
Simultaneous detection of the two main proliferation driving EBV encoded proteins, EBNA-2 and LMP-1...
11
Research paper Simultaneous detection of the two main proliferation driving EBV encoded proteins, EBNA-2 and LMP-1 in single B cells Abu E. Rasul a, ⁎, Noémi Nagy a , Ebba Sohlberg b , Mónika Ádori a , Hans-Erik Claesson c , George Klein a , Eva Klein a, ⁎⁎ a Department of Microbiology, Tumor and Cell Biology (MTC), Karolinska Institutet, Nobels väg 16, S-171 77 Stockholm, Sweden b Department of Immunology, Wenner-Gren Institute, Svante Arrheniusväg 20C F5 S-106 91, Stockholm University, Stockholm, Sweden c Department of Medical Biochemistry and Biophysics, Karolinska Institutet, Nobels väg 16, S-171 77 Stockholm, Sweden article info abstract Article history: Received 21 February 2012 Received in revised form 8 August 2012 Accepted 8 August 2012 Available online 18 August 2012 Epstein Barr virus (EBV) is carried by almost all adults, mostly without clinical manifestations. Latent virus infection of B lymphocytes induces activation and proliferation that can be demonstrated in vitro. In healthy individuals, generation of EBV induced malignant proliferation is avoided by continuous immunological surveillance. The proliferation inducing set of the virally encoded genes is expressed exclusively in B cells in a defined differentiation window. It comprises nine EBV encoded nuclear proteins, EBNA 1–6, and three cell membrane associated proteins, LMP-1, 2A and 2B, designated as latency Type III. Outside this window the expression of the viral genes is limited. Healthy carriers harbor a low number of B lymphocytes in which the viral genome is either silent or expresses one virally encoded protein, EBNA-1, latency Type I. In addition, EBV genome carrying B cells can lack either EBNA-2 or LMP-1, latency Type IIa or Type IIb respectively. These cells have no inherent proliferation capacity. Detection of both EBNA-2 and LMP-1 can identify B cells with growth potential. We devised therefore a method for their simultaneous detection in cytospin deposited cell populations. Simultaneous detection of EBNA-2 and LMP-1 was reported earlier in tissues derived from infectious mononucleosis (IM), postransplantation lymphoproliferative disorders (PTLD) and from “humanized” mice infected with EBV. We show for the first time the occurrence of Type IIa and Type IIb cells in cord blood lymphocyte populations infected with EBV in vitro. Further, we confirm the variation of EBNA-2 and LMP-1 expression in several Type III lines and that they vary independently in individual cells. We visualize that in Type III LCL, induced for plasmacytoid differentiation by IL-21 treatment, EBV protein expression changes to Type IIa (EBNA-2 negative LMP-1 positive). We also show that when the proliferation of EBV infected cord blood lymphocyte culture is inhibited by the immunomodulator, PSK, the majority of the cells express latency Type IIa pattern. These results show that by modifying the differentiation state, the proliferating EBV positive B cells can be “curbed”. Type IIa expression is a characteristic for EBV positive Reed–Sternberg (R/S) cells in EBV positive Hodgkin's lymphomas. For survival and proliferation, the R/S cells require the contribution of the in vivo microenvironment. Consequently, Type IIa lines could not be established from Hodgkin's lymphoma in vitro. We propose that these experimental cultures can be exploited for study of the Type IIa cells. © 2012 Elsevier B.V. All rights reserved. Keywords: EBV Immunofluorescence EBNA-2 LMP-1 Type IIa Type IIb 1. Introduction Almost all humans carry the Epstein Barr virus (Thorley- Lawson and Allday, 2008; Young and Rickinson, 2004). In the Western World primary infection occurs most often in Journal of Immunological Methods 385 (2012) 60–70 ⁎ Corresponding author. Tel.: +46 8 524 86761. ⁎⁎ Corresponding author. Tel.: +46 8 524 86774; fax: +46 8 330498. E-mail addresses: [email protected] (A.E. Rasul), [email protected] (E. Klein). 0022-1759/$ – see front matter © 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.jim.2012.08.008 Contents lists available at SciVerse ScienceDirect Journal of Immunological Methods journal homepage: www.elsevier.com/locate/jim
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Simultaneous detection of the two main proliferation driving EBV encoded proteins, EBNA-2 and LMP-1...

Journal of Immunological Methods 385 (2012) 60–70
Contents lists available at SciVerse ScienceDirect
Journal of Immunological Methods
j ourna l homepage: www.e lsev ie r .com/ locate / j im
Research paper
Simultaneous detection of the two main proliferation driving EBV encodedproteins, EBNA-2 and LMP-1 in single B cells
Abu E. Rasul a,⁎, Noémi Nagy a, Ebba Sohlberg b, Mónika Ádori a, Hans-Erik Claesson c,George Klein a, Eva Klein a,⁎⁎a Department of Microbiology, Tumor and Cell Biology (MTC), Karolinska Institutet, Nobels väg 16, S-171 77 Stockholm, Swedenb Department of Immunology, Wenner-Gren Institute, Svante Arrheniusväg 20C F5 S-106 91, Stockholm University, Stockholm, Swedenc Department of Medical Biochemistry and Biophysics, Karolinska Institutet, Nobels väg 16, S-171 77 Stockholm, Sweden
a r t i c l e i n f o
⁎ Corresponding author. Tel.: +46 8 524 86761.⁎⁎ Corresponding author. Tel.: +46 8 524 86774; fa
E-mail addresses: [email protected] (A.E. Rasul),(E. Klein).
0022-1759/$ – see front matter © 2012 Elsevier B.V. Ahttp://dx.doi.org/10.1016/j.jim.2012.08.008
a b s t r a c t
Article history:Received 21 February 2012Received in revised form 8 August 2012Accepted 8 August 2012Available online 18 August 2012
Epstein Barr virus (EBV) is carried by almost all adults, mostly without clinical manifestations.Latent virus infection of B lymphocytes induces activation and proliferation that can bedemonstrated in vitro. In healthy individuals, generation of EBV induced malignant proliferationis avoided by continuous immunological surveillance. The proliferation inducing set of the virallyencoded genes is expressed exclusively in B cells in a defined differentiationwindow. It comprisesnine EBV encoded nuclear proteins, EBNA 1–6, and three cell membrane associated proteins,LMP-1, 2A and 2B, designated as latency Type III. Outside this window the expression of the viralgenes is limited. Healthy carriers harbor a low number of B lymphocytes in which the viralgenome is either silent or expresses one virally encoded protein, EBNA-1, latency Type I. Inaddition, EBV genome carrying B cells can lack either EBNA-2 or LMP-1, latency Type IIa or TypeIIb respectively. These cells have no inherent proliferation capacity. Detection of both EBNA-2 andLMP-1 can identify B cells with growth potential. We devised therefore a method for theirsimultaneous detection in cytospin deposited cell populations. Simultaneous detection of EBNA-2and LMP-1 was reported earlier in tissues derived from infectious mononucleosis (IM),postransplantation lymphoproliferative disorders (PTLD) and from “humanized” mice infectedwith EBV.We show for the first time the occurrence of Type IIa and Type IIb cells in cord blood lymphocytepopulations infected with EBV in vitro. Further, we confirm the variation of EBNA-2 and LMP-1expression in several Type III lines and that they vary independently in individual cells.We visualizethat in Type III LCL, induced for plasmacytoid differentiation by IL-21 treatment, EBV proteinexpression changes to Type IIa (EBNA-2 negative LMP-1 positive). We also show that when theproliferation of EBV infected cord blood lymphocyte culture is inhibited by the immunomodulator,PSK, the majority of the cells express latency Type IIa pattern. These results show that by modifyingthe differentiation state, the proliferating EBVpositive B cells can be “curbed”. Type IIa expression is acharacteristic for EBV positive Reed–Sternberg (R/S) cells in EBV positive Hodgkin's lymphomas. Forsurvival and proliferation, the R/S cells require the contribution of the in vivo microenvironment.Consequently, Type IIa lines could not be established fromHodgkin's lymphoma in vitro.We proposethat these experimental cultures can be exploited for study of the Type IIa cells.
© 2012 Elsevier B.V. All rights reserved.
Keywords:EBVImmunofluorescenceEBNA-2LMP-1Type IIaType IIb
x: +46 8 [email protected]
ll rights reserved.
1. Introduction
Almost all humans carry the Epstein Barr virus (Thorley-Lawson and Allday, 2008; Young and Rickinson, 2004). In theWestern World primary infection occurs most often in

61A.E. Rasul et al. / Journal of Immunological Methods 385 (2012) 60–70
adolescents. Clinical signs of the infection can be absent.Alternatively it can cause uncharacteristic mild symptoms ortypical infectious mononucleosis, IM with varied symptom-atology and severity (Straus et al., 1993). Some of thesymptoms of IM reflect activation of innate immunity thatprotects the host from the immediate danger of the infection(Klein and Claesson, 2010). This danger stems from the efficienttransforming potential of the virus for its main target cell, the Blymphocyte. This can lead to malignancy unless controlled bythe immune system, as evidenced by conditions that areaccompanied by immunosuppression like postransplantationlymphoproliferative disorders (PTLD) (Swerdlow et al., 2008).Whether or not the primary infection is causing symptoms, it isalways followed by the virus carrier state that is easilydetectable by sustained virus specific immunity, antibodiesand cell mediated reactions to viral structural proteins and tovirally encoded cellular proteins (Klein, 1994; Rickinson andMoss, 1997; Yao et al., 1985).
The expression of the viral genes varies with the differen-tiation of the cell which in turn determines the fate of the cell(Klein et al., 2007; Thorley-Lawson, 2001). This is the key of thehealthy lifelong carrier state where memory B cells harbor thepartly or completely silent virus genome (Babcock et al., 2000).
The details of the virus-B cell interactionwere clarified by invitro experiments. Resting B cells can be infected with EBV andeventually become activated and induced for proliferation(Küppers, 2003; Pope et al., 1968). Nine virally encoded genesare expressed in these cells that can establish as immortalizedlymphoblastoid cell lines, LCLs. This set of proteins constitutesthe “growth program” also referred to as latency Type III (Roweet al., 1992). It comprises six virally encoded nuclear, (EBNA-1,2, 3A, 3B, 3C and LP) and three membrane associated proteins,LMP-1, LMP-2a and LMP-2b. EBNA-2 and LMP-1 are pivotal forimposing inherent proliferation capacity. Due to the require-ment of B cell specific transcription factors, the growthprogram is expressed only in B lymphocytes and in a defineddifferentiationwindow. Outside this window the expression ofviral genomemay be silent or express only one virally encodedprotein, EBNA-1, latency Type I. In addition, EBV genome caringB cells can lack either EBNA-2 or LMP-1, latency type IIa or IIbrespectively. These cells have no inherent proliferation capacity(Klein et al., 2007).
Visualization of EBNA-2 and LMP-1 by immuneperoxidaseand by immunofluorescence, IF was reported earlier in tissuesderived from IM and PTLD and also in EBV infected “human-ized”mice (Brink et al., 1997; Kurth et al., 2000, 2003;Ma et al.,2011; Niedobitek et al., 1997; Oudejans et al., 1995). Thesestudies showed considerable independent variation in theexpression of EBNA-2 and LMP-1 in Type III cells and detectedcells with the restricted Type IIa and Type IIb, LMP-1 positiveEBNA-2 negative and EBNA-2 positive LMP-1 negative respec-tively, expression pattern.
The first infection leads to a lifelong carrier state. Therefore,the danger of proliferating Type III cells has to be constantlyeliminated. Their recognition by adaptive immunologicalmechanisms was thought to be mainly responsible for thesurveillance. When Type III cells arise in healthy individualswith EBV specific adaptive immunity, cytotoxic CD8 lympho-cytes recognize virally encoded proteins and the activationmarkers on the transformed cells (Khanna et al., 1999; Pearsonet al., 1970).
Expression of both EBNA-2 and LMP-1 is the marker ofB cells with inherent proliferative capacity. Their solitaryexpression identifies EBV positive B cells with differentbiological behavior. Therefore, a reliable method for theirsimultaneous detection is warranted. We devised the methodfor simultaneous detection of EBNA-2 and LMP-1 in cytospindeposited cell population. The results shown here validate themethod by confirmation of earlier findings in several experi-mental systems. We showed differences in the pattern ofEBNA-2 and LMP-1 expression in various Type III lines. Weconfirmed and visualized loss of EBNA-2 in LCL cells inducedfor plasmacytoid differentiation.
Furthermore, we showed that in vitro EBV infected cellpopulations in which proliferation was inhibited by immuno-modulator treatment were made up mainly of Type IIa cells.We detected for the first time non-proliferating EBV genomecarrying cells exhibiting latency Type IIa and Type IIb in thecultures of cord blood lymphocyte populations early afterinfection in vitro.
2. Materials and methods
2.1. Cell Lines
We used cell lines with Type III EBV expression: LCLsderived from in vitro EBV infected cord blood, M-LCL (newlydeveloped), and CBM1-Ral-STO (Ernberg et al., 1989); in vitroEBV infected isolated CD19+ B cells from adult blood, LCL 07.08(Nagy et al., 2012); CLL cells which are EBV infected in vitro,HG3 and HG6 (Rosén et al., 2012); EBV positive lines that grewfrom explanted CLL cells, Mec-2 (Stacchini et al., 1999);Burkitt's lymphoma (BL) derived line, Mutu III cl.99 (Gregoryet al., 1990). BJABwas used as EBV negative control (Klein et al.,1974). The cells were cultured in RPMI 1640 supplementedwith 10% heat-inactivated FCS, 100 units/ml penicillin, and100 μg/ml streptomycin in humidified incubator at 37 °C and5% CO2.
2.2. Immunofluorescence staining
The cells were deposited on glass slides in a cytospincentrifuge and fixed in acetone–methanol (2:1) for 30 min. ForEBNA-2 and LMP-1 double staining, following rehydration inbalanced salt solution (BSS) for 1 h, the slides were incubatedwith mouse monoclonal antibody (mAb) for EBNA-2, PE2(IgG1, Leica Biosystem NovoCastra Ltd, dilution 1:50) at roomtemperature for 60 min. With subsequent washing in BSS, theslides were exposed to goat anti-mouse IgG1-Alexa Fluor 488(Invitrogen, dilution 1:200) for 30 min. After washing in BSS,monoclonal anti LMP-1 Ab, S-12 (IgG2a, culture supernatant,dilution 1:50) was added and the slides were incubated for60 min. Followed by washing in BSS, goat anti-mouse IgG2-
a-Alexa Fluor 594 (Invitrogen, dilution 1:200) was used todetect LMP‐1 with incubation for 30 min. Finally after washingin BSS, the slides were mounted with Vectashield containingDAPI (Vector laboratories, Inc.).
For Ki-67 and LMP-1 double staining, a similar protocol wasused. The slides were first incubated with rabbit anti Ki-67(Dakopatts AB, 1:50) for 1 h at room temperature and then itwas visualized by Goat anti-rabbit IgG, Alexa Fluor 488

62 A.E. Rasul et al. / Journal of Immunological Methods 385 (2012) 60–70
(Invitrogen, 1:200). Thereafter, the staining for LMP-1 was doneas described above.
Images were generated with a Leitz DM RB microscope(Leica Microsystems) using a 40×/1.25–0.75 N.A. oil immer-sion objective lens. Images were captured with Hamamatsudual-mode cooled charged-coupled-device camera (C48880)and Hipic 6.4.0 software (Hamamatsu Photonics) as 8-bituncompressed TIFF files. Pictures were edited for optimal colorcontrast by using Photoshop CS3 (Adobe systems).
2.3. Immunoblotting
The cells were lysed in sodium dodecyl sulfate (SDS)gel-loading buffer. Aliquots corresponding to 1.5×105 cellswere loaded in each lane. Immunoblotting was performed asdescribed previously (Kis et al., 2010). EBNA-2 was detectedwith the mAB, clone PE2. For detection of LMP-1 the mousemonoclonal antibody S-12 and the monoclonal cocktail CS1-4(Novocastra Laboratories, UK) were used. As a control forprotein loading, mAb specific for β-actin, clone AC-15 (Sigma–Aldrich) was used.
2.4. IL-21 treatment
CBM1-Ral-STO cells were cultured at a concentration of0.16×106 cells/ml with the addition of 100 ng/ml IL-21(PeproTech EC, UK). Every second day, the cultures werereadjusted to 0.16×106 cells/ml with fresh IL-21 at the samedose.
2.5. In vitro EBV infection and culture of cord blood lymphocytes
Mononuclear cells were isolated from heparinized cordblood by Ficoll–Hypaque density centrifugation. They wereincubated with B95-8 virus containing supernatant in ahumidified incubator at 37 °C and 5% CO2 for one and halfhour with shaking at every 30 min. Thereafter the cells werewashed and re-suspended in complete RPMI 1640 at aconcentration of 1×106 cells per ml. PSK was applied at aconcentration of 100 μg/ml (Kureha Chemical Ind. Co., Tokyo).The 5-lypoxygenase inhibitor, BW B70C (a kind gift from Dr.Lawrie G Garland, The Welcome Foundation Ltd, Unitedkingdom) was used at a concentration of 1 μM.
2.6. Flow cytometry
Ex vivo and cultured cells (7 day old) were washed in coldPBS containing 2% FCS and then stained with FITC-, or PE-, orPE-Cy5-conjugatedmouse anti-humanmonoclonal antibodies.Mouse monoclonal CD3, CD4, CD8 and CD19 were purchasedfrom Becton Dickinson (San Jose, Ca). Ten thousand eventswere collected on a FACScan flow cytometer, and the resultswere analyzed using CELLQUEST (Becton Dickinson) software.
3. Results
3.1. EBNA-2 and LMP-1 expression in Type III cell lines
Both EBNA-2 and LMP-1 were clearly detectable by IF in allcells of Type III lines (Figs. 1 and 2). The panel included LCLsestablished by in vitro infection of normal B lymphocytes and of
B-CLL cells, B-CLL cells from a patient in prolymphocytoidtransformation that acquired EBV in vivo and one Burkitt'slymphoma derived cell line (see Material and methods). Thespecificity of the staining was validated with the EBV negativeline, BJABwhich did not show stainingwith the same procedure.
The pattern and intensities of the staining of the twoproteins differed considerably in the cell lines. The populationswere heterogeneous and the EBNA-2 and LMP-1 expressionsvaried independently. The normal B cell derived LCLs wereparticularly heterogeneous. In comparison to EBNA-2, theLMP-1 expression showed more variation in intensity andpattern. EBNA-2 staining was confined to the nucleus. LMP-1staining was mainly connected with the cell membrane andpolarized. In the Mutu III and Mec-2 lines, LMP-1 staining wasmainly intracellular and diffuse (Figs. 1 and 2). Some cells inthe normal B cell derived lines, M-LCL and LCL 07.08, stainedvery intensely for LMP-1 and therefore exposure time had to beadjusted at the expense of theweakly stained cells which couldappear negative on the picture, but in visual scoring negativecells were not found. On the other hand, due to the weakstaining of LMP-1 inMutu III cl.99 cells, recording required longexposure time (Fig. 1). Thus the intensity of staining in thedifferent samples cannot be compared.
Asmentioned above, LMP-1 staining of the BL derivedMutuIII cl.99 cells was extremely weak. This is in accord with theearlier report about LMP-1 expression in the BL derived Type IIIlines, being considerably lower in comparison to normal B cellderived LCLs (Masucci et al., 1987). The HG3 and HG6 lineshave been established at different times by in vitro infection ofCLL cells collected from a patient and frozen preserved inseveral aliquot. The LMP-1 expression patterns in HG3 andHG6differed slightly; it was more polarized in HG6 cells (Fig. 2).
Immunoblotting substantiated the IF results, with oneexception (Fig. 3). For detection of LMP-1, two antibodieswere used; CS1-4 and S-12 (both in immunoblotting but onlyS-12 in IF, see Material and methods). According to Brink etal. (1997), the sensitivity of S-12 is higher. In our experi-ments CS1-4 detected LMP-1 by immunoblotting in all of thelines tested; while S-12 detected LMP-1 in the Mec-2 cells byIF but not in the blot. Because of high sensitivity of S-12antibody, we used considerably shorter time to visualize theblot. Prolonged exposure gave strong backgroundmaking thebands indistinguishable. Very weak or absence of band mightbe due to disturbed conformation of the protein in the lysate.
3.2. Ki-67 and LMP-1 expression in LCL cells and in in vitroinfected CBMC
We applied the staining protocol established for EBNA-2and LMP-1 for detection of Ki-67, the proliferation markertogether with LMP-1 (Gerdes et al., 1983). Our results were inline with an earlier report showing that in the LCL population,cells with high levels of LMP-1 incorporated less BrdU (Lam etal., 2004). In theM-LCL culture, high proportion of the cells thathad strong LMP-1 expression was negative for Ki-67 (Fig. 4).This negative correlation was not absolute however, some ofthe Ki-67 positive cells strongly expressed LMP-1 as well. Theapparently LMP-1 negative cells were weak expressers; asmentioned above in order to compensate the strongly stainedcells, the exposure time had to be reduced at the expense ofweakly expressing cells.
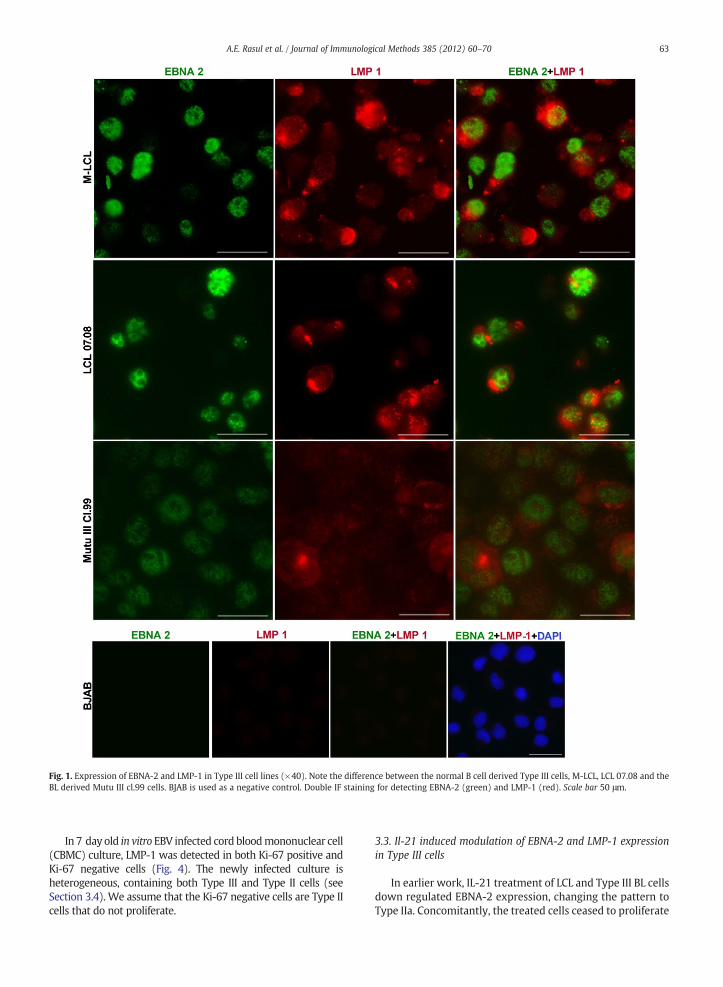
Fig. 1. Expression of EBNA-2 and LMP-1 in Type III cell lines (×40). Note the difference between the normal B cell derived Type III cells, M-LCL, LCL 07.08 and theBL derived Mutu III cl.99 cells. BJAB is used as a negative control. Double IF staining for detecting EBNA-2 (green) and LMP-1 (red). Scale bar 50 μm.
63A.E. Rasul et al. / Journal of Immunological Methods 385 (2012) 60–70
In 7 day old in vitro EBV infected cord bloodmononuclear cell(CBMC) culture, LMP-1 was detected in both Ki-67 positive andKi-67 negative cells (Fig. 4). The newly infected culture isheterogeneous, containing both Type III and Type II cells (seeSection 3.4). We assume that the Ki-67 negative cells are Type IIcells that do not proliferate.
3.3. Il-21 induced modulation of EBNA-2 and LMP-1 expressionin Type III cells
In earlier work, IL-21 treatment of LCL and Type III BL cellsdown regulated EBNA-2 expression, changing the pattern toType IIa. Concomitantly, the treated cells ceased to proliferate
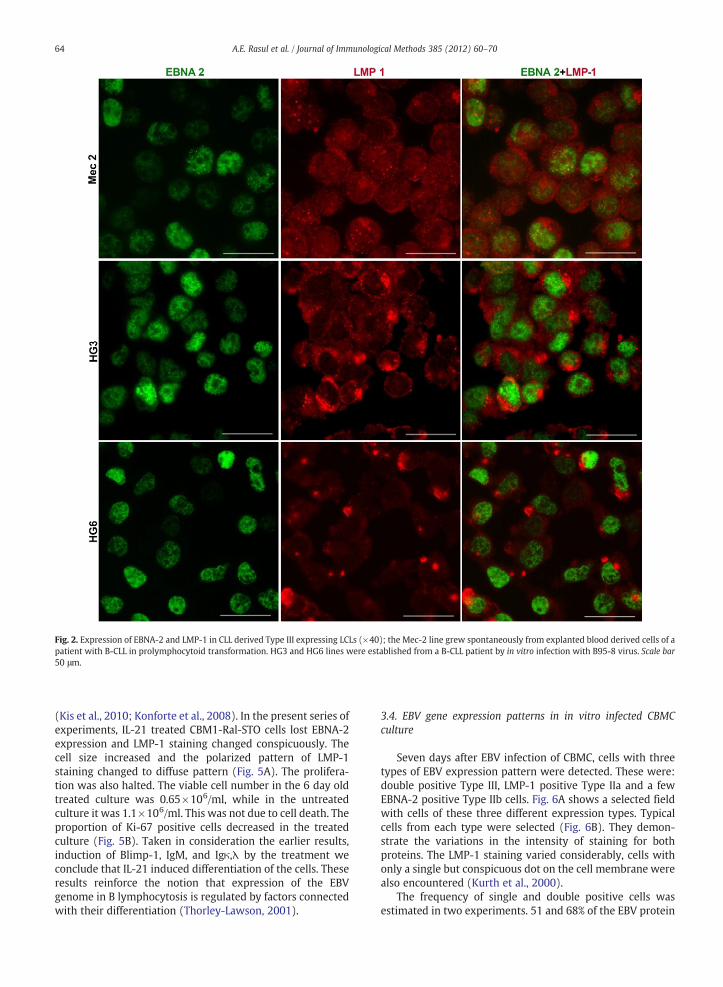
Fig. 2. Expression of EBNA-2 and LMP-1 in CLL derived Type III expressing LCLs (×40); the Mec-2 line grew spontaneously from explanted blood derived cells of apatient with B-CLL in prolymphocytoid transformation. HG3 and HG6 lines were established from a B-CLL patient by in vitro infection with B95-8 virus. Scale bar50 μm.
64 A.E. Rasul et al. / Journal of Immunological Methods 385 (2012) 60–70
(Kis et al., 2010; Konforte et al., 2008). In the present series ofexperiments, IL-21 treated CBM1-Ral-STO cells lost EBNA-2expression and LMP-1 staining changed conspicuously. Thecell size increased and the polarized pattern of LMP-1staining changed to diffuse pattern (Fig. 5A). The prolifera-tion was also halted. The viable cell number in the 6 day oldtreated culture was 0.65×106/ml, while in the untreatedculture it was 1.1×106/ml. This was not due to cell death. Theproportion of Ki-67 positive cells decreased in the treatedculture (Fig. 5B). Taken in consideration the earlier results,induction of Blimp-1, IgM, and Igκ,λ by the treatment weconclude that IL-21 induced differentiation of the cells. Theseresults reinforce the notion that expression of the EBVgenome in B lymphocytosis is regulated by factors connectedwith their differentiation (Thorley-Lawson, 2001).
3.4. EBV gene expression patterns in in vitro infected CBMCculture
Seven days after EBV infection of CBMC, cells with threetypes of EBV expression pattern were detected. These were:double positive Type III, LMP-1 positive Type IIa and a fewEBNA-2 positive Type IIb cells. Fig. 6A shows a selected fieldwith cells of these three different expression types. Typicalcells from each type were selected (Fig. 6B). They demon-strate the variations in the intensity of staining for bothproteins. The LMP-1 staining varied considerably, cells withonly a single but conspicuous dot on the cell membrane werealso encountered (Kurth et al., 2000).
The frequency of single and double positive cells wasestimated in two experiments. 51 and 68% of the EBV protein
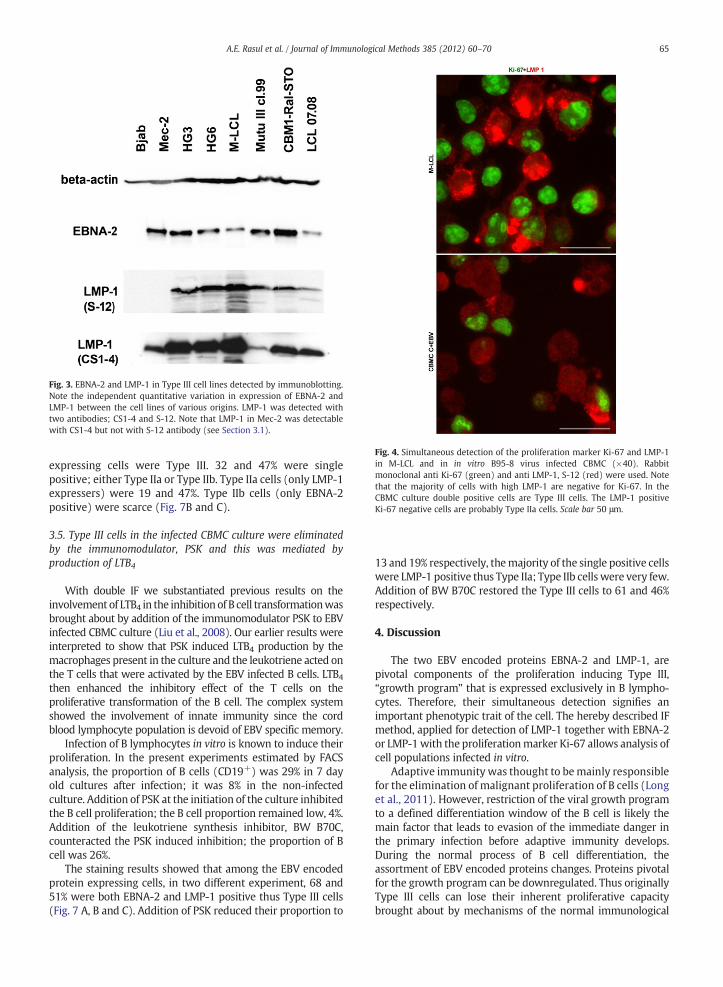
Fig. 3. EBNA-2 and LMP-1 in Type III cell lines detected by immunoblotting.Note the independent quantitative variation in expression of EBNA-2 andLMP-1 between the cell lines of various origins. LMP-1 was detected withtwo antibodies; CS1-4 and S-12. Note that LMP-1 in Mec-2 was detectablewith CS1-4 but not with S-12 antibody (see Section 3.1).
Fig. 4. Simultaneous detection of the proliferation marker Ki-67 and LMP-1in M-LCL and in in vitro B95-8 virus infected CBMC (×40). Rabbitmonoclonal anti Ki-67 (green) and anti LMP-1, S-12 (red) were used. Notethat the majority of cells with high LMP-1 are negative for Ki-67. In theCBMC culture double positive cells are Type III cells. The LMP-1 positiveKi-67 negative cells are probably Type IIa cells. Scale bar 50 μm.
65A.E. Rasul et al. / Journal of Immunological Methods 385 (2012) 60–70
expressing cells were Type III. 32 and 47% were singlepositive; either Type IIa or Type IIb. Type IIa cells (only LMP-1expressers) were 19 and 47%. Type IIb cells (only EBNA-2positive) were scarce (Fig. 7B and C).
3.5. Type III cells in the infected CBMC culture were eliminatedby the immunomodulator, PSK and this was mediated byproduction of LTB4
With double IF we substantiated previous results on theinvolvement of LTB4 in the inhibition of B cell transformationwasbrought about by addition of the immunomodulator PSK to EBVinfected CBMC culture (Liu et al., 2008). Our earlier results wereinterpreted to show that PSK induced LTB4 production by themacrophages present in the culture and the leukotriene acted onthe T cells that were activated by the EBV infected B cells. LTB4then enhanced the inhibitory effect of the T cells on theproliferative transformation of the B cell. The complex systemshowed the involvement of innate immunity since the cordblood lymphocyte population is devoid of EBV specific memory.
Infection of B lymphocytes in vitro is known to induce theirproliferation. In the present experiments estimated by FACSanalysis, the proportion of B cells (CD19+) was 29% in 7 dayold cultures after infection; it was 8% in the non-infectedculture. Addition of PSK at the initiation of the culture inhibitedthe B cell proliferation; the B cell proportion remained low, 4%.Addition of the leukotriene synthesis inhibitor, BW B70C,counteracted the PSK induced inhibition; the proportion of Bcell was 26%.
The staining results showed that among the EBV encodedprotein expressing cells, in two different experiment, 68 and51% were both EBNA-2 and LMP-1 positive thus Type III cells(Fig. 7 A, B and C). Addition of PSK reduced their proportion to
13 and 19% respectively, themajority of the single positive cellswere LMP-1 positive thus Type IIa; Type IIb cells were very few.Addition of BW B70C restored the Type III cells to 61 and 46%respectively.
4. Discussion
The two EBV encoded proteins EBNA-2 and LMP-1, arepivotal components of the proliferation inducing Type III,“growth program” that is expressed exclusively in B lympho-cytes. Therefore, their simultaneous detection signifies animportant phenotypic trait of the cell. The hereby described IFmethod, applied for detection of LMP-1 together with EBNA-2or LMP-1with the proliferationmarker Ki-67 allows analysis ofcell populations infected in vitro.
Adaptive immunity was thought to bemainly responsiblefor the elimination of malignant proliferation of B cells (Longet al., 2011). However, restriction of the viral growth programto a defined differentiation window of the B cell is likely themain factor that leads to evasion of the immediate danger inthe primary infection before adaptive immunity develops.During the normal process of B cell differentiation, theassortment of EBV encoded proteins changes. Proteins pivotalfor the growth program can be downregulated. Thus originallyType III cells can lose their inherent proliferative capacitybrought about by mechanisms of the normal immunological
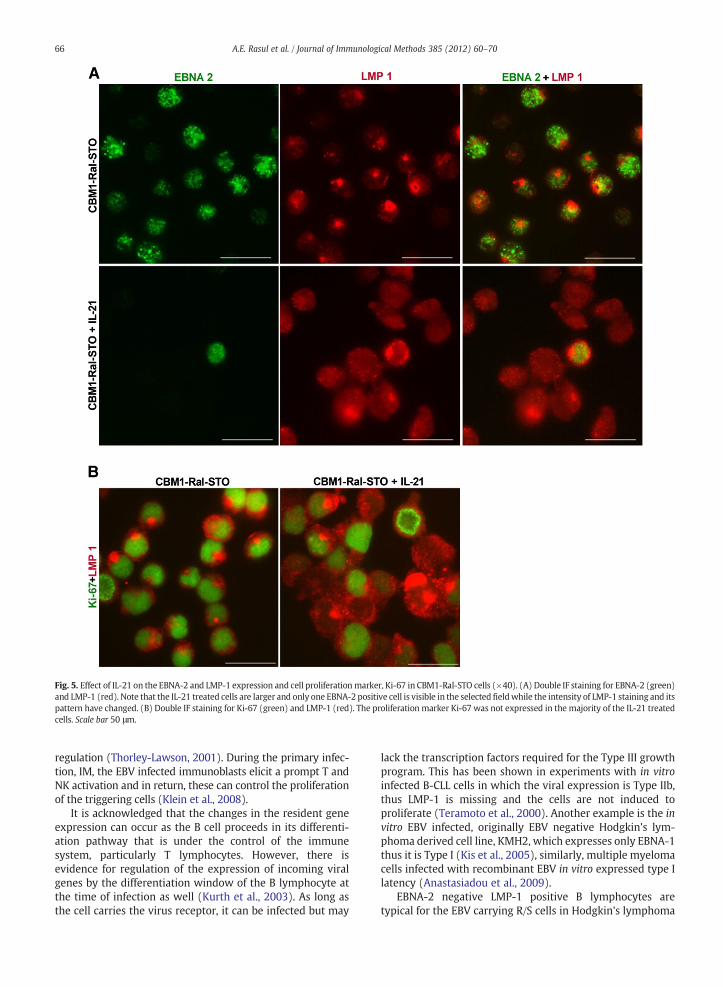
Fig. 5. Effect of IL-21 on the EBNA-2 and LMP-1 expression and cell proliferationmarker, Ki-67 in CBM1-Ral-STO cells (×40). (A) Double IF staining for EBNA-2 (green)and LMP-1 (red). Note that the IL-21 treated cells are larger and only one EBNA-2 positive cell is visible in the selected fieldwhile the intensity of LMP-1 staining and itspattern have changed. (B) Double IF staining for Ki-67 (green) and LMP-1 (red). The proliferation marker Ki-67 was not expressed in the majority of the IL-21 treatedcells. Scale bar 50 μm.
66 A.E. Rasul et al. / Journal of Immunological Methods 385 (2012) 60–70
regulation (Thorley-Lawson, 2001). During the primary infec-tion, IM, the EBV infected immunoblasts elicit a prompt T andNK activation and in return, these can control the proliferationof the triggering cells (Klein et al., 2008).
It is acknowledged that the changes in the resident geneexpression can occur as the B cell proceeds in its differenti-ation pathway that is under the control of the immunesystem, particularly T lymphocytes. However, there isevidence for regulation of the expression of incoming viralgenes by the differentiation window of the B lymphocyte atthe time of infection as well (Kurth et al., 2003). As long asthe cell carries the virus receptor, it can be infected but may
lack the transcription factors required for the Type III growthprogram. This has been shown in experiments with in vitroinfected B-CLL cells in which the viral expression is Type IIb,thus LMP-1 is missing and the cells are not induced toproliferate (Teramoto et al., 2000). Another example is the invitro EBV infected, originally EBV negative Hodgkin's lym-phoma derived cell line, KMH2, which expresses only EBNA-1thus it is Type I (Kis et al., 2005), similarly, multiple myelomacells infected with recombinant EBV in vitro expressed type Ilatency (Anastasiadou et al., 2009).
EBNA-2 negative LMP-1 positive B lymphocytes aretypical for the EBV carrying R/S cells in Hodgkin's lymphoma
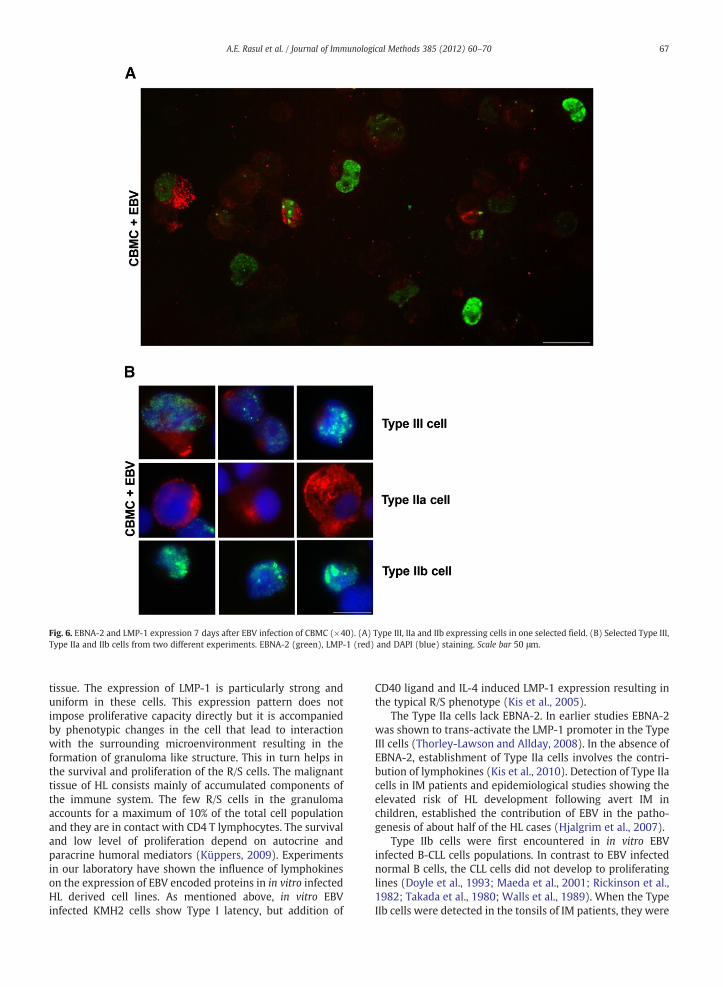
Fig. 6. EBNA-2 and LMP-1 expression 7 days after EBV infection of CBMC (×40). (A) Type III, IIa and IIb expressing cells in one selected field. (B) Selected Type III,Type IIa and IIb cells from two different experiments. EBNA-2 (green), LMP-1 (red) and DAPI (blue) staining. Scale bar 50 μm.
67A.E. Rasul et al. / Journal of Immunological Methods 385 (2012) 60–70
tissue. The expression of LMP-1 is particularly strong anduniform in these cells. This expression pattern does notimpose proliferative capacity directly but it is accompaniedby phenotypic changes in the cell that lead to interactionwith the surrounding microenvironment resulting in theformation of granuloma like structure. This in turn helps inthe survival and proliferation of the R/S cells. The malignanttissue of HL consists mainly of accumulated components ofthe immune system. The few R/S cells in the granulomaaccounts for a maximum of 10% of the total cell populationand they are in contact with CD4 T lymphocytes. The survivaland low level of proliferation depend on autocrine andparacrine humoral mediators (Küppers, 2009). Experimentsin our laboratory have shown the influence of lymphokineson the expression of EBV encoded proteins in in vitro infectedHL derived cell lines. As mentioned above, in vitro EBVinfected KMH2 cells show Type I latency, but addition of
CD40 ligand and IL-4 induced LMP-1 expression resulting inthe typical R/S phenotype (Kis et al., 2005).
The Type IIa cells lack EBNA-2. In earlier studies EBNA-2was shown to trans-activate the LMP-1 promoter in the TypeIII cells (Thorley-Lawson and Allday, 2008). In the absence ofEBNA-2, establishment of Type IIa cells involves the contri-bution of lymphokines (Kis et al., 2010). Detection of Type IIacells in IM patients and epidemiological studies showing theelevated risk of HL development following avert IM inchildren, established the contribution of EBV in the patho-genesis of about half of the HL cases (Hjalgrim et al., 2007).
Type IIb cells were first encountered in in vitro EBVinfected B-CLL cells populations. In contrast to EBV infectednormal B cells, the CLL cells did not develop to proliferatinglines (Doyle et al., 1993; Maeda et al., 2001; Rickinson et al.,1982; Takada et al., 1980; Walls et al., 1989). When the TypeIIb cells were detected in the tonsils of IM patients, they were
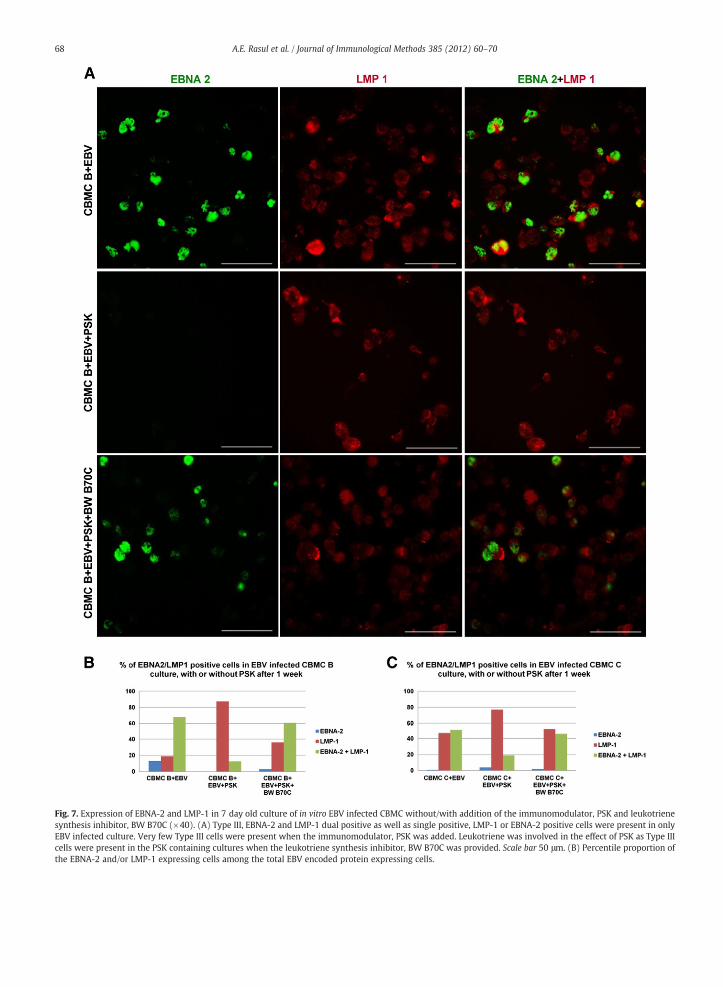
Fig. 7. Expression of EBNA-2 and LMP-1 in 7 day old culture of in vitro EBV infected CBMC without/with addition of the immunomodulator, PSK and leukotrienesynthesis inhibitor, BW B70C (×40). (A) Type III, EBNA-2 and LMP-1 dual positive as well as single positive, LMP-1 or EBNA-2 positive cells were present in onlyEBV infected culture. Very few Type III cells were present when the immunomodulator, PSK was added. Leukotriene was involved in the effect of PSK as Type IIIcells were present in the PSK containing cultures when the leukotriene synthesis inhibitor, BW B70C was provided. Scale bar 50 μm. (B) Percentile proportion ofthe EBNA-2 and/or LMP-1 expressing cells among the total EBV encoded protein expressing cells.
68 A.E. Rasul et al. / Journal of Immunological Methods 385 (2012) 60–70

69A.E. Rasul et al. / Journal of Immunological Methods 385 (2012) 60–70
assumed to identify a transitory stage of the newly infected Bcell (Kurth et al., 2000). The molecular events regulating thisunusual expression pattern of the latent EBV genes are notyet known. Rarely, EBV positive cell lines were establishedfrom CLL cells. But these are typical LCLs and thus Type IIIcells (Rosén et al., 2012; Stacchini et al., 1999). CLL derivedLCLs that grew out spontaneously from the explanted bloodof CLL patient have probably arisen from some CLL cells thatchange their differentiation window under the culturecondition.
By visualization of LMP-1 and the proliferation markerKi-67, we confirmed that majority of the strong LMP-1 ex-presser cells in LCL population do not proliferate (Lam et al.,2004). This was assessed earlier by BrdU incorporation relatedto the level of LMP-1 expression in EBV carrying lymphoblasts(Lam et al., 2004).
We confirmed our earlier finding that showed loss ofEBNA-2 and the intensified LMP-1 expression in Type III cells,when they were induced for plasmacytoid differentiation byIL-21 treatment (Kis et al., 2010). The present experimentsdetected a conspicuous change in the pattern of LMP-1 stainingas well. In the untreated cells the staining was polarized andlocalized mainly on the cell membrane while it was homoge-neous in the enlarged and differentiated cells.
We have shown earlier in EBV infected cord bloodlymphocyte culture that the proliferation of B lymphocyteswas inhibited when an immunomodulator, PSK acting onmacrophages was added and proved that leukotriene produc-tion was involved in this effect (Liu et al., 2008). Weinterpreted the results as activation of T cells for cytotoxicfunction by solublemacrophage products, thus elimination of Bcells with proliferative potential. Our present results showedthat the PSK containing cultures were mainly made up of TypeIIa cells. Thus suggest that the cessation of B cell proliferationcould be attributed, at least in part, to downregulation ofEBNA-2 by products of activated T cells (Nagy et al., 2012). Theestablishment of Type IIa EBV infected B cell population, evenfor a short period, by various means can be exploited for theirfurther characterization, particularly in relation to the EBVpositive R/S cells.
In summary, we visualized in various experimental systemsthe simultaneous expression of the EBV encoded proteins,LMP-1 and EBNA-2 in single cells. Some of the results validatethe method and substantiate earlier findings. Use of themethod detected new aspects in the complex interaction ofEBV with its target that can be pursued in future experiments.
Acknowledgments
We thank the Department of Obstetrics and Gynecology,Karolinska University Hospital, for the provision of cord bloodsamples. This work was supported by the Swedish CancerSociety and by the Cancer Research Institute (New York, NY)/Concern Foundation (Los Angeles, CA).
References
Anastasiadou, E., Vaeth, S., Cuomo, L., Boccellato, F., Vincenti, S., Cirone, M.,et al., 2009. Epstein–Barr virus infection leads to partial phenotypicreversion of terminally differentiated malignant B cells. Cancer Lett.284, 165.
Babcock, G.J., Hochberg, D., Thorley-Lawson, D.A., 2000. The expressionpattern of Epstein–Barr virus latent genes in vivo is dependent upon thedifferentiation stage of the infected B cell. Immunity 13, 497.
Brink, A.T.P., Dukers, D.F., van den Brule, A.J.C., Oudejans, J.J., Middeldorp,J.P., Meijer, C.J.L.M., Jiwa, M., 1997. Presence of Epstein–Barr viruslatency type III at the single cell level in post-transplantationlymphoproliferative disorders and AIDS related lymphomas. J. Clin.Pathol. 50, 911.
Doyle, M.G., Catovsky, D., Crawford, D.H., 1993. Infection of leukaemic Blymphocytes by Epstein–Barr virus. Leukemia 7, 1858.
Ernberg, I., Falk, K., Minatovits, J., P., busson, Tursz, T., et al., 1989. The role ofmethylation in the phenotype-dependent modulation of Epstein–barrnuclear antigen 2 and latent membrane protein genes in cells latentlyinfected with Epstein–Barr virus. J. Gen. Virol. 70, 2989.
Gerdes, J., Schwab, U., Lemke, H., Stein, H., 1983. Production of a mousemonoclonal antibody reactive with a human nuclear antigen associatedwith cell proliferation. Int. J. Cancer 31, 13.
Gregory, C.D., Rowe, M., Rickinson, A.B., 1990. Different Epstein–Barr virus-Bcell interactions in phenotypically distinct clones of a Burkitt lymphomacell line. J. Gen. Virol. 71, 1481.
Hjalgrim, H., Smedby, K.E., Rostgaard, K., et al., 2007. Infectious mononucleosis,childhood social environment, and risk of Hodgkin lymphoma. Cancer Res.67, 2382.
Khanna, R., Moss, D.J., Burrows, S.R., 1999. Vaccine strategies againstEpstein–Barr virus associated diseases: lessons from studies on cytotoxicT-cell-mediated immune regulation. Immunol. Rev. 170, 49.
Kis, L.L., Nishikawa, J., Takahara, M., Nagy, N., Matskova, L., Takada, K., et al.,2005. In vitro EBV-infected sub line of KMH2, derived from Hodgkinlymphoma, expresses only EBNA-1, while CD40 ligand and IL-4 induceLMP-1 but not EBNA-2. Int. J. Cancer 113, 937.
Kis, L.L., Salamon, D., Persson, E.K., Nagy, N., Scheeren, F.A., Spits, H., Klein, G.,Klein, E., 2010. Il-21 imposes a type II EBV gene expression on type IIIand type I B cells by the repression of C- and activation of LMP-1-promoter. Proc. Natl. Acad. Sci. USA 107, 872–877.
Klein, G., 1994. Epstein–Barr virus strategy in normal and neoplastic B cells.Cell 77, 791–793.
Klein, E., Claesson, H.E., 2010. The central role of leukotriene B4 in Epstein–Barrvirus activated innate immunity in vitro. Curr. Immunol. Rev. 6, 23.
Klein, G., Lindahl, T., Jondal, M., Leibold, W., Menezes, J., Nilsson, K., Sundstrom,C., 1974. Continuous lymphoid cell lines with characteristics of B Cells(bone-marrow-derived), lacking the Epstein–Barr virus genome andderived from three human lymphomas. PNAS 71, 3283.
Klein, E., Kis, L.L., Klein, G., 2007. Epstein–Barr virus infection in humans:from harmless to life endangering virus–lymphocyte interactions.Oncogene 26, 1297.
Klein, E., Liu, A., Claesson, H.E., 2008. Activation of innate immunity byleukotriene B4 inhibits EBV induced B-cell transformation in cord bloodderived mononuclear cultures. Immunol. Lett. 116, 174.
Konforte, D., Simard, N., Paige, C.J., 2008. Interleukin-21 regulates expressionof key Epstein–Barr virus oncoproteins, EBNA2 and LMP1, in infectedhuman B cells. Virology 374, 100.
Küppers, R., 2003. B cells under influence: transformation of B cells byEpstein–Barr virus. Nat. Rev. Immunol. 3, 801.
Küppers, R., 2009. The biology of Hodgkin's lymphoma. Nat. Rev. Cancer9, 15.
Kurth, J., Spieker, T., Wustrow, J., Strickler, J.G., Rajewsky, K., Küppers, R.,2000. EBV-infected B cells in infectious mononucleosis: viral strategiesfor spreading in the B cell compartment and establishing latency.Immunity 13, 485.
Kurth, J., Hansmann, M.L., Rajewsky, R., Küppers, R., 2003. Epstein–Barr virusinfected B cells expanding in germinal centers of infectiousmononucleosispatients do not participate in the germinal center reaction. PNAS 100,4730.
Lam, N., Sandberg, M.L., Sugden, B., 2004. High physiological levels of LMP1result in phosphorylation of eIF2α in Epstein–Barr virus-infected cells.J. Virol. 78, 1657.
Liu, A., Claesson, H.E., Mashid, Y., Klein, G., Klein, E., 2008. Leukotriene B4
activates T cell that inhibit B-cell proliferation in EBV-infected cordblood-derived mononuclear cell cultures. Blood 111, 2693.
Long, H.M., Taylor, G.S., Rickinson, A.B., 2011. Immune defence against EBVand EBV-associated disease. Curr. Opin. Immunol. 23, 258.
Ma, S.D., Hegde, S., Young, K.H., Sullivan, R., Rajesh, D., Zhou, Y., et al., 2011. Anew model of Epstein–Barr virus infection reveals an important role forearly lytic viral protein expression in the development of lymphomas.J. Virol. 85, 165.
Maeda, A., Bandobashi, K., Nagy, N., Teramoto, N., Gogol·k, P., Pokrovskaja,K., SzÈkely, L., Bjˆrkholm, M., Klein, G., Klein, E., 2001. Epstein–Barr viruscan infect B-chronic lymphocytic leukaemia cells but it does notorchestrate the cell cycle regulatory proteins. J. Hum. Virol. 4, 227.

70 A.E. Rasul et al. / Journal of Immunological Methods 385 (2012) 60–70
Masucci, M.G., Torsteinsdottir, S., Colombani, C., Brautbar, C., Klein, E., Klein,G., 1987. Down-regulation of HLA antigens and of the Epstein–Barrvirus-encoded latent membrane protein in Burkitt lymphoma lines.Proc. Natl. Acad. Sci. 84, 4567.
Nagy, N., Ádori, M., Rasul, A., Heuts, F., Salamon, D., Ujvári, D., Madapura, H.S.,Leveau, B., Klein, G., Klein, E., 2012. Soluble factors produced by activatedCD4+ T cells modulate EBV latency. Proc. Natl. Acad. Sci. 109, 1512.
Niedobitek, G., Agathanggelou, A., Herbst, H., Whitehead, L., Wright, D.H.,Young, L.S., 1997. Epstein–Barr virus (EBV) infection in infectiousmononucleosis: virus latency, replication and phenotype. J. Pathol.182, 151.
Oudejans, J.J., Jiwa, M., van den Brule, A.J.C., Grasser, F.A., Horstman, A., Vos, W., etal., 1995. Detection of heterogeneous Epstein–Barr virus gene expressionpatterns within individual post-transplantation lymphoproliferative disor-ders. Am. J. Pathol. 147, 923.
Pearson, G., Dewey, F., Klein, G., Henle, G., Henle, W., 1970. Relation betweenneutralization of Epstein–Barr virus and antibodies to cell membraneantigens induced by the virus. J. Natl. Cancer Inst. 45, 989.
Pope, J.H., Horne, M.K., Scott, W., 1968. Transformation of foetal humanleukocytes in vitro by filtrates of a human leukaemic cell line containingherpes-like virus. Int. J. Cancer 3, 857.
Rickinson, A.B., Moss, D.J., 1997. Human cytotoxic T lymphocyte responses toEostein–Baar virus infection. Annu. Rev. Immunol. 15, 405.
Rickinson, A.B., Finerty, S., Epstein, M.A., 1982. Interaction of Epstein–Barrvirus with leukaemic B cells in vitro. I. Abortive infection and rare cellline establishment from chronic lymphocytic leukaemic cells. Clin. Exp.Immunol. 50, 347.
Rosén, A., Bergh, A.C., Gogolák, P., Evaldsson, C., Myhrinder, A.L., Hellqvist, E.,Rasul, A., et al., 2012. Lymphoblastoid cell line with B1 cell character-istics established from a chronic lymphocytic leukaemia clone by in vitroEBV infection. OncoImmunology 1, 18.
Rowe, M., Lear, A.L., Croom-Carter, D., Davies, A.H., Rickinson, A.B., 1992.Three pathways of Epstein–Barr virus gene activation from EBNA1-positive latency in B lymphocytes. J. Virol. 66, 122.
Stacchini, A., Argano, M., Vallario, A., Circosta, P., Gottardi, D., Faldella, A., et al.,1999. MEC1 and MEC2: two new cell lines derived from B-chroniclymphocytic leukaemia in prolymphocytoid transformation. Leuk. Res.23, 127.
Straus, S.E., Cohen, J.I., Tosato, G., Meier, J., 1993. Epstein–Barr virus infections,biology, pathogenesis, and management. Ann. Intern. Med. 118, 45.
Swerdlow, S.H., Webber, S.A., Chadburn, A., et al., 2008. Post-transplantLymphoproliferative Disorders (PTLD). WHO classification of tumors ofhaematopoitic and lymphoid tissue. IARC Press, Lyon, In. pp 343.
Takada, K., Yamamoto, K., Osato, T., 1980. Analysis of the transformation ofhuman lymphocytes by Epstein–Barr virus. II. Abortive response ofleukemic cells to the transforming virus. Intervirology 13, 223.
Teramoto, Gogolak, N.P., Nagy, N., Maeda, A., Kvarnung, K., T., Bjorkholm, etal., 2000. Epstein–Barr virus-infected B-chronic lymphocyte leukemiacells express the virally encoded nuclear proteins but they do not enterthe cell cycle. J. Hum. Virol. 3, 125.
Thorley-Lawson, D.A., 2001. Epstein–Barr virus: exploiting the immunesystem. Nat. Rev. Immunol. 1, 75.
Thorley-Lawson, D.A., Allday, M.J., 2008. The curious case of the tumor virus:50 years of Burkitt's lymphoma. Nat. Rev. Microbiol. 6, 913.
Walls, E.V., Doyle, M.G., Patel, K.K., Allday, M.J., Catovsky, D., Crawford, D.H.,1989. Activation and immortalization of leukaemic B cells by Epstein–Barr virus. Int. J. Cancer 44, 846.
Yao, Q.Y., Rickinson, A.B., Epstein, M.A., 1985. A re-examination of theEpstein–Barr virus carrier state in healthy seropositive individuals. Int.J. Cancer 35, 35.
Young, L.S., Rickinson, A.B., 2004. Epstein–Barr virus: 40 years on. Nat. Rev.Cancer 4, 757.




















