
Serotonin regulates an acyl-CoA-binding protein (ACBP) gene expression in the midgut of Rhodnius...
7
Serotonin regulates an acyl-CoA-binding protein (ACBP) gene expression in the midgut of Rhodnius prolixus Michele Alves-Bezerra a,1 , David Majerowicz a, 1 , Luciano A.M. Grillo a, b , Hugo Tremonte c , Caroline B. Almeida a , Glória R.C. Braz c, e , Mauro Sola-Penna d , Gabriela O. Paiva-Silva a, e , Katia C. Gondim a, e, * a Instituto de Bioquímica Médica, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil b Escola de Enfermagem e Farmácia, Universidade Federal de Alagoas, Alagoas, Brazil c Departamento de Bioquímica, Instituto de Química, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil d Departamento de Fármacos, Faculdade de Farmácia, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil e Instituto Nacional de Ciência e Tecnologia em Entomologia Molecular, Brazil article info Article history: Received 6 November 2009 Received in revised form 16 December 2009 Accepted 6 January 2010 Keywords: ACBP Lipid Acyl-CoA ester Serotonin 5-HT Rhodnius prolixus abstract Acyl-CoA esters have many intracellular functions, acting as energy source, substrate for metabolic processes and taking part in cell signaling. The acyl-CoA-binding protein (ACBP), a highly conserved 10 kDa intracellular protein, binds long- and medium-chain acyl-CoA esters with very high affinity, directing them to specific metabolic routes and protecting them from hydrolysis. An ACBP gene sequence was identified in the genome of Rhodnius prolixus. This ACBP gene (RpACBP-1) was expressed in all analyzed tissues and quantitative PCR showed that expression was highest in posterior midgut. In this tissue, ACBP gene expression increased in the first day after blood meal (w10-fold) and then decreased to unfed levels in the seventh day after meal. Injection of serotonin (5-hydroxytryptamine; 5-HT), a neu- roamine released in the hemolymph after the start of feeding, increased the expression of this gene in the midgut of unfed females, reaching levels similar to those observed in fed insects. This effect of injected 5-HT was inhibited by spiperone, an antagonist of 5-HT mammalian receptors, that was also able to block the physiological increase in RpACBP-1 expression observed after feeding. Injection of cholera toxin or dibutyryl-cAMP also resulted in the stimulation of this gene expression. These data reveal a transcriptional regulatory mechanism in R. prolixus, that is triggered by 5-HT. In this way, a novel role for 5-HT is proposed, as a regulator of ACBP gene expression and, consequently, taking part in the control of lipid metabolism. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction Although lipids are the main energy source for demanding metabolic processes, as flight and oogenesis, little is known about how these hydrophobic macronutrients and their intermediate metabolites, like fatty acyl-CoA esters, are maintained soluble in the cytosol. Lipid-binding proteins, as the fatty acid-binding protein (FABP) and the acyl-CoA-binding protein (ACBP), have a crucial role in this process. As ACBP shows an affinity for acyl-CoA that is four orders of magnitude higher than FABP, this protein is considered very important for the regulation of lipid metabolism in the cells (Faergeman and Knudsen, 1997). ACBP is a 10 kDa protein, highly conserved and present in all eukaryotes and some prokaryotes (Burton et al., 2005; Faergeman et al., 2007). This protein can bind long- and medium-chain acyl- CoA esters with high affinity and specificity (Rosendal et al., 1993), participates in the transport of acyl-CoA inside cells (Schjerling et al., 1996) and protects these lipids from hydrolysis (Schroeder et al., 2005), forming an intracellular pool of acyl-CoA. ACBP is also apparently involved in fatty acid elongation, sphingolipid synthesis, protein sorting and vesicle traffic(Gaigg et al., 2001; Faergeman et al., 2004). Furthermore, it has been shown that the presence of ACBP can regulate the activity of enzymes involved in lipid metabolism, as acetyl-CoA carboxylase and mitochondrial acyl-CoA synthetase (Rasmussen et al., 1993), glycerol-3-phosphate acyl- transferase (Collison et al., 2005) and hormone sensitive lipase (Hu et al., 2005). Although ACBP gene has all features of * Corresponding author at: Instituto de Bioquímica Médica, Universidade Federal do Rio de Janeiro, 21941-902, Rio de Janeiro, RJ, Brazil. Tel.: þ55 21 2562 6785; fax: þ55 21 2270 8647. E-mail address: [email protected] (K.C. Gondim). 1 The first two authors contributed equally to this work. Contents lists available at ScienceDirect Insect Biochemistry and Molecular Biology journal homepage: www.elsevier.com/locate/ibmb 0965-1748/$ e see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.ibmb.2010.01.002 Insect Biochemistry and Molecular Biology 40 (2010) 119e125
Transcript of Serotonin regulates an acyl-CoA-binding protein (ACBP) gene expression in the midgut of Rhodnius...

lable at ScienceDirect
Insect Biochemistry and Molecular Biology 40 (2010) 119e125
Contents lists avai
Insect Biochemistry and Molecular Biology
journal homepage: www.elsevier .com/locate/ ibmb
Serotonin regulates an acyl-CoA-binding protein (ACBP) gene expressionin the midgut of Rhodnius prolixus
Michele Alves-Bezerra a,1, David Majerowicz a,1, Luciano A.M. Grillo a,b, Hugo Tremonte c,Caroline B. Almeida a, Glória R.C. Braz c,e, Mauro Sola-Penna d,Gabriela O. Paiva-Silva a,e, Katia C. Gondim a,e,*
a Instituto de Bioquímica Médica, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazilb Escola de Enfermagem e Farmácia, Universidade Federal de Alagoas, Alagoas, BrazilcDepartamento de Bioquímica, Instituto de Química, Universidade Federal do Rio de Janeiro, Rio de Janeiro, BrazildDepartamento de Fármacos, Faculdade de Farmácia, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazile Instituto Nacional de Ciência e Tecnologia em Entomologia Molecular, Brazil
a r t i c l e i n f o
Article history:Received 6 November 2009Received in revised form16 December 2009Accepted 6 January 2010
Keywords:ACBPLipidAcyl-CoA esterSerotonin5-HTRhodnius prolixus
* Corresponding author at: Instituto de Bioquímicado Rio de Janeiro, 21941-902, Rio de Janeiro, RJ, Brafax: þ55 21 2270 8647.
E-mail address: [email protected] (K.C. Gondi1 The first two authors contributed equally to this
0965-1748/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.ibmb.2010.01.002
a b s t r a c t
Acyl-CoA esters have many intracellular functions, acting as energy source, substrate for metabolicprocesses and taking part in cell signaling. The acyl-CoA-binding protein (ACBP), a highly conserved10 kDa intracellular protein, binds long- and medium-chain acyl-CoA esters with very high affinity,directing them to specific metabolic routes and protecting them from hydrolysis. An ACBP gene sequencewas identified in the genome of Rhodnius prolixus. This ACBP gene (RpACBP-1) was expressed in allanalyzed tissues and quantitative PCR showed that expression was highest in posterior midgut. In thistissue, ACBP gene expression increased in the first day after blood meal (w10-fold) and then decreased tounfed levels in the seventh day after meal. Injection of serotonin (5-hydroxytryptamine; 5-HT), a neu-roamine released in the hemolymph after the start of feeding, increased the expression of this gene inthe midgut of unfed females, reaching levels similar to those observed in fed insects. This effect ofinjected 5-HT was inhibited by spiperone, an antagonist of 5-HT mammalian receptors, that was also ableto block the physiological increase in RpACBP-1 expression observed after feeding. Injection of choleratoxin or dibutyryl-cAMP also resulted in the stimulation of this gene expression. These data reveala transcriptional regulatory mechanism in R. prolixus, that is triggered by 5-HT. In this way, a novel rolefor 5-HT is proposed, as a regulator of ACBP gene expression and, consequently, taking part in the controlof lipid metabolism.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Although lipids are the main energy source for demandingmetabolic processes, as flight and oogenesis, little is known abouthow these hydrophobic macronutrients and their intermediatemetabolites, like fatty acyl-CoA esters, are maintained soluble inthe cytosol. Lipid-binding proteins, as the fatty acid-binding protein(FABP) and the acyl-CoA-binding protein (ACBP), have a crucial rolein this process. As ACBP shows an affinity for acyl-CoA that is fourorders of magnitude higher than FABP, this protein is considered
Médica, Universidade Federalzil. Tel.: þ55 21 2562 6785;
m).work.
All rights reserved.
very important for the regulation of lipid metabolism in the cells(Faergeman and Knudsen, 1997).
ACBP is a 10 kDa protein, highly conserved and present in alleukaryotes and some prokaryotes (Burton et al., 2005; Faergemanet al., 2007). This protein can bind long- and medium-chain acyl-CoA esters with high affinity and specificity (Rosendal et al., 1993),participates in the transport of acyl-CoA inside cells (Schjerlinget al., 1996) and protects these lipids from hydrolysis (Schroederet al., 2005), forming an intracellular pool of acyl-CoA. ACBP is alsoapparently involved in fatty acid elongation, sphingolipid synthesis,protein sorting and vesicle traffic (Gaigg et al., 2001; Faergemanet al., 2004). Furthermore, it has been shown that the presenceof ACBP can regulate the activity of enzymes involved in lipidmetabolism, as acetyl-CoA carboxylase andmitochondrial acyl-CoAsynthetase (Rasmussen et al., 1993), glycerol-3-phosphate acyl-transferase (Collison et al., 2005) and hormone sensitive lipase(Hu et al., 2005). Although ACBP gene has all features of

2 The Rhodnius prolixus genome was sequenced and assembled by the GenomeSequencing Center at Washington University School of Medicine in St. Louis, USA.
M. Alves-Bezerra et al. / Insect Biochemistry and Molecular Biology 40 (2010) 119e125120
a housekeeping gene (Mandrup et al., 1992), its transcription isregulated by both sterol regulatory element-binding protein(SREBP) and peroxisome proliferator-activated receptor (PPAR) inmammals hepatocytes (Sandberg et al., 2005) and adipocytes(Swinnen et al., 1998; Helledie et al., 2002; Neess et al., 2006). Inmouse brain, some reports have shown that ACBP expression canbe induced by psychological stress (Katsura et al., 2002), nitricoxide (Katsura et al., 2004), androgens and glucocorticoids(Compere et al., 2006), and morphine (Shibasaki et al., 2007),although how this expression increase occurs remain unclear.
In insects, ACBP expression was shown in a variety of species(Snyder and Feyereisen, 1993; Kolmer et al., 1994; Matsumoto et al.,2001; Liu et al., 2005; Sieglaff et al., 2005; Wang et al., 2008) and itseems to be involved in lipid and energetic metabolism and in thesynthesis of ecdysteroids. ACBP is present in tissues with highenergy demand in Drosophila melanogaster (Kolmer et al., 1994),and the knockdown of this protein expression, by RNA interference,reduced the size of lipid droplets and the amount of triacylglycerolstored in pheromone glands in Bombyx mori (Ohnishi et al., 2006).In the tobacco hornworm Manduca sexta, ACBP level in the midgutis highest at times of active feeding, what is consistent with itspossible role in the uptake of lipid nutrients from the diet (Snyderand Van Antwerpen,1997). Although variations in the expression ofinsect ACBP genes were described, mechanisms involved in thisregulation are basically unknown. So far, this has only been studiedin the silkmoth B. mori, where ACBP expression in the pheromonegland is induced by a humoral factor, identified as a b-D-glucosyl-O-L-tyrosine (Ohnishi et al., 2005).
In Rhodnius prolixus, a hematophagous hemipteran, vector ofChagas disease, triacylglycerols present in blood meal are hydro-lyzed in the midgut lumen by a triacylglycerol-lipase activity andreleased fatty acids are absorbed by the midgut epithelium (Grilloet al., 2007). Inside these cells, fatty acids are used for the synthesisof other lipids, such as diacylglycerols and phospholipids (Grilloet al., 2007) that are transferred to the major circulating lipoproteinin the hemolymph, lipophorin, and then distributed to other tissues(Atella et al., 1995; Coelho et al., 1997; Grillo et al., 2007). Duringthese events in the cytosol of midgut cells, the absorbed lipids andtheir intermediate metabolites must be solubilized, protected andtransported, and nothing is known about the possible role of ACBPin this process.
The event of feeding is under strong modulation of serotonin(5-hydroxytryptamine, 5-HT) in many invertebrates and also inmammals (Simansky, 1996; Orchard, 2006). In insects, this mole-cule can act either via serotonergic neurons or as a neurohormone(Lange et al., 1989). In R. prolixus 5-HT is released in the hemolymphwithin minutes of the onset of gorging (Lange et al., 1989; Barrettet al., 1993). This neurohormone coordinates feeding-relatedphysiological events, like diuresis and muscle contraction in sali-vary glands and posterior midgut (Orchard, 2006) and appears tobind to specific receptors, as antagonists of mammalian 5-HTreceptors decrease its effects (Maddrell et al., 1991; Ali and Orchard,1994; Grieco and Lopes, 1997). In this report, an ACBP gene ofR. prolixus (RpACBP-1) was identified and its expression profile wasanalyzed. It was highest expressed in the midgut at the first dayafter blood meal. As this induction appears to be regulated by thefeeding event, we investigated the role of 5-HT as a modulator ofthis increase in RpACBP-1 gene expression.
2. Material and methods
2.1. Insects
Insects were taken from a colony of R. prolixus maintained at28 �C and 80e90% relative humidity, under a photoperiod of 12 h
light/12 h dark. Adult insects were fed on live rabbit blood at3-week intervals, starting 15 days after moulting. Experimentalinsects were adult females after the third meal, or starving femalesthree weeks after the second blood meal (unfed condition).
2.2. RpACBP-1 gene identification and annotation
RpACBP-1 gene was identified in R. prolixus genome2 tracearchives database, using an ACBP sequence found in a midgut cDNAlibrary (GenBank accession no. EU233793) as query in a Blastnsearch (Altschul et al., 1997). Blastn hit was used as seed sequenceto assemble the complete RpACBP-1 gene sequence using GenSeedsoftware (Sobreira and Gruber, 2008) and the genome tracearchives database. The obtained gene sequencewas confirmed afterthe release of the R. prolixus genome assembly version 1.0. Codingregions were localized using Blastx (Altschul et al., 1997). Splicingsites were predicted by NetGene 2.0, using Caenorhabditis elegansnetwork (Brunak et al., 1991; Hebsgaard et al., 1996) and tran-scription initiation site was predicted by Neural Network PromoterPrediction (Reese, 2001).
2.3. Tissue isolation
For RpACBP-1 expression analysis in different tissues, adultfemales were dissected four days after feeding and the followingtissues were obtained: anterior midgut, posterior midgut, ovary, fatbody and salivary glands. For expression analysis of RpACBP-1 inposterior midgut during digestion, females were dissected beforefeeding and at different days after blood meal. To investigate therole of possible modulators of RpACBP-1 gene expression, theposterior midguts of females injected with different substanceswere dissected 24 h after each treatment, as described below.
2.4. 5-HT treatment
Unfed females were injected with the help of a 10 ml syringe(Hamilton Company, Reno, USA) with 2 or 4 pmol of 5-hydroxy-tryptamine (SigmaeAldrich Co, USA) diluted in 1 ml of Milli-Q water(Millipore, Bedford, USA). Females injected with 1 ml of Milli-Qwater were used as experimental control. The insects weredissected 24 h after injection and total RNA was extracted fromposterior midgut.
2.5. Spiperone treatment
Unfed females were injected with 4 pmol of 5-hydroxytrypta-mine diluted in 2 ml of Milli-Q water in the presence or absence of 9or 100 pmol of spiperone (SigmaeAldrich Co, USA). Femalesinjected with 2 ml of Milli-Q water were used as experimentalcontrol. The insects were dissected 24 h after injection and totalRNA was extracted from posterior midgut. Alternatively, unfedfemales were injected with 9 pmol of spiperone diluted in 2 ml ofMilli-Q water or with 2 ml of Milli-Q water as control. Insects werefed immediately after injection, were dissected 24 h later and totalRNA was extracted from posterior midgut.
2.6. Dibutyryl-cyclic AMP treatment
Unfed females were injected with 5 or 10 nmol of dibutyryl-cyclic AMP (Calbiochem, San Diego, USA) diluted in 1 ml of 0.2%dimethyl sulfoxide (Me2SO; Merck KGaA, Darmstadt, Germany).

M. Alves-Bezerra et al. / Insect Biochemistry and Molecular Biology 40 (2010) 119e125 121
Females injected with 1 ml of 0.2% Me2SO were used as experi-mental control. Insects were dissected 24 h after injection and totalRNA was extracted from posterior midgut.
2.7. Cholera toxin treatment
Unfed females were injected with 10, 50 or 100 nmol of choleratoxin (SigmaeAldrich Co, USA) diluted in 1 ml of Milli-Q water.Females injected with 1 ml of Milli-Q water were used as experi-mental control. Insects were dissected 24 h after injection and totalRNA was extracted from posterior midgut.
2.8. RNA isolation
The dissected tissues were washed in cold deionized water andtotal RNA was isolated from samples using TRIzol� Reagent (Invi-trogen Corporation, Carlsbad, USA) according to the manufacturer'sinstructions. Total RNA concentrations were determined spectro-photometrically at 260 nm. One microgram of RNA was treatedwith RNase-free DNaseI (Fermentas International Inc., Burlington,Canada) and used to synthesize cDNA samples using High-CapacitycDNA Reverse Transcription Kit (Applied Biosystems, Foster City,USA).
2.9. Reverse Transcriptase-PCR (RT-PCR)
RT-PCR reactions were carried out using Taq DNA Polymerase(Fermentas International Inc., Burlington, Canada) under thefollowing conditions: one cycle for 2 min at 94 �C, followed by 30cycles of 30 s at 94 �C, 30 s at 50 �C and 30 s at 72 �C, and then a finalextension at 72 �C for 10 min. RpACBP-1 PCR amplification wasperformed using the following primers: RpACBP1 F 50- CTGGGGACTGTAATACGAG -30 (forward) and RpACBP1 R 50- TCCATAAGATGCAATCAGTG -30 (reverse). These primers were designed basedon the RpACBP-1 identified in the cDNA library of midgut ofR. prolixus. Actin amplification was used as positive control withthe following primers: RpAct F 50-AGTAGCTGCATGGGTTGTAG-30
(forward) and RpAct R 50-CAACATACATTGCTGGACTG-30 (reverse).These primers were designed based on an actin cDNA sequencedescribed elsewhere (Ribeiro et al., 2004). The above primers weredesigned using Primer3 software (Rozen and Skaletsky, 2000). PCRproducts were subjected to 2% agarose gel eletrophoresis andvisualized under UV light.
2.10. Quantitative PCR (qPCR)
For qPCR, an Applied Biosystems ABI Prism 7500 Real-Time PCRSystem (Applied Biosystems, Foster City, USA) and the SYBR GreenPCR Master Mix (Applied Biosystems, Foster City, USA) were usedunder the following conditions: one cycle for 10 min at 95 �C,followed by 50 cycles of 15 s at 95 �C and 45 s at 60 �C. RpACBP-1PCR amplification was performed using the following primers:RpACBP1RT F 50- GGGGACTGTAATACGAGCAA -30 (forward) andRpACBP1RT R 50- TTCAATCCATAAGATGCAATCA -30 (reverse). Allassays were done in triplicate and actin expression was used fornormalization. Primers used for actin PCR amplification: RpActRTF 50-CCATGTACCCAGGTATTGCT-30 (forward) and RpActRT R50-ATCTGTTGGAAGGTGGACAG-30 (reverse).
2.11. Statistical analyses of qPCR results
Relative expression and DDCt values were calculated from Ct(cycle threshold) values obtained on qPCR as described elsewhere(Norgard et al., 2006). The DDCt mean values obtained fromexperiments were submitted to Grubb's test to detect outliers
(Burns et al., 2005) and the comparison between different condi-tions was made using ANOVA followed by Tukey's multiplecomparison test.When only two conditions were compared (Fig. 5),unpaired Student's t test was used. Differences were consideredsignificant at P < 0.05. The relative expression values (2�DDCt) wereused only for graphic construction. All statistical analyses wereperformed using Prism 4.0 software (GraphPad Software, SanDiego, USA).
3. Results
In R. prolixus, lipids from bloodmeal are absorbed by the midgutepithelium, where they are metabolized (Grillo et al., 2007).Probably, ACBP takes part in the transport of acyl-CoA required bypathways of lipid biosynthesis.
An ACBP cDNA sequence was identified in a cDNA library fromR. prolixus midgut (GenBank accession no. EU233793). It was usedas query to search R. prolixus genome trace archives database usingBlastn algorithm. The best hit was subjected to GenSeed softwareand this analysis resulted in an assembled contig with about 3 kbnamed RpACBP-1. This sequence was found in R. prolixus genomein the contig 1271.1 (GenBank accession no. ACPB01048817.1)between positions 332 and 3509. The gene has 3 introns, one ofthem in the 50 untranslated region, and codes a protein with 89amino acids (Fig. 1). Comparison of this sequence with ACBPs fromyeast, arthropods and mouse showed high identity (46e69%) andsimilarity (62e83%) among them (Table 1). Alignment of RpACBP-1and ACBPs from other organisms (Table 1) showed that this proteinhas conserved residues, mainly in the central region (data notshown), including those identified as important for binding acyl-CoA esters (Fig. 1) (Kragelund et al., 1999).
In order to identify the sites where RpACBP-1 is expressed,different tissues of females were dissected and tested for thepresence of RpACBP-1 transcripts by RT-PCR. After a blood meal, anadult of R. prolixus takes about two weeks to complete digestionand oviposition. Females were dissected at 4th day after bloodmeal, when they are metabolically very active, and expressioncould be detected in all tissues analyzed: fat body, salivary glands,anterior midgut, posterior midgut and ovaries (Fig. 2A).
Subsequent analysis by qPCR revealed that the highest level ofRpACBP-1 expression was found in posterior midgut (Fig. 2B). Inaddition, the expression profile of RpACBP-1 in this tissue wasdetermined during the first week after feeding, as blood digestionoccurred (Fig. 3). This expression was maximal in the first day afterblood meal (about 10-fold compared to unfed females) and, in theconsecutive days, it returned to levels found in unfed insects (Fig. 3).
In adults of R. prolixus, 5-HT concentration in the hemolymphincreases three-fold as soon as feeding starts and returns to basallevels 15e20 min later (Barrett et al., 1993). In order to investigatea possible role of 5-HT in the transcriptional activation mechanismof RpACBP-1, unfed females were injected with this neurohormoneand RpACBP-1 mRNA levels were determined in the posteriormidgut (Fig. 4A). It was observed that an injection of 4 pmolresulted in an expression level 5-fold higher than the control. Thiseffect was abolished by the simultaneous injection of spiperone(Fig. 4B), a classical antagonist of some subtypes of 5-HT receptorsin vertebrate systems (Hoyer et al., 1994; Metwally et al., 1998).When 100 pmol of spiperone was injected, its inhibitory effect wasnot observed, and the stimulation of RpACBP-1 expression by 5-HTactivation occurred (Fig. 4B). This is an already known phenom-enon, as spiperone may have inverse agonist activity at highconcentrations (Miller et al., 1997). Moreover, spiperone was alsoable to abolish the effect triggered by feeding on RpACBP-1expression. When females were injected with spiperone beforefeeding, the increase in this gene transcription, normally observed

Table 1Comparison between amino acid sequences of ACBP from Rhodnius prolixus andother organisms.
Accession numbera Species Identity (%)b Similarity (%)b
XP_001664028 Aedes aegypti 65 83XP_001237266 Anopheles gambiae 67 82XP_394745 Apis mellifera 61 76XP_001845393 Culex quinquefasciatus 68 84NP_609187 Drosophila melanogaster 66 81ACU82846 Fenneropenaeus chinensis 65 80ABM55609 Maconellicoccus hirsutus 61 81XP_001605849 Nasonia vitripennis 69 83ACX53980 Rhipicephalus sanguineus 64 75XP_974824 Tribolium castaneum 61 81NP_011551.1 Saccharomyces cerevisiae 46 62NP_031856.1 Mus musculus 54 71
a Accession numbers for amino acid sequences obtained from GenBank.b Percentages of identity and similarity were calculated using Blastp algorithm.
Fig. 2. Expression analysis of RpACBP-1 gene in different tissues. Four days after bloodmeal, adult females were dissected and total RNA was extracted from tissues. RNAsamples were treated with DNaseI and RT-PCR and qPCR were carried out usingspecific primers designed to RpACBP-1 sequence. Actin amplification was used aspositive and endogenous control. FB: fat body; SG: salivary glands; AM: anteriormidgut; PM: posterior midgut; OV: ovary. (A) RT-PCR products were subjected toagarose gel electrophoresis; the image is representative from 3 independent experi-ments. (B) In qPCR analysis, RpACBP-1 expression levels are relative to fat body values(1.0). Results are mean � S.E.M. for 4 independent determinations. (**): P < 0.01against all other tissues.
Fig. 1. Nucleotide and amino acid sequences of the Rhodnius prolixus RpACBP-1. TheRpACBP-1 gene sequence was obtained from Rhodnius prolixus genome trace archivesand assembled by GenSeed as described in Material and Methods. The promoter regionis underlined. Nucleotides in bold indicate exons and polyadenylation site is in italic.The asterisk indicates the stop codon. Residues that are important for acyl-CoA bindingare boxed.
M. Alves-Bezerra et al. / Insect Biochemistry and Molecular Biology 40 (2010) 119e125122
after blood meal, was inhibited (Fig. 5). This result reinforces therole of 5-HT as a physiological modulator of RpACBP-1 expression,as its antagonist was able to reverse the natural stimulus triggeredby feeding.
As previously described (Barrett et al., 1993), there is a 5-HT-induced elevation of cAMP content in anterior midgut in R. prolixus.To determine if cAMP is involved in the observed activation ofRpACBP-1 expression by 5-HT, unfed females were injected withdibutyryl-cAMP, a permeant cAMP analog (Fig. 6). This treatmentwas able to increase RpACBP-1 mRNA levels in comparison to
Fig. 3. RpACBP-1 gene expression profile in posterior midgut at different days afterblood meal. Adult females, before feeding and at different days after blood meal, weredissected and total RNA was extracted from posterior midgut. RNA samples weretreated with DNaseI and then used for cDNA synthesis. Differences between cDNAlevels were determined by qPCR. RpACBP-1 expression levels are relative to day0 (unfed) values (1.0). Results are mean � S.E.M. for 4e5 determinations. (**) and (*):P < 0.01 and P < 0.05 against values for unfed females, respectively.
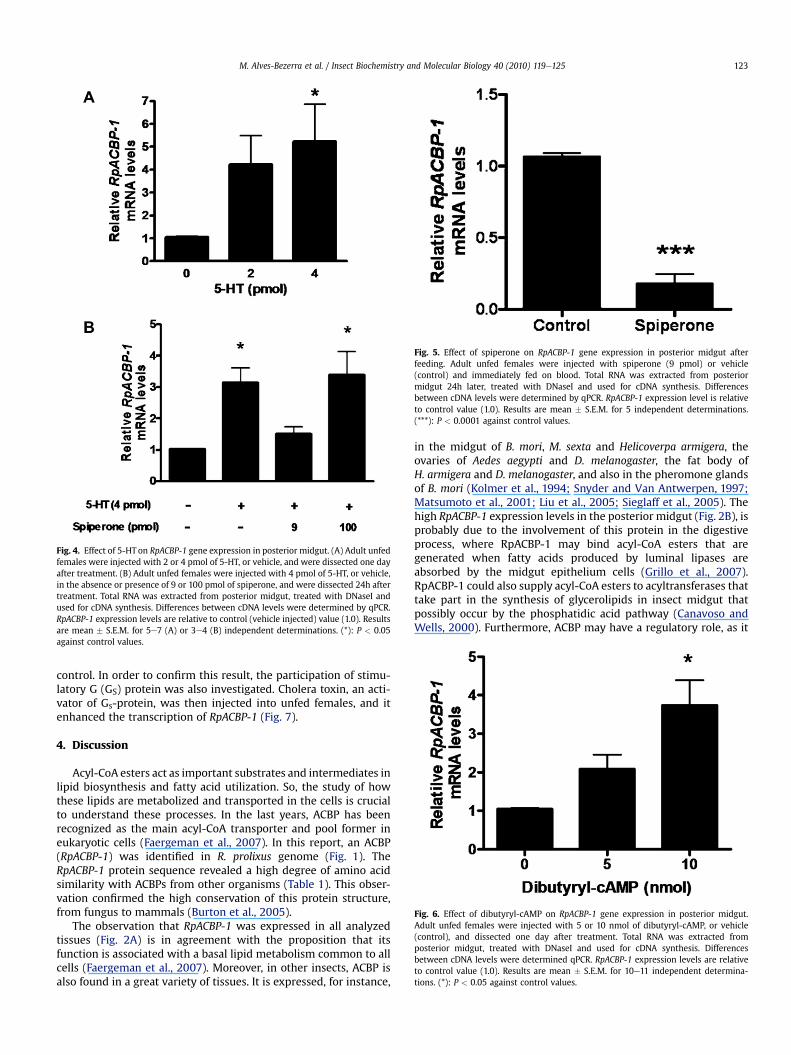
Fig. 4. Effect of 5-HT on RpACBP-1 gene expression in posterior midgut. (A) Adult unfedfemales were injected with 2 or 4 pmol of 5-HT, or vehicle, and were dissected one dayafter treatment. (B) Adult unfed females were injected with 4 pmol of 5-HT, or vehicle,in the absence or presence of 9 or 100 pmol of spiperone, and were dissected 24h aftertreatment. Total RNA was extracted from posterior midgut, treated with DNaseI andused for cDNA synthesis. Differences between cDNA levels were determined by qPCR.RpACBP-1 expression levels are relative to control (vehicle injected) value (1.0). Resultsare mean � S.E.M. for 5e7 (A) or 3e4 (B) independent determinations. (*): P < 0.05against control values.
Fig. 5. Effect of spiperone on RpACBP-1 gene expression in posterior midgut afterfeeding. Adult unfed females were injected with spiperone (9 pmol) or vehicle(control) and immediately fed on blood. Total RNA was extracted from posteriormidgut 24h later, treated with DNaseI and used for cDNA synthesis. Differencesbetween cDNA levels were determined by qPCR. RpACBP-1 expression level is relativeto control value (1.0). Results are mean � S.E.M. for 5 independent determinations.(***): P < 0.0001 against control values.
Fig. 6. Effect of dibutyryl-cAMP on RpACBP-1 gene expression in posterior midgut.Adult unfed females were injected with 5 or 10 nmol of dibutyryl-cAMP, or vehicle(control), and dissected one day after treatment. Total RNA was extracted fromposterior midgut, treated with DNaseI and used for cDNA synthesis. Differencesbetween cDNA levels were determined qPCR. RpACBP-1 expression levels are relativeto control value (1.0). Results are mean � S.E.M. for 10e11 independent determina-tions. (*): P < 0.05 against control values.
M. Alves-Bezerra et al. / Insect Biochemistry and Molecular Biology 40 (2010) 119e125 123
control. In order to confirm this result, the participation of stimu-latory G (GS) protein was also investigated. Cholera toxin, an acti-vator of Gs-protein, was then injected into unfed females, and itenhanced the transcription of RpACBP-1 (Fig. 7).
4. Discussion
Acyl-CoA esters act as important substrates and intermediates inlipid biosynthesis and fatty acid utilization. So, the study of howthese lipids are metabolized and transported in the cells is crucialto understand these processes. In the last years, ACBP has beenrecognized as the main acyl-CoA transporter and pool former ineukaryotic cells (Faergeman et al., 2007). In this report, an ACBP(RpACBP-1) was identified in R. prolixus genome (Fig. 1). TheRpACBP-1 protein sequence revealed a high degree of amino acidsimilarity with ACBPs from other organisms (Table 1). This obser-vation confirmed the high conservation of this protein structure,from fungus to mammals (Burton et al., 2005).
The observation that RpACBP-1 was expressed in all analyzedtissues (Fig. 2A) is in agreement with the proposition that itsfunction is associated with a basal lipid metabolism common to allcells (Faergeman et al., 2007). Moreover, in other insects, ACBP isalso found in a great variety of tissues. It is expressed, for instance,
in the midgut of B. mori, M. sexta and Helicoverpa armigera, theovaries of Aedes aegypti and D. melanogaster, the fat body ofH. armigera and D. melanogaster, and also in the pheromone glandsof B. mori (Kolmer et al., 1994; Snyder and Van Antwerpen, 1997;Matsumoto et al., 2001; Liu et al., 2005; Sieglaff et al., 2005). Thehigh RpACBP-1 expression levels in the posterior midgut (Fig. 2B), isprobably due to the involvement of this protein in the digestiveprocess, where RpACBP-1 may bind acyl-CoA esters that aregenerated when fatty acids produced by luminal lipases areabsorbed by the midgut epithelium cells (Grillo et al., 2007).RpACBP-1 could also supply acyl-CoA esters to acyltransferases thattake part in the synthesis of glycerolipids in insect midgut thatpossibly occur by the phosphatidic acid pathway (Canavoso andWells, 2000). Furthermore, ACBP may have a regulatory role, as it

Fig. 7. Effect of cholera toxin on RpACBP-1 gene expression in posterior midgut. Adultunfed females were injected with 10, 50 or 100 nmol of cholera toxin, or vehicle(control), and dissected one day after treatment. Total RNA was extracted fromposterior midgut, treated with DNaseI and used for cDNA synthesis. Differencesbetween cDNA levels were determined by qPCR. RpACBP-1 expression levels arerelative to control value (1.0). Results are mean � S.E.M. for 3 independent determi-nations. (*): P < 0.05 against control values.
M. Alves-Bezerra et al. / Insect Biochemistry and Molecular Biology 40 (2010) 119e125124
was shown in rat T-lymphocytes that this proteinmay stimulate theactivity of glycerol-3-phosphate acyltransferase, which catalysesthe limiting step in the phosphatidic acid pathway (Collison et al.,2005). The importance of RpACBP-1 for the digestive process ofR. prolixus is also sustained by the observation that its transcrip-tional level was maximal at the first day after meal (Fig. 3), whenlipid metabolism in the midgut is very high. At this moment, theactivity of the luminal triacylglycerol-lipase and the consequentgeneration and absorption of free fatty acids are maximal (Grilloet al., 2007). Accordingly, the midgut also has a very high capacityto transfer lipids to the circulating lipoprotein, lipophorin, in thefirst days after blood meal (Coelho et al., 1997).
In other insect species, ACBP presence is also associated withtissues involvedwith high energy demand. InM. sextamidgut, ACBPmRNA and protein levels are highest at times of active feedingduring the final larval instar (Snyder and Van Antwerpen, 1997).Similarly, inH. armigera, ACBP gene expression in themidgut is veryhigh between the first and second day after the emerging of sixthlarval instar, when insects have a great feeding activity (Liu et al.,2005; Wang et al., 2008). The present results reinforce the hypoth-esis that this protein is involved with the intake of lipids frommeal.
The transcription of RpACBP-1 was stimulated after blood meal(Fig. 3), when there is also an increase in the hemolymphaticconcentration of 5-HT (Barrett et al., 1993). In R. prolixus, thissubstance acts as a neurohormone, and is released in the hemo-lymph a few minutes after the start of feeding by gorging stimulus(Lange et al., 1989). It coordinates several feeding-related physio-logical processes like cuticle plasticization, salivation, increased gutmotility and heart rate, and diuresis (Orchard, 2006). In addition tothese effects, that in general have short response times and are notat genomic levels, in this study we have observed that 5-HT wasable to enhance the transcription of RpACBP-1 in the midgut ofR. prolixus (Fig. 4A).
Spiperone was previously described to prevent other 5-HTeffects in R. prolixus (Maddrell et al., 1991) but, at present, R. prolixus5-HT receptors are still uncharacterized and new studies arerequired to identify and classify them, and to describe their struc-tural and pharmacological properties. In mammals, spiperonebinds, with high affinity, to 5-HT receptors of type 5-HT1A, 5-HT2Aand 5-HT7 (Zifa and Fillion, 1992; Ruat et al., 1993; Metwally et al.,1998). Spiperone inhibited the stimulatory effect of 5-HT on
RpACBP-1 expression and, so, if this only property is considered forcomparison with mammalian 5-HT receptors, R. prolixus receptormay be similar to the above mentioned receptor types, but a clas-sification for invertebrate receptors still needs to be well estab-lished. However, the fact that spiperone was able to reverse thephysiological stimulus of feeding on RpACBP-1 expression (Fig. 5),strongly reinforced the importance of 5-HT in this regulatorymechanism. To our knowledge, B. mori is the only other insectwhere the regulation of ACBP gene transcription was studied. Inthis case, a hemolymphatic factor, identified as b-D-glucosyl-O-L-tyrosine was shown to trigger it (Ohnishi et al., 2005).
The fact that the injection of either dibutyryl-cAMP or choleratoxin into unfed females caused the increase of RpACBP-1 expres-sion (Figs. 6 and 7) indicates that 5-HT affects gene expression bya mechanism that involves cAMP as second messenger, which issynthesized by adenylyl cyclase, after activation by a Gs-protein.However, the complete pathway that connects cAMP with theincrease in RpACBP-1 expression, including possible kinases andtranscription factors, is still unknown.
It has been demonstrated that, in insects, cAMP is the secondmessenger for 5-HT, when different effects of this neurohormoneare considered. In R. prolixus, during post-prandial diuresis, therates of fluid absorption by the anterior midgut and of fluidsecretion by the Malpighian tubules are stimulated by 5-HT,via a cAMP-dependent mechanism (Farmer et al., 1981; Maddrellet al., 1991). This was also demonstrated for the regulation ofa (NaþþKþ)ATPase activity in Malpighian tubules of this insect(Grieco and Lopes, 1997). In addition, 5-HT stimulates secretion insalivary glands of the blowfly Calliphora erytrocephala and regulatescontractions in locust mandibular closer muscles through theincrease in cAMP concentrations (Heslop and Berridge, 1980;Baines et al., 1990).
In insects, especially in R. prolixus, althoughmany effects of 5-HTon the acquisition and digestion of blood meal have been described(Orchard, 2006), none of them occur at the level of gene expressioncontrol or is related to lipid metabolism. So, this study providesnew insights on 5-HT action, as it is described as a modulator ofACBP gene expression and, probably, has a role in the regulation oflipid flux in this insect.
Acknowledgements
The authors thank Dr. Pedro L. Oliveira for a critical reading ofthis manuscript, Lilian S. C. Gomes and Heloisa S. L. Coelho forexcellent technical assistance and José de S. Lima Junior and LitianeM. Rodrigues for insect care.
This work was supported by grants from Conselho Nacionalde Desenvolvimento Científico e Tecnológico (CNPq), Fundaçãode Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) andInstituto Nacional de Ciência e Tecnologia em EntomologiaMolecular (INCT-EM).
References
Ali, D.W., Orchard, I., 1994. Characterization of dopamine and serotonin receptors onthe salivary glands of the locust, Locusta migratoria. Biog. Amines 10, 195e212.
Altschul, S.F., Madden, T.L., Schäffer, A.A., Zhang, J., Zhang, Z., Miller, W., Lipman, D.J.,1997. Gapped BLAST and PSI-BLAST: a new generation of protein databasesearch programs. Nucleic Acids Res. 25, 3389e3402.
Atella, G.C., Gondim, K.C., Masuda, H., 1995. Loading of lipophorin particles withphospholipids at the midgut of Rhodnius prolixus. Arch. Insect Biochem. Physiol.30, 337e350.
Baines, R.A., Tyrer, N.M., Downer, R.G., 1990. Serotoninergic innervation of thelocust mandibular closer muscle modulates contractions through the elevationof cyclic adenosine monophosphate. J. Comp. Neurol. 294, 623e632.
Barrett, F.M., Orchard, I., Tebrugge, V., 1993. Characteristics of serotonin-inducedcyclic AMP elevation in the integument and anterior midgut of the blood-feeding bug, Rhodnius prolixus. J. Insect Physiol. 39, 581e587.

M. Alves-Bezerra et al. / Insect Biochemistry and Molecular Biology 40 (2010) 119e125 125
Brunak, S., Engelbrecht, J., Knudsen, S., 1991. Prediction of human mRNA donor andacceptor sites from the DNA sequence. J. Mol. Biol. 220, 49e65.
Burns, M.J., Nixon, G.J., Foy, C.A., Harris, N., 2005. Standardisation of data from real-time quantitative PCR methods e evaluation of outliers and comparison ofcalibration curves. BMC Biotechnol. 5, 31.
Burton, M., Rose, T.M., Faergeman, N.J., Knudsen, J., 2005. Evolution of the acyl-CoAbinding protein (ACBP). Biochem. J. 392, 299e307.
Canavoso, L.E., Wells, M.A., 2000. Metabolic pathways for diacylglycerol biosyn-thesis and release in the midgut of larval Manduca sexta. Insect Biochem. Mol.Biol. 30, 1173e1180.
Coelho, H.S.L., Atella, G.C., Moreira, M.F., Gondim, K.C., Masuda, H., 1997. Lipophorindensity variation during oogenesis in Rhodnius prolixus. Arch. Insect Biochem.Physiol. 35, 301e313.
Collison, L.W., Kannan, L., Onorato, T.A., Knudsen, J., Haldar, D., Jolly, C.A., 2005.Aging reduces glycerol-3-phosphate acyltransferase activity in activated ratsplenic T-lymphocytes. Biochim. Biophys. Acta 1687, 164e172.
Compere, V., Ouellet, J., Luu-The, V., Dureuil, B., Tonon, M.C., Vaudry, H., Labrie, F.,Pelletier, G., 2006. Role of androgens and glucocorticoids in the regulation ofdiazepam-binding inhibitor mRNA levels in male mouse hypothalamus. BrainRes. 1119, 50e57.
Faergeman, N.J., Feddersen, S., Christiansen, J.K., Larsen, M.K., Schneiter, R.,Ungermann, C., Mutenda, K., Roepstorff, P., Knudsen, J., 2004. Acyl-CoA-bindingprotein, Acb1p, is required for normal vacuole function and ceramide synthesisin Saccharomyces cerevisiae. Biochem. J. 380, 907e918.
Faergeman, N.J., Knudsen, J., 1997. Role of long-chain fatty acyl-CoA esters in theregulation of metabolism and in cell signalling. Biochem. J. 323, 1e12.
Faergeman, N.J., Wadum, M., Feddersen, S., Burton, M., Kragelund, B.B., Knudsen, J.,2007. Acyl-CoA binding proteins; structural and functional conservation over2000 MYA. Mol. Cell. Biochem. 299, 55e65.
Farmer, J., Maddrell, S.H.P., Spring, J.H., 1981. Absorption of fluid by the midgut ofRhodnius. J. Exp. Biol. 94, 301e316.
Gaigg, B., Neergaard, T.B.F., Schneiter, R., Hansen, J.K., Faergeman, N.J., Jensen, N.A.,Andersen, J.R., Friis, J., Sandhoff, R., Schroder, H.D., Knudsen, J., 2001. Depletionof acyl-coenzyme-A-binding protein affects sphingolipid synthesis and causesvesicle accumulation and membrane defects in Saccharomyces cerevisiae. Mol.Biol. Cell 12, 1147e1160.
Grieco, M.A.B., Lopes, A.G., 1997. 5-Hydroxytryptamine regulates the (NaþþKþ)ATPase activity in Malpighian tubules of Rhodnius prolixus: evidence forinvolvement of G-protein and cAMP-dependent protein kinase. Arch. InsectBiochem. Physiol. 36, 203e214.
Grillo, L.A.M., Majerowicz, D., Gondim, K.C., 2007. Lipid metabolism in Rhodniusprolixus (Hemiptera: Reduviidae): role of a midgut triacylglycerol-lipase. InsectBiochem. Mol. Biol. 37, 579e588.
Hebsgaard, S.M., Korning, P.G., Tolstrup, N., Engelbrecht, J., Rouzé, P., Brunak, S.,1996. Splice site prediction in Arabidopsis thaliana pre-mRNA by combininglocal and global sequence information. Nucleic Acids Res. 24, 3439e3452.
Helledie, T., Grontved, L., Jensen, S.S., Kiilerich, P., Rietveld, L., Albrektsen, T.,Boysen, M.S., Nohr, J., Larsen, L.K., Fleckner, J., Stunnenberg, H.G., Kristiansen, K.,Mandrup, S., 2002. The gene encoding the Acyl-CoA-binding protein is acti-vated by peroxisome proliferator-activated receptor gamma through an intronicresponse element functionally conserved between humans and rodents. J. Biol.Chem. 277, 26821e26830.
Heslop, J.P., Berridge, M.J., 1980. Changes in cyclic AMP and cyclic GMP concen-trations during the action of 5-hydroxytryptamine on an insect salivary gland.Biochem. J. 192, 247e255.
Hoyer, D., Clarke, D.E., Fozard, J.R., Hartig, P.R., Martin, G.R., Mylecharane, E.J.,Saxena, P.R., Humphrey, P.P., 1994. International union of pharmacology clas-sification of receptors for 5-hydroxytryptamine (serotonin). Pharmacol. Rev. 46,157e203.
Hu, L.P., Deeney, J.T., Nolan, C.J., Peyot, M.L., Ao, A., Richard, A.M., Luc, E.,Faergeman, N.J., Knudsen, J., Guo, W., Sorhede-Winzell, M., Prentki, M.,Corkey, B.E., 2005. Regulation of lipolytic activity by long-chain acyl-coenzymeA in islets and adipocytes. Am. J. Physiol. Endocrinol. Metab. 289, E1085eE1092.
Katsura, M., Mohri, Y., Shuto, K., Tsujimura, A., Ukai, M., Ohkuma, S., 2002.Psychological stress, but not physical stress, causes increase in diazepambinding inhibitor (DBI) mRNA expression in mouse brains. Brain Res. Mol. BrainRes. 104, 103e109.
Katsura, M., Shuto, K., Mohri, Y., Tsujimura, A., Shibata, D., Tachi, M., Okuma, S.,2004. Continuous exposure to nitric oxide enhances diazepam binding inhibitormRNA expression in mouse cerebral cortical neurons. Brain Res. Mol. Brain Res.124, 29e39.
Kolmer, M., Roos, C., Tirronen, M., Myohanen, S., Alho, H., 1994. Tissue specificexpression of the diazepam binding inhibitor in Drosophila melanogaster:cloning, structure, and localization of the gene. Mol. Cell. Biol. 14, 6983e6995.
Kragelund, B.B., Poulsen, K., Andersen, K.V., Baldursson, T., Krøll, J.B., Neergård, T.B.,Jepsen, J., Roepstorff, P., Kristiansen, K., Poulsen, F.M., Knudsen, J., 1999.Conserved residues and their role in the structure, function, and stability ofacyl-coenzyme A binding protein. Biochem. (Mosc.) 38, 2386e2394.
Lange, A.B., Orchard, I., Michael Barrett, F., 1989. Changes in haemolymph serotoninlevels associated with feeding in the blood-sucking bug, Rhodnius prolixus.J. Insect Physiol. 35, 393e399.
Liu, M., Zhang, T.Y., Xu, W.H., 2005. A cDNA encoding diazepam-binding inhibitor/acyl-CoA-binding protein in Helicoverpa armigera: molecular characterizationand expression analysis associated with pupal diapause. Comp. Biochem.Physiol. C 141, 168e176.
Maddrell, S.H., Herman,W.S.,Mooney, R.L., Overton, J.A.,1991. 5-Hydroxytryptamine:a second diuretic hormone in Rhodnius prolixus. J. Exp. Biol. 156, 557e566.
Mandrup, S., Hummel, R., Ravn, S., Jensen, G., Andreasen, P.H., Gregersen, N.,Knudsen, J., Kristiansen, K., 1992. Acyl-CoA-binding protein, diazepam bindinginhibitor gene and pseudogenes: a typical housekeeping gene family. J. Mol.Biol. 228, 1011e1022.
Matsumoto, S., Yoshiga, T., Yokoyama, N., Iwanaga, M., Koshiba, S., Kigawa, T.,Hirota, H., Yokoyama, S., Okano, K., Mita, K., Shimada, T., Tatsuki, S., 2001.Characterization of acyl-CoA-binding protein (ACBP) in the pheromone gland ofthe silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 31, 603e609.
Metwally, K.A., Dukat, M., Egan, C.T., Smith, C., DuPre, A., Gauthier, C.B., Herrick-Davis, K., Teitler, M., Glennon, R.A., 1998. Spiperone: influence of spiro ringsubstituents on5-HT2Aserotonin receptor binding. J.Med. Chem.41, 5084e5093.
Miller, K.J., Mariano, C.L., Cruz, W.R., 1997. Serotonin 5HT2A receptor activationinhibits inducible nitric oxide synthase activity in C6 glioma cells. Life Sci. 61,1819e1827.
Neess, D., Kiilerich, P., Sandberg, M.B., Helledie, T., Nielsen, R., Mandrup, S., 2006.ACBP e a PPAR and SREBP modulated housekeeping gene. Mol. Cell. Biochem.284, 149e157.
Norgard, O., Kvaloy, J.T., Farmen, R.K., Heikkila, R., 2006. Error propagation in relativereal-time reverse transcriptionpolymerase chain reactionquantificationmodels:the balance between accuracy and precision. Anal. Biochem. 356, 182e193.
Ohnishi, A., Hull, J.J., Matsumoto, S., 2006. Targeted disruption of genes in theBombyx mori sex pheromone biosynthetic pathway. Proc. Natl. Acad. Sci. U S A103, 4398e4403.
Ohnishi, A., Koshino, H., Takahashi, S., Esumi, Y., Matsumoto, S., 2005. Isolation andcharacterization of a humoral factor that stimulates transcription of the acyl-CoA-binding protein in the pheromone gland of the silkmoth, Bombyx mori.J. Biol. Chem. 280, 4111e4116.
Orchard, I., 2006. Serotonin: a coordinator of feeding-related physiological events intheblood-gorgingbug,Rhodnius prolixus. Comp.Biochem.Physiol.144A, 316e324.
Rasmussen, J.T., Rosendal, J., Knudsen, J., 1993. Interaction of acyl-CoA-bindingprotein (ACBP) on processes for which acyl-CoA is a substrate, product orinhibitor. Biochem. J. 292, 907e913.
Reese, M.G., 2001. Application of a time-delay neural network to promoter anno-tation in the Drosophila melanogaster genome. Comput. Chem. 26, 51e56.
Ribeiro, J.M., Andersen, J., Silva-Neto,M.A., Pham, V.M., Garfield,M.K., Valenzuela, J.G.,2004. Exploring the sialome of the blood-sucking bug Rhodnius prolixus. InsectBiochem. Mol. Biol. 34, 61e79.
Rosendal, J., Ertbjerg, P., Knudsen, J., 1993. Characterization of ligand binding toacyl-CoA-binding protein. Biochem. J. 290, 321e326.
Rozen, S., Skaletsky, H.J., 2000. Primer3 on the WWW for general users and for biol-ogist programmers. In: Krawetz, S.,Misener, S. (Eds.), Bioinformaticsmethods andprotocols: methods in molecular biology. Humana Press, Totowa, pp. 365e386.
Ruat,M., Traiffort, E., Leurs, R., Tardivel-Lacombe, J., Diaz, J., Arrang, J.M., Schwartz, J.C.,1993. Molecular cloning, characterization, and localization of a high-affinityserotonin receptor (5-HT7) activating cAMP formation. Proc. Natl. Acad. Sci. U S A90, 8547e8551.
Sandberg, M.B., Bloksgaard, M., Duran-Sandoval, D., Duval, C., Staels, B., Mandrup, S.,2005. The gene encoding Acyl-CoA-binding protein is subject to metabolic regu-lation by both sterol regulatory element-binding protein and peroxisome pro-liferator-activated receptor alpha in hepatocytes. J. Biol. Chem. 280, 5258e5266.
Schjerling, C.K., Hummel, R., Hansen, J.R., Borsting, C., Mikkelsen, J.M.,Kristiansen, K., Knudsen, J., 1996. Disruption of the gene encoding the acyl-CoA-binding protein (ACB1) perturbs acyl-CoA metabolism in Saccharomycescerevisiae. J. Biol. Chem. 271, 22514e22521.
Schroeder, F., Huang, H., Hostetler, H.A., Petrescu, A.D., Hertz, R., Bar-Tana, J., Kier, A.B.,2005. Stability of fatty acyl-coenzyme A thioester ligands of hepatocytenuclear factor-4a and peroxisome proliferator-activated receptor-a. Lipids 40,559e568.
Shibasaki, M., Katsura, M., Torigoe, F., Honda, T., Sumimoto, A., Tsujimura, A.,Ohkuma, S., 2007. Increase in diazepam binding inhibitor expression by sus-tained morphine exposure is mediated via mu-opioid receptors in primarycultures of mouse cerebral cortical neurons. J. Neurosci. Res. 85, 2971e2980.
Sieglaff, D.H., Duncan, K.A., Brown, M.R., 2005. Expression of genes encodingproteins involved in ecdysteroidogenesis in the female mosquito, Aedes aegypti.Insect Biochem. Mol. Biol. 35, 471e490.
Simansky, K.J., 1996. Serotonergic control of the organization of feeding and satiety.Behav. Brain Res. 73, 37e42.
Snyder, M.J., Feyereisen, R., 1993. A diazepam binding inhibitor (DBI) homolog fromthe tobacco hornworm, Manduca sexta. Mol. Cell. Endocrinol. 94, R1eR4.
Snyder, M.J., Van Antwerpen, R., 1997. Cellular distribution, levels, and function ofthe diazepam-binding inhibitor/acyl-CoA-binding protein in last instar Man-duca sexta midgut. Cell Tissue Res. 288, 177e184.
Sobreira, T.J.P., Gruber, A., 2008. Sequence-specific reconstruction from fragmentarydatabases using seed sequences: implementation and validation on SAGE,proteome and generic sequencing data. Bioinformatics 24, 1676e1680.
Swinnen, J.V., Alen, P., Heyns, W., Verhoeven, G., 1998. Identification of diazepam-binding inhibitor/acyl-CoA-binding protein as a sterol regulatory element-binding protein-responsive gene. J. Biol. Chem. 273, 19938e19944.
Wang, J.L., Wang, J.X., Zhao, X.F., 2008. Molecular cloning and expression profiles ofthe acyl-CoA-binding protein gene from the cotton bollworm Helicoverpaarmigera. Arch. Insect Biochem. Physiol. 68, 79e88.
Zifa, E., Fillion, G., 1992. 5-Hydroxytryptamine receptors. Pharmacol. Rev. 44,401e458.




















