
Sodium-dependent influx of orthophosphate in mammalian non-myelinated nerve
20
J. Physiol. (1976), 260, pp. 667-686 667 With 6 text-figurea Printed in Gréât Britain SODIUM-DEPENDENT INFLUX 0F ORTHOPHOSPHATE IN MAMMALIAN NON-MYELINATED NERVE BY BEATRICE ANNEE, J. FERRERO, P. JIROUNEK, G. J. JONES, A. SALAMIN ANDR. W. STRAUB From thé Department de Pharmacologie, Ecole de Médecine, CH-1211 Genève 4, Switzerland (Received 20 February 1976) SUMMARY 1. The rate of uptake of radïophosphate was measured in desheathed vagus nerves of rabbits mounted in an apparatus where thé incubating solution flowed along thé préparation. 2. Replacement of thé Na of thé Locke by either choline or Tris slowed thé rate of uptake to about 10% of its value in Na; with K it was slowed to 20%; and with Li to 50%. 3. Measurements of thé rate of uptake at différent extracellular phos- phate concentrations showed that in choline-Locke thé influx of phosphate was proportional to thé extracellular phosphate concentration, while in Locke thé rate of inward flow showed a tendency to saturation with in- creasing phosphate concentrations. 4. Extracts of nerves after 45 min exposure to labelled solutions showed for différent phosphate concentrations a slowing of thé labelling of ATP, ADP and creatine-phosphate (CrP) and inorganic phosphate (Pi) when Locke was replaced by choline-Locke. 5. A similar slowing was found when, at 0-2 niM phosphate, thé labelling of thèse compounds was measured at différent times : application of cho- line-Locke reduced thé rate of incorporation of 32P to about 15 % of thé rate in Locke. 6. In préparations that were loaded with radiophosphate and then washed with inactive solution, and thé effluent fractionated, over 90 % of thé radioactivity was found in thé inorganic phosphate fraction. 7. The effiux of 32P showed an initial exponential phase with a time constant of 20-30 min and a much alower one. The slow efflux had a rate constant of 0-0014 min"1 in 0-2 mM external phosphate. 8. Increasing thé external phosphate concentration increased thé efflux. 9. Prolonged incubation in. choline-Locke reduced thé efflux.
Transcript of Sodium-dependent influx of orthophosphate in mammalian non-myelinated nerve

J. Physiol. (1976), 260, pp. 667-686 667With 6 text-figureaPrinted in Gréât Britain
SODIUM-DEPENDENT INFLUX 0F ORTHOPHOSPHATE INMAMMALIAN NON-MYELINATED NERVE
BY BEATRICE ANNEE, J. FERRERO, P. JIROUNEK,G. J. JONES, A. SALAMIN AND R. W. STRAUB
From thé Department de Pharmacologie, Ecole de Médecine, CH-1211Genève 4, Switzerland
(Received 20 February 1976)
SUMMARY
1. The rate of uptake of radïophosphate was measured in desheathedvagus nerves of rabbits mounted in an apparatus where thé incubatingsolution flowed along thé préparation.
2. Replacement of thé Na of thé Locke by either choline or Tris slowedthé rate of uptake to about 10% of its value in Na; with K it was slowedto 20%; and with Li to 50%.
3. Measurements of thé rate of uptake at différent extracellular phos-phate concentrations showed that in choline-Locke thé influx of phosphatewas proportional to thé extracellular phosphate concentration, while inLocke thé rate of inward flow showed a tendency to saturation with in-creasing phosphate concentrations.
4. Extracts of nerves after 45 min exposure to labelled solutions showedfor différent phosphate concentrations a slowing of thé labelling of ATP,ADP and creatine-phosphate (CrP) and inorganic phosphate (Pi) whenLocke was replaced by choline-Locke.
5. A similar slowing was found when, at 0-2 niM phosphate, thé labellingof thèse compounds was measured at différent times : application of cho-line-Locke reduced thé rate of incorporation of 32P to about 15 % of thérate in Locke.
6. In préparations that were loaded with radiophosphate and thenwashed with inactive solution, and thé effluent fractionated, over 90 % ofthé radioactivity was found in thé inorganic phosphate fraction.
7. The effiux of 32P showed an initial exponential phase with a timeconstant of 20-30 min and a much alower one. The slow efflux had a rateconstant of 0-0014 min"1 in 0-2 mM external phosphate.
8. Increasing thé external phosphate concentration increased théefflux.
9. Prolonged incubation in. choline-Locke reduced thé efflux.

668 B. ANNEE AND OTHERS
10. The influx of phosphate was calculated for différent external phos-phate concentrations using thé rate of uptake of radiophosphate measured45 min after thé application of isotope and thé corresponding efflux ratecoefficients and thé intracellular spécifie activities of thé labelled com-pounds.
11. A correction was also introduced for diffusion, using thé 'limitedbiophase model'.
12. The Na-sensitive influx, i.e. thé différence between influx in Lockeand influx in choline-Locke, showed saturation kinetics.
13. A Lineweaver-Burk plot for Na-sensitive uncorrected influxesgave a wmax of 16-7 /<mole/kg wet wt. min and an apparent Km of 0-42 mM ;for thé corrected fluxes thèse values were 18-2 and 0-36.
14. It is concluded that a large part of phosphate influx and some of théphosphate efflux is mediated by a spécifie Na-dependent phosphate trans-port system.
15. This system seems to be présent also in other types of nervoustissues and probably in many types of animal cells.
INTRODUCTION
Préviens experiments (Anner, Ferrero, Jirounek & Straub, 1975)showed that thé influx of orthophosphate into nerve fibres of thé rabbitvagus is slow, much slower than its rate of rnetabolic turnover. Further,thé influx was found to increase with increasing extracellular phosphate,with a tendency to saturation at high external phosphate concentrations.
Observations in other tissues (Chambers, 1963; Harrîson & Harrison,1963) suggest that thé uptake of phosphate is decreased when thé extra-cellular Na is removed, so that it seemed possible that thé influx of phos-phate in nerve may require thé présence of Na ions. Experiments in rabbitvagus (Anner, Ferrero, Jirounek & Straub, 1973a) showed a slowing of théuptake of radiophosphate when Na was replaced by choline. Similarrésulta were obtained in pike olfactory nerve fibres (Straub, Ferrero,Jirounek, Jones & Salamin, 1975). The présent paper describes thé Na-dependent phosphate influx in more détail. In addition, thé efflux ofradiophosphate from thé vagus was measured in Locke and Na-freesolution.
METHODS
The techniques used wore similar to those of thé previous paper (Anner et al.1975). In brief, for measurements of thé rate of uptake of radiophosphate, a de-sheathed cervical vagus nerve of a rabbit was mounted in a narrow polyethylenetube that was perfused with Locko or modified Locke. After a period of équilibra-tion of about 1 hr, a srnall amount of radiophosphate was added to thé perfusion

Na-DEPENDENT PHOSPHATE INFLUX 669fluid and thé radioactivity of a given length of thé préparation arid its surroundingfluid recorded during several hours. The rate of uptake of radiophosphate wasmcasurod from. thèse records.
In a number of experimcnts, after various times of exposure to labelled solution,thé préparations were removed from thé apparatus after a brief wash with phosphateand tracer-free solution. The inorganic phosphate and thé organîc phosphorylatedcompounds were then extracted, separated by chromatography and thé amounts ofeach eompound and its radioactivity were measured as dcscribed previously (Aimeret al. 1975).
For measurements of thé efflux, thé préparations were loaded with radiophos-phate as in thé influx experiments. After a loading period of 1-3 hr thé radioactivesolution was replaced by inactive Locke or modified Locke, which flowed at i ml./min, and thé emuent of thé apparatus collected in glass vials and counted. At théend of thé experiments thé nerves were removed, weighed, dried in an oven at60° C for 1-2 hr, re-weighed and then dissolved in 0-2 ml. concentratcd nitric acid.Distilled water was then added and thé total counts determined, if necessary afterdilution.
Locke contained (mia) : NaCl, 154; KC1, 5-6; CaCl2, 0-9; MgCl2, 0-5; Tris, 10;glucose, 5 and, in addition, Na2HPÛ4 and NaH2PO4; thé phosphate concentrationindicated in thé text refers to thé total concentration of orthophosphate (referredto also as phosphate or PJ. The pH was adjusted with HC1. In choline-Locke,NaCl was replaced by choline chloride and thé phosphates added as K salts with acorresponding réduction in KC1. Tris-Locke and Li-Locke were prepared along thésame lines. K-isethionate-Locke was made up in such a way that its [K] x [Cl]product was near to that of Locke, in order to avoid volume changes of thé prépara-tion. It contained (mai): K, isethionate, 157 (neutralized with a few drops oflN-H3SO4; KC1, 3-2; CaCl2, 0-9; MgCl2, 0-5; Tris, 10; glucose, 5; N-H3SO4 foradjusting thé pH; and KH2PO4 and K2HPO4 according to thé final phosphateconcentration required. In ail solutions thé pH was brought to 7-40 after thé additionof thé phosphates. Unless otherwise stated, thé température was 37° C.
RESULTS
Influx
Effects of replacing Na with différent cations
Fig. 1 shows a record of an experiment where thé uptake of radiophos-phate into a desheathed rabbit vagus was studied. Immediately after théapplication of radioactive Locke there was a rapid increase in thé recordedradioactivity, due to thé filling of thé tube and thé extracellular space.Afterwards there was a slower uptake of radiophosphate which, as shownpreviously (Anner et al. 1975), équilibrâtes with a superficial phosphatefraction with a time constant of about 13min, and with intracellularphosphate with a time constant of about 11 hr at thé external phosphateconcentration used in this experiment (0-6 mM). In thé experiment ofFig. 1 thé uptake in Locke was followed for 15 min; afterwards thé Na ofthé Locke was replaced by choline. When this solution was applied, thé rateof uptake was slowed to about 10%of its value in thé présence of Na. Therate of uptake remained at this low value as long as Na was absent. Upon

670 B. ANNER AND OTHERS
reintroduction of Na thé rate recovered to a value that was slightly higherthan before thé application of thé choline-Locke and later returned toapproximately thé initial value. A similar effect was found when Na wasreplaced by Tris. The slowing was then about thé same as in choline-Locke(Fig. 1). When instead of Locke, K-isethionate-Locke was used, thé rate
Choline K isethionate Tris
20 min
Fig. 1. Effect of N'a withdrawal on rate of phosphate uptake. Record ofradioactivity of préparation and surrounding médium shows filling of tubeand extracellular space after application of labellod Locke (fast upstroke)followed by uptake into rierve fibres. The uptake is reversibly inhibited whenLocke is replaced by choline-Locke, K-isethionate Locke, or Tris-Locke.Return to label-free Locke shows loss of radioactivity from extracellularspace and tube {fast downstroke). Ail solutions coritained 0-6 mM ortho-phosphate, pH 7-40. 50 counta/sec correspond to uptake of 0-5m-mole phos-phate/kg wet nerve; température 37° C.
of uptake was only lowered to about 20% of its value in Locke (Fig. 1).In both cases thé uptake recovered after thé introduction of Locke.
Fig. 2 shows an experiment where thé effects of choline-Locke, K-isethionate-Locke and Tris-Locke were studied in a différent way. In thiscase thé préparation was first perfused with Locke solution, labelledLocke was then applied for 25 min and afterwards normal label-freeLocke. Next, thé préparation was perfused with choline-Locke for about10 min, labelled choline-Locke was then introduced for 25 min and théradiophosphate was washed out with choline-Locke during 5 min. The

Na-DEPENDENT PHOSPHATE INFLUX 671
same procédure was then repeated with K-isethionate-Locke and Tris-Locke. Fig. 2 shows that whenever Na was absent, thé uptake was slowed,thé slowing being largest after replacement of Na with choline or withTris, and somewhat smaller in K-isethionate-Locke. In two experirnentswith Li-Locke, at 0-2 mM phosphate, thé uptake was slowed by 50%.
D mm
10 min
Fig. 2. Uptake of radio phosphate by vagus nerve kept in Locke or in Na-free solutions. Records of radioactivity of préparation and surroundingmédium show filling of tube and extracellular space after application oflabelled solutions (fast upstroke), followed by uptake into nerve fibres, andloss of radioactivity from tube and extracellular space after return tolabel-free solution (fast downstroke). The perfusing fluids were (.4) Locke,(B) choline-Locke, (C) K-isethionate-Locke, (D) Tris-Locke, (E) Locke.The solutions were applied on thé séquence shown; after each record thépréparation was perfused with label-free solution, first for 5 min with thésolution used and then for another 5 min with thé next solution. Ail solu-tions contained 0-2 IÏIM orthophosphate, pH 7-40, 100 counts/sec correspondto uptake of 280 /«noie phosphate/kg wet nerve; température 37° C.
Effect of Na withdrawal at différent extracellular phosphate concentrations
Since thé application of radiophosphate leads to thé labelling of a'superficial' fraction and to influx into thé intracellular compartment (seeAnnereJaZ. 1975), experirnents like thoseof Figs. 1 and 2, though suggesting

672 B. ANNER AND OTHERS
a dependence of phosphate uptake on extracellular Na, do not allow aquantitative estimate of thé Na dependence of thé phosphate influx. Inother experiments therefore thé rate of uptake of radioactivity wasmeasured after 45 min exposure to radiophosphate Locke at a time whenthé 'superficial' fraction was almost completely labelled. Similar experi-ments were then repeated in choline-Locke, for it was found that in thissolution too, a 'superficial' fraction is labelled approximately exponen-tially with a time constant of 15-20 min. Fig. 3, which illustrâtes thé
20
10
1 2 3Extracellular phosphate concentration (mn)
Fig. 3. Phosphate influx at différent extracellular phosphate concentra-tions, in Locke (0) and choline-Locke (O)- Influx was taken as propor-tional to rate of uptake of radiophosphate at 45 min after application ofisotope. The différence between uptake in Locke and uptake in choline-Locke, which corresponds to thé Na-dependent uptake, is also indicated(C)- Bars or diameter of signs indicate S.E., errors for Na-dependent fluxare not indicated; thé lines were drawn by eye. Température 37° C,
results obtained at varions extracellular phosphate concentrations, showsthat at ail concentrations tested, thé phosphate influx in Locke was muchfaster than in choline-Locke. In choline-Locke thé rate of uptake wasalmost proportional to thé extracellular phosphate concentration anddid not show a tendency to approach a constant rate, as in Locke. Bysubstracting thé influx in choline-Locke from thé influx in Locke, thé rateof Na-dependent uptake is found. It shows a pronounced tendency tosaturation with increasing extracellular phosphate (see Fig. 3).

Na-DEPENDENT PHOSPHATE INFLUX 673
Effect of Na-wiihdrawal on labelling of nucleotides
Another démonstration that Na affects thé uptake of radiophosphate isobtained when thé labelling of nucleotides is measured in Locke and incholine-Locke.
In a first séries of experiments nerves were incubated for 40 min inLocke or choline-Locke with différent phosphate concentrations. After a
TABLE 1. Effect of choline-Locke and température on thé labelling of nucleotides,creatine-phosphate and extracted inorganic phosphate, after incubation of 45 min
0-10-20-40-61-01-53-05-0
0-10-20-40-61-01-53-0
0-10-20-40-61-01-53-05-0
Locko37° CA
(m-mole/kg)
0-1020-0760-1500-1270-1480-2250-2420-356
0-0240-0230-0300-0400-0870-0390-060
0-1220-1690-2210-3400-3720-3730-6700-805
Choline-Locke37° C
B(m-mole/
kg)
0-006
0-0100-017
0-1060-126
Locke 5° CC
(m-mole/kg)
Labelled' ATP
0-00230-00260-030
< 0-005
< 0-0013< 0-026< 0-043
'Labelled' ADP
< 0-00070-003 <0-0013
0-005o-oii
0-02G
< 0-0020
< 0-0097<0-019
'Labelled'
0-OG5
0-0580-160
0-5850-410
O'OSO0-1140-0380-052
0-2160-590
BfA
0-079
0-0790-115
0-4380-354
0-213
0-130
0-1250-126
0-430
0-202
0-385
0-1700-430
0-8730-509
0-473
CIA
0-0230-0340-200
< 0-040
<0-160< 0-058
0-091 Mean
< 0-029< 0-057
< 0-050
< 0-248<0-316
0-140 Mean
0-2460-6750-1720-153
0-3220-733
0-384 Mean
Concentrations are given in m-rnole/kg wet wt., for calculation of 'labelled'amounts, see Methods.
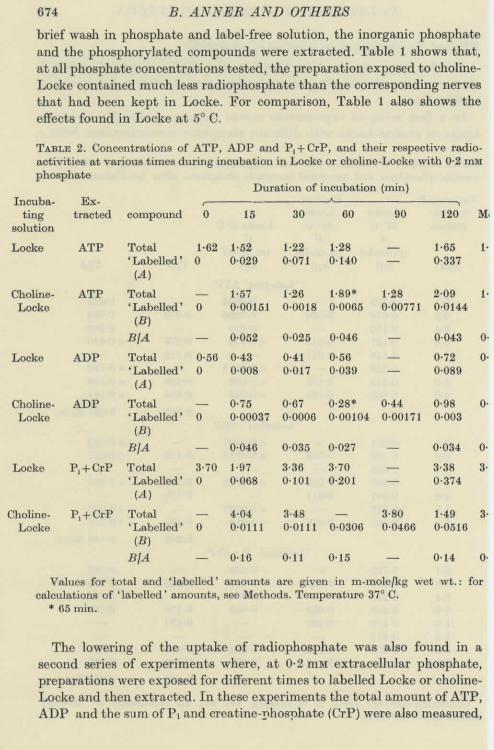
674 B. ANNER AND OTHERS
brief wash in phosphate and label-free solution, thé inorganic phosphateand thé phosphorylated compounds were extracted. Table 1 shows that,at ail phosphate concentrations tested, thé préparation exposed to choline-Locke contained much less radiophosphate than thé corresponding nervesthat had been kept in Locke. For comparison, Table 1 also shows théeffects found in Locke at 5° C.
TABLE 2. Concentrations of ATP, ADP and P, + CrP, and their respective radio-activities at various times during incubation in Locke or choline-Locke with 0-2 mMphosphate
Duration of incubation (min)Incuba-
tingsolution
Locko
Choline-Locke
Locko
Choline-Locke
Locke
Choline-Locke
J-JJV-
tracted
ATP
ATP
ADP
ADP
PI + CrP
PI + CrP
compound
Total'Labelled'
(A)Total' Labelled '
(B)
BIA
Total' Labelled '
(A)Total' Labelled '
(B)
BIA
Total' Labelled '
(A)Total' Labelled '
(B)
BIA
r^0
1-620
—0
—
0-560
—0
—
3-700
—0
_
15
1-520-029
1-570-00151
0-052
0-430-008
0-750-00037
0-046
1-970-068
4-040-0111
0-16
30
1-220-071
1-260-0018
0-025
0-410-017
0-670-0006
0-035
3-360-101
3-480-0111
0-11
60
1-280-140
1-89*0-0065
0-046
0-560-039
0-28*0-00104
0-027
3-700-201
—0-0306
0-15
i90 120
1-650-337
1-28 2-090-00771 0-0144
0-043
0-720-089
0-44 0-980-00171 0-003
0-034
3-380-374
3-80 1-490-0466 0-0516
— 0-14
M.
1-
0-
Values for total and 'labclled' amounts are given in m-mole/kg wet wt.: forcalculations of 'labelled' amounts, sec Methods. Température 37° C.
* 65 min.
The lowerîng of thé uptake of radiophosphate was also found in asecond séries of experiments where, at 0-2 mM extracellular phosphate,préparations were exposed for différent times to labelled Locke or choline-Locke and then extracted. In thèse experiments thé total amount of ATP,ADP and thé sum of Pi and creatine-phosphate (CrP) were also measured,

Na-DEPENDENT PHOSPHATE INFLUX 675
so that thé spécifie activity of thèse compounds could be determined. Theresults are shown in Table 2 and thé spécifie activities are plotted againsttime in Fig. 4. Both Table 2 and Fig. 4 demonstrate thé slowing of labellingwhen Na-free solution was used. The results obtained for ATP are prob-ably thé most reliable, since thé values for ADP are relatively small andthé values for inorganic phosphate are presumably somewhat uncertain,as some of thé membrane-bound phosphate may be included in thisfraction.
20 r
15
£ 10
60 120 0Time (min)
60 120
Fig. 4. Relative spécifie activities of ATP ( + and <>), ADP <; • and Q),and thé suin of Pl and creatine phosphate (0 and Q) extracted from vagiisnerve during incubation in Locke (full symbols) and choline-Locke (opensymbols). Activities wero calculated from Table 2 on assumption that 1radie-phosphate exchanged per molécule of extracted compound. Tem-pérature 37° C.
Our observation, that thé ATP level does not change when Locke is rcplaced bycholine-Locke is somewhat surprising, in view of thé finding of Baker (1965, Table10) that thé intracellular P, concentration increases in crab nerve, after applicationof choline sea water. It must, however, be kept in iniiid that in thé experiments ofBaker, thé nerves were stimulated before thé application of thé Na-free solution, sothat their intracellular Na concentration was high, and thé additional stimulationof thé Na-K pump by thé lowering of extracellular Na could then greatly increasethé ATP utilization. In our experiments, on thé other hand, thé internai Na was notraised by stimulation and began to decrease as soon as choline-Locko was applied.Similarly, Baker (1965) mentions that thé effect of choline sea water was much re-iuced in unstimulated nerves.

676 B, ANNER AND OTHERS
EjfluxEfflux in Locke
In a first séries of expérimenta thé efflux was measured in préparationsthat had been loaded with radiophosphate during exposure to Locke with0-2 IÏÎM phosphate and that were then washed with inactive solution withthé same phosphate concentration. The experiments showed that théefflux first fell rapidly and then much more slowly. A semilogarithmicplot of thé efflux against time showed two fairly distinct exponential
10 r
60 120 180
Time (min)240 300
Fig. 5. Fraction of radioactivity lost (ordinate) from vagus nerve loadedwith radiophosphate and then washed with label-free Locke (0 ) or choline-Locke (O) î abscissa indicates time after beginning of washing. AU solu-tions contained 0-2 mai phosphate. Température 37° C.
phases, one with a time constant for la'belled phosphate loss of 20-30 minand a much slower one. It is probable that thèse phases correspond to thé'superficial' and thé intracellular fractions seen in thé influx experiments.
When thé fraction of radioactivity lost per minute is calculated (seeKeynes & Ritchie, 1965) and then plotted against time (Fig. 5) it can beseen that 1 hr after thé beginning of thé washing thé fraction lost remainsat a fairly constant value of about 0-001 min-1. Since thé main intça-

Na-DEPENDENT PHOSPHATE INFLUX 677
cellular phosphate fractions equilibrate with rate constants of thé order of0-1 min""1 (see Anner et al. 1975), thé fraction lost per minute, which isabout 100 times smaller than this value, can be considered to be thé rateconstant of thé efflux.
In other experiments solutions with 0-02, 0-04, 0-08, 0-6, 1, 2 and 3 mMphosphate were used for thé labelling and thé washout. Thèse experi-ments showed that thé rate of phosphate efflux increased with increasingextracelhilar phosphate (see Table 3). Further, for phosphate concentra-tions above 0-2 mM thé fraction of radioactivity lost per minute tooklonger to approach a fairly steady level than with low phosphate con-centrations.
Our results can be compared with observations in squid axons where thé efflux of32P was measured after a micro-injection of labelled phosphate into thé axoplasm.In thé experiments of Tasaki, Teorell & Spyropoulos (1961) thé initial rate constantof 0-3 x 10~3 min"1 fell after 1 hr to about half; in thé experiments of Caldwell,Hodgkin, Keynes & Shaw (1964) thé initial rate constant of 0-06-0-22 x 10~3 min-1
also decreased considerably during thé washout. In both séries of exporimonts thésurrounding solution was phosphate-froc. Compared to our values, thèse rate con-stants are much larger, îf thé différences in axon diameter and température aretaken into account. On thé other hand, thé falling of thé efflux rate constant, whichthèse authors attribute to thé incorporation of radiophosphate into varions com-pounds, is similar to our observations.
Chromatography of efflux. In some experiments thé efiTuent of théapparatus was collected as described above and then diluted with distilledwater and passed through thé chromatography columns that were usedfor thé séparation of différent phosphate fractions. After elution, morethan 95 % of thé total radioactivity was found in thé fraction that con-tained inorganic phosphate and CrP. Further, thé CrP of thé effluent wasmeasured by an enzymatic-nuorimetric method (see Greengard & Straub,1959). It was found in thèse experiments, kindly performed for us by DrM. Chmouliovsky, that at most 2 % of thé phosphate in thé effluent couldbe accounted for by CrP. In conclusion then, at least 90% of thé radio-phosphate of thé efflux is found in thé form of inorganic phosphate.
Efflux in choline-Locke
Fig. 5 also shows an experiment in which, thé efflux in choline-Lockewas measured. In this and similar expérimenta thé préparations wereloaded with radiophosphate by exposure to Locke. Compared to théefflux in Locke, thé efflux in choline-Locke was first slightly faster andlater definitely slower than in thé présence of sodium. Similar results wereobtained at other phosphate concentrations (Table 3). It is évident fromTable 3 that prolongea exposure to choline-Locke produced a slowing ofthé efflux of phosphate from thé préparation. The immédiate effect of thé

678 B. ANNER AND OTHERS
application of choline-Locke, which is quite différent from thé late slowingdoes not appear to be caused by an increase in intracellular Pi, since wedid not find any change in thé ATP level during exposure to choline-Locke (see Table 2). The transient increase in phosphate efflux is describedin more détail elsewhere (Straub, Ferrero, Jirounek, Jones & Salamin,1976).
Calculation of thé true phosphate influx
Correction for efflux of radiophosphate
As mentioned, thé initial influx of radiophosphate could not be measuredbecause of thé rapid labelling of a 'superficial' fraction. The influxes of
TABLE 3. Measured phosphate influxes, rate constant of phosphate efflux, andcorrected phosphate influxes
Phosphateconcn. ofsolution
(mM)
Measuredphosphate
influx{/(mole/kg. min)
Effluxrate
constant(x l -000 /min) (b)
Relativespécifieactivityafter 45
min incu-bation (fi)
Locke
Relativeefflux ofradio-activity
(/(mole/kg /per min)
Correctedinflux
(/(mole/kg, min)*
(A) (B)
0-020-040-080-20-6123
0-040-20-6123
0-8401-853-034-63
10-7813-2715-7718-74
0-3170-3951-181-392-644-34
0-670-690-831-421-691-571-682-36
0-560-580-620-710-490'49
—
—0-0390-0720-106
—0-129
Choline -Locke—
0-00890-00980-024
—0-084
—
— .0-380-84MB
__2-10
— .0-0360-0420-12__
0-28
— •
—5-01
11-6214-42
— .20-84
—0-4311-171-51
—4-62
0-8661-913-154-93
11-6314-2417-0120-84
0-3250-4051-161-442-74-43
The measured influxes were obtainod from thé rates of uptake of radioactivity at45 min after thé start of incubation in radioactive solution. The efflux rate con-stants are mean values for thé period of efflux from 100 to 250 min. The relativespécifie activities (fi) are mean values for ATP, ADP, P, and CrP, taken fromTable 1, with thé assumptioii that two phosphates of ATP and one phosphate ofADP inay be labelled, and usîng values for thé total arnounts of thèse compoundsgiven in Armer et al. (1975). The relative efflux of radioactivity was calculated fromthé formula fi x b x Pt, where Pt, thé total size of thé internai phosphate pool wastaken as 2 x ATP + ADP + F, + CrP = 6-9 m-mole/kg wet wt. In column A for thécorrected phosphate influx, thé relative effluxes hâve been added to thé measuredinfluxes {sce Text, formula (3)). In columri B, thé corrected influxes wero calculatedas described in thé Text, formula (7 ) .

Na-DEPENDENT PHOSPHATE INFLUX 679
Fig. 3 and thé second column of Table 3 were taken from rates of uptakemeasured when thé labelling of thé 'superficial' fraction was almost com-plète. Thèse rates are smaller than thé initial rates, by an amount corre-sponding to thé loss of radiophosphate. Knowing thé rates of uptake and ofloss, thé exact influx can be calculated.
The net rate of uptake of radioactivity is equal to thé différence between uptakeand loss :
dP* P*
-AT ° ̂ -^E <"{compare eqn. (9) of Hodgkin, 1951), where / and E are thé influx and efflux ofphosphate, respectively, expressed horc as mole/kg wet wt. min; a0 is thé externalspécifie activity; Pt is thé size of thé internai phosphate pool, in mole/kg. Trans-forming eqn. (1) :
1 dP* 1I = -- + _a0 dt a0
where E has been replaced by b x Pt; 6 is thé efflux rate coefficient. If fi is thé relativespécifie activity of thé internai pool of phosphate (Anner et al. 1975), fi — P*/(P txaJ, and:
Phosphate influxes corrected using eqn. (3) are given in column A of Table 3.Alternati vely, eqn. (1) is integrated from thé beginning of thé incubation to thé
time of measu rement. If influx and efflux are constant:
*. (4)f dP* = a0It-~ fJO -^t Jfl
Since thé labellmg of thé internai phosphate pool is almost linear during this timo(sec Fig. 4) :
P'^a^-^P*. (5)Rearranging :
P* =a0/*/(l + J&*). (6)Inserting P*} into eqn. (2):
«0 dt
Influxes caleulatcd using eqn. (4) are givcn in column B of Table 3. The two methodsof ad j usting thé measured phosphate influxes gave almost identical results {Table 3).The advantagc of thé second method was that it could be used for ail phosphateconcentrations.
Correction for diffusion
In order to know thé exact relation between thé extracellular phos-phate concentration and thé phosphate influx, a correction for thé diffu-sion of phosphate between thé bulk of thé solution and thé axonal mem-branes has to be introduced. This correction was made using thé 'limitedbiophase model' (see Rang, 1966; Colquhoun &• Ritchie, 1972; Colquhoun,Henderson & Ritchie, 1972).

680 B. ANNER AND OTHERS
TABLE 4. Corrections for thé diffusion of phosphate in thé vagus nerve
PhosphatePhosphate influxconcn. of from Table 3solution (/«noie/
(mia) kg. min)
extracellular space
0-020-040-080-20-6123
0-040-20-6123
0-8661-913-165-01
11-6214-4217-0120-84
0-3250-4311-171-512-704-62
Phosphateinflux
(/«noie/kg. min)
4-624-765-739-80
11-6610-8311-5916-28
Relativeextracellular
spécifieactivity
Locke0-5330-6890-7870-8430-9310-9610-9780-979
Correctedphosphate
influx{/«noie/kg. min)
1-622-774-005-94
12-4815-0117-3821-27
Calculatedextracellularphosphate
concentration(mM)
0-0310-0480-0870-2150-5970-9841-982-98
Choline- Locke3-864-004-284-903-383-38
0-7320-9290-9740-9810-9940-996
0-4440-4641-201-542-724-64
0-0530-2130-6111-012-003-00
The phosphate influxes in thé first column are taken from Table 3, column A(where available, otherwise column B). The effluxes hâve been calculated as 6 x.Pt,where 6 is thé eflflux rate constant (Table 3) and Pt is thé size of thé internai phos-phate pool, 6-9 m-mole/kg wet wt. (see legend to Table 3). The relative spécifieactivities in thé extracellular space are relative to those in thé external solution andwere calculated using formula (12) of thé Text. Dividing thé phosphate influxesfrom Table 3 by thé relative spécifie activities gave thé corrected influxes in thé fifthcolumn. The average extracellular phosphate concentrations were calculated usingformula (9) of thé Text.
The model postulâtes a single compartment, thé biophase, containing receptorsites, separated from thé external solution by a simple diffusion barrier. We con-sider first thé period of équilibration in Locke with a given phosphate concentration(<70). The concentration in thé biophase (G) is given by:
where k is thé rate constant for exchange between thé biophase and thé externalsolution and F thé volume of thé biophaso. / and E are unknown functions of G, sincethé expérimental values are thé solutions of thé model. However, because thé secondterm is small, thé solution of eqn. (8) is close to an exponentîal with rate constantk. A steady state is attained in a time considerably less than thé équilibration period(see below). Putting thé right-hand side of eqn. (8) equal to zéro:
k(Ctl-C)=-(I-E). (9)

Na-DEPENDENT PHOSPHATE INFLUX 681When radioactive Locke is applied, thé concentration of radioactive phosphate
in thé biophase (C*) is given by:
dC* 1 C*
where thé efflux of radioactivity at thé time of measurement of uptake is ignored (itmakes a small further correction only; cf. Table 3); C* is thé external concentrationof radioactivity and C is given by eqn. (9). The solution of eqn. (10) is an exponentialwith a rate constant which is less than k. A quasi-steady state is reached during théinitial period of incubation when thé concentration of radioactivity in thé biophaseis such that thé net influx from outside is compensated by thé axonal uptake. Put-ting thé left-hand side of eqn. (10) equal to zéro:
1 <7*kiCÎ-O*) = 1. (11)
V C
Eqns. (9) and ( 11 ) are solved for a — C""/C, thé spécifie activity in thé biophase ; and
Corrected influxes are obtained from thé measured influxes by dividing by thé factoron thé right of eqn. (12), using values of E obtained from thé efflux expérimenta.The corresponding biophase phosphate concentrations are then given by eqn. (9).The volume of thé biophase was taken as thé whole of thé extracellular space,0-6 l./kg wet wt. (Keynes & Ritchie, 1965). The time constant for équilibration ofthis space with an external solution is 4-5 min for mannitol, at 20° C (Colquhounet al. 1972). Since phosphate, at 37° C, diffuses about twice as fast as mannitol at20° C, k was taken to be (\ 4-5)-1 = 0-44 min-1.
Eqns. (12) and (9) were used to calculate thé corrected influxes and thé averageextracellular phosphate concentrations for experiments in Locke and choline-Locke(Table 4). For thé latter, thé Na-free solution was applied 10 min before thé labelledsolution. This time was suffïcient to obtaîn a new equilibrium for thé average extra-cellular phosphate concentration.
To test thé correctness of thé calculations, and also tofollowthe time courses of théattainment of steady-state conditions, eqns. (8) and (10) were solved using ananalogue computer. Parameters for thé biophase were as given; thé dependence ofinflux and efflux on thé extracellular phosphate concentration were approximatedby two Michaelis-Menten functions, with parameters adjusted to give a close corre-spondence to thé corrected expérimental values. The steady-state average extra-cellular phosphate concentrations obtained from thé computer simulation were, onaverage, within 5% of those given in Table 4; thé steady states were obtained intimes close to 10 min, thé time needed for almost complète équilibration of théextracellular space with thé external solution by simple diffusion with rate constant0-44 min"1. The correction factors for thé influxes obtained from thé computersimulation were within a few per cent of those given in Table 4.
The biophase may not correspond with thé whole of thé extracellular space ; itmay be restricted to thé mesaxonal space. The effect of thé biophase, however, isdetermined by thé value of thé fraction IjkV. For thé total extracellular space, thisfraction is (0-44 x 0-6)"1 = 3-79 min.kg wet wt. l."1. The mesaxonal space lias amuch smaller volume, about 1 % of thé total nerve volume, but thé time constantfor équilibration of this space with thé rest of thé extracellular space is of théorder of milliseconds (Greengard & Straub, 1958). Thus 1/fcF is of thé order of

682 B. ANNER AND OTHERS(1-OOOxO'Ol) 1 = 0-1 min.kg wet wt. l."1, and correction for diffusion into thémesaxonal space is at least an order of magnitude smaller than that for thé totalextracellular space.
Fig. G A shows a plot of thé expérimental data for thé influxes; thé dataobtained after correction for efflux; and thé data calculated with correc-tions for both efflux and diffusion. The Na-dependent influx was analysedby a Lineweaver-Burk plot (Fig. 6_B), which shows a reasonably good fitto a straight line for thé expérimental data. This conclusion was thé samefor thé corrected data, though thé points fell close to a différent straight
v
20 r
10
50
Fig. 6. A, expérimental and corrected phosphate fluxes at différent phos-phate concentrations, in Locke (full symbols) and choline-Locke (opensymbols). Expérimental values (% and O) wore replotted from Fig. 3.Influxes, v, in /^mole/kg . min after correction for efflux of radioactivity(A and A), and after correction for diffusion (• and Q) are taken fromTables 3 and 4. Continuous and dashed curves are plots of thé function
for corrected and expérimental influxes, rospectively, where G is thé phos-phate concentration. The first terni represents saturable, Na-dependentinflux ; maximum velocities, v^^ and apparent dissociation constant Km
were obtained from Fig. 6B. The second, linear, term represents Na-independent influx; constant A wasl-5xlO~3 and 1-43 xlO~3 1. /kg. wetwt. min, respectively. Symbols are omîtted when thoy overlap with expéri-mental data points. B, Lineweaver-Burk plot of Na-dependent fraction ofphosphate influx, for expérimental data (O) and values corrected forofflux and diffusion ( f] ). The lines were drawn by hand. Maximal velocitiesare 16-7 and 18-2 /«noie /kg. wet wt. min for thé expérimental and correcteddata, and apparent dissociation constants 0-42 and 0-36 mai, respectively.Symbols for corrected data are omitted at high phosphate concentrations,whore they overlap with expérimental data points.

Na-DEPENDENT PHOSPHATE INFLUX 683line (Fig. 6B), The maximum velocities and apparent dissociation con-stants obtained from thé inverse plots were used to construct thé curvesshown in Fig. 6^4 ; thé influxes in Na-free solution were taken to increaselinearly with thé phosphate concentration. Fig. §A and B show that théphosphate influx has two cornponents, a linear, Na-independent com-ponent, and a Na-dependent, saturable component with simple Michaelis-Menten kinetics. The resuit of applying thé corrections described aboveis only to change slightly thé values of thé parameters of thèse twocomponents.
DISCUSSION
The finding that thé rate of uptake of radiophosphate is greatly de-creased in rabbit vagus when thé extracellular Na is replaced by choline,Tris, Li or K suggests that a large proportion of thé phosphate influx dé-pends on thé présence of extracellular Na. For thé replacement of Na bycholine thé argument can be carried further, for in tins case, thé labellingof intracellular nucleotides was also decreased, and in separate experi-ments it was found that, far from increasing thé efflux of phosphate(which could hâve caused a decrease in thé rate of uptake), thé phosphateefflux was actually reduced by prolonged exposure to choline. When K orLi was used, thé decrease in uptake was smaller than when Na was re-placed by choline or Tris, suggesting that K or Li ions might to some ex-tent replace Na ions for thé influx of phosphate.
The Na dependence of phosphate uptake has also been observed inother types of nervous tissue. A lowering of phosphate uptake after re-placing Na by choline was found in pike olfactory nerve (Straub et al.1975), and phosphate influx measurements based on analysis of extrudedaxoplasm showed that in squid giant axons thé phosphate influx is re-duced to about 1/10 when Na is replaced by choline or Li (P. C. Caldwell,Personal communication). The Na dependence of phosphate influx mayalso explain thé observation of Abood & Koyama (1963), who found a de-crease of phosphate incorporation into ATP in desheathed bullfrog sciaticnerve in Ringer with high K (presumably obtained by replacing Na by K).
The Na-dependent phosphate uptake appears to be also présent inother tissues. The decrease in phosphate uptake observed by Chambers(1963)inseaurchineggsandby Harrison & Harrison (1963) in rat intestineon removal of external Na hâve already been mentioned. Moreover, théexperiments of Siegenthaler, Belsky & Goldstein (1967) in a marinefungus showed a large decrease in phosphate uptake when NaCI was re-placed by KC1 or LiCl and more recently it has been suggested that théphosphate absorption in rat rénal proximal convoluted tubule (Baumann,Rouffignac, Roinel, Rumrich & Ullrich, Ï975), and thé phosphate uptake

684 B. ANNEE AND OTHERS
in cultured tumour cells (Brown & Lamb, 1975) are Na-dependent trans-port processes. Thèse observations then give ground to belief that Na-dependent phosphate transport is présent in a gréât variety of tissues.
It is therefore of interest to study this transport System in more détail.The relation between influx and extracellular phosphate shows that théSystem is saturable. Further it is known that phosphate transport is in-hibited by arsenate, as shown for instance in sea urchin eggs by Chambers& Whiteley (1966) and in rabbit vagus in our earlier studies (Armer,Ferrero, Jirounek & Straub,19736), while other anions (e.g. nitrate andsulphate, unpublished experiments) do not interfère to any gréât extent.
At présent it is not clear whether thé phosphate transport System isdifférent from thé Na-dependent transport System for sugars or amino-acids (see Schultz & Curran, 1970), but in view of thé spécifie effect ofarsenate on phosphate fluxes thé phosphate transport appears to bemediated by a sépara te System.
From thé experiments of Table 3 it is clear that Na ions affect not onlythé influx, but that thé phosphate efflux is also dépendent on thé présenceof Na ions. The lowering of thé efflux found during exposure to Na-freesolution could be due to thé absence of extracellular Na, but more likely,as suggested by thé slow development of thé lowering of thé efflux, a de-crease in intracellular Na may be responsible for thé decrease in phosphateefflux. Such a decrease of thé Na content has been found in this tissueduring exposure to Na-free solution (Wespi, 1968). A more elaborate studyon thé relation between intracellular PI, Na, and phosphate efflux, how-ever, still needs to be done.
A Lineweaver-Burk plot of thé expérimental data for thé Na dépendentinflux gives a fairly straight line with a vmax of 16-7 /^mole/kg wet wt. minand a Km of 0-42 mM; with corrections for efflux and diffusion thé corre-sponding values are 18-2 and 0-36. Thèse data apply to thé steady-stateconditions : when sudden changes in thé extracellular solutions are pro-duced, thé efflux markedly déviâtes from thé steady-state condition. TheNa-dependent transport System may thus be more complicated than asimple enzymatic reaction (see, for example, Schultz & Curran, 1970). Themain complication probably cornes from thé fact that thé fluxes not onlydépend on thé extracellular phosphate and Na concentration but also,because of backward reactions, on thé internai concentrations of phos-phate and Na. The influence of internai Na is suggested by thé efflux ex-periments, and a combination of both Na-dependent and passive effluxcan be seen when thé transient effects of changes in thé external Na on théefflux are studied (see Ferrero, Jirounek, Jones, Salamin & Straub, 1976;Straub et al. 1976).

Na-DEPENDENT PHOSPHATE INFLUX 685
We are gratoful to Mrs M. Moosmayer for compétent technical assistance through-out tlie experiments and to thé S.N.S.F. for grants nos. 3.0890.73 and 3.478.0.75,
REFERENCES
ABOOD, L. G. & KOYAMA, I. (1965). Phosphate incorporation in desheathed nerves:effocts of potassium and calcium ions. Science, N.Y. 141, 1277-1278.
ANNER, B., FERRERO, J., JIROUNEK, P. & STRAUB, R. W. (1973a). Inhibition ofintracellular orthophosphate uptake in rabbit vagus nerve by Na withdrawal andlow température. J. Physiol. 232, 47-487*.
ANNEB, B., FEBRERO, J., JIBOUNEK, P. & STRAUB, R. W. (19736). Na-dependentphosphate influx into mammalian iiorve fibres. Experientia 29, 740.
ANNER, B., FERRERO, J., JIBOUNEK, P. & STRAUB, R. W. (1975). Uptake of ortho-phosphate by rabbit vagus nerve nbre_s. J. Physiol. 247, 759-771.
BAKER, P. F. (1965). Phosphorus metabolism of intact crab nerve and its relationto tho active transport of ions. J. Physiol. 180, 383-423.
BAUMANN, K., DE ROUFFIGNAC, C., ROINEL, N., RUMRICH, G. & ULLRICH, K. J.(1975). Rénal phosphate transport: inhomogeneity of local proximal transportrates and sodium dependence. P/lugers Arch. ges. Physiol. 356, 287-297.
BROWN, K. D. & LAMB, J. F. (1975). Na-dependont phosphate transport in cul-tured cells. J. Physiol. 251, 58-59P.
CALDWELL, P. C-, HODGKIN, A. L., KEYNES, R. D. & SHAW, T. I. (1964). The rateof formation and turnover of phosphorus compounds in squid giant axons.J. Physiol. 171, 119-131.
CHAMBERS, E. L. (1963). Rôle of cations in phosphate transport by fertilized seaurchin eggs. Fedn Proc. 22, 331.
CHAMBERS, E. L. & WHITELEY, A. H. (1966). Phosphate transport in fertilized seaurchin eggs. J. cell. comp. Physiol. 68, 289-308.
COLQUHOUN, D., HENDERSON, R. & RITCHIE, J. M. (1972). Tho binding of labelledtetrodotoxin to non-myelinated nerve fibres. J. Physiol. 227, 95-216.
COLQUHOUN, D. & RITCHIE, J. M. (1972). The kinetics of thé interaction betweentetrodotoxin and mammalian non-myelinated nerve fibres. Molec. Pharmacol. 8,285-292.
FERRERO, J., JIROUNEK, P., JONES, G. J., SALAMIN, A. & STRAUB, R. W. (1976).Effects of ions and température on thé efflux of orthophosphate from rabbitvagus nerve fibres. J. Physiol. 254, 64-65.P.
GREENGARD, P. & STRAUB, R. W, (1958). After-potentials in mammalian non-myelinated nerve fibres. J. Physiol. 144, 442-462.
GREENGARD, P. & STRAUB, R. W. (1959). Effect of frequency of electrical stimula-tion on thé concentration of intermediary metabolites in mammalian noii-myelinated fibres. J. Physiol. 148, 353-361.
HARRISON, H. E. & HARRISON, H. C. (1963). Sodium, potassium, and intestinaltransport of glucose, 1-tyrosine, phosphate, and calcium. Am. J. Physiol. 205,107-111.
HODGKIN, A. L. (1951). The ionic basis of electrical activity in nerve and muscle,Biol. Rev. 26, 339-409.
KEYNES, R. D. & RITCHIE, J. M. (1965). The movements of labelled ions in mam-malian non-myelinated nerve fibres. J. Physiol. 179, 333-367.
RANG, H. P. (1966). The kinetics of action of acetylcholine antagonists in smoothmuscle. Proc. R. Soc. B 164, 488-510.
SCHULTZ, S. G. & CURRAN, P. F. (1970). Coupled transport of sodium and organicsolutions. Physiol. Rev. 50, 637-718.

686 B. ANNEE AND OTHERS
SIEOBNTHALER, P. A., BELSKY, M. M. & GOLDSTEIN, S. (1967). Phosphate uptakeinanobligately marine fungus: a spécifie requirement for sodium.Science, N. Y. 155,93-94.
STRA.UB, R. W., FERRERO, J., JIKOUNEK, P., JONES, G. J. & SALAMIN, A. (1975).Na-dependent transport of orthophosphate in vertebrate non-myelinated nervesat différent pH. Abstr. 6th Int. Congr. Pharmacol., Helsinki, p. 367.
STRAUB, R. W., FERRERO, J., JIROUNEK, P., JONES, G. J. & SALAMIN, A. (1976).Transmembranal transport of inorganic phosphate and its implication in somediseases. In Membranes and Diseases, éd. BOLIS, L., HOFFMAN, J. F. & LEAF, A.New York : Raven Press.
TASAKI, I., TEOEELL, T. & SPYBOPOULOS, O.S. (1961). Movement of radioactivetracers across squid axon membrane. Am. J. Physiol. 200, 11-22.
WESPI, H. H. (1968). Transport actif et mouvements passifs de potassium, sodiumet lithium dans des fibres non-myelinisées de lapin. Thèse No 3117, Faculté deMédecine, Université de Genève.




















