Size-dependent cellular toxicity of silver nanoparticles
11
Size-dependent cellular toxicity of silver nanoparticles Tae-Hyun Kim, 1 Meeju Kim, 2 Hyung-Seok Park, 2 Ueon Sang Shin, 1,2 Myoung-Seon Gong, 1,2,3 Hae-Won Kim 1,2,4 1 Institute of Tissue Regeneration Engineering (ITREN), Dankook University, South Korea 2 Department of Nanobiomedical Science and WCU Research Center, Dankook University, South Korea 3 Department of Chemistry, Dankook University, South Korea 4 Department of Biomaterials Science, School of Dentistry, Dankook University, South Korea Received 6 August 2011; revised 4 November 2011; accepted 29 November 2011 Published online 5 February 2012 in Wiley Online Library (wileyonlinelibrary.com). DOI: 10.1002/jbm.a.34053 Abstract: Silver nanoparticles (AgNPs) have found a variety of uses including biomedical materials; however, studies of the cytotoxicity of AgNPs by size effects are only in the beginning stage. In this study, we examined the size-dependent cellular toxicity of AgNPs using three different characteristic sizes ( 10, 50, and 100 nm) against several cell lines includ- ing MC3T3-E1 and PC12. The cytotoxic effect determined based on the cell viability, intracellular reactive oxygen species generation, lactate dehydrogenase release, ultra- structural changes in cell morphology, and upregulation of stress-related genes (ho-1 and MMP-3) was fairly size- and dose-dependent. In particular, AgNPs stimulated apoptosis in the MC3T3-E1 cells, but induced necrotic cell death in the PC12 cells. Furthermore, the smallest sized AgNPs (10 nm size) had a greater ability to induce apoptosis in the MC3T3- E1 cells than the other sized AgNPs (50 and 100 nm). These data suggest that the AgNPs-induced cytotoxic effects against tissue cells are particle size-dependent, and thus, the particle size needs careful consideration in the design of the nanopar- ticles for biomedical uses. V C 2012 Wiley Periodicals, Inc. J Biomed Mater Res Part A: 100A: 1033–1043, 2012. Key Words: silver nanoparticles, cell toxicity, in vitro, size dependence, endocytosis How to cite this article: Kim T-H, Kim M, Park H-S, Shin US, Gong M-S, Kim H-W. 2012. Size-dependent cellular toxicity of silver nanoparticles. J Biomed Mater Res Part A 2012:100A:1033–1043. INTRODUCTION Silver nanoparticles (AgNPs) are one of the most widely used nanomaterials in personal care products, dressings as treat- ments for external wounds, ointments, and surgical instru- ments because of their effective antibacterial activity. 1–3 Furthermore, the application of AgNPs has been expanded in biological and medical sciences such as drug and gene delivery vehicles, biosensors, implantable materials, and bone prostheses. 4,5 However, these various applications increase human exposure to AgNPs and thus elevate the potential risk related to their toxicity. For this reason, the toxicity of AgNPs is considered to be a critical factor when using them for biomedical purposes. 6,7 Several studies have reported that AgNPs showed signif- icant toxicity against various cell types. For example, AgNP induced cytotoxicity in macrophages 8 and human T cells. 9 A decrease in cell viability and antiproliferative activity in response to treatment with AgNPs was also observed in neuron, liver, skin, and lung epithelial cells. 10–14 Kim et al. 15 reported the subchronic oral toxicity and tissue accumula- tion of AgNPs in rats for 90 days. In their study, histopatho- logical investigations revealed a higher incidence of bile duct hyperplasia, necrosis, fibrosis, and pigmentation in treated animals. There was also a dose-dependent accumu- lation of silver in tissues. Other studies have shown that the AgNPs can pass through the epithelia of the respiratory tract and across the blood stream directly. 16 AgNPs also can be taken up by nerve endings and directly accessed to the CNS. 17 It is generally accepted that AgNPs express their toxicity through the generation of reactive oxygen species (ROS). 13 Increased cellular ROS levels are associated with the induc- tion of genetically programmed cell death (apoptosis), as well as necrotic cell death in several cell lines. 18 Apoptosis eliminates damaged or unwanted cells via nuclear conden- sation, membrane blebbing, and DNA fragmentation in the process of development or in response to infection or DNA damage. 19,20 In NIH3T3 fibroblasts, AgNPs were found to have cytotoxic effects and induce apoptosis, the release of cytochrome c into the cytoplasm from mitochondria, and translocation of Bax into the mitochondria. 21 In addition to the generation of ROS, AgNPs induced the depletion of Correspondence to: H.-W. Kim; e-mail: [email protected] Contract grant sponsor: Priority Research Centers Program; contract grant number: 2009-0093829 Contract grant sponsor: WCU (World Class University) Program through the National Research Foundation (NRF), Ministry of Education, Science and Technology; contract grant number: R31-10069 V C 2012 WILEY PERIODICALS, INC. 1033
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Size-dependent cellular toxicity of silver nanoparticles

Size-dependent cellular toxicity of silver nanoparticles
Tae-Hyun Kim,1 Meeju Kim,2 Hyung-Seok Park,2 Ueon Sang Shin,1,2 Myoung-Seon Gong,1,2,3
Hae-Won Kim1,2,4
1Institute of Tissue Regeneration Engineering (ITREN), Dankook University, South Korea2Department of Nanobiomedical Science and WCU Research Center, Dankook University, South Korea3Department of Chemistry, Dankook University, South Korea4Department of Biomaterials Science, School of Dentistry, Dankook University, South Korea
Received 6 August 2011; revised 4 November 2011; accepted 29 November 2011
Published online 5 February 2012 in Wiley Online Library (wileyonlinelibrary.com). DOI: 10.1002/jbm.a.34053
Abstract: Silver nanoparticles (AgNPs) have found a variety
of uses including biomedical materials; however, studies of
the cytotoxicity of AgNPs by size effects are only in the
beginning stage. In this study, we examined the size-dependent
cellular toxicity of AgNPs using three different characteristic
sizes (� 10, 50, and 100 nm) against several cell lines includ-
ing MC3T3-E1 and PC12. The cytotoxic effect determined
based on the cell viability, intracellular reactive oxygen
species generation, lactate dehydrogenase release, ultra-
structural changes in cell morphology, and upregulation of
stress-related genes (ho-1 and MMP-3) was fairly size- and
dose-dependent. In particular, AgNPs stimulated apoptosis in
the MC3T3-E1 cells, but induced necrotic cell death in the
PC12 cells. Furthermore, the smallest sized AgNPs (10 nm
size) had a greater ability to induce apoptosis in the MC3T3-
E1 cells than the other sized AgNPs (50 and 100 nm). These
data suggest that the AgNPs-induced cytotoxic effects against
tissue cells are particle size-dependent, and thus, the particle
size needs careful consideration in the design of the nanopar-
ticles for biomedical uses. VC 2012 Wiley Periodicals, Inc. J Biomed
Mater Res Part A: 100A: 1033–1043, 2012.
Key Words: silver nanoparticles, cell toxicity, in vitro, size
dependence, endocytosis
How to cite this article: Kim T-H, Kim M, Park H-S, Shin US, Gong M-S, Kim H-W. 2012. Size-dependent cellular toxicity of silvernanoparticles. J Biomed Mater Res Part A 2012:100A:1033–1043.
INTRODUCTION
Silver nanoparticles (AgNPs) are one of the most widely usednanomaterials in personal care products, dressings as treat-ments for external wounds, ointments, and surgical instru-ments because of their effective antibacterial activity.1–3
Furthermore, the application of AgNPs has been expanded inbiological and medical sciences such as drug and genedelivery vehicles, biosensors, implantable materials, andbone prostheses.4,5 However, these various applicationsincrease human exposure to AgNPs and thus elevate thepotential risk related to their toxicity. For this reason, thetoxicity of AgNPs is considered to be a critical factor whenusing them for biomedical purposes.6,7
Several studies have reported that AgNPs showed signif-icant toxicity against various cell types. For example, AgNPinduced cytotoxicity in macrophages8 and human T cells.9 Adecrease in cell viability and antiproliferative activity inresponse to treatment with AgNPs was also observed inneuron, liver, skin, and lung epithelial cells.10–14 Kim et al.15
reported the subchronic oral toxicity and tissue accumula-tion of AgNPs in rats for 90 days. In their study, histopatho-
logical investigations revealed a higher incidence of bileduct hyperplasia, necrosis, fibrosis, and pigmentation intreated animals. There was also a dose-dependent accumu-lation of silver in tissues. Other studies have shown that theAgNPs can pass through the epithelia of the respiratorytract and across the blood stream directly.16 AgNPs alsocan be taken up by nerve endings and directly accessed tothe CNS.17
It is generally accepted that AgNPs express their toxicitythrough the generation of reactive oxygen species (ROS).13
Increased cellular ROS levels are associated with the induc-tion of genetically programmed cell death (apoptosis), aswell as necrotic cell death in several cell lines.18 Apoptosiseliminates damaged or unwanted cells via nuclear conden-sation, membrane blebbing, and DNA fragmentation in theprocess of development or in response to infection or DNAdamage.19,20 In NIH3T3 fibroblasts, AgNPs were found tohave cytotoxic effects and induce apoptosis, the release ofcytochrome c into the cytoplasm from mitochondria, andtranslocation of Bax into the mitochondria.21 In addition tothe generation of ROS, AgNPs induced the depletion of
Correspondence to: H.-W. Kim; e-mail: [email protected]
Contract grant sponsor: Priority Research Centers Program; contract grant number: 2009-0093829
Contract grant sponsor: WCU (World Class University) Program through the National Research Foundation (NRF), Ministry of Education, Science
and Technology; contract grant number: R31-10069
VC 2012 WILEY PERIODICALS, INC. 1033

antioxidant glutathione (GSH), mitochondrial dysfunction,and the release of lactate dehydrogenase (LDH).11,22–25 LDHis a cytoplasmic enzyme that exists in all major organ sys-tems. When necrotic cell death occurs or cell membranesare damaged, LDH in the cell is released into the extracellu-lar environment; therefore, the extracellular presence ofLDH is used as a marker of cell damage or cell death.26
These studies have mainly highlighted that AgNPs havesignificant toxic effects on cells that primarily occur in adose-dependent manner. However, there have been fewsystematic reports on the size dependence of the cellulartoxicity of AgNPs. One important feature of nano-sizedmaterials such as nanoparticles is that the biologicalresponses to these compounds vary greatly with their size.The particle size of nanoparticles plays a critical role inthe cellular uptake processes from the intracellular mem-brane to the nuclear membrane.27–29 For example, goldnanoparticles enter the HeLa cells via receptor-mediatedendocytosis in a size-dependent manner.30 Yuan et al.31
also reported that the size of hydroxyapatite (HA) nano-particles had a predominant effect on HA nanoparticle-induced cytotoxicity, apoptosis, and the levels of apoptoticproteins in human hepatoma HepG2 cells. Based on cyto-toxicity studies of AgNPs, the size of the AgNPs should bea critical factor involved in determining the cell responses.Examination of the size-dependent cellular toxicity ofAgNPs will provide useful information and rationaleregarding their specific uses because many recent studieshave concentrated on using AgNPs for the delivery vehicleof genes and drugs. As such, studies have focused on thedevelopment of AgNPs with sizes as small as possible,even less than 10 nm, for those purposes.
In this study, we examined the size-dependent cellulartoxicity of AgNPs using three different characteristic sizes. Aseries of cell lines including osteoblastic MC3T3-E1 cellsand PC12 pheochromocytoma cells were used to investigatethe effects of those AgNPs on their biological responses,such as the regulation of proliferation, ROS production, LDHrelease, apoptosis induction, and stress-related gene expres-sion for further biomedical uses of the AgNPs.
MATERIALS AND METHODS
Preparation and characterization of AgNPsAgNPs with sizes of about 10 (AgNPs-10) and 100 nm(AgNPs-100) were purchased from InkTec Co. (Asan, Korea)and Sigma-Aldrich (catalog No. 576832-5G with purity of99.5%), respectively. In addition, 50 nm Ag-NPs (AgNPs-50)were prepared by reduction of an isopropyl alcohol solution(10 mL) of Ag-complex with 2-ethylhexyl amine and 2-ethyl-hexylammonium carbamate (Ag ¼ 10 wt %; InkTec Co.) and0.1 g hydrazine at room temperature. Before use, theAg-complex solution was diluted with 200 mL isopropylalcohol. During slow addition of hydrazine solution in10 mL of isopropyl alcohol, a brown (or dark green) colorslowly appeared, indicating the formation of AgNPs. Afterbeing allowed to stand for several hours, the AgNPs hadprecipitated onto the bottom of the flask. The AgNPs werethen separated by centrifugation and decantation of the
supernatant, after which they were washed with ethanol.Morphological characterization of the purchased and pre-pared AgNPs was conducted by transmission electronmicroscopy (TEM; JEM1010, JEOL, Japan). The size of thenanoparticles was then measured and averaged based onthe TEM images. The phase of the AgNPs was characterizedby X-ray diffraction (XRD; Ultima IV, Rigaku, Japan) usingCuKa radiation at k ¼ 1.54056 Å.
Cell lines and cultureThe mouse preosteoblast cell line, MC3T3-E1, rat adrenalmedulla derived cell line, PC12, human cervical cancer cellline, HeLa, and CHO cells derived from the ovaries ofChinese hamsters were purchased from the American TypeCulture Collection (Rockville, MD) and maintained at 37�Cin an atmosphere of 5% O2 in Dulbecco’s modified Eagle’smedium (Welgene, Korea) containing 10% fetal bovineserum (Gibco) and 1% penicillin-streptomycin.32,33 Themedium was replaced three times per week, and the cellswere passaged at subconfluency. A 100 lL aliquot of thecells prepared at a density of 1 � 105/mL was plated ineach well of 96-well plates. After culture for 24 h, the cul-ture medium was refreshed with that containing AgNPs pre-pared at specific concentrations (10, 20, 40, 80, and 160lg/mL). The final quantities of AgNPs contained in eachsample were thus 1, 2, 4, 8, and 16 lg. After incubation forfurther 24, 48, or 72 h, the cells were collected and ana-lyzed for viability, gene expression, and morphological andbiochemical changes.
Assessment of cell toxicityFollowing each treatment of the AgNPs, 10 lL of the cellcounting kit-8 (CCK-8; Dojindo, Japan) solution was addedto each well and the cells were incubated for another 4 h at37�C. At the end of each treatment, 100 lL of each cell cul-ture supernatant were collected and measured at 450 nmusing an iMark microplate reader (BioRad).
Assay of intracellular ROS levelThe production of ROS was analyzed by flow cytometryusing 20,70-dichlorofluorescein diacetate (H2DCF-DA; Molecu-lar Probes, OR). The hydrogen peroxide (H2O2) produced bycells oxidizes H2DCF-DA to the highly fluorescent compound,20,70-dichlorofluorescein (DCF). In this study, 10 lM DCF-DAwas added to cells that were treated with AgNPs for 48 hand then incubated for 30 min at 37�C. The harvested cellswere then washed twice with phosphate buffered saline(PBS) and measured by FACSCAN flow cytometry (BD Bio-sciences). Data were acquired and analyzed using the Cell-Quest Pro software (BD Biosciences).
LDH release assayThe LDH release assay, which quantifies the LDH releasedby dead cells or cells with damaged membranes into the su-pernatant, was used to quantify the cell cytotoxicity or celldeath. The released LDH was measured using a LDH Cyto-toxicity Detection Kit (Takara Bio, Japan) according to themanufacturer’s guidelines. Briefly, cells previously seeded
1034 KIM ET AL. SIZE-DEPENDENT CELLULAR TOXICITY OF AgNPS

into a 96-well culture plate were treated with AgNPs for48 h. After AgNP treatment, 100 lL of the supernatants ineach well were transferred to a new 96-well plate. LDHassay solution (100 lL) was then added to each well, afterwhich the samples were incubated in the dark for 30 min.The total LDH released was then measured and analyzedusing an iMark microplate reader (Bio-rad). The data arepresented as a percentage of the control. Cells cultured for48 h in the presence of 1% Triton X-100 were used as apositive control.
Quantitative real-time polymerase chain reaction(qRT-PCR) for stress-related gene expressionThe expression of the stress-related genes, heme oxygenase-1 (ho-1) and matrix metalloproteinase-3 (MMP-3), byMC3T3-E1 and PC12 cells treated with different sizedAgNPs for 48 h was confirmed by qRT-PCR. The first strandcDNA was synthesized from the total RNA (2 lg) using aSuperScript first strand synthesis system for qRT-PCR (Invi-trogen) according to the manufacturer’s instructions. Thereaction mixture was made up to 50 lL. Realtime PCR wasconducted using SYBR GreenER qPCR SuperMix reagents(Invitrogen) and a Bio-Rad iCycler. The relative transcriptquantities were calculated using the DDCt method withGAPDH as the endogenous reference gene amplified fromthe samples. The sequences of primer sets used in theexperiment are summarized in Table I.
TEM observationFor the TEM examination, cells treated with AgNPs for 48 hwere harvested and fixed in a mixture of 2% paraformalde-hyde and 2.5% glutaraldehyde with 0.2M PBS for 8 h at pH7.2. Postfixation was conducted using 1% osmium tetroxidein PBS for 2 h. Subsequently, the fixed cells were dehy-drated in ascending concentrations of ethanol (70%, 80%,90%, 95%, and 100%) and embedded in EMbed 812 resin(EMS, PA) via propylene oxide. Ultrathin sections wereobtained from an ultramicrotome (Leica, IL) and were dou-ble stained with uranyl acetate and lead citrate. The stainedsections on the grids were then examined with a H7000TEM (Hitachi, Japan) at 80 kV.
Fluorescein isothiocyanate (FITC)-Annexin V andpropidium iodide double staining for apoptosisdetectionFITC-Annexin V and propidium iodide (PI) double stain wasused to detect the apoptosis induced by AgNPs. The PC12and MC3T3-E1 cells treated with AgNPs for 48 h were har-vested and washed with cold PBS (4�C). The washed cellswere then stained using an FITC Annexin V Apoptosis Detec-tion Kit (BD Pharmingen, NJ). Briefly, the cells were resus-pended in 1 mL of 1� binding buffer at a concentration of1 � 106 cells/mL. Next, 5 lL of FITC Annexin V and 5 lL ofPI were added per 100 lL of the cell suspension (1 � 105
cells). After gentle vortexing, the cells were incubated for 15min at room temperature in the dark. Subsequently, 400 lLof 1� binding buffer was added to each tube and analyzedusing a FACSCAN flow cytometer (BD Biosciences). The dataacquired for 15,000 cells in each sample were analyzedusing the CellQuest Pro software (BD Biosciences).
Statistical analysisThree independent experiments were conducted, and theresults were expressed as the means and one standard devi-ations from the triplicate experiments. Statistical analysiswas performed by one-way analysis of variance and p-val-ues less than 0.05 were considered significant.
RESULTS
Characteristics of AgNPsThe morphologies of AgNPs with three different sizes wereobserved by TEM, as shown in Figure 1(A). The size distri-bution of the nanoparticles is shown in the inset. The AgNPsshowed relatively narrow distributions and a clear differ-ence in their sizes based on the TEM images from a smallsample population. The average sizes calculated from theTEM images (as summarized in Table II) were 8.6 (63.2),46 (68.0), and 75 (626) nm, respectively, for AgNP-10,AgNP-50, and AgNP-100. AgNP-10 and AgNP-50 were spher-ical in shape although a fraction (�10%) of the AgNP-100was somewhat elongated. All the AgNPs showed a typicalsilver phase based on the XRD pattern. A characteristic XRDpattern on AgNP-50 is shown in Figure 1(B), which revealeddiffraction peaks at 2y values of 38.00� , 44.4� , 64.5� , and77.5�. These values can be readily indexed to (111), (200),(220), and (310) reflections of silver crystal.
Effect of the AgNPs on cytoxicity in different cell linesThe effects of AgNPs with three different sizes and varyingdoses (from 10 to 160 lg/mL) on the cellular toxicity wereexamined in the MC3T3-E1, PC12, HeLa, and CHO cell lines.Cell proliferation assay was conducted at 24, 48, and 72 hafter treatment with AgNP-10, 50, and 100. Data arepresented with respect to the control group without thenanoparticles treatment. Clear size- and dose-dependent cel-lular toxicity was observed for all tested cell lines (Fig. 2).In particular, MC3T3-E1 and PC12 cells were shown to bemore sensitive to the change in dose during prolonged cultureperiods. Moreover, the largest nanoparticles (AgNP-100)
TABLE I. Primer Sequences Used in this Study
GeneName
Primer Sequence (50–30)
MC3T3-E1 Cell PC12 Cell
ho-1 GCCTGCTAGCCTGGTGCAAG
GCCTGCTAGCCTGGTGCAAG
AGCGGTGTGATGAGCTACTGG
AGCGGTGTCTGGGATGAGCTA
MMP-3 TGGAGATGCTCACTTTGACG
TTGTCCTTCGATGCAGTCAG
GCCTTGGCTGAGTGGTAGAG
AGACGGCCAAAATGAAGAGA
GAPDH GGGAAGCCCATCACCATCTT
AACCTGCCAAGTATGATG
CGGCCTCACCCCATTTG
GGAGTTGCTGTTGAAGTC
ORIGINAL ARTICLE
JOURNAL OF BIOMEDICAL MATERIALS RESEARCH A | APR 2012 VOL 100A, ISSUE 4 1035

showed the highest toxicity against cells, particularly MC3T3-E1 and PC12.
Analysis of ROS production in MC3T3-E1 and PC12 cellsThe cellular toxicity of the AgNPs was also examined basedon the ROS generation of the cells, particularly for theMC3T3-E1 and PC12 cell lines. Both cells were treated withvarious sized AgNPs at concentrations of 10, 20, 40, and80 lg/mL for 48 h, and the ROS generating cells were thenassessed by flow cytometry. The nanoparticle treated groupsshowed significantly increased populations of cells generat-ing ROS (Fig. 3). Additionally, the ROS production increasedas the size and concentration of the AgNPs increased.
Specifically, when the MC3T3-E1 cells were treated withAgNP-100 at 80 lg/mL, the ROS generation was as high as64.3%, while the ROS production of cells with AgNP-10 andAgNP-50 was 52.9% and 61%, respectively.
Analysis of cell toxicity by LDH activity assayThe cytotoxicity of the AgNPs was examined by an assay ofthe released LDH activity of the MC3T3-E1 and PC12 celllines [Fig. 4(A,B), respectively]. When the nanoparticlesdose was low (10 and 20 lg/mL), the LDH activity levels ofthe nanoparticles of all three sizes were in the range of105%–120% of the negative control. When the dose washigh (40 and 80 lg/mL), particularly in the large-sized
FIGURE 1. (A) TEM images of AgNPs with three different sizes and (B) the characteristic XRD pattern of AgNPs used in the experiment.
1036 KIM ET AL. SIZE-DEPENDENT CELLULAR TOXICITY OF AgNPS

nanoparticles (AgNP-100), the LDH levels increased signifi-cantly to 150%–190% of that of the negative control, whichwere close to the values of the positive control. The LDHactivity also demonstrated that both cell lines responded tothe AgNPs in a size- and dose-dependent fashion.
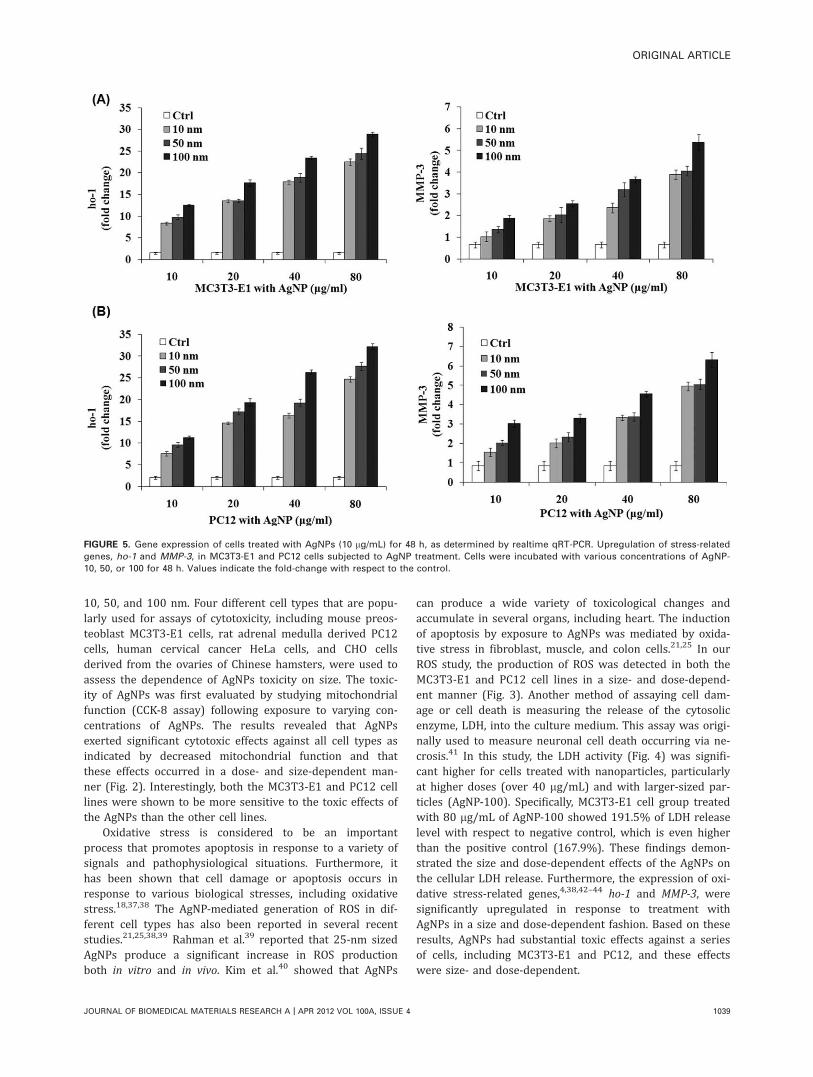
QRT-PCR analysis of stress-responsive genesThe cytotoxic effects of the AgNP were further estimated bythe expression of genes. It is well known that nanomaterialsinduce several stress-response genes that are related tooxidative stress and protein homeostasis. We analyzed theho-1 and MMP-3 genes, which are typically involved in thesebiological responses. AgNPs led to upregulated ho-1 andMMP-3 expression in a dose and size-dependent manner forthe MC3T3-E1 cell line [Fig. 5(A)]. A similar gene inductionpattern was observed in PC12 cells [Fig. 5(B)], althoughthe stress-related gene induction was slightly higher in thisline.
TABLE II. Average Particle Sizes of the AgNPs Calculated
Based on the TEM Observation
AgNP-10 AgNP-50 AgNP-100
Mean 6 std 8.6 6 3.2 46 6 8.0 75 6 26
FIGURE 2. Cytotoxicity of the AgNPs, as determined using a CCK-8 kit. MC3T3-E1, PC12, HeLa and CHO cells were treated with AgNP-10, 50,
and 100 at varying concentrations (from 10 to 160 lg/mL) for different lengths of time (24, 48, and 72 h). Cell toxicity was found to be size- and
dose-dependent in all test cell lines. Data represent the means 6 one standard deviation for n ¼ 4.
ORIGINAL ARTICLE
JOURNAL OF BIOMEDICAL MATERIALS RESEARCH A | APR 2012 VOL 100A, ISSUE 4 1037

Ultrastructural characteristics of AgNP-treated cellsThe ultrastructural changes in MC3T3-E1 and PC12 cellstreated with 10 lg/mL AgNPs for 48 h were observed byTEM (Fig. 6). The results revealed that many AgNPs wereaccumulated in the cytoplasm of both cell types. The AgNPsseemed to induce apoptosis in the MC3T3-E1 cells[Fig. 6(A)] because cells were shown to have nucleus bleb-bing and fragmentation, a typical feature of apoptotic cells.The large vacuoles appeared as a result of the destructionof cell organelles, but the cytoplasm structure and cellmembrane was maintained until the cells died. In contrast,the membranes and a number of organelles were disruptedin PC12 cells [Fig. 6(B)].
Size-dependent apoptosis induction of AgNPsTo assess the apoptosis induced by the AgNPs, the treatedcells were double stained with FITC Annexin V and PI andthen analyzed by flow cytometry. Externalization of phos-phatidyl serine (PS) from the inner to the outer cell mem-brane is one of the strongest indicators of early apoptosis.Therefore, Annexin V, a calcium-dependent phospholipid-binding protein with a high affinity for PS, can be used asan effective marker for the apoptosis. PI, a nonspecific DNAintercalating agent, can detect necrotic cells because it ispermeable only through the damaged cell membranes but isexcluded by the membranes of living cells or early apoptoticcells. As shown in Figure 7(A), almost no apoptotic cellswere detected in the untreated control MC3T3-E1 group.However, there were increases of about 5.05%, 2.49%, and1.11% with respect to the control in the early apoptoticpopulation of MC3T3-E1 cells treated with 10 lg/mL AgNP-
10, 50, and 100, respectively, demonstrating that the induc-tion of apoptosis was greater in the smaller AgNPs. Con-versely, the percentage of apoptotic cells was not signifi-cantly higher in the PC12 cells subjected to any of the AgNPtreatments [Fig. 7(B)].
DISCUSSION
With the considerable development of nanomaterials target-ing medical applications, there has been general concernregarding their potential toxicity based on the anticipatedhealth risk associated with ultrafine and nano-sized par-ticles.34 Thus, it is important to investigate the toxicity andthe mechanism of action by which such toxicity occurs.Although the therapeutic use of AgNPs is still in the initialphase, the antimicrobial effects of silver ions (e.g., AgNO3)have been exploited in medical treatments for decades.Silver ions have primarily been applied topically to preventor cure infections; consequently, investigations of its toxicityhave focused on dermal exposure. However, AgNPs haverecently been studied in terms of drug delivery and target-ing, which would involve intracellular application of theAgNPs.35,36 Owing to its potential use as a tool for drugdelivery, it is crucial to investigate the effects of AgNPs ontissue cells.
Furthermore, it is well documented that the size ofnanoparticles has great effects on their cellular uptake andensuing functional impact.31 In this study, we conducted adetailed investigation of the cytotoxic effects of the size ofAgNPs using three different sizes; with averages of about
FIGURE 3. Effects of AgNPs on ROS production in the cell line of (A)
MC3T3-E1 and (B) PC12 during treatment for 48 h. Flow cytometric
analysis of the cells in response to AgNPs showed the generation of
ROS dose- and size-dependently.
FIGURE 4. Effects of AgNPs on LDH production in the cell line of (A)
MC3T3-E1 and (B) PC12 during treatment for 48 h. Flow cytometric
analysis of cells in response to treatment with AgNPs showed that
the generation of LDH was dose- and size-dependent.
1038 KIM ET AL. SIZE-DEPENDENT CELLULAR TOXICITY OF AgNPS
10, 50, and 100 nm. Four different cell types that are popu-larly used for assays of cytotoxicity, including mouse preos-teoblast MC3T3-E1 cells, rat adrenal medulla derived PC12cells, human cervical cancer HeLa cells, and CHO cellsderived from the ovaries of Chinese hamsters, were used toassess the dependence of AgNPs toxicity on size. The toxic-ity of AgNPs was first evaluated by studying mitochondrialfunction (CCK-8 assay) following exposure to varying con-centrations of AgNPs. The results revealed that AgNPsexerted significant cytotoxic effects against all cell types asindicated by decreased mitochondrial function and thatthese effects occurred in a dose- and size-dependent man-ner (Fig. 2). Interestingly, both the MC3T3-E1 and PC12 celllines were shown to be more sensitive to the toxic effects ofthe AgNPs than the other cell lines.
Oxidative stress is considered to be an importantprocess that promotes apoptosis in response to a variety ofsignals and pathophysiological situations. Furthermore, ithas been shown that cell damage or apoptosis occurs inresponse to various biological stresses, including oxidativestress.18,37,38 The AgNP-mediated generation of ROS in dif-ferent cell types has also been reported in several recentstudies.21,25,38,39 Rahman et al.39 reported that 25-nm sizedAgNPs produce a significant increase in ROS productionboth in vitro and in vivo. Kim et al.40 showed that AgNPs
can produce a wide variety of toxicological changes andaccumulate in several organs, including heart. The inductionof apoptosis by exposure to AgNPs was mediated by oxida-tive stress in fibroblast, muscle, and colon cells.21,25 In ourROS study, the production of ROS was detected in both theMC3T3-E1 and PC12 cell lines in a size- and dose-depend-ent manner (Fig. 3). Another method of assaying cell dam-age or cell death is measuring the release of the cytosolicenzyme, LDH, into the culture medium. This assay was origi-nally used to measure neuronal cell death occurring via ne-crosis.41 In this study, the LDH activity (Fig. 4) was signifi-cant higher for cells treated with nanoparticles, particularlyat higher doses (over 40 lg/mL) and with larger-sized par-ticles (AgNP-100). Specifically, MC3T3-E1 cell group treatedwith 80 lg/mL of AgNP-100 showed 191.5% of LDH releaselevel with respect to negative control, which is even higherthan the positive control (167.9%). These findings demon-strated the size and dose-dependent effects of the AgNPs onthe cellular LDH release. Furthermore, the expression of oxi-dative stress-related genes,4,38,42–44 ho-1 and MMP-3, weresignificantly upregulated in response to treatment withAgNPs in a size and dose-dependent fashion. Based on theseresults, AgNPs had substantial toxic effects against a seriesof cells, including MC3T3-E1 and PC12, and these effectswere size- and dose-dependent.
FIGURE 5. Gene expression of cells treated with AgNPs (10 lg/mL) for 48 h, as determined by realtime qRT-PCR. Upregulation of stress-related
genes, ho-1 and MMP-3, in MC3T3-E1 and PC12 cells subjected to AgNP treatment. Cells were incubated with various concentrations of AgNP-
10, 50, or 100 for 48 h. Values indicate the fold-change with respect to the control.
ORIGINAL ARTICLE
JOURNAL OF BIOMEDICAL MATERIALS RESEARCH A | APR 2012 VOL 100A, ISSUE 4 1039

To investigate the interaction of AgNP with cells, localiza-tion studies of AgNPs were conducted by TEM after exposingthe cells to AgNPs at various sizes and doses. Ultra-thin sec-tions of AgNPs-treated cells visualized under TEM showedthe presence of electron dense, aggregated regions in thecytoplasm (Fig. 6). Lopotko et al.45 reported that interactionof nanoparticles with cells leads to the formation of nanopar-ticle clusters in the cytoplasm. The most active redox cellorganelles and nanoparticles that are taken up into cells areprimarily localized to mitochondria.46,47 The presence ofAgNPs inside the mitochondria could lead to alterations inROS production, thereby interfering with cellular antioxidantdefenses.25
The ultrastructural characteristics of apoptosis is mainlyfeatured by cell shrinkage, condensation of the chromatin tothe margins of the nuclear membrane, nuclear breakdown,cell membrane blebbing, and eventual fragmentation of thecell into apoptotic bodies that are engulfed by neighboringcells or phagocytes.48 Conversely, the changes in the organ-elles are known to be relatively minor. Necrosis, on theother hand, occurs when the cell is sacrificed and is charac-terized by cell swelling, formation of microvesicles, andbreakage of the cell membrane, which allows leakage of thecytoplasm into the intercellular space.49 In this study, theultrastructure of the MC3T3-E1 cells treated with AgNPsshowed chromatin condensation and nucleus blebbing andfragmentation. In contrast, AgNPs induced necrosis in thePC12 cells. These findings indicate that the cytotoxic effectsof nanoparticles need to be differently interpreted based onwhether they occur via apoptosis or necrosis.
It is well known that oxidant stress can lead to apopto-sis or necrosis.50 During apoptosis, ROS is produced, whichprecedes the loss of mitochondria membrane potential, therelease of cytochrome c, the activation of caspase 3, and theinduction of mitochondrial-dependent apoptosis. Thus, ROSseems to play an essential role in the upstream of mitochon-dria.51 To assess the mode of cell death induced by AgNPs,namely whether it was apoptotic or necrotic, we stained thetreated cells with FITC annexin V and PI and then subjectedthem to flow cytometric analysis. FACS revealed that theAgNPs induced apoptosis in MC3T3-E1 cells, primarily inresponse to the AgNP-10, although some apoptotic cellswere also observed in response to treatment with AgNP-50and AgNP-100. Moreover, the apoptotic behavior did notseem to be dose-dependent except for the case in largestnanoparticles (AgNP-100). Conversely, AgNPs primarilyinduced necrosis in the PC12 cells, without showing any sig-nificant levels of apoptosis. The induction of apoptosis byAgNPs was previously reported in mouse NIH3T-3 cells.21
Furthermore, apoptosis-related genes were found to bealtered in the liver of mice that were treated with AgNPs.52
Similarly, the AgNPs used herein are believed to be involvedin the mechanism of apoptosis in the mouse-derivedMC3T3-E1 cells.
With regard to the size-dependent apoptotic action, it isbelieved that the small nanoparticles are likely taken upthrough the intracellular membrane to the nucleus pores,which differs from the uptake process in larger particles.Several previous studies of the correlation between nano-particle size and cellular uptake have demonstrated thatnanoparticles with a diameter of 50 nm are readily takenup by mammalian cells at higher rates and concentrationsthan other sizes via an endocytotic mechanism.27,53 How-ever, when the nanoparticles are smaller than 50 nm, theytend to form clusters and aggregate during cellular uptake.For nanoparticles larger than 50 nm, the slower receptordiffusion rate was found to lead to much fewer nanopar-ticles being taken up by cells.27 Furthermore, living cellshave nuclear pore complexes (NPCs) in the nuclear mem-brane, and the diameter of NPCs is largely 20–50 nm, whichvaries depending on cell type.54,55 To enter the cell nucleus,
FIGURE 6. Ultrastructural characteristics of (A) MC3T3-E1 and (B)
PC12 cells treated with AgNPs (10 lg/mL) for 48 h. Untreated MC3T3-
E1 cells (A-a) have a rounded nucleus (N) and cell organelles in the
cytoplasm. Treatment with 10 lg/mL of AgNP-10 (A-b), 50 (A-c), or
100 (A-d) resulted in the accumulation of AgNPs in the cytoplasm
(arrows) and typical apoptotic characteristics including chromatin con-
densation, nucleus blebbing, and vacuole (V) formation. Control PC12
cells (B-a) showed rounded nuclei and several cell organelles in the
cytoplasm. Treatment with 10 lg/mL of AgNP-10 (B-b), 50 (B-c), or
100 (B-d) resulted in the accumulation of AgNPs in the cytoplasm
(arrows) and necrosis features. AgNP treated cells showed cleaved or
chromatin condensed nucleus and degraded organelles and numbers
of vacuoles (B-c and d). Cell membrane was ruptured, and cell organ-
elles were destroyed in the AgNP-100 treated PC12 cells (B-d). Scale
bar indicates 4 lm.
1040 KIM ET AL. SIZE-DEPENDENT CELLULAR TOXICITY OF AgNPS

nanoparticles should be small enough to pass the NPCs onthe nuclear membrane. Pante and Kann56 have found thatgold particles with a diameter close to 39 nm could betranslocated into the cell nucleus. Li et al.57 reported thatAgNPs with a diameter of around 5 and 20 nm elevated theROS level and induced apoptosis in human cells, but thosewith diameters greater than 50 nm could not do so. Like-wise, the significantly upregulated apoptosis in the AgNP-10, but not in the AgNP-50 or AgNP-100 treated cells, isalso understandable.
These findings indicate that the use of nanoparticles asa drug delivery vehicle should be carefully designed, partic-ularly in the context of their size. This also corresponds tothe development of AgNPs. According to the target sites, i.e.,the nucleus or cytoskeletal organelles, the sizes should beeither decreased to a few tens of nanometers or maintainedat sizes that do not readily pass through the nuclearmembrane. The nucleus is a desirable target because of thepresence of genomic DNA. In addition, numerous nuclearproteins actively participate in several key cellular processes,such as DNA replication, recombination, RNA transcription,DNA damage and repair, genomic alterations, and cell cyclecontrol.58 Therefore, the smaller sized nanoparticles at agiven composition may be more effectively delivered to the
nucleus. However, this should be discussed in the same con-text as the cytotoxic action of the nanoparticles in case theyelicit substantial toxicity as AgNPs do. The possible geneticdisorders manipulated by the nucleus-translocated ultrafine-sized toxic nanoparticles are not so satisfactory althoughthey seemingly elicit less cellular toxicity that can be inter-preted from the routine cell viability tools. This study high-lights the importance of the size of AgNPs, not just theirdose, on the cytotoxicity mechanisms. Furthermore, theresults presented here indicate that the cytotoxicity needs tobe interpreted using appropriate assay tools.
CONCLUSIONS
We demonstrated that AgNPs with sizes of approximately10, 50, and 100 nm have the potential to exert cytotoxiceffects against several cell lines, including MC3T3-E1 andPC12. The cellular toxicity was confirmed by the cell viabil-ity, ROS generation, LDH release, and expression of stress-related genes (ho-1 and MMP-3) and was found to be fairlysize- and dose-dependent. In particular, the cytotoxicresponse to AgNPs was cell-dependent, with apoptosisoccurring in MC3T3-E1 and necrosis in PC12. Moreover, thesmallest sized AgNPs (10 nm size) had a greater apoptoticeffect against the MC3T3-E1 cells than other-sized AgNPs
FIGURE 7. Induction of apoptosis in MC3T3-E1 and PC12 cells as determined by Annexin V-FITC and PI double staining. (A) Representative dot
plots of Annexin V/PI staining of MC3T3-E1 cells untreated (control) and treated with 10 lg/mL of AgNP-10, 50, and 100. Lower left quadrant con-
tains the vital (double negative) population. Lower right quadrant contains the early apoptotic (Annexin V positive/PI negative) population.
Upper left quadrant contains the necrotic cell (Annexin V negative/PI positive) population and upper right contains the apoptotic population. (B)
Percentage of the apoptotic MC3T3-E1 and PC12 cells after treatment with different sizes and concentrations of AgNP as measured by Annexin
V-FITC and PI double staining.
ORIGINAL ARTICLE
JOURNAL OF BIOMEDICAL MATERIALS RESEARCH A | APR 2012 VOL 100A, ISSUE 4 1041

(50 and 100 nm). It is believed that the difference in theeffects of these particles is associated in part with theircellular uptake processes from the cell membranes to thenuclear pores. These findings suggest that the design ofnanomaterials including AgNPs requires special care, partic-ularly with respect to their size, to gain satisfactory actionsfor target sites as well as to reduce cellular toxicity.
ACKNOWLEDGMENT
The kind support of IBST in Dankook University isacknowledged.
REFERENCES1. Lewinski N, Colvin V, Drezedk R. Cytotoxicity of nanoparticles.
Small 2008;4:26–49.
2. Faunce T, Watal A. Nanosilver and global public health: Interna-
tional regulatory issues. Nanomedicine 2010;5:617–632.
3. Margaret IP, Lui SL, Poon VK, Lung I, Burd A. Antimicrobial activ-
ities of silver dressings: An in vitro comparison. J Med Microbiol
2006;55:59–63.
4. Miura N, Shinohara Y. Cytotoxic effects and apoptosis induction
by silver nanoparticles in Hela cells. Biochem Biophys Res Com-
mun 2009;390:733–737.
5. Ahmed M, Alsalhi MS, Siddiqui MK. Silver nanoparticle applica-
tions and human health. Clin Chim Acta 2010;411:1841–1848.
6. Park E-J, Yi J, Kim Y, Choi K, Park K. Silver nanoparticles induce
cytotoxicity by a Trojan-horse type mechanism. Toxicol In Vitro
2010;24:872–878.
7. Yon JN, Lead JR. Manufactured nanoparticles: An overview of
their chemistry, interactions and potential environmental implica-
tions. Sci Total Environ 2008;400:396–414.
8. Yen HJ, Hsu SH, Tsai CL. Cytotoxicity and immunological
responses of gold and silver nanoparticles of different sizes.
Small 2009;5:1553–1561.
9. Eom HJ, Choi J. p38 MAPK activation, DNA damage, cell cycle
arrest and apoptosis as mechanisms of toxicity of silver nanopar-
ticles in Jurkat T cells. Environ Sci Technol 2010;44:8337–8342.
10. Yang Z, Liu ZW, Allaker RP, Reip P, Oxford J, Ahmad Z, Ren G.
A review of nanoparticle functionality and toxicity on the central
nervous system. J R Soc Interface 2010;7:411–422.
11. Hussain SM, Hess KL, Gearhart JM, Geiss KT, Schlager JJ. In
vitro toxicity of nanoparticles in BRL 3A rat liver cells. Toxicol In
Vitro 2005;19:975–983.
12. Samberg ME, Oldenburg SJ, Monteiro-Riviere NA. Evaluation of
silver nanoparticle toxicity in skin in vivo and keratinocytes in
vitro. Environ Health Perspect 2010;118:407–413.
13. Foldbjerg R, Olesen P, Hougaard M, Dang DA, Hoffmann HJ,
Autrup H. PVP-coated silver nanoparticles and silver ions induce
reactive oxygen species, apoptosis and necrosis in THP-1 mono-
cytes. Toxicol Lett 2009;190:156–162.
14. Asharani PV, Hande MP, Valiyaveettil S. Anti-proliferative activity
of silver nanoparticles. BMC Cell Biol 2009;17:10–65.
15. Kim YS, Song MY, Park JD, Song KS, Ryu HR, Chung YH, Chang
HK, Lee JH, Oh KH, Kelman BJ, Hwang IK, Yu IJ. Subchronic oral
toxicity of silver nanoparticles. Part Fibre Toxicol 2010;7:20–31.
16. Medina C, Santos-Martinez MJ, Radomski A, Corrigan OI, Radom-
ski MW. Nanoparticles: Pharmacological and toxicological signifi-
cance. Br J Pharmacol 2007;150:552–558.
17. Oberd€orster G, Sharp Z, Atudorei V, Elder A, Gelein R, Kreyling
W, Cox C. Translocation of inhaled ultrafine particles to the brain.
Inhal Toxicol 2004;16:437–445.
18. Ott M, Gogvadze V, Orrenius S, Zhivotovsky B. Mitochondria,
oxidative stress and cell death. Apoptosis 2007;12:913–922.
19. Chipuk JE, Bouchier-Hayes L, Kuwana T, Newmeyer DD, Green
DR. PUMA couples the nuclear and cytoplasmic proapoptotic
function of p53. Science 2005;309:1732–1735.
20. Gopinath P, Gogoi SK, Sanpui P, Paul A, Chattopadhyay A, Ghosh
SS. Signaling gene cascade in silver nanoparticle induced apopto-
sis. Colloids Surf B 2010;77:240–245.
21. Hsin YH, Chen CF, Huang S, Shih TS, Lai PS, Chueh PJ. The apopto-
tic effects of nanosilver is mediated by a ROS- and JNK-dependent
mechanism involving the mitochondrial pathway in NIH3T3 cells.
Toxicol Lett 2008;179:130–139.
22. Carlson C, Hussain SM, Schrand AM, Braydich-Stolle LK, Hess KL,
Jones RL, Schlager JJ. Unique cellular interaction of silver nano-
particles: Size dependent generation of reactive oxygen species. J
Phys Chem B 2008;112:13608–13619.
23. Nowrouzi A, Meghrazi K, Golmohammadi T, Golestani A, Ahma-
dian S, Shafiezadeh M, Shajary Z, Khaghani S, Amiri AN. Cytotox-
icity of subtoxic AgNP in human hepatoma cell line (HepG2) after
long-term exposure. Iran Biomed J 2010;14:23–32.
24. Arora S, Jain J, Rajwade JM, Paknikar KM. Cellular responses
induced by silver nanoparticles: In vitro studies. Toxicol Lett 2008;
179:93–100.
25. Arora S, Jain J, Rajwade JM, Paknikar KM. Interactions of silver
nanoparticles with primary mouse fibroblasts and liver cells. Toxi-
col Appl Pharmacol 2009;236:310–318.
26. Wang Q, Zheng XL, Yang L, Shi F, Gao LB, Zhong YJ, Sun H, He
F, Lin Y, Wang X. Reactive oxygen species-mediated apoptosis
contributes to chemosensitization effect of saikosaponins on cis-
platin-induced cytotoxicity in cancer cells. J Exp Clin Cancer Res
2010;29:159–167.
27. Chithrani BD, Chan WCW. Elucidating the mechanism of cellular
uptake and removal of protein-coated gold nanoparticles of differ-
ent sizes and shapes. Nano Lett 2007;7:1542–1550.
28. Jiang W, Kim BYS, Rutka JT, Chan WCW. Nanoparticle-mediated
cellular response is size-dependent. Nat Nanotechnol 2008;3:
145–150.
29. Chen M, Mikecz A. Formation of nucleoplasmic protein aggre-
gates impairs nuclear function in response to SiO2 nanoparticles.
Exp Cell Res 2005;305:51–62.
30. Chithrani BD, Ghazani AA, Chan CW. Determining the size and
shape dependence of gold nanoparticle uptake into mammalian
cells. Nano Lett 2006;6:662–668.
31. Yuan Y, Liu C, Qian J, Wang J, Zhang Y. Size-mediated cytotoxicity
and apoptosis of hydroxyapatite nanoparticles in human hepatoma
HepG2 cells. Biomaterials 2010;31:730–740.
32. Oh SA, Kim SH, Won JE, Kim JJ, Shin US, Kim HW. Effects ongrowth and osteogenic differentiation of mesenchymal stem cellsby the zinc-added sol-gel glass granules. J Tissue Eng 2010;2010:Article ID 475260, 10 p.
33. Jin GZ, Kim M, Shin US, Kim HW. Effect of carbon nanotube
coating of aligned nanofibrous polymer scaffolds on the neurite
outgrowth of PC-12 cells. Cell Biol Int 2011;35:741–745.
34. Oberdorster G, Oberdorster E, Oberdorster J. Nanotoxicology: An
emerging discipline evolving from studies of ultrafine particles.
Environ Health Perspect 2005;113:823–839.
35. Boucher W, Stern JM, Kotsinyan V, Kempuraj D, Papaliodis D,
Cohen MS, Theoharides TC. Intravesical nanocrystalline silver
decreases experimental bladder inflammation. J Urol 2008;179:
1598–1602.
36. Gopinath P, Gogoi SK, Chattopadhyay A, Ghosh SS. Implications
of silver nanoparticle induced cell apoptosis for in vitro gene ther-
apy. Nanotechnology 2008;19:75–104.
37. Rana SV. Metals and apoptosis: Recent developments. J Trace
Elem Med Biol 2008;22:262–284.
38. Ueda S, Masutani H, Nakamura H, Tanaka T, Ueno M, Yodoi J.
Redox control of cell death. Antioxid Redox Signal 2002;2004:
405–414.
39. Rahman MF, Wang J, Patterson TA, Saini UT, Robinson BL, New-
port GD, Murdock RC, Schlager JJ, Hussain SM, Ali SF. Expres-
sion of genes related to oxidative stress in the mouse brain after
exposure to silver-25 nanoparticles. Toxicol Lett 2009;187:15–21.
40. Kim YS, Kim JS, Cho HS, Rha DS, Kim JM. Twenty-eight-day oral
toxicity, genotoxicity and gender-related tissue distribution of sil-
ver nanoparticles in Sprague–Dawley rats. Inhal Toxicol 2008;20:
575–583.
41. Lobner D. Comparison of the LDH and MTT assays for quantifying
cell death: Validity for neuronal apoptosis? J Neurosci Methods
2000;96:147–152.
42. Kondoh M, Inoue Y, Atagi S, Futakawa N, Higashimoto M, Sato
M. Specific induction of metallothionein synthesis by mitochon-
drial oxidative stress. Life Sci 2001;69:2137–2146.
1042 KIM ET AL. SIZE-DEPENDENT CELLULAR TOXICITY OF AgNPS

43. Sato M, Bremner I. Oxygen free radicals and metallothionein.
Free Radic Biol Med 1993;14:325–337.
44. Takahashi T, Morita K, Akagi R, Sassa S. Heme oxygenase-1: A
novel therapeutic target in oxidative tissue injuries. Curr Med
Chem 2004;11:1545–1561.
45. Lopotko DO, Lukianova-Hleb EY, Oraevsky AO. Clusterization of
nanoparticles during their interaction with living cells. Nanomedi-
cine 2007;2:241–253.
46. Foley S, Crowley C, Smaihi M, Bonfils C, Erlanger BF, Seta P.
Cellular localisation of a water-soluble fullerene derivative.
Biochem Biophys Res Commun 2002;294:116–119.
47. Li N, Sioutas C, Cho A, Schmitz D, Misra C, Sempf J. Ultrafine
particulate pollutants induce oxidative stress and mitochondrial
damage. Environ Health Perspect 2003;111:455–460.
48. Urne AG, Vaux DL. Molecular and clinical aspects of apoptosis.
Pharmacol Ther 1996;72:37–50.
49. Boulares AH, Yakovlev AG, Ivanova V, Stoica BA, Wang G, Iyer S.
Role of poly-(ADP-ribose) polymerase (PARP) cleavage in apopto-
sis. Caspase-3 resistant PARP mutant increases rates of apoptosis
in transfected cells. J Biol Chem 1999;274:22932–22940.
50. Curtin JF, Donovan M, Cotter TG. Regulation and measurement
of oxidative stress in apoptosis. J Immunol Methods 2002;265:
49–72.
51. Herrera B, Alvarez AM, S�anchez A, Fern�andez M, Roncero C,
Benito M, Fabregat I. Reactive oxygen species (ROS) mediates the
mitochondrial-dependent apoptosis induced by transforming
growth factor b in fetal hepatocytes. FASEB J 2001;15:741–751.
52. Cha K, Hong HW, Choi YG, Lee MJ, Park JH, Chae HK, Ryu G,
Myung H. Comparison of acute responses of mice livers to short-
term exposure to nano-sized or micro-sized silver particles. Bio-
technol Lett 2008;30:1893–1899.
53. Shukla R, Bansal V, Chaudhary M. Biocompatibility of gold nano-
particles and their endocytotic fate inside the cellular compart-
ment: A microscopic overview. Langmuir 2005;21:10644–10654.
54. Fahrenkrog B, Aebi U. The nuclear pore complex: Nucleocytoplas-
mic transport and beyond. Nat Rev Mol Cell Biol 2003;4:757–766.
55. Wente SR. Gatekeepers of the nucleus. Science 2000;288:1374–
1377.
56. Pante N, Kann M. Nuclear pore complex is able to transport macro-
molecules with diameters of -39 nm. Mol Biol Cell 2002;13:425–434.
57. Li Y, Tian X, Lu Z, Yang C, Yang G, Zhou X, Yao H, Zhu Z, Xi Z,
Yang X. Mechanism for alpha-MnO2 nanowire-induced cytotoxic-
ity in Hela cells. J Nanosci Nanotechnol 2010;10:397–404.
58. Koh JY, Choi DW. Quantitative determination of glutamate medi-
ated cortical neuronal injury in cell culture by lactate dehydrogen-
ase efflux assay. J Neurosci Methods 1987;20:83–90.
ORIGINAL ARTICLE
JOURNAL OF BIOMEDICAL MATERIALS RESEARCH A | APR 2012 VOL 100A, ISSUE 4 1043