliceous component, comprisingradiolarian tests and diatom frustules. Our ob-servations suggest that small, soft-shelled mono-thalamous foraminifera are abundant in allabyssal deep-sea areas where bottom water oxygenis not severely depleted.
4.3. Geographical and bathymetric distributions
We agree with Gray (2002) regarding the needfor more studies of deep-sea diversity at largerspatial scales. In order to assess the scale ofregional diversity, it is necessary to establish theranges of individual species. This, in turn, dependson being able to consistently recognise species indifferent samples. While many hard-shelled for-aminifera (e.g. members of the orders Rotaliida,Buliminida, Miliolida and Textulariida) can beeasily identified, the consistent recognition of soft-shelled monothalamous species is more proble-matic. Our study suggests that at least some soft-shelled morphospecies have broad geographicaldistributions at abyssal depths (Table 4). As aresult, the turnover rate of species, and henceregional diversity, may be relatively low, althoughlocal or sample diversity is often high. It remains

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–53 49
to be determined, however, whether these widelydistributed morphospecies constitute single entitiesat the genetic level.Several of the morphospecies in our samples are
particularly notable. The three Nodellum-likeforms (x1–3) are recorded at all or most of ourstations. Saccamminid sp. 5 is found at all sitesexcept the Atacama Trench and is very similar to aform reported from the upper bathyal (B900mwater depth) western Mediterranean (Heinz,1999). Saccamminid sp. 4 is also widely distrib-uted. Needle-shaped specimens assigned to ?Chit-
inosiphon spp. from the bathyal (1345m depth)Porcupine Seabight (NE Atlantic) are very similarto those from hadal depths (7800m) in theAtacama Trench. The same form has beenrecognised recently in the Weddell Sea (Goodayand Pawlowski, unpublished). Many commondeep-water calcareous foraminiferans also havewide geographical ranges (e.g., van Morkhovenet al., 1986; Douglas and Woodruff, 1981; Murray,1991). Widely distributed abyssal agglutinatedforaminiferans were reported by Schr .oder et al.(1988) who identified 38 species, including mem-bers of the genera Hormosina, Reophax, Lagen-
ammina and Rhabdammina, that occurred insamples from the abyssal central North Pacificand Northwestern Atlantic (Nares Abyssal Plain).If true, this would represent a sharp contrast withshallow-water species, many of which have re-stricted geographical ranges (Buzas and Culver,1991; Culver and Buzas, 1995, 1998).There is similar evidence for broad geographical
ranges among some abyssal metazoans. Cosmo-politan distributions are more common amongprotobranch bivalves at water depths >2000m inthe Atlantic Ocean than at shallower sites, possiblybecause conditions are more uniform (Allen andSanders, 1996). Similarly, a decrease in phenotypicvariation occurs among populations of somegastropod species in the NW Atlantic. This mayreflect reduced habitat complexity and selectivepressure at abyssal depths, together with anincrease in the relative (but not absolute) abun-dance of species with planktotrophic development(Etter and Rex, 1990; Rex and Etter, 1990).Genetic divergence also appears to be higher oncontinental slopes than on abyssal plains (Etter
et al., 2000). One factor that probably influencesthe geographical ranges of species is their abilityto disperse (e.g. Scheltema, 1988; Bouchet andWar!en, 1994). For example, nematode species,which have limited mobility, lack dispersive larvaeand complete their life cycle within the sediment,are currently presumed to have narrow ranges(Lambshead, 1993). On the other hand, anincrease in gastropod species ranges with waterdepth in the NW Atlantic is accompanied by anincrease in the relative abundance of species withplanktotrophic larvae (Rex and War"en, 1990; Rexand Etter, 1990). Foraminiferans, particularlysmall taxa, can be dispersed fairly easily bycurrents; juveniles and small adults may beentrained by current velocities of 10–15 cm s�1
(Alve, 1999). Entrainment will be easiest in thecase of epifaunal species, but also possible forinfaunal species that move towards the surface toreproduce (Kitazato et al., 2000). When combinedwith considerable geological longevity (Milleret al., 1992), it seems likely that foraminiferalspecies are able to colonise all parts of the oceanfloor where conditions are suitable (Alve andGoldstein, 2002). Fossil evidence for a geologicallyinstantaneous spread of newly evolved foramini-feran species around the North American con-tinent (Culver and Buzas, 1998) is consistent withan ability to disperse quickly over considerabledistances.Species with very broad bathymetric ranges are
more problematic than those with wide horizontaldistributions. Pressure effects on cell physiologyand biochemistry impose depth limits on manyanimal species (Somero, 1991, 1992). Recenttheoretical studies suggest that speciation occursmore readily along environmental gradients thanin settings without spatial structure (Doebell andDieckmann, 2003). Evidence for substantial popu-lation differentiation among metazoans on con-tinental margins also challenges the integrity ofsuch species. Chase et al. (1998) found thatcontinental slope (o2500m water depth) and rise(>2500m) populations of the bivalve Deminucula
atacellana were genetically distinct over a hor-izontal distance of only 134 km, despite the gooddispersion capabilities of this species. Considerablegenetic differentiation exists within upper bathyal

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–5350
(457–1102m water depth) populations of thegastropod Frigidoalvania brychia on the NWAtlantic slope (Quattro et al., 2001). Examples ofpossible ‘cryptic speciation’ (genetically distinctspecies that are morphologically very similar) arereported from continental slopes; for example,among bathyal (o3200m water depth) popula-tions of the scavenging amphipod Eurythenes
gryllus (France and Kocher, 1996) and fourmolluscan morphospecies from 500–5000m depthin the NW Atlantic (Etter et al., 1999). Theseobservations on metazoans cannot be extrapolatedautomatically to protists which are likely to exhibitgreater intraspecific genetic variation than metazo-ans. Nevertheless, it would be unwise to regardsoft-shelled foraminiferal morphotypes that rangefrom abyssal or hadal to bathyal water depths assingle genetic entities unless there is compellingevidence to do so. The existence of slightmorphological variability among some of ourmorphospecies (or ‘forms’) (e.g. Figs. 3I–L)suggests that they probably include several sepa-rate species. Slight differences also exist betweenotherwise identical calcareous species living atwidely different water depths (Gooday and Alve,2001).While it is possible that some of the soft-shelled
morphospecies are endemic (i.e. confined to aparticular sampling area, ocean basin or ocean)this is difficult to demonstrate conclusively. Thepresence or absence of a species at a particular sitemay reflect environmental controls (Loubere andFariduddin, 1999; Saidova, 2000), or it may simplybe an artifact of undersampling (Glover et al.,2001). Nevertheless, certain allogromiids occurconsistently in the Pacific but not in other oceans.One example is Resigella moniliforme (Fig. 5C), aspecies that was first described by Resig (1982)from 4010m water depth in the eastern EquatorialPacific and occurs at all five of our abyssal Pacificsites (Table 4). Another example may be the smallResigella-like form with elongate chambers (Fig.5D) that is abundant at the Atacama Trench site(Sabbatini et al., 2002) and occurs rarely at Station64 (sample B). Neither of these forms has beenobserved in our Atlantic or Indian Ocean material.Among hard-shelled monothalamous foraminifer-ans, Jugimurammina stellaperuta Zheng, a large,
tubular species with distinctive longitudinal inter-nal ridges, is reported from the NE Pacific(Quinterno, 1995; Levin et al., 2001b, as ?Rhab-
dammina sp.) and from the NW Pacific (Zheng andFu, 2001) but appears to be absent in the Atlantic.Similarly, the Pacific xenophyophore fauna in-cludes numerous species, particularly members ofthe order Stannomida, that are not present in theAtlantic (Tendal, 1972, 1996). The Atlantic andPacific Oceans have different geological historiesand oceanographic characterisitcs and they exhibita number of clear faunal differences at highertaxonomic levels (Levin and Gooday, 2003). Atleast to some extent, biogeographic patternsamong small, soft-shelled species are likely toreflect the unique characteristics of these verydifferent oceans. We suspect that an exhaustivestudy, employing molecular as well as morpholo-gical and morphometric approaches, may revealexamples of endemism among small, soft-shelledforaminiferal species.
5. Conclusions
The systematic study of monothalamous, agglu-tinated and organic-walled, deep-sea foraminifer-ans is in its infancy. These poorly known protists,many of which have soft, delicate tests, present animportant challenge to researchers investigatingthe scale and pattern of biodiversity on the oceanfloor. Small agglutinated spheres (family Psammo-sphaeridae) are particularly difficult to analysebecause they provide few taxonomic charactersand sometimes exhibit considerable morphologicalvariability. Despite these problems, our studysuggests that monothalamous foraminiferans areat least as abundant and diverse in abyssal Pacifichabitats as they are in comparable settings in theAtlantic and Indian Oceans. Resigella- and No-
dellum-like morphotypes are a distinctive elementin all these assemblages. Abyssal monothalamousfaunas often contain cosmopolitan morphospecies.Some of these are probably single genetic entities,but we suspect that future investigations will revealothers to consist of several species that aremorphologically closely similar. These wide-ran-ging morphospecies may therefore provide good

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–53 51
targets for future investigations addressing popu-lation differentiation at the molecular level amongdeep-sea foraminifera. It will also be important forsuch studies to establish whether the converse istrue, i.e. whether morphological divergence existsamong species that are genetically very close.
Acknowledgements
Collaboration between AJG and HK wasfacilitated by a Fellowship from the JapaneseSociety for the Promotion of Science. Part of thisresearch was supported by Grants-in-Aid from theMinistry of Education, Science and Sports ofJapan, Fundamental Research B (Grant numbers11440154 and 14340156). We thank Elisabeth Alveand Jan Pawlowski for their comments on earlierdrafts of the manuscript. This work was influencedby the Working Group on Deep-Sea Biodiversityof the National Center for Ecological Analysis andSynthesis, a centre funded by the NSF (DEB-9421535), the University of California, SantaBarbara, and the State of California.
References
Allen, J.A., Sanders, H.L., 1996. The zoogeography, diversity
and origin of the deep-sea protobranch bivalves of the
Atlantic: the epilogue. Progress in Oceanography 38,
95–153.
Alve, E., 1999. Colonization of new habitats by benthic
foraminifera: a review. Earth Sciences Reviews 46, 167–185.
Alve, E., Goldstein, S.T., 2002. Resting stage in benthic
foraminiferal propagules: a key feature for dispersal?
Evidence from two shallow water species. Journal of
Micropalaeontology 21, 95–96.
Bernstein, B.B., Hessler, R.R., Smith, R., Jumars, P.A., 1978.
Spatial dispersion of benthic Foraminifera in the abyssal
central North Pacific. Limnology and Oceanography 23,
401–416.
Bouchet, P., War!en, A., 1994. Ontogenetic migration and
dispersal of deep-sea gastropod larvae. In: Young, C.M.,
Eckelbarger, K.J. (Eds.), Reproduction, Larval Biology,
and Recruitment of the Deep-Sea Benthos. Columbia
University Press, New York, pp. 98–117.
Brady, H.B., 1879. Notes on some of the reticularian
Rhizopoda of the ‘Challenger’ Expedition. 1. On new and
little known arenaceous types. Quarterly Journal of Micro-
scopical Sciences 21, 31–71.
Brady, H.B., 1884. Report on the Foraminifera dredged by
H.M.S. Challenger during the years 1873–1876: Report of
the Scientific Results of the Voyage of H.M.S. Challenger,
1873–1876. Zoology 9, 1–814, pls 1–115.
Buzas, M.A., Culver, S.J., 1991. Species diversity and dispersal
of foraminifera. BioScience 41, 483–489.
Chase, M.R., Etter, R.J., Rex, M.A., Quattro, J.M., 1998.
Bathymetric patterns of genetic differentiation in a deep-sea
protobranch bivalve, Deminucula atacellana. Marine Biol-
ogy 131, 301–308.
Culver, S.J, Buzas, M.A., 1995. The effects of anthropogenic
habitat disturbance, habitat destruction, and global warm-
ing on shallow marine benthic foraminifera. Journal of
Foraminiferal Research 25, 204–211.
Culver, S.J, Buzas, M.A., 1998. Patterns of occurrence
of benthic foraminifera in time and space. In: Donovan,
S.K., Paul, C.R.C. (Eds.), The Adequacy of the
Fossil Record. Cambridge University Press, Cambridge,
pp. 207–226.
Cushman, J.A., 1910. A monograph of foraminifera of the
North Pacific Ocean. Part. 1, Astrorhizidae and Lituolidae.
Bulletin of the United States National Museum 71 (1),
1–134.
Cushman, J.A., 1911. A monograph of foraminifera of the
North Pacific Ocean. Part. 2, Textulariidae. Bulletin of the
United States National Museum 71 (2), 1–134.
Cushman, J.A., 1913. A monograph of foraminifera of the
North Pacific Ocean. Part. 3, Lagenidae. Bulletin of the
United States National Museum 71 (3), 1–125.
Cushman, J.A., 1914. A monograph of foraminifera of the
North Pacific Ocean. Part. 4, Chilostomellidae. Bulletin of
the United States National Museum 71 (4), 1–46.
Cushman, J.A., 1915. A monograph of foraminifera of the
North Pacific Ocean. Part. 5, Rotaliidae. Bulletin of the
United States National Museum 71 (5), 1–81.
Cushman, J.A., 1917. A monograph of foraminifera of the
North Pacific Ocean. Part. 6, Miliolidae. Bulletin of the
United States National Museum 71 (6), 1–108.
Doebell, M., Dieckmann, U., 2003. Speciation along environ-
mental gradients. Nature 421, 259–264.
Douglas, R.G., Woodruff, F., 1981. Deep-sea benthic for-
aminifera. In: Emiliani, E. (Ed.), The Oceanic Lithosphere.
The Sea, Vol. 7. Wiley, New York, pp. 1233–1327.
Earland, A., 1933. Foraminifera. Part II. South Georgia.
Discovery Reports 7, 27–138, pls 1–7.
Etter, R.J., Mullineaux, L.S., 2000. Deep-sea communities. In:
Bertness, M.D., Gaines, S., Hay, M. (Eds.), Marine
Ecology. Sinauer, Sunderland, MA, pp. 367–393.
Etter, R.J., Rex, M.A., 1990. Population differentiation
decreases with depth in deep-sea gastropods. Deep-Sea
Research 37, 1251–1261.
Etter, R.J., Rex, M.A., Chase, M.C., Quattro, J.M., 1999. A
genetic dimension to deep-sea biodiversity. Deep-Sea
Research I 46, 1095–1099.
Etter, R.J., Rex, M.A., Chase, M.C., Quattro, J.M., Zardus,
J.D., 2000. Population differentiation decreases with depth
in deep-sea protobranch bivalves. In: Abstracts, Nineth

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–5352
Deep-Sea Biology Symposium, June 2000, Galway, Ireland,
p. 23.
France, S.C., Kocher, T.D., 1996. Geographic and bathymetric
patterns of mitochondrial 16S rRNA sequence divergence
among deep-sea amphipods, Eurythenes gryllus. Marine
Biology 126, 633–643.
Glover, A., Paterson, G., Bett, B., Gage, J., Sibuet, M.,
Scheader, M., Hawkins, L., 2001. Patterns in poly-
chaete abundance and diversity from the Madeira
Abyssal Plain, northeast Atlantic. Deep-Sea Research I
48, 217–236.
Gooday, A.J., 1986a. Soft-shelled foraminifera in meiofaunal
samples from the bathyal northeast Atlantic. Sarsia 71,
275–287.
Gooday, A.J., 1986b. Meiofaunal foraminiferans from the
bathyal Porcupine Seabight (northeast Atlantic): size
structure, standing stock, taxonomic composition, species
diversity and vertical distribution in the sediment. Deep-Sea
Research 33, 1345–1373.
Gooday, A.J., 1990. Tinogullmia riemanni sp. nov. (Allogro-
miina; Foraminiferida), a new species associated with
organic detritus in the deep sea. Bulletin British Museum
Natural History, (Zoology) 56, 93–103.
Gooday, A.J., 1994. The biology of deep-sea foraminifera: a
review of some advances and their applications in paleo-
ceanography. Palaios 9, 14–31.
Gooday, A.J., 1996. Epifaunal and shallow infaunal forami-
niferal communities at three abyssal NE Atlantic sites
subject to differing phytodetritus input regimes. Deep-Sea
Research I 43, 1395–1421.
Gooday, A.J., 2002. Organic-walled allogromiids: their occur-
rence, abundance and diversity in marine habitats. Journal
of Foraminiferal Research 32, 384–399.
Gooday, A.J., Alve, E., 2001. Morphological and ecological
parallels between sublittoral and abyssal foraminiferal
species in the NE Atlantic: a comparison of Stainforthia
fusiformis and Stainforthia sp. Progress in Oceanography
50, 261–283.
Gooday, A.J., Lambshead, P.J.D., 1989. Influence of seasonally
deposited phytodetritus on benthic foraminiferal popula-
tions in the bathyal northeast Atlantic: the species response.
Marine Ecology Progress Series 58, 53–67.
Gooday, A.J., Carstens, M., Thiel, H., 1995. Micro- and
nanoforaminifera from abyssal Northeast Atlantic sedi-
ments: a preliminary report. Internationale Revue gesamten
Hydrobiologie 80, 361–383.
Gooday, A.J., Bett, B.J., Shires, R., Lambshead, P.J.D., 1998.
Deep-sea benthic foraminiferal diversity in the NE Atlantic
and NW Arabian sea: a synthesis. Deep-Sea Research II 45,
165–201.
Gooday, A, J., Bernhard, J.M., Levin, L.A., Suhr, S.B., 2000.
Foraminifera in the Arabian Sea oxygen minimum zone and
other oxygen deficient settings: taxonomic composition,
diversity, and relation to metazoan faunas. Deep-Sea
Research II 47, 25–54.
Gooday, A.J., Kitazato, H., Hori, S., Toyofuku, T., 2001.
Monothalamous soft-shelled foraminifera at an abyssal site
in the North Pacific: a preliminary report. Journal of
Oceanography 57, 377–384.
Grassle, J.F., Maciolek, N.J., 1992. Deep-sea species rich-
ness: regional and local diversity estimates from quantit-
ative bottom samples. American Naturalist 139, 313–341.
Gray, J.S., 2002. Species richness of marine soft sediments.
Marine Ecology Progress Series 244, 285–297.
Heinz, P., 1999. Response of deep-sea benthic foraminifera to
simulated phytodetritus pulses under laboratory conditions.
T .ubinger Mikropal.aontolgische Mitteilungen No. 20,
126pp.
Hori, S. (2001). Deep-sea benthic foraminiferal assemblages
at three abyssal areas: comparison among the North
Pacific, Equatorial Pacific and North Atlantic assem-
blages. Unpublished undergraduate thesis of the
Institute of Geosciences, Shizuoka University, 36pp.,
24 pls.
Kajitani, Y., 1997. Summary of the Japanese Environmental
Study for Manganese Nodules Development. In: Pro-
ceedings of International Symposium on Environmental
Studies for Deep-Sea Mining (1997), Tokyo, Japan,
pp. 20–21.
Kawaguchi, K. (Ed.), (1998). Preliminary report of the R/V
Hakuho Maru Cruise KH-97-2. Ocean Research Institute,
University of Tokyo, 157pp.
Kitazato, H., Shirayama, Y., Nakatsuka, T., Fujiwara, S.,
Shimanaga, M., Kato, Y., Okada, Y., Kanda, J., Yamaoka,
A., Masuzawa, T., Suzuki, K., 2000. Seasonal phytodetritus
deposition and responses of bathyal benthic foraminiferal
populations in Sagami Bay: preliminary results from
‘Project Sagami 1996–1999’. Marine Micropaleontology
40, 135–149.
Kurbjeweit, F., Schmiedl, G., Schiebel, R., Hemleben, Ch.,
Pfannkuche, O., Wallmann, K., Sch.afer, P., 2000. Distribu-
tion, biomass and diversity of benthic foraminifera in
relation to sediment geochemistry in the Arabian Sea.
Deep-Sea Research II 47, 2913–2955.
Lambshead, P.J.D., 1993. Recent developments in marine
benthic biodiversity research. Oc!eanis 19, 5–24.
Levin, L.A., Gooday, A.J., 2003. The deep Atlantic Ocean. In:
Tyler, P.A. (Ed.), The Deep Seafloor Ecosystems of the
World (editor-in-chief D.W. Goodall). Elsevier, Amster-
dam, Lausanne, New York, Oxford, Shannon, Singapore,
Tokyo, pp. 111–179.
Levin, L.A., Etter, R.J., Rex, M.A., Gooday, A.J., Smith, C.R.,
Pineda, J., Stuart, C.T., Hessler, R.R., Pawson, D., 2001a.
Environmental influences on regional deep-sea species
diversity. Annual Review of Systematics and Ecology 32,
51–93.
Levin, L.A., Gooday, A.J., James, D.W., 2001b. Dressing up
for the deep: agglutinated protists adorn an irregular urchin.
Journal of the Marine Biological Association of the United
Kingdom 81, 881–882.
Loeblich, A.R., Tappan, H. (1964). Protista 2. Sarcodina chiefly
‘Thecamoebians’ and Foraminiferida. In: Moore, R.C.
(Ed.), Treatise on Invertebrate Paleontology, Part C, 2 vols.
pp. 1–900.

ARTICLE IN PRESS
A.J. Gooday et al. / Deep-Sea Research I 51 (2004) 33–53 53
Loeblich, A.R., Tappan, H., 1987. Foraminiferal Genera and
their Classification. Vol. 1–2. Van Nostrand Reinhold, New
York.
Loubere, P., Fariduddin, M., 1999. Benthic foraminifera and
the flux of organic carbon to the seabed. In: Sen Gupta,
B.K. (Ed.), Modern Foraminifera. Kluwer Academic
Publishers, Dordrecht, Boston, London, pp. 181–199.
Miller, K.G., Katz, M.E., Berggren, W.A., 1992. Cenozoic
deep-sea benthic foraminifera: a tale of two turnovers. In:
Studies in Benthic Foraminifera BENTHOS ‘90, Sendai,
Tokyo University Press, Tokyo, pp. 67–75.
Murray, J.W., 1991. Ecology and Palaeoecology of Benthic
Foraminifera. Longman Scientific and Technical, New
York, 397pp.
Pawlowski, J., Fahrni, J.F., Brykczynksa, U., Habura, A.,
Bowser, S.S., 2002. Molecular data reveal high taxonomic
diversity of allogromiid foraminifera in Explorers Cove
(McMurdo Sound, Antarctica). Polar Biology 25, 96–105.
Quattro, J.M., Chase, M.R., Rex, M.A., Grieg, T.W., 2001.
Extreme mitochondrial DNA divergence within populations
of the deep-sea gastropod Frigidoalvania brychia. Marine
Biology 139, 1107–1113.
Quinterno, P.J., 1995. Quaternary foraminifers from the
Escanaba Trough, Northeast Pacific Ocean. In: Morton,
J.L. et al. (Eds.), Geologic, Hydrothermal, and Biological
Studies at Escanaba Trough, Gorda Ridge, Offshore
Northern California. US Geological Survey Bulletin, 2022,
pp. 337–359.
Resig, J.M., 1982. Nodellum moniliforme, Ammomarginulina
hadalensis and Favocassidulina subfavus, three new species of
Recent deep-water benthic foraminifera. Journal of Paleon-
tology 56, 977–982.
Rex, M.A., Etter, R.J., 1990. Geographical variation and
population differentiation in two deep-sea snails Bentho-
mangelia antonia and Benthonella tenella. Deep-Sea Re-
search 37, 1229–1249.
Rex, M.A., War"en, A., 1990. Planktotrophic development in
deep-sea prosobranch snails from the western North
Atlantic. Deep-Sea Research 29, 171–184.
Sabbatini, A., Morigi, C., Negri, A., Gooday, A., 2002. Soft-
shelled foraminifera from a hadal site (7800m water depth)
in the Atacama Trench (SE Pacific): preliminary observa-
tions. Journal of Micropalaeontology 21, 131–135.
Saidova, Kh. M. (1970). Benthic foraminifers of the Kurile-
Kamchatka Trench area. In: Bogorov (Ed.), Fauna of the
Kurile-Kamchatka Trench and its Environment. Academy
of Sciences of the USSR. Proceedings of the Shirshov
Institute of Oceanology 86, 144–173.
Saidova, Kh. M. (1975). Benthonic Foraminifera of the Pacific
Ocean, 3 vols. P.P. Shirshov Institut Okeanologii, Akade-
miya Nauk SSSR, Moscow.
Saidova, Kh.M., 1981. Recent foraminiferal communities of the
Pacific Abyssal Plains. Oceanology 21, 259–262.
Saidova, Kh.M., 2000. Facial variability of benthic foraminif-
eral communities of the open regions of the ocean.
Oceanology 40, 66–72.
Scheltema, A., 1988. Planktonic and nonplanktonic develop-
ment among prosobranch gastropods and its relationship to
the geographical range of species. In: Ryland, J.S., Tyler,
P.A. (Eds.), Genetics, Reproduction, and Distributions of
Marine Organisms. Olsen & Olsen, Fredensborg, Denmark,
pp. 183–188.
Schr .oder, C.J., 1986. Deep-water arenaceous foraminifera in
the northwest Atlantic Ocean. Canadian Technical Report
of Hydrography and Ocean Sciences 71, 1–191.
Schr .oder, C.J., Scott, D.B., Medioli, F.S., Bernstein, B.B.,
Hessler, R.R., 1988. Large agglutinated foraminifera:
comparison of assemblages from central North Pacific and
western North Atlantic (Nares Abyssal Plain). Journal of
Foraminiferal Research 18, 25–41.
Sen Gupta, B.K., 1999. Systematics of modern Foraminifera. In:
Sen Gupta, B.K. (Ed.), Modern Foraminifera. Kluwer
Academic Publishers, Dordrecht, Boston, London, pp. 7–36.
Smith, P.B., 1973. Foraminifera of the North Pacific Ocean.
United State Geological Survey Professional Publication
766, 1–17.
Snelgrove, P.V.R., Smith, C.R., 2002. A riot of species in an
environmental calm: the paradox of the species-rich deep-
sea floor. Oceanography and Marine Biology 40, 311–342.
Somero, G.N., 1991. Adaptations to high hydrostatic pressure.
Annual Review of Physiology 1992 54, 557–577.
Somero, G.N., 1992. Biochemical ecology of deep-sea animals.
Experimentia 48, 537–543.
Tendal, O.S., 1972. A monograph of the Xenophyophoria.
Galathea Report 12, 7–103.
Tendal, O.S., 1996. Synoptic checklist and bibliography of the
Xenophyophorea (Protista), with a zoogeographical survey
of the group. Galathea Report 17, 79–101.
Tendal, O.S., Hessler, R.R., 1977. An introduction to the
biology and systematics of Komokiacea. Galathea Report
14, 165–194.
Thalmann, H.E., Bermudez, P.J., 1954. Chitinosiphon, a new
genus of the Rhizamminidae. Contributions from the
Cushman Foundation for Foraminiferal Research 5, 53–54.
Van Morkhoven, F.P.C.M., Berggren, W.A., Edwards, A.S.,
1986. Cenozoic cosmopolitan deep-water foraminifera.
Bulletin des Centres de Recherches Exploration-Production
Elf-Aquitaine, Memoire 11, 419pp.
Witbaard, R., Duineveld, G.C.A., Van der Weele, J.A.,
Berghuis, E.M., Reyss, J.L., 2000. The benthic response to
seasonal deposition of phytodetritus at the Porcupine
Abyssal Plain in the North East Atlantic. Journal of Sea
Research 43, 15–31.
Zheng, S., Fu, Z. (2001). Fauna Sinica. Phylum Granuloreti-
culosa, Class Foraminiferea, Agglutinated Foraminifera.
Science Press, Beijing, 788pp., 122 pls.




















