
Simultaneous evaluation of clastogenicity, aneugenicity and toxicity in the mouse micronucleus assay...
9
Mutation Research, 282 (1992) 159-167 159 © 1992 Elsevier Science Publishers B.V. All rights reserved 0165-7992/92/$05.00 MUTLET 00674 Simultaneous evaluation of clastogenicity, aneugenicity and toxicity in the mouse micronucleus assay using immunofluorescence G. Krishna, R. Fiedler and J.C. Theiss Molecular Toxicology Section, Department of Pathology and Experimental Toxicology, Parke-Davis Pharmaceutical Research Division, Warner-Lambert Company, Ann Arbor, MI 48105, USA (Received 19 February 1992) (Accepted 2 March 1992) Keywords: Antikinetochore antibody technique; Clastogenicity evaluation; Aneugenicity evaluation; Toxicity evaluation; Micronu- cleus assay; Mouse; Aneuploidy; Cyclophosphamide; Vincristine Summary An improved antikinetochore antibody technique was established in the mouse micronucleus assay to simultaneously evaluate toxicity, clastogenicity and aneugenicity induced by various test agents. The procedure involved the use of cellulose column fractionated cytospun slides for analysis. The staining method consisted of sequential treatment of slides with crest serum, fluorosceinated goat-antihuman and swine-antigoat antibodies, and propidium iodide. In this method, polychromatic erythrocytes (PCEs, dark red), normochromatic erythrocytes (NCEs, green), chromosome(s)/fragments/micronuclei (orange), and kinetochores (yellow), are identified using the same filter setting under blue excitation (440-490 nm) with a barrier filter at 520 nm. Using this method, three agents, cyclophosphamide, X-rays and vincristine were tested for micronucleus/aneuploidy induction and bone marrow toxicity. The aneugen, vincristine, and clastogens, X-rays and cyclophosphamide, induced predominantly kinetochore positive (K + ) and negative (K-) micronucleated PCEs, respectively. At the doses tested, cyclophosphamide caused a slight but statistically significant decrease in PCEs in females, and other agents did not produce any severe bone-marrow toxicity in either male or female mice. These results are comparable with the results reported in the literature on these compounds with various methods and thus demonstrate the usefulness of this assay in distinguishing clastogenicity from aneugenicity and in evaluating toxicity. Short-term tests have been widely used in the genotoxicity evaluation of test compounds. The in vivo micronucleus assay has played a prominent role in mutagenicity testing (Heddle et al., 1983; Arlett et al., 1984; Mavournin et al., 1990; Ashby, Correspondence Dr. C. Krishna, Molecular Toxicology Sec- tion, Parke-Davis Pharmaceutical Research, Warner-Lambert Company, 2800 Plymouth Road, Ann Arbor, MI 48105, USA. 1983, 1986). During erythropoiesis, the micronu- clei are known to be formed by the lagging cen- tric/acentric chromosome fragments and/or whole chromosomes. Thus, the in vivo micronu- cleus assay can potentially detect agents which cause chromosome breaks and agents which act on spindle. The routine micronucleus assay (Schmid, 1975, 1976; Heddle et al., 1983), how- ever, can not differentiate between the two mech- anisms of micronucleus formation.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Simultaneous evaluation of clastogenicity, aneugenicity and toxicity in the mouse micronucleus assay...

Mutation Research, 282 (1992) 159-167 159 © 1992 Elsevier Science Publishers B.V. All rights reserved 0165-7992/92/$05.00
MUTLET 00674
Simultaneous evaluation of clastogenicity, aneugenicity and toxicity in the mouse micronucleus assay using immunofluorescence
G. Krishna, R. Fiedler and J.C. Theiss Molecular Toxicology Section, Department of Pathology and Experimental Toxicology, Parke-Davis Pharmaceutical Research Division,
Warner-Lambert Company, Ann Arbor, MI 48105, USA
(Received 19 February 1992) (Accepted 2 March 1992)
Keywords: Antikinetochore antibody technique; Clastogenicity evaluation; Aneugenicity evaluation; Toxicity evaluation; Micronu- cleus assay; Mouse; Aneuploidy; Cyclophosphamide; Vincristine
Summary
An improved antikinetochore antibody technique was established in the mouse micronucleus assay to simultaneously evaluate toxicity, clastogenicity and aneugenicity induced by various test agents. The procedure involved the use of cellulose column fractionated cytospun slides for analysis. The staining method consisted of sequential treatment of slides with crest serum, fluorosceinated goat-antihuman and swine-antigoat antibodies, and propidium iodide. In this method, polychromatic erythrocytes (PCEs, dark red), normochromatic erythrocytes (NCEs, green), chromosome(s)/fragments/micronuclei (orange), and kinetochores (yellow), are identified using the same filter setting under blue excitation (440-490 nm) with a barrier filter at 520 nm. Using this method, three agents, cyclophosphamide, X-rays and vincristine were tested for micronucleus/aneuploidy induction and bone marrow toxicity. The aneugen, vincristine, and clastogens, X-rays and cyclophosphamide, induced predominantly kinetochore positive (K + ) and negative ( K - ) micronucleated PCEs, respectively. At the doses tested, cyclophosphamide caused a slight but statistically significant decrease in PCEs in females, and other agents did not produce any severe bone-marrow toxicity in either male or female mice. These results are comparable with the results reported in the literature on these compounds with various methods and thus demonstrate the usefulness of this assay in distinguishing clastogenicity from aneugenicity and in evaluating toxicity.
Short-term tests have been widely used in the genotoxicity evaluation of test compounds. The in vivo micronucleus assay has played a prominent role in mutagenicity testing (Heddle et al., 1983; Arlett et al., 1984; Mavournin et al., 1990; Ashby,
Correspondence Dr. C. Krishna, Molecular Toxicology Sec- tion, Parke-Davis Pharmaceutical Research, Warner-Lambert Company, 2800 Plymouth Road, Ann Arbor, MI 48105, USA.
1983, 1986). During erythropoiesis, the micronu- clei are known to be formed by the lagging cen- tric/acentric chromosome fragments and/or whole chromosomes. Thus, the in vivo micronu- cleus assay can potentially detect agents which cause chromosome breaks and agents which act on spindle. The routine micronucleus assay (Schmid, 1975, 1976; Heddle et al., 1983), how- ever, can not differentiate between the two mech- anisms of micronucleus formation.

160
In recent years, several investigators at various laboratories have successfully used an im- munofluorescence method under in vitro condi- tions (Degrassi and Tanzarella, 1988; Hennig et al., 1988; Thomson and Perry, 1988; Eastmond and Tucker, 1989a,b; Fenech and Morley, 1989; Channarayappa et al., 1991; Antoccia et al., 1991; Rudd et al., 1991; Krishna et al., 1992). This method involves the use of an antikinetochore antibody (obtained from the serum of patients with a particular form of scleroderma pigmento- sum, CREST syndrome) to determine whether the micronuclei contain kinetochores, inferring the presence of entire chromosome(s) or centric fragment(s).
In vivo assessment and identification of aneu- ploidy is one of the important phases of genotoxi- city evaluation (Cimino et al., 1985; Liang and Satyaprakash, 1985; Dellarco et al., 1986; Onfelt, 1986; Miller and Adler, 1989; Vanderkerken et al., 1989; Wang Xu and Adler, 1990; Adler et al., 1991; Parry and Parry, 1991). Aneuploidy-induc- ing agents (aneugens) generally produce micronu- clei containing whole chromosome(s) or centric fragment(s) and clastogens produce micronuclei containing predominantly acentric fragments (Parry and Parry, 1987; Rizzoni et al., 1989). The use of antikinetochore antibody allows these two types of genotoxic agents to be distinguished.
Recently, a few investigators have reported the use of antikinetochore antibody under in vivo conditions to differentiate aneugens from clasto- gens (Gudi et al., 1990; Miller and Adler, 1990; Miller et al., 1991). In these studies, the mouse micronucleus assay was utilized and micronuclei were scored in total erythrocytes. However, in the routine acute micronucleus assay, the micronuclei are evaluated in polychromatic erythrocytes (PCEs). The prior studies could not differentiate between PCEs and normochromatic erythrocytes (NCEs). Gudi et al. (1990) reported the use of whole bone marrow for standard smear prepara- tion. The presence of nucleated cells in the whole bone marrow interfere with accurate evaluation of micronuclei because of background fluores- cence.
In the present study, we have made an attempt to improve the antikinetochore-antibody ap- proach described earlier for the in vivo mouse
micronucleus assay. To eliminate nucleated cells and to minimize background fluorescence, a cel- lulose column cell fractionation (Romagna, 1988; Romagna and Staniforth, 1989; Krishna et al., 1990a, 1991) method was utilized. Further, the sequential treatment of slides with crest serum (antikinetochore antibody), fluorescein-conju- gated goat antihuman and swine-antigoat anti- bodies, and propidium iodide was used. This method helps differentiate PCEs from NCEs and facilitates analysis of micronuclei for the presence or absence of kinetochore(s) in PCEs under one filter setting. This procedure is easy and can be adapted for routine testing. To validate this tech- nique, vincristine sulfate (an aneugen), X-rays (a clastogen), and cyclophosphamide (a clastogen) were used. The study was performed in both male and female mice in order to assess differences between sexes. The results of these studies are reported in this paper.
Materials and methods
Chemicals / radiation Cyclophosphamide (CAS No. 50-18-0) was ob-
tained from Sigma Chemical Company, St. Louis, MO). Vincristine sulfate was purchased from Aldrich (Milwaukee, WI). Polyoxyethylenesorbi- tan monolaurate (Tween 20) and propidium io- dide were obtained from Sigma. An X-ray source (320 KV, 17 cm target, half value thickness of 2.1 mm copper) was used for whole animal X-ray treatment at a dose rate of 200 rad/min. The dose selection for various test agents was based on the literature (Gudi et al., 1990; Mavournin et al., 1990; Krishna et al., 1990b) and our prelimi- nary studies. A kinetochore-positive control serum for the antinuclear antibody test was purchased from Antibodies, Inc. (Davis, CA). This primary antibody is highly specific for a centromere-asso- ciated epitope which is assumed to be the kineto- chore. The fluoresceinated goat antihuman and swine antigoat y-globulins were purchased from Boehringer Mannheim Biochemicals (Indianapo- lis, IN).
Animals. 6-8-week-old CD-1 male and fe- male mice obtained from Charles River Breeding Laboratories (Portage, MI), were housed individ-

ually in stainless steel wire mesh cages and accli- matized for 1 week before dosing. The animals were observed before initiation of the study to ensure that they were healthy. Only animals found to be in a clinically acceptable condition were assigned to the study. During the study, food (Purina Certified Rodent Chow ® 5002) was of- fered ad libitum in stainless steel food containers. Water was supplied ad libitum by an automatic watering system. An automatic timer provided the animal room with an altering 12-h cycle of light and darkness. Each experimental group con- sisted of 3-4 animals per sex.
Chemical treatment. Single doses of vin- cristine (0.1 and 0.2 mg/kg) or cyclophosphamide (40 mg/kg) were injected intraperitoneally at a dose volume of 10 ml/kg. Saline (0.9% NaC1) at a dose volume of 10 mg/kg served as control.
Animal sacrifice, bone-marrow cell isolation and slide preparation. Bone-marrow cells for mi- cronucleus assessment were isolated and slides were prepared as described by Schmid (1975, 1976), Ashby and Mohammed (1986), Romagna and Staniforth (1989), and Krishna et al. (1990a). Briefly, mice were sacrificed by CO 2 asphyxiation 24 h postdose and the femurs excised immedi- ately. Skin and muscle tissue were trimmed from both femurs and the femurs were severed with bone snips. The bone marrow was flushed gently from the channel into a tube with fetal bovine serum (approx. 3 ml/mouse). Cell clumps were broken up by pipetting, and debris was removed by gravity sedimentation for approximately 5 min. The cell suspension was placed on a cellulose column and washed using Hanks' balanced salt solution (HBSS, without phenol red). The eluant was spun at 800 x g for 10 min and the super- natant discarded. The pellet was resuspended in an appropriate amount of HBSS and slides were prepared using a cytocentrifugation method. All slides were fixed in methanol and stained using an immunofluorescent method described below.
Antibody labeling procedure Slides were soaked in PBS-0.01% Tween 20
for 4 min, after which the excess fluid was drained (care was taken to keep slides moist throughout
161
the procedure). The antikinetochore antibody so- lution was diluted with an equal volume of PBS, and 50 ~1 of this solution was placed on each slide. The slides were coverslipped and placed in an incubator (37°C, humidified) for 1 h. Follow- ing incubation, the coverslips were removed and slides were rinsed in PBS for 3 min with frequent agitation. After a second rinse with fresh PBS, slides were rinsed with PBS-0.1% Tween 20 for 2 min and the excess fluid was drained. A 50-/~1 aliquot of fluoresceinated goat-antihuman anti- body previously diluted 1 : 20 with PBS was placed on each slide. The slides were coverslipped, incu- bated for 50 min, rinsed, and excess fluid was drained as before. These slides were labeled again with 50/zl of a second antibody, fluoresceinated swine-antigoat antibody, previously diluted 1:20 in PBS. Slides were coverslipped and incubated for 50 min. Following incubation, the slides were rinsed with two changes of PBS and excess fluid was drained. Slides were placed in 1 /xg/ml pro- pidium iodide solution (in PBS) for 40 sec and rinsed once in PBS for 1-2 sec. Finally, twO drops of antifade solution were placed onto each slide and the slides were coverslipped. The slides were blind coded and either scored immediately or stored in the refrigerator (in dark) up to a week until scoring was completed.
Scoring procedure For scoring, an American Optical microscope
equipped with epifluorescence was used. The slides were scored under blue excitation (440-490 nm) with a barrier filter combination at 520 nm. In this method, PCEs stain dark red, NCEs stain green, chromosome(s)/fragment(s)/micronuclei stain orange, and kinetochore stain yellow. For micronucleus analysis in PCEs, 2000 PCEs per animal were analyzed. When a PCE with a mi- cronucleus was noted, the micronucleus was clas- sified to be either kinetochore negative ( K - , absence of yellow spot in the micronucleus), or positive (K + , presence of one or more yellow spots in the micronucleus). When more than one micronucleus per PCE was noted, the presence of at least one K + micronucleus was used as a criterion to classify a cell as a micronucleated PCE (MNPCE) with K + micronucleus. The mor- phological criteria regarding micronucleus scor-

162
Fig. 1. Photomicrographs of cellulose column fractionated cytospun mouse bone-marrow cells stained with antikinetochore antibody, fluoresceinated antibodies, and propidium iodide; (a) A microscopic field (1200 ×) showing polychromatic erythrocyte (PCE) and normochromatic erythrocyte (NCE) differentiation and a PCE with a kinetochore-negative micronucleus (arrow), (b) The same microscopic field as in (a) but the cells were restained with Wright's Giemsa, showing PCE and NCE differentiation
(1000 x ), (c) A microscopic field showing two PCEs with kinetochore-positive micronuclei(arrows).
ing in a P C E were similar to earlier studies (Schmid, 1975, 1976; Hedd le et al., 1983; Ashby and M o h a m m e d , 1986; M a c G r e g o r et al., 1987; Mavournin et al., 1990). To evaluate potent ial drug toxicity to the bone marrow, propor t ions of PCEs were coun ted based on 400 total erythro- cytes (TEs) per animal.
Statistical analysis of data The square roots of arcsine t ransformed data
were used to improve the homogenei ty of vari- ance and to bet ter comply with the normali ty assumption. The analysis of variance procedure was used to compare each t rea tment group with vehicle control group. Separa te statistical analy-

163
$ [ llmK+ ~K. ~7
M 3 :: ~
NC P ~7 : E2
i
1 : :
, /
o d-~d 0 CP X-RAY
5 - /
4
%
M 3- N P C E 2 - 8
m r + O z - I
VC 0.1 VC 0.2 0 CP X-ItAY VC'0 .1 VC'0 .2
TREATMENT
/
iiiiiiiiiiiiii~!iii~iliiii iiii!iiiiiiiili!iii!iii!i! !iiiii!ili!i!iiiiiiiii!iii
...... iiiiiii!iiiiii!!iiii!ill i i ii :iiii i iil iiiiiiiiiiiii!iiiiiiiil
Iiii!iii iii !ii!iiii! iiil ii iiii ii iiii! ii ii ~ii~ !: ~iii!~ii~i~iiiii!i~i!i! i
iii!iiiiiiiililili i iiill
TREATMENT
Fig. 2. (a) Kinetochore-positive (K + ) and kinetochore-negative ( K - ) micronucleated polychromatic erythrocyte response induced by various test agents in male mice. (b) Kinetochore-positive (K + ) and kinetochore-negative ( K - ) micronucleated polychromatic erythrocyte response induced by various test agents in female mice, 0 = vehicle control (saline); CP = cyclophosphamide (40
mg/kg); X-rays (200 rad/animal); VC = vincristine (0.1 or 0.2 mg/kg).
ses were conducted for each sex, taken separately and combined. The primary measures were 'the frequency of PCEs in the erythrocyte population, the frequency of MNPCEs in the PCE popula- tion, and the proportion of K + and K - MN- PCEs.
Results Differentiation of PCEs and NCEs and ap-
pearance of kinetochore(s) within the micronuclei of mouse bone-marrow erythrocytes using the antikinetochore antibody technique described in the "Materials and Methods" section is shown in Fig. 1. Fig. la shows PCE and NCE differentia- tion under a microscopic field (1200 × ) and a MNPCE. The micronucleus in the PCE is K - . Fig. lb shows the same microscopic field as in Fig. la, but the slide was rinsed and restained with Wright's Giemsa (1000 × ). Fig. lc shows a microscopic field with two MNPCEs which are K + and one of these MNPCEs contains two kinetochore spots in a micronucleus. These pho- tos, taken under one filter-setting, demonstrate the usefulness of the technique for routine test- ing.
Using this technique in mice, saline controls and three test agents were evaluated for the percent PCEs in the bone-marrow erythrocyte population, the frequency of MNPCEs, and the presence of kinetochore spots in the MNPCEs (Table 1). The saline controls, contained 1.63 and 1.25 MNPCEs per 1000 PCEs in males and fe- males, respectively. Among these cells (combined sex), 78% were K - and 22% were K + .
The clastogens, cyclophosphamide and X-rays at the doses tested, clearly increased MNPCE frequencies in both sexes (Fig. 2a, b). For cy- clophosphamide essentially all of the MNPCEs were K - (100% in males, 99.5% in females). For X-rays, approximately 93% were K - in both sexes. For cyclophosphamide, a slight but statisti- cally significant decrease in % PCEs was ob- served in females. However, the % PCEs in cy- clophosphamide-treated males and X-ray-treated males and females did not differ significantly from control values (Fig. 3).
The aneugen vincristine also clearly induced dose-related increases in MNPCE frequencies in both sexes (Fig. 2a, b). In males, approximately 87% of MNPCEs were K + for both doses (0.1 or 0.2 mg/kg) tested. In females, 82% at 0.1 mg/kg

TA
BL
E 1
IND
UC
TIO
N O
F
MIC
RO
NU
CL
EI
IN
MO
US
E
BO
NE
-MA
RR
OW
P
OL
YC
HR
OM
AT
IC
ER
YT
HR
OC
YT
ES
F
OL
LO
WIN
G
TR
EA
TM
EN
T
WIT
H
VA
RIO
US
AG
EN
TS
Tre
atm
ent
Do
se
Nu
mb
er o
f M
ales
F
emal
es
(mg
/kg
) an
imal
s %
PC
Es
a M
NP
CE
s b
/10
00
PC
Es
% P
CE
s
per
sex
±
SD
T
ota
l K
+
c K
- d
± S
D
MN
PC
Es/
10
00
PC
Es
To
tal
K +
K
-
Co
ntr
ol
0 4
55
.81
±1
2.3
1
.63
±
1.2
0.3
8±
0.5
1.
25+
_ 1.
0 5
8.4
4±
5.
8 1
.25
±
1.0
0.2
5±
0.3
Cy
clo
ph
osp
ham
ide
40
4 5
5.1
9±
12
.5
27
.13
± 1
1.5
e 0
27
.13
± 1
1.5
e 5
4.0
0±
2.
6 c
24
.25
±
4.7
e 0
.13
±0
.3
X-R
ays
200
4 5
6.5
0±
14.
0 4
6.8
8±
14.
6 e
3.3
8±
2.6
e
43
.50
± 1
4.0
~ 5
7.0
0±
6.
3 3
3.6
3+
3.
1e
2.3
8+
2.9
(rad
)
Vin
cris
tine
0.
1 3
58
.42
±
8.0
21
.00
±
2.5
e 18
.17+
2.1
e 2
.83
±
0.8
53
.75
±
9.1
12
.00
±1
0.5
9
.83
±8
.5 e
V
incr
isti
ne
0.2
3 48
.58_
+
7.1
32
.83
±
7.6
e 2
8.6
7±
9.0
e
4.1
7+
1.
6 5
5.0
8±
13.
7 4
2.0
0±
6.
5 e
31
.50
±2
.3 e
1.0
0±
0.9
24
.13
±4
.7 e
31
.25
±3
.5 e
2.1
7±
2.1
1
0.5
0±
4.4
e
400
cell
s fo
r %
PC
Es
and
2000
PC
Es
for
MN
PC
E w
ere
sco
red
per
ani
mal
. a
Pol
ychr
omat
ic e
ryth
rocy
tes.
b
Mic
ronu
clea
ted
PC
Es.
c
Kin
etoc
hore
-pos
itiv
e M
NP
CE
. d
Kin
etoc
hore
-neg
ativ
e M
NP
CE
. c
p <
0.0
5.
70
60
4ales Females J
50
% 40
P C E 30 S
20
10
0 0 CP X-RAY VC 0.1 V C 0 . 2
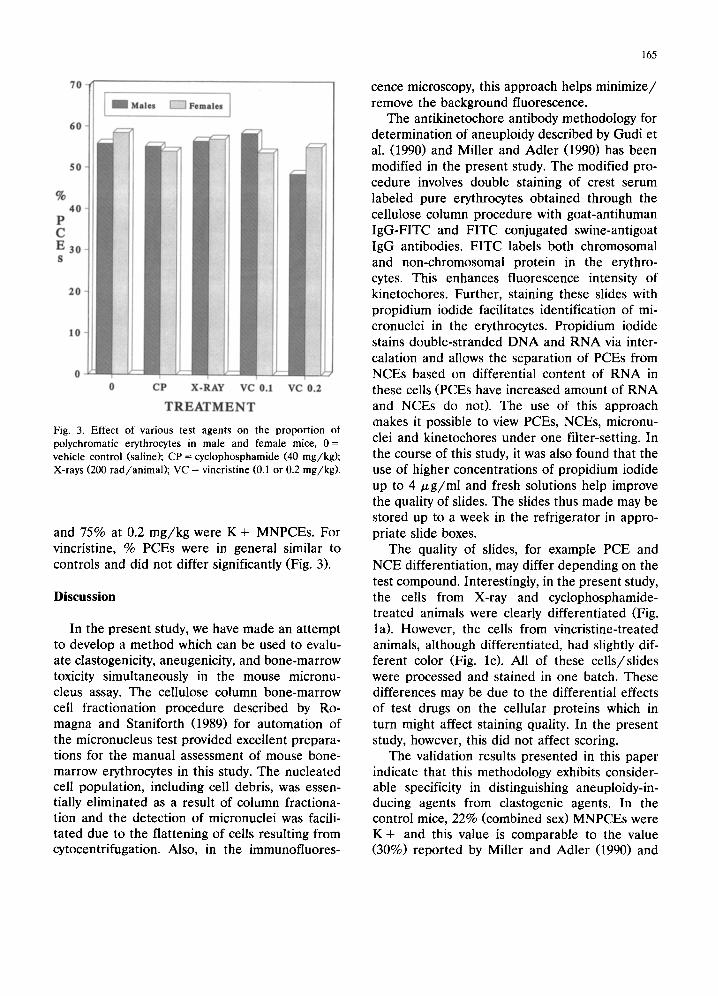
T R E A T M E N T Fig. 3. Effect of various test agents on the proportion ot polychromatic erythrocytes in male and female mice, 0 = vehicle control (saline); CP = cyclophosphamide (40 mg/kg) ; X-rays (200 rad /an imal ) ; VC = vincristine (0.1 or 0.2 mg/kg) .
and 75% at 0.2 mg/kg were K + MNPCEs. For vincristine, % PCEs were in general similar to controls and did not differ significantly (Fig. 3),
Discussion
In the present study, we have made an attempt to develop a method which can be used to evalu- ate clastogenicity, aneugenicity, and bone-marrow toxicity simultaneously in the mouse micronu- cleus assay. The cellulose column bone-marrow cell fractionation procedure described by Ro- magna and Staniforth (1989) for automation of the micronucleus test provided excellent prepara- tions for the manual assessment of mouse bone- marrow erythrocytes in this study. The nucleated cell population, including cell debris, was essen- tially eliminated as a result of column fractiona- tion and the detection of micronuclei was facili- tated due to the flattening of cells resulting from cytocentrifugation. Also, in the immunofluores-
165
cenee microscopy, this approach helps minimize/ remove the background fluorescence.
The antikinetochore antibody methodology for determination of aneuploidy described by Gudi et al. (1990) and Miller and Adler (1990) has been modified in the present study. The modified pro- cedure involves double staining of crest serum labeled pure erythrocytes obtained through the cellulose column procedure with goat-antihuman IgG-FITC and FITC conjugated swine-antigoat IgG antibodies. FITC labels both chromosomal and non-chromosomal protein in the erythro- cytes. This enhances fluorescence intensity of kinetochores. Further, staining these slides with propidium iodide facilitates identification of mi- cronuclei in the erythrocytes. Propidium iodide stains double-stranded DNA and RNA via inter- calation and allows the separation of PCEs from NCEs based on differential content of RNA in these cells (PCEs have increased amount of RNA and NCEs do not). The use of this approach makes it possible to view PCEs, NCEs, micronu- clei and kinetochores under one filter-setting. In the course of this study, it was also found that the use of higher concentrations of propidium iodide up to 4 /~g/ml and fresh solutions help improve the quality of slides. The slides thus made may be stored up to a week in the refrigerator in appro- priate slide boxes.
The quality of slides, for example PCE and NCE differentiation, may differ depending on the test compound. Interestingly, in the present study, the cells from X-ray and cyclophosphamide- treated animals were clearly differentiated (Fig. la). However, the cells from vincristine-treated animals, although differentiated, had slightly dif- ferent color (Fig. lc). All of these cells/slides were processed and stained in one batch. These differences may be due to the differential effects of test drugs on the cellular proteins which in turn might affect staining quality. In the present study, however, this did not affect scoring.
The validation results presented in this paper indicate that this methodology exhibits consider- able specificity in distinguishing aneuploidy-in- ducing agents from clastogenic agents. In the control mice, 22% (combined sex) MNPCEs were K + and this value is comparable to the value (30%) reported by Miller and Adler (1990) and

166
Miller et al. (1991) in total erythrocytes from male mice. However, Gudi et al. (1990) did not find any K + micronuclei among total erythro- cytes in control male mice.
Based on the results in this study, cyclophos- phamide appears to be a pure clastogen. This compound is an alkylating agent known to di- rectly interact with DNA as an electrophile. Tin- well and Ashby (1991) recently described a mor- phological criterion to distinguish aneugens from clastogens, wherein cyclophosphamide produced primarily normal micronuclei and thus, inferring pure clastogenic nature of this antineoplastic drug. The present study clearly demonstrates this activity. X-Rays in the present study predomi- nantly produced MNPCEs devoid of kinetochores (93%). This observation is similar to an earlier study which reported an absence of kinetochores in 97% of total micronucleated erythrocytes ana- lyzed at 200 rad (based on whole body irradia- tion) tested (Gudi et al., 1990).
Vincristine, an aneugen, produced predomi- nantly MNPCEs containing K + spots, in a dose- dependent fashion. In both sexes, on the average, over 80% of MNPCEs were K + . A similar mag- nitude of aneugenicity was also noted for this compound in an earlier study which utilized im- munofluorescence to distinguish such activity (Gudi et al., 1990). The aneugenic activity for this compound was also consistent and evident in a morphological study (Tinwell and Ashby, 1991) which reported predominantly aberrant, large and crescent type micronuclei.
Differentiation of PCEs and NCEs in this study made it possible to also evaluate toxicity of test drugs to the bone marrow. Assessment of bone- marrow toxicity is an essential endpoint in the micronucleus assay. X-Rays at 200 rad tested, did not produce any obvious toxicity. However, cy- clophosphamide produced a slight but statistically significant decrease in % PCEs in females but not in males. Cyclophosphamide, at 40 mg/kg, has been routinely used as a positive control compound in the micronucleus assay in our labo- ratory with no or minimal toxicity (Krishna et al., 1990b). Vincristine, in general, did not produce any severe toxicity at the doses (0.1 or 0.2 mg/kg) tested. In an earlier study, Tinwell and Ashby (1991) also did not notice any toxicity up to 0.5
mg/kg following i.p. treatment of male mice with vincristine, which supports the results of the pre- sent study.
In summary, the data reported here demon- strate the usefulness of the modified antikineto- chore antibody staining technique in the mouse micronucleus assay for routine testing to simulta- neously assess toxic, clastogenic and aneugenic effects. This technique may be used with other tissues or species.
References
Adler I.-D., U. Kliesch, P. van Hummelen and M. Kirsch- Voiders (1991) Mouse micronucleus tests with known and suspect spindle poisons: results from two laboratories, Mutagenesis, 6, 47-53.
Antoccia, A., F. Degrassi, A. Battistoni, P. Ciliutti and C. Tanzarella (1991) In vitro micronucleus test with kineto- chore staining: evaluation of test performance, Mutagene- sis, 6, 319-324.
Arlett, C.F., J. Ashby, R.J. Fiedler and D. Scott (1989) Meet- ing report: Micronuclei: origins, applications and method- ologies - - a workshop sponsored by the Health and Safety Executive held in Manchester, May 23-25, 1988, Mutagen- esis, 4, 482-485.
Ashby, J. (1983) The unique role of rodents in the detection of possible human carcinogens and mutagens, Mutation Res., 115, 117-213.
Ashby, J. (1986) The prospects for a simplified and interna- tionally harmonized approach to the detection of possible human carcinogens and mutagens, Mutagenesis, 1, 3-16.
Ashby, J., and R. Mohammed (1986) Slide preparation and sampling as a major source of variability in the mouse micronucleus assay, Mutation Res., 123, 61-118.
Channarayappa, Nath J., and T. Ong (1991) Study of clasto- genic and aneuploidogenic effects of cigarette smoke con- densate, mitomycin C, and vincristine sulfate, Environ. Mol. Mutagen., 17 (Suppl. 19), 15.
Cimino, M.L, R.R. Tice and J.C. Liang (1986) Aneuploidy in mammalian somatic cells in vivo, Mutation Res., 167, 107-122.
Degrassi, F., and C. Tanzarella (1988) Immunofluorescent staining of kinetochore in micronuclei: a new assay for the detection of aneuploidy, Mutation Res., 203, 339-345.
Dellarco, V.L., K.H. Mavournin and M.D. Waters (1986) Aneuploidy data review committee: summary compilation of chemical data base and evaluation of test methodology, Mutation Res., 167, 149-169.
Eastmond, D.A., and J.D. Tucker (1989a) Identification of aneuploidy-inducing agents using cytokinesis-blocked hu- man lymphocytes and an antikinetochore antibody, Envi- ron. Mol. Mutagenesis, 13, 34-43.
Eastmond, D.A., and J.D. Tucker (1989b) Kinetochore local- ization in micronucleated cytokinesis-blocked Chinese

hamster ovary cells: a new and rapid assay for identifying aneuploidy-inducing agents, Mutation Res., 224, 517-525.
Fenech, M., and A.A. Morley (1989) Kinetochore detection in micronuclei: an alternative method for measuring chromo- some loss, Mutagenesis, 4, 98-104.
Gudi, R., S.S. Sandhu and R.S. Athwal (1990) Kinetochore identification in micronuclei in mouse bone-marrow ery- throcytes: an assay for the detection of aneuploidy-induc- ing agents, Mutation Res., 234, 263-268.
Heddle, J.A., M. Hite, B. Kirkhart, K. Mavournin, J.T. Mac- Gregor, G.W. Newell and M.F. Salamone (1983) The induction of micronuclei as a measure of genotoxicity, Mutation Res., 123, 61-118.
Hennig, U.G.G., N.L. Rudd and D.I. Hoar (1988) Kineto- chore immunofluorescence in micronuclei: a rapid method for the in situ detection of aneuploidy and chromosome breakage in human fibroblasts, Mutation Res., 203, 405- 414.
Krishna, G., D. Brott, G. Urda, L. Vanderwiele, J. Zandee, M. Mckeel, V. Ciaravino, M. Kropko and J. Theiss (1990a) Comparison of micronucleus quantitation in mixed cell populations with enucleated cell preparations quantified manually and using flow cytometry, Environ. Mol. Muta- gen., 15 (Suppl. 17), 32.
Krishna, G., M. Kropko and J. Theiss (1990b) Dimethylni- trosamine-induced micronucleus formation in mouse bone marrow and spleen, Mutation Res., 242, 345-351.
Krishna, G., M. Kropko, V. Ciaravino and J. Theiss (1991) Simultaneous micronucleus and chromosome aberration assessment in the rat, Mutation Res., 264, 29-35.
Krishna, G., R. Fiedler and J. Theiss (1992) In vitro use of antikinetochore antibody (AKA) for simultaneous analysis of chromosome damage and aneuploidy in V79 Chinese hamster lung cells, Environ. Mol. Mutagen., Abstract.
Liang, J.C., and K.L. Satyaprakash (1985) Induction of aneu- ploidy by mitotic arrestants in mouse bone marrow, Muta- tion Res., 155, 61-70.
MacGregor, J.T., J.A. Heddle, M. Hire, B.H. Margolin, C. Ramel, M.F. Salamone, R.R. Tice and D. Wild (1987) Guidelines for the conduct of micronucleus assays in mammalian bone-marrow erythrocytes, Mutation Res., 189, 103-112.
Mavournin, K.H., D.H. Blakey, M.C. Cimino, M.F. Salamone and J.A. Heddle (1990) The in vivo micronucleus assay in mammalian bone marrow and peripheral blood. A report of the US Environmental Protection Agency Gene-Tox Program, Mutation Res., 239, 29-80.
Miller, B.M. and I.-D. Adler (1989) Suspect spindle poisons: analysis of C-mitotic effects in mouse bone-marrow cells, Mutagenesis, 4, 208-215.
Miller, B.M. and I.-D. Adler (1990) Application of antikineto-
167
chore antibody staining (CREST staining) to micronuclei in erythrocytes induced in vivo, Mutagenesis, 5, 411-415.
Miller, B.M., H.F. Zitzelsberger, H.UI.G. Weier and I.-D. Adler (1991) Classification of micronuclei in murine ery- throcytes: immunofluorescent staining using CREST anti- bodies compared to in situ hybridization with biotinylated gamma satellite DNA, Mutagenesis, 6, 297-302.
Onfelt, A. (1986) Mechanistic aspects on chemical induction of spindle disturbances and abnormal chromosome num- bers, Mutation Res., 168, 249-300.
Parry, J.M., and E.M. Parry (1987) Comparison of tests for aneuploidy, Mutation Res., 181, 267-287.
Parry, J.M., and E.M. Parry (1991) The EEC aneuploidy project: an introduction, Mutagenesis, 6, 48, Abstract.
Rizzoni, M., C. Tanzarella, B. Gustavino, F. Degrassi, A. Guarino and E. Vitagliano (1989) Indirect mitotic nondis- junction in Ficia faba and Chinese hamster cells, Chromo- soma, 97, 339-346.
Romagna, F. (1988) Current issues in mutagenesis and car- cinogenesis; fractionation of a pure PE and NE population from rodent bone marrow, Mutation Res., 206, 307-309.
Romagna, F. and C.D. Staniforth (1989) The automated bone-marrow micronucleus test, Mutation Res., 213, 91- 104.
Rudd, N.L., S.E. Williams, M. Evans, U.G.G. Hennig and D.I. Hoar (1991) Kinetochore analysis of micronuclei al- lows insights into the actions of colcemid and mitomycin C, Mutation Res., 261, 57-68.
Schmid, W. (1975) The micronucleus test, Mutation Res., 31, 9-15.
Schmid, W. (1976) The micronucleus test for cytogenetic analysis, in: A. Hollaender (Ed), Chemical Mutagens, Principles and Methods for their Detection, Vol. 4, Aca- demic Press, New York, pp. 31-53.
Thomson, E.J., and P.E. Perry (1988) The identification of micronucleated chromosomes: a possible assay for aneu- ploidy, Mutagenesis, 3, 415-418.
Tinwell, H., and J. Ashby (1991) Micronucleus morphology as a means to distinguish aneugens and clastogens in the mouse bone-marrow micronucleus assay, Mutagenesis, 6, 193-198.
Vanderkerken, K., Ph. Vanparys, L. Verschaeve and M. Kirsch-Volders (1989) The mouse bone-marrow micronu- cleus assay can be used to distinguish aneugens from clastogens, Mutagenesis, 4, 6-11.
Wang, Xu., and I.-D. Adler (1990) Clastogenic effects of known and suspect spindle poisons studied by chromo- some analysis in mouse bone-marrow cells, Mutagenesis, 5, 371-374.
Communicated by J.M. Gentile




















