
Sex steroid hormone fluctuations and morphological changes of the reproductive system of the female...
15
JOURNAL OF EXPERIMENTAL ZOOLOGY 289:33–47 (2001) © 2001 WILEY-LISS, INC. Sex Steroid Hormone Fluctuations and Morphological Changes of the Reproductive System of the Female of Octopus vulgaris Throughout the Annual Cycle ANNA DI COSMO, 1 * CARLO DI CRISTO, 1 AND MARINA PAOLUCCI 2 1 Department of Zoology, University of Naples, Federico II, 80134 Napoli, Italy 2 Faculty of Science, University of Sannio, 82100 Benevento, Italy ABSTRACT Sex steroids (17β-estradiol and progesterone) and morphological variations of the reproductive system of the female of Octopus vulgaris from the Bay of Naples were followed over a period of 2 years. The increase in the ovary weight was independent of body weight as demon- strated by the gonado-somatic index (GSI). Both 17β-estradiol and progesterone have been de- tected in the ovary of O. vulgaris, and their concentrations changed in correlation with the ovarian development. No 17β-estradiol or progesterone was found in the hemolymph. 3β-Hydroxysteroid dehydrogenase activity has been detected in the ovary, indicating that in the female of O. vulgaris the reproductive system is a source of sex steroid hormones. According to the morphological changes of the ovary, the ovarian cycle can be divided into the following phases: previtellogenesis; early vitellogenesis, full vitellogenesis and late vitellogenesis. The morphological changes of the ovi- ducts and oviducal glands throughout the reproductive cycle were in accordance with their role in the transport and secretion of gelatinous coat covering the eggs, as well as in sperm storage and sperm reactivation during fertilization. J. Exp. Zool. 289:33–47, 2001. © 2001 Wiley-Liss, Inc. The reproductive strategy of the female of Oc- topus vulgaris, like many cephalopods, consists of a single spawning over a short period of time near the end of the life (Mangold, ’87). The ovary, the oviducts, and the oviducal glands enlarge as sexual maturity approaches. The ovary is a single structure, which contains oocytes at different ma- turity stages (Wells, ’78). Ovarian maturation has been demonstrated to be under the control of the optic glands, which are essential for the develop- ment and maturation of the oocytes during the last phase of ovarian maturity, when yolk is pro- duced by follicle cells and taken up by growing oocytes (Wells and Wells, ’75). Eggs are attached individually to shelters by means of oviducal glands secretion and are protected by females. As a rule, females die after eggs hatch (Boletzky, ’81), showing sexual behavior of high complexity, as is observed with other cephalopods (Sauer et al., ’97). Although there are several reviews on the struc- ture of the reproductive system in the main or- ders of cephalopods (Fields, ’65; Arkhipkin, ’83), a detailed description of the reproductive system, its development, and function has been reported only for decapods such as the squid Loligo opale- scens and Thysanoteuthis rhombus and cuttlefish Sepia officinalis (Thompsett, ’39; von Boletsky, ’83; Kueta et al., ’93). Among octopuses, a general de- scription of the male and female reproductive sys- tem is available only for O. vulgaris (Froesch and Marthy, ’75; Guerra, ’75; Wells, ’78; Smale and Buchan, ’81; Arkhipkin, ’92), Octopus dofleini mar- tini (Mann et al., ’70), and Octopus bimaculoides (Peterson, ’59), while more extensive information on maturity stage determination is available only for Eledone cirrhosa (Boyle and Chevis, ’91, ’92), Octopus mimus (Olivares et al., ’94), and Eledone moschata (Ezzeddine-Najai, ’97). Recently, sex steroid hormones (17β-estradiol, progesterone, androgens), sex binding proteins, and 3β-hydroxysteroid dehydrogenase, a key en- zyme in steroidogenesis, have been reported in the reproductive system of the male of O. vulgaris (Di Grant sponsor: MURST. *Correspondence to: Anna Di Cosmo, Department of Zoology, Uni- versity of Naples, Federico II, via Mezzocannone 8, 80134 Napoli, Italy. E-mail: [email protected] Received 9 January 2000; Accepted 7 June 2000
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Sex steroid hormone fluctuations and morphological changes of the reproductive system of the female...

JOURNAL OF EXPERIMENTAL ZOOLOGY 289:33–47 (2001)
© 2001 WILEY-LISS, INC.
Sex Steroid Hormone Fluctuations andMorphological Changes of the ReproductiveSystem of the Female of Octopus vulgarisThroughout the Annual Cycle
ANNA DI COSMO,1* CARLO DI CRISTO,1 AND MARINA PAOLUCCI2
1Department of Zoology, University of Naples, Federico II, 80134Napoli, Italy
2Faculty of Science, University of Sannio, 82100 Benevento, Italy
ABSTRACT Sex steroids (17β-estradiol and progesterone) and morphological variations of thereproductive system of the female of Octopus vulgaris from the Bay of Naples were followed over aperiod of 2 years. The increase in the ovary weight was independent of body weight as demon-strated by the gonado-somatic index (GSI). Both 17β-estradiol and progesterone have been de-tected in the ovary of O. vulgaris, and their concentrations changed in correlation with the ovariandevelopment. No 17β-estradiol or progesterone was found in the hemolymph. 3β-Hydroxysteroiddehydrogenase activity has been detected in the ovary, indicating that in the female of O. vulgaristhe reproductive system is a source of sex steroid hormones. According to the morphological changesof the ovary, the ovarian cycle can be divided into the following phases: previtellogenesis; earlyvitellogenesis, full vitellogenesis and late vitellogenesis. The morphological changes of the ovi-ducts and oviducal glands throughout the reproductive cycle were in accordance with their role inthe transport and secretion of gelatinous coat covering the eggs, as well as in sperm storage andsperm reactivation during fertilization. J. Exp. Zool. 289:33–47, 2001. © 2001 Wiley-Liss, Inc.
The reproductive strategy of the female of Oc-topus vulgaris, like many cephalopods, consists ofa single spawning over a short period of time nearthe end of the life (Mangold, ’87). The ovary, theoviducts, and the oviducal glands enlarge assexual maturity approaches. The ovary is a singlestructure, which contains oocytes at different ma-turity stages (Wells, ’78). Ovarian maturation hasbeen demonstrated to be under the control of theoptic glands, which are essential for the develop-ment and maturation of the oocytes during thelast phase of ovarian maturity, when yolk is pro-duced by follicle cells and taken up by growingoocytes (Wells and Wells, ’75). Eggs are attachedindividually to shelters by means of oviducalglands secretion and are protected by females. Asa rule, females die after eggs hatch (Boletzky, ’81),showing sexual behavior of high complexity, as isobserved with other cephalopods (Sauer et al., ’97).Although there are several reviews on the struc-ture of the reproductive system in the main or-ders of cephalopods (Fields, ’65; Arkhipkin, ’83),a detailed description of the reproductive system,its development, and function has been reportedonly for decapods such as the squid Loligo opale-
scens and Thysanoteuthis rhombus and cuttlefishSepia officinalis (Thompsett, ’39; von Boletsky, ’83;Kueta et al., ’93). Among octopuses, a general de-scription of the male and female reproductive sys-tem is available only for O. vulgaris (Froesch andMarthy, ’75; Guerra, ’75; Wells, ’78; Smale andBuchan, ’81; Arkhipkin, ’92), Octopus dofleini mar-tini (Mann et al., ’70), and Octopus bimaculoides(Peterson, ’59), while more extensive informationon maturity stage determination is available onlyfor Eledone cirrhosa (Boyle and Chevis, ’91, ’92),Octopus mimus (Olivares et al., ’94), and Eledonemoschata (Ezzeddine-Najai, ’97).
Recently, sex steroid hormones (17β-estradiol,progesterone, androgens), sex binding proteins,and 3β-hydroxysteroid dehydrogenase, a key en-zyme in steroidogenesis, have been reported in thereproductive system of the male of O. vulgaris (Di
Grant sponsor: MURST.*Correspondence to: Anna Di Cosmo, Department of Zoology, Uni-
versity of Naples, Federico II, via Mezzocannone 8, 80134 Napoli,Italy. E-mail: [email protected]
Received 9 January 2000; Accepted 7 June 2000

34 A. DI COSMO ET AL.
Cosmo and Di Cristo, ’96; D’Aniello et al., ’96).17β-Estradiol and progesterone receptors havebeen characterized and immunolocalized in thereproductive system of the female of O. vulgaris(Di Cosmo et al., ’98a,b). Such findings supportthe hypothesized parallelism with the known hor-monal regulation system operating in vertebrates(Wells and Wells, ’59, ’75). This view is also sus-tained by the observation that, in the male of O.vulgaris, gonad may be a source of sex steroid hor-mones (D’Aniello et al., ’95). The parallel betweenthe reproductive physiology of O. vulgaris and ver-tebrates is not a recent discovery. Indeed, back in1959, Wells and Wells identified the optic glandsin O. vulgaris as the endocrine structures involvedin the gonadal maturation and hypothesized thatthey are under the control of the central nervoussystem (CNS). In favor of such a hypothesis, twoneuropeptides, the gonadotropin releasing factor(GnRH), known to be involved in the modulationof pituitary in vertebrates (King and Millar, ’95),and the FMRF amide, a molluscan neuromodu-lator (Cottrel et al., ’83, ’84), have been localizedin the nervous lobes that control the activity ofthe optic gland in O. vulgaris (Di Cosmo and DiCristo, ’98).
In this paper we show the presence of 17β-es-tradiol, progesterone, and 3β-hydroxysteroid de-hydrogenase in the ovary of the O. vulgaris. Ourfindings suggest a possible correlation between sexsteroid hormones and the morphological variationsof the reproductive system (ovary, oviducts, andoviducal glands) in this species during the repro-ductive cycle.
MATERIALS AND METHODSAnimals
About 100 females of O. vulgaris of the harborof Naples were collected from October to July dur-ing 2 years (1996–1998). In O. vulgaris from theharbor of Naples, eggs are laid in June and fe-males die about a month after the deposition. To-tal body weight and fresh gonad weight weremeasured in all specimens. The ratio between theovary weight and the body weight is referred toas gonadal somatic index (GSI). After capture, fe-males were maintained in aquarium tanks witha recirculating seawater system as described byLickey et al. (’70) and Moliner and Cakaza (’84)and under natural photoperiod (8:16, L:D) for amaximum of 5 days. Water temperature was setat a value similar to the one of the water in theharbor of Naples (13–15°C during winter months
and 19–20°C during summer months). Animalswere anesthetized on ice. Soon after the sacrificethe ovary, the proximal and distal regions of theoviducts, and oviducal glands were extracted andprocessed separately. Hemolymph was drawn bya 1-mm capillary inserted into the dorsal aortaand centrifuged at 800g for 15 min at 4°C. Thepellet was discharged, and the supernatant wascentrifuged at 105,000g for 4 hr at 4°C to elimi-nate the hemocyanin. At the end of centrifuga-tion the supernatant appeared to be composed ofthree layers of different color. Only the upperlayer, which was transparent, was collected andstored at –70°C until use.
Tissue preparation for histologyThe ovaries, the proximal and the distal portion
of the oviducts, and the oviducal glands were fixedin 4% paraformaldehyde in sea water, dehydrated,embedded in Histowax (Reichert-Jung, GmbH, Ger-many) and sectioned at 7 µm, mounted on coatedslides, and stained with hematoxylin/eosin.
From females caught in October and in July,a small piece of the ovary was collected andsoaked in 30% sucrose in sea water at 4°C, fro-zen, and sectioned by cryostat in slices of 10–15µm. Slices were coated on slides pre-treated withBio-Bond (Biocell) and dried for 5 min at roomtemperature.
Enzymatic histochemistry of 3b-hydroxysteroid dehydrogenase
Slices of ovary were washed in 0.1 M cacody-late buffer, pH 7.4, for 10 min at 4°C and then for5 min at room temperature. Slices were incubatedat 37°C for 1–3 hr in a reaction mixture consist-ing of 0.1 M cacodylate buffer, pH 7.4, containing0.2 µg/ml of steroid DHA, dissolved in diethylether, 0.5 mM βNAD, and 0.15 mM nitro-BT tet-razolium salt. Sections were then washed in wash-ing buffer, fixed for 10 min in formaldehydevapors, and mounted in glycerin. Controls wereroutinely run in all experiments omitting DHA,βNAD, or both.
Tissue extraction for radioimmunoassayOvaries were weighed and extracted with a mix-
ture of methanol and distilled water (70–30%). Tis-sues were homogenized with a glass potter andincubated at 75°C overnight. The next day sampleswere centrifuged at 2,000g for 30 min at room tem-perature, and supernatants collected and storedat 4°C. The radioimmunoassay was performedwithin 24 hr.

SEX STEROID LEVELS AND SEXUAL CYCLE IN O. VULGARIS 35
RadioimmunoassaySteroid (17β-estradiol and progesterone) concen-
trations in hemolymph and ovary extracts weremeasured by radioimmunoassay. The followingsensitivities were registered: 17β-estradiol, 5 pg(intrassay variability, 9%; interassay variability,13%); progesterone, 7 pg (intrassay variability, 8%;interassay variability, 10%).
The antisera were provided by G. Bolelli (Phys-iopathology of Reproduction Service, University ofBologna, Italy). Tritium-labeled steroids were pur-chased from Amersham International (Bucking-ham, England), and unlabeled steroids wereobtained from Sigma (St. Louis, MO).
STATISTICSAll data are expressed as mean ± SD. Data were
statistically analyzed by one-way ANOVA followedby the Duncan’s test.
RESULTS17b-Estradiol and progesterone in the
ovary and hemolymphThe concentrations of 17β-estradiol and proges-
terone in the ovary fluctuated throughout the year
and showed parallel profiles (Fig. 1). 17β-Estra-diol levels were always lower than those of proges-terone. Both steroids showed the lowest levelsfrom October to April and reached a peak in May(P < 0.01), which was followed by a decrease inJune. Neither 17β-estradiol nor progesterone wasdetected in the hemolymph throughout the repro-ductive cycle.
General morphology of thereproductive system
Female O. vulgaris have a single ovary with twooviducts opening on either side of the midlineabout halfway along the mantle cavity. The ovi-ducal gland lies about halfway along the oviductwhich is divided into two regions: the proximaloviduct, stretching from the ovary to the oviducalgland and the distal region, from the oviducalgland to the opening into the mantle cavity (Wells,’60; Wells and Wells, ’75; Di Cosmo et al., ’98).
Annual variations of the ovaryIn the months of October, November, and De-
cember the ovary contained small oocytes with adiameter between 80 and 110 µm developing fromthe germinal epithelium which was much en-
specimens analyzed was as follows: October = 4; November =4; December = 3; January = 3; February = 3; March = 3;April = 3; May = 8; June = 5; July = 3. Asterisks over barsindicate statistically significant values.
Fig. 1. 17β-Estradiol (9) and progesterone (9) levels inthe ovary and ovary weight (—) of O. vulgaris throughoutthe annual cycle. Values are reported as mean ± SD. Theovary weight values are reported as mean. The number of

36 A. DI COSMO ET AL.
folded. The oocytes remained attached by stalksto the underlying connective tissue, hanging inbunches from the anterior wall of the ovisac (Fig.2A). Oocytes were either spherical or ovoidal, andthe nucleus was spherical and centrally placedinto the dense, finely granular ooplasm and con-tained an eccentric nucleolus. Rarely was theremore than one nucleolus. Oocytes were sur-rounded by a double layer of follicle cells: the in-ner layer was cuboidal and secretory, the outerlayer was thin and flattened epithelium (the oo-cyte and the associated follicle cells are definedas follicle) (Fig. 2B). In the months of Januaryand February, the nucleus in the oocyte wasspherical and centrally placed, with a scarcely vis-ible nucleolus. Oocytes were surrounded by cuboi-
dal follicle cells. However, in this period the fol-licle cells start enfolding into the ooplasm. At thisstage 70% of oocytes did not show any fold, 3%showed three folds, 20% showed four folds, and4% and 3% were respectively characterized by fiveand six folds. Oocytes with folds showed a diam-eter of between 110 and 200 µm. The ooplasm wascompact in those oocytes without folds and withthree folds and was vacuolated in the oocytes withfour, five, and six folds. An extensive blood capil-lary system enveloped the follicle (Fig. 3A,B). Inthe months of March and April oocytes showed adiameter between 200 and 400 µm. Within theoocyte the nucleus was long and narrow. The fol-licle cells were columnar, with prominent and elon-gated nuclei. It is likely that in this period a phase
Fig. 2. Transverse section of the ovary of the month ofNovember (October and December gave similar results). A:ovary contains small oocytes developing from the germinalepithelium (original magnification ×100). B: transverse sec-tion of follicles at a higher magnification, showing the layerof follicle cells surrounding the oocytes (the oocyte and theassociated follicle cells are defined as follicle) (arrows); ge,germinal epithelium (original magnification ×280).
Fig. 3. Transverse section of the ovary of the month of Janu-ary (February gave similar results), A: showing oocytes sur-rounded by follicle cells starting to enfold: f0 = follicles withoutfolds; f3 = follicles with three folds; f4 = follicles with four folds(original magnification ×150). B: transverse section at a highermagnification of a follicle with four folds and one fold startingto enfold (original magnification ×320). Blood capillaries run-ning at the base of the folds are visible (arrow).

SEX STEROID LEVELS AND SEXUAL CYCLE IN O. VULGARIS 37
of synthesis probably of yolk proteins begins, withan increase of the basophilia of the nucleus withinthe follicle cells (Fig. 4). At this stage 35% of oo-cytes were without folds, 50% showed four folds,and 15% of oocytes showed five folds or six folds.In the month of May oocytes showed an averagediameter between 400 and 600 µm. Within theoocyte the nucleus was long and narrow. The fol-licle cells were columnar. At this stage folds werenarrower and extended less deeply into the oo-cyte with respect to the previous period (Fig. 5);20% of the oocytes did not show any fold, 70%showed four folds, and 10% had five and six folds.The ooplasm was strongly vacuolated in those oo-cytes with folds. In the month of June the oocyteswith four folds underwent a profound transforma-tion with a total regression of the folds, and thewhole ooplasm was filled with yolk (Fig. 6A). Atthis stage the diameter of the oocytes was between600 and 1000 µm. Oocytes with five and six foldsshowed strongly vacuolated ooplasm (Fig. 6B).
Annual variations of the oviductsIn the months of October, November, and De-
cember, the proximal and distal regions of the ovi-ducts were characterized by thin and thick walls,respectively, and a single cuboidal epithelium. Themucosa was arranged in irregular folds, whichwere finger-like with an irregular shape in theproximal region and extended deeply into the lu-men, whereas in the distal region folds wereshorter and protruded less deeply into the lumen(Fig. 7A,B). The epithelial cells of both regionswere ciliated and had basophilic, small, and round,
Fig. 4. Transverse section of the ovary of the month ofMarch (April gave similar results), showing follicles with fourfolds and vacuolated ooplasm filled with yolk; y, yolk; (origi-nal magnification ×170).
Fig. 5. Transverse section of the ovary of the month ofMay, showing follicles with four folds narrower and less deepthan during the previous period; y, yolk (original magnifica-tion ×200).
Fig. 6. Transverse section of a follicle with four folds ofthe month of June, showing in A: detail of a regressed fold(arrows) and ooplasm filled with yolk (original magnification×400). B: follicle with five folds and one fold starting to en-fold and strongly vacuolated ooplasm; y, yolk (original mag-nification ×150).
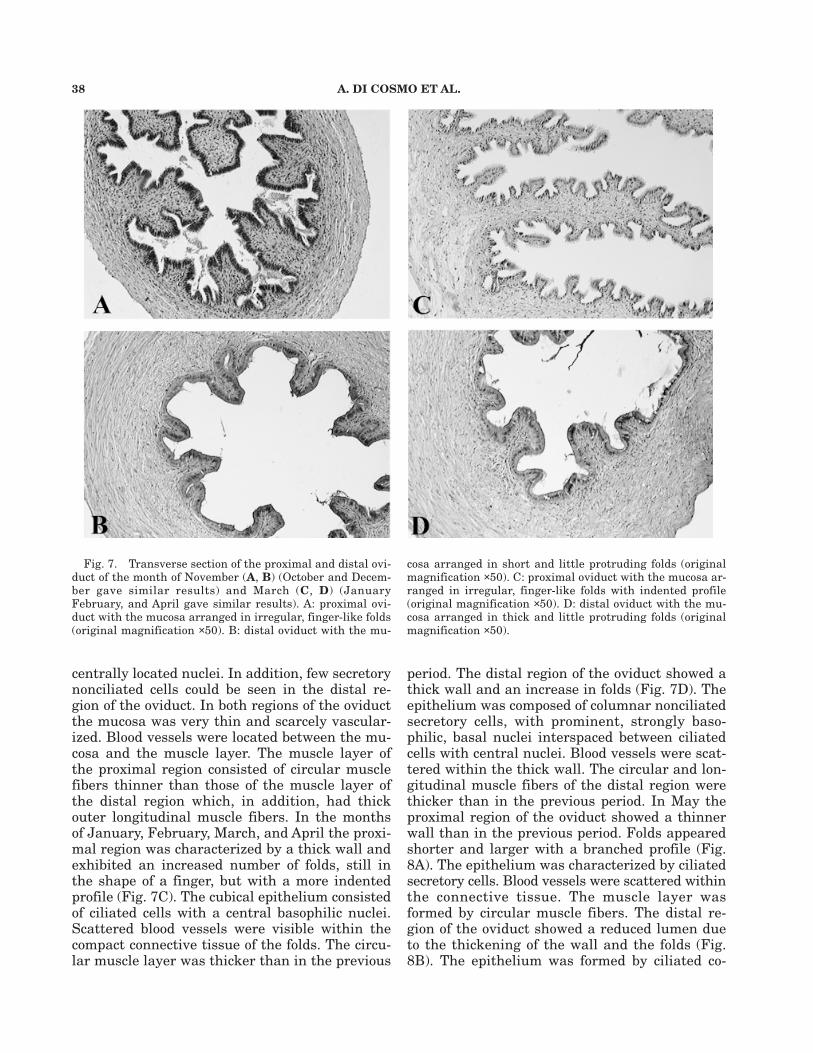
38 A. DI COSMO ET AL.
centrally located nuclei. In addition, few secretorynonciliated cells could be seen in the distal re-gion of the oviduct. In both regions of the oviductthe mucosa was very thin and scarcely vascular-ized. Blood vessels were located between the mu-cosa and the muscle layer. The muscle layer ofthe proximal region consisted of circular musclefibers thinner than those of the muscle layer ofthe distal region which, in addition, had thickouter longitudinal muscle fibers. In the monthsof January, February, March, and April the proxi-mal region was characterized by a thick wall andexhibited an increased number of folds, still inthe shape of a finger, but with a more indentedprofile (Fig. 7C). The cubical epithelium consistedof ciliated cells with a central basophilic nuclei.Scattered blood vessels were visible within thecompact connective tissue of the folds. The circu-lar muscle layer was thicker than in the previous
period. The distal region of the oviduct showed athick wall and an increase in folds (Fig. 7D). Theepithelium was composed of columnar nonciliatedsecretory cells, with prominent, strongly baso-philic, basal nuclei interspaced between ciliatedcells with central nuclei. Blood vessels were scat-tered within the thick wall. The circular and lon-gitudinal muscle fibers of the distal region werethicker than in the previous period. In May theproximal region of the oviduct showed a thinnerwall than in the previous period. Folds appearedshorter and larger with a branched profile (Fig.8A). The epithelium was characterized by ciliatedsecretory cells. Blood vessels were scattered withinthe connective tissue. The muscle layer wasformed by circular muscle fibers. The distal re-gion of the oviduct showed a reduced lumen dueto the thickening of the wall and the folds (Fig.8B). The epithelium was formed by ciliated co-
Fig. 7. Transverse section of the proximal and distal ovi-duct of the month of November (A, B) (October and Decem-ber gave similar results) and March (C, D) (JanuaryFebruary, and April gave similar results). A: proximal ovi-duct with the mucosa arranged in irregular, finger-like folds(original magnification ×50). B: distal oviduct with the mu-
cosa arranged in short and little protruding folds (originalmagnification ×50). C: proximal oviduct with the mucosa ar-ranged in irregular, finger-like folds with indented profile(original magnification ×50). D: distal oviduct with the mu-cosa arranged in thick and little protruding folds (originalmagnification ×50).

SEX STEROID LEVELS AND SEXUAL CYCLE IN O. VULGARIS 39
lumnar cells with basal nuclei. The muscle layerwas thicker than in the previous period and wascharacterized by inner circular fibers and outer lon-gitudinal fibers. In June the proximal region ofthe oviduct showed two different types of folds.Some were enlarged and dramatically irregular inshape, interspaced by little finger-like folds, andboth were surrounded by fluid mucous material(Fig. 8C). The epithelium was formed by ciliatedand nonciliated secretory cells. The muscle layerwas formed by circular muscle fibers, and blood ves-sels were dispersed within the connective tissue.The distal region exhibited maximal development,showing a thick wall and long folds, protrudingwithin the lumen, which appeared to be filled withdense mucous material (Fig. 8D). The epitheliumwas formed by empty secretory nonciliated cells.The morphological variations of the oviducts areschematically reported in Fig. 9.
Annual variations of the oviducal glandsThe oviducal glands are arranged halfway along
the oviducts. The analysis of the serial transversesections allows us to divide the oviducal glandsinto two regions: the former stretched from theproximal part of the oviduct to the central cavityconstituted by an enlargement of the lumen of theoviduct and extended for about two-thirds of thegland. It contained the spermathecae embeddedin compact connective tissue crossed by muscularfibers that were actually the wall of the oviductitself and that were surrounded by a glandularcompartment divided into an inner and an outerregion, with respect to the lumen of the oviduct.The latter region stretched from the central cav-ity to the distal part of the oviduct and extendedfor about one-third of the gland. Within this re-gion, the spermathecae were lacking and the glan-dular components converged and opened with
Fig. 8. Transverse section of the proximal and distal ovi-duct of the months of May (A, B) and June (C, D). A: proxi-mal oviduct with the mucosa arranged in irregular, finger-likefolds with short (small arrow), large (medium arrow), andbranched profile (large arrow) (original magnification ×50).B: distal oviduct with the mucosa arranged in thick and pro-truding folds (original magnification ×50). C: proximal ovi-
duct with the mucosa arranged in two different kinds of folds,one large and irregular in shape and the other little and fin-ger-like profile (arrows) (original magnification ×50). D: dis-tal oviduct with the mucosa arranged in maximally enlargedfolds, protruding into a lumen filled with mucous material(original magnification ×50); mm, mucous material (originalmagnification ×120).

40 A. DI COSMO ET AL.
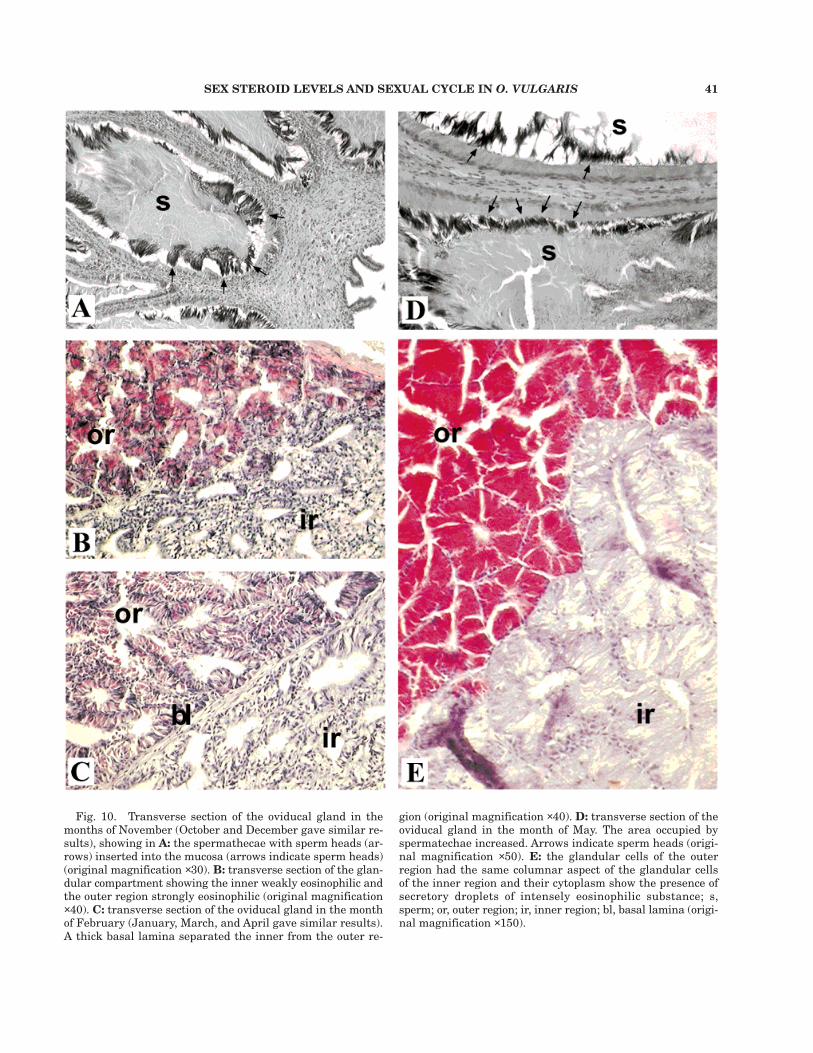
ducts into the lumen of the central cavity. In themonths of October, November, and December thespermathecae showed simple ciliated cubical epi-thelium with basophilic elongated basal nuclei.The cytoplasm was weakly eosinophilic. Abundantsperm were crypted and immobilized in the sper-mathecae epithelium, with the heads insertedwithin the mucosa, while the tails were not clearlyvisible due to their being crowded together witha dense and weakly eosinophilic material, ob-served in the central part of the lumen of the sper-mathecae (Fig. 10A). The glandular compartmentlooked like a composite tubular gland with a com-pact texture and narrow lumens. The two regions(inner and outer regions) were not separated by a
basal lamina. The cytoplasm of the cells of theinner region was weakly eosinophilic, rich in vacu-oles, and showed elongated basophilic and basalnuclei. In the strongly eosinophilic cytoplasm ofthe cells of the outer region, vacuoles were rarelyseen. Basophilic, elongated nuclei were basallyplaced (Fig. 10B). In the months of January, Feb-ruary, March, and April no changes could be seenin the morphology of spermathecae or in the dis-tribution of the spermatozoa. On the other hand,the glandular component appeared less compactthan in the previous period, with wide lumensamong tubular components. A thick basal laminaseparated the inner from the outer region. Theeosinophilic cytoplasm of the cells of the inner re-
Fig. 9. Schematic drawing of the morphological variationsof the proximal and distal oviduct and the oviducal gland,from October to July. On the left is represented the oviductand the oviducal gland as they lay within the body cavity.On the right, transverse sections of the proximal and distaloviduct and the oviducal gland are represented, showing their
morphological changes. Drawings of the transverse sectionswere obtained by camera lucida at the level indicated by thelines on the oviduct and the oviducal gland on the left; sp,spermathecae; ir, inner region; or, outer region; l, lumen; w,oviduct wall. Drawings are to scale.
SEX STEROID LEVELS AND SEXUAL CYCLE IN O. VULGARIS 41
Fig. 10. Transverse section of the oviducal gland in themonths of November (October and December gave similar re-sults), showing in A: the spermathecae with sperm heads (ar-rows) inserted into the mucosa (arrows indicate sperm heads)(original magnification ×30). B: transverse section of the glan-dular compartment showing the inner weakly eosinophilic andthe outer region strongly eosinophilic (original magnification×40). C: transverse section of the oviducal gland in the monthof February (January, March, and April gave similar results).A thick basal lamina separated the inner from the outer re-
gion (original magnification ×40). D: transverse section of theoviducal gland in the month of May. The area occupied byspermatechae increased. Arrows indicate sperm heads (origi-nal magnification ×50). E: the glandular cells of the outerregion had the same columnar aspect of the glandular cellsof the inner region and their cytoplasm show the presence ofsecretory droplets of intensely eosinophilic substance; s,sperm; or, outer region; ir, inner region; bl, basal lamina (origi-nal magnification ×150).
42 A. DI COSMO ET AL.
gion showed less elongated basophilic nuclei andmore vacuoles than in the previous period. How-ever, the cytoplasm of the outer region with scarcevacuoles appeared less strongly eosinophilic thanin the previous period and with basophilic, elon-gated nuclei (Fig. 10C). In May, the area occupiedby spermathecae increased and the epitheliumappeared strongly eosinophilic with basal andsmall spherical basophilic nuclei. Spermatozoawere arranged in the same position with respectto the previous period, while the material in thelumen of the spermathecae appeared strongly eosi-nophilic (Fig. 10D). The glandular compartmentalso increased in size. The inner region epithe-lium, weakly eosinophilic, became columnar withsmall, basophilic nuclei, basally placed. The epi-thelial cells showed fewer vacuoles than in theprevious period and were filled with a transpar-ent substance. The glandular cells of the outerregion had the same columnar aspect of the in-ner region cells and their cytoplasm, which wasnot vacuolated, showed the presence of secretorydroplets of intensely eosinophilic substance. Thesmall, basophilic nuclei were basally arranged(Fig. 10E). In June, no changes in the morphol-
ogy of either spermathecae or the glandular com-partment could be seen. The whole oviducal gland,however, appeared to be larger. The morphologi-cal variations of the oviducal gland are schemati-cally reported in Fig. 9.
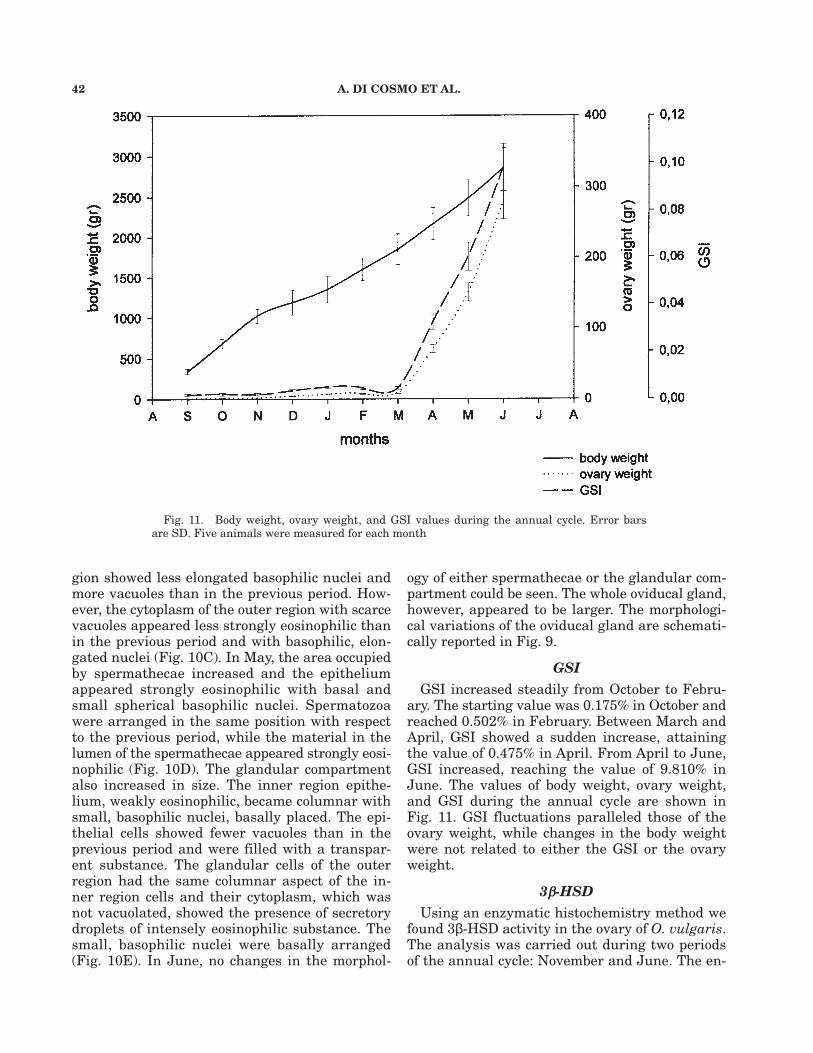
GSIGSI increased steadily from October to Febru-
ary. The starting value was 0.175% in October andreached 0.502% in February. Between March andApril, GSI showed a sudden increase, attainingthe value of 0.475% in April. From April to June,GSI increased, reaching the value of 9.810% inJune. The values of body weight, ovary weight,and GSI during the annual cycle are shown inFig. 11. GSI fluctuations paralleled those of theovary weight, while changes in the body weightwere not related to either the GSI or the ovaryweight.
3b-HSDUsing an enzymatic histochemistry method we
found 3β-HSD activity in the ovary of O. vulgaris.The analysis was carried out during two periodsof the annual cycle: November and June. The en-
Fig. 11. Body weight, ovary weight, and GSI values during the annual cycle. Error barsare SD. Five animals were measured for each month

SEX STEROID LEVELS AND SEXUAL CYCLE IN O. VULGARIS 43
Fig. 12. Histochemistry of 3β-hydroxysteroid dehydroge-nase. A: cryostat transverse slice of the ovary in the monthsof November (original magnification ×120) and B: showingthe enzymatic activity confined in the follicular cells surround-ing and enfolding the oocytes (arrows) in the month of June(original magnification ×400). C: control experiment cryostattransverse slice of the ovary in the month of November show-ing nonspecific products of reaction (large arrows) due to theprecipitation of NBT salts (see Materials and Methods) (origi-nal magnification ×150).
zymatic activity was localized, in both periods, inthe follicle cells (Fig. 12A,B). Control experimentswere performed in both periods. Nonspecific reac-tion products were observed in the shape of scat-tered granules of variable size and intensity ofcolor, due to the precipitation of NBT salts dur-ing the incubation time (Fig. 12C).
DISCUSSIONIn this paper we present data on 17β-estradiol
and progesterone levels, and morphological changesof the reproductive system (ovary, oviducts, andoviducal glands) of the female of O. vulgaris,throughout the reproductive cycle. The reproduc-tive system of the female of O. vulgaris increasedin size and weight regularly from October to June,and the increase does not seem to be correlatedto the weight of the animal, as indicated by theGSI values. Our data are in agreement with theobservations carried out by Buckley (’76) on O. vul-garis. As a common feature in cephalopods, folliclecells proliferate during the annual cycle, resultingin deep enfolding into the oocyte (Boucaud-Camouet al., ’91; Ikeda et al., ’94). We observed that in O.vulgaris, from October to June, oocytes increasedin both diameter and in number of folds. Althoughthe maximum number of folds present in a singleoocyte was six, the highest percentage of oocyteshad four folds. Moreover, oocytes with four foldswere the only ones that reached a maturationalstage such that the whole ooplasm was occupiedby yolk, with a contemporary regression of theheight and width of the enfolding. On the con-trary, oocytes with five and six folds never attainedsuch a maturational stage, but always showed deepfolds and scarce yolk, showing the typical aspectof atresia. This suggests that a morphological fea-ture is indicative of an achieved functional matu-rity. On the grounds of such observations, wepropose to divide the ovarian cycle of O. vulgarisinto four main phases: (i) previtellogenesis, whenthe majority of oocytes does not show any fold,and the ooplasm is compact; (ii) early vitellogen-esis, when the follicle cells start enfolding in theooplasm of the oocyte and yolk granule deposi-tion within the ooplasm begins: oocytes withoutfolds are still present in the ovary; however, themajority of oocytes that undergo the vitellogen-esis show four folds; (iii) full vitellogenesis, whenthe majority of oocytes has four folds and the oop-lasm is almost completely filled with yolk gran-ules; and (iv) late vitellogenesis, when the diameterof the oocyte is unchanged with respect to the pre-

44 A. DI COSMO ET AL.
been postulated and sustained by experimentalobservations (Perez and Callard, ’89). Among in-vertebrates 17β-estradiol and progesterone seemto play less clearly differentiated roles in repro-duction. It seems that 17β-estradiol is involved invitellogenesis in crustaceans (Couch et al., ’87; VanBeek and De Loof, ’88; Quinitio et al., ’91), and astimulatory effect of 17α-hydroxyprogesterone onvitellogenin secretion has been demonstrated inthe shrimp Penaeus japonicus (Yano, ’87). Proges-terone stimulates ovarian development in theprawn Parapenaeopsis hardwickii (Kulkarni et al.,’79) and the shrimp Metapenaeus ensis (Yano, ’85).In the snail Helix pomatia, progesterone andoestone-acetate stimulate ovogenesis (Csaba andBierbauer, ’79). Differently from crustaceans andinsects, where vitellogenin is synthesized into thefat bodies and transported through the hemo-lymph to the ovary (Engelman, ’94), in Octopus ithas been clearly established that the follicle cellssynthesize and transfer yolk proteins into the oop-
Fig. 13. Schematic drawing of the morphological changesin follicles of O. vulgaris. Follicles of the previtellogenic phase(PV; A, B); follicles of the early vitellogenic phase (EV; B, C);
follicles of the full vitellogenic phase (FV; D, E); follicles oflate vitellogenic phase (LV; E, F).
vious period, but folds are now dramatically re-gressed and the ooplasm is completely filled withyolk (Fig. 13).
In the ovary of O. vulgaris 17β-estradiol andprogesterone concentrations changed in relationto the ovarian development during the reproduc-tive cycle. Both steroids were low during theprevitellogenesis and increase at the onset of vi-tellogenesis, reaching a peak during the full vi-tellogenesis. Their increase prior to vitellogenesisseems to suggest their involvement in yolk syn-thesis, an aspect also sustained by their decreasewhen the vitellogenesis has come to an end. Invertebrates, the main steroid involved in yolk syn-thesis is the 17β-estradiol (Wallace, ’85), althoughother factors have been proved to play an activerole (Ho et al., ’81, ’85). Progesterone is regardedas the hormone of viviparity, due to its action inpreparing the genital tract to receive and shelterthe embryo (Callard et al., ’92). Recently, a novelrole in suppressing the vitellogenin synthesis has

SEX STEROID LEVELS AND SEXUAL CYCLE IN O. VULGARIS 45
lasm (O’Dor and Wells, ’73, ’75). In O. vulgaristhe factor(s) controlling the onset of vitellogenesisappear to be associated with increased secretionof a yet unidentified gonadotropic hormone fromthe optic gland (Wells and Wells, ’59, ’72, ’75).However, we believe that a local effect, exertedon follicle cells, in the modulation of vitellogen-esis might be carried out by either 17β-estradiolor progesterone or both, as suggested by the pres-ence of such steroids within the ovary when thevitellogenesis is active. 17β-Estradiol and proges-terone may act through a specific receptor, whosepresence within the follicle cells has been alreadyreported in this species (Di Cosmo et al., ’98a,b).Both an autocrine and paracrine role is sustain-able in light of 17β-estradiol and progesterone ab-sence from the hemolymph. It is important to notethat gonads are not only a target organ but alsoa source of steroids, as proved by the presence of3β-HSD in the ovary (present data) and testis ofO. vulgaris (D’Aniello et al., ’96).
Morphological changes in the oviduct and ovi-ducal gland observed throughout the reproductivecycle in O. vulgaris, along with 17β-estradiol andprogesterone fluctuations, suggest that both ste-roids might work in synergy to sustain the growthand differentiation of the female reproductivesystem in this species. In oviparous vertebrates17β-estradiol promotes the proliferation and dif-ferentiation of the epithelial cells, while progest-erone is active in albumen secretion and shellproduction (Ratia, ’79; Joensuu, ’90; Joensuu et al.,’92). In O. vulgaris, during the egg-laying period,the oviducal glands produce two different sub-stances which, when combined together, surroundthe eggs in a gelatinous covering. The polymer-ization of such gelatinous material is preventedby a mucous substance secreted by the proximaltract of the oviduct that, in this phase of the re-productive period, increases both the number andthe surface area of epithelial folds (Wells, ’78). Aswe report here, both the oviducts and the ovidu-cal glands undergo profound changes that may berelated to the functions of this structure. Indeed,the oviduct increases in number and surface offolds and secretes substances which are likely tobe involved in facilitating eggs’ descent. We havealready demonstrated that the proximal and thedistal tracts of the oviduct are target organs of17β-estradiol and progesterone, as demonstratedby the presence of specific receptors (Di Cosmo etal., ’98a,b). Progesterone might be responsible forthe production of lubricating material and formuscle contractions as sustained by the observa-
tion that the oviduct in this species is richly in-nervated (Young, ’67). Froesch and Marty (’75)suggest that in octopuses, when the eggs reachthe central cavity of the oviducal glands, they areretained and wait for sperm to be mobilized andreleased from spermathecae. The morphologicalchanges of the oviducal glands in O. vulgaris arein agreement with their role in sperm mobiliza-tion. We may hypothesize that the oviducal glandshave complex functions, as suggested by the twoglandular compartments which are characterizedby different staining properties, in agreement withthe supposed twofold role played by these glandsin secreting substances which harden into an egg-shell and store the sperm (Wells, ’78). We observedthat the difference between the two compartmentsbecomes evident during the full vitellogenesis,when sperm begins to mobilize, sustaining thatthe central cavity of the glands may be the site offertilization (Froesch and Marthy, ’75). Moreover,we cannot exclude that this phenomenon may behormonally controlled, as suggested by the pres-ence of progesterone receptors in the glandularregion (Di Cosmo et al., ’98b).
In conclusion, in this study we report that 17β-estradiol and progesterone fluctuations paralleledthe morphological changes of the reproductive sys-tem of the female of O. vulgaris during the repro-ductive cycle, suggesting that, in this species, thereproductive system is under hormonal control.
LITERATURE CITEDArkhipkin AI. 1992. Reproductive system structure, develop-
ment, and function in cephalopods with a new general scalefor maturity stages. J Northwest Fish Sci 12:63–74.
Arkhipkin AI, Nigmatullin CM, Sabirov RM, Shilin KD. 1983.Morphology and characteristic features of sexual systemfunctioning in Thysanotheuthis rhombus squid. In: System-atics and ecology of cephalopod molluscs. Leningrad: NaukaPress. p 59–61.
Boucaud-Camou E, Kueta N, Boismery J, Medhioub A. 1991.The sexual cycle of Sepia officinalis L. from the bay of Seine.In: Boucaud-Camou E, editor. La seiche, 1st InternationalSymposium on the Cuttlefish Sepia. Caen: Centre de Pub-lications de l’Université de Caen. p 141–151.
Boyle PR, Chevis D. 1991. Changes in follicle cells epithe-lium nuclei at the onset of vitellogenesis in the octopusEledone cirrhosa. Bull Mar Sci 49:373–378.
Boyle PR, Chevis D. 1992. Egg development in the octopusEledone cirrhosa. J Zool (London) 227:623–638.
Callard IP, Fileti LA, Perez LE, Sorbera LA, Giannoukos G,Klosterman LL, Tsang P, McCracken J. 1992. Role of thecorpus luteum and progesterone in the evolution of verte-brate viviparity. Am Zool 32:264–275.
Cottrel GA, Schot LPC, Dockray GJ. 1983. Identification andprobable role of a single neurone containing the neuropep-tide Helix FMRF-amide. Nature 304:638–640.
Cottrel GA, Davies NW, Green KA. 1984. Multiple action of

46 A. DI COSMO ET AL.
a molluscan cardioexcitatory neuropeptide and related pep-tides on identified Helix neurones. J Physiol (London)356:315–333.
Couch EF, Hagino N, Lee JW. 1987. Changes in estradiol andprogesterone immunoreactivity in tissue of the lobster,Homarus americanus, with developing and immature ova-ries. Comp Biochem Physiol A 87:765–770.
Csaba G, Bierbauer J. 1979. Effect of oestrogenic, and andro-genic and gestagenic hormones on the gametogenesis (oo-genesis and spermatogenesis) in the snail Helix pomatia.Acta Biol Med Ger 38(8):1145–1148.
D’Aniello A, Di Cosmo A, Di Cristo C, Fisher G. 1995. D-As-partate in the male and female reproductive system of Oc-topus vulgaris Lam. Gen Comp Endocrinol 100:69–72.
D’Aniello A, Di Cosmo A, Di Cristo C, Assisi L, Botte V, DiFiore MM. 1996. Occurrence of sex steroid hormones andtheir binding proteins in Octopus vulgaris Lam. BiophysBiochem Res Commun 227:782–788.
Di Cosmo A, Di Cristo C. 1996. Endocrine control of the re-production of Octopus vulgaris. Proceedings of the 57th Na-tional Meeting of the Italian Zoological Union. p 104.
Di Cosmo A, Di Cristo C. 1998. Neuropeptidergic control ofthe optic gland of Octopus vulgaris: FMRF-amide and GnRHimmunoreactivity. J Comp Neurol 398:1–12.
Di Cosmo A, Paolucci M, Di Cristo C, Ciarcia G. 1998a.Molecole leganti l’estradiolo nel sistema riproduttivo dellafemmina di Octopus vulgaris. Proceedings of the 59th Na-tional Meeting of the Italian Zoological Union. p 76.
Di Cosmo A, Paolucci M, Di Cristo C, Botte V, Ciarcia G.1998b. Progesterone receptor in the reproductive system ofthe female of Octopus vulgaris: characterization andimmunolocalization. Mol Reprod Dev 50:451–460.
Engelman F. 1994. Invertebrates: hormone-regulated gonadalactivity. In: Epple A, Scanes CG, Stentson MH, editors.Perspectives in comparative endocrinology. Ottawa, Canada:Academic Press. p 36–40
Ezzeddine-Najai S. 1997. Maturation sexuelle d’Eledonemoschata (Cephalopoda, Octopoda) du golfe de Gabès. VieMilieu 47:69–76
Fields WG. 1965. The structure, development, food relations,reproduction, and life history of the squid Loligo opalescensBerry. Fish Bull Calif Dept Fish Game 131:1–108
Froesch D, Marthy HJ. 1975 The structure and function ofthe oviducal gland in octopods (Cephalopoda). Proc R SocLondon B 188:95–101.
Guerra A. 1975. Determination of the different stages of thesexual development of the common octopus by a maturityindex. Invest Pesq 39:397–416.
Ho SM, Danko D, Callard IP. 1981. Effect of exogenous es-tradiol-17β on plasma vitellogenin levels in male and fe-male Chrysemys and its modulation by testosterone andprogesterone. Gen Comp Endocrinol 43:413–421.
Ho SM, Wangh LJ, Callard IP. 1985. Sexual differences inthe in vitro induction of vitellogenesis in the turtle: role ofthe pituitary and growth hormone. Comp Biochem Physiol81B:467–472.
Ikeda Y, Sakurai Y, Shimazaki K. 1993. Fertilizing capacityof squid (Todarodes pacificus) spermatozoa collected fromvarious sperm storage sites, with special reference to therole of gelatinous substance from oviducal gland in fertili-zation and embryonic development. Invert Reprod Dev23:39–44.
Joensuu TK. 1990. Chick oviduct differentiation. The effectof estrogen and progesterone on the expression of progest-erone receptor. Cell Differ Dev 30:207–218
Joensuu TK, Niemela A, Kunnas T, Salomaa S, Alho H, ViljaP, Ylikomi T, Kulomaa M, Tuohimaa P. 1992. Inducibility ofthe avidin gene by progesterone is suppressed during es-trogen-induced cytodifferentiation. J Steroid Biochem MolBiol 43:609–618.
King JA, Millar RP. 1995. Evolutionary aspects of gonadotro-pin-releasing hormone and its receptor. Cell Mol Neurobiol15:5–23.
Kueta N, Boucaud-Camou E, Renou AM. 1993. Gonadotropicmitogenic activity of the optic gland of the cuttlefish, Sepiaofficinalis, during sexual maturation. J Mar Biol Assoc UK75:391–404.
Kulkarni GK, Nagabhushanam R, Joshi P. 1979. Effect ofprogesterone on ovarian maturation in a marine penaeidprawn Parapenaeopsis hardwickii. Indian J Exp Biol17:986–987.
Lickey ME, Emigh RL, Randle FR. 1970. A recirculating seawater aquarium system for inland laboratories. Mar Biol7:149–152.
Mangold K. 1989. Reproduction. In: Boyle PR, editor. Cepha-lopod life cycles II. Orlando, FL: Academic Press. 441 p.
Mann T, Martin AW, Thiersch JB. 1970. Male reproductivetract, spermatophores, and spermatophoric reaction in thegiant octopus of the North Pacific, Octopus dofleini mar-tini. Proc R Soc London B 175:31–61.
Moliner A, Cakaza N. 1984. Tecnica para el mantenimientode Octopus vulgaris en acuarios marinos con circulacioncerrada de agua. Inf Tec Inst Esp Oceanogr 28:1–11
O’Dor RK, Wells MJ. 1973. Yolk protein synthesis in the ovaryof Octopus vulgaris and its control by the optic gland gona-dotropin. J Exp Biol 59:665–674.
O’Dor RK, Wells MJ. 1975. Control of yolk protein synthesisby Octopus gonadotropin in vivo and in vitro (effects of Oc-topus gonadotropin). Gen Comp Endocrinol 27:129–135.
Olivares AP, Zuniga OR, Retamales EN. 1994. Sexual matu-rity scale for Octopus from II region, Chile. Estud Oceanol13:89–91.
Perez LE, Callard IP. 1989. Evidence for progesterone inhibi-tion of vitellogenesis in the skate (Raja erinacea). Am Zool29(4):78A.
Peterson RP. 1959. The anatomy and histology of the reproduc-tive system of Octopus bimaculoides. J Morphol 104:61–87.
Quinitio ET, Yamauchi K, Hara A, Fuji A. 1991. Profiles ofprogesterone- and estradiol-like substances in the hemo-lymph of female Pandalus kessleri during an annual repro-ductive cycle. Gen Comp Endocrinol 81:343–348.
Ratia J, Elo H, Tuohimaa P. 1979. Differentiation of imma-ture chick oviduct by progesterone and diethylstilbestrol.In: Vermeulen A, Klopper A, Sciarra F, Jungblut P, LernerL, editors. Research on steroids. Amsterdam: North-HollandPublishing Company. p 371–379.
Sauer WH, Roberts MJ, Lipinski MR, Smale MJ, Hanlon RT,Webber DM, O’Dor RK. 1997. Choreography of the squid’s“nuptial dance.” Biol Bull 192(2):203–207.
Smale MJ, Buchan PR. 1981. Biology of Octopus vulgaris ofthe East Coast of South Africa. Mar Biol 65:1–12.
Tompsett DH. 1939. In: Daniel RJ, editor. Sepia. Liverpool,England: The University Press of Liverpool.
Van Beek E, De Loof, A 1988. Radioimmunological determina-tions of the concentrations of six C21, C19, and C18 steroidsduring the reproductive cycle of female Artemia sp. (Crusta-cea: Anostaraca). Comp Biochem Physiol A 89:595–599.
von Boletsky S. 1981. Reflexions sur les strategies de re-production chez les chephalopodes. Bull Soc Zool Fr 106:293–304.

SEX STEROID LEVELS AND SEXUAL CYCLE IN O. VULGARIS 47
von Boletzky S. 1983. Sepia officinalis. In: Boyle PR, editor.Cephalopod life cycles, vol. 1. Orlando, FL: Academic Press.p 31–52.
Wallace RA. 1985. Vitellogenesis and oocyte growth: non-mam-malian vertebrates. In: Browder L, editor. Developmentalbiology: a comprehensive synthesis, vol. I, Oogenesis. NewYork: Plenum Press. p 127–177.
Wells, MJ. 1978. Octopus: physiology and behaviour of anadvanced invertebrate. London: Chapman and Hall. p1–417.
Wells MJ, Wells J. 1959. Hormonal control of sexual matu-rity in Octopus. J Exp Biol 36:1–33.
Wells MJ, Wells J. 1972. Sexual displays and mating of Octo-
pus vulgaris Cuvier and O. cyanea Gray and attempts toalter performance by manipulating the glandular conditionof the animals. Anim Behav 20:293–308.
Wells MJ, Wells J. 1975. Optic gland implants and their ef-fects on the gonads of Octopus. J Exp Biol 62:579–588.
Yano I. 1985. Induced ovarian maturation and spawning ingreasyback shrimp, Metapenaeus ensis, by progesterone.Aquaculture 47:223–229.
Yano I. 1987. Effect of 17α-hydroxyprogesterone on vitel-logenin secretion in kuruma prawn, Penaeus japonicus.Aquaculture 61:49–57.
Young JZ. 1967 The visceral nerves of Octopus. Phil Trans RSoc London B 253:1–22.




















