
Self-Renewing Osteoprogenitors in Bone Marrow Sinusoids Can Organize a Hematopoietic...
39
Self-Renewing Osteoprogenitors in Bone Marrow Sinusoids Can Organize a Hematopoietic Microenvironment Benedetto Sacchetti, 1,3,8 Alessia Funari, 3,4,8 Stefano Michienzi, 1,3 Silvia Di Cesare, 3 Stefania Piersanti, 3 Isabella Saggio, 2,3,5 Enrico Tagliafico, 6 Stefano Ferrari, 6 Pamela Gehron Robey, 7 Mara Riminucci, 3,4 and Paolo Bianco 1,3, * 1 Deparment of Experimental Medicine 2 Department of Genetics & Molecular Biology La Sapienza University, 00161 Rome, Italy 3 Biomedical Science Park San Raffaele, 00128 Rome, Italy 4 Department of Experimental Medicine, University of L’Aquila, 67100 L’Aquila, Italy 5 Institute for Molecular Biology and Pathology, National Research Council, Rome, Italy 6 Department of Biochemistry, University of Modena, 59100 Modena, Italy 7 Craniofacial and Skeletal Diseases Branch, National Institute of Dental and Craniofacial Research, National Institutes of Health, Department of Health and Human Services, Bethesda, MD 20892, USA 8 These authors contributed equally to this work. *Correspondence: p.bianco@flashnet.it DOI 10.1016/j.cell.2007.08.025 SUMMARY The identity of cells that establish the hemato- poietic microenvironment (HME) in human bone marrow (BM), and of clonogenic skeletal progenitors found in BM stroma, has long re- mained elusive. We show that MCAM/CD146- expressing, subendothelial cells in human BM stroma are capable of transferring, upon trans- plantation, the HME to heterotopic sites, coinci- dent with the establishment of identical suben- dothelial cells within a miniature bone organ. Establishment of subendothelial stromal cells in developing heterotopic BM in vivo occurs via specific, dynamic interactions with develop- ing sinusoids. Subendothelial stromal cells re- siding on the sinusoidal wall are major pro- ducers of Angiopoietin-1 (a pivotal molecule of the HSC ‘‘niche’’ involved in vascular remodel- ing). Our data reveal the functional relationships between establishment of the HME in vivo, es- tablishment of skeletal progenitors in BM sinu- soids, and angiogenesis. INTRODUCTION Bone marrow (BM) is the only permanent hematopoietic organ in humans. During organogenesis, hematopoiesis is established through interaction of blood-borne hemato- poietic stem cells (HSCs) with a local stroma of mesenchy- mal lineage established during ossification (reviewed in Bianco and Robey, 2000). The critical role of BM stroma for homing and long-term maintenance of hematopoiesis in mammalian bone was classically demonstrated by transfer of the hematopoietic microenvironment (HME) to an ectopic site upon in vivo transplantation of either BM fragments or BM stromal cells (BMSCs; Friedenstein et al., 1974; Maniatis et al., 1971; Tavassoli and Crosby, 1968). Concurrent formation of heterotopic bone in the same systems first revealed that progenitors of skeletal tissues (bone, cartilage, fibrous tissue, adipocytes) are in- cluded in postnatal BM stroma (Friedenstein, 1990), a no- tion that later evolved into the current concept that skele- tal stem cells (SSCs [Bianco and Robey, 2004], also known as stromal or ‘‘mesenchymal’’ stem cells) coexist with HSCs in BM. However, the identity of cells that estab- lish the heterotopic HME, as well as that of putative SSCs, has remained elusive. Anatomically, BM stroma includes virtually all nonhematopoietic cells types found in the BM microenvironment (osteoblasts, endothelial cells, fi- broblasts, reticular cells [Weiss, 1976]), each of which, in principle, may play a critical role in hematopoiesis. For ex- ample, the establishment of an HSC ‘‘niche’’ (Moore and Lemischka, 2006) in the mouse BM has been ascribed to osteoblasts residing on bone surfaces (Arai et al., 2004; Calvi et al., 2003; Zhang et al., 2003), as well as to endothelial cells lining sinusoids (Kiel et al., 2005), sug- gesting a potential multiplicity of physiologically important microenvironments. Establishment of hematopoiesis within heterotopic os- sicles formed by transplantation of BM stroma reflects the interaction of circulating HSCs and progenitors of the host with a locally established HME. Therefore, trans- plantation of BMSCs represents an appealing model of BM organogenesis in which to identify HME-establishing cells. The value of the system has been limited, however, 324 Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc.
Transcript of Self-Renewing Osteoprogenitors in Bone Marrow Sinusoids Can Organize a Hematopoietic...

Self-Renewing Osteoprogenitors in BoneMarrow Sinusoids Can Organizea Hematopoietic MicroenvironmentBenedetto Sacchetti,1,3,8 Alessia Funari,3,4,8 Stefano Michienzi,1,3 Silvia Di Cesare,3 Stefania Piersanti,3
Isabella Saggio,2,3,5 Enrico Tagliafico,6 Stefano Ferrari,6 Pamela Gehron Robey,7 Mara Riminucci,3,4
and Paolo Bianco1,3,*1Deparment of Experimental Medicine2Department of Genetics & Molecular BiologyLa Sapienza University, 00161 Rome, Italy3Biomedical Science Park San Raffaele, 00128 Rome, Italy4Department of Experimental Medicine, University of L’Aquila, 67100 L’Aquila, Italy5Institute for Molecular Biology and Pathology, National Research Council, Rome, Italy6Department of Biochemistry, University of Modena, 59100 Modena, Italy7Craniofacial and Skeletal Diseases Branch, National Institute of Dental and Craniofacial Research, National Institutes of Health,
Department of Health and Human Services, Bethesda, MD 20892, USA8These authors contributed equally to this work.
*Correspondence: [email protected]
DOI 10.1016/j.cell.2007.08.025
SUMMARY
The identity of cells that establish the hemato-poietic microenvironment (HME) in humanbone marrow (BM), and of clonogenic skeletalprogenitors found in BM stroma, has long re-mained elusive. We show that MCAM/CD146-expressing, subendothelial cells in human BMstroma are capable of transferring, upon trans-plantation, the HME to heterotopic sites, coinci-dent with the establishment of identical suben-dothelial cells within a miniature bone organ.Establishment of subendothelial stromal cellsin developing heterotopic BM in vivo occursvia specific, dynamic interactions with develop-ing sinusoids. Subendothelial stromal cells re-siding on the sinusoidal wall are major pro-ducers of Angiopoietin-1 (a pivotal molecule ofthe HSC ‘‘niche’’ involved in vascular remodel-ing). Our data reveal the functional relationshipsbetween establishment of the HME in vivo, es-tablishment of skeletal progenitors in BM sinu-soids, and angiogenesis.
INTRODUCTION
Bone marrow (BM) is the only permanent hematopoietic
organ in humans. During organogenesis, hematopoiesis
is established through interaction of blood-borne hemato-
poietic stem cells (HSCs) with a local stroma of mesenchy-
mal lineage established during ossification (reviewed in
Bianco and Robey, 2000). The critical role of BM stroma
324 Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc.
for homing and long-term maintenance of hematopoiesis
in mammalian bone was classically demonstrated by
transfer of the hematopoietic microenvironment (HME)
to an ectopic site upon in vivo transplantation of either
BM fragments or BM stromal cells (BMSCs; Friedenstein
et al., 1974; Maniatis et al., 1971; Tavassoli and Crosby,
1968). Concurrent formation of heterotopic bone in the
same systems first revealed that progenitors of skeletal
tissues (bone, cartilage, fibrous tissue, adipocytes) are in-
cluded in postnatal BM stroma (Friedenstein, 1990), a no-
tion that later evolved into the current concept that skele-
tal stem cells (SSCs [Bianco and Robey, 2004], also
known as stromal or ‘‘mesenchymal’’ stem cells) coexist
with HSCs in BM. However, the identity of cells that estab-
lish the heterotopic HME, as well as that of putative SSCs,
has remained elusive. Anatomically, BM stroma includes
virtually all nonhematopoietic cells types found in the
BM microenvironment (osteoblasts, endothelial cells, fi-
broblasts, reticular cells [Weiss, 1976]), each of which, in
principle, may play a critical role in hematopoiesis. For ex-
ample, the establishment of an HSC ‘‘niche’’ (Moore and
Lemischka, 2006) in the mouse BM has been ascribed
to osteoblasts residing on bone surfaces (Arai et al.,
2004; Calvi et al., 2003; Zhang et al., 2003), as well as to
endothelial cells lining sinusoids (Kiel et al., 2005), sug-
gesting a potential multiplicity of physiologically important
microenvironments.
Establishment of hematopoiesis within heterotopic os-
sicles formed by transplantation of BM stroma reflects
the interaction of circulating HSCs and progenitors of
the host with a locally established HME. Therefore, trans-
plantation of BMSCs represents an appealing model of
BM organogenesis in which to identify HME-establishing
cells. The value of the system has been limited, however,

by lack of markers suitable for visualization of specific
stromal cell types other than mature osteoblasts. Whereas
the donor origin of mature osteoblasts and the host origin
of hematopoietic cells colonizing the ‘‘ossicle’’ have been
directly proven (Krebsbach et al., 1997; Kuznetsov et al.,
1997), the host origin of the endothelial cells has
only been surmised, and donor or host origin of stromal
cells proper (e.g., reticular cells) has never been demon-
strated.
Similar limitations have prevented definition of the pre-
cise identity and properties of clonogenic progenitors
found in BM stroma. Whereas the multipotency of a frac-
tion of human skeletal progenitors has been demonstrated
(Kuznetsov et al., 1997), the ability to self-renew has not
been formally determined for any subset of stromal cells,
detracting from the very claim of their ‘‘stemness’’ and
at odds with the general popularity gained by the concept
of ‘‘mesenchymal’’ stem cells. Self-renewal implies re-
constitution of the same compartment of phenotypically
defined progenitor cells in vivo. As applied to stromal pro-
genitor cells, this postulates that (1) the identity/pheno-
type of explanted progenitor cells is defined and (2) cells
with identical phenotype and properties are demonstrated
within the tissues formed de novo following in vivo trans-
plantation. Whereas multiple markers expressed in clono-
genic stromal cells from human BM have been proposed
(Barry et al., 1999; Deschaseaux and Charbord, 2000;
Gronthos et al., 1999; Shi and Gronthos, 2003; Simmons
and Torok-Storb, 1991; Vogel et al., 2003; Zannettino
et al., 2003), none of them have been used (and most are
not suitable) for in situ identification of cells prior to expla-
nation and after in vivo transplantation.
In this study, we show that a specific phenotype de-
fines stromal progenitors in human BM that regenerate
bone and stroma and establish the HME in vivo. Marked
by high expression of melanoma-associated cell adhe-
sion molecule, MCAM/CD146 (Shih, 1999), this pheno-
type is shared by subendothelial cells of BM sinusoids
in situ and is not duplicated by osteoblastic cells capable
of forming heterotopic bone, but not an HME, in vivo. By
following the fate of transplanted, human CD146+ cells
during organogenesis of heterotopic BM, we document
their stepwise regeneration into CD146+ subendothelial
cells in de novo formed BM, from which human
CD146+ clonogenic cells can ultimately be isolated in
culture. Our data suggest self-renewal of CD146+ osteo-
progenitors in vivo as an integral part of angiogenic
events whereby sinusoids are established in vivo prior
to hematopoiesis. Consistent with their nature as suben-
dothelial cells, CD146+ osteoprogenitors, but not their
differentiated osteoblastic progeny, express Angiopoie-
tin-1 (Ang-1), a pivotal regulator both of vascular remod-
eling (Suri et al., 1996) and of the HSC niche (Arai et al.,
2004). Our data anatomically and functionally identify
clonogenic skeletal progenitors with the capacity to
self-renew, reveal their identity as cells that transfer the
HME in vivo, and link their establishment and regenera-
tion in BM to angiogenesis.
RESULTS
BMSCs, But Not Other Osteogenic Cell Strains,Transfer the HME In VivoThe ability to establish an HME was evaluated by trans-
plantation of cell strains derived from BMSCs from normal
hematopoietic BM, human trabecular bone cells (HTBs),
periosteal cells (PCs), fibrotic bone marrow of fibrous dys-
plasia of bone (FD; OMIM#174800), and as controls, mus-
cle fibroblasts (MFs) and skin fibroblasts (SFs). By in vitro
differentiation assays commonly used to characterize
‘‘mesenchymal’’ stem cells, all strains (except SFs) gener-
ated adipocyte-like cells or ‘‘mineralization nodules’’ (data
not shown). When transplanted subcutaneously into im-
munocompromised mice, control cell strains (MFs and
SFs) failed to generate either bone or BM (12/12 trans-
plants), while bone-derived cell strains (HTBs and PCs)
generated bone but no heterotopic BM (12/12 transplants
each strain). As previously reported (Bianco et al., 1998),
FD cells, derived from fibrotic, nonhematopoietic BM,
also formed bone but not BM (data not shown). Only
BMSCs derived from hematopoietic BM established
both bone and hematopoietic tissue (12/12 transplants)
(Figure 1A).
CD146 Expression Distinguishes BMSCsfrom Other Osteogenic StrainsCharacterization of the same cell strains using a panel of
markers (Table S1), including putative markers of ‘‘mesen-
chymal’’ stem cells (CD49a, CD63, CD90, CD105,
CD140b, CD146, STRO-1, and alkaline phosphatase
[ALP]), revealed that high levels of CD63 distinguished
cell strains endowed with osteogenic potential in vivo
(BMSCs, HTBs, PCs, and FD cells) from nonosteogenic
MFs and SFs. Only a characteristic ‘‘high/bright’’ expres-
sion of CD146 distinguished BMSCs from all other (osteo-
genic and nonosteogenic) strains, which, in contrast, were
all noted for a ‘‘low/dim’’ expression of CD146 (Figures 1B
and S1A).
All BM Colony Forming Unit-Fibroblasts and TheirClonal Progeny, But Not All BM ‘‘Stromal’’ Cells,Express CD146Primary cultures of BMSCs were established either as
nonclonal (high density, > 1.6 3 105 cells/cm2) cultures
or by plating BM cell suspensions at clonal density in order
to obtain discrete colonies (colony forming unit-fibroblast
[CFU-F] cultures) (Friedenstein, 1980). Randomly selected
colonies (n = 22) were individually harvested from primary
CFU-F cultures from two donors. Multiclonal strains (n = 3)
were generated by pooling all primary colonies obtained in
CFU-F cultures from three donors (29, 13, and 11 colo-
nies, respectively). CD146 was expressed at high levels
in �99% of cells in 22/22 clones and in 3/3 multiclonal
strains (Figure 2A). In contrast, only �30% of the cells
were CD146+ in 3 nonclonal cultures (Figures 2A and
S1B). FACS analysis revealed that high numbers of
CD146�CD105+ or CD146�ALP+ cells (putatively mature
Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc. 325

osteoblastic cells) were included in nonclonal cultures
(data not shown). Hence, establishing CFU-F cultures
from unfractionated BM, while selecting for clonogenic
cells, also selected for CD146+ cells at the same time.
Figure 1. High Expression of CD146 Distinguishes BMSC,
Capable of Establishing the HME, from Other Osteogenic or
Nonosteogenic Cell Populations
(A) Histology of transplants of human skin fibroblasts (SFs), periosteal
cells (PCs), human trabecular bone cells (HTBs), and bone marrow
stromal cells (BMSCs) harvested at 8 weeks. Neither bone nor BM
formed in transplants of SFs. Bone, but not marrow, formed in trans-
plants of PCs and HTBs. Both bone and marrow formed in transplants
of BMSCs. hac, hydroxyapatite carrier; bm, bone marrow; ft, fibrous
tissue, H&E.
(B) FACS analysis of ‘‘mesenchymal stem cell’’ markers in cultures of
SFs, MFs, HTBs, PCs, FD cells, and BMSCs grown under identical
conditions up to passage 3. CD146high/bright expression distinguishes
BMSCs from all other strains (CD146low/dim). No other single marker
is expressed differentially in BMSCs compared to all other strains.
326 Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc.
These cells initiate the CFU-F cultures from which
CD146high/bright BMSC strains are generated.
All CFU-Fs Are Found, and Most Are Recovered,in the CD45�CD146+ Fraction of BMNCsBy FACS analysis, CD146+ cells accounted for 0.11% ±
0.02% of total human BM nucleated cells (BMNCs). Con-
sistent with the known expression of CD146 in certain he-
matopoietic cells (e.g., T cells; Pickl et al., 1997),�20% of
CD146+ cells coexpressed CD45 (not shown). CD45�
cells within the BMNC suspensions were therefore mag-
netically separated prior to immunoselection of CD146+
cells. CD146+ cells were enriched�10-fold (1.2% ± 0.9%)
in the CD45� fraction compared to BMNCs. By dual-label
FACS analysis (Figure 2B), CD45�CD146+ cells coex-
pressed ALP (78%–85%) and CD105 (>95%). In contrast,
only �2% of CD45�CD146+ cells coexpressed CD34,
suggesting that >95% of CD45�CD146+ cells in human
BM are not endothelial cells (ECs). CD45� cells were sep-
arated into CD146+ and CD146� fractions by FACS, and
colony forming efficiency (CFE) assays were conducted
on both fractions (Figure 2C, Tables S2 and S3). An aver-
age of 2.6 ± 0.8 CFU-Fs/102 cells were observed in the
CD146+ fraction, which translates into an enrichment of
8.3 3 102 compared to BMNCs and a recovery of �80%
of the total CFU-Fs found in unfractionated BM (Table
S3). No colonies were formed by CD45�CD146� cells
plated at the same or higher density (Figure 2C, Tables
S2 and S3). When sorted and subjected to CFE assays,
total CD34+ cells and the CD146+CD34+ fraction failed to
generate fibroblastic colonies (Table S4).
Adventitial Reticular Cells Are the In SituCounterpart of CD146+ CFU-Fs in Human BMIn human BM in situ, no labeling for CD146 was observed
in erythroid or myeloid cells, megakaryocytes, endothe-
lial cells, adipocytes, osteoblasts, osteocytes, or endos-
teal cells. Within hematopoietic tissue, CD146 labeling
was restricted to adventitial reticular cells (ARCs; Bianco
and Boyde, 1993; Tavassoli and Friedenstein, 1983;
Weiss, 1976; Westen and Bainton, 1979). ARCs formed
a subendothelial (adventitial) layer in sinusoidal walls,
and projected reticular processes that associated with
hematopoietic cells, away from sinusoidal walls
(Figure 2D). Like the CD146+ CFU-Fs from human BM,
CD146+ ARCs coexpressed endoglin (CD105) and ALP
(both of which were also expressed by additional cell
types in BM) in situ, but not CD34, CD31, or CD45 (not
shown).
Cultured CD146+ Cells Display Mural CellPropertiesUndifferentiated CD146+ BMSCs expressed transcripts
of early osteogenic progenitors but not of mature osteo-
blasts. Of note, HSC niche-related transcripts (Jagged-1,
N-cadherin, CXCL12 [Sugiyama et al., 2006], and SCF
[Duncan et al., 2005; Nagasawa et al., 1996; Zhang et al.,
2003]) were highly expressed (Figure S2). Consistent with

Figure 2. CD146+ Stromal Cells Are the CFU-Fs and Derive from Adventitial Reticular Cells(A) FACS analyses of randomly selected clones, formed in primary CFU-F cultures (4 clones representative of 22). CD146 is highly expressed in vir-
tually all cells in each clone. Representative FACS analyses of multiclonal (29 pooled colonies) and nonclonal primary cultures of BMSCs. Whereas
CD146 is highly expressed in virtually all cells in a multiclonal strain, only �30% of cells are CD146+ in a nonclonal primary culture.
(B) Dual-label FACS analyses of magnetically separated CD45�marrow cells. CD45�CD146+ cells coexpress CD105 (�99%) and alkaline phospha-
tase (ALP; �80%). Only a minor fraction coexpresses CD34.
(C) CD146+ and CD146� fractions within CD45� cells were separated by FACS. One hundred cells were plated in 100 mm dishes (1.6 cells/cm2).
Clonogenic cells were found only in the CD45�CD146+ fraction.
(D) Sections of human bone marrow immunolabeled for CD146 counterstained with hematoxylin. Labeling is restricted to adventitial cells of sinusoids
(double arrows), extending ‘‘reticular’’ processes (single arrows) among hematopoietic cells.
their origin from ARCs, cultured CD146+ cells failed to ex-
press endothelial markers or to differentiate into ECs when
exposed to specific conditions. In contrast, they ex-
pressed a host of markers of subendothelial cells (mural
cells/pericytes, MC [Jain, 2003], such as a-smooth muscle
actin, NG2, calponin 1 and 3, PDGFRb; Figure S2). Factors
known to regulate mural cell growth and phenotype dur-
ing the maturation phase of angiogenesis (Hirschi and
D’Amore, 1996; Jain, 2003), such as FGF-2 and TGF-b1,
modulated the mural cell phenotype and the proliferation
of CD146+ cells, in an opposite fashion (Figure S3).
Whereas FGF-2 stimulated the proliferation of CD146+
cells, TGF-b1 inhibited it, consistent with the effects of
these factors on mural cells (Hirschi and D’Amore, 1996;
Jain, 2003). When cocultured with endothelial cells,
CD146+ stromal cells behaved like mural cells; i.e., they
directed the assembly and timed remodeling of pseudo-
vascular structures in vitro, in sharp contrast with con-
trol, nondescript fibroblasts or unrelated cell types
(Figure S4).
Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc. 327
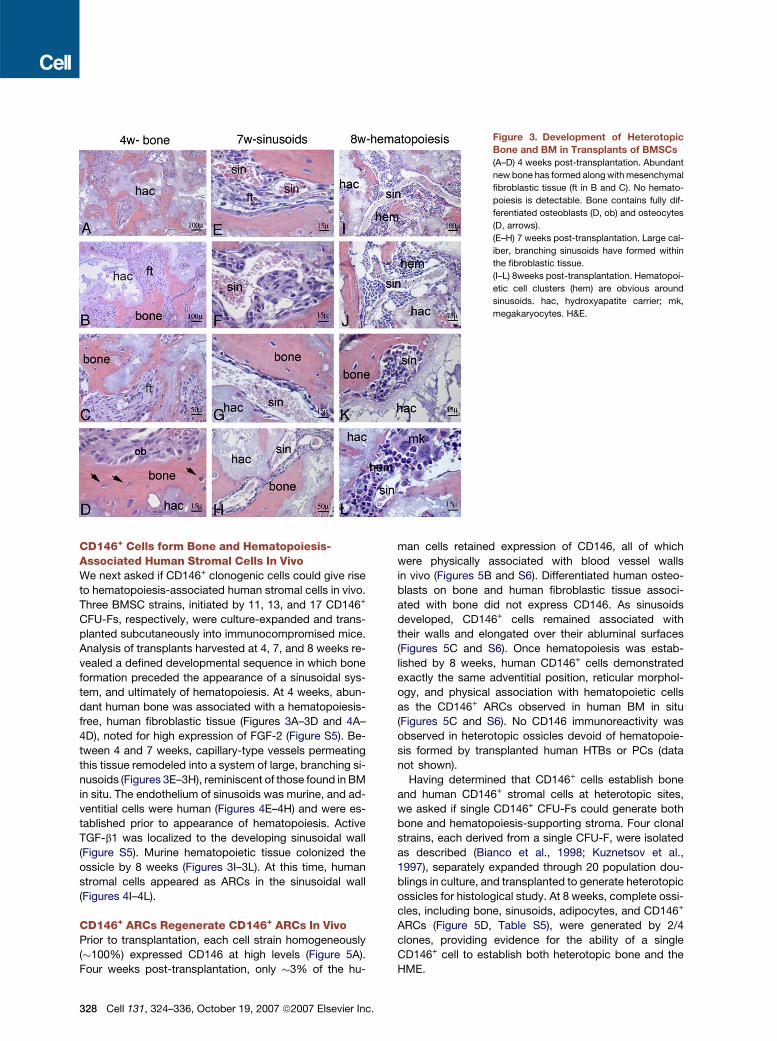
Figure 3. Development of Heterotopic
Bone and BM in Transplants of BMSCs
(A–D) 4 weeks post-transplantation. Abundant
new bone has formed along with mesenchymal
fibroblastic tissue (ft in B and C). No hemato-
poiesis is detectable. Bone contains fully dif-
ferentiated osteoblasts (D, ob) and osteocytes
(D, arrows).
(E–H) 7 weeks post-transplantation. Large cal-
iber, branching sinusoids have formed within
the fibroblastic tissue.
(I–L) 8weeks post-transplantation. Hematopoi-
etic cell clusters (hem) are obvious around
sinusoids. hac, hydroxyapatite carrier; mk,
megakaryocytes. H&E.
CD146+ Cells form Bone and Hematopoiesis-Associated Human Stromal Cells In VivoWe next asked if CD146+ clonogenic cells could give rise
to hematopoiesis-associated human stromal cells in vivo.
Three BMSC strains, initiated by 11, 13, and 17 CD146+
CFU-Fs, respectively, were culture-expanded and trans-
planted subcutaneously into immunocompromised mice.
Analysis of transplants harvested at 4, 7, and 8 weeks re-
vealed a defined developmental sequence in which bone
formation preceded the appearance of a sinusoidal sys-
tem, and ultimately of hematopoiesis. At 4 weeks, abun-
dant human bone was associated with a hematopoiesis-
free, human fibroblastic tissue (Figures 3A–3D and 4A–
4D), noted for high expression of FGF-2 (Figure S5). Be-
tween 4 and 7 weeks, capillary-type vessels permeating
this tissue remodeled into a system of large, branching si-
nusoids (Figures 3E–3H), reminiscent of those found in BM
in situ. The endothelium of sinusoids was murine, and ad-
ventitial cells were human (Figures 4E–4H) and were es-
tablished prior to appearance of hematopoiesis. Active
TGF-b1 was localized to the developing sinusoidal wall
(Figure S5). Murine hematopoietic tissue colonized the
ossicle by 8 weeks (Figures 3I–3L). At this time, human
stromal cells appeared as ARCs in the sinusoidal wall
(Figures 4I–4L).
CD146+ ARCs Regenerate CD146+ ARCs In VivoPrior to transplantation, each cell strain homogeneously
(�100%) expressed CD146 at high levels (Figure 5A).
Four weeks post-transplantation, only �3% of the hu-
328 Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc.
man cells retained expression of CD146, all of which
were physically associated with blood vessel walls
in vivo (Figures 5B and S6). Differentiated human osteo-
blasts on bone and human fibroblastic tissue associ-
ated with bone did not express CD146. As sinusoids
developed, CD146+ cells remained associated with
their walls and elongated over their abluminal surfaces
(Figures 5C and S6). Once hematopoiesis was estab-
lished by 8 weeks, human CD146+ cells demonstrated
exactly the same adventitial position, reticular morphol-
ogy, and physical association with hematopoietic cells
as the CD146+ ARCs observed in human BM in situ
(Figures 5C and S6). No CD146 immunoreactivity was
observed in heterotopic ossicles devoid of hematopoie-
sis formed by transplanted human HTBs or PCs (data
not shown).
Having determined that CD146+ cells establish bone
and human CD146+ stromal cells at heterotopic sites,
we asked if single CD146+ CFU-Fs could generate both
bone and hematopoiesis-supporting stroma. Four clonal
strains, each derived from a single CFU-F, were isolated
as described (Bianco et al., 1998; Kuznetsov et al.,
1997), separately expanded through 20 population dou-
blings in culture, and transplanted to generate heterotopic
ossicles for histological study. At 8 weeks, complete ossi-
cles, including bone, sinusoids, adipocytes, and CD146+
ARCs (Figure 5D, Table S5), were generated by 2/4
clones, providing evidence for the ability of a single
CD146+ cell to establish both heterotopic bone and the
HME.

Figure 4. Development of Human Stro-
mal Cells in the Chimeric Heterotopic
BM—Immunolabeling for Human Mito-
chondria
(A–D) 4 weeks post-transplantation. The fibro-
blastic tissue (ft), bone-forming osteoblasts on
bone surfaces (D, ob), and osteocytes within
newly formed bone (D, arrows) are human.
(E–H) 7 weeks post-transplantation. ECs lining
sinusoids (s) are murine (F–H, small arrows),
whereas adventitial cells are human (F–H, large
arrows).
(I–L) 8 weeks post-transplantation. Osteocytes
within bone are human. Hematopoietic cells in
newly formed marrow spaces (hem) and ECs
lining sinusoids (J–K, small arrows) are murine.
Stromal cells at the abluminal side of sinusoids,
or interspersed among hematopoietic cells,
are human (J–L, large arrows), as are bone-lin-
ing cells (L, double small arrow). hac, hydroxy-
apatite carrier.
Secondary Passage of CD146+ CFU-FsHaving shown that CD146+ cells could give rise to hetero-
topic stromal cells with anatomy and phenotype identical
to those of the originally explanted cells, we sought evi-
dence that CFU-Fs could be secondarily passaged as fur-
ther indication of the self-renewal capacity of CD146+
stromal cells. Additional transplants were generated with
cell strains originating from a limited number of CFU-Fs
(10 and 12, respectively, in two experiments). At 8 weeks,
transplants were harvested and collagenase-digested to
generate single-cell suspensions. Total human cells
were magnetically sorted, either after short-term culture
(Figure 6A) or directly from the fresh cell suspension
(Figure 6B), based on hCD44 expression, which allowed
for recovery of �2 3 104 and �1.5 3 104 human cells, re-
spectively (Table S6). Aliquots of 2 3 103 and 5 3 103 cells
were plated at clonal density. One and six discrete colo-
nies were observed and harvested at 2 weeks, which
translates into an estimated number of secondary CFU-
Fs of 10 and 18, respectively, in the original cell suspen-
sions (Table S6). FACS analysis showed homogeneous,
high expression of CD146 in all colonies (Figures 6A and
6B).
In separate experiments (Figures 6C and 6D), trans-
plants generated by five clonal strains, each of which
was individually expanded from single CFU-Fs, were
similarly collagenase-digested to generate cell suspen-
sions. Greater than 1 3 106 total cells were obtained in
2/5 transplants generated by the progeny of a single
CFU-F. A 2–5 3 105 aliquot from each of the five cell sus-
pensions liberated by collagenase was used for FACS
analysis and revealed a frequency of hCD146+ cells of
0.09%–0.4% (Table S6). From the two collagenase-re-
leased populations with the largest numbers of cells,
the remaining aliquots were magnetically sorted directly
for human CD146+ cells, resulting in �2.1 3 103 and
�4.0 3 103 cells, and plated in culture at a density of
1.6 cells/cm2. This resulted in the generation of 3 and 2
secondary CFU-Fs, which translates into an estimated
total number of 4 and 3 assayable CFU-Fs from the orig-
inal total cell suspension, respectively. No colonies were
generated from CD146� cells, plated at the same or
higher (160 cells/cm2) density. All of the colonies gener-
ated by the secondary CFU-Fs expressed CD146 at
high levels (Figures 6C and 6D). Thus, heterotopic ossi-
cles generated either by multiple CFU-Fs or by single
CFU-Fs contained a number of assayable CD146+
CFU-Fs similar to or greater than the number of originally
explanted, culture-initiating CD146+ CFU-Fs.
Regulated Production of Ang-1by CD146+ Stromal CellsAng-1 (a ligand of the Tie-2 tyrosine kinase receptor spe-
cifically expressed in ECs and HSCs [Davis et al., 1996;
Dumont et al., 1992; Hsu et al., 2000; Iwama et al., 1993;
Sato et al., 1993]) is produced by MCs in development
Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc. 329

Figure 5. Self-Renewal of CD146+ Cells In Vivo
(A) FACS analysis prior to transplantation. Homogeneous expression of high levels of CD146.
(B) Serial sections of transplants of the same strain at 4 weeks, stained for CD146 and human mitochondria, respectively. The vast majority of human
cells in the ossicle, including osteoblasts (red arrows) are CD146�. CD146+ cells are restricted to an adventitial layer in vessel walls.
(C) Development of CD146+ ARCs in the heterotopic BM. At 4 weeks, small mononuclear CD146+ cells associate with blood vessels. At 7 weeks,
elongated CD146+ cells are found over and around sinusoids. At 8 weeks, reticular processes of CD146+ cells establish contacts with hematopoietic
cells (h). mk, megakaryocyte; blue arrow, endothelium; red arrows, hematopoietic cells between endothelial and CD146+ cells.
(D) Transplantation of a single CFU-F derived cell strain. At 14 days, the colony formed by a single CFU-F was isolated with a cloning cylinder and
individually expanded. Bottom left, FACS analysis of the expanded clone at passage 3, demonstrating the homogeneous expression of CD146
(and the coexpression of ALP in a subset of the clonal population). Top right, histology of the heterotopic ossicle (8 weeks) formed by the expanded
clonal population. Bone, sinusoids (sin), adipocytes (ad), and hematopoiesis (hem) are shown. Bottom right, human CD146+ adventitial reticular cells
in the same heterotopic ossicle.
and regulates microvessel assembly and remodeling in
mice (Suri et al., 1996). Reported to be produced by oste-
oblasts, Ang-1 has also been implicated as a key compo-
nent of the HSC niche in postnatal murine bone (Arai et al.,
2004). Since our data implicated CD146+ cells in estab-
lishment of both the sinusoidal wall structure and the
HME in vivo, we investigated Ang-1 expression. In human
BM in situ, Ang-1 immunoreactivity was restricted to
ARCs with no signal over bone surfaces where differenti-
ated osteoblasts reside (Figure 7A). In the heterotopic os-
sicles prior to the establishment of hematopoiesis, Ang-1
was localized to mesenchymal cells around sinusoids but
330 Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc.
not to osteoblasts (Figure 7B). Once hematopoiesis was
established, ARCs adjacent to sinusoids expressed Ang-
1, whereas Tie-2 was localized to sinusoidal endothelium
and to rare, small cells of hematopoietic habit, adjacent to
the sinusoidal wall (Figure 7C). Undifferentiated CD146+
BMSCs produced abundant Ang-1 in vitro but did not ex-
press Tie-2. When BMSCs were induced to differentiate
into osteoblasts in vitro, both Ang-1 and CD146 expres-
sion were potently downregulated (Figures 7D–7G), in
agreement with our in vivo data. Interestingly, gene knock-
down of either CD146 or Ang-1 in CD146+ stromal cells
significantly interfered with their ability to direct the

Figure 6. Secondary Passage of CD146+ CFU-Fs
CD146+ CFU-Fs were recovered from heterotopic ossicles generated by transplanting the progeny of either a limited number of CFU-Fs (A, n = 10; B,
n = 12) or a single CFU-F (C and D). In (A) and (B), hCD90+/hCD44+ cells were sorted after short-term culture (A) or directly from the collagenase-
generated cell suspension (B). All colonies obtained by plating sorted cells at clonal density were homogeneously CD146high. In (C) and (D), CD146+
cells were sorted directly from the cell suspension obtained from single CFU-F-generated ossicles. All colonies (C, 3 colonies; D, 2 colonies) obtained
by replating the sorted cells at clonal density expressed CD146 at high levels. No colonies were obtained from the CD146� population (not shown).
assembly and remodeling of pseudovascular structures in
vitro (Figure S7), suggesting that both CD146 and Ang-1
expressed in stromal cells may participate in stromal-
endothelial interactions. Both CD146 and Ang-1 were also
downregulated by FGF-2 (Figure S8) in the context of the
complex effects exerted by this factor on the growth and
differentiation of CD146+ cells in culture. Notably, in trans-
plants generated with FGF-2-treated CD146+ cells, the
ability of the latter to establish CD146+ stromal cells and
the HME in vivo was abrogated, while their ability to
form bone remained unscathed (Figure S8), indicating
that the two abilities can be dissociated from one another
in a single-cell population otherwise competent to both
functions.
DISCUSSION
The notion that BM includes skeletal progenitor (stem)
cells and the notion that BM stroma provides cues for
homing, maintenance, proliferation, and maturation of
hematopoietic progenitors both emanate from the same
classical transplantation experiments. The multipotency
of at least a subset of CFU-Fs supports the view that a sec-
ond type of stem cell (skeletal [Bianco and Robey, 2004],
stromal [Owen and Friedenstein, 1988], or ‘‘mesenchy-
mal’’ [Caplan, 1991]) coexists with HSCs in BM. Due to the
lack of markers suited to bridge the gap between in situ,
ex vivo, and in vivo observations, the in situ counterpart
of CFU-Fs has previously remained unknown, and the
Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc. 331

Figure 7. Expression of Ang-1 and Tie-2
(A) Ang-1 immunoreactivity in sections of human BM is restricted to ARCs (arrows) and absent over bone surfaces. ad, adipocytes.
(B) Localization of Ang-1 to ARCs in heterotopic ossicles. Prior to establishment of hematopoiesis (4 weeks post-transplantation), Ang-1 immunore-
active-mesenchymal cells reside in presumptive marrow spaces in the vicinity of developing sinusoids. After establishment of hematopoiesis, retic-
ular cells in the hematopoietic space and in the vicinity of sinusoids express Ang-1. Osteoblasts are not labeled. ob, osteoblasts; hac, hydroxyapatite
carrier; sin, sinusoid; arrows, ARCs.
(C) Expression of Tie-2 in heterotopic ossicles is restricted to sinusoidal endothelium (blue arrows) and to rare small mononuclear cells adjacent to the
luminal or abluminal side of endothelium (black arrows).
(D and E) Expression and regulation of CD146 and Ang-1 in CD146+ cells induced to differentiate into osteoblasts in vitro. PD, pre-differentiation; M,
mineralization conditions. (D) Matrix mineralization. (E) Upregulation of markers of mature osteoblasts (BSP, osteocalcin) and downregulation of
CD146 and Ang-1 mRNAs (qPCR, fold change over PD). Error bars indicate ± standard deviation (SD).
332 Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc.

very self-renewal of stromal progenitors, a defining char-
acteristic of stem cells, has not been formally demon-
strated. Expression of high levels of CD146, a cell adhesion
molecule of the immunoglobulin superfamily expressed
in a restricted range of normal cells (Shih, 1999), identifies
all ex vivo assayable CFU-Fs, and a specific subset of stro-
mal cells in situ. Explantable CFU-Fs exhibit the same
phenotype as adventitial reticular cells (ARCs), which re-
side in bone marrow sinusoids next to the endothelial
layer, strongly indicating that ARCs are in fact the cells
explanted ex vivo as CFU-Fs. We have now shown that
following transplantation of CD146+ stromal cells, a small
subset retain CD146 expression, dynamically associate
with developing sinusoids, and eventually regenerate het-
erotopic human cells with the anatomy and phenotype of
ARCs.
We have also shown that transplantation of cell popula-
tions derived from either a limited number of CD146+ CFU-
Fs or single CD146+ CFU-Fs results in the re-establish-
ment, in the heterotopic ossicles, of CD146+ CFU-Fs
that can be secondarily passaged and directly assayed.
By providing evidence for the ability of CD146+ stromal
cells to function as self-renewing, clonogenic skeletal pro-
genitors, our data outline the long sought anatomical iden-
tity of SSCs (‘‘mesenchymal’’ stem cells) in human BM and
a crucial feature of their phenotype. While our data provide
evidence for the self-renewal and multipotency of CD146+
CFU-Fs, a larger-scale study would be necessary to accu-
rately determine the actual frequency of in vivo assayable,
multipotent, and self-renewing clonogenic progenitors
within our population of phenotype-defined cells. Even
though such frequency would appear to be high based
on our data (50%), the relative weight of inherent biologi-
cal heterogeneity, versus heterogeneity relative to the
specific experimental assay and its constraints (including
culture and transplantation conditions), remains to be
further analyzed.
As portrayed in our system, self-renewal of adventitial
reticular cells originally explanted as CD146+ CFU-Fs is in-
scribed into dynamic organogenic events, which are part
of the stepwise reconstitution, in vivo of the HME. The es-
tablishment of subendothelial ARCs at heterotopic mar-
row sinusoids involves the interaction of transplanted cells
with host endothelial cells and the remodeling and matu-
ration of sinusoidal vessels. A lead to the identification of
mechanisms dictating the establishment of skeletal pro-
genitors in BM during organogenesis, and their regenera-
tion in our model, can be found in this context. In the mat-
uration phase of angiogenesis, the mitotic quiescence of
the perivascular mesenchymal cells that are recruited to
a subendothelial mural cell fate is induced via direct inter-
action with endothelial cells and may be mediated by TGF-
b1, locally activated at the interface of endothelial and
subendothelial cells (Antonelli-Orlidge et al., 1989; Jain,
2003; Sato and Rifkin, 1989). Conceivably, the establish-
ment of quiescent skeletal progenitors at the sinusoidal
wall during organogenesis may depend on mechanisms
similar to those establishing mitotically quiescent mural
cells in other tissues, a view that is consistent with our
in vivo and ex vivo data.
How the ability to generate differentiated bone-forming
cells and bone tissue relates to the ability to support he-
matopoiesis (both shared by the BM stroma as a whole)
has long remained elusive. Recent data suggest that oste-
oblasts (differentiated bone-forming cells residing on
bone surfaces) directly maintain a niche for HSCs (Calvi
et al., 2003; Moore and Lemischka, 2006; Zhang et al.,
2003), a view that would easily account for the fact that
transplantation of BM stroma leads to the formation of he-
matopoiesis-accommodating bone. In search for the spe-
cific cell type in BM stroma that is capable of establishing
the HME at heterotopic sites, however, we have shown
that this ability is not synonymous with the ability to gener-
ate differentiated osteoblasts and bone tissue in vivo.
Cell strains originating from CD146high/bright clonogenic
progenitors in BM form bone and transfer the HME in vivo.
CD146low/dim cell strains originating from specific anatom-
ical compartments of bone other than BM space (trabec-
ular bone or periosteum) do establish differentiated oste-
oblasts and bone upon in vivo transplantation but do not
transfer the HME. Furthermore, in a cell population com-
petent to form bone and establish the HME in vivo, the
two functions can be experimentally dissociated from
one another, as seen, for example, as the effect of FGF-
2 treatment in our data.
Establishment of the HME at heterotopic sites occurs
via a defined developmental sequence in which bone for-
mation regularly precedes the appearance of a heterotopic
bone marrow stroma and ultimately of heterotopic hema-
topoiesis. Whereas the establishment of osteoblasts and
bone may be necessary as part of this developmental se-
quence, additional events precede the establishment of
hematopoiesis within bone at heterotopic sites: the re-
modeling of the local vasculature into a sinusoidal system
and the establishment of a CD146+ stromal population at
the sinusoidal wall. Although these events involve the in-
teraction of different cell types, including both donor-
and host-derived (endothelial) cells in our system, trans-
planted CD146+ stromal progenitors critically contribute
to their unfolding in vivo. Of note, neither a sinusoidal sys-
tem nor a local population of human CD146+ stromal cells
are established in vivo when human CD146� cell popula-
tions are transplanted that are competent to generate
bone and osteoblasts in vivo but unable to transfer the
HME.
The link between establishment of subendothelial cells
in BM and establishment of the HME finds in Ang-1 (Davis
et al., 1996) an important molecular correlate. As the
(F) Western analysis demonstrating depletion of CD146 and Ang-1 protein and ELISA demonstrating depletion of secreted Ang-1. m, medium; ID,
immunodepleted medium; IP, immunoprecipitated medium.
(G) Tie-2 expression in HUVEC and lack of Tie-2 expression in BMSCs and BMSCs exposed to endothelial differentiation medium (BMSC-E).
Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc. 333

ligand of the Tie-2 receptor that is specifically expressed
in ECs and HSCs, Ang-1 plays pivotal roles both in angio-
genesis and hematopoiesis. Distinct from the growth-pro-
moting effects of VEGF, the role of Ang-1 in angiogenesis
is specifically related to establishment of MCs and to re-
modeling of vascular plexuses (Suri et al., 1996). Consid-
ering that in development Ang-1 is both a product and
a regulator of pericytes/mural cells, Ang-1 expression in
BM CD146+ cells is consistent with their overall ‘‘mural’’
cell phenotype and subendothelial position. We have
shown that in human BM and in heterotopic ossicles in
vivo, human stromal cells are major producers of Ang-1,
and Ang-1 production is regulated in vitro when stromal
cells are induced to differentiate into osteoblasts or ex-
posed to angiogenesis-regulating factors, such as FGF-
2, that act in vivo on the peri-endothelial mesenchyme.
Vascular remodeling is a significant component of BM or-
ganogenesis. This process establishes a unique system of
large, slow flow sinusoids conducive for bidirectional cell
traffic between BM and peripheral blood. As portrayed
in our in vivo transplantation system, timed remodeling
of capillaries into sinusoids, before the establishment of
hematopoiesis, is coupled to the physical association of
CD146+ subendothelial cells with nascent or growing ves-
sels. Ang-1 may contribute to sinusoid remodeling in post-
natal BM when systemically delivered (Hattori et al., 2001),
and we have observed that BMSC-directed patterning and
remodeling of pseudovascular structures in vitro is altered
by Ang-1 (and CD146) gene silencing. This suggests that
Ang-1 (and CD146), expressed locally in BMSCs, may be
part of the molecular machinery regulating vascular remod-
eling through a local interaction of endothelial and suben-
dothelial cells, which may contribute to the organization
of the unique vascular structure of the bone marrow.
Current evidence suggests that Ang-1 also directly con-
tributes to HSC regulation (Arai et al., 2004) by interacting
with HSC-expressed Tie-2. A body of evidence also sug-
gests that endosteal (Calvi et al., 2003; Zhang et al., 2003)
and sinusoidal surfaces (Kiel et al., 2005), but also
CXCL12-expressing ‘‘reticular’’ cells within the hemato-
poietic space (Sugiyama et al., 2006), may represent spe-
cific sites of HSC regulation (‘‘niches’’). Our data show that
CD146+ stromal progenitors indeed physically coincide
with reticular cells and express CXCL12, Ang-1, and mul-
tiple other gene products that have been implicated in
HSC regulation. As osteoblast progenitors, CD146+ stro-
mal cells generate osteoblasts, which form bone and are
regarded as critical components of an endosteal HSC
niche. As (self-renewing) progenitors of sinusoidal adven-
titial reticular cells, CD146+ stromal cells contribute to the
organization, and become an integral part, of the structure
of sinusoidal walls, in the vicinities of which HSCs have
been directly localized (Kiel et al., 2005). Skeletal progen-
itors residing over sinusoids may thus contribute to hema-
topoietic regulation within the BM—either directly at the
sinusoidal wall where they reside as adventitial reticular
cells or through their osteoblastic progeny at endosteal
surfaces. For example, CD146+ subendothelial cells ex-
334 Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc.
pressing HSC regulators such as Ang-1 or CXCL12 would
be strategically positioned to facilitate the homing of blood
borne hematopoietic progenitors to the marrow environ-
ment or to contribute to a steady-state sinusoidal niche
where HSCs can be localized. By generating, or contribut-
ing to, functionally distinct cell types (osteoblasts and
ARCs) and structures (bone surfaces and sinusoidal ablu-
minal surfaces), CD146+ skeletal progenitors play a pivotal
role in the development of the HME, as recapitulated in our
in vivo system, and contribute to establishing and organiz-
ing the very diversity of physiologically important and spa-
tially distinct microenvironments within the BM. Our data
also indicate that properties of CD146+ subendothelial
cells, such as Ang-1 expression, that are relevant to he-
matopoietic regulation may be modulated when skeletal
progenitors are themselves recruited to cell proliferation
or osteogenic differentiation or exposed to vasculogenic
cues. These cellular events are inscribed, in vivo, in funda-
mental organogenic processes such as skeletal growth,
lifelong bone remodeling, and adaptation, to which skele-
tal progenitors critically contribute. Hence, the complex
interplay of osteogenesis and hematopoiesis in develop-
ment, physiology, and disease may be seen as rooted
into a unique functional interplay of two systems of pro-
genitor/stem cells that takes place in the bone marrow
environment at specific sites.
EXPERIMENTAL PROCEDURES
Reagents
Antibodies for cell sorting and flow cytometry are listed in Table S1 and
for immunohistochemistry and western blotting in Table S7. Primers
for RT-PCR are listed in Table S8.
Culture and Characterization of Cell Strains
BMSCs, PCs, HTBs, and stromal cells from the nonhematopoietic
bone marrow of fibrous dysplastic bone (FD) were isolated by estab-
lished methods (Bianco et al., 1998; Kuznetsov et al., 1997; Miura
and O’Driscoll, 1998; Robey and Termine, 1985). Samples were ob-
tained with informed consent per institutionally approved protocols.
Human foreskin fibroblasts (SFs) were from A. Orecchia, IDI, Rome,
Italy. Human muscle fibroblasts (MFs) were from G. Cossu, DIBIT-
HSR, Milan, Italy. After primary culture, all strains were cultured under
identical conditions in a-MEM (Invitrogen)/20% FBS, 100 U/ml penicil-
lin, and 100 mg/ml streptomycin, prior to analysis. Expression of
markers was assessed using a FACSCalibur flow cytometer and Cell-
Quest software (Becton Dickinson Biosciences, San Diego, CA).
In vitro differentiation assays were done by established methods
(Bianco et al., 2006).
Cell Sorting
1 3 107 freshly isolated BMNCs were resuspended in HBSS/30 mM
HEPES (Sigma, St. Louis, MO), 100 U/ml penicillin, 100 mg/ml strepto-
mycin, 1% BSA (Sigma) and incubated on ice for 30 min. Cells were
pelleted in HBSS/2 mM EDTA, 1% BSA, resuspended in 1 ml blocking
buffer, and incubated with anti-CD45 conjugated magnetic beads (Mil-
tenyi Biotec, Auburn, CA) for 20 min on ice. Cells were separated into
CD45� and CD45+ fractions using a MiniMACS magnetic column sep-
aration unit per the manufacturer’s instructions (Miltenyi). CD45� cells
were incubated with PE-conjugated anti-CD146 antibody, and
CD146+ and CD146� cells were separated using a FACS DIVAntageSE
flow cytometer (BD Biosciences Labware, San Diego, CA).

CFE Assays and CFU-F Cultures
CFE was assessed as described (Kuznetsov et al., 1997). Plating den-
sities were 0.1–10 3 103/cm2 for total BMNCs and 1.6–1.6 3 104 cells/
cm2 for separated fractions of BM cells. Colonies (>50 cells) were
counted after 14 days. Multiclonal strains were established by passag-
ing all colonies obtained in primary CFU-F cultures. Individual colonies
(clones) were isolated from primary CFU-F cultures using cloning cyl-
inders (Kuznetsov et al., 1997). Nonclonal BMSC cultures were ob-
tained by passaging primary cultures established at nonclonal density
(>105 total nucleated cells/cm2).
Histology
Heterotopic ossicles were processed as reported (Bianco et al., 1998).
Sections of human iliac crest biopsies (three subjects) with normal BM
were cut from archival paraffin blocks on file in our department. Human
specificity of the CD146 antibody was verified on sections of mouse
bone/BM and of heterotopic ossicles formed by murine BMSCs (Kuz-
netsov et al., 2004). Immunolocalization was performed using standard
immunoperoxidase (DAB reaction) and sections were counterstained
with hematoxylin.
In Vivo Transplantation
In vivo transplantation of different cell strains was performed as
reported (Krebsbach et al., 1997; Kuznetsov et al., 1997). All animal
procedures were approved by the relevant institutional committee.
2 3 106 cells were allowed to attach to hydroxyapatite/tricalcium
phosphate particles (40 mg, 100–200 mm; Zimmer, Warsaw IN) and
embedded in a fibrin gel. Carrier-cell constructs, and carrier alone
as control, were transplanted subcutaneously into 8- to 15-week-
old female nih/nu/xid/bg mice (Harlan-Sprague Dawley, Inc., Indianap-
olis, IN).
Secondary Passage of CD146+ CFU-Fs
Cell cultures were initiated either from a limited number of CFU-Fs (10
and 12 in two experiments) or from single CFU-Fs (n = 5). Heterotopic
ossicles were harvested at 8 weeks, washed in HBSS/30 mM HEPES,
100 U/ml penicillin, 100 mg/ml streptomycin, and digested twice with
100 U/ml Chlostridium histolyticum type II collagenase (Invitrogen) in
PBS/3 mM CaCl2 for 40 min at 37�C. 5 3 105 cells obtained from the
two digestions were used for FACS analysis of hCD146 expression.
Cell suspensions derived from multiclonal generated ossicles were
used to magnetically separate human cells based on hCD44 expres-
sion using MiniMacs (Miltenyi), either after short-term culture or di-
rectly from the fresh cell suspension. hCD44+ cells (�20,000 and
�15,000 in two experiments) were recovered, resuspended in me-
dium, and 2,000 and 5,000 cells, respectively, were plated in culture
at clonal density (1.6 cells/cm2). Cultures were scored for colony for-
mation at 2 weeks. The discrete colonies obtained were harvested
and analyzed by FACS for expression of hCD90, hCD44, and
hCD146. Cell suspensions obtained by collagenase digestion of ossi-
cles generated by transplanting the progeny of single CFU-Fs were
used to sort hCD146+ cells directly. These were then replated at clonal
density (1.6 cells/cm2) to assay for secondary CFU-Fs. The discrete
colonies obtained were harvested and analyzed for expression of
hCD90, hCD44, and hCD146.
Supplemental Data
Supplemental Data include Supplemental Results, Supplemental Ex-
perimental Procedures, eight figures, and nine tables and can be found
with this article online at http://www.cell.com/cgi/content/full/131/2/
324/DC1/.
ACKNOWLEDGMENTS
This work was supported by AIRC, Telethon and MIUR of Italy (to P.B.),
and in part by the DIR/NIDCR of the IRP/NIH (P.G.R.).
Received: August 10, 2006
Revised: May 29, 2007
Accepted: August 6, 2007
Published: October 18, 2007
REFERENCES
Antonelli-Orlidge, A., Saunders, K.B., Smith, S.R., and D’Amore, P.A.
(1989). An activated form of transforming growth factor beta is pro-
duced by cocultures of endothelial cells and pericytes. Proc. Natl.
Acad. Sci. USA 86, 4544–4548.
Arai, F., Hirao, A., Ohmura, M., Sato, H., Matsuoka, S., Takubo, K., Ito,
K., Koh, G.Y., and Suda, T. (2004). Tie2/angiopoietin-1 signaling regu-
lates hematopoietic stem cell quiescence in the bone marrow niche.
Cell 118, 149–161.
Barry, F.P., Boynton, R.E., Haynesworth, S., Murphy, J.M., and Zaia, J.
(1999). The monoclonal antibody SH-2, raised against human mesen-
chymal stem cells, recognizes an epitope on endoglin (CD105). Bio-
chem. Biophys. Res. Commun. 265, 134–139.
Bianco, P., and Boyde, A. (1993). Confocal images of marrow stromal
(Westen-Bainton) cells. Histochemistry 100, 93–99.
Bianco, P., and Robey, P.G. (2000). Marrow stromal stem cells. J. Clin.
Invest. 105, 1663–1668.
Bianco, P., and Robey, P.G. (2004). Skeletal stem cells. In Handbook
of Stem Cells, R. Lanza, ed. (New York: Academic Press), pp. 415–
424.
Bianco, P., Kuznetsov, S.A., Riminucci, M., Fisher, L.W., Spiegel, A.M.,
and Robey, P.G. (1998). Reproduction of human fibrous dysplasia of
bone in immunocompromised mice by transplanted mosaics of normal
and Gsalpha-mutated skeletal progenitor cells. J. Clin. Invest. 101,
1737–1744.
Bianco, P., Kuznetsov, S.A., Riminucci, M., and Robey, P.G. (2006).
Post-natal skeletal stem cells. Methods Enzymol. 419, 117–149.
Calvi, L.M., Adams, G.B., Weibrecht, K.W., Weber, J.M., Olson, D.P.,
Knight, M.C., Martin, R.P., Schipani, E., Divieti, P., Bringhurst, F.R.,
et al. (2003). Osteoblastic cells regulate the haematopoietic stem cell
niche. Nature 425, 841–846.
Caplan, A.I. (1991). Mesenchymal stem cells. J. Orthop. Res. 9, 641–
650.
Davis, S., Aldrich, T.H., Jones, P.F., Acheson, A., Compton, D.L., Jain,
V., Ryan, T.E., Bruno, J., Radziejewski, C., Maisonpierre, P.C., and
Yancopoulos, G.D. (1996). Isolation of angiopoietin-1, a ligand for
the TIE2 receptor, by secretion-trap expression cloning. Cell 87,
1161–1169.
Deschaseaux, F., and Charbord, P. (2000). Human marrow stromal
precursors are alpha 1 integrin subunit-positive. J. Cell. Physiol. 184,
319–325.
Dumont, D.J., Yamaguchi, T.P., Conlon, R.A., Rossant, J., and Breit-
man, M.L. (1992). tek, a novel tyrosine kinase gene located on mouse
chromosome 4, is expressed in endothelial cells and their presumptive
precursors. Oncogene 7, 1471–1480.
Duncan, A.W., Rattis, F.M., Di Mascio, L.N., Congdon, K.L., Pazianos,
G., Zhao, C., Yoon, K., Cook, J.M., Willert, K., Galano, N., and Reya, T.
(2005). Integration of Notch and Wnt signalling in hematopoietic stem
cell maintenance. Nat. Immunol. 6, 314–322.
Friedenstein, A.J. (1980). Stromal mechanisms of bone marrow: clon-
ing in vitro and retransplantation in vivo. Hamatol. Bluttransfus. 25, 19–
29.
Friedenstein, A.J. (1990). Bone marrow osteogenic stem cells. In Cal-
cium Regulation and Bone Metabolism, D.V. Cohn, F.H. Glorieux, and
T.J. Martin, eds. (Cambridge, UK: Elsevier), pp. 353–361.
Friedenstein, A.J., Chailakhyan, R.K., Latsinik, N.V., Panasyuk, A.F.,
and Keiliss-Borok, I.V. (1974). Stromal cells responsible for transferring
Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc. 335

the microenvironment of the hemopoietic tissues. Cloning in vitro and
retransplantation in vivo. Transplantation 17, 331–340.
Gronthos, S., Zannettino, A.C., Graves, S.E., Ohta, S., Hay, S.J., and
Simmons, P.J. (1999). Differential cell surface expression of the
STRO-1 and alkaline phosphatase antigens on discrete developmental
stages in primary cultures of human bone cells. J. Bone Miner. Res. 14,
47–56.
Hattori, K., Dias, S., Heissig, B., Hackett, N.R., Lyden, D., Tateno, M.,
Hicklin, D.J., Zhu, Z., Witte, L., Crystal, R.G., et al. (2001). Vascular en-
dothelial growth factor and angiopoietin-1 stimulate postnatal hemato-
poiesis by recruitment of vasculogenic and hematopoietic stem cells.
J. Exp. Med. 193, 1005–1014.
Hirschi, K.K., and D’Amore, P.A. (1996). Pericytes in the microvascula-
ture. Cardiovasc. Res. 32, 687–698.
Hsu, H.C., Ema, H., Osawa, M., Nakamura, Y., Suda, T., and Nakauchi,
H. (2000). Hematopoietic stem cells express Tie-2 receptor in the mu-
rine fetal liver. Blood 96, 3757–3762.
Iwama, A., Hamaguchi, I., Hashiyama, M., Murayama, Y., Yasunaga,
K., and Suda, T. (1993). Molecular cloning and characterization of
mouse TIE and TEK receptor tyrosine kinase genes and their expres-
sion in hematopoietic stem cells. Biochem. Biophys. Res. Commun.
195, 301–309.
Jain, R.K. (2003). Molecular regulation of vessel maturation. Nat. Med.
9, 685–693.
Kiel, M.J., Yilmaz, O.H., Iwashita, T., Terhorst, C., and Morrison, S.J.
(2005). SLAM family receptors distinguish hematopoietic stem and
progenitor cells and reveal endothelial niches for stem cells. Cell
121, 1109–1121.
Krebsbach, P.H., Kuznetsov, S.A., Satomura, K., Emmons, R.V.,
Rowe, D.W., and Robey, P.G. (1997). Bone formation in vivo: compar-
ison of osteogenesis by transplanted mouse and human marrow stro-
mal fibroblasts. Transplantation 63, 1059–1069.
Kuznetsov, S.A., Krebsbach, P.H., Satomura, K., Kerr, J., Riminucci,
M., Benayahu, D., and Robey, P.G. (1997). Single-colony derived
strains of human marrow stromal fibroblasts form bone after transplan-
tation in vivo. J. Bone Miner. Res. 12, 1335–1347.
Kuznetsov, S.A., Riminucci, M., Ziran, N., Tsutsui, T.W., Corsi, A.,
Calvi, L., Kronenberg, H.M., Schipani, E., Robey, P.G., and Bianco,
P. (2004). The interplay of osteogenesis and hematopoiesis: expres-
sion of a constitutively active PTH/PTHrP receptor in osteogenic cells
perturbs the establishment of hematopoiesis in bone and of skeletal
stem cells in the bone marrow. J. Cell Biol. 167, 1113–1122.
Maniatis, A., Tavassoli, M., and Crosby, W.H. (1971). Origin of osteo-
genic precursor cells in extramedullary marrow implants. Blood 38,
569–575.
Miura, Y., and O’Driscoll, S.W. (1998). Culturing periosteum in vitro: the
influence of different sizes of explants. Cell Transplant. 7, 453–457.
Moore, K.A., and Lemischka, I.R. (2006). Stem cells and their niches.
Science 311, 1880–1885.
Nagasawa, T., Hirota, S., Tachibana, K., Takakura, N., Nishikawa, S.,
Kitamura, Y., Yoshida, N., Kikutani, H., and Kishimoto, T. (1996). De-
fects of B-cell lymphopoiesis and bone-marrow myelopoiesis in mice
lacking the CXC chemokine PBSF/SDF-1. Nature 382, 635–638.
Owen, M., and Friedenstein, A.J. (1988). Stromal stem cells: marrow-
derived osteogenic precursors. Ciba Found. Symp. 136, 42–60.
336 Cell 131, 324–336, October 19, 2007 ª2007 Elsevier Inc.
Pickl, W.F., Majdic, O., Fischer, G.F., Petzelbauer, P., Fae, I., Waclavi-
cek, M., Stockl, J., Scheinecker, C., Vidicki, T., Aschauer, H., et al.
(1997). MUC18/MCAM (CD146), an activation antigen of human T lym-
phocytes. J. Immunol. 158, 2107–2115.
Robey, P.G., and Termine, J.D. (1985). Human bone cells in vitro. Cal-
cif. Tissue Int. 37, 453–460.
Sato, T.N., Qin, Y., Kozak, C.A., and Audus, K.L. (1993). Tie-1 and tie-2
define another class of putative receptor tyrosine kinase genes ex-
pressed in early embryonic vascular system. Proc. Natl. Acad. Sci.
USA 90, 9355–9358.
Sato, Y., and Rifkin, D.B. (1989). Inhibition of endothelial cell move-
ment by pericytes and smooth muscle cells: activation of a latent
transforming growth factor-beta 1-like molecule by plasmin during
co-culture. J. Cell Biol. 109, 309–315.
Shi, S., and Gronthos, S. (2003). Perivascular niche of postnatal mes-
enchymal stem cells in human bone marrow and dental pulp. J. Bone
Miner. Res. 18, 696–704.
Shih, I.M. (1999). The role of CD146 (Mel-CAM) in biology and pathol-
ogy. J. Pathol. 189, 4–11.
Simmons, P.J., and Torok-Storb, B. (1991). Identification of stromal
cell precursors in human bone marrow by a novel monoclonal anti-
body, STRO-1. Blood 78, 55–62.
Sugiyama, T., Kohara, H., Noda, M., and Nagasawa, T. (2006). Mainte-
nance of the hematopoietic stem cell pool by CXCL12-CXCR4
chemokine signaling in bone marrow stromal cell niches. Immunity
25, 977–988.
Suri, C., Jones, P.F., Patan, S., Bartunkova, S., Maisonpierre, P.C.,
Davis, S., Sato, T.N., and Yancopoulos, G.D. (1996). Requisite role
of angiopoietin-1, a ligand for the TIE2 receptor, during embryonic
angiogenesis. Cell 87, 1171–1180.
Tavassoli, M., and Crosby, W.H. (1968). Transplantation of marrow to
extramedullary sites. Science 161, 54–56.
Tavassoli, M., and Friedenstein, A. (1983). Hemopoietic stromal micro-
environment. Am. J. Hematol. 15, 195–203.
Vogel, W., Grunebach, F., Messam, C.A., Kanz, L., Brugger, W., and
Buhring, H.J. (2003). Heterogeneity among human bone marrow-de-
rived mesenchymal stem cells and neural progenitor cells. Haemato-
logica 88, 126–133.
Weiss, L. (1976). The hematopoietic microenvironment of the bone
marrow: an ultrastructural study of the stroma in rats. Anat. Rec.
186, 161–184.
Westen, H., and Bainton, D.F. (1979). Association of alkaline-phospha-
tase-positive reticulum cells in bone marrow with granulocytic precur-
sors. J. Exp. Med. 150, 919–937.
Zannettino, A.C., Harrison, K., Joyner, C.J., Triffitt, J.T., and Simmons,
P.J. (2003). Molecular cloning of the cell surface antigen identified by
the osteoprogenitor-specific monoclonal antibody, HOP-26. J. Cell.
Biochem. 89, 56–66.
Zhang, J., Niu, C., Ye, L., Huang, H., He, X., Tong, W.G., Ross, J.,
Haug, J., Johnson, T., Feng, J.Q., et al. (2003). Identification of the hae-
matopoietic stem cell niche and control of the niche size. Nature 425,
836–841.

Cell, Volume 131
Supplemental Data
Self-Renewing Osteoprogenitors in Bone
Marrow Sinusoids Can Organize
a Hematopoietic Microenvironment Benedetto Sacchetti, Alessia Funari, Stefano Michienzi, Silvia Di Cesare, Stefania Piersanti, Isabella Saggio, Enrico Tagliafico, Stefano Ferrari, Pamela Gehron Robey, Mara Riminucci, and Paolo Bianco
Supplemental Results BMSCs direct assembly and remodeling of pseudovascular structures in vitro through CD146 and Ang-1 To further probe the function of CD146+ BMSCs as MCs, and the potential role of CD146 and Ang-1 in their interaction with ECs, an in vitro system designed to assay the ability of ECs and MCs to assemble into pseudovascular structures was used [cord assembly assay (Darland and D'Amore, 2001)]. In this assay, ECs cultured in isolation at low cell density failed to form pseudovascular cords. BMSCs efficiently directed assembly of 3D pseudovascular cords in which ECs and BMSCs colocalized (Suppl Fig 4). No such structures formed when SFs or two epithelial cell lines were substituted for either CD146+ BMSCs or HUVEC (not shown). Silencing expression of either CD146 or Ang-1 in BMSCs strikingly altered the assembly, pattern and remodeling of pseudovascular cords. CD146 silencing in BMSCs (Suppl Fig 7), but not in ECs (not shown), and strikingly altered secondary remodeling of the primary lattice between 6-24h, leading to formation of a stable pattern of long and thin cords (Suppl Fig 7), which did not undergo timed remodeling and regression. Ang-1 silencing resulted in the near-complete abrogation of primary pseudovascular cords (Suppl Fig 7), suggesting that formation of pseudovascular cords at low cell density was dependent not only on BMSCs, but also on BMSC-produced Ang-1. Factors regulating microvessel assembly regulate CD146 and Ang-1 expression in BMSCs During angiogenesis and vessel maturation, FGF-2 promotes MC proliferation and PDGF-BB recruits presumptive MCs to the vessel wall. TGF-β, activated at sites of EC-MC contact, induces their mitotic quiescence (Jain, 2003). Both FGF-2 and TGF-β were highly expressed in the heterotopic ossicles prior to the onset of hematopoiesis. FGF-2 was expressed in fibroblastic cells and osteoblasts. Consistent with the activation of latent TGF-β at sites of EC-MC contacts, mature TGF-β was localized to the wall of developing sinusoids (Suppl Fig 5). Consistent with their general effects on MCs (Jain, 2003), TGF-β and FGF-2 modulated the proliferation and phenotype of cultured CD146+ BMSCs in opposite ways. FGF-2 stimulated cell proliferation and attenuated the MC phenotype. TGF-β inhibited proliferation and preserved or enhanced features of a MC phenotype, and PDGF-BB had similar effects (Suppl Fig 3, Suppl Fig 8). In this context, expression of CD146 and Ang-1 were essentially abrogated by FGF-2, reduced by PDGF-BB, and left unchanged by TGF-β (Suppl Fig 8).

FGF-2-treated BMSCs do not transfer the HME We then asked if in vitro effects of FGF-2 would also be associated with a functional effect in vivo. BMSCs expanded under standard conditions or with 5ng/ml FGF-2 were transplanted in vivo. Control strains established heterotopic bone and BM (6/6 transplants). FGF-2 treated cells formed heterotopic ossicles that included abundant bone, but were devoid of sinusoids, hematopoiesis and CD146+ stromal cells (6/6 transplants; Suppl Fig 8). Therefore, FGF-2 stimulation of BMSCs prior to transplantation apparently dissociated their ability to form bone from their ability to transfer the HME in vivo.
Supplemental Experimental Procedures Gene expression profiling and data analysis Total cellular RNA was isolated from cell populations using RNeasy RNA isolation kit (Qiagen, Valencia, CA) following manufacturer’s recommendations. Disposable RNA chips (Agilent RNA 6000 Nano LabChip kit) were used to determine the concentration and purity/integrity of RNA samples using Agilent 2100 bioanalyzer. cDNA synthesis, biotin-labeled target synthesis, HG-U133 plus 2.0 GeneChip (Affymetrix, Santa Clara, CA) arrays hybridization, staining and scanning were performed according to the standard protocol supplied by Affymetrix. The amount of a transcript mRNA (signal) was determined by the Affymetrix GeneChip Operative Software (GCOS) 1.2 absolute analysis algorithm as already described (Liu et al 2002). All expression values for the genes in the GCOS absolute analyses were determined using the global scaling option. Alternatively, probe level data were converted to expression values using robust multi-array average (RMA) procedure (Irizarry et al 2003). Perfect Match (PM) values were background adjusted, normalized using invariant set normalization, and log transformed. The RMA generated data were uploaded onto GeneSpringTM software version 7.3 using the log2 transformation procedure. A “per chip” and a “per gene” normalization were achieved by dividing each signal for the 50.0th percentile of all above-10 signals in that sample and by the median of its values in all samples. For growth factor treatment experiments, a “per gene” normalization was achieved by dividing each signal of treated samples for the signal in untreated sample. Hierarchical agglomerative clustering was performed in GeneSpring™ using Pearson’s correlation coefficient and average-linkage as distance and linkage methods. Functionally oriented gene lists were obtained through the use of Ingenuity Pathways Analysis (Ingenuity® Systems, www.ingenuity.com). All microarray data have been submitted to NCBI-GEO MIAME compliant public database (Accession number GSE6460).
RT-PCR Total RNA was extracted using TRIZOLTM RNA isolation system (Invitrogen) per the manufacturer’s instructions. cDNA was synthesized using 3µg of RNA, 150ng of random hexamers, and 50 units of SuperScript II Reverse Transcriptase (Invitrogen) in a total volume of 20µl. The target cDNA sequences were amplified in standard PCR reactions using Platinum® PCR SuperMix in agreement to the manufacturer’s instructions. After a denaturation step at 94°C for 2 minutes, the reactions were run for 21-23 cycles (GAPDH) or 23-33 cycles (for other genes) at the following temperatures: 94°C, 30 sec; 56 °C (GAPDH) or 55 °C (other genes), 30 sec; 72°C 40 sec, with a final extension at 72°C for 3 minutes. Primers used for RT-PCR are listed in Suppl Table 8. Amplified PCR fragments were electrophoresed on 2% agarose gels and stained with ethidium bromide.

Quantitative real-time PCR analysis of endothelial and mural cell markers Quantitative real-time PCR (qRT-PCR) experiments were carried out using an ABI PRISM 7000 Sequence Detection System (Applied Biosystems, Gaithersburg, MD). Taqman oligonucleotides (Assay-on-Demand) for glyceraldehyde-3-phosphate dehydrogenase (GAPDH), cadherin 5, type 2, VE-cadherin (CDH5), vascular endothelial growth factor receptor 2 (KDR), CD34 molecule (CD34) transcript variant 1, prominin 1, actin alpha 2 smooth muscle aorta (ACTA2) [Assay ID: Hs00426835_g1], chondroitin sulfate proteoglycan 4 (CSPG4) [Assay ID: Hs00426981_m1], caldesmon 1 (CALD1) [Assay ID: Hs00189021_m1], calponin 2 (CNN2) [Assay ID: Hs00854264_s1] transcript variant 1, desmin (DES) [Assay ID: Hs00157258_m1], collagen, type IV alpha 1 (COL4A1) [Assay ID: Hs00266237_m1], collagen type IV alpha 2 (COL4A2) [Assay ID: Hs00300500_m1], platelet-derived growth factor receptor beta polypeptide (PDGFRB) [Assay ID: Hs00182163_m1], endothelial differentiation sphingolipid G-protein-coupled receptor 1 (EDG1) [Assay ID: Hs00173499_m1], integrin, alpha 1 (ITGA1) [Assay ID: Hs00235030_m1], endoglin (Osler-Rendu-Weber syndrome 1) (ENG) [Assay ID: Hs00164438_m1] were from TaqMan® Assays-on-demand Gene expression products (Applied Biosystems). Ct values were normalized with those obtained from the amplification of GAPDH.
Sorting and CFE assay of CD34+ cells 1x107 freshly isolated bone marrow nucleated cells (BMNCs) were resuspended in HBSS containing 30mM HEPES (Sigma, St Louis, MO), 100U/ml penicillin, 100µg/ml streptomycin), 1% BSA and incubated on ice for 30 minutes. The cells were then pelleted in HBSS supplemented with 2mM EDTA (Sigma), 1% BSA (Sigma), resuspended in 1 ml blocking buffer and incubated with PE-conjugated anti-CD146 clone P1H12 and anti-CD34 Clone 581 antibodies, and CD146+/CD34+, CD146-/CD34+ fractions were separated using a FACS DIVAntageSE flow cytometer (BD Biosciences Labware, San Diego, CA). In a separate experiment, cells were incubated with anti-CD34 clone QBEND/10 conjugated magnetic beads (Miltenyi Biotec, Auburn, CA) for 20 min on ice. Cells were separated into CD34- and CD34+ fractions using a MiniMACS magnetic column separation unit (Miltenyi) as per the manufacturer’s instructions. CFE assays were conducted by plating cells at different densities and scoring the formation of CFU-Fs at 14 days (see material and methods) in comparison with unsorted cells.
Endothelial differentiation assay MCAM/CD146-expressing BMSCs from primary CFU-F cultures were plated at 2x104 cells/cm2 in fibronectin-coated culture plates and cultured with two different protocols (A and B). Protocol A was as described in Reyes et al 2002. Briefly, cells were cultured in low glucose DMEM/MCDB 201(Sigma) with 10ng/ml VEGF, ITS, dexamethasone, ascorbic acid. Protocol B involved culture in Clonetics Endothelial Cell Basal medium (Cambrex) supplemented with 5% FBS, 10-4 M ascorbic acid, 1ng/ml VEGF, 10ng/ml FGF2, 10ng/ml EGF, 20ng/ml Long R3 IGF-1, 22.5µg/ml heparin, 200ng/ml hydrocortisone and 100 U penicillin, and 1,000 U streptomycin (all from Cambrex). After 10 days, expression of CD14 (LPS receptor), CD31 (PECAM-1), CD34, CD133, CD144 (VE-cadherin), KDR (VEGFR2), CD62E (E-selectin) markers was assessed by using a FACSCalibur flow cytometer and CellQuest software (Becton Dickinson Biosciences, San Diego, CA, USA). Expression of Tie-2 was evaluated by western blot analysis. The DiI-Ac-LDL staining kit was purchased from Biomedical Technologies (Stoughton, Massachusetts, USA) and the assay was performed per the manufacturer’s recommendations. Human umbilical vein endothelial cells (HUVEC) were used as a positive control.

Microvascular cord assembly Endothelial cells (ECs-HUVEC and HMVEC-d) were grown in Clonetics EGM-2 BulletKit and EGM-2-MV BulletKit (Cambrex Corporation, Walkersville, NJ). BMSCs and ECs were labeled with PKH67 and PKH26 respectively per the manufacturer’s instructions (Sigma). Cultures of ECs, CD146+ BMSCs, or SFs alone, and co-cultures of ECs and CD146+ BMSCs or SFs were established on Growth Factor Reduced MatrigelTM as reported (Darland and D'Amore, 2001). 50,000 cells/well were plated on 200µl MatrigelTM in 24 well plates in phenol red free-DMEM (Invitrogen), 2% FBS. Pseudovascular cord formation was monitored at 3, 6, 9, 12, 24 and 48hrs. Cord length and number were measured in 30 random images for each experimental point and compared by ANOVA).
Confocal microscopy Confocal fluorescence images of co-cultures of HUVEC and BMSCs were obtained using the Leica TCS SP5 confocal laser scanning microscopy system (Leica Microsystems, Mannheim, Germany) using the HeNe 543 nm for and the Ar 488 nm for visualizing the red and green vital fluorochromes respectively. Stacks of 512 x 512 pixel optical sections were collected with 0.117 µm interval using HCX Plan Apo 63X oil, NA 1.40 objective.
Lenti-viral vectors for CD146 and Ang-1 silencing Short hairpin (sh) sequences (19 nt) targeted to human CD146 exon 6, 8 and 15 and targeted to human Ang-1 exon 1, 2/3, 4 and 9 were designed using algorithms in the public domain (http://www.ambion.com/techlib/misc/siRNA_finder.html), submitted to BLAST analysis to exclude off-target annealing, and custom-synthesized (Operon Biotechnologies GmbH, Cologne, Germany). The control 19 nt sequence was designed to not match any sequence in the human genome. The shRNA duplexes were cloned into ClaI/MluI sites of the pLVTHM-eGFP lenti-viral transfer vector (from D. Trono, Ecole Politechnique, Genève Switzerland; maps at http://www.tronolab.com), downstream of the H1 promoter. Lenti-viral vectors were produced as described (Piersanti et al., 2006), by transfecting 293T cells with the transfer vector, the packaging vector pCMV-dR8.74 and the VSV-G envelope vector pMD2G (http://www.tronolab.com). ECs and BMSCs were infected with each lentivirus as described (Piersanti et al., 2006). Efficiency and efficacy were assessed by western blot analysis and FACS (CD146) or ELISA (Ang-1). The lenti-viral vectors encoding shRNA targeted to CD146 exon 15 (LV-shCD146) and to Ang-1 exon 1 (LV-siAngpt-1) were chosen as the most effective and used for experiments at an MOI of 1 (Suppl Fig 5D-F).
Expression and regulation of CD146 and Ang-1 Osteogenic differentiation of BMSCs was induced as described (Bianco et al. 2006). FGF-2, TGF-β (Sigma) or PDGF-BB (R&D Systems) were added to cultures for 1wk (10ng/ml). Total RNA was extracted using TRIZOLTM per the manufacturer's instructions (Invitrogen). cDNA was synthesized using 3µg of RNA, 150ng of random hexamers, and 50 units of SuperScript II Reverse Transcriptase (Invitrogen) in a total volume of 20µl. Real-time PCR was performed in an ABI PRISM 7000 Detection System (Applied Biosystems, Foster City, USA) using TaqMan oligonucleotides (Assay on Demand) for human Ang-1, CD146, Cbfa1/Runx2, Bone Sialoprotein, Osteocalcin, and GAPDH (Applied Biosystems). Delta-delta Ct values were normalized to GAPDH. Proteins were extracted as described (Wedegaertner et al., 1996), separated on NuPAGE Novex 4-12% Bis-Tris gels (Invitrogen) under reducing conditions, and

transferred onto PVDF membrane (Invitrogen). Immunoblotting was performed with 150ng/ml anti-human Ang-1, Tie-2, and anti-CD146 (1:2,500) and anti-actin (1:15,000). Ang-1 in cell-conditioned medium was immunoprecipitated and western blot analysis was performed under native conditions. The concentration of Ang-1 in conditioned medium was assessed by Quantikine (Human Ang-1 Immunoassay; R&D Systems) according to the manufacturer’s instructions.
Figure S1. A) Frequency of CD146+ cells in different cell strains. Data are derived from FACS analyses of 3 different cultures for each strain at passage 3, and are expressed as mean + SD. The frequency of CD146+ cells observed in BMSC (bone marrow stromal cell) cultures, obtained from CFU-F (Colony Forming Unit-Fibroblast) primary cultures is significantly higher than in all other strains (ANOVA). FD, stromal cells from the non-hematopoietic, fibrotic marrow of fibrous dysplasia; HTB, human trabecular bone cells; PE, periosteal cells; MF, muscle fibroblasts; SF, skin fibroblasts. B) Frequency of CD146+ cells in single clones (n=3), multi-clonal cultures (n=3), and non-clonal cultures (n=3) of BMSCs. Single clonal and multi-clonal strains express CD146 homogeneously, whereas a significantly lower proportion of cells express CD146 in non-clonal BMSC cultures. Data are expressed as mean + SD and compared by ANOVA.


Figure S2. Mural cell phenotype of cultured CD146+ cells A) Graphic representation of hybridization intensity (signal) on Affymetrix GeneChips of mural cell markers, endothelial markers, genes related to early and late osteogenesis, and of genes expressed by the hematopoietic niche. The BMSC samples used were from 4 different donors (1-4) and are represented in the columns. Sample 3 is from the same cell strain used for the FACS analysis and in vivo transplantation, which is shown in Fig 4. The signal value was calculated using the absolute analysis algorithm of the Affymetrix GeneChip Operative Software (GCOS). The signal intensity gradient is shown at bottom. CD146+ cells express VEGF, and mural cell genes (α-smooth muscle actin (ACTA2), calponin 1 and 3 (CNN1,3), caldesmon 1 (CALD1), leiomodin 1 (LMOD1), PDGFR-β, myosin regulatory protein (MRCL3), the pericyte marker NG2 (CSPG4) (Ozerdem, 2001) and, at low levels, desmin (DES), whereas they do not express endothelial markers (CD31/PECAM, von Willebrand Factor (VWF), CD133 (PROM1), VEGFR2 (KDR), VE-cadherin (CDH5), CD34, and Flt-1. They express markers of early, immature osteogenic cells such as RUNX2, COL1A1 and COLA12, osteonectin (SPARC), osteoblastic cadherin (CDH11), and alkaline phosphatase (ALP)), but do not express markers of mature osteoblasts (late osteogenic – osterix (SP7), bone sialoprotein (IBSP), osteocalcin (BGLAP) and osteopontin (SSP1). They also express genes related to the HSC niche (Jagged-1 (JAG1), Kit ligand (KITLG), N-cadherin (CDH2), and CXCL12). B) RT-PCR analysis of expression of a selection of the same genes. Lane 1, HUVEC; Lanes 2 and 3, two different strains of CD146+ BMSCs; Lane 4, negative control. Endothelial markers (VE-cadherin, Flk-1, CD34, CD133) are not expressed in CD146+ BMSCs. All of them are expressed in HUVEC (CD133 at low levels). The integrin α1 subunit (ITGA1) and endoglin (ENG) are expressed both in endothelial cells and CD146+ cells. Markers of mural cells such as the pericyte marker NG2 (CSPG4) (Ozerdem et al 2001), desmin and the PDGF receptor β are expressed in CD146+ BMSCs and not in HUVEC. Low levels of EDG1, a shingosine-1-phosphate receptor expressed in both endothelial cells and mural cells and required for the establishment of mural cells (Liu et al 2000, Kluk and Hla 2002 #22), are expressed in BMSCs. C) Lack of endothelial differentiation potential of CD146+ BMSCs. CD146+ BMSCs from primary CFU-F cultures were maintained in standard medium or cultured under endothelial differentiating conditions (Reyes et al, 2002). Expression of endothelial markers was analyzed by FACS after 10 days in control and treated BMSCs (BMSC-E), and in HUVEC maintained in their medium as a positive control. No endothelial marker [CD34, CD31, Flk-1, CD133, VE-cadherin, E-Selectin (CD62E)] was expressed in BMSCs or up-regulated as an effect of exposure to endothelial differentiation medium. Identical results (not shown) were obtained using a different endothelial growth medium (see Suppl. Experimental Procedures). HUVEC expressed CD31, Flk-1, VE-cadherin, but CD34 and CD133 expression were barely detectable by FACS analysis. Exposure to endothelial differentiation medium down regulated CD146 expression in BMSCs. D) Functional analysis of endothelial phenotype of the same cell strains. Uptake of DiI-Ac-LDL, a known activity of endothelial cells (Reyes et al 2002, Steinberg et al 1985), was observed in HUVEC, but not in BMSCs maintained in standard medium or in endothelial differentiation medium (BMSC-E).


Figure S3. Proliferation and phenotype of CD146+ BMSCs are modulated by mural cell regulating factors A) Effects of FGF-2, TGF-β, and PDGF-BB on BMSC growth (population doublings, mean + SEM, n=3) in the absence of serum and B) in the presence of serum. C) Analysis of expression of 1140 genes positively regulating progression along the cell cycle, which were selected using the Ingenuity Pathway Analysis application (the gene list is provided in Suppl. Table 8). The vast majority of these genes are up regulated after FGF-2 treatment, and down regulated or not changed after TGFβ or PDGF treatment. D) Graphic representation of hybridization intensity (signal) on Affymetrix GeneChips of mural cell markers. FGF-2 treatment down regulates most mRNAs characteristic of the mural cell phenotype, including α-smooth muscle actin (ACTA2), caldesmon 1 (CALD1), calponins (CNN 1-3), desmin (DES), the PDGFR-β (PDGFRB), and basement membrane proteins such as SMOC1 and COL4A1. Of note, FGF-2 treatment up regulates expression of VEGF, and of a group of sphingosine-1-phosphate receptors that are co-expressed in endothelial cells and mural cells (Kluk and Hla, 2002). EDG1, which mediates a proliferative and migratory response in mural cells (Kluk and Hla, 2002), is required for the establishment of mural cells in the mouse (Liu et al. 2000). TGF-β, which stabilizes mural cells and nascent vessels (Jain, 2003), has primarily the opposite effects. E) qPCR validation of array results, a selection of mRNAs is shown. F) FACS analysis. FGF-2 down regulates CD146, the PDGFR-β (which is involved in the recruitment of mural cells to neovessel walls), endoglin (CD105, a TGF-β co-receptor), α1 integrin [CD49a (which binds to collagen type IV)]. PDGF-BB has similar effects. None of these markers are down regulated by TGF-β, which in contrast down regulates CD63 and promotes the expression of α-SM actin. Expression of Thy-1 (CD90) and ALP, different “markers” of “mesenchymal stem cells,” are left unchanged by all treatments.


Figure S4. CD146+ cells function as mural cells in co-culture with endothelial cells A) Human ECs and SFs do not self-aggregate on MATRIGELTM in culture by themselves. CD146+ BMSCs form 2D sheets. B) Pseudovascular cords are formed in co-cultures of ECs with CD146+ BMSCs, but not with SFs. C) Confocal microscopy of pseudovascular cords. A selection of optical sections out of a stack of 117 images (top), and two orthogonal z-sections (bottom) are shown to demonstrate the position of BMSCs (green) to the side and the top of endothelial cells (red).
Figure S5. FGF-2 was expressed in fibroblastic cells and osteoblasts in the heterotopic ossicles prior to establishment of hematopoiesis (two left panels). Localization of active TGF-beta is observed along the walls and in the vicinity of the developing sinusoids (large arrows, middle and right panels), and in the newly deposited bone matrix, adjacent to osteoblasts (small arrows, lower middle panel).
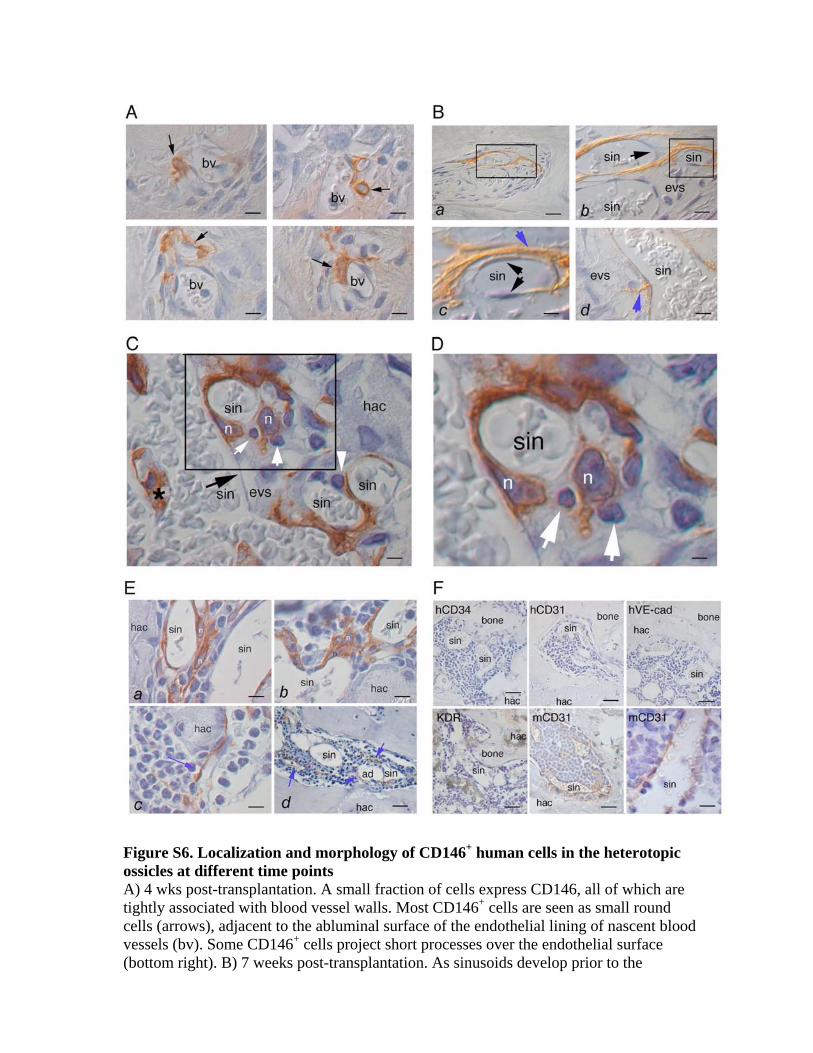
Figure S6. Localization and morphology of CD146+ human cells in the heterotopic ossicles at different time points A) 4 wks post-transplantation. A small fraction of cells express CD146, all of which are tightly associated with blood vessel walls. Most CD146+ cells are seen as small round cells (arrows), adjacent to the abluminal surface of the endothelial lining of nascent blood vessels (bv). Some CD146+ cells project short processes over the endothelial surface (bottom right). B) 7 weeks post-transplantation. As sinusoids develop prior to the

establishment of hematopoiesis, CD146+ cells elongate to cover their abluminal surface. Ba), CD146+ cells associated with one sinusoid. Due to warping of the sinusoid across the sectioning plane, two sections of the sinus wall and lumen are visible (box). Bb) An elongated CD146+ cells wraps around the vessel with a spiral course from bottom to top side, but is clearly located outside of the endothelial layer. Arrow indicates the endothelial nuclei. Bc) Higher magnification of the area boxed in Bb. The endothelial wall and the nuclei of two endothelial cells are clearly seen (black arrows). The CD146+ cell (blue arrow) resides in the extravascular space. Bd) A different sinusoid, and a lateral process projected from an adventitial CD146+ cell into the extravascular space (evs). Note the contact between the adventitial cell process and a single small mononuclear cell. C) High magnification, and D) detail, of a developing marrow space at 7 weeks post-transplantation. A large sinus (sin) and several collateral branches containing red blood cells are shown. The endothelial wall of the large sinus is identified by the black arrow. Multiple CD146+ adventitial reticular cells, and the nuclei (n) of two of them are clearly located in the extra-vascular space (evs) between the sinus wall and the hydroxyapatite carrier (hac). White arrows (C,D) indicate small round cells of hematopoietic habit, in close vicinity to CD146+ cells and their processes. One of them (C, white arrowhead) occupies a cleavage plane between the endothelial wall and the adjacent reticular cell process. The asterisk indicates the cross-section of a point of branching of the main sinus, in which another CD146+ cell is located. E) 8 weeks post-transplantation. Morphology and position of CD146+ cells in the heterotopic marrow spaces undergoing hematopoietic seeding. Panels a,b demonstrate multiple CD146+ cells and hematopoietic cells. The nuclei (n) of at least three CD146+ cells are visible. They are clearly located outside of the sinusoid walls, as are the reticular process shown in panel c (blue arrow). Panel d shows a low power overview of a fully repopulated heterotopic marrow space in which CD146+ cells are seen both over sinus walls and away from them (blue arrows). F) Immunolabeling of 8wk transplant sections for human and murine endothelial markers. No labeling is observed in the sinusoidal wall, or outside of the sinusoids, for hCD34, hCD31, hVE-cadherin, or KDR, and hE-selectin or vWF (not shown). Uniform and restricted labeling of the sinusoid wall is observed with an antibody to murine CD31. Similar results were obtained at 4wk post-transplantation (not shown). Nuclear counterstain with hematoxylin. A-D, Nomarski optics.

Figure S7. CD146 and Ang-1 gene knockdown in CD146+ BMSCs A-C) Lenti-viral vector-mediated RNA interference in CD146+ BMSCs. CD146 (exon 15) and angiopoietin-1 siRNA sequences and control siRNA are shown. A) Schematic drawing of lenti-viral transfer vector in which the siRNA and control sequences were cloned. The expresssion of RNA interference sequences is controlled by the H1

promoter; the vectors contain an internal marker (eGFP) under transcriptional control of the EF-1α promoter. B) Western analysis of CD146 expression in mock-treated, control vector-infected (LV-siCtr), and LV-siCD146 infected BMSCs, demonstrating specific depletion of CD146 protein in LV-siCD146 infected BMSCs. C) Ang-1 ELISA in BMSCs infected with LV-siControl and LV-siAng-1 (exon 1) vectors, demonstrating depletion of secreted Ang-1 in cells with LV-directed expression of an Ang-1 specific RNA interfering sequence. D) Assembly and remodeling of pseudovascular cords in co-cultures of BMSCs and HUVEC. BMSCs and HUVEC generate a primary lattice of well-formed pseudovascular cords at 6h. Subsequent remodeling leads to cord elongation and ultimately disassembly and regression by 24h. CD146 silencing (siCD146) in BMSCs leads to a rudimentary lattice of short cords at 6h, which then grow into a system of long and thin cords that remain stable by 24h. Ang-1 silencing (siANG1) in BMSCs reduces the number of pseudovascular cords ab initio, with predominant formation of sheets and clumps. E) Cord length and numbers in co-cultures of BMSC and HUVEC cells expressed as mean + SD and compared by ANOVA.

Figure S8. Effects of MC-regulating factors on CD146 and Ang-1 expression in BMSCs, and on transfer of the HME Effects of TGF-β, FGF-2 and PDGF-BB on BMSC expression of CD146 and Ang-1 as assessed by qPCR (A), western analysis (B) and ELISA (C). Data are expressed as mean ± SD (p < 0.01). Both CD146 and Ang-1 are down regulated by FGF-2. D) Heterotopic ossicles formed by FGF-2-treated and control BMSCs. 8wk transplants of FGF-2 treated cells include bone. Hematopoiesis (hem), sinusoids (sin), or CD146+ cells (arrows) are only established in non-FGF-2 transplants. hac, HA/TCP carrier; m, megakaryocyte.

Supplemental References
Darland, D. C., and D'Amore, P. A. (2001). TGF beta is required for the formation of capillary-like structures in three-dimensional cocultures of 10T1/2 and endothelial cells. Angiogenesis 4, 11-20. Irizarry, R. A., Hobbs, B., Collin, F., Beazer-Barclay, Y. D., Antonellis, K. J., Scherf, U., and Speed, T. P. (2003). Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 4, 249-264. Jain, R. K. (2003). Molecular regulation of vessel maturation. Nat Med 9, 685-693. Kluk, M. J., and Hla, T. (2002). Signaling of sphingosine-1-phosphate via the S1P/EDG-family of G-protein-coupled receptors. Biochim Biophys Acta 1582, 72-80. Liu, W. M., Mei, R., Di, X., Ryder, T. B., Hubbell, E., Dee, S., Webster, T. A., Harrington, C. A., Ho, M. H., Baid, J., and Smeekens, S. P. (2002). Analysis of high density expression microarrays with signed-rank call algorithms. Bioinformatics 18, 1593-1599. Liu, Y., Wada, R., Yamashita, T., Mi, Y., Deng, C. X., Hobson, J. P., Rosenfeldt, H. M., Nava, V. E., Chae, S. S., Lee, M. J., et al. (2000). Edg-1, the G protein-coupled receptor for sphingosine-1-phosphate, is essential for vascular maturation. J Clin Invest 106, 951-961. Ozerdem, U., Grako, K. A., Dahlin-Huppe, K., Monosov, E., and Stallcup, W. B. (2001). NG2 proteoglycan is expressed exclusively by mural cells during vascular morphogenesis. Dev Dyn 222, 218-227. Piersanti, S., Sacchetti, B., Funari, A., Di Cesare, S., Bonci, D., Cherubini, G., Peschle, C., Riminucci, M., Bianco, P., and Saggio, I. (2006). Lentiviral transduction of human postnatal skeletal (stromal, mesenchymal) stem cells: in vivo transplantation and gene silencing. Calcif Tissue Int 78, 372-384. Reyes, M., Dudek, A., Jahagirdar, B., Koodie, L., Marker, P. H., and Verfaillie, C. M. (2002). Origin of endothelial progenitors in human postnatal bone marrow. J Clin Invest 109, 337-346. Steinberg, D., Pittman, R. C., and Carew, T. E. (1985). Mechanisms involved in the uptake and degradation of low density lipoprotein by the artery wall in vivo. Ann N Y Acad Sci 454, 195-206. Wedegaertner, P. B., Bourne, H. R., and von Zastrow, M. (1996). Activation-induced subcellular redistribution of Gs alpha. Mol Biol Cell 7, 1225-1233.

Table S1. Antibodies used for cell sorting and flow cytometry
Antigen Type
Label
Clone/Cat#
Distributor
CD14 (LPS receptor) MC PE M5E2 BD Biosciences, CD31 (PECAM-1) MC PE WM59 BD Biosciences, CD33 (P67) MC PECy5 WM53 BD Biosciences, CD41a MC PE HIP8 BD Biosciences CD44 MC PE 515 BD Biosciences CD34 (gp 105-120) MC FITC 581 BD Biosciences CD45 (LCA, T200) MC FITC HI30 BD Biosciences CD49a (VLA-1a, a1 integrin) MC NC SR84 BD Biosciences CD61 (GP IIIa, b3 integrin) MC FITC VI-PL2 BD Biosciences CD63 (LIMP, MLA1, gp55) MC PE H5C6 BD Biosciences CD71 (T9, Transferrin receptor) MC PECy5 M-A712 BD Biosciences CD90 (Thy-1) MC PECy5 5E10 BD Biosciences CD105 (Endoglin) MC NC 266 BD Biosciences CD117 (c-kit) MC PECy5 YB5.B8 BD Biosciences CD133 (AC133) MC NC 170411 R&D Systems CD140b (PDGF receptor beta) MC NC 28 BD Biosciences CD144 (VE-cadherin, cadherin5) MC PE TEA1/31 Beckman Coulter CD146 (MUC-18, S-endo 1) MC PE P1H12 BD Biosciences α-smooth muscle actin MC NC 1A4 R&D Systems ALP MC NC B4-78 R&D Systems STRO-1 MC NC Stro-1 R&D Systems KDR (Flk-1, VEGF R2) PC APC 89106 R&D Systems Flt-1 (VEGF R1) PC PE 49560 R&D Systems CD62E (E-Selectin) MC FITC 1.2B6 Chemicon NGF receptor MC PE C40-1457 BD Biosciences MC, mouse monoclonal; PC, rabbit polyclonal antiserum; PE, phycoerythrin; FITC, fluorescein isothiocyanate; NC, non-conjugated; APC, allophycocyanin; PECy5, Phycoerythrin-Cy5

Table S2. CFE (CFU-Fs) for unsorted bone marrow nucleated cells (BMNCs) and sorted fractions
Cells plated
Cell
density (cells/cm2)
CFU-F
Unsorted CD45+ CD45- CD45-/CD146- CD45-/CD146+ 1x102 1.6 0 0 0 0 2.6 + 0.8 5x103 84.7 0 0 0 0 ND 5x105 8.4x103 12.9 + 2.3 0 55+8 0 ND
CFU-F, Colony Forming Unit-Fibroblast. Data are expressed as mean + SEM and indicate an
enrichment factor (CFECD45-/CD146+ /CFEunsorted) of 8.3 x 102 in the sorted population compared
to unfractionated BMNCs.

Table S3. Recovery of CFU-Fs in the CD45-/CD146+ fraction of bone marrow cells
Unsorted CD45-/CD146+
#/106 BMNCs 25.8 + 4.6 20.8 + 8.5 Recovery (%) 100 80.6
Enrichment 8.3 x 102
BMNCs, Total bone marrow nucleated cells. Data obtained from CFE assays (table 2) are normalized to 106 BMNCs, based on a 0.08% frequency of CD45-/CD146+ cells in total BMNCs.
Table S4. Clonogenic efficiency (CFU-F) of BM CD34+ cells
A) CFE for bone marrow cells sorted for CD34 (MACS) Cells plated
Cell density (cells/cm2)
CFU-F
Unsorted CD34+ CD34- 1x102 1.6 0 0 0 3x103 50 0 0 0 3x104 50x102 0 0 0 5x105 8.3x103 14.9 + 1.3 ND 6+2
B) CFE for bone marrow cells sorted for CD146 and CD34 (FACS)
Cells plated
Cell density (cells/cm2)
CFU-F
Unsorted CD146+/CD34+ CD146-/CD34+ 1x102 1.6 0 0 0 3x103 48 0 0 0 3x104 4.8x103 0 ND 0 5x105 8.4x103 9+2 ND ND
CFU-F, Colony Forming Unit-Fibroblast. Data are expressed as mean + SEM.

Table S5. Transplantation of single CD146+ CFU-F derived strains
Clone
Bone Human osteoblasts
Marrow adipocytes
Sinusoids Hematopoiesis
CD146+ cells
A27 – 4 + + + + A27 – 5 - - - - A27 - 9 + + + + A27 - 11 - - - -
Table S6. Secondary passage of CD146+ CFU-Fs
iCFU-F Ossicles Cells
recovered (x106)
hCells recovered
CD146+
(%) CD44+ plated
CD146+ plated
sCFU-F detected
(CD146+)
sCFU-F Total
10 8 2.0 2x104 0.2 2x103 ND 1 10 12 6 1.5 1.5x104 0.3 5x103 ND 6 18 1 4 2.0 0.14 ND 2100 3 4 1 1 0.4 0.09 ND ND ND ND 1 1 0.3 0.16 ND ND ND ND 1 4 1.5 0.4 ND 4000 2 3 1 1 0.5 ND ND ND ND ND
iCFU-F, culture-initiating CFU-Fs. Ossicles, number of heterotopic ossicles generated (1 ossicle 2x106 transplanted cells). Cells recovered, total human/murine cells released by collagenase. hCells recovered, absolute number of hCD44/hCD90 double positive cells. CD146+(%), CD146+ fraction in the total recovered population. CD44+ plated, number of CD44+ sorted human cells used for CFU assay. CD146+ plated, number of CD146+ sorted human cells used for CFU assay. sCFU-F detected, secondary CFU-F, directly assayed. sCFU-F Total, secondary CFU-Fs, normalized for the number of recovered cells
Table S7. Antibodies used for immunohistochemistry and western blotting
Antigen Type Cat# Distributor Tie2 GP AF313 R&D Systems CD146 (IHC) MM NCL-CD146 Novocastra CD146 (Western blot) GP SC-18942 Santa Cruz Biotechnology Human Mitochondria MM MAB-1273 Chemicon CD105 (Endoglin) MM 555690 BD Biosciences Alkaline Phosphatase (LF-47) RP --- Gift from LW Fisher, NIDCR, NIH Ang-1 MM MAB923 R&D Systems Human CD34 MM H7168 Dako Human E-selectin MM BBA16 R&D Systems CD150 MM NCL-SLAM Novocastra FGFR3 MM Sc-13121 Santa Cruz Biotechnology TGFβ1 MM MAB1032 Chemicon FGF2 RP Sc-79 Santa Cruz Biotechnology Human VE-cadherin MM MAB1989 Chemicon Murine CD31 RaM 550274 BD Biosciences Human CD31 MM 08-1425 Histoline KDR GP AF357 R&D Systems Actin MC sc-8432 Santa Cruz Biotechnology
MC, mouse monoclonal; RP, rabbit polyclonal; GP, goat polyclonal; RaM, rat monoclonal; IHC, Immunohistochemistry

Table S8. Primers for RT-PCR
Name Sequence Description and GenBank accession number Product
size
GAPDH-F 5'-CGG GAA GCT TGT GAT CAA TGG-3'
GAPDH-R 5'-GGC AGT GAT GGC ATG GAC TG-3'
Human glyceraldehyde-3-phosphate dehydrogenase
[M17851] 358
VE-Cadherin s 5'-GAC TGA CCA TCA TGC CCT CT-3'
VE-Cadherin as 5'-GAA CAT CTG CCC CTT CTC AG-3'
Homo sapiens cadherin 5, type 2, VE-cadherin (vascular
epithelium)(CDH5) [NM_001795] {Son, 2006 #17} 400
Flk1-KDR-VEGFR2 s 5'-GTG ACC AAC ATG GAG TCG TG-3'
Flk1-KDR-VEGFR2 as 5'-CCA GAG ATT CCA TGC CAC TT-3'
Homo sapiens vascular endothelial growth factor receptor
2 (KDR) [AF063658] {Son, 2006 #17} 660
CD31 for 5'-GCC TTC AAC AGA GCC AAC CA -3'
CD31 rev 5'-CGG CAT CAG GGA CAG CTT T -3'
Homo sapiens platelet/endothelial cell adhesion molecule
(CD31 antigen) (PECAM1) [NM_000442] 451
CD34 for 5'-TGA GTT TGC TGC CTT CTG GG-3'
CD34 rev 5'-CCG CAC AGC TGG AGG TCT TA-3'
Homo sapiens CD34 molecule (CD34), transcript variant 1
[NM_001025109] 534
CD133 for 5'-CGC GTG ATT TCC CAG AAG AT-3'
CD133 rev 5'-ATC GCT GTT GCC ATG GAC TT-3' Homo sapiens prominin 1 [NM_006017] 586
ACTA2hu-for 5’-ACT GGC ATC GTG CTG GAC TC-3’
ACTA2hu-rev 5’-CCA GCA GAC TCC ATC CCG AT-3’
Homo sapiens actin, alpha 2, smooth muscle, aorta
(ACTA2) [NM_001613] 374
CSPG4hu-for 5’-GCA GAG GTC TAC GCT GGG AAT-3’
CSPG4hu-rev 5’-TGG AGG TGG GCA CGA AAG T-3’
Homo sapiens chondroitin sulfate proteoglycan 4
[NM_001897] 470
CALD1hu-for 5’-TCG AAG CAG AAA GAA TCG CC-3’
CALD1hu-rev 5’-CTT CAG CAT CCC TCC AAT CAT T-3’
Homo sapiens caldesmon 1 (CALD1), transcript variant 1
[NM_033138] 486
CNN2hu-for 5’-GGG AAG GCC AAG ACT AAG GG-3’
CNN2hu-rev 5’-TAG CCC ATC TGC AGG GAC A-3’
Homo sapiens calponin 2 (CNN2), transcript variant 1
[NM_004368] 368
DEShu-for 5’-GCA TGA AGA GGA GAT CCG TGA-3’
DEShu-rev 5’-GTC CTG GTA CTC GCG CAG AT-3’ Homo sapiens desmin (DES) [NM_001927] 445
COL4A1hu-for 5’-AGG GCA TGC CTG GTA TTG G-3’
COL4A1hu-rev 5’-CAC AAG GAA GCC GTG ATC AAC-3’
Homo sapiens collagen, type IV, alpha 1 (COL4A1)
[NM_001845] 479
COL4A2hu-for 5’-CCA TCC AGC CAG GTT GCA TA-3’
COL4A2hu-rev 5’-CAT CCC TTG GCT TCC TCT GAA T-3’
Homo sapiens collagen, type IV, alpha 2 (COL4A2)
[NM_001846] 413
PDGFRBhu-for 5’-CAG ATC AAT GTC CCT GTC CGA-3’
PDGFRBhu-rev 5’-AAA GGG CAA GGA GTG TGG C-3’
Homo sapiens platelet-derived growth factor receptor, beta
polypeptide (PDGFRB) [NM_002609] 357
EDG1hu-for 5’-CTT GAG CGA GGC TGC GGT-3’
EDG1hu-rev 5’-GGG TCG GTG GAA TTT CTT GG-3’
Homo sapiens endothelial differentiation, sphingolipid G-
protein-coupled receptor, 1 (EDG1) [NM_001400] 410
ITGA1hu-for 5’-AAG TGC AAC AAG TGA CAG CGA-3’
ITGA1hu-rev 5’-TTG CAG TCC AGA ATT GTG CC-3’ Homo sapiens integrin, alpha 1 (ITGA1) [NM_181501] 428
ENGhu-for 5’-TGC ACT TGG CCT ACA ATT CCA-3’
ENGhu-rev 5’-TCT CCA GTG GTC CAG ATC TGC-3’
Homo sapiens endoglin (Osler-Rendu-Weber syndrome 1)
(ENG) [NM_000118] 481

Table S9. Genes regulating cell cycle progression (Figure S3) – Probe IDs
1552263_at 1552264_a_at 1552329_at 1552637_at 1552982_a_at 1553096_s_at 1553112_s_at 1553113_s_at 1553387_at 1553530_a_at 1553574_at 1554155_at 1554264_at 1554365_a_at 1554411_at 1554420_at 1554600_s_at 1554726_at 1554768_a_at 1554874_at 1554906_a_at 1555004_a_at 1555016_at 1555086_at 1555155_at 1555167_s_at 1555259_at 1555278_a_at 1555378_at 1555416_a_at 1555514_a_at 1555758_a_at 1555814_a_at 1555826_at 1556043_a_at 1556057_s_at 1556213_a_at 1556227_at 1556228_a_at 1556256_a_at 1556338_at 1556339_a_at 1556595_at 1556621_a_at 1556931_at 1557193_at 1557452_at 1557543_at 1557555_at 1557718_at 1557769_at 1557813_at 1557905_s_at 1557964_at 1558208_at 1558475_at 1559218_s_at 1559227_s_at 1559282_at 1559307_s_at 1559529_at 1559622_at 1559633_a_at 1559634_at 1559856_s_at 1559936_at 1560074_at 1560445_x_at 1561042_at
1561144_at 1561690_at 1561867_at 1562031_at 1562168_at 1562390_at 1562454_at 1563101_at 1563468_at 1563709_at 1564520_s_at 1564521_x_at 1564836_at 1565483_at 1565693_at 1565868_at 1566043_at 1566105_at 1566485_at 1567213_at 1567214_a_at 1567443_x_at 1568012_at 1568943_at 1569073_x_at 1569487_at 1569508_at 1569642_at 1570350_at 1570352_at 1570507_at 1598_g_at 1861_at 200010_at 200020_at 200040_at 200043_at 200063_s_at 200098_s_at 200602_at 200622_x_at 200623_s_at 200653_s_at 200655_s_at 200658_s_at 200659_s_at 200678_x_at 200695_at 200709_at 200712_s_at 200749_at 200750_s_at 200772_x_at 200792_at 200812_at 200813_s_at 200816_s_at 200836_s_at 200877_at 200913_at 200932_s_at 200943_at 200944_s_at 200953_s_at 200956_s_at 200957_s_at 201012_at 201041_s_at 201077_s_at
201111_at 201112_s_at 201131_s_at 201155_s_at 201190_s_at 201191_at 201192_s_at 201202_at 201234_at 201331_s_at 201371_s_at 201372_s_at 201390_s_at 201423_s_at 201424_s_at 201456_s_at 201457_x_at 201458_s_at 201464_x_at 201465_s_at 201466_s_at 201473_at 201479_at 201482_at 201488_x_at 201502_s_at 201528_at 201529_s_at 201533_at 201558_at 201601_x_at 201652_at 201693_s_at 201697_s_at 201700_at 201725_at 201755_at 201758_at 201763_s_at 201774_s_at 201783_s_at 201853_s_at 201856_s_at 201857_at 201865_x_at 201866_s_at 201877_s_at 201939_at 201975_at 201979_s_at 201983_s_at 201984_s_at 202014_at 202094_at 202095_s_at 202102_s_at 202107_s_at 202149_at 202154_x_at 202160_at 202183_s_at 202191_s_at 202216_x_at 202221_s_at 202224_at 202226_s_at 202240_at 202246_s_at 202248_at
202268_s_at 202281_at 202288_at 202312_s_at 202327_s_at 202329_at 202340_x_at 202424_at 202431_s_at 202454_s_at 202455_at 202474_s_at 202514_at 202515_at 202516_s_at 202520_s_at 202530_at 202531_at 202580_x_at 202587_s_at 202588_at 202642_s_at 202645_s_at 202647_s_at 202652_at 202670_at 202672_s_at 202686_s_at 202687_s_at 202688_at 202717_s_at 202726_at 202762_at 202763_at 202770_s_at 202819_s_at 202820_at 202834_at 202859_x_at 202870_s_at 202892_at 2028_s_at 202906_s_at 202907_s_at 202927_at 202934_at 203050_at 203055_s_at 203065_s_at 203079_s_at 203084_at 203085_s_at 203132_at 203175_at 203198_at 203214_x_at 203297_s_at 203298_s_at 203301_s_at 203332_s_at 203362_s_at 203381_s_at 203382_s_at 203411_s_at 203448_s_at 203449_s_at 203460_s_at 203533_s_at 203547_at
203554_x_at 203601_s_at 203602_s_at 203665_at 203666_at 203685_at 203688_at 203692_s_at 203693_s_at 203694_s_at 203716_s_at 203717_at 203725_at 203740_at 203749_s_at 203777_s_at 203787_at 203792_x_at 203793_x_at 203805_s_at 203806_s_at 203844_at 203845_at 203847_s_at 203848_at 203853_s_at 203882_at 203966_s_at 203973_s_at 203975_s_at 203976_s_at 203989_x_at 204009_s_at 204020_at 204021_s_at 204026_s_at 204032_at 204039_at 204053_x_at 204054_at 204092_s_at 204131_s_at 204132_s_at 204164_at 204170_s_at 204188_s_at 204206_at 204244_s_at 204246_s_at 204252_at 204262_s_at 204267_x_at 204312_x_at 204313_s_at 204346_s_at 204420_at 204444_at 204457_s_at 204489_s_at 204490_s_at 204510_at 204531_s_at 204566_at 204621_s_at 204622_x_at 204641_at 204642_at 204650_s_at
204686_at 204696_s_at 204752_x_at 204781_s_at 204784_s_at 204802_at 204812_at 204822_at 204826_at 204827_s_at 204831_at 204853_at 204857_at 204880_at 204891_s_at 204901_at 204907_s_at 204908_s_at 204947_at 204958_at 205015_s_at 205032_at 205034_at 205046_at 205051_s_at 205067_at 205178_s_at 205207_at 205215_at 205225_at 205266_at 205393_s_at 205394_at 205396_at 205397_x_at 205398_s_at 205449_at 205451_at 205500_at 205516_x_at 205526_s_at 205548_s_at 205623_at 205646_s_at 205655_at 205672_at 205681_at 205754_at 205782_at 205841_at 205842_s_at 205867_at 205868_s_at 205883_at 205892_s_at 205895_s_at 206036_s_at 206054_at 206075_s_at 206219_s_at 206267_s_at 206316_s_at 206332_s_at 206430_at 206474_at 206486_at 206492_at 206503_x_at 206543_at

206544_x_at 206552_s_at 206569_at 206598_at 206661_at 206665_s_at 206675_s_at 206693_at 206714_at 206783_at 206847_s_at 206848_at 206923_at 207002_s_at 207023_x_at 207029_at 207039_at 207042_at 207113_s_at 207178_s_at 207188_at 207243_s_at 207326_at 207361_at 207382_at 207428_x_at 207556_s_at 207591_s_at 207614_s_at 207630_s_at 207749_s_at 207766_at 207782_s_at 207808_s_at 207821_s_at 207822_at 207823_s_at 207828_s_at 207845_s_at 207848_at 207849_at 207851_s_at 207937_x_at 207943_x_at 207978_s_at 208000_at 208033_s_at 208071_s_at 208079_s_at 208080_at 208121_s_at 208351_s_at 208372_s_at 208403_x_at 208414_s_at 208510_s_at 208530_s_at 208644_at 208652_at 208656_s_at 208676_s_at 208697_s_at 208711_s_at 208712_at 208721_s_at 208722_s_at 208774_at 208791_at 208792_s_at 208793_x_at 208796_s_at 208820_at
208823_s_at 208889_s_at 208937_s_at 208965_s_at 208966_x_at 208992_s_at 209014_at 209026_x_at 209102_s_at 209112_at 209113_s_at 209115_at 209151_x_at 209152_s_at 209157_at 209165_at 209172_s_at 209189_at 209239_at 209260_at 209318_x_at 209331_s_at 209332_s_at 209350_s_at 209359_x_at 209361_s_at 209364_at 209375_at 209408_at 209414_at 209416_s_at 209421_at 209425_at 209464_at 209529_at 209591_s_at 209632_at 209633_at 209642_at 209644_x_at 209658_at 209659_s_at 209665_at 209687_at 209716_at 209798_at 209811_at 209827_s_at 209835_x_at 209863_s_at 209875_s_at 209878_s_at 209891_at 209895_at 209896_s_at 209952_s_at 209960_at 209961_s_at 209967_s_at 209974_s_at 209999_x_at 210001_s_at 210038_at 210039_s_at 210051_at 210052_s_at 210141_s_at 210186_s_at 210187_at 210211_s_at 210226_at 210229_s_at
210232_at 210317_s_at 210334_x_at 210338_s_at 210362_x_at 210365_at 210407_at 210416_s_at 210440_s_at 210477_x_at 210512_s_at 210513_s_at 210557_x_at 210559_s_at 210570_x_at 210607_at 210644_s_at 210649_s_at 210655_s_at 210695_s_at 210716_s_at 210719_s_at 210728_s_at 210734_x_at 210755_at 210756_s_at 210765_at 210766_s_at 210776_x_at 210829_s_at 210845_s_at 210858_x_at 210889_s_at 210894_s_at 210916_s_at 210930_s_at 210962_s_at 210973_s_at 210984_x_at 210996_s_at 210997_at 210998_s_at 211036_x_at 211080_s_at 211087_x_at 211110_s_at 211124_s_at 211143_x_at 211154_at 211179_at 211193_at 211234_x_at 211251_x_at 211259_s_at 211277_x_at 211284_s_at 211297_s_at 211300_s_at 211318_s_at 211338_at 211373_s_at 211419_s_at 211504_x_at 211506_s_at 211518_s_at 211519_s_at 211527_x_at 211547_s_at 211553_x_at 211554_s_at 211559_s_at 211561_x_at
211584_s_at 211589_at 211596_s_at 211607_x_at 211620_x_at 211621_at 211671_s_at 211711_s_at 211714_x_at 211808_s_at 211814_s_at 211831_s_at 211839_s_at 211851_x_at 211865_s_at 211921_x_at 211924_s_at 211949_s_at 211951_at 211968_s_at 211969_at 211985_s_at 212014_x_at 212020_s_at 212021_s_at 212022_s_at 212023_s_at 212036_s_at 212046_x_at 212063_at 212072_s_at 212076_at 212079_s_at 212080_at 212086_x_at 212097_at 212152_x_at 212167_s_at 212171_x_at 212239_at 212240_s_at 212257_s_at 212271_at 212312_at 212330_at 212377_s_at 212533_at 212549_at 212550_at 212566_at 212567_s_at 212610_at 212672_at 212688_at 212781_at 212783_at 212825_at 212899_at 213006_at 213093_at 213095_x_at 213134_x_at 213136_at 213137_s_at 213147_at 213217_at 213226_at 213286_at 213305_s_at 213337_s_at 213385_at 213419_at
213476_x_at 213490_s_at 213551_x_at 213560_at 213579_s_at 213655_at 213710_s_at 213720_s_at 213731_s_at 213795_s_at 213799_s_at 213921_at 213966_at 213976_at 213977_s_at 213987_s_at 214022_s_at 214063_s_at 214083_at 214108_at 214119_s_at 214239_x_at 214360_at 214384_s_at 214426_x_at 214427_at 214442_s_at 214508_x_at 214513_s_at 214637_at 214683_s_at 214710_s_at 214794_at 214853_s_at 214866_at 214908_s_at 214953_s_at 215037_s_at 215195_at 215308_at 215346_at 215483_at 215508_at 215509_s_at 215552_s_at 215638_at 215705_at 215711_s_at 215714_s_at 215719_x_at 215731_s_at 215773_x_at 215828_at 215878_at 215879_at 215988_s_at 216038_x_at 216041_x_at 216056_at 216091_s_at 216178_x_at 216205_s_at 216212_s_at 216237_s_at 216248_s_at 216252_x_at 216275_at 216277_at 216321_s_at 216326_s_at 216491_x_at 216624_s_at
216647_at 216854_at 216862_s_at 216953_s_at 216955_at 216969_s_at 216974_at 217010_s_at 217113_at 217178_at 217190_x_at 217208_s_at 217254_s_at 217299_s_at 217373_x_at 217389_s_at 217523_at 217620_s_at 217782_s_at 217786_at 217802_s_at 217850_at 217988_at 218117_at 218234_at 218252_at 218284_at 218308_at 218350_s_at 218386_x_at 218414_s_at 218467_at 218529_at 218750_at 218803_at 218875_s_at 218878_s_at 218902_at 218917_s_at 219028_at 219077_s_at 219105_x_at 219157_at 219257_s_at 219264_s_at 219350_s_at 219370_at 219592_at 219769_at 219810_at 219813_at 219966_x_at 220266_s_at 220658_s_at 220789_s_at 220792_at 221085_at 221264_s_at 221282_x_at 221308_at 221496_s_at 221520_s_at 221548_s_at 221557_s_at 221558_s_at 221580_s_at 221586_s_at 221691_x_at 221841_s_at 221891_x_at 221918_at 221965_at

222013_x_at 222033_s_at 222034_at 222043_at 222164_at 222176_at 222243_s_at 222258_s_at 222292_at 222305_at 222374_at 222424_s_at 222728_s_at 222757_s_at 223163_s_at 223198_x_at 223302_s_at 223322_at 223329_x_at 223330_s_at 223381_at 223502_s_at 223519_at 223545_at 223586_at 223802_s_at 223819_x_at 223828_s_at 223850_at 224065_at 224185_at 224187_x_at 224204_x_at 224309_s_at 224385_s_at 224387_at 224471_s_at 224620_at 224657_at 224753_at 224784_at 224806_at 224889_at 224891_at 224999_at 225097_at 225115_at 225116_at 225297_at 225368_at 225531_at 225532_at 225662_at 225665_at 225740_x_at 225742_at 225912_at 225945_at 226036_x_at 226213_at 226315_at 226323_at 226400_at 226442_at 226534_at 226705_at 226880_at 226917_s_at 226981_at 226994_at 227249_at 227404_s_at
227684_at 228037_at 228052_x_at 228081_at 228243_at 228484_s_at 228729_at 228758_at 228814_at 228822_s_at 228848_at 228899_at 228904_at 229216_s_at 229353_s_at 229408_at 229657_at 229703_at 229729_at 229756_at 229825_at 230170_at 230219_at 230229_at 230393_at 230562_at 230938_x_at 231168_at 231177_at 231198_at 231237_x_at 231370_at 231481_at 231673_at 231693_at 231767_at 231777_at 231801_at 231938_at 232120_at 232210_at 232231_at 232266_x_at 232331_at 232375_at 232379_at 232431_at 232466_at 232524_x_at 232541_at 232546_at 232551_at 232614_at 232640_at 232660_at 232876_at 232925_at 232967_at 233011_at 233223_at 233254_x_at 233313_at 233314_at 233530_at 233545_at 233629_at 233674_at 233752_s_at 233783_at 233833_at 234159_at 234211_at
234215_at 234393_at 234411_x_at 234838_at 234863_x_at 234895_at 235066_at 235135_at 235226_at 235392_at 235419_at 235429_at 235582_at 235602_at 235737_at 235739_at 235753_at 235764_at 235785_at 235858_at 235926_at 236025_at 236094_at 236341_at 236495_at 236512_at 236520_at 236645_at 236669_at 236675_at 236729_at 236814_at 236859_at 236934_at 237015_at 237030_at 237181_at 237239_at 237376_at 237424_at 237548_at 237571_at 237837_at 237985_at 238208_at 238339_x_at 238405_at 238486_at 238757_at 239000_at 239121_at 239124_at 239156_at 239248_at 239249_at 239448_at 239555_at 239582_at 239585_at 239692_at 239804_at 239826_at 239829_at 239908_at 239939_at 240019_at 240033_at 240129_at 240278_at 240360_at 240370_at
240491_at 240543_at 240618_at 240621_at 240630_at 240674_at 240732_at 240752_at 240792_at 240793_at 240953_at 241038_at 241069_at 241387_at 241524_at 241545_x_at 241724_x_at 241756_at 241791_at 241824_at 241855_s_at 241876_at 241959_at 241982_at 241983_at 242099_at 242106_at 242152_at 242194_at 242240_at 242320_at 242359_at 242362_at 242425_at 242560_at 242567_at 242599_at 242679_at 242693_at 242743_at 242939_at 242952_at 243002_at 243296_at 243624_at 243640_x_at 243655_x_at 243677_at 243_g_at 244023_at 244035_at 244054_at 244225_x_at 244263_at 244373_at 244536_at 244550_at 244658_at 244721_at 244790_at 244836_at 244839_at 244862_at 244869_at 33148_at 33322_i_at 33323_r_at 33850_at 34449_at 35150_at 37028_at
37433_at 38158_at 38290_at 38707_r_at 39402_at 40148_at 40225_at 40273_at 48825_at




















