
Chapter 4. The hypothesis of eolian loess origin - the physical aspects

ELSEVIER Geoderma 76 (1997) 221-252
C, EODEI~MA
Small scale secondary C a C O 3 accumulations in selected sections of the European loess belt.
Morphological forms and potential for paleoenvironmental reconstruction
J. Becze-Defi.k ", R. Langohr ~', E.P. Verrecchia b.* " Soil &'ience Unit. Unit'ersit3" of Ghent. Krijgslaan 281. 9000 Ghent. Belgium
h IJMR 5561 CNRS. Universit~ de Bourgogne. 6. Bd Gabriel. 21(XX) Dijon. France
Received 28 November 1995: accepted 16 December 1996
Abstract
Eighteen important Quaternary loess paleosoil sites have been studied across the European loess belt. They included approximately 50 buried soils, the over and /or underlying loess and the present-day surface soils (where possible). From the numerous types of secondary CaCO 3 accumulations recognized, only the small scale accumulations are discussed in this paper. including calcified root cells, CaCO~ hypocoatings, needle-fiber calcite, powdery calcite coatings. earthworm biospheroliths, and calcite pseudomorphs on gypsum. Most of the features studied are found in the rhizosphere microecosystem. Each of these features showed a unique relationship with organic matter, micro-organism and plant distribution, moisture and leaching processes. For example, the exceptional abundance of calcified root cells is an indicator of a long stability of soil surface during favorable climatic conditions. Needle-fiber calcite is related to biomineralization of saprophytes in soils with sufficient moisture and decomposing organic matter. The presence of powdery calcite coatings covering clay and Mn coatings in non-calcareous soils indicates a change from leaching to non-leaching soil moisture regimes. Earthworm biospheroids provide arguments tbr a relative stability of the soil surface, without rapid burial processes or drastic, prolonged water stress. Finally, gypsum pseudomorphs are indicators of changes in local climatic parameters. In addition to a discussion of the genesis and the dynamics of the various accumulations, several examples are given to show how these accumulations can be used as indicators of a changing environment, such as renewed loess deposition, the extent of drought or varying intensity of leaching during the year. To conclude, various forms of small scale secondary CaCO 3 accumula- tions are common in the loess paleosoils and each type has the potential to contribute to a better
' Corresponding author. Fax: + 33 3 8039-6387.
0016-7061/97/$17.00 © 1997 Elsevier Science B.V. All rights reserved. PII S()O I 6-706 I (96)00 106- I

222 J. Bec'ze-Defk et al. / Geoderma 76 ¢ 1997) 221-252
understanding of the evolution of the paleoenvironment. Therefore. it is proposed that their individual presence or absence throughout the pedostratigraphical levels and throughout the (paleo)topography should be recorded at all scales of observation.
K¢'ywords: Europe; Quaternary: micromorphology: kv...ss; calcite: biomineralization: rhizosphere: paleosol
1. Introduction
Calcium carbonate accumulations are common secondary features in loess. Related to physico-chemical processes and biota activity, they record part of the pedogenetic history of the loess soil-sedimentary complexes. Detailed examination of loess paleosoils across Europe showed that there are numerous types of secondary CaCO 3 accumula- tions. Among these only small scale accumulations will be discussed. These accumula- tions (less than a few millimetres in size) and their fabric are best observed using a microscope. Although most of the different types of small scale accumulations can be recognized in the field with a hand lens, they are commonly described in general terms, such as filaments (Gile et al.. 1966), pseudomycelium (Bronger, 1975; Bronger et al., 1994), pseudomycelia (Kaemmerer et al., 1991), or kalkmycel (Boenigk et al., 1994).
The primary aim of this study is to present a typology of the small scale accumula- tions in terms of their fabric, mineralogy, and origin. They include calcified root cells. hypocoatings, needle-fiber calcite, powdery coatings, and earthworm biospheroids. Field data, complemented with meso- and micromorphological observations and laboratory data, permitted an overview of the distribution of these accumulations in paleosoils. The second aim is to discuss the origin of these various types of accumulations and their role in the pedogenesis. Finally, an attempt is made to evaluate their potential role in the stratigraphic record of the loess paleosoils and their climatic and environmental signifi- cance.
2. Stratigraphic position and description of the soil-sedimentary sequences
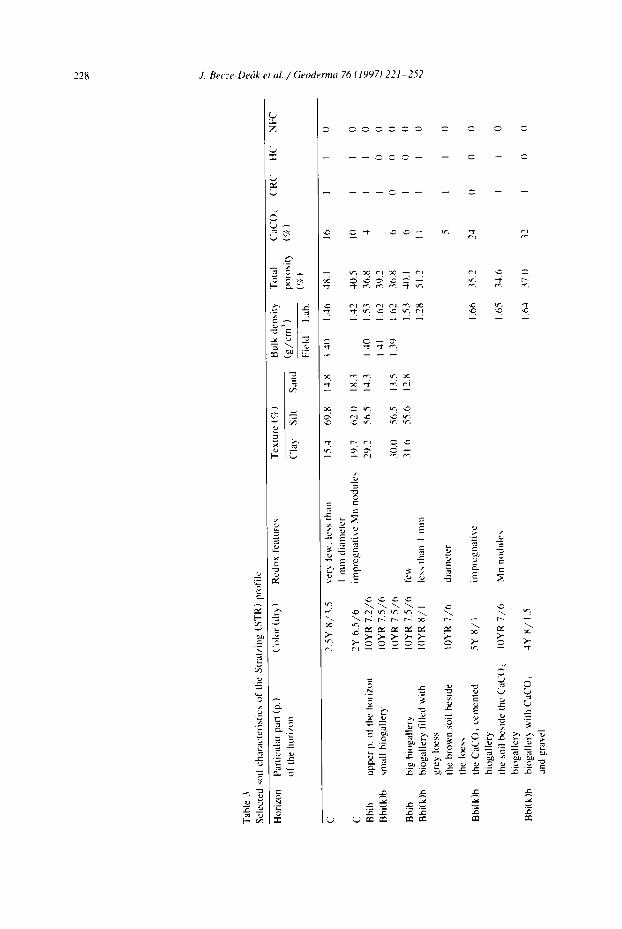
Eighteen Quaternary loess paleosoil sites of the Last Glacial and Last Interglacial periods have been studied across the European loess belt (Fig. 1). Five of the sites were selected for more detailed laboratory investigation because of the abundance of the small scale accumulations or because of the presence of specific types of features. A simplified sketch (Fig. 2) shows these five profiles and the distribution of the various small scale accumulations. Tables 1-5 give a brief characterization of the main soil characteristics. The reference profiles are situated in Wallertheim (Germany-two pro- files), Stratzing (Austria), Dolni Vestonice (Czech Republic) and Mitoc (Romania).
Since the 1920s, the Wallertheim (WA) brickyard has been known for its rich Middle Palaeolithic bones and lithic artifacts (Schmidtgen and Wagner, 1928). Two profiles were selected:
Site 1: the paleosoil selected for the particular abundance of calcified root cells is exposed at the bottom of the upper part of the brickyard (WA I). It is part of a 150 cm

J. Becze-Dedk et al. / Geoderma 76 (1997) 221-252 223
- ' 4 %
sea -- y
Fig. 1. Location of the observed and/or sampled sites across the European loess bell. The sampled profiles are part of the Ioess-paleosoil sequences and/or correspond to the present-day surface soils on loess. Geographic distribution: Belgium: (1) Bruyelle, (2) Roborst; Germany: (3) T6nchesberg, (,1) Metternich, (5) Wallertheim: Austria: (6) Stratzing, (7) G6ttweich. (8) Paudorf, (9) Hollabrun, (10) Stillfried: Czech Republic: (11) Dolni Vestonice: Slol'aki Republic: (12) Nitra, (13) Borovce: Hungary: (14) Basaharc, (15) Mende, (16) Paks. (17) Debrecen; Romania: (18) Mitts.
thick loess sequence on top of a 20-50 cm thick gravel deposit overlying the Tertiary marl substratum.
Site 2: the present-day surface soil (WA 2), a Chernozem, developed in Late Weichselian loess (Leser et al., 1975; Blume, 1986).
In the Stratzing brickyard, a brown paleosoil (STR) is situated a few meters below a Gravettian find layer (Neugebauer-Maresch, 1993). The soil could correspond to the Last Interglacial and probably was situated on a former straight mid-slope position. The upper part of the soil is non-humiferous, slightly calcareous, and shows abundant traces of very strong bioturbation (Bbi) with biogalleries commonly 10-20 cm and locally up to 50 cm in diameter. These galleries are infiiled with material from both overlying and underlying horizons. The recarbonated reddish clay accumulation horizon is underlain by a CaCO 3 cemented horizon, which is on top of a loess sequence several meters thick.
In the Dolni Vestonice brickyard, the (DV) Last Interglacial soil (Klima and Kukla, 1968) is situated in a toeslope position. The soil is characterized by: (1) a humiferous, slightly calcareous surface horizon; (2) a thick well developed clay accumulation (Bt) horizon with a non-calcareous matrix, but prominent CaCO 3 coatings that cover clay, as well as manganese coatings; (3) a very sharp transition to the calcareous parent loess: and (4) below this boundary, CaCO 3 nodules, up to 10 cm in diameter.
In the Mitoc profile (MI). a sequence with three weak humiferous horizons situated between the Gravettian and Aurignatian occupation levels (Haesaerts, 1993) contains peculiar carbonate accumulations in a non-calcareous material. The site is situated on the lower part of a mid-slope. The humiferous horizons are very faint but it was possible to

224 g. Becze-De6k et al. / Geodermtt 76 119971 221-252
/ - - - ~ ~ ~ - T v ~ , ' - ~ - - - , • , ~ , , , , . ,,-,. _ ,. - ,. ,.i,~,.,,~.~,.~, ,,~,,-,..~:~-,. , _,.
~ . ' ! , , ' _ , ~ _ o ' ': " ' , ' , 4 - - : " , ' , ' , , ' q ~ ' t ~ , ' , ' - ' "' ' " , - ' , " v ' ~ ' , / , " - ~ " ~ " n , , " ~ ~ ~ , ~ 1 ' ' - ' ' I : " ~ " " ' " ~ " : ~ : ' w ~ , v " ,%~,d" ' " - , ~ ¢ ¥ ~ " ~ " " " ' .1
.s~ o, ~' ~, ~, g, g' ~, o, $, o, g, ~, ~ g,
S ' - - i , ' :
~-~ ,-., - ' ~' Z' ~' f _ ' ° ' ° ' ~' ~' .- o' ~' o' ~' ~ ~ g' ~ ~ ~ ,-., ,.-,
~'-~ ~. ~ .~'~ ~ , - , ~ ~, v u ,I
,o
.~ ~ . _ z :=; .~ ..~
z
' ",-_: '_, I ~ , . , ~ s _ ~
~. ~ ~ . ~ . ~ ' ~ ~ ~ , - ~
~ ' ~ ~L.;, ~ ~ '~ '~ "~ ~ " : 0 II ~ .I
z o . . . . ~ ~ ~ .~. .~ .~ .~ ~ , ~, :~ ~ ~o.~'~' ,~ ~, ~ ~ ~ ...,- ii
_ ~ ~ r- ...., .= ~ '~
u u ~ ~ ~ u u u u u
, - - : i,~,:, , :i l ,: i , ,_,--, . : , , . . . . . . ", ~ ! -- -. ~, ' , , . , ~ ' , ' . . . . , , " . . : ,,, _ ' - - '- . ,
£-~ z,o,
~ . " ~ . " ". ' ~ - . . ,*",-I ~ o' o' o' ~' o' ~' o' o'
~ . ~ =
• ~ ~ .~ x -~ ".=
~qb
,~,,. - ~ - _ ~ ~ C
, E < ~ , ~
~ . t. ~

J. Becze-Defk et al. / Geoderma 76 (1997) 221-252 225
N
i
~ z m
v ~
~ . ~
> , > > .
d d
i
f,
,~ ~ Z t ' ' '
= ,f,.# •
F- .~ = 2 N ' ~
"r" ~
• E = -
,~ .~_
= ~ . ~
~-~ ~ . . . ~ ~"

226 J. Be ( ' z e -De fk et al. / G e o d e r m a 76 (1997) 2 2 1 - 2 5 2
.¢ C -- rJ rl -- --
a )
y.
<
¢-q
Y:
~z
un "U-.
--r -.(
r i -r
- ~ v 4 t t - , t o ' , t t ~, .

J. Becze-De6k et al. / Geoderma 76 (1997) 221-252 227
follow them all across the excavation. Both the parent loess and the humiferous horizons indicate bioturbation. The amount of secondary carbonates is very small. Gypsum rosettes are present in the lower part of the sequence (3Cb, 4Ab, 4Cb horizons).
3. Materials and methods
Care was taken to ascertain a continuity in recording the data, from the field down to the ultramicroscopic observations. This procedure permitted a close link to be main- tained between laboratory data and field observations.
Most of the sites are well known for their importance in archaeological research. Whenever possible, the present-day surface soil was also sampled. For the selected features, special attention was given to their presence or absence, their spatial variability. and their distinction from features related to diagenetic processes (macroscale). Observa- tions were facilitated by selecting sites with large exposures. The traditional observation of lithostratigraphic records on fresh vertical sections was completed with horizontal sections, whenever possible. All samples collected from pedostratigraphic layers were undisturbed, oriented clods, mostly of at least fist size, except for very friable horizons. where metal KubiEna boxes were used to keep the samples intact.
Samples for thin section preparation were cut from the clods, which were previously observed under the stereomicroscope (mesoscale). This procedure is crucial to maintain the link between the traditional micromorphological study of the soil thin sections and the field observations (e.g., Targulian et al., 1974; Langohr and Pajares, 1983: Callot and Jaillard, 1987). Calcium carbonate and manganese tests (using HCI and H 2 0 2,
respectively) were made to characterize small accumulation features in the soil matrix. After selection and preliminary classification of the features at the macro- and
mesoscale, 54 2.5 × 4.5 cm and 49 6 × 9 cm thin sections were prepared and studied using the optical polarizing microscope (microscale). The micromorphological terms used are according to Bullock et al. (1985). Part of the thin sections were not covered to allow observations with a scanning electron microscope (SEM-JEOL 8400). SEM (ultramicroscale) observations were made on gold coated bulk samples and carbon coated thin sections. Backscattering electron imaging was used on thin sections and the electron dispersive spectroscopy (EDS) microprobe provided measurements of major elements (Ca, Si, K, Fe, AI).
Semi-quantitative estimations of the distribution of the carbonate accumulations were done by comparative observations of both undisturbed samples and thin sections. The abundance of the features was quantified according to tables in Bullock et al. (1985, pp. 24-25; this paper Fig. 2, and Tables 1-5). An exception was made for the earthworm biospheroliths, for which only the presence was noted (the maximum abundance being two individuals per thin section).
Total porosity distribution was measured by mercury porosimetry (Micromeretics Micropore-sizer) on selected samples after drying at 40°C for 24 h. Laboratory bulk densities were calculated based on measurements of the total porosity and apparent skeletal density. Field bulk densities were measured on 24-h oven-dried samples (3 ring samples of 100 cm 3 for each value). Texture was determined on the organic matter and
228 J. Becze-De6k et al. / Geoderma 76 (1997) 221-252
.#
#
. r -
- - t " l r . ~ ,
-'3
~ J
~_~ ~ .
, . ~ , ~ , . c , c
>. ~. >.. >,. >- >.. >..
~=

J. Becze-Dehk et al, / Geoderma 76 (1997) 221-252 2 2 9
u"' , r e " ¢ ' I " I " 3C ~ ~ ~C - - " I "
3 0 ,,+*', 3 ~ i..+. _ _ +,+., 0 , .
I t ' . t r ,
~ - - - - ,+q ~ - - ,-,~ ,--i ~ .~ %, ~c
..1- ,+.. +~
,'--! r,-: :3", <'+ ~".
,~ =~,,,-.,,,,. ,.,.~ ~£ t.-:
t ' l e.+", ~'~
-** ~-~
~+, " ,C +~"', +e~. t " + ~C t#-, I t- , e+'-
t ' , l - -
--r " - t ~
~+, t r , t r , , t r ,
"+', - - ' q " t
¢'+'I r - I - - ¢ ' z
.E
• - ' m ~ - ~ = ~
- - - - - - ¢ " 1 - -
E. •
r ~ r ~ j
=
E
5 ,-B_

230 J. Becze-Dedk et al. / Geoderma 76 (1997) 221-252
m;
> .~.
> "7-
o
'V--.
~7 ~ A
0 ~
e~, e+- ~-, "1" t ~
e g
,6 . . . . ,o ,e ,< ~ -%
¢a 9

J. Becze-Dehk et a l . / Geoderma 76 (1997) 221-252 231
~ ~ ~ - - - - 0 ~
+X
M", - - ,¢,. _.E
"+'. I
-i
--
I
e,
I
E
);
I-
: + =+++ ... ~ ~
_~ ~ 0 +~-
-- ~ ~ ~ i ~ . ~ - . _ ~ . - + ~+ + .+ .+ + + ~ +
+ , + + : ~ . _ b ,"= = .-'5 >
++~ + + + + .. ; - ) - + + + ~ \ - + \

232 J. Becze- l )e ( tk el al. / G e o d e r m a 76 (1997) 2 2 1 - 2 5 2
t r ,
+.,.J
+_
++.,
+"3
C
,.-i + ~ ~ ,
'++-. ~ "~ t+ "r',
.-j. r+-, t++--, t~- ,'r+', ' ~ ,++" l "++ .+. ,++-: ~ ,+,": ~ ~:
.~ ~ ~ ':: r ~ .+.i
• ~ r++ r'! . I - t~ , ~_+ t+,,.-,
..-+ ~ ~ , ~
t.r'. tr+
~+ r~, " t" P+". " P ~ i
,d .d ,~ ,d .~ ~ r.J .d

J. Becze-Dedk et al. / Geoderrm~ 76 (1997) 221-252 233
CaCO 3 free samples (after H,O, and HCI treatment). Fractions > 50 txm were dry-sieved and < 50 ixm fractions were determined using the pipette method (Gee and Bauder, 1986). The total CaCO 3 content was determined using the pressure-calcimeter method (Nelson, 1982).
4. Descriptive typology and origin of the small scale secondary CaCO 3 features
The study showed that the small scale secondary CaCO 3 accumulations occur in more numerous morphological forms than suspected in the field or commonly described in the literature, in this section the morphology and the related distribution of these features will be described at various magnification scales. These data are complemented with discussions on the possible origin and environmental significance of the accumula- tions.
4.1. Calcif ied root cells
Observed with a hand lens, calcified roots are bright calcareous pore fillings of mostly I-ram diameter, filling straight or curved channels. The length of these features only exceptionally exceeds 1 cm. In calcareous loess, they appear in high contrast to the groundmass: they are surrounded by a < 1 mm thick, browner decalcified rim (see below). They are composed of equant or slightly elongated, mostly polyhedral-shaped extremely bright crystals of calcite that occur either as densely packed individual crystals or as tubes formed of fused crystals. Some of these tubes can be very resistant and remain intact after wet sieving of soil fragments on a 200-1xm mesh.
The features described here correspond to both 'crystal tubes' described by Kubii~na (1938, p. 188), Brewer (1964, p. 287), and Bal (1975, p. 165), and 'rhizoliths' resulting from root petrification, according to Klappa (1980). More recently, Jaillard (1987) and Jaillard et al. (1991) have described similar features in calcareous soils from France and referred to them as "calcified root cells".
An extensive study on calcified root cells was done by Jaillard (1987), who described the biomineralization process of roots mainly in strongly calcareous soils. Calcium carbonate in the soil matrix is dissolved by a H +/HCO 3 exchange and the excretion of organic acids. The available Ca 2. is taken up by the root, absorbed by the cells and accumulates in the vacuole where it precipitates as calcium carbonate. The growth mechanism of vacuolian crystals explains 95% of the mass of the calcified cells, whereas the outer part of the cells mineralizes during plasmolysis (the tendency of the plasmolemme to shrink away from the cell wall being due to water stress; Milne and Milne, 1959, p. 43; Jaillard, 1983). This process leads to cortex cell mineralization (parenchyma cells of the plant roots), forming the calcified root cells and explaining the tubular shape. Calcification of the apex is a process rarely observed, occurring mainly in well drained soils (Jaillard, 1985).
At the microscale, the shape of the calcified roots can be straight, curved or branching, being composed of equigranular calcite grains. The individual crystals are either loosely packed or grouped in a close arrangement along the length of the feature. In the latter case, they often form tubes with a hollow central part (Fig. 3a) when cut in

"34 J. llecze-Dec~k el cd. / Geoderm~ 76 t 1997) 221-2.52
tt
• ~ ; :~.0 " P v
• ~ "" ~ . ' I
!
. . # ,
~'2. : • " "
~. ~ ~~ ~ .~ : ~"~ I , i B ~
I " - , ~ r , . # Z , . ~ 1 P" L " , "~ ], , o--~'., r=.N I~
,: , :~'~.~_~.,~ ~ a ~ : ? "N'- , , ... . . ~ . ~ , . ._~ ,~ ~ :
~ ~ ~ ~ ,

J. Becze-De6k et al. / Geoderraa 76 (1997) 221-252 235
their longitudinal section. In cross-section, the feature has a ring structure: a hollow center is surrounded by a rim of calcified cells (Fig. 3b). In calcareous material, the tubes are often surrounded by a calcium carbonate depletion hypocoating of I to a few mm thick.
The cells are independent but the fabric of the assemblage retains the root tissue morphology (Fig. 3b,c). When disturbed the crystal tubes are reduced to clusters of a few cells or to single cells isolated in the groundmass. The individual calcified cells are 40 to 100 txm long on average, and rarely up to 120 Ixm. In cross section, the undisturbed tubes are made up of a succession of layers of cells (from the outer contact with the soil matrix toward the hollow center) in the shape of tablets, polygons, ovoid-cylinders and tablets again (Fig. 3a to Fig. 4b). The ovoid shape is the most common and the best preserved.
The calcified cells can be mono- or polycrystalline, as noted by Jaillard et al. (1991). A singularity is the occurrence of spherulitic and fibro-radial crystals (Fig. 3c,d), similar to those described by Brewer (1964, p. 319) as crystic spherulitic fabric. These peculiar cells do not seem to be related to a specific position in the tube. Under crossed polarizers, each cell is surrounded by a thin rim of cryptocrystalline calcite of less than I ~m thick (Fig. 3d), which appears smooth under the scanning microscope (Fig. 4a). Nevertheless, the cells' surface can have irregularities. Casts of wall structures or microdomes (2 la, m) of cell plasmodesms are preserved during mineralization (Fig. 4a), which can occur even in the intercellular space, leading to complex shaped crystals (Fig. 4b). Cells can also undergo dissolution, either along the crystal lattice or leading to more irregular, scraped or perforated surfaces (Fig. 4b). This weathering of calcified cells is related to the action of yeast or endolithic micro-organisms (Jaillard et al.. 1991), or simply to dissolution by soil solution.
The mineralogy of the cells is clearly identified under the EDS microprobe as low-magnesium calcite. The main elementary component of the cells is calcium. The calcium is not associated with notable amounts of AI, K, S, or Fe, but occasionally inside the cells, the amount of silica is high in a few spots. This is related to the crystallization of silica spheroids inside the cells (similar to ones in Fig. 3d), a feature common in Gramineae (grass) roots (Jaillard, 1983). In addition to the root morphology, the presence of silica spheroids may indicate the type of vegetation.
Calcified root cells were observed in the field at all 18 loess sites and most pedostratigraphic levels, regardless of their age, position in the landscape or field determined texture. However, they do not occur in the alluvial plain deposits of the Last Interglacial at the Wallertheim site. No relationship with the present-day surface can be
Fig. 3. (a) Longitudinal section of a calcified root. Cells form a tube with a hollow central part. Wallertheim site. profile PI. horizon Bk2b, crossed polarized light (XPL). (b) Cross section of calcified rc~t. Wallertheim site. profile P2, horizon Bbi(k), XPL. (c) Well preserved calcified root cells. Are visible in XPL: (I) their monocrystalline nature (2) locally their fibro-radial character. In this case, the cells show a black cross of extinction, emphasizing the fibro-radial character. Wallertheim site, profile P2. horizon Bbi(k), XPL. (d) Detail of calcified root cells. The fibro-radial structure is obvious. ( I ) Presence of siliceous spherulites in root cells is typical for Gramineae. (2) Note birefringent rim of cryptocrystalline calcite. Note also the various shapes of the cells (polygons, ovoids). Wallertheim site, profile P2. horizon C(k), XPL.

236 J. Becze-Defk et al. / (_;,eoderma 76 f' 1997) 221-252
. f f ~ " , . /"
~ .~,. ~ . ~ l :1
@ .
Fig. 4. SEM picture of a calcified root cell. (a) Close-up of one cell. Note presence of microdomes (arrows). Wallerthcim site. profile PI, horizon Bk2b. (b) Calcified rc~t cells with dissolution features: ( 1 ) irregular-shaped and perforated surfaccs due to endolithic micro-organisms. (2) dissolution along fibro-radial crystals: (3J calcified root cells with mineralization in the intercellular space. Wallerlheim site. profile PI. horizon Bk2b.
tound at any of the sites. However. undisturbed calcified roots are present in the soil matrix and in the kratovinas of the Chemozems in the Forest-Steppe region of Russia. In the loess calcareous parent material, they may represent up to 1-2% volume and they are approximately uniformly distributed throughout the sections. In the paleosoils, they have a more uneven distribution and locally they can be very abundant (up to 10ck, Fig. 2). They also occur in partially or totally decalcified soil horizons. In the latter case. either they are relict (just above the decalcification boundary, partially dissolved-DV) or they are related to the overlying calcareous sediment (DV upper part, and MI). They have been observed also in the Early Glacial. humiferous soil-sedimentary complex of the Wallertheim site, where the soil matrix is largely non-calcareous, but contains some small calcareous marl fragments. The latter observations indicate that such accumula-

J. Becze-Defk et aL / Geoderma 76 (1997) 221-252 237
tions can occur in soils with a non-calcareous matrix, the calcium being provided by water flowing laterally or percolating through the soil. Afterwards, they remain pre- served as long as the percolating solution is Ca saturated.
a / \.o
/Y \°+"
100 so 60 s a n d 40 20 0
10£1 0
6°3 \ ,o
o/ . I i - I " I " ~ loo
loc ~o eo s a n d 40 2:) o
Fig. 5. Ternary diagrams showing: (a) relationship between texture of the studied soil-sedimentary complexes and the amount of calcified root cells observed (as indicated in Table 1Table 2Table 3Table 4Table 5): + , no calcified root cells (CRC)" O, few CRC ( < 2 area.C/¢) - D, common CRC (3-5 area%); ,',, abundant CRC (6-10 area %); (b) relationship between texture of the studied soil sedimentary complexes and the amount of CaCO3 hypocoatings observed (as indicated in Table ITable 2Table 3Table 4Table 5): +. no hypocoatings (HC): O, few HC (2 vol. %); D, common HC (3-5 vol.~). According to our data, there is no clear relationship between the texture of the material and the amount of calcified root cells or CaCO 3 hypocoatings. This observation indicates that in the loess soil-sedimentary complex the texture does not influence their distribution.

238 J. Becze-De6k et al. / Geoderma 76 (1997) 221-252
Regarding related distribution, in most of the cases the calcified root cells in the loess soil-sedimentary complexes are present as individual accumulations in pores without any other CaCO 3 accumulation features. There is one exception, in the calcareous parent loess of Dolni Vestonice (DV), where calcified cells are observed in pores with CaCO~ hypocoatings. In this case, no decalcification rim is observable. Occasionally. they are associated with needle-fiber calcite.
There seems to be no particular relationship between the amount of calcified cells and the texture of the soil studied (Fig. 5a). The porosity measurements show that there is no major difference in the total porosity of the samples with or without the calcified root cells (Tables I-5). In addition, according to our data, there is no correlation between the abundance of the calcified root cells and the redoximorphic features observed (Tables 1-5).
Beside the sections studied here (about 50 soils with the under- and overlying loess), calcified root cells have been observed in many other loess sections around the world (e.g., China: Bronger et al., 1994: Xiaomin et al., 1994; Kemp et al., 1995: Russia: Bal, 1975; Poland: Konecka-Betley et al.. 1986: Slovakia: Klukanova and Sajgalik. 1994) although their exact origin has not always been identified. The presence of calcified root cells throughout the calcareous loess sections, regardless of their age and landscape position and the absence of any relationship with the present-day surface soils examined in this study, indicate that their lbrmation is related with the period of loess deposition. In present-day surface soils, calcified root cells have been observed mainly in regions with a Mediterranean moisture regime, i.e., a pronounced warm, dry season (Herrero and Porta, 1987: Jaillard, 1987" Jaillard et al.. 1991: ) or with an aridic moisture regime (Sehgal and Stoops. 1972; Courty and Fedoroff, 1985; Hachicha et al., 1987). Ducloux and Butel (1983) described them also in the wetter environment of southwest France, but with a period of 2 months pronounced summer dryness. This distribution suggests that biomineralization occurs under a climatic regime with a pronounced dry season, which is in agreement with the environment of loess deposition (Mticher, 1986, p. 51) and of Forest-Steppe pedogenesis (as observed by the authors in the Kursk region, Russia).
In loess and loess paleosoils, the length of the accumulations (rarely > I cm) suggests that only part of the plant roots are active in the formation of calcified roots. Traces of silica, observed with optical microscope and confirmed with EDS support the argument that at least some grass species contribute to their formation, i.e.. G r a m i n e a e
are known to commonly accumulate silica. The uneven distribution and the relative scarcity of calcified root cells are most probably related to a combination of various factors, such as the rate of loess accumulation and the scarcity of plant species that are able to accumulate CaCO 3. However, their exceptional abundance in profile 1 of the Wallertheim site (Fig. 2) is considered as an indication of a possible longer stability of this soil surface under favorable climatic conditions.
4.2. H y p o c o a t i n g s
CaCO 3 impregnations of the soil matrix around pores have been observed in the field and with the binocular in many sections (Fig. 2). In loess, they take the form of accumulations a few millimeters thick coating pores, which are a few millimeters in

J. Becze-Derk et al. / Geoderma 76 (1997) 221-252 239
diameter and mostly < 1 cm long. Observations with the microscope show that these impregnations are composed of micritic crystals.
According to Wieder and Yaalon (1982), hypocoatings are the result of rapid precipitation of CaCO 3 from the water suction and desiccating effect due to root metabolism. Their hypothesis is based on the fact that the hypocoatings are frequently associated with 'root remnants'. Brewer (1964) proposed two hypotheses for the formation of CaCO 3 hypocoatings. They are due either to evaporation of a Ca-rich solution from the soil matrix or to precipitation from soil solutions percolating along the pores and penetrating into the soil matrix. The latter hypothesis also has been suggested by Sehgal and Stoops (1972). Courty and Fedoroff (1985), Courty (1990) and Kemp (1995). These publications do not mention the role of vegetation in the dynamics of hypocoating formation. To our knowledge, no experiments were carried out in order to confirm any of the hypotheses. In experiments on the waste products involved in processing bauxite, Thompson et al. (1991) describe micritic hypocoatings on void walls as a result of several wetting and drying cycles. Micritic coatings of channels as a result of evaporating soil solutions, observed in pedozoological experiments, also have been described by Jeanson (1967).
Regarding the distribution of the hypocoatings in the sections studied, they seem to be more abundant in loess parent material than in the calcareous or non-calcareous paleosoils (DV, STR, see Fig. 1). They are present even in the Mitoc profile (Fig. 2), where the soil matrix is non-calcareous. They also occur in the calcareous deposits of the Last Interglacial alluvial plain at the Wallertheim site. In the parent loess of the Stratzing and Dolni Vestonice profiles, hypocoatings associated with bundles of needle- fiber calcite in pores have been observed in several thin sections. According to our data, it seems that the amount of hypocoatings cannot be correlated to the texture of the material (Fig. 5b) or to the total porosity (Tables 1-5). These data suggest that the genesis of hypocoatings is most probably related to local environmental parameters. The presence of needle-fiber calcite in the pores with hypocoatings in loess and paleosoils (STR, DV) indicates the possible influence of vegetation in their formation and most probably also during the loess deposition. Indeed, needles are related to calcified fungal filaments that consume the organic matter (Verrecchia and Verrecchia, 1994).
In present-day surface soils, CaCO 3 hypocoatings have been commonly described in arid and semiarid regions (Sehgal and Stoops, 1972; Courty et al., 1987; Drees and Wilding, 1987; Courty, 1990; Monger et al., 1991), and also in areas with fluctuating water tables (Sehgal and Stoops, 1972). In addition to the sections studied here. hypocoatings have been observed and described in many other loess sections throughout the world (e.g., Argentina: Imbellone and Teruggi, 1987; China: Kowalinski et al., 1972; Guo and Fedoroff, 1990; Xiaomin et al., 1994; Kemp et al., 1995; Canada: Mermut et al., 1990; The Netherlands: Slager et al., 1978).
The presence of C a C O 3 hypocoatings throughout the loess sections, regardless ot" their age and geographical position, suggests that they are probably related to a period of loess buildup, also indicating the aridity of the environment. The simultaneous presence of hypocoatings and calcified root cells might be the consequence of the fact that the various plants are reacting differently to the environmental parameters. Moreover, the presence of hypocoatings in environments with more pronounced evidence of moist

240 J. Bec-e-Defk et al. / Geoderma 76 t 1997) 221-252
conditions, such as the Last Interglacial alluvial plain deposits ot" the Wallertheim site. indicates that the hypocoatings can possibly form in a more moist phase of the soil-sedimentary evolution. In future, the systematic record of the related distribution of these accumulations might enable the differentiation of the fluctuations of the moisture regime during the evolution of the soil-sedimentary complexes.
4.3. Needle-tiber cah'ite
In the present-day calcareous surface soils (Wallertheim. Borovce, Stillfried, Nitra, Debrecen, Fig. 1), needle-fiber calcite is the most common type of calcite in the transition zone between the surface humiferous A horizon and the subsurface C horizon, gradually vanishing in the deeper part of the soil. In these soils, the needle-fiber calcite is associated with large pores, the crystals forming random meshes and convoluted bundles (Fig. 6a). Needle-fiber calcite also has been observed relatively deep in the parent loess of the Strazing and Dolni Vestonice profiles (Fig. 6b). In the calcareous loess, they have been observed several times in channels surrounded by CaCO~ hypocoatings. Locally, they were observed in pores with calcified root cells.
These needles (Fig. 6a,b) are mainly of the MA and occasionally of MB types of Verrecchia and Verrecchia (1994). The MA type needles are long and smooth, com- posed by a variable number of individual fibers, most commonly two. They have a mean length of 15-20 p,m and are less than 1 txm wide (mean around 0.5 ~tm). The MB type needles are serrated-edged and of approximately the same length as the MA type but thicker (up to 2 ~m). These two types are lound together inside the pore network with a random fabric.
KubiEna (1938), who described needle-fiber calcite in loess from Lower Austria and Hungary. already suggested that organic matter greatly favors the formation of needles. The origin of MA needle-fiber calcite is related to fungal biomineralization inside mycelia bundles (Callot et al., 1985a,b). Released after decay of the organic matter of the fungal walls, they may undergo a secondary precipitation of calcium carbonate forming serrated plates (Verrecchia and Verrecchia, 1994). In conclusion, the needle-fiber calcite is thought to be related to saprophyte biomineralization. As such, they are indicators of the presence of decaying organic matter in the soil and they are also indicators of sufficient soil moisture. This may explain why their occurrence is mostly restricted to two layers. The first one is the lower part of the thick humiferous surface horizon of the Chernozem soil type. In this layer, a considerable amount of organic matter is present in combination with relatively moist conditions. This part of the humiferous horizons dries out less than the upper part and some moisture may even be drawn up from the lower soil horizons by capillary suction. In Chernozems of Slovakia, Curlik (1985) also found a strong correlation between the occurrence of needle-shaped calcite and the evaporation regime of the soils in various topographic positions. But he does not consider the process of biomineralization as part of their genesis. The second occurrence is in the calcareous parent loess, once again indicating the presence of vegetation during loess buildup. The needles can be preserved as long as the percolating soil solution is Ca saturated and the loess is out of the reach of intense bioturbation. Absence of needle-fiber calcite in humiferous soil-sedimentary complexes such as the

J. Becze-De6k et al. / Geoderma 76 (1997) 22 I-252 241
Fig. 6. (a) Random mesh of needle-fiber calcite in present-day surface soil. Wallertheim site. profile P2. horizon ABbi(k), XPL. (b) Random mesh of needle-fiber calcite of MB type in the loess. Stratzing profile. horizon C(k,~b, SEM picture.
Early Glacial complex at the Wallertheim site is interpreted as an additional argument for the humus being deposited with the original sediment rather than as corresponding to an in situ, biotic accumulation of fresh organic matter in soil, which would enhance the fungal activity. This humiferous character of the sediment is also confirmed by traces of preserved stratified organic rich pedorelicts, observed at the microscale. A later dissolu- tion is not probable because there are secondary carbonates in this soil-sedimentary complex and the overlying deposit is calcareous loess.
4.4. Powder3' coatings
CaCO 3 coatings with a dull powdery appearance along the ped faces and channels (visible in the field) have been observed in several of the profiles (DV. TO, Fig. 1). The

242 J. Be~'-e-Dedk et al. / Geoderma 76 (1997) 221 - 252
Dolni Vestonice site has been selected for more detailed study (Fig. 2). At this site. the accumulations are related to the overlying calcareous soil-sedimentary complex. In the clay accumulation horizon, they are on top of the clay and manganese coatings (Fig. 7a). Under crossed polarizers, micritic coatings or infillings and locally hypocoatings associ- ated with channels and ped faces are visible. Occasionally, needle-shaped crystals are observable at the edge of the accumulations. The SEM shows that they are composed ol" needles, mainly with serrated plates (Fig. 7b, MB and P type, Verrecchia and Verrec- chia, 1994). Despite their similarity in morphology to needle-fiber calcite at the ultramicroscale, the powdery coatings will be discussed separately because of their different environmental significance. While the random meshes and convoluted bundles in larger pores (described above) are associated with the soil surface in which they
..-. =, , , t ,~ , , ik 1 0 b t m ~ f 7
Fig. 7. (a) Powdery CaCO~ coatings on top of the clay (C) and Mn (M) coatings (am~w), observed with binocular. Dolni Vestonice profile, horizon Btk(b)22. (b) SEM picture of powdery CaCO+ coating, showing the needle-fiber shape of MB and P type (Verrecchia and Verrecchia. 1994"1. 2500×.

J. Becze-Derk et al. / Geoderma 76 (1997) 22 I- 252 243
occur, the powdery coatings reflect the influence of pedogenesis related to the overlying soil surfaces, after the soil was buried. Because the soil matrix is non-calcareous, and as there is no evidence of a water table, the source of the CaCO 3 is probably related to the overlying calcareous loess. The position of the powdery coatings in the profile, on channel walls and ped faces, corresponds with places where roots passed after the soil was buried. The needle morphology indicates biomineralization in the fungal sheath (Callot et al., 1985a,b), probably associated with the decaying organic matter. The serrated plates on the needles have been interpreted as secondary physico-chemical precipitations (Verrecchia and Verrecchia, 1994). The presence of powdery coatings in a non-calcareous soil, where they cover clay and Mn coatings, indicates a change in the amount and chemistry of water percolating through the profile. They signal drier conditions, characterized by a non-leaching soil moisture regime, i.e., continental or Mediterranean.
4.5. Sparitic biospheroids (earthworm biospheroids)
These spheroids have mostly ellipsoidal shapes and diameters, ranging from 350 p.m to 1000 ~m in length and from 200 Ixm to 600 ~m in width. They are composed of pure calcite crystals, visible only in thin section (Fig. 8a). The outer boundary of the crystals is sharp and smoothly undulating. In most cases, there is a thin dark layer (probably rich in organic matter) lining the external boundary. The internal fabric resembles a geode, composed of drusic centripetal calcite crystals, the core being occupied by smaller crystals. They usually are found in biochannels or in the soil matrix and occasionally in earthworm casts. This particular type of spheroid is observed in both profiles at Wallertheim (Fig. 2), which are influenced by the present-day pedogenesis. However, similar accumulations are present in the Last Interglacial and Early Glacial soils at the Wallertheim and TiSnchesberg sites (Fig. 1).
The origin of these accumulations can be related to the calciferous glands of earthworms. Described for the first time by Darwin (1881), they form in the calciferous glands of worms, "pass right through the intestine and are excreted in casts" and "are not peculiar to any major taxonomic groups of earthworms" (Lee, 1985, p. 225-226; Bal, 1977, p. 105). They are "composed of a central cortex of fine soft calcite crystals and an outer zone of larger, harder calcite crystals" (Wiecek and Messenger, 1972, p. 479) articulated in a tight unit (Ponomareva. 1948). They are approximately 0.4 to 2 mm in diameter, with an average weight of 1.5 mg (Wiecek and Messenger, 1972). Biomineralization occurs in the calcigeneous cells of the epithelium that "secrete amorphous calcium carbonate which crystallizes in the lumen of the oesophageal pouch to form spheroliths of calcite, which are excreted into the oesophagus" (Lee, 1985, p. 224). This process could be a reaction against calcium toxicity in lime-rich soils (Crang et al., 1968: Bal. 1977: Simkiss and Wilbur, 1989) and/or contribute to the pH buffering of body fluids during high environmental carbon dioxide (Simkiss and Wilbur, 1989). Nevertheless, in an experiment on a calcium-free soil, Ponomareva (1948) obtained calcite nodules after 47 days by feeding worms on clover and cabbage leaves. The production of calcite was explained by concentration of calcium from the leaves. Therefore. the process of biomineralization is extremely rapid and apparently influenced

244 J. Bec:e-I)e6k et a l . / Geoderma 76 (1997) 221-252
Fig. 8. (a) Earthworm biospheroid, sho~,:ing it,', ~lycrystalline and sparitic composition. The internal structure ix determined by the drusic centripetal calcite crystals. The outcr boundary is undulatory, marked b}. the rounded edge of the individual crystals. WA site. profile Pl(Bk2b), XPI,. (b) Rosette of calcite pseudornorph of gypsum observed with binocular. Mitoc profile, horizon 4Ch.
by a protein matrix in which mineralization occurs (Nakahara and Bevelander, 1969). The presence of this protein-rich envelope can explain the presence of a thin organic layer around the spheroids observed in the selected soils (compare Fig. 8a with Fig. 4 in Nakahara and Bevelander, 1969).
These biospheroids indicate the presence of earthworms, which is obvious in the case of the present-day Chernozem type surface soils with numerous earthworm galleries. When, through diagenesis, clear traces of earthworm activity are absent in the buried paleosoils, the presence of the earthworm biospheroids may provide an argument for a

J. Becze-Derk et al. / Geoderma 76 (1997) 221-252 245
relative stability of the soil-sedimentary environment: no rapid burial process and no drastic prolonged water stress. Indeed, one of the most important environmental requirements for earthworms is adequate moisture, because their water conservation mechanisms are poorly developed (Lee, 1985). They can also give an indication of the in situ character of the soil-sedimentary complex, because the spheroids break when transported (including the outer boundary of the crystals) and the organic coating disappears. The presence/absence of these characteristics may permit a distinction to be made as to whether the spheroids formed in the soil or if they are part of the sediment.
Other secondary CaCO3 accumulations related to earthworm activities mentioned in the literature are: ( l) micritic coatings deposited on the walls of the earthworm channels by the evaporating soil solution (Jeanson, 1964, 1967), and (2) sparitic calcite crystals ('intercalary crystals', Bal. 1977) in the soil matrix. In the sections studied, none of these features have been observed or distinguished from the other features recognized.
Courty et al. (1989) have described calcareous granules with internal radial fabric and sizes similar to those described above, which they interpreted as representing the internal shell of slugs (Arionids). However, the natural occurrence of slugs is related to environments of high humidity and low calcium availability (Barnes. 1980), their internal shell being an adaptation to these environmental conditions. Therefore, with the present state of knowledge we attribute the biospheroids observed in loess paleosoils to earthworms.
4.6. Gypsum pseudomorphs
Bright white, lens-shaped crystals, mostly reacting with HCI, have been observed with the binocular throughout the non-calcareous soil-sedimentary complex of the Mitoc site (Fig. 2). These crystals (mostly 150-200 txm, but up to 500 ~m long) form: (1) rosettes, a few mm in diameter (Fig. 8b); (2) occasionally tubules along channels; and (3) locally individual lenses. Commonly. they are partially coated by manganese and often display serrated edges. Under crossed polarizers, these lens-shaped crystals show high interference colors, typical of calcite. However, occasionally greyish interference colors have been observed. The internal fabric of these crystals is often poikilotopic. Starting approximately from the first humiferous horizon (2(A)b) downward (Fig. 2), gypsum rosettes and tubular accumulations have been observed both in the field and at the microscale. The gypsum crystals also have a poikilotopic internal fabric. In this part of the sequence, the lenticular crystals are less frequent. In the upper part of the sequence, no gypsum has been observed, but the lenticular rosettes are more common and larger. The shape of the crystals and their optical characteristics suggest that they are calcite pseudomorphs of gypsum. The microprobe analyses show that these crystals are calcite, but with traces of sulphur, confirming this hypothesis. Transformation of gypsum to calcite has been described, and experimentally proved by Lattman and Lauffenburger (1974), and explained by the action of bacteria in a reducing environ- ment. In the soil-sedimentary complex of the Mitoc site with abundant evidence of biological activity, the influence of the bacteria is very probable. Moreover, the association with manganese coatings and the presence of redoximorphic features (see Table 5) are indicators of a temporary reducing environment, favoring the transforma-

246 d. Bee':e-Derek et al. / Geodenna 76 11997) 221-252
tion by the bacteria. Nevertheless, the physico-chemical transformation is also possible, but the available data do not permit more detailed interpretation.
To our knowledge, this is the first time that such accumulations are described in loess paleosoiis. Although calcite pseudomorphs of gypsum have been presumed, based on crystal morphology and staining methods in studies of soils in Texas (Drees and Wilding, 1987), no data were mentioned on the nature of the process that is involved in the transformation. According to Drees and Wilding (1987), gypsum crystals also occur lower down in the sequence.
In the case of the Mitoc sequence, the related distribution of calcite pseudomorphs and gypsum crystals can be used as an indicator of change in the local climatic parameters. Although initially the evolution of the non-calcareous soil-sedimentary complex was characterised by more aridic conditions (gypsum formation), later the moisture regime was less aridic with at least some water percolating through (from above or laterally), dissolving the gypsum and favoring the growth of the calcite pseudomorphs.
5. Discussion and conclusion
Most of the features described in this study belong to the particular microecosystem of the rhizosphere. Roots of some plants (including grass species) calcify and form calcified root cell structures. In other plant species or under different environmental conditions, roots suck water from the soil solution in such a way that they supersaturate the medium around the roots with calcium carbonate, leading to hypocoatings. Earth- worms eat the carbonate-rich ground and/or organic tissues and excrete calcium carbonate spheroids in their casts. Fungi that consume organic matter can also mineral- ize and produce various amounts of needle-fiber calcite, which accumulates in the soil pores. All of these calcium carbonate redistributions concern relatively low amounts of CaCO~. A very particular aspect among these accumulations is the relatively large size of the calcite crystals as compared to the micritic matrix of the loess. Moreover, they can form in a very short period: crystals of calcified root cells after 24 h (Jaillard. 1992), earthworm biospheroliths in 47 days (Ponomareva, 1948), bacterial transformation of gypsum to calcite in 17 days (Lattman and Lauffenburger, 1974). As such, these features are very sensitive environmental recorders. That these biogenic processes do not produce more important quantities of secondary calcium carbonate is probably due to the fact that the environment of loess deposition is characterized by limited biological activity (patchy vegetation, short growing season). Biomineralization can further be inhibited by loess deposition, burying the surface, and moving the active microecosystem higher in the stratigraphy. The systematic presence of these various torms of secondary carbonates in the loess soil-sedimentary complexes indicates that, indeed "'loess is not just an accumulation of dust", as suggested by P6csi (1990).
The distribution of the various accumulations in the loess soil-sedimentary complexes can give important indications about both the minor and major events of change in the environment. Bioturbation of calcified root cells will leave clusters of few cells or isolated crystals in the groundmass. This can be interpreted as an indication of the

J. Becze-Defk et al. / Geoderma 76 (1997) 22 I - 252 247
temporary absence of loess deposition. The external shape of the earthworm bio- spheroliths allows the detection of erosion-sedimentation processes. Change in the moisture regime, i.e., transition from non-leaching environment of the loess deposition to leaching environment, is detectable through the dissolution features or by observation of the related distribution of the secondary carbonates versus the decalcification bound- ary. However, in terms of the environmental significance of small scale secondary CaCO 3, most can be related to the moisture status during the soil evolution, with little information about the temperature gradients. All these observations underline the importance of a pedosedimentary approach in investigations of the environmental changes recorded in the loess soil-sedimentary sequences, as concluded by Kemp (1995).
The slow buildup of loess on a vegetated surface, with patchy distribution of various plant species in a relatively dry climate, can best explain the uneven distribution of calcified root cells. The existing observations give no clear relationship of the related distribution of the calcified root cells and the hypocoatings. It is not excluded that they are formed by different plant species, possibly active under different environmental conditions. This can occur even on a seasonal basis or be induced by the topographic position. The absence of earthworm biospheroliths in the loess sequences might be related to prolonged water stress or too high a sedimentation rate. The distribution of needle-fiber calcite is an indicator of available decaying organic matter as well as of the absence, at a particular depth, of a severe prolonged drought. In a subsequent leaching environment, some of the secondary carbonates may disappear, but again the depth of their absence along toposequences can provide important information about intensity and dynamics of the various climatic parameters. The switch from leaching to non-leach- ing conditions is also recorded by the occurrence of calcified root cells, hypocoatings. and powdery coatings in the horizons with evidence of former decalcification.
Fig. 9 is a schematic representation o1" possible lbrms of microscale carbonate accumulations under various changing environments. The gradual buildup of loess deposits on vegetated surfaces is marked by the scattered distribution of calcified root cells and/or hypocoatings and/or needle-fiber calcite (Fig. 9a). The absence of loess deposition is marked by a relative increase in density of calcified root cells and/or hypocoatings and/or needle-fiber calcite compared to the under- and overlying loess. Bioturbation of these accumulations (best observed with a polarizing microscope) is another argument for longer absence of loess input (Fig. 9b). The presence of needle-fiber calcite, associated with more humiferous horizons, indicates that there was sufficient moisture lbr at least seasonal vegetation growth. In such environments earthworm biospheroliths may also be present. These features will be present as long as leaching conditions do not occur. Moreover, the depth of occurrence of needle-fiber calcite might give an indication of the extent of severe drought and/or seasonal leaching (Fig. 9c). Once the moisture regime changes from non-leaching to leaching, the microscale carbonates will be partially or totally dissolved. When totally dissolved, their former presence can be traced by their related distribution with the decalcification boundary (Fig. 9d). The gradual buildup of the new loess deposits is again marked by scattered distributions of calcified root cells, hypocoatings, needle-fiber calcite (Fig. 9el-e3). Moreover, the change from a leaching to a non-leaching environment is clearly marked

2 4 8 Y. Be(ze-l)ec'~k et al. / Geoderma 76 (1997~ 221-252
, . . \ .. Ob
2"
q
z
8~
z~
I
I
°°Oo ~.
""" ' ~ (~I,
e~
7.
Z
I
°° ]
• X i .... "°'. ~ , i ''~
N '~ ~ ;~'~ I
J
~0
O0
c O
E t,
• ~ b-: = . ' 2 - ~ o
= : , l ~,
, - - _
~'=
v '--- h

J. Becze-De6k et al. / Geoderma 76 (1997) 221-252 249
by secondary carbonates, which are found on both sides of the decalcification boundary and in the decalcified matrix (Fig. 9e3).
In conclusion, small scale secondary CaCO 3 accumulations are common in the loess paleosoils. There are various forms of such accumulations, each of them having a potential to contribute to the understanding the evolution of the (paleo)environment. Many of the studies mentioning secondary calcium carbonate accumulations focus on micromorphological observations. At this scale the various types of accumulations are recognized, but very rarely a clear link is made between the origin and their distribution. For a better understanding of the environmental significance of these accumulations, we propose that their individual presence or absence throughout the pedostratigraphic levels and throughout the (paleo)topography should be recorded at all scales of observation.
Acknowledgements
Field work was greatly facilitated by the generous help of the archaeologists Dr. N.J. Conard, Dr. V. ChiricS., Dr. D. Stassikova, Dr. J. Tejral, and Dr. C. Neugebauer-Maresch. This text presents research results of the Federal Services for Scientific, Technical and Cultural Matters, Project SC-004 initiated by the Belgian State, Prime Minister's Office, Science Policy Programming. The scientific responsibility is assumed by its authors. The micromorphoiogical observations were performed with equipment provided by two grants from the Belgian National Science Foundation in 1990 and 1991 to Dr. R. Langohr. Dr. E. Verrecchia contributed to this study during a post-doctoral year in Ghent University, funded by the EEC grant No. ERB4001GT931740. The authors would like to thank Dr. G. Stoops and Dr. D.H. Yaalon for their helpful comments and critical reading of the manuscript.
References
Bal. L.. 1975. Carbonate in soil: A theoretical consideration on, and proposal for its fabric analysis. 2. Crystal tubes, intercalary crystals, K fabric. Neth. J. Agric. Sci., 23: 163-176.
Bal. L., 1977. The formation of carbonate nodules and intercalary crystals in the soils by the earthworm Lumbricus rubellus. Pedobiologia, 17: 102-106.
Barnes. R.D., 1980. Invertebrate Zoology. Sauders College. Philadelphia. 1089 pp. Blume, H.P., 1986. Soil near Wallertheim. In: Mitteilungen der Deutschen Bodenkundlichen Gesellschaft, 46.
Guidebook Tour A. Soils and Landscapes in Northern and Southern Germany. pp. 171-173. Boenigk. W.. Frechen. M. and Weidenfeller. M.. 1994. Die mittel- und obenpleistoz~.ne Deckschichtenfolge im
Naturschutzgebiet 'Eiszeitliches L6ssprofil" in Koblenz-Metternich. Mainzer Geowiss. Mitt., 23: 287-320. Brewer. R.. 1964. Fabric and Mineral Analysis of Soils. Wiley. New York. 470 pp. Bronger. A.. 1975. Pal~iobtiden als Klimazugen-dargestellt an L~ss-Boden-Abfolgen des Karpatenbeckens.
Eiszeitalter Gegenwart. 26:131 - 154. Bronger, A.. Bruhn-Lobin. N. and Heinkele. Th., 1994. Micromorphology of paleosols-genetic and paleoenvi-
ronmental deduction: case studies from central China. south India, NW Morocco and the Great Plains of the USA. In: A.J. Ringrose-Voase and G.S. Humpreys (Editors). Soil Micromorphology: Studies in Management and Genesis. Proc. IX Int. Working Meeting on Soil Micromorphology. Townsville, Australia, July 1992. Developments in Soil Science 22. Elsevier. Amsterdam. pp. 187-206.

250 J. Be('ze-Defk et al. / Geoderma 76 ( 1997~ 221-252
Bullock. P., Fedoroff. N.. Jongerius. A.. Stoops. G. and Tursma. T.. 1985. Handb(}ok for Soil Thin Section Description. Waine Research Publications, Wolderhampton, 152 pp.
(_'allot, G. and Jaillard, B., 1987. Apports de la Ioupe binoculaire a I'etude des interfaces sol/racine et sol/champignon. In: N. Fedoroff. l,.M. Bresson and M.A. Courty (Editors). Soil Micromorpholog). Proc. VIIth Int. Working Meeling on Soil Micromorphology. Paris-July 1985. Association Fran(~aise pour l'Etude du Sol, Paris. pp. ?3-?9.
Callot, G.. Guyon. A. and Mousain. D.. 1985a. Inler-relations entre aiguilles de calcite el hyphes myc0.1iens. Agronomie. 5(3): 209-216.
Callot. G.. Mousain. D. and Plassard, C., 1985b. Concentrations de carbonate de calcium ,~ur les patois des hyphes myc~liens. Agronomie. 5(2): 143-150.
Court). M.A.. 1990. Pedogenesis on Holocene calcareous parent materials under semi-arid conditions (Ghaggar Plain. NW India). In: L.A. Douglas (l'klitor). Soil Micromorphology: a Basic and Applied Science. Proc. Vlllth Int. Working Meeting of Soil Micromorphology. San Antonio. Texa,~. July 1988. Elsevier. Amsterdam. pp. 361-366.
Court~. M.A. and Fedoroff. N.. 1985. Micromorphology of recent and buried soils in a semiarid region of Northwestern India. Gee, derma, 35: 287-332.
Courty. MA. . Dhir. P. and Raghavan. tt.. 1987. Microfabrics of calcium carbonate accumulations in arid soils of Western India. In: N. Fedoroff. I , .M Bresson and MA. Courty (Editors), Soil Micromorphology. Prcx:. Vllth lnt. Working Meeting on Soil Micromorphology. Paris. Jul,, 1985. Association Frangaise pour I'l~tude du Sol, Paris, pp. 227-235.
Courty. M.A., Goldberg. P. and Macphail. R . 1989. Soils and Micromorphology in Archeology. Cambridge University Press. Cambridge, 344 pp.
(:rang. R.t':.. Holsen. R.C. and Hilt. J.B., 1968. Calcite production in Mit()chondria of earthworm calciferous glands. Bioscience, 18(4): 299-301.
Curlik, J.. 1985. Processes of carbonatizalion in soils. In: Soils in relation to the phytomass production and landscape formation. Trans. 6th Czechoslm.'ak Soil Science Conf.. Nitra. 2: 352-357.
Darwin. C.R.. 1881. The formation of vegetable mould through the action of worms, with observations on their habits Murray, l,ondon. In: Lee, K.E. 1985. pp. 226.
Drees. L.R. and Wilding, I_..P.. 1987. Micromorphic record and interpretations of carbonate forms m the Rolling Plains of Texas. Geoderma. 40: 157-175.
I)ucloux. J. and Butel. P., 1983. Micromorphology of calcretes in a slope deposit in the Poitevine Plain. France. In: P. Bull(xzk and C.P. Murphy (Editors). Soil Micromorpholog'.. Vol. 2: Soil Genesis. A B Academic Publishers, Berkhamsted. pp. 637-646.
Gee, G.W. and Bauder. J.M.. 1986. Particle size anal)sis. In: A. Klute (Editor): Meth(v, Js of Soil Anal'.,ses. Part I, Physical and Mineralogical Methods. American Society of Agronorns.. Soil Science Society ot America, Publisher, Madison. Wisconsin, pp. 399-441.
Gile. l,.lt., Peterson. F.F. and Grossman. R.B.. 1966. Morphological and genetic sequences of carbonate accumulation in desert soils. Soil Sci.. 11)1(5): 347-360.
Guo, ZT. and Fedoroft'. N.. 1990. Genesis of calcium carl'xmate in k~'ss and in paleosoils in central China. In: L.A. Douglas (Editor), Soil Micromorphology: a Basic and Applied Science. Proc. VIIhh Int. Working Meeting of Soil Micromorphology. San Antonio. Texas. July 1988. Elsevier. Amsterdam. pp. 355-359.
Hachicha. M.. Stoops, G. and M'Hiri. A.. 1987. Aspects micromorphologiques de I'ew)lution des sols de lunettes argileuses en Tunisie. In: N. Fedoroff, L.M. Bresson and M.A. Courty (Editors). Soil Micromor- phology. Prec. Vlhh International Working Meeting on Soil Micmmorphology, Paris. July 1')85. Ass(x:ia- lion Fran~aise r~mr I'l~tude du Sol. Paris, pp. 193-197.
Haesaerts, P.. 1993. Stratigraphie du gisement paleolithique de Mitoc Malul Galben. Prehistoire Europeenne. 3: 67-71.
Herrero. J. and Porta, J.. 1987. Gypsiferous soils in the North-East of Spain. In: N. Fedoroff. I,.M. Bresson and M.A. Courty (Editors). Soil Micromorphology. Proc. Vlhh Int. Working Meeting on Soil Micromor- phology, Paris, July 1985. Association Fran~aise pour I'[~tude du Sol. Paris. pp. 187-192.
[mbellone, P.A. and Teruggi, M.E.. 1987. Discontinuous calcretes in I(ycssic paleosols near La Plata. Argentina. In: N. Fedoroff. I,.M. Bresson and M.A. Courty (Editors), Soil Micromorphology. Prec. Vlhh Int. Working Meeting on Soil Micromorpholog~, Paris. July 1985. Association Fran~aise pour l'Etude du Sol. Paris, pp. 625-630.

J. Becze-De6k et al. / Geoderma 76 (1997) 221-252 251
Jaillard, B., 1983. Mise en 6vidence de la calcitisation des cellules corticales des racines des Gramin~ees en milieu carbonate. C. R. Acad. Sci., Paris. 297(2): 293-296.
Jaillard. B., 1985. Activite racinaire et rhizostructures en milieu carbonate. Pedologie. XXXV(3): 297-313. Jaillard, B., 1987. Les structures rhizomorphes calcaires: module de r6organization des min6raux du sol par les
racines. Institut National de la Recherche Agronomique l,aboratoire de Science du Sol, Montpellier, 221
PP- Jaillard, B.. 1992. Calcification des cellules corticales des racines en milieu calcaire. Bull. Soc. Bot. Ft.. 139,
Actual. Bot.. I : 41-46. Jaillard, B., Guyon. A. and Maurin. A.F., 1991. Structure and composition of calcified roots and their
identification in calcareous soils. Geoderma, 50: 197-210. Jeanson. C.. 1964. Micromorphologie et pddozoologie exp~rimentale: Contribution h 1"6tude sur plaques
minces de grande dimensions d'un sol artificiel structur6 par les Lombricides. In: A. Jongerius (Editor): Soil Micromorphology. Pro<:. Second Int. Working-Meeting on Soil Micromorphology. Amhem, The Netherlands, September 1964. Elsevier. Amsterdam. pp. 47-55.
Jeanson. C.. 1967. Migrations chimiques dans un sol artificiel: etude micromorphologique. Geoderma. I: 325-345.
Kaemmerer. M. Revel, J.-C. and Barlier. J.-F., 1991. Formation des amas friables et des n~xlules calcaires dans des sols argileux en r6gion temp~r~e et semi-aride. Sci. Sol. 29(1 ): I - 12.
Kemp. R.A.. 1995. Distribution and genesis of calcitic pedoleatures within a rapidly aggradating Ioess-paleo- sol sequence in China. Geoderma. 65:303-316.
Kemp, R.A., Derbyshire, E.. Xingmin. M., Fahu. Ch., Baotin, P.. 1995. Pedosedimentary reconstruction of thick Ioess-paleosol sequences near Lanzhou in North-Central China. Quat. Res., 43: 30-45.
Klappa. C.F., 1980. Rhizoliths in terrestrial carbonates: classification, recognition, genesis and significance. Sedimentology, 27: 613-629.
Klima, B. and Kukla. J.. 1968. Detailquerprofile durch die Hauptwand der Lehmgrube der verlassenen Ziegelei in Dolni Vestonice. In: J. Demek and J. Kukla (Editors). 1969. Periglazialzone. L6ss und Pal~iolithikum der Tschechoslowakei. pp. 102.
Klukanova. A. and Sajgalik, J.. 1994. Changes in loess fabric caused by collapse: an experimental study. Quat. Intern., 24: 35-39.
Konecka-Betley, K.. Czepinska-Kaminska. D. and Zagorski. Z.. 1986. Development and properties of paleosols in the Loess Section at Sandomierz (SE Poland). Annales Universitatis Mariae Curie-Skl<)do- woska. Lublin Poland. XLI(12): 203-212.
Kowalinski. S., Pons, L.J. and Singer, S.. 1972. Micromorphological comparison of three soils derived from loess in different climatic regions. Geoderma. 7: 141-158.
Kubi~na, W.U. 1938. Micropedology. Collegiate Press. Ames, Iowa, 243 pp. Langohr. R. and Pajares. G., 1983. The chronosequence of pedogenetic processes in Fraglossudalfs of the
Belgian loess belt. In: P. Bullock and C.P. Murphy (Editors). Soil Micromorphology. Vol. 2: Soil Genesis. A B Academic Publishers, Berkhamsted, pp. 503-511.
Lattman. L.H. and Lauffenburger, S.K., 1974. Prop~sed role of gypsum in the lormation of caliche. Z. Geomorphol., N.F.. Suppl. Bd. 20: 140-149.
Lee, K.E., 1985. Earthworms. Their Ecology and Relationships with Soils and Land Use. Academic Press Australia, Sydney, 411 pp.
Leser, V.H.. Maqsud. B. and Maqsud, N.. 1975. Sp~tglaziale bis friihholozfine Steppebodenbildung und Klimaentwicklung im siidlichen Rheinhessischen Tafel- und Hiigelland. Eiszeitaltcr Gegenwart, 26: 119-130.
Mermut. A.R.. Rostard, H.P.W. and Arnaud, St. R.J.. 1990. Micromorphological studies of some loess soils in Western Saskatchewan. Canada. In: L.A. Douglas (Editor), Soil Micromorphology: a Basic and Applied Science. Pro,..'. Vllltb International Working Meeting of Soil Micromorphology. San Antonio. Texas. July 1988. Elsevier, Amsterdam. pp. 309-315.
Milne. kJ . and Milne. M., 1959. Plant Life. Prentice-Hall. Englewood Cliffs. N.J., 283 pp. Monger. H.C.. Daugherty, L.A. and Gile. L.. 1991. A Microscopic Examination of Pedogenetic Calcite in an
Aridisol of Southern New Mexico. In: W.D. Nettleton (Editor). Occurrence. Characteristics, and Genesis of Carbonate. Gypsum, and Silica Accumulations in Soils. SSSA Special Publication Number 26. Soil Science Society of America, Inc.. Madison, Wisconsin. pp. 37-60.

252 J. Becze-Defk et al. / Geoderma 76 (/997) 221-252
Mticher. H.J., 1986. Loess: a critical review. In: H.J. Miicher (Editor). Aspects of Loess and l,oess derived Slope Deposits: an Experimental and Micromorphological Approach. Fysisch Geografisch en Bodemkundig Laboratorium, Universiteit van Amsterdam. pp. 19-100.
Nakahara. H. and Bevelander. G., 1969. An electron microscope and autoradiographic study of the calciferous glands of the earthworm. Lumhricus terrestris. Calc. Tiss. Res.: 193-201.
Nelson, R., E.. 1982. Carbonate and Gypsum. In: A.L. Page (Editor). Methods of Soil Analysis. Part 2. Chemical and Microbiological Propreties. American Society of Agronomy. Soil Science Society of America. Publisher. Madison, Wisconsin. pp. 181-196.
Neugebauer-Maresch. Ch.. 1993. Siedlungswesen. In: Ch. Neugebauer-Maresch (Editor). Altsteinzeit im Osten Osterreichs. Wissenschaftliche Schriftenreihe, Niederi.isterreich. Verlag Niedert~sterreichisches Pressehaus. St. P61ten-Wien, pp. 56-64.
P6csi, M.. 1990. Loess is not just the accumulation of dust. Quat. Intern.. 7 /8 : 1-21. Ponomareva. S.l., 1948. The rate of formation of calcite in soils by earthworms. Rep. Acad. Sci. USSR. 61(3):
505-507. Schmidtgen, O. and Wagner. W., 1928. Eine altpaRiolithische Jagdstelle bei Wallertheim in Rheinhessen.
Notizblatt des Vereines ftir Erdkunde und der Hessischen Geologischen Landesanstalt zu Darmstadt, V(I I): 1-31.
Sehgal, JL. and Stc~ps. G.. 1972. Pedogenetic calcite accumulation in arid and semi-arid regions of the Indo-Gangetic alluvial plain of Erstwhile Punjab (India) - their morphology and origin. Geoderma. 8: 59-72.
Simkiss, K. and Wilbur, K.M.. 1989. Biomineralization. Academic Press, San Diego, 337 pp. Slager. S.. Jongmans, A.G.. Miedema. R. and Ports, L.J., 1978. Fossil and recent soil f~rmation in Late
Pleistocene loess deposits in the southern part of the Netherlands. Neth J. Agric. Sci.. 26: 326-335. Soil Survey Staff. 1994. Keys to Soil Taxonomy. U.S.D.A-S.C.S.. U.S. Gov. Print Office. 306 pp. Targulian. V.O.. Birina, A.G., Kulikov. A.V., Sokolova, T.A. and Tselischeva, L.K.. 1974. Arrangement.
composition and genesis of sod-pale podzolic soil derived from mantle loess. Morphological investigation. Xth Inter. Congress Soil Science. Moskow, 47 pp.
Thompson, T.L.. Hossner. L.R. and Wilding. L.P.. 1991. Micromorphology of calcium carbonate in bauxite processing waste. Geoderma. 48: 31-42.
Verrecchia. E.P. and Verrecchia. K.E., 1994. Needle-fiber calcite: a critical review and a proposed classifica- tion. J. Sediment. Res., A64(3): 650-664.
Wiecek, C.S. and Messenger. A.S.. 1972. Calcite contributions by earthworms to Forest Soils in Northern Illinois. Soil Sci. Soc. Am. Proc.. 36: .-I-78-480.
Wieder, M. and Yaalon, DH., 1982. Micromorphological fabrics and developmental stages of carbonate nodular forms related to soil characteristics. Geoderma, 28: 203-220.
Xiaomin. F., Jijun. L.. Derbyshire. E., Fitzpatrick, E.A. and Kemp, RA., 1994. Micromorphology of the Beiyuan loess-paleosol sequence in Gansu Province. China: geomorphological and paleoenvironmental significance. Palaeogeogr. Palaeoclimatol. Palaeoecol.. II1: 289-303.
Copyright © 2022 FDOKUMEN




















