
EBV-positive diffuse large B-cell lymphoma of the elderly: A case series from Peru
40
doi:10.1182/blood-2013-03-489708 Prepublished online May 6, 2013; Chi Young Ok, Thomas G. Papathomas, L. Jeffrey Medeiros and Ken H. Young EBV-positive diffuse large B-cell lymphoma of the elderly http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: digital object identifier (DOIs) and date of initial publication. the indexed by PubMed from initial publication. Citations to Advance online articles must include final publication). Advance online articles are citable and establish publication priority; they are appeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet Copyright 2011 by The American Society of Hematology; all rights reserved. 20036. the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.org From
Transcript of EBV-positive diffuse large B-cell lymphoma of the elderly: A case series from Peru

doi:10.1182/blood-2013-03-489708Prepublished online May 6, 2013;
Chi Young Ok, Thomas G. Papathomas, L. Jeffrey Medeiros and Ken H. Young EBV-positive diffuse large B-cell lymphoma of the elderly
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 1 -
EBV-Positive Diffuse Large B-Cell Lymphoma of the Elderly
- A New Provisional Entity of Lymphoma in the WHO Classification
Chi Young Ok1, Thomas G. Papathomas2, L. Jeffrey Medeiros1, Ken H. Young1¶
1Department of Hematopathology, The University of Texas MD Anderson Cancer Center,
Houston, Texas, USA; 2Department of Pathology, Erasmus MC University Medical Center
Rotterdam, Rotterdam, The Netherlands
Running title: EBV-positive DLBCL of the elderly
Key words: Epstein-Barr virus, diffuse large B-cell lymphoma, elderly, CD30, NF-κB,
LMP1, EBNA1, EBNA2, Akt, PI3K, Jak-Stat
¶Correspondence:
Ken H. Young, MD, PhD
The University of Texas MD Anderson Cancer Center
Department of Hematopathology
1515 Holcombe Boulevard
Houston, Texas 77030-4009
Phone: 1-713-745-2598
Email: [email protected]
Blood First Edition Paper, prepublished online May 6, 2013; DOI 10.1182/blood-2013-03-489708
Copyright © 2013 American Society of Hematology
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 2 -
Abstract
Epstein-Barr virus (EBV) positive diffuse large B-cell lymphoma (DLBCL) of the elderly,
initially described in 2003, is a provisional entity in the 2008 World Health Organization
classification system, and is defined as an EBV-positive monoclonal large B-cell proliferation
that occurs in patients > 50 years of age and in whom there is no known immunodeficiency or
history of lymphoma. These tumors are more common in Asia, but also occur in North
America and Europe at a low frequency. These neoplasms exhibit a morphologic continuum,
from polymorphous to monomorphous, but morphologic features do not correlate with
prognosis as all patients have a clinically aggressive course. Most EBV-positive DLBCL of the
elderly patients have an activated B-cell immunophenotype and are characterized by prominent
NF-κB activation. Cytogenetic complexity is usually low. In this review, we comprehensively
delineate the data emerging from analyses of EBV latency program, microRNA-mediated EBV
viral oncogenesis, functional genomics of EBV and its biology, as well as differential diagnosis
challenge for EBV-positive DLBCL of the elderly. It is hoped that the improved understanding
of these tumors will lead to development of novel therapeutic approaches, enhance the
effective clinical trials and improve the prognosis.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 3 -
Introduction
The 2008 World Health Organization (WHO) classification of tumors of hematopoietic and
lymphoid tissues has recognized a new provisional entity designated as Epstein-Barr virus-
positive diffuse large B-cell lymphoma (DLBCL) of the elderly. This tumor is defined as an
EBV-positive monoclonal large B-cell lymphoproliferative disorder arising in
immunocompetent patients > 50 years.1 To denote patient age in the definition was a means
of emphasizing the tendency of these tumors to arise in patients of advanced age, but this age
cutoff is clearly arbitrary, and EBV-positive DLBCL of the elderly has been reported rarely in
younger patients.
Since the initial description of 22 patients with EBV-positive DLBCL of the elderly in 2003
by Oyama et al,2 knowledge of EBV-mediated mechanisms of oncogenesis, microRNA
profiles in EBV-positive lymphomas and novel efficient therapeutics for patients with these
neoplasms have been developed. In this review, we discuss the epidemiology,
clinicopathologic features, and differential diagnoses of EBV-positive DLBCL of the elderly.
We also review novel molecular findings including host and virus-induced miRNAs, and
some novel therapeutic approaches.
Epidemiology
The median age of patients with EBV-positive DLBCL of the elderly is 71 years (range, 50-
91), however, younger patients can be affected.3 There is a slight male predominance with a
male to female ratio of 1.4 to 1.4-10 There is a higher prevalence of EBV-positive DLBCL
among East Asians (8.7-11.4%)11-13 compared with less than 5% in Western countries5,6
(Table 1). Meanwhile, Beltran et al4 has reported the highest frequency of EBV-positive
DLBCL in a Peruvian population, 14.9%, using a criterion of ≥ 20% cells with positive EBV-
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 4 -
encoded RNA (EBER) expression. Of note, there are no uniform criteria for the percentage of
EBV-positive cells in EBV-positive DLBCL and this is a limitation when assessing disease
prevalence. The fact that differences in cutoffs for EBER positivity clearly impact the
frequency of EBV positive DLBCL was respectively shown by Wada et al and others.13-15
These authors emphasize the need for establishing uniform criteria for EBV positivity, either
>20%, >50%, or almost all tumor cells. Hofscheier et al interpreted EBER-positive DLBCL
with <20% EBER as clonally unrelated EBV-transformed B-cells or secondary EBV infection
in an established B-cell clone.7 The definitive criterion for EBV positivity in EBV-positive
DLBCL is remained to be agreed.
Epidemiologic features correlate with geographical differences in the literature. In contrast
with East Asian series, cases in Mexico display a relatively younger age at presentation, with
predominantly nodal disease and rare expression of the EBNA2.7 Recent evidence casts a
doubt on geographic and/or ethnic variation in the prevalence of EBV-positive DLBCL of the
elderly. Contrary to earlier reports from the Far East, a report from Northern China showed a
frequency of EBV positivity in only 8 of 212 (3.8%) in patients >50 years old with DLBCL
(using a cutoff of >50% EBER positivity).9
Pathogenetic aspects
From an immunologic viewpoint, aging can be characterized as a dysregulated relationship
between inflammatory and inflammation-neutralizing processes, resulting in a low-grade
chronic pro-inflammatory state.16 This imbalance might exert other potential effects promoting
lymphomagenesis. Inflammation induces radical oxygen species which can cause
dysregulation of critical oncogenic pathways, such as p53, retinoblastoma (Rb), NF-κB, and
mitogen-activated protein (MAP) kinases.17
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 5 -
It has been postulated that EBV-positive DLBCL of the elderly might be caused by the
senescence of the immune system, as a part of the normal aging process, based largely on
shared features with immunodeficiency-associated lymphoproliferative disorders (LPDs).
These shared features include EBV infection, similar EBV latency pattern, morphologic
similarities, and presence of monoclonal T-cell populations.8,12,18,19 Of note,
immunosenescence is not simply the result of a progressive decline of all immune functions,
but rather refers to a continuous remodeling process, whereby the adaptive immunity is
preferentially affected compared with innate immunity.20,21 In this context, B-cell diversity
decreases with age and is characterized by clonal expansions of B cells in vivo.22
Concurrently, the T-cell compartment shows characteristic modifications, such as: (i) a
reduction of the absolute number of total T lymphocytes, including helper/inducer (CD4+) and
suppressor/cytotoxic (CD8+) cell counts; (ii) a progressive decline of the näive T-cell
repertoire with a parallel decline in T-cell receptor repertoire diversity; and (iii) a progressive
expansion of oligoclonal CD28- T cells, particularly among the CD8+ T-cell subset.20,21
Particularly, Ouyang et al23 showed a significantly greater frequency of EBV-specific
receptor-carrying cells within the CD8+ T-cell subset as well as a lower frequency of EBV
antigen-specific interferon (IFN)-gamma-producing T cells in the elderly, suggesting clonal
expansion of dysfunctional EBV-specific cells to age. Whether oligoclonal or monoclonal T-
cell populations in patients with EBV-positive DLBCL of the elderly reflects a physiological
T-cell response to EBV infection8 or antedate the development of the LPD stemming from an
underlying deficiency in T-cell function18 is unknown.
EBV is a gamma-herpes virus that has a tropism for lymphocytes. The virus infects and
replicates within epithelial cells and most viral particles are cleared by cytotoxic T cells.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 6 -
However, EBV survives in a host cell by establishing latency as an episome (a double-
stranded circular form of the genome) in memory B-cells. In latent infection, EBV uses only a
limited number of genes to maintain its genome and evades the host immune reaction. Based
on different patterns of viral gene usage, latency can be classified as type III, type II, or type I.
Type III is characterized by the expression of all six EBV nuclear antigens (EBNA1, 2, 3A,
3B, 3C, and LP), three latent membrane proteins (LMP1, 2A and 2B), and EBER (Figure 1A-
B). Type III latency is observed in patients with infectious mononucleosis or a subset of post-
transplant lymphoproliferative disorders (PTLD). Type II latency is associated with
expression of EBNA1, LMP1, LMP2A, and EBER and is commonly seen in classical
Hodgkin lymphoma and a subset of PTLD. In type I latency, only EBNA1 and EBER are
expressed; this pattern is characteristic of Burkitt lymphoma.
EBV-positive DLBCL of the elderly shares a similar EBV latency patterns (type II or III) with
PTLD.24 Using reverse transcription polymerase chain reaction (RT-PCR) method, Nguyen-
Van et al19 showed that many EBV-positive DLBCL of the elderly and EBV-positive PTLD
resembling DLBCL shared features of EBV latency type III. A relatively high frequency of
EBNA2 expression (28% to 32%) has been observed in several studies.2,12 However,
Hofscheier et al found history of immunosuppression or immunodysfunction in 4/17 cases
with EBNA2 expression, emphasizing that care should be taken in the diagnosis of EBNA2-
positive DLBCL cases.7
EBV-positive DLBCL of the elderly is characterized by prominent classical and alternative
NF-κB pathway activation, with most cases displaying an activated B-cell (ABC)
immunophenotype.8 Whereas NF-κB activation has been known in DLBCL, mostly in the
ABC subtype,25 the association between NF-κB activation and the presence of EBV infection
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 7 -
in DLBCL is much stronger.8 Interestingly, Montes-Moreno et al found NF-κB activation not
only in the ABC but also in the germinal center B-cell (GCB) subtype of EBV-positive
DLBCL, suggesting an additional role for EBV in NF-κB activation independent of the cell of
origin. This finding could be related to c-Rel (a subunit of NF-κB transcription factor)
amplification, which has been observed exclusively in the GCB immunophenotype of DLBCL
on the basis of gene expression profiling.
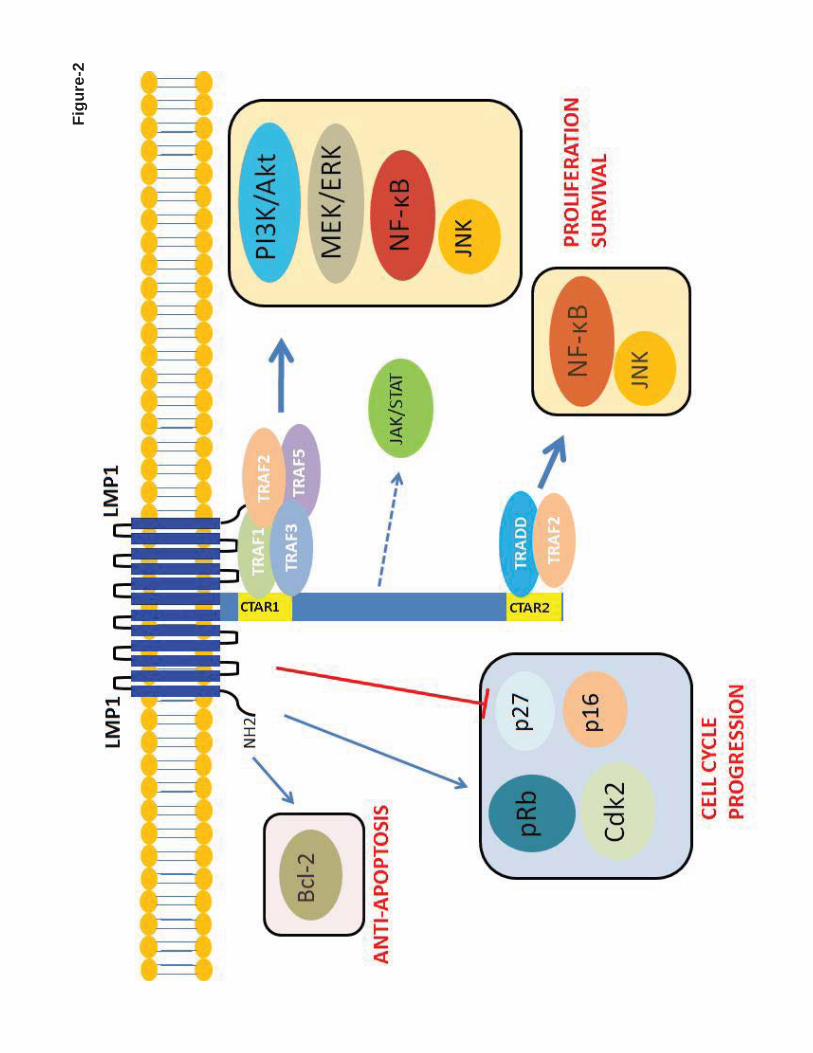
The oncogenic mechanisms of EBV are thought to be attributable predominantly to LMP1
(Figure 2). LMP1 (MW 63kDa) is an integral membrane protein that consists of three different
domains. The N-terminus cytoplasmic domain orientates LMP1 protein and binds to the
plasma membrane. The long C-terminus cytoplasmic tail contains signaling activity. A six-
transmembrane loop between the C- and N-terminus domains provides a platform for self-
aggregation and oligomerization. LMP1 functionally mimics CD40, which is expressed
constitutively on the B-cell membrane involving in B-cell activation and proliferation. Of
note, the function of LMP1 is ligand-independent and is constitutively activated as a result of
self-aggregation and oligomerization.26 The C-terminal tail has two distinct domains, C-
terminal activation regions 1 and 2 (CTAR1 and 2), which activate the NF-κB pathway.27
LMP1 aggregates provide a platform for CTAR1 and CTAR2 to interact with downstream
molecules (Figure 1-C).28-30 LMP1 can also activate the c-Jun N-terminal kinase (JNK)/AP-1,
MAPK, and phosphatidylinositol 3-kinase (PI3K)-AKT pathways.31-33
LMP1 also deregulates the cell cycle. Everly et al showed that LMP1 increases expression of
cyclin-dependent kinase 2 (Cdk2), phosphorylates retinoblastoma (Rb) protein, and inhibits
expression of p27Kip1.34 The p16INK4a-Rb pathway has been shown to be hindered by LMP1.35
LMP1 also increases expression of Bcl-2 to prevent latently EBV-infected cells from
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 8 -
undergoing apoptosis.36 Of note, virus and host transcriptome analyses have revealed an
unexpected coordination between viral lytic reactivation and cellular pathways involved in B-
cell expansion as well as tumor promotion,37 supporting a role for the lytic cycle in virus-
mediated oncogenesis while further highlighting the complexity of interactions between EBV
and host.38
Clinical features
EBV-positive DLBCL of the elderly is characterized by higher age distribution and an
aggressive clinical course with a median survival of two years in Asian patients (Figure 3A-
B).4,7,8,10,12,13,18 Initial reports emphasized that EBV-positive DLBCL of the elderly commonly
involved extranodal sites.2,13 Site of primary extranodal involvement include the skin, soft
tissue, bones, nasal cavity, pharynx/hypopharynx, tonsils, tongue, lung, pleura, stomach, liver,
spleen, peritoneum, cecum and bone marrow.4-8,10,39 Subsequently, several studies showed that
lymph node involvement is very common, in up to 70% of patients.4-8,10,39
EBV-positive DLBCLs, including EBV-positive DLBCL of the elderly, respond more poorly
to treatment with a poorer outcome compared with patients who have EBV-negative DLBCL.
Therefore, novel therapeutic agents are needed to treat patients with EBV-positive DLBCL of
the elderly.13 Montes-Moreno et al8 compared the survival distributions of age-related EBV-
positive DLBCL cases and a control cohort either selecting ABC-type DLBCL cases or
patients > 60 years of age and subsequently confirmed that patients with EBV-positive
DLBCL of the elderly have a poorer overall survival and progression-free survival than
patients with ABC-type EBV-negative DLBCL in older European patients (Figure 3C-D). In
comparison, a negative impact of EBV infection in DLBCL survival was not observed in the
North America patients. There was a trend toward higher percentage of stage III/IV disease
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 9 -
and bone marrow involvement (85% vs 57% and 46% vs 16%, p = 0.052), but no unique
clinical features were found in EBV-positive DLBCL patients (Figure 3E-F). However,
significantly worse survival is observed in EBV-positive DLBCL patients if CD30 expression
is also present (Figure 3G-H).
Recently, Dojcinov et al18 expanded the spectrum of EBV-positive DLBCL. They analyzed
122 cases of EBV-positive B-cell LPDs in adults with no other identifiable cause of
immunosuppression and classified them into four diagnostic categories: (1) reactive lymphoid
hyperplasia; (2) polymorphic extranodal lymphoproliferative disease; (3) polymorphic nodal
lymphoproliferative disease; and (4) DLBCL. In these four groups the five-year disease-
specific survival decreases progressively (100%, 93%, 57%, and 25%, respectively) while the
frequency of monoclonality increases (16%, 33%, 63%, and 56%, respectively). It needs to be
emphasized that these categories are not fixed as patients can progress from reactive
hyperplasia to polymorphic or monomorphic EBV-positive LPD.40 With respect to
morphology, some EBV-positive DLBCL of the elderly display a mixed pattern with
polymorphic and monomorphic areas8 or polymorphic cases eventually progress to the
monomorphic subtype of EBV-positive DLBCL of the elderly5,41.
Two additional entities have been described that further add to the spectrum of EBV-positive
LPDs. EBV-positive mucocutaneous ulcer is a term used recently to designate small-volume
extranodal polymorphic lymphoproliferative disease, particularly involving mucosal sites and
skin, and characterized by an indolent clinical course and good prognosis.18,42 Plasmablastic
lymphoma of the elderly is another newly recognized entity with a relatively indolent clinical
course and better prognosis compared with other age-related EBV-positive B-cell LPDs24.
Plasmablastic lymphoma of the elderly shares features with age-related EBV-positive B-cell
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 10 -
LPDs, including older age, immunosenescence, EBV infection, HIV negativity, and an ABC
immunophenotype, although contrasting features include a higher of extranodal infection and
MYC translocations and EBV type I latency pattern.24
Histopathologic Features
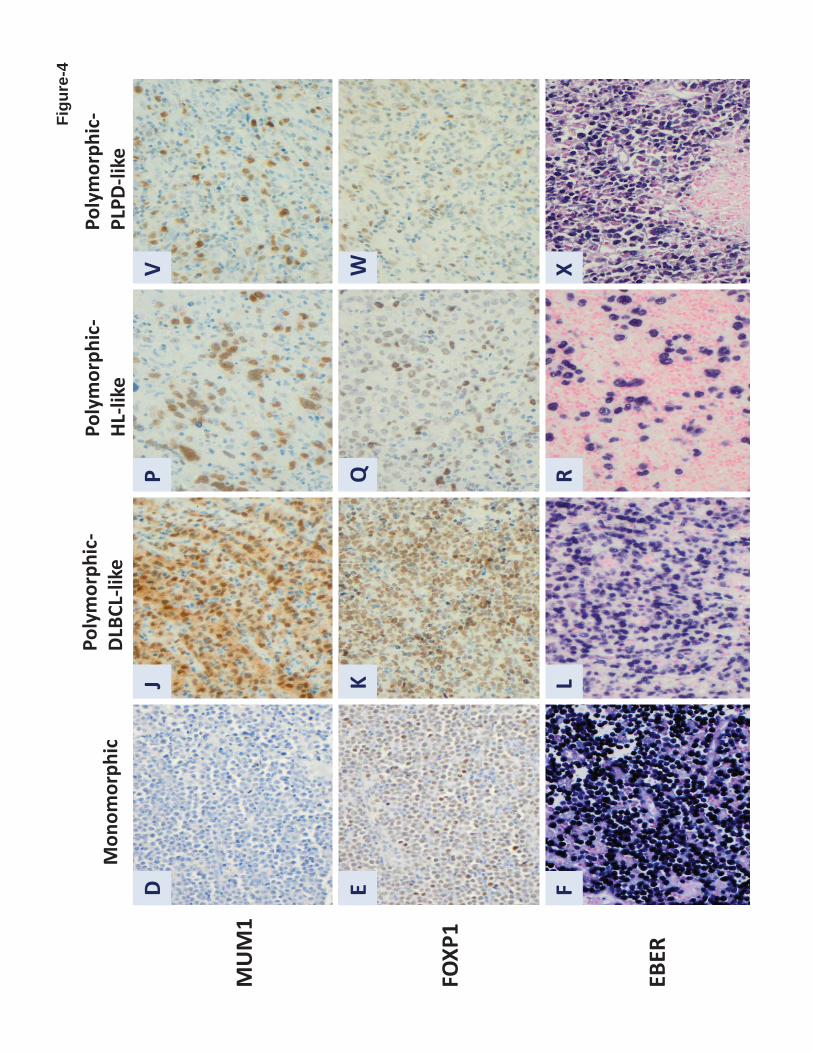
Two morphologic subtypes of EBV-positive DLBCL of the elderly have been recognized,
polymorphic and monomorphic. Both subtypes may include large transformed cells or
immunoblasts as well as Hodgkin and Reed-Sternberg (HRS)-like giant cells and may
demonstrate increased mitotic activity and areas of geographic necrosis.1 The polymorphic
subtype displays a broad range of B-cell maturation, and lesions are composed of centroblasts,
immunoblasts and plasmablasts with a variable component of admixed reactive cells,
including small lymphocytes, plasma cells, histiocytes and epithelioid histiocytes.1 Montes-
Moreno et al8 subdivided the polymorphic EBV-positive DLBCL of the elderly into three
subgroups based on the relative proportion of large neoplastic cells and presence of HRS-like
cells: (1) canonical large B-cell neoplasm (high density of large neoplastic cells and scattered
cells with HRS-like features); (2) DLBCL with Hodgkin lymphoma-like features (lower
density of neoplastic cells with HRS-like features); and (3) DLBCL with polymorphic LPD-
like features (low density of neoplastic cells without HL-like features). The monomorphic
subtype of EBV-positive DLBCL of the elderly is composed of monotonous sheets of large
transformed B cells (Figure 4).1 Although this distinction is helpful for recognizing the
spectrum, these morphologic variants do not impart clinical or prognostic significance.1 Cases
of EBV-positive DLBCL of the elderly also can have a mixed pattern with intermingled
polymorphic/and monomorphic areas8, suggesting that the subtypes represent two ends of a
morphologic spectrum.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 11 -
Immunophenotype
The neoplastic cells are of B-cell lineage as they express the pan B-cell antigens CD19, CD20,
CD22, CD79a and PAX5 and are negative for pan T-cell antigents. Immunoglobulin light
chain restriction may be difficult to demonstrate, except in cases with immunoblastic or
plasmablastic features in which cytoplasmic Ig can be assessed. Plasmacytoid cases can be
weakly positive or negative CD20.1 EBV-positive DLBCL of the elderly usually has an ABC
immunophenotype being MUM1/IRF4+ and CD10- and usually BCL6-. BCL-2 and CD30 are
usually positive; CD15 is negative.1,8 Ki-67 generally shows a high proliferation index.8 By
definition, the neoplastic cells are EBV-positive and accordingly display EBER, LMP1 in >
90% and EBNA2 in approximately 15% and 30% of cases.1,43 The concordance rate between
EBER and LMP1 can reach 80% to 90%, but is not uniformly consistent.
The proportion of EBV-positive DLBCL cases among all DLBCL patients increases with
age,12,44 which is analogous to the ABC subtype that also increases with age.45 These findings
are in accord with recent data indicating that aging is an important determinant of lymphoma
biology.46 In an effort to explain the proportional increase of ABC DLBCL with age,
Mareschal et al45 speculated that either a change in B-cell population during aging or on the
putative pathological specificity of EBV-positive DLBCL of the elderly. Not surprisingly,
most cases of EBV-positive DLBCL of the elderly displayed a striking shift to an ABC
immunophenotype with prominent activation of NF-κB pathway.8
Differential Diagnosis
The major entities in the differential diagnosis of EBV-positive DLBCL of the elderly are
summarized in Table 2. In part, EBV-positive DLBCL of the elderly is a diagnosis of
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 12 -
exclusion. If there is a known cause of immunodeficiency, these tumors are better classified as
post-transplant LPDs, other iatrogenic immunodeficiency-associated LPDs, etc. Other forms of
EBV-positive DLBCL such as lymphomatoid granulomatosis, DLBCL associated with chronic
inflammation, primary effusion lymphoma, and plasmablastic lymphoma are also considered in
the differential diagnosis. Reactive diseases such as infectious mononucleosis of the elderly
and chronic active EBV infection also may be considered based on the morphologic findings
but clinical history and serologic testing are helpful.
Genetic and miRNA profile
Only a few genetic studies on cases of EBV-positive DLBCL of the elderly have been
performed.8,18 The immunoglobulin genes are monoclonally rearranged in most cases, with
clonality of EBV also usually detectable using EBV terminal repeat regions probes and
molecular techniques.1,5,7,8,18,47 IgH-mediated translocations are uncommon (~15%).8,18
However, these analyses were restricted to single loci (IGH, IGK, IGL, PAX5, C-MYC, BCL2
and BCL6). A single case with a t(9;14)(p13;q32) translocation involving the PAX5 gene has
been reported.48 Given the presence of a low number of genomic aberrations in EBV-positive
DLBCL of the elderly, it has been suggested that immunosenescence coupled with the EBV
oncogenic properties are sufficient and additional chromosomal alterations are therefore
usually not needed for lymphomagenesis.8
EBV was the first virus in which miRNAs were identified and 44 mature EBV-miRNAs are
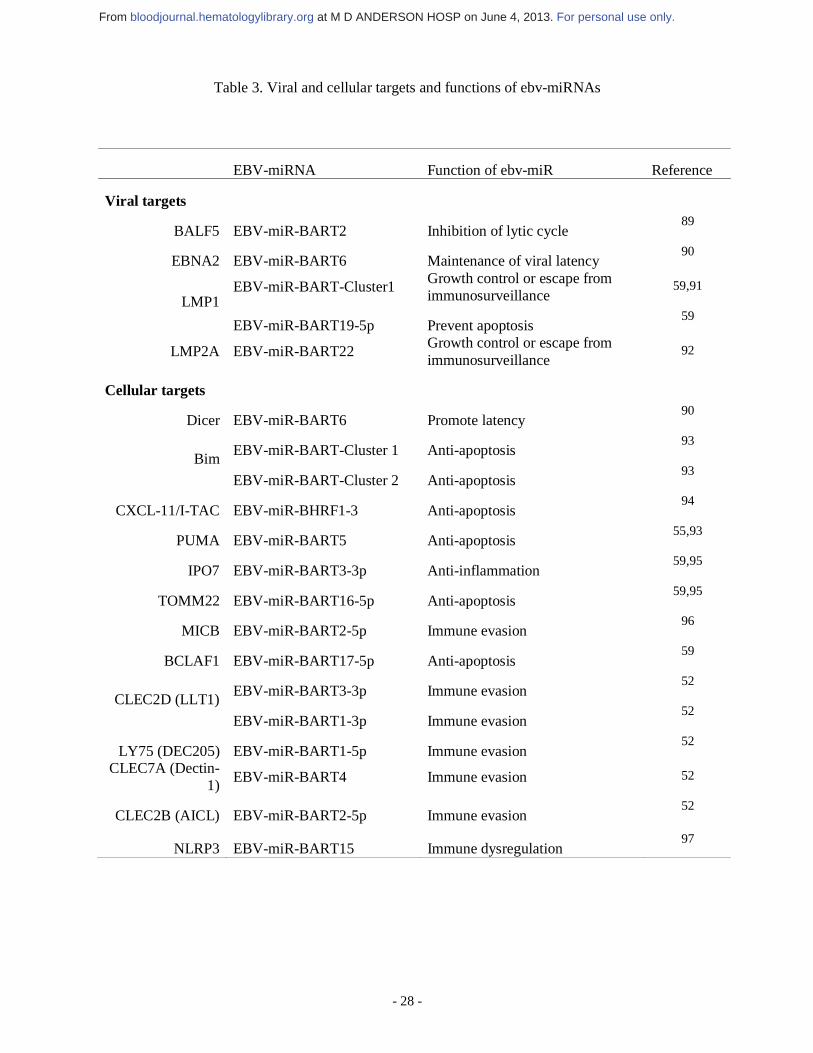
currently known.49 A detailed list of EBV-miRNAs is summarized in Table 3. EBV-miRNAs
can regulate viral and cellular targets, and they generally target host transcripts during latent
infection (Table 3).50,51 Although viral miRNA constitutes only ~2% of all miRNA in EBV-
positive DLBCL, the viral miRNAs share seed sequence homology with cellular miRNA. For
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 13 -
example, EBV-miR-BART5 and EBV-miR-BART1-3p share similarity with cellular miR-18
(miR-17-92 cluster) and miR-29a/b, respectively.52,53 Furthermore, viral miRNAs share seed
sequence homology among themselves.50 Moreover, individual EBV-miRNAs can target
multiple mRNAs, suggesting that many other targets are actually affected and await further
identification.54
Several studies have shown that EBV-miRNAs contribute to EBV-induced oncogenesis.
EBV-miR-BART5 targets and degrades p53-up-regulated modulator of apoptosis, a pro-
apoptotic protein.55 EBV-miR-BHRF1 promotes B-cell transformation and proliferation and
prevents apoptosis.56 EBV-miR-BART9 and BART17-5p can downregulate BCL6
expression.57 Considering the fact that BCL6 represses NF-κB, downregulation of BCL6 by
EBV-miRNA might indirectly contribute to NF-κB activation.57,58 Riley et al59 showed that
numerous EBV-miR-Bam HI-A region rightward transcripts (BARTs) likely target at least
132 apoptotic mRNAs in latency type III infection, highlighting their collective influence on
apoptosis regulation.
It also becomes evident that EBV-miRNAs have evolved to target multiple cellular pathways
rather than a single pathway. Pathway analysis has predicted that there are several EBV-
miRNA targeted genes in at least 20 specific pathways, including p53 feedback loops, B-cell
activation, oxidative stress response, inflammation mediated by chemokines or the cytokine
signaling pathway, the PI3K pathway and apoptosis (Figure 6).52
Interestingly, cellular miRNAs are modulated by viral proteins. Mir-155 has been shown in
DLBCL, especially in ABC subtype, and can be induced by LMP1 via the NF-κB pathway.60
Of note, miR-155 has been shown to co-target INPP5D (SHIP1), a key target implicated in
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 14 -
lymphomagenesis, with EBV-miRNAs.59,61 Mir-34a is induced by LMP1 via the NF-κB
pathway and promotes the growth of EBV-transformed cells.62 EBNA2 has been shown to
induce miR-21 and downregulate miR-146a.63 Overexpression of miR-21 has been found to
promote tumor progression and invasion in solid tumors via targeting PTEN.64,65 Mir-146a has
ben shown to inhibit the NF-κB pathway in the NK/T-cell lymphoma cell lines.66 Therefore,
EBNA2 can provide extra signals via miRNAs to enhance the NF-κB and Akt pathways,
thereby providing survival/proliferation benefits.
Novel therapeutic approaches
Novel therapeutic approaches need to be considered for patients with EBV-positive DLBCL
of the elderly. Possible therapeutic approaches include: (1) EBV-specific adoptive
immunotherapy; (2) microRNA-targeted therapy; (3) combination therapy based on EBV lytic
phase induction followed by exposure of the tumor cells to anti-herpesvirus drugs; and (4)
targeting specific signaling pathways, including the NF-κB pathway (Table 4 and Figure 6).
EBV-specific adoptive immunotherapy, in which EBV-specific cytotoxic T-lymphocytes
(CTLs) are ex vivo activated and expanded and then reinfused seem promising.67 Using this
approach, EBV-specific CTLs could be engineered to recognize and lyse tumor cell targets via
chimeric receptors, while maintaining their ability to proliferate in response to EBV target
antigens and accordingly destroy virus-infected cells.68 Optimal tumor eradication appears to
require proper target antigen selection, co-stimulatory signalling and the ability of chimeric
antigen receptor-modified T-cells to traffic, persist, and retain function following adoptive
transfer.69 Recent evidence has suggested that both EBV-positive monomorphic PTLD that
resemble DLBCL and EBV-positive DLBCL of the elderly share features consistent with type
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 15 -
III EBV latency, including immunodominant EBNA3A protein expression, indicating that
such an approach may be feasible.19
Given the aforementioned complex interplay between host and viral-encoded miRNAs in
EBV-mediated oncogenesis, targeting miRNA as a therapeutic approach appears to be
promising. Various approaches have been proposed: (1) anti-miRNA oligonucleotides are
inhibitory molecules blocking the interactions between miRNAs and their target mRNAs by
interacting directly to the target miRNA; (2) miRNA mask is a sequence with perfect
complementarity to the binding site for a miRNA in the target mRNA, which can form a
duplex with the target mRNA with higher affinity and block the access of the miRNA; (3)
miRNA sponges are synthetic decoys that contain multiple binding sites for a miRNA of
interest, preventing the interaction between miRNA and its targets; (4) small molecule
inhibitors against specific miRNAs can specifically inhibit miRNA synthesis; (5) restoring
activity of miRNAs with oncosuppressive functions; and (6) using miRNA to sensitize tumors
to known and efficient cancer therapeutic modalities.70
Another potential approach could be combining induction of EBV lytic phase with subsequent
exposure to anti-herpesvirus drugs.71 Lytic phase inducers may refer to phorbol esters, DNA
methylase transferase inhibitors, histone deacetylase (HDAC) inhibitors, proteasome
inhibitors, B-cell receptor-blocking antibodies, chemotherapeutic drugs and cellular
miRNAs.71,72 Although proof of principle of a combination therapy approach has been
documented in a clinical trial,73 the optimal pharmacological lytic phase inducer remains to be
determined. It has been recently shown that other HDAC inhibitors such as panobinostat
appear to be efficient and potent for the treatment of EBV-associated lymphomas.74
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 16 -
Given that the EBV-positive DLBCL of the elderly is an aggressive post-germinal center B-
cell neoplasm characterized by prominent NF-κB activation8, targeting the NF-κB pathway
constitutes a rational therapeutic approach (Table 4). Bortezomib, a proteasome inhibitor, has
been found to induce apoptosis in EBV lymphoblastoid cell lines and to reduce canonical and
non-canonical activities of NF-κB pathway.75 A recent phase I/II clinical trial with bortezomib
plus R-CHOP in DLBCL abrogated the adverse outcome in non-GCB and showed similar
survival between GCB and non-GCB patients.76 Bay 11-7085, a pharmacological inhibitor of
NF-κB activity, or NF-κB/p65 small interfering RNA (siRNA), can inhibit DLBCL growth in
vitro via induction of apoptosis.25 In contrast with bortezomib, the latter strategies of NF-κB
inhibition down-regulate expression of key NF-κB regulated gene products, including Bcl-2
and Bcl-XL.25,75 NF-κB essential modulator-binding domain peptide has been shown to inhibit
NF-κB target gene expression and reduce tumor burden in an in vivo model as well.77
Recent evidence indicates a critical function of PI3K-PDK1 signaling upstream of NF-κB in
ABC-DLBCL cells.78 PI3K inhibition reduced NF-κB activity and decreased the expression of
NF-κB target genes in ABC-DLBCL cell lines, providing a rationale for the pharmacological
use of PI3K inhibitors for treating patients with ABC-type DLBCL.78 In addition, LMP1 has
been implicated in the activation of the PI3K/Akt signaling pathway, which lies upstream of
mTOR.79 Experimental evidence stemming either from dual PI3K/mTOR inhibitors or a single
mTOR inhibitor further reinforces the putative exploitation of LMP1/LMP2A-activated
PI3K/Akt/mTOR signaling pathway.80 Another attractive target for developing antiviral or
antitumor strategies is EBNA1 owing to its essential role in maintaining the EBV genome and
the consistent expression of EBNA1 in all proliferating EBV-positive cells.81 Sun et al showed
that Hsp90 inhibitors block the outgrowth of EBV-infected malignant cells in vitro and in vivo
through an EBNA1-dependent mechanism.81 Notably, Hsp90 inhibitors have been found to
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 17 -
suppress NF-κB activity.82 A CD30-directed antibody-drug conjugate could be a good option
given frequent CD30 expression in EBV-positive DLBCL patients. A recent trial Brentuximab
Vedotin in 2 cases of EBV-positive DLBCL did not show promising result, but small size trial
might be not representative.83
Conclusions
We have provided a comprehensive review of EBV-positive DLBCL of the elderly discussing
its epidemiologic, pathogenetic and clinicopathologic features, differential diagnosis, and
proposed some comments about potential novel therapeutic approaches for patients with this
disease. Currently, an arbitrary cutoff of 50 years is used to define this entity in the current
WHO classification, but patients younger than 50 years can also be affected. Although these
tumors were originally reported as usually being extranodal, it is clear that nodal presentation
is also seen. Most cases of EBV-positive DLBCL of the elderly have an activated B-cell
immunophenotype. Assessment for EBV infection in tissue biopsy specimen can be easily
performed by in situ hybridization analysis for EBER, but other methods of EBV detection
including PCR can be used. Caution should be exercised before rendering this particular
diagnosis as other causes of EBV-positive DLBCL must be excluded and there can be no
known explanation for immunodeficiency. Clearly, a better understanding of the molecular
genetic basis of EBV-positive DLBCL of the elderly is needed. Further studies are required to
address the heterogeneous EBV latency status and miRNA expression profile of EBV-positive
DLBCL of the elderly, as well as the role of the NF-κB pathway.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 18 -
Acknowledgments
This work was supported by an MD Anderson Pathology Fellowship Award (C.Y.O); The
University of Texas MD Anderson Cancer Center Institutional R & D Fund, an Institutional
Research Grant Award, an MD Anderson Lymphoma SPORE Research Development
Program Award, an MD Anderson Myeloma SPORE Research Development Program Award,
and MD Anderson Collaborative Research Funds with Daiichi Sankyo Pharmaceuticals, High-
Throughput Molecular Diagnostics and Roche Molecular System (K.H.Y.). This work was
also partially supported by National Cancer Institute and National Institute of Health
(R01CA138688, 1RC1CA146299, P50CA136411 and P50CA142509).
Authorship Contributions:
C.Y.O., T.P., L.J.M., and K.H.Y. designed the study, analyzed data, and wrote the paper.
CONFLICTS OF INTEREST DISCLOSURE
The authors declare no conflicts of interest.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 19 -
References
1. Nakamura S, Jaffe ES, Swerdlow SH eds. EBV positive diffuse large B-cell lymphoma of the elderly. In: Swerdlow SH, Campo E, Harris NL, et al. eds. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues. Lyon: International Agency for Research on Cancer; 2008. 2. Oyama T, Ichimura K, Suzuki R, et al. Senile EBV+ B-cell lymphoproliferative disorders: a clinicopathologic study of 22 patients. Am J Surg Pathol. 2003;27(1):16-26. 3. Beltran BE, Morales D, Quinones P, Medeiros LJ, Miranda RN, Castillo JJ. EBV-positive diffuse large b-cell lymphoma in young immunocompetent individuals. Clin Lymphoma Myeloma Leuk. 2011;11(6):512-516. 4. Beltran BE, Castillo JJ, Morales D, et al. EBV-positive diffuse large B-cell lymphoma of the elderly: a case series from Peru. Am J Hematol. 2011;86(8):663-667. 5. Gibson SE, Hsi ED. Epstein-Barr virus-positive B-cell lymphoma of the elderly at a United States tertiary medical center: an uncommon aggressive lymphoma with a nongerminal center B-cell phenotype. Hum Pathol. 2009;40(5):653-661. 6. Hoeller S, Tzankov A, Pileri SA, Went P, Dirnhofer S. Epstein-Barr virus-positive diffuse large B-cell lymphoma in elderly patients is rare in Western populations. Hum Pathol.2010;41(3):352-357. 7. Hofscheier A, Ponciano A, Bonzheim I, et al. Geographic variation in the prevalence of Epstein-Barr virus-positive diffuse large B-cell lymphoma of the elderly: a comparative analysis of a Mexican and a German population. Mod Pathol. 2011;24(8):1046-1054. 8. Montes-Moreno S, Odqvist L, Diaz-Perez JA, et al. EBV-positive diffuse large B-cell lymphoma of the elderly is an aggressive post-germinal center B-cell neoplasm characterized by prominent nuclear factor-kB activation. Mod Pathol. 2012;25(7):968-982. 9. Pan Y, Meng B, Zhang H, et al. Low incidence of Epstein-Barr virus-positive diffuse large B-cell lymphoma of the elderly in Tianjin, northern China. Leuk Lymphoma. 2012;[Epub ahead of print]. 10. Uner A, Akyurek N, Saglam A, et al. The presence of Epstein-Barr virus (EBV) in diffuse large B-cell lymphomas (DLBCLs) in Turkey: special emphasis on 'EBV-positive DLBCL of the elderly'. APMIS. 2011;119(4-5):309-316. 11. Kuze T, Nakamura N, Hashimoto Y, Sasaki Y, Abe M. The characteristics of Epstein-Barr virus (EBV)-positive diffuse large B-cell lymphoma: comparison between EBV(+) and EBV(-) cases in Japanese population. Jpn J Cancer Res. 2000;91(12):1233-1240. 12. Oyama T, Yamamoto K, Asano N, et al. Age-related EBV-associated B-cell lymphoproliferative disorders constitute a distinct clinicopathologic group: a study of 96 patients. Clin Cancer Res. 2007;13(17):5124-5132. 13. Park S, Lee J, Ko YH, et al. The impact of Epstein-Barr virus status on clinical outcome in diffuse large B-cell lymphoma. Blood. 2007;110(3):972-978. 14. Wada N, Ikeda J, Hori Y, et al. Epstein-barr virus in diffuse large B-Cell lymphoma in immunocompetent patients in Japan is as low as in Western Countries. J Med Virol.2011;83(2):317-321. 15. Chuang SS, Ichinohasama R, Yang CC, et al. Multicentric primary intestinal EBV-positive diffuse large B cell lymphoma of the elderly presenting with perforation. Int J Hematol. 2010;91(3):534-538. 16. Franceschi C, Capri M, Monti D, et al. Inflammaging and anti-inflammaging: a systemic perspective on aging and longevity emerged from studies in humans. Mech Ageing Dev. 2007;128(1):92-105. 17. Hussain SP, Hofseth LJ, Harris CC. Radical causes of cancer. Nat Rev Cancer.2003;3(4):276-285.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 20 -
18. Dojcinov SD, Venkataraman G, Pittaluga S, et al. Age-related EBV-associated lymphoproliferative disorders in the Western population: a spectrum of reactive lymphoid hyperplasia and lymphoma. Blood. 2011;117(18):4726-4735. 19. Nguyen-Van D, Keane C, Han E, et al. Epstein-Barr virus-positive diffuse large B-cell lymphoma of the elderly expresses EBNA3A with conserved CD8 T-cell epitopes. Am J Blood Res. 2011;1(2):146-159. 20. Sansoni P, Vescovini R, Fagnoni F, et al. The immune system in extreme longevity. Exp Gerontol. 2008;43(2):61-65. 21. Hakim FT, Gress RE. Immunosenescence: deficits in adaptive immunity in the elderly. Tissue Antigens. 2007;70(3):179-189. 22. Gibson KL, Wu YC, Barnett Y, et al. B-cell diversity decreases in old age and is correlated with poor health status. Aging Cell. 2009;8(1):18-25. 23. Ouyang Q, Wagner WM, Walter S, et al. An age-related increase in the number of CD8+ T cells carrying receptors for an immunodominant Epstein-Barr virus (EBV) epitope is counteracted by a decreased frequency of their antigen-specific responsiveness. Mech Ageing Dev. 2003;124(4):477-485. 24. Liu F, Asano N, Tatematsu A, et al. Plasmablastic lymphoma of the elderly: a clinicopathological comparison with age-related Epstein-Barr virus-associated B cell lymphoproliferative disorder. Histopathology. 2012;[Epub ahead of print]. 25. Bavi P, Uddin S, Bu R, et al. The biological and clinical impact of inhibition of NF-kappaB-initiated apoptosis in diffuse large B cell lymphoma (DLBCL). J Pathol.2011;224(3):355-366. 26. Gires O, Zimber-Strobl U, Gonnella R, et al. Latent membrane protein 1 of Epstein-Barr virus mimics a constitutively active receptor molecule. EMBO J. 1997;16(20):6131-6140. 27. Huen DS, Henderson SA, Croom-Carter D, Rowe M. The Epstein-Barr virus latent membrane protein-1 (LMP1) mediates activation of NF-kappa B and cell surface phenotype via two effector regions in its carboxy-terminal cytoplasmic domain. Oncogene.1995;10(3):549-560. 28. Eliopoulos AG, Young LS. LMP1 structure and signal transduction. Semin Cancer Biol. 2001;11(6):435-444. 29. Izumi KM, Kieff ED. The Epstein-Barr virus oncogene product latent membrane protein 1 engages the tumor necrosis factor receptor-associated death domain protein to mediate B lymphocyte growth transformation and activate NF-kappaB. Proc Natl Acad Sci U S A. 1997;94(23):12592-12597. 30. Mainou BA, Everly DN, Jr., Raab-Traub N. Unique signaling properties of CTAR1 in LMP1-mediated transformation. J Virol. 2007;81(18):9680-9692. 31. Dawson CW, Tramountanis G, Eliopoulos AG, Young LS. Epstein-Barr virus latent membrane protein 1 (LMP1) activates the phosphatidylinositol 3-kinase/Akt pathway to promote cell survival and induce actin filament remodeling. J Biol Chem. 2003;278(6):3694-3704. 32. Roberts ML, Cooper NR. Activation of a ras-MAPK-dependent pathway by Epstein-Barr virus latent membrane protein 1 is essential for cellular transformation. Virology.1998;240(1):93-99. 33. Eliopoulos AG, Waites ER, Blake SM, Davies C, Murray P, Young LS. TRAF1 is a critical regulator of JNK signaling by the TRAF-binding domain of the Epstein-Barr virus-encoded latent infection membrane protein 1 but not CD40. J Virol. 2003;77(2):1316-1328. 34. Everly DN, Jr., Mainou BA, Raab-Traub N. Induction of Id1 and Id3 by latent membrane protein 1 of Epstein-Barr virus and regulation of p27/Kip and cyclin-dependent kinase 2 in rodent fibroblast transformation. J Virol. 2004;78(24):13470-13478.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 21 -
35. Ohtani N, Brennan P, Gaubatz S, et al. Epstein-Barr virus LMP1 blocks p16INK4a-RB pathway by promoting nuclear export of E2F4/5. J Cell Biol. 2003;162(2):173-183. 36. Henderson S, Rowe M, Gregory C, et al. Induction of bcl-2 expression by Epstein-Barr virus latent membrane protein 1 protects infected B cells from programmed cell death. Cell.1991;65(7):1107-1115. 37. Arvey A, Tempera I, Tsai K, et al. An atlas of the Epstein-Barr virus transcriptome and epigenome reveals host-virus regulatory interactions. Cell Host Microbe. 2012;12(2):233-245. 38. White RE, Groves IJ, Turro E, Yee J, Kremmer E, Allday MJ. Extensive co-operation between the Epstein-Barr virus EBNA3 proteins in the manipulation of host gene expression and epigenetic chromatin modification. PLoS One. 2010;5(11):e13979. 39. Nimubona S, Bernard M, De Guibert S, et al. Age-related Epstein-Barr virus-associated B-cell lymphoproliferative disorders in elderly white patients. Histopathology.2010;57(2):320-323. 40. de la Hera Magallanes AI, Montes-Moreno S, Hernandez SG, et al. Early phase of Epstein-Barr virus (EBV)-positive diffuse large B cell lymphoma of the elderly mimicking EBV-positive reactive follicular hyperplasia. Histopathology. 2011;59(3):571-575. 41. Shiozawa E, Saito B, Yamochi-Onizuka T, et al. Senile EBV-associated B-cell lymphoproliferative disorder of indolent clinical phenotype with recurrence as aggressive lymphoma. Pathol Int. 2007;57(10):688-693. 42. Dojcinov SD, Venkataraman G, Raffeld M, Pittaluga S, Jaffe ES. EBV positive mucocutaneous ulcer--a study of 26 cases associated with various sources of immunosuppression. Am J Surg Pathol. 2010;34(3):405-417. 43. Adam P, Bonzheim I, Fend F, Quintanilla-Martinez L. Epstein-Barr virus-positive diffuse large B-cell lymphomas of the elderly. Adv Anat Pathol. 2011;18(5):349-355. 44. Shimoyama Y, Yamamoto K, Asano N, Oyama T, Kinoshita T, Nakamura S. Age-related Epstein-Barr virus-associated B-cell lymphoproliferative disorders: special references to lymphomas surrounding this newly recognized clinicopathologic disease. Cancer Sci.2008;99(6):1085-1091. 45. Mareschal S, Lanic H, Ruminy P, Bastard C, Tilly H, Jardin F. The proportion of activated B-cell like subtype among de novo diffuse large B-cell lymphoma increases with age. Haematologica. 2011;96(12):1888-1890. 46. Klapper W, Kreuz M, Kohler CW, et al. Patient age at diagnosis is associated with the molecular characteristics of diffuse large B-cell lymphoma. Blood. 2012;119(8):1882-1887. 47. Shimoyama Y, Asano N, Kojima M, et al. Age-related EBV-associated B-cell lymphoproliferative disorders: diagnostic approach to a newly recognized clinicopathological entity. Pathol Int. 2009;59(12):835-843. 48. Ohno H, Nishikori M, Haga H, Isoda K. Epstein-Barr virus-positive diffuse large B-cell lymphoma carrying a t(9;14)(p13;q32) translocation. Int J Hematol. 2009;89(5):704-708. 49. Pfeffer S, Zavolan M, Grasser FA, et al. Identification of virus-encoded microRNAs. Science. 2004;304(5671):734-736. 50. Imig J, Motsch N, Zhu JY, et al. microRNA profiling in Epstein-Barr virus-associated B-cell lymphoma. Nucleic Acids Res. 2011;39(5):1880-1893. 51. Barth S, Meister G, Grasser FA. EBV-encoded miRNAs. Biochim Biophys Acta. 2011;1809(11-12):631-640. 52. Skalsky RL, Corcoran DL, Gottwein E, et al. The viral and cellular microRNA targetome in lymphoblastoid cell lines. PLoS Pathog. 2012;8(1):e1002484. 53. Babu SG, Ponia SS, Kumar D, Saxena S. Cellular oncomiR orthologue in EBV oncogenesis. Comput Biol Med. 2011;41(10):891-898. 54. Marquitz AR, Raab-Traub N. The role of miRNAs and EBV BARTs in NPC. Semin Cancer Biol. 2012;22(2):166-172.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 22 -
55. Choy EY, Siu KL, Kok KH, et al. An Epstein-Barr virus-encoded microRNA targets PUMA to promote host cell survival. J Exp Med. 2008;205(11):2551-2560. 56. Seto E, Moosmann A, Gromminger S, Walz N, Grundhoff A, Hammerschmidt W. Micro RNAs of Epstein-Barr virus promote cell cycle progression and prevent apoptosis of primary human B cells. PLoS Pathog. 2010;6(8):e1001063. 57. Martin-Perez D, Vargiu P, Montes-Moreno S, et al. Epstein-Barr virus microRNAs repress BCL6 expression in diffuse large B-cell lymphoma. Leukemia. 2012;26(1):180-183. 58. Perez-Rosado A, Artiga M, Vargiu P, Sanchez-Aguilera A, Alvarez-Barrientos A, Piris M. BCL6 represses NFkappaB activity in diffuse large B-cell lymphomas. J Pathol.2008;214(4):498-507. 59. Riley KJ, Rabinowitz GS, Yario TA, Luna JM, Darnell RB, Steitz JA. EBV and human microRNAs co-target oncogenic and apoptotic viral and human genes during latency. EMBO J. 2012;31(9):2207-2221. 60. Gatto G, Rossi A, Rossi D, Kroening S, Bonatti S, Mallardo M. Epstein-Barr virus latent membrane protein 1 trans-activates miR-155 transcription through the NF-kappaB pathway. Nucleic Acids Res. 2008;36(20):6608-6619. 61. O'Connell RM, Chaudhuri AA, Rao DS, Baltimore D. Inositol phosphatase SHIP1 is a primary target of miR-155. Proc Natl Acad Sci U S A. 2009;106(17):7113-7118. 62. Forte E, Salinas RE, Chang C, et al. The Epstein-Barr virus (EBV)-induced tumor suppressor microRNA MiR-34a is growth promoting in EBV-infected B cells. J Virol.2012;86(12):6889-6898. 63. Rosato P, Anastasiadou E, Garg N, et al. Differential regulation of miR-21 and miR-146a by Epstein-Barr virus-encoded EBNA2. Leukemia. 2012;26(11):2343-2352. 64. Zhang BG, Li JF, Yu BQ, Zhu ZG, Liu BY, Yan M. microRNA-21 promotes tumor proliferation and invasion in gastric cancer by targeting PTEN. Oncol Rep. 2012;27(4):1019-1026. 65. Xiong B, Cheng Y, Ma L, Zhang C. MiR-21 regulates biological behavior through the PTEN/PI-3 K/Akt signaling pathway in human colorectal cancer cells. Int J Oncol.2013;42(1):219-228. 66. Paik JH, Jang JY, Jeon YK, et al. MicroRNA-146a downregulates NFkappaB activity via targeting TRAF6 and functions as a tumor suppressor having strong prognostic implications in NK/T cell lymphoma. Clin Cancer Res. 2011;17(14):4761-4771. 67. Martorelli D, Muraro E, Merlo A, et al. Exploiting the interplay between innate and adaptive immunity to improve immunotherapeutic strategies for Epstein-Barr-virus-driven disorders. Clin Dev Immunol. 2012;2012:931952. 68. Rossig C, Bollard CM, Nuchtern JG, Rooney CM, Brenner MK. Epstein-Barr virus-specific human T lymphocytes expressing antitumor chimeric T-cell receptors: potential for improved immunotherapy. Blood. 2002;99(6):2009-2016. 69. Curran KJ, Pegram HJ, Brentjens RJ. Chimeric antigen receptors for T cell immunotherapy: current understanding and future directions. J Gene Med. 2012;14(6):405-415. 70. Kunej T, Godnic I, Horvat S, Zorc M, Calin GA. Cross talk between microRNA and coding cancer genes. Cancer J. 2012;18(3):223-231. 71. Ghosh SK, Perrine SP, Faller DV. Advances in Virus-Directed Therapeutics against Epstein-Barr Virus-Associated Malignancies. Adv Virol. 2012;2012:509296. 72. Ellis-Connell AL, Iempridee T, Xu I, Mertz JE. Cellular microRNAs 200b and 429 regulate the Epstein-Barr virus switch between latency and lytic replication. J Virol.2010;84(19):10329-10343.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 23 -
73. Perrine SP, Hermine O, Small T, et al. A phase 1/2 trial of arginine butyrate and ganciclovir in patients with Epstein-Barr virus-associated lymphoid malignancies. Blood.2007;109(6):2571-2578. 74. Ghosh SK, Perrine SP, Williams RM, Faller DV. Histone deacetylase inhibitors are potent inducers of gene expression in latent EBV and sensitize lymphoma cells to nucleoside antiviral agents. Blood. 2012;119(4):1008-1017. 75. Zou P, Kawada J, Pesnicak L, Cohen JI. Bortezomib induces apoptosis of Epstein-Barr virus (EBV)-transformed B cells and prolongs survival of mice inoculated with EBV-transformed B cells. J Virol. 2007;81(18):10029-10036. 76. Ruan J, Martin P, Furman RR, et al. Bortezomib plus CHOP-rituximab for previously untreated diffuse large B-cell lymphoma and mantle cell lymphoma. J Clin Oncol.2011;29(6):690-697. 77. Gaurnier-Hausser A, Patel R, Baldwin AS, May MJ, Mason NJ. NEMO-binding domain peptide inhibits constitutive NF-kappaB activity and reduces tumor burden in a canine model of relapsed, refractory diffuse large B-cell lymphoma. Clin Cancer Res.2011;17(14):4661-4671. 78. Kloo B, Nagel D, Pfeifer M, et al. Critical role of PI3K signaling for NF-kappaB-dependent survival in a subset of activated B-cell-like diffuse large B-cell lymphoma cells. Proc Natl Acad Sci U S A. 2011;108(1):272-277. 79. Hatton O, Martinez OM, Esquivel CO. Emerging therapeutic strategies for Epstein-Barr virus+ post-transplant lymphoproliferative disorder. Pediatr Transplant. 2012;16(3):220-229. 80. Vaysberg M, Balatoni CE, Nepomuceno RR, Krams SM, Martinez OM. Rapamycin inhibits proliferation of Epstein-Barr virus-positive B-cell lymphomas through modulation of cell-cycle protein expression. Transplantation. 2007;83(8):1114-1121. 81. Sun X, Barlow EA, Ma S, et al. Hsp90 inhibitors block outgrowth of EBV-infected malignant cells in vitro and in vivo through an EBNA1-dependent mechanism. Proc Natl Acad Sci U S A. 2010;107(7):3146-3151. 82. Boll B, Eltaib F, Reiners KS, et al. Heat shock protein 90 inhibitor BIIB021 (CNF2024) depletes NF-kappaB and sensitizes Hodgkin's lymphoma cells for natural killer cell-mediated cytotoxicity. Clin Cancer Res. 2009;15(16):5108-5116. 83. Jacobsen ED, Advani RH, Oki Y, et al. A Phase 2 Study of Brentuximab Vedotin in Patients with Relapsed or Refractory CD30-Positive Non-Hodgkin Lymphomas: Interim Results. 2012 ASH Meeting Abstract. 2012;#2746. 84. Hristov AC. Primary cutaneous diffuse large B-cell lymphoma, leg type: diagnostic considerations. Arch Pathol Lab Med. 2012;136(8):876-881. 85. Kodama K, Massone C, Chott A, Metze D, Kerl H, Cerroni L. Primary cutaneous large B-cell lymphomas: clinicopathologic features, classification, and prognostic factors in a large series of patients. Blood. 2005;106(7):2491-2497. 86. Kojima M, Kashimura M, Itoh H, et al. Epstein-Barr virus-related reactive lymphoproliferative disorders in middle-aged or elderly patients presenting with atypical features. A clinicopathological study of six cases. Pathol Res Pract. 2007;203(8):587-591. 87. Kojima M, Suzuki K, Shimizu S, Masawa N, Nakamura N. Lymph node lesion of acute infectious mononucleosis in the elderly : a case report. J Clin Exp Hematop. 2011;51(2):151-153. 88. Wang E, Papavassiliou P, Sebastian S. A malignant lymphoma with histological features and immunophenotypic profile intermediate between EBV-positive diffuse large B-cell lymphoma and EBV-positive classical Hodgkin lymphoma in a 67-year-old female: a "gray zone" lymphoma associated with Epstein-Barr virus in the elderly. Pathol Res Pract.2012;208(6):363-367.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 24 -
89. Barth S, Pfuhl T, Mamiani A, et al. Epstein-Barr virus-encoded microRNA miR-BART2 down-regulates the viral DNA polymerase BALF5. Nucleic Acids Res.2008;36(2):666-675. 90. Iizasa H, Wulff BE, Alla NR, et al. Editing of Epstein-Barr virus-encoded BART6 microRNAs controls their dicer targeting and consequently affects viral latency. J Biol Chem.2010;285(43):33358-33370. 91. Lo AK, To KF, Lo KW, et al. Modulation of LMP1 protein expression by EBV-encoded microRNAs. Proc Natl Acad Sci U S A. 2007;104(41):16164-16169. 92. Lung RW, Tong JH, Sung YM, et al. Modulation of LMP2A expression by a newly identified Epstein-Barr virus-encoded microRNA miR-BART22. Neoplasia.2009;11(11):1174-1184. 93. Marquitz AR, Mathur A, Nam CS, Raab-Traub N. The Epstein-Barr Virus BART microRNAs target the pro-apoptotic protein Bim. Virology. 2011;412(2):392-400. 94. Xia T, O'Hara A, Araujo I, et al. EBV microRNAs in primary lymphomas and targeting of CXCL-11 by ebv-mir-BHRF1-3. Cancer Res. 2008;68(5):1436-1442. 95. Dolken L, Malterer G, Erhard F, et al. Systematic analysis of viral and cellular microRNA targets in cells latently infected with human gamma-herpesviruses by RISC immunoprecipitation assay. Cell Host Microbe. 2010;7(4):324-334. 96. Nachmani D, Stern-Ginossar N, Sarid R, Mandelboim O. Diverse herpesvirus microRNAs target the stress-induced immune ligand MICB to escape recognition by natural killer cells. Cell Host Microbe. 2009;5(4):376-385. 97. Haneklaus M, Gerlic M, Kurowska-Stolarska M, et al. Cutting edge: miR-223 and EBV miR-BART15 regulate the NLRP3 inflammasome and IL-1beta production. J Immunol.2012;189(8):3795-3799. 98. Kieff E, Rickinson A. Fields Virology. Vol. 2 (ed 5): Wolters Kluwer Health/Lippincott Williams & Wilkins; 2007. 99. Fielding CA, Sandvej K, Mehl A, Brennan P, Jones M, Rowe M. Epstein-Barr virus LMP-1 natural sequence variants differ in their potential to activate cellular signaling pathways. J Virol. 2001;75(19):9129-9141. 100. Perepu U, Dogan A, Maurer M, et al. EBV(+) Diffuse Large B Cell Lymphoma Is Infrequent in Upper Central United States and Lacks Unique Clinical Characteristics or Adverse Prognosis Compared to EBV (-) Counterparts: Results From University of Iowa/Mayo Clinic SPORE. 2012 ASH Meeting Abstract. 2012;#1604.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 25 -
Table 1. Frequency of EBV positivity with different cutoffs in B-cell lymphoproliferative
disorder among different countries
Country EBER+ cases %
EBER cutoff
20%
EBER cutoff
50% Diagnosis Ref
Japan 13/114 11.4 not specified not specified DLBCL 11
Japan 156/1,792 8.7 n/a 156/1,792 (8.7%) B-LPD* 12
Japan 5/460* 1.1 11/460 (2.4%) 5/460 (1.1%) DLBCL 14
Korea 34/380 8.9 34/380 (8.9%) n/a DLBCL 13
Korea 14/468 3.0 n/a 14/468 (3.0%) DLBCL 15
China 10/212 4.7 > 50% DLBCL DLBCL 9
Turkey 12/178* 6.7 not specified not specified DLBCL 10
Peru 28/188* 14.9 28/188 (14.9%) 17/188 (9.0%) DLBCL 4
Mexico 9/134 6.7 9/134 (6.7%) 9/134 (6.7%) DLBCL 7
Germany 4/167 2.4 4/167 (2.4%) 4/167 (2.4%) DLBCL 7
S/I/A 8/258 3.1 7/258 (2.7%) 5/258 (1.9%) DLBCL 6
USA
5 patients
(2002-2007) n/a not specified not specified DLBCL 5
Saudi
Arabia 16/217 7.4 not specified not specified DLBCL 25
*EBER positive rate was re-calculated in cases with an asterisk after excluding
immunodeficiency-associated LPD (including PTLD and HIV-associated lymphoma),
autoimmune-associated LPD, and non-DLBCL patients from each study.
** EBER: EBV-encoded RNA; LPD: lymphoproliferative disorder; S/I/A: Switzerland, Italy,
and Austria; Ref: reference; n/a: not applicable; DLBCL: diffuse large B-cell lymphoma
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

26
Tabl
e 2.
Diff
eren
tial d
iagn
ostic
con
side
ratio
ns o
f EB
V-p
ositi
ve D
LBC
L of
the
elde
rly
Entit
y Si
te
Arc
hite
ctur
e Im
mun
ophe
noty
pe
Spec
ific
findi
ngs
Ligh
t cha
in
Clo
nalit
y
EBV
-pos
itive
DLB
CL,
eld
erly
N
(70%
)
E (3
0%)
Effa
ced
Pan
B m
arke
rs+
CD
10- /B
CL6
- /MU
M1+ ,
EBER
+ ,CD
30+/
-
Geo
grap
hic
necr
osis
M
onot
ypic
M
onoc
lona
l
EBV
-ass
ocia
ted
reac
tive
atyp
ical
lym
phop
rolif
erat
ions
/infe
ctio
us
mon
onuc
leos
is in
eld
erly
N
Not
effa
ced
Mix
ed B
- and
T-c
ell n
atur
e of
HR
S-lik
e ce
lls (C
D3+ /2
0+ /30+ )
EBER
+
n/a
Poly
typi
c Po
lycl
onal
EBV
-pos
itive
cla
ssic
al H
odgk
in
lym
phom
a
N
Effa
ced
CD
20- /C
D79
a- /Pax
5w+
CD
30+ /C
D15
+ , EB
ER+/
-
HR
S ce
lls in
reac
tive
back
grou
nd
Poly
typi
c Po
lycl
onal
DLB
CL,
NO
S, a
napl
astic
var
iant
N
Ef
face
d Pa
n B
mar
kers
+
EBER
- , CD
30+
Sinu
soda
l or c
ohes
ive
patte
rn
Mon
otyp
ic
Mon
oclo
nal
T-ce
ll/hi
stio
cyte
-rich
larg
e B
-cel
l
lym
phom
a
N
Effa
ced
Pan
B m
arke
rs+
EBER
- , CD
30+/
- , CD
15-
Scat
tere
d B
cel
ls in
ben
ign
T-ce
lls/h
istio
cyte
s
back
grou
nd
Mon
otyp
ic
Mon
oclo
nal
Prim
ary
cuta
neou
s DLB
CL,
leg
type
E (S
kin)
D
erm
al
infil
tratio
n
Pan
B-c
ell m
arke
rs+ , E
BER
-
BC
L2+ /M
UM
1+ /FO
XP1
+
Diff
use
shee
ts o
f
cent
robl
asts
and
Mon
otyp
ic
Mon
oclo
nal
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

27
imm
unob
last
s
Lym
phom
atoi
d gr
anul
omat
osis
E po
lym
orph
ic
infil
tratio
n
EBER
+ , LM
P1-
Ang
ioin
vasi
on, H
RS-
like
cells
in re
activ
e ba
ckgr
ound
Non
-
info
rmat
ive
Mon
oclo
nal
Plas
mab
last
ic ly
mph
oma
of th
e
elde
rly
N (5
0%)
E (5
0%)
Effa
ced
CD
38+ , C
D13
8+ , MU
M1+ , C
D20
-
EBER
+ , LM
P1- , E
BN
A2-
Plas
mab
last
ic m
orph
olog
y
EBV
late
ncy
I
Mon
otyp
ic
n/a
Ang
ioim
mun
obla
stic
T-c
ell
lym
phom
a
N
Effa
ced
CD
10+ , C
XC
L13+ , P
D1+
FDC
exp
ansi
on
Poly
typi
c M
onoc
lona
l
in T
-cel
ls
Perip
hera
l T-c
ell l
ymph
oma,
not
othe
rwis
e sp
ecifi
ed
N
Effa
ced
CD
3+/- w
ith p
artia
l los
s of T
-
mar
kers
, CD
30+/
- , CD
15-/+
EBER
-/+
Larg
e at
ypic
al c
ells
clea
r cyt
opla
sm+/
-
Poly
typi
c M
onoc
lona
l
in T
-cel
ls
* Pa
n B
-cel
l mar
kers
: CD
19, C
D20
, CD
22; S
ite N
: nod
al; S
ite E
: ext
rano
dal;
EBV
: Eps
tein
Bar
r viru
s; E
BER
: EB
V-e
ncod
ed R
NA
; DLB
CL:
diff
use
larg
e B
-
cell
lym
phom
a; H
RS:
Hod
gkin
Ree
d-St
ernb
erg;
n/a
: non
-ava
ilabl
e
** R
efer
ence
s 1,
25,4
2-44
,46,
82,8
4-88
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom
- 28 -
Table 3. Viral and cellular targets and functions of ebv-miRNAs
EBV-miRNA Function of ebv-miR Reference
Viral targets
BALF5 EBV-miR-BART2 Inhibition of lytic cycle 89
EBNA2 EBV-miR-BART6 Maintenance of viral latency 90
LMP1EBV-miR-BART-Cluster1 Growth control or escape from
immunosurveillance 59,91
EBV-miR-BART19-5p Prevent apoptosis 59
LMP2A EBV-miR-BART22 Growth control or escape from immunosurveillance
92
Cellular targets
Dicer EBV-miR-BART6 Promote latency 90
Bim EBV-miR-BART-Cluster 1 Anti-apoptosis 93
EBV-miR-BART-Cluster 2 Anti-apoptosis 93
CXCL-11/I-TAC EBV-miR-BHRF1-3 Anti-apoptosis 94
PUMA EBV-miR-BART5 Anti-apoptosis 55,93
IPO7 EBV-miR-BART3-3p Anti-inflammation 59,95
TOMM22 EBV-miR-BART16-5p Anti-apoptosis 59,95
MICB EBV-miR-BART2-5p Immune evasion 96
BCLAF1 EBV-miR-BART17-5p Anti-apoptosis 59
CLEC2D (LLT1) EBV-miR-BART3-3p Immune evasion 52
EBV-miR-BART1-3p Immune evasion 52
LY75 (DEC205) EBV-miR-BART1-5p Immune evasion 52
CLEC7A (Dectin-1) EBV-miR-BART4 Immune evasion 52
CLEC2B (AICL) EBV-miR-BART2-5p Immune evasion 52
NLRP3 EBV-miR-BART15 Immune dysregulation 97
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 29 -
Table 4. Differentially expressed genes in EBV-positive ABC DLBCL vs. EBV-negative ABC DLBCL
A. Genes upregulated in EBV-positive ABC-DLBCL
Gene functional categories
No. of genes
Representative genes
NF-κB target 11 PRDM1, CYLD, FYN, TNFAIP3, FTH1, LITAF
Cell cycle regulation 4 CYTIP, PPM1A, TMEM30A, MRPL41
Anti-apoptosis 4 MDM2, DNAJC5, SERPINB9, GSTO1
Tumor progression 5 LAMP1, PTP4A2, DYNLRB1, ASAP1, ERLEC1
Cell proliferation 3 TXN, PDXK, GLUL
Cell adhesion and cytoskeleton
6 GABARAP, WIPF, AOC3, DST, SLAMF7, C1orf38
Intracellular trafficking 4 TMED2, RAB27A, M6PR, FLOT1
Immune response 2 SAMHD1, GBP2
Cell metabolism 12 GPD2, P4HB, GNS, RPN2, APOL6, LASS6
Transcription factors 2 ARNTL, TLE3
Others and unknown 17 RPN2, SLC31A1, M6PR, SETD3, KIAA2013, TPCN2
B. Genes downregulated in EBV-positive ABC-DLBCL
Gene functional categories
No. of genes
Representative genes
Pro-apopototic 2 TBX5, CUL3
Immune response 2 ENPP3, CD79B
Cell metabolism 3 GATM, MOCOS, GALNT6
ECM, adhesion, cytoskeleton
10 CDHR1, CDHR3, CDH11, TRIP6, CLDN10, SMAGP
Signaling and transcription factors
3 CHN2, STYXL1, SPIB
Others and unknown 49 SPINK5, MYO3A, PHF14, LRRC39, MAGEB3, MGP
* ECM: extracellular matrix
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 30 -
Figure legends
Figure 1. Schematic diagram of linear EBV genome (~172 Kb).
(A), In proportion to their size, largely unique (U1-U5), internal repeat (IR1-IR4) and terminal repeat
(TR) sequence domains are demonstrated. Ori P (indicated in red), the origin for latent infection EBV
episome replication, has plasmid maintenance and DNA replication activity. Location of exons for
latent EBV-induced membrane proteins and nuclear proteins and the size of each gene are shown. Of
note, LMP1 transcription has an opposite direction to the other gene transcripts. LMP2 can only be
transcribed when EBV exists as episome because the gene spans across the TR. LMP2A is transcribed
from the exon 1, whereas LMP2B is transcribed from the exon 2. EBNAs are transcribed from
differently spliced primary EBNA transcript. In latency III, EBNA1 transcription begins from Cp or
Wp, whereas it starts from Qp in latency I or II. EBNA-LP has variable sizes due to variable numbers
of repetitive exons.98
(B), Schematic diagram of circular EBV genome. The U1-U5, IR1-IR4, TR, and Ori P sequence
domains are indicated. Open reading frames for latent proteins are shown. Of note, the size of each
open reading frame does not illustrate the size of each latent protein.98
(C), Amino acid sequence of LMP1. C-terminal activating regions (CTARs) are indicated in red.
TRAF-binding motif in CTAR1 and TRADD-binding motif are indicated in blue with an underbar in
the CTAR1 and CTAR2 domains, respectively. Numbers on the right denote the order of the amino
acid sequence.99 Cp: C promoter, Qp: Q promoter, Wp: W promoter, LMP: latent membrane protein,
EBNA: Epstein-Barr virus nuclear antigen, EBNA-LP: EBNA leader protein, EBER: Epstein-Barr
virus-encoded RNA
Figure 2. Schematic diagram of EBV-mediated oncogenic signalling pathway activation in EBV-
positive diffuse large B-cell lymphoma of the elderly. LMP1 has a six-transmembrane domain with a
long cytoplasmic C-terminal chain. It can self-aggregate to activate itself constitutively and provide a
platform to interact with downstream molecules. LMP1 provides a proliferation signal via activating
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 31 -
NF-κB, PI3K/Akt, MEK-ERK, and JNK-AP-1 MAP kinase pathways. It also gives signal for cell
cycle progression by enhancing cyclin-dependent kinase 2 and phosphorylation of retinoblastoma
protein and by inhibiting p16 and p27. LMP1 can also activate bcl-2 to provide an anti-apoptotic
signal. CTAR1 can directly interact with TRAF1, 2, 3, and 5 to activate NF-κB, PI3K/Akt, MEK-
ERK, and JNK-AP-1 MAP kinase pathways. CTAR2 needs TRADD to interact with TRAF2 to
activate NF-κB. Both canonical and non-canonical pathways of NF-κB are activated to give
proliferation signal to the nucleus. There are 3 putative JAK3-binding motifs between CTAR1 and
CTAR2. However, this finding was not reproduced by others.26-36 AP-1: activator protein 1, CTAR:
C-terminal activation region, ERK: extracellular signal-regulated kinases, JAK3: Janus kinase 3, JNK:
Jun amino-terminal kinases, MEK: MAPK (mitogen-activated protein kinase)/ERK kinase, PI3K:
phosphatidylinositol 3-kinase, TRADD: tumor necrosis factor receptor type 1-associated DEATH
domain, TRAF: tumor necrosis factor receptor-associated factor
Figure 3. Geographical variations of overall survival and progression-free survival in EBV positive
diffuse large B-cell lymphoma of the elderly. (A and B), EBV positive DLBCL of the elderly showed
worse overall survival and progression-free survival in Korean patients13; (C and D), EBV positive
DLBCL of the elderly were more aggressive compared to > 60 year-old patients with ABC subtype
and GCB subtype in European patients8. (E and F), In North America, EBV positive DLBCL of the
elderly showed no difference in overall survival and progression-free survival in comparison to EBV
negative DLBCL patients100. (G and H), In North America, EBV+CD30+ DLBCL of the elderly
showed significant difference in overall survival and progression-free survival in comparison to either
EBV+CD30- or EBV-D30+ DLBCL patients.
Figure 4. Morphologic variants and immunophenotypic profiling in EBV-positive diffuse large B-cell
lymphoma of the elderly. (A-F), The monomorphic subtype is characterized by monotonous sheets of
large transformed B cells. (G-L), Polymorphic DLBCL-like shows canonical large B-cell neoplasm
morphology, characterized by a high density of large neoplastic cells and scattered cells with (Reed-
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

- 32 -
Sternberg) RS-like and Hodgkin-like features. (M-R), The polymorphic HL-like image, the image
with DLBCL with Hodgkin lymphoma-like features, shows a lower density of neoplastic cells with
RS-like and Hodgkin-like features. (S-X), The polymorphic PLPD-like subtype is DLBCL with
polymorphic lymphoproliferative disorder (LPD)-like features. It is characterized by a low density of
neoplastic cells without HL-like features. EBV-positive DLBCL of the elderly is predominantly the
ABC subtype. Interestingly, the monomorphic case presented in this figure shows the GCB subtype
(A-F). All polymorphic subtypes show the ABC-DLBCL molecular phenotype.
Figure 5. Gene expression profiling in the DLBCL ABC subtype was performed according to EBV
status. The EBV-positive case shows a distinct gene expression pattern compared to cases with EBV
negativity, with overexpression of multiple components of the NF-κB pathway.
Figure 6. Therapeutic modulation of EBV infection and associated signaling pathways. Strategies to
inactivate EBV infection or EBV associated oncogenic pathways have been explored in lymphoma
cells. Many therapeutic targets have been identified. (1) Conventional chemotherapeutic agents or
radiation target DNA. (2) Different strategies disrupting miRNAs can be offered. (3) EBV-specific
cytotoxic T-cells are engineered and re-infused to patients as immunoregulatory therapy approach. (4)
Lytic cycle of EBV is induced by a couple of agents and anti-herpetic agents targeting virus in lytic
cycle are added. Agents targeting (5) NF-κB pathway, (6), PI3K/Akt pathway, (7) PKC or (8) MAPK
pathway can be tried. Monoclonal antibodies, Brentuximab vedotin and Rituximab, are available
targeting, (9) CD30 and (10) CD20, respectively.
For personal use only. at M D ANDERSON HOSP on June 4, 2013. bloodjournal.hematologylibrary.orgFrom

EB
V G
enom
e, 1
72 k
b
Figu
re-1
Figu
re-2
x

100
80
60
40
20
0
100
80
60
40
20
0
100
80
60
40
20
0
100
80
60
40
20
0
20 40 60 80 100 120 140 0 20 40 60 80 100 120 140 0
A
C
E
G
B
D
F
H
Survival Time (months)
Surv
ival
Tim
e (%
)
EBV-CD30+ (n=65)
EBV+CD30+ (n=14)
EBV-CD30+ (n=65)
EBV+CD30+ (n=14)
EBV+ (n=13)
EBV- (n=171)
EBV+ (n=13)
EBV- (n=171)
EBV- (n=346)
EBV+ (n=34)
EBV- (n=346)
EBV+ (n=34)
EBV-, GCB (n=162)
EBV-, ABC (n=78)
EBV+ DLBCL (n=38)
EBV-, GCB (n=162)
EBV-, ABC (n=78)
EBV+ DLBCL (n=38)
P=0.026 P=0.018
P < 0.001 P < 0.001
P = 0.14 P = 0.14
P < 0.0001 P < 0.0001
Overall Survival Progression Free Survival
Figure-3

A B C
G
H I
M
N
O
S T U
Mon
omor
phic
Po
lym
orph
ic-
DLBC
L-lik
e Po
lym
orph
ic-
HL-li
ke
Poly
mor
phic
-PL
PD-li
ke
H &
E
CD10
BCL6
Figu
re-4
D E F
J K L
P Q
R
V W
X
MU
M1
FOXP
1
EBER
Mon
omor
phic
Po
lym
orph
ic-
DLBC
L-lik
e Po
lym
orph
ic-
HL-li
ke
Poly
mor
phic
-PL
PD-li
ke
Figu
re-4

Figu
re-5
EB
V+D
LBC
L EB
V- DLB
CL

Figu
re-6




















