
The cuticle proteins of Drosophila melanogaster: Stage specificity
Upload
independentCategory
view
3download
0
General and Comparative Endocrinology 191 (2013) 137–145
Contents lists available at SciVerse ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen
Selective elimination/RNAi silencing of FMRF-related peptides and theirreceptors decreases the locomotor activity in Drosophila melanogaster
0016-6480/$ - see front matter � 2013 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.ygcen.2013.05.023
⇑ Corresponding author.E-mail address: [email protected] (I. Kiss).
Brigitta Kiss a, Tamás Szlanka a,b, Ágnes Zvara a, Michal Zurovec d, Michal Sery d, Štefan Kakaš d,Beáta Ramasz a, Zoltán Heged}us c, Tamás Lukacsovich e, László Puskás a, Adrien Fónagy f, István Kiss a,⇑a Institute of Genetics, Biological Research Centre, Hungarian Academy of Sciences, Temesvári krt. 62, H-6726 Szeged, Hungaryb Institute of Biochemistry, Biological Research Centre, Hungarian Academy of Sciences, Temesvári krt. 62, H-6726 Szeged, Hungaryc Institute of Biophysics, Biological Research Centre, Hungarian Academy of Sciences, Temesvári krt. 62, H-6726 Szeged, Hungaryd Biology Centre, Czech Academy of Sciences, Branisovská 31, 37005 Ceské Budejovice, Czech Republice Dept. of Developmental and Cell Biology, University of California, Irvine, USAf Plant Protection Institute, Centre for Agricultural Research, Hungarian Academy of Sciences, Herman Ottó u. 15, H-1022 Budapest, Hungary
a r t i c l e i n f o a b s t r a c t
Article history:Available online 11 June 2013
Keywords:Drosophila melanogasterFMRF-related peptidesG protein-coupled receptorsRNAi-silencingUAS-reaper ablationLocomotor activity
Five neuropeptide genes are classified in the FMRF-related (FaRP) group: the Fmrf, dromyosuppressin(Dms), drosulfakinin (Dsk), neuropeptide F (npf) and short neuropeptide F (sNPF) genes coding for 8, 1, 2,1 and 4 peptides, respectively. In order to compare their effects on the locomotor activity of Drosophilaadults, we made RNAi knockdown of the peptides and their specific receptor genes. In addition, we con-structed Gal4 drivers with three distinct parts of the Fmrf gene’s 50 regulatory sequence (RS8-Gal4, RS11-Gal4, RS17-Gal4), and used them to ablate FMRF-positive neurons inducing apoptosis by expressing thereaper (rpr) gene. We examined the locomotor activity of flies by measuring the mean velocity of move-ment (MVM) following repeated air-puffs. Locomotor activity was decreased by RNAi knockdowninduced in the CNS by the elav-Gal4 driver. According to the MVM curve profiles, RNAi knockdown mosteffectively decreased the velocity when the DmsR-1 and DmsR-2 genes were silenced together (DmsR-1-RNAi/elav-Gal4; DmsR-2-RNAi/+). Similar effect was observed in Dsk-RNAi/ elav-Gal4; DskR-2-RNAi/+, whilemoderate effects were found in three other combinations (Fmrf-RNAi/elav-Gal4; FR-RNAi/+, Dms-RNAi/elav-Gal4;DmsR-2-RNAi/+, CCKLR-17D1-RNAi/elav-Gal4; CCKLR-17D3-RNAi/+), and weak effect in DmsR-2-RNAi/elav-Gal4; DmsR-1-RNAi/+. Male and female flies were not different in this respect. In the cell abla-tion experiment, the MVM profiles of the female flies were different from the controls when the UAS-rprtransgene was driven by RS8-Gal4 or RS17-Gal4. The RS11-Gal4 and Fmrf-Gal4 drivers were ineffective. Inthe males only the RS17-Gal4 showed a weak effect. RNAi silencing of the FaRP and FaRP-receptor geneseffectively decreased the startle-induced locomotor activity of flies. Ablation of FMRF-positive neurons bythe RS8-Gal4 and/or RS17-Gal4 drivers also decreased the flies’ activity.
� 2013 Elsevier Inc. All rights reserved.
1. Introduction melanogaster, the genetically best known organism among the
Specific neurons in the central nervous system (CNS) produceand secrete peptides and biogenic amines. Some of these neuro-peptides reach the target organ through the circulation and actas neurohormones while others remain within the CNS and modifythe activity of other neurons as neuromodulators. Neuropeptidesconstitute a large group of signaling molecules. They influencemany important processes of life in all Metazoans, like neurotrans-mission, nociception, feeding and metabolism, energy and osmotichomeostasis, learning and memory, etc. Insects as well use a pleth-ora of neuropeptides to control and modulate physiological, devel-opmental and behavioral events. The fruitfly Drosophila
higher Eukaryotes, offers an excellent model system for studyingthe action mechanism of neuropeptides.
1.1. Drosophila neuropeptides and their GPCR receptors
The availability of Drosophila genome sequence (Adams et al.,2000), has given a large impetus to the neuropeptide field as well:within a few years, many of the putative peptide and receptorgenes have been identified in the fruitfly (Hewes and Taghert,2007; Nässel and Winther, 2010). Bioinformatic approach pre-dicted 119 peptide precursor genes in the Drosophila genome.Out of them, at least 42 genes may code for ‘‘prepropeptide’’ pro-teins containing one or more copies of identical and/or closely re-lated peptide sequences flanked by proteolytic cutting sites. Thepeptidomic analysis has verified 46 neuropeptides which are de-rived from 19 of these precursor genes (Clynen et al., 2010).

138 B. Kiss et al. / General and Comparative Endocrinology 191 (2013) 137–145
Almost all the receptors which bind peptide ligands belong tothe G protein-coupled receptors (GPCRs). GPCRs represent one ofthe largest protein family, their genes comprise 1.5% of theDrosophila genome (Brody and Cravchik, 2000). Currently about160 GPCRs are known in Drosophila, of which 44 are known to bindpeptide ligands (Hewes and Taghert, 2007; Hauser et al., 2006a,b;Nässel and Winther, 2010). However, much less is known aboutthe signaling pathways beyond the neuropeptide ligands and thecognate receptors.
1.2. FMRFamide-related neuropeptides (FaRPs) in Drosophilamelanogaster
The Drosophila Fmrf gene codes for a prepropeptide which con-tains six peptide sequences sharing the characteristic FMRF aminoacid sequence at the C-terminus, and two other peptides withmore divergent C-terminal sequences (Wegener and Gorbashov,2008; Nässel and Winther, 2010). In addition to FMRF, neuropep-tides derived from four other genes have the RF motif at theirC-terminal end. Traditionally, these five neuropeptide genes areclassified as the FMRF-related group (FaRP) comprising the Fmrf(CG2346), dromyosuppressin (Dms, CG6440), drosulfakinin (Dsk,CG18090), neuropeptide F (npf, CG10342) and short neuropeptide F(sNPF, CG13968) genes (Nässel and Winther, 2010, and Flybase(http://flybase.org). The Dms and npf genes code for one peptideeach, while sNPF and Dsk code for five and three peptides,respectively. The DSK1 and DSK2 peptides have been verified bio-chemically, DSK0 is putative because it has not been confirmed asan expressed peptide. Interestingly, Palmer et al. (2007) recentlyfound that the DSK0 peptide induced crop contractions. All thesepeptides have a conserved C-terminus with the terminal RF-amidesequence, and a divergent N terminus which could be responsiblefor the specific biological effects.
The FaRP peptides have characteristic myoactivities, but theyare not related functionally: their target organs, effective thresh-olds, time-course and expression patterns are different (Nicholset al., 2002). Drosophila FMRF peptides generally stimulate musclecontraction in the heart, crop, foregut and body wall (Nichols,2003. DMS is a myoinhibitory peptide decreasing e.g. the move-ment of the heart and adult foregut/crop (Kaminski et al., 2002).DSK peptides stimulate the contractions of the heart and crop (Nic-hols et al., 2002, 2009), and increase larval locomotion activity andinduces escape response (Chen et al., 2012). DSKs and their CCKLRreceptors are required for the growth of the neuromuscular junc-tions (Chen and Ganetzky, 2012).
All the FaRPs are expressed in the CNS in neurons arranged inbilaterally symmetric patterns, which are characteristic for eachneuropeptide (Santos et al., 2007; Park et al., 2008; Nässel andWinther, 2010). The FMRF peptides are expressed in the DrosophilaCNS in 17 various cell types, in about 60 neurons (Schneider et al.,1991, 1993). Even peptides which are derived from the same FMRFprepropeptide protein by differential processing can show differ-ent tissue-specific expression: the SDNFMRF, DPKQDFMRF andTPAEDFMRF peptides are expressed in similar patterns in the thor-aco-abdominal ganglion, but TPAEDFMRF is additionally expressedin the adult lateral protocerebrum and the gut (Nichols et al.,1999). DMS is expressed in the CNS but could be detected by im-muno-staining in the crop and gut (‘‘brain-gut peptide’’, McCor-mick and Nichols, 1993; Merte and Nichols, 2002). DSK is alsoexpressed in the CNS (Nichols and Lim, 1996). DSK is expressedin some of the DILP (Drosophila insulin-like peptide)-producingneurons which have axonal connection to neurohemal areas inthe corpora cardiaca, aorta and the anterior intestine. DILP andDSK together control the feeding behavior as hormonal satiety sig-nals (Söderberg et al., 2012). NPF and sNPF are expressed both inthe CNS and the midgut (Veenstra, 2009). sNPF is also expressed
in several thousand CNS neurons and consequently exerts a broadspectrum of modulatory effects (Nässel et al., 2008).
The determination of the neuronal cell fate and the neuropep-tide expression patterns in the CNS are regulated by specificenhancers binding transcription factors in the genes’ regulatoryregions (Benveniste and Taghert, 1999; Vanden Broeck, 2001).Specification of the identity of single neurons in the CNS is a multi-step process which builds up a combinatorial code of the activatedenhancers (Baumgardt et al., 2007). GPCR receptors of the neuro-peptides and neurohormones are expressed on a broader scale inthe target cells and organs (Cazzamali and Grimmelikhuijzen,2002; Duttlinger et al., 2002; Nichols, 2003; Bendena et al., 2012).
The experiments described here address the effects exerted bysome of the FaRP peptides (FMRF, DMS, DSK) and receptors onthe locomotor activity of Drosophila adults. For this purpose, weused two genetic approaches (i) RNAi silencing of FMRF-relatedpeptides and their receptors: and (ii) construction of new Fmrf-Gal4 drivers which represent specific parts of the FMRF spatialexpression in the CNS, and their application to remove the corre-sponding neural cells by reaper (rpr)-induced apoptosis. The resultsshowed that these treatments decreased the locomotor activity ofthe flies in general.
2. Materials and methods
2.1. Drosophila strains
Drosophila stocks and crosses were maintained on standardcornmeal–yeast–agar medium. The experiments were performedat 25 �C. The RNAi stocks silencing the FaRPs and receptor geneswere obtained from the VDRC Stock Center (Vienna, http://stock-center.vdrc.at) and the NIGFly collection (Kyoto, http://www.shi-gen.nig.ac.jp), their identification numbers are given in Table 1.The other mutant and balancer stocks were received from theBloomington Drosophila Stock Center (http://flystocks.bio.indi-ana.edu). The Fmrf-Gal4 driver strain was a kind gift from P.H. Tag-hert. All of the Drosophila stocks used were on the homozygousw1118 background.
2.2. RNAi silencing of FaRPs and their receptors
We have built a RNAi-based genetic system in which the FaRPgenes and their specific receptors can be silenced in pairwise com-binations, to achieve a maximal loss-of-function effect. In thesestocks the RNAi transgene is driven by the Gal4-inducible UAS pro-moter. The test combinations contained an UAS-Dicer-2 construct(to facilitate cleaving the double-stranded RNA and the formationof the RNA-induced silencing complex RISC) on the X chromosome,as well as one or two UAS-RNAi constructs on the 2nd and/or 3rdchromosome, respectively, silencing different genes (a neuropep-tide and/or its specific receptor or two related receptors). Theelav-Gal4 and Act5-Gal4 drivers are on the 2nd and 3rd chromo-somes, respectively. The test combinations were produced by stan-dard crosses, and carried the RNAi transgenes and the Gal4 driversin heterozygous condition. Genotypes of the test combinationswere: Dcr-2/+; UAS-RNAi/+; Act5C-Gal4/+ or Dcr-2/+; UAS-RNAi/+;Act5C-Gal4/UAS-RNAi as well as Dcr-2/+; elav-Gal4/UAS-RNAi orDcr-2/+; elav-Gal4/UAS-RNAi; UAS-RNAi/+. We tested the efficiencyof the RNAi gene silencing in the test combinations by measuringthe specific mRNA levels through quantitative RT-PCR (for the pri-mer sequences see Table S11). The amount of mRNA was signifi-cantly decreased in all cases (Tables S8 and S9).
2.3. Construction of new Fmrf-Gal4 drivers
Short sequences RS 8, RS 11 and RS 17 in the regulatory region ofFmrf gene were PCR-amplified from genomic DNA of Oregon R flies

Table 1Phenotypic effects of ubiquitous RNAi silencing of FMRFa-related peptide andreceptor genesa.
Silenced gene Genetic combinations:UAS-RNAi stock ID numberb
F1 phenotype(25 �C)c
Fmrf (CG2346) KK 103981 (2nd) LFR (CG2114) NIG 2114R-1 (3rd) VDms (CG6440) KK 108760 (2nd) LDmsR-1 (CG8985) KK101845 (2nd) Ld
GD 9370 (3rd) VDmsR-2 (CG43745) KK 109513 (2nd) L
GD 49953 (3rd) VDsk (CG18090) KK 106592 (2nd) VCCKLR-17D1 (CG42301) KK 100760 (2nd) L
GD 7231 (3rd) VCCKLR-17D3 (CG32540) KK 102039 (2nd) V
GD 9154 (3rd) VFmrf; FR KK103981; NIG 2114R-1 LDms; DmsR-2 KK108760; GD49953 LDmsR-1; DmsR-2 KK101845; GD49953 LDmsR-2; DmsR-1 KK109513; GD9370 LDsk; CCKLR-17D1 KK106592; GD7231 VCCKLR-17D3; CCKLR-17D1 KK102039; GD7231 VCCKLR-17D1; CCKLR-17D3 KK100760; GD9154 L
KK and GD denote RNAi transgenic stocks from the VDRC stock center (http://stockcenter.vdrc.at); NIG denotes NIGFLY stock (http://www.shigen.nig.ac.jp/fly/nigfly/indec.jsp). RNAi transgene-bearing chromosomes are given in parentheses.
a Abbreviations: L = lethal; V = viable; Genetic symbols (Flybase, http://fly-base.org): Dms = Dromyosuppressin; DmsR-1, -R2 = Dms-Receptor-1, -2; Dsk = Dro-sulfakinin; CCKLR-17D1 = Cholecystokinin-like Receptor at 17D1; CCKLR-17D3 = Cholecystokinin-like Receptor at 17D3; Fmrf = Fmrf-amide; FR = Fmrfa-Recep-tor; CG gene annotation numbers are in parentheses.
b Genotypic combinations: Dcr-2/+;UAS-RNAi/+; Act5C-Gal4/+ or Dcr-2/+;UAS-RNAi/+; Act5C-Gal4/ UAS-RNAi.
c Crosses made at 18 �C or 29 �C (not shown) gave results similar to those at25 �C.
d Few weak escapers at 18 �C with distorted wings and disorganized movement.
B. Kiss et al. / General and Comparative Endocrinology 191 (2013) 137–145 139
with the help of Phusion DNA Polymerase (Finzymes) and primerscarrying extensions of BamHI and NotI recognition sites. For the ex-act chromosomal locations of the regulatory regions amplified, thesequences of the PCR primers and the PCR programs see Tables S1–S3, respectively. PCR products were purified from agarose gels withQIAquick Gel Extraction Kit (QIAGEN) according to the manufac-turer’s instructions, digested with BamHI and NotI restriction endo-nucleases (Fermentas) and ligated (with T4 DNA Ligase,Fermentas) into pENTR1A plasmid, digested by the same enzymes.After transformation of the ligated DNA into competent bacterialcells, positive clones were selected and subjected to plasmid puri-fication (High-Speed Plasmid Mini Kit, Qiagene). PCR-amplified re-gions of the purified plasmids were sequenced with an ABI Prism3100 sequence analyzer and the resulted data were compared withthat of deposited in FlyBase (http://flybase.org). Clones containingno PCR-induced errors were selected and in a Gateway™ (GIB-COBRL) exchange reaction the regulatory regions were introducedinto the pBPGUw plasmid using LR Clonase™ Mix (Invitrogene) en-zyme according to the manufacturer’s instructions. After transfor-mation, plasmid DNA purified from the resulted clones were testedwith restriction endonuclease digestion to confirm proper locationand orientation of the regulatory regions. Plasmid DNA of positiveclones was used for injections. All DNA manipulations were doneaccording to standard protocols (Sambrook and Russell, 2001).Transgenic strains were produced by injecting the vector DNA intonos-int; attP/attP embryos (Ni et al., 2008). The nos-int integrasemediates the insertion of the vector into the attP ‘‘landing plat-form’’ on the 3rd chromosome assuring strong and uniformexpression.
According to Benveniste and Taghert (1999), these regulatorysequences direct the gene expression in specific subsets of FMRF-positive neurons. In our constructs, the orientation of the cloned
sequences relative to Gal4 was the same as in the genomic Fmrf50 region. We used the new Gal4 drivers to target the expressionof an UAS-rpr transgene to these specific neural cells which wereconsequently killed by apoptosis.
2.4. Immuno-staining and confocal microscopy of GFP and cleavedcaspase 3 expression
Brains with the thoraco-abdominal ganglion of late 3rd instarlarvae were dissected and fixed in 4% paraformaldehyde for20 min at room temperature. They were washed three times inPBS + 0.1% Triton-X-100 (PBST) followed by blocking for 1 h at25 �C in PBST + 0.2% BSA (PBS-BT). Then the samples were incu-bated overnight at 4 �C with the primary anti-GFP or anti-caspase3 antibody (AB) (Invitrogene A6455, 1000� diluted) in PBS-BT. Thebrains were then washed three times in PBST for 20 min each, andincubated for 2 h at room temperature with the secondary anti-body, washed three times in PBST for 20 min each and finally inPBS for 10 min. The samples were kept in glycerol:PBS (1:1) for5 min, and finally covered in glycerol:PBS (4:1). The preparationswere examined under an Olympus FluoView FV1000 confocal laserscanning microscope (Olympus Life Science Europa GMBH, Ham-burg, Germany). The following antibodies were used: GFP-specificGoat IgG by R&D System, Cat. No. AF4240, 40� diluted; anti-cas-pase 3 (cleaved) antibody from rabbit, Cell Signaling, Cat. No.9661S, 250� diluted in PBST. For the secondary stainings: anti-goatantibody from donkey (Molecular Probes A11055, Alexa Fluor 488)diluted 600�; anti-rabbit IgG from donkey (Invitrogen, Cat. No.A31573) diluted 600�. Double stainings were made with mixturesof the appropriate primary and secondary antibodies.
2.5. Locomotion test
To examine the locomotor activity of adult flies, we used theRepetitive Startle-induced Hyperactivity (ReSH) test as describedby Lebestky et al. (2009), with slight modifications. Briefly, theexperimental device consisted of eight horizontally arrangedtransparent glass tubes (160 mm long with 14 mm internal diam-eter) each connected to a pressurized air container (five bars) andfixed onto a plastic board, which is placed under a Logitech Web-Cam Pro 9000 video camera. Flies were selected under CO2 at least1 day prior to the experiments. For the test, ten 4–5 day old flies,males or females separately, were transferred into each tube. Fliesof the same genotype were placed into two parallel tubes, and al-lowed to calm down for 10 min Then two air puffs were deliveredin a 1 s interval which pushed the flies to the rear end of the tubesclosed with a fine mesh. The video-recording began 1 min beforethe first air puff and lasted for 4 min afterwards. After finishingthe first cycle, the flies were maintained in the tubes for 10 min,after which the experiment was repeated. To avoid the differencesgenerated by the circadian rhythm, the tests were made in latemorning or early afternoon. Three cycles were made with eachgroup of flies. Four experiments (each repeated in three cycles)were performed with different flies of each genotype, i.e. 80 maleand female flies for every genotype were tested altogether. Basedon the video records, the average velocity (mm/s) of the flies wascalculated by the Drosana custom computer program (developedby Stefan Kakaš) in 0.1–0.2 s intervals, and used to construct curvesrepresenting the changing mean velocity of movement (MVM) intime.
2.6. Negative geotaxis test
With no previous anesthetization, ten 4–5 day old flies, males orfemales separately, were transferred into glass tubes (160 mm longwith 14 mm internal diameter), and were allowed to calm down

140 B. Kiss et al. / General and Comparative Endocrinology 191 (2013) 137–145
for 10 min. The tubes were placed into a rack in vertical position,and the flies were shaken off by knocking the rack twice to the ta-ble manually. Leaving the rack in vertical position, photographswere taken at the 10th, 20th and 30th seconds, and the flies werecounted according to their distribution by height along the tube.
3. Results and discussion
3.1. Construction of FMRF expression pattern-specific Gal4 drivers
Benveniste and Taghert (1999) described 50 upstream DNA se-quences regulating the spatial expression in the CNS of the Fmrfgene. When they cloned these DNA fragments into a vector up-stream of the b-galactosidase gene, they found the staining patternreproducing specific parts of the FMRF spatial expression. Based onthis information, we amplified three such DNA sequences by PCR,cloned them into the pBPGUw vector upstream to the Gal4 codingsequence, and made transgenic Drosophila strains (RS8-Gal4, RS11-Gal4, RS17-Gal4) carrying these constructs on the 3rd chromosomein the attP ‘‘landing platform’’. By making use of the UAS-GFP repor-ter gene, we tested the Gal4 expression in these stocks. As it wasshown by the GFP fluorescence and GFP-specific immuno-staining,the expression pattern in the larval CNS of RS8-Gal4, RS11-Gal4 andRS17-Gal4 reproduced the patterns described by Benveniste andTaghert (1999) for the same regulatory sequences (Fig. 1). How-ever, additional GFP-positive neurons were also detected in allthree cases, especially in the case of RS8-Gal4. This might be ex-plained by the enhanced visibility of the GFP expression providedby the immuno-staining with GFP-specific antibody. Another dif-ference between our constructs and those of Benveniste and Tag-hert (1999) was in the relative orientation of the clonedsequences. In all three constructs of ours the orientation of the reg-ulatory DNA segment relative to the Gal4 coding part was the sameas in the genomic sequence. In the case of Benveniste and Taghert(1999), however, the regulatory sequences corresponding to RS8and RS11 were cloned in the reverse orientation. Benveniste andTaghert (1999) attributed the observed expression patterns tothe effect of enhancers, the activity of which is not influenced bytheir relative position and polarity. However, the possible effectof other ‘‘non-enhancer’’ sequences in the cloned DNA segmentsmight be sensitive to their polarity relative to the coding region,and this could also be responsible for the ectopic expression.Unraveling this problem needs further studies.
3.2. Ubiquitous RNAi silencing of FaRP peptides and their receptors
Silencing the gene expression by double-stranded RNA seldomgenerates a null-activity phenotype, some residual gene activityremains in most cases resulting in a ‘‘hypomorhic’’ phenotype.However, the effect becomes stronger by increasing the RNAitransgene copy number and/or the temperature up to 29 �C. Atthe same time, the silencing effect is largely independent of thedosage (copy number) of the silenced gene, and it results in a dom-inant loss-of-function phenotype.
In the first experiment, the stocks carrying RNAi constructs to si-lence the FaRP neuropeptide and receptor genes were crossed to theAct5-Gal4 driver on the 2nd chromosome which induces a ubiqui-tous and continuous expression of the double-stranded inhibitoryRNA. As Table 1 shows, the ubiquitous silencing of the Fmrf, Dms,DmsR-1, DmsR-2, CCKLR-17D1 and CCKLR-17D3 genes resulted incomplete lethality, most of the animals died as pharate adults (datanot shown). The FR-RNAi and Dsk-RNAi flies remained viable andfertile. The lethal effect was observed with the KK series of theVDRC mutants where the silencing construct is inserted into a stan-dard ‘‘landing site’’ (attP2, Ni et al., 2008) on the 2nd chromosome
ensuring a uniform strong silencing effect. The same constructs, in-serted on the 3rd chromosome at random sites (GD series), did notshow lethality even at 29 �C, probably due to the lower level of theRNAi effect. It should be noted that, as the VDRC collection has noFR-RNAi transgene in the strongly expressing KK series, we silencedthe FR receptor by a randomly inserted RNAi construct (NIG 2114R-1) from the NIG-Fly collection, with a possibly lower expressionrate. It remains to be seen whether a stronger silencing of the FRgene would also result in lethality.
While the main source of the neuropeptides are the neurons inthe CNS, some of them were found in other organs as well: FMRFpeptides are also expressed in the gut (Nichols et al., 1999). DMScould be detected by immuno-staining in the crop and gut(‘‘brain-gut peptide’’, McCormick and Nichols, 1993; Merte andNichols, 2002) as well as the heart and the male accessory gland(FlyATLAS, the Drosophila gene expression atlas by Chintapalli,Wang and Dow, http://flyatlas.org). NPF and sNPF are expressedboth in the CNS and the midgut (Veenstra, 2009). Their receptorsare also expressed in other tissues/organs: according to the FlyAT-LAS database, the sNPF-R and CCKLR-17D3 genes show a low butbroad-scale expression in the larval and adult internal organs,while DmsR-1 and NPFR1 have a higher expression in the cropand Malpighian tubules, respectively. The generalized silencing ofthe receptors could be responsible for the lethal phenotype. Thisseems to be supported by the observation that when the samestrains were crossed to the elav-Gal4 pan-neuronal driver, andtherefore the silencing effect was restricted to the CNS, none ofthe above combinations were lethal (data not shown). This sug-gests that the cause of the lethality is outside of the CNS.
By P transposon remobilization, we isolated intragenic dele-tions in the DmsR-1 and Fmrf genes (unpublished; data not shown).Although these deficiencies apparently eliminated the gene func-tion, the homozygous mutant flies proved to be fully viable andfertile (unpublished observation). Liu (2008) also reported that anull-activity Fmrf gene mutant isolated by him proved to be fullyviable and fertile. There is an obvious contradiction between theviability of the DmsR-1 and Fmrf deletion mutants and the lethalityof the DmsR-1-RNAi; Act5-Gal4 and Fmrf-RNAi; Act5-Gal4 combina-tions. This suggests the possibility of an ‘‘off-target’’ effect, a cross-reaction of the RNAi machinery which can also destroy unspecificmRNAs if those have a sequence motif identical with or closely re-lated to the target sequence (Kulkarni et al., 2006; Echeverri et al.,2006). Clarification of this problem needs further studies.
3.3. RNAi silencing of FaRPs and FaRP receptors in the CNS candecrease locomotor activity
Flies having the RNAi constructs for a FaRP and its specificreceptor were crossed to the elav-Gal4 driver strain. The offspringcarrying the RNAi transgenes and the elav driver showed normalviability and fertility, and was subjected to the ReSH test as de-scribed above. The results are shown in Fig. 2 for the female flies.(Similar effects were observed with the males, see Fig. S1.) In gen-eral, suppressing the FARPs and/or their receptors significantly(p < 0.01, Student t-test) decreased the MVM in most of the geneticcombinations tested (see Tables S4 and S6). The MVM decreasewas observed at both the rest activity before the air puffs andthe puff-induced activity. The largest difference between theRNAi-reduced MVM and the control values was found when bothDMS-receptor genes (DmsR-1 and DmsR-2) were silenced in paral-lel. As for the absolute values, three of the tested combinations(silencing transgenes on the 2nd; 3rd chromosomes: Dms-RNAi/elav-Gal4; DmsR-2-RNAi /+, DmsR-1-RNAi /elav-Gal4; DmsR-2-RNAi/+, Dsk-RNAi /elav-Gal4; CCKLR-17D1-RNAi /+) showed the largestMVM decrease. In the two other cases (Fmrf-RNAi/elav-Gal4;FR-RNAi /+ and DmsR-2-RNAi/elav-Gal4; DmsR-1-RNAi/+) the MVM

Fig. 1. Transcription activating patterns of different regulatory sequences (A–C: RS11, D–F: RS8 and G–I: RS17) of the Fnrf gene in the larval CNS. Confocal micrographs (A, Dand G) and their schematic representations (B, E and H) of immuno-stained UAS-GFP expressing cells in the brain and the ventral ganglion of third instar larvae. Schematicrepresentations of beta-galactosidase expressions (C, F and I) in larval CNS, controlled by RS11, RS8 and RS17 were drawn after Benveniste and Taghert, 1999. Positions of theregulatory sequences within the Fmrf gene 50 region and their orientations relative to the reporter genes are shown above each schematic panel. In the schemes, red and greenpoints represent FMRF-positive neurons described by Benveniste and Taghert (1999) and/or observed in this study, respectively.
B. Kiss et al. / General and Comparative Endocrinology 191 (2013) 137–145 141
decrease was less. For the CCKLR-17D1-RNAi/elav-Gal4;CCKLR-17D3-RNAi/+ silencing combination, the difference from the con-trols was not so remarkable.
As mentioned above (see Section 2.2), the transgenes on the 2ndchromosome are inserted in the attP2 platform and have anuniformly strong expression, while the transgenes on the 3rd
chromosome have a varying, probably weaker expression due tothe different position effects at the different chromosomal inser-tion sites. Although both the DmsR-1-RNAi /elav-Gal4; DmsR-2-RNAi/+ and DmsR-2-RNAi /elav-Gal4;DmsR-1-RNAi/+ combinations si-lenced the DmsR-1 and DmsR-2 genes together (Fig. 2C and D),the MVM effect was stronger when the DMSR-1-RNAi transgene

Fig. 2. RNAi knock-down of FaRP and FaRP receptor genes effectively changes the startle (air puff)-induced locomotor activity of Drosophila adult females. The curvesrepresent the changes in time of the mean velocity of movement (MVM) before and after the air-puffs. Blue and green: control curves; red: RNAi knock-down curve. Flygenotypes are indicated in the upper right corner of each panel. Abbreviations: Dcr = Dicer; DskR-1 = CCKLR-17D3, DskR-2 = CCKLR-17D1, RNAi = UAS-RNAi, Gal4-inducibletransgene producing double-stranded RNA; elav = Gal4 driver.
142 B. Kiss et al. / General and Comparative Endocrinology 191 (2013) 137–145
was inserted in the attP2 platform on the 2nd chromosome and theDmsR-2-RNAi was on the 3rd (Fig. 2C and D). We think that the dif-ferences in the chromosomal position effects may be responsiblefor the variation between the MVM curves. These results showthat, at least in the combinations tested, the FaRP neuropeptidesand their receptors can modify the locomotion activity and stressresponse of the fruitfly adults.
Previous studies have already suggested that most FaRP pep-tides have myotropic effects (Nichols et al., 2002; Kahsai et al.,2010). They can influence e.g. the gut and crop contraction (Dutt-linger et al., 2002), the heart rate (Nichols, 2006), and the locomo-tion activity and stress response (Nässel and Wegener, 2011; Chenet al., 2012). Clark and his co-workers (2008) showed that, at leastin the case of DPKQDFMRFmide, the most abundant peptide coded
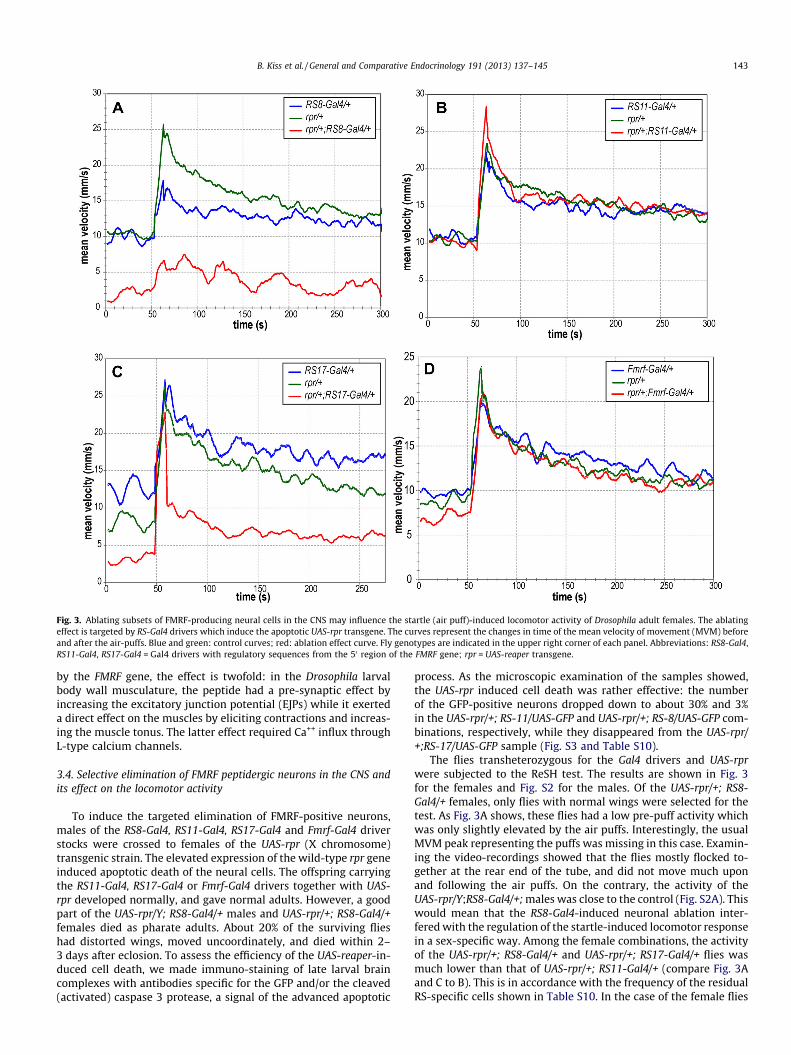
Fig. 3. Ablating subsets of FMRF-producing neural cells in the CNS may influence the startle (air puff)-induced locomotor activity of Drosophila adult females. The ablatingeffect is targeted by RS-Gal4 drivers which induce the apoptotic UAS-rpr transgene. The curves represent the changes in time of the mean velocity of movement (MVM) beforeand after the air-puffs. Blue and green: control curves; red: ablation effect curve. Fly genotypes are indicated in the upper right corner of each panel. Abbreviations: RS8-Gal4,RS11-Gal4, RS17-Gal4 = Gal4 drivers with regulatory sequences from the 50 region of the FMRF gene; rpr = UAS-reaper transgene.
B. Kiss et al. / General and Comparative Endocrinology 191 (2013) 137–145 143
by the FMRF gene, the effect is twofold: in the Drosophila larvalbody wall musculature, the peptide had a pre-synaptic effect byincreasing the excitatory junction potential (EJPs) while it exerteda direct effect on the muscles by eliciting contractions and increas-ing the muscle tonus. The latter effect required Ca++ influx throughL-type calcium channels.
3.4. Selective elimination of FMRF peptidergic neurons in the CNS andits effect on the locomotor activity
To induce the targeted elimination of FMRF-positive neurons,males of the RS8-Gal4, RS11-Gal4, RS17-Gal4 and Fmrf-Gal4 driverstocks were crossed to females of the UAS-rpr (X chromosome)transgenic strain. The elevated expression of the wild-type rpr geneinduced apoptotic death of the neural cells. The offspring carryingthe RS11-Gal4, RS17-Gal4 or Fmrf-Gal4 drivers together with UAS-rpr developed normally, and gave normal adults. However, a goodpart of the UAS-rpr/Y; RS8-Gal4/+ males and UAS-rpr/+; RS8-Gal4/+females died as pharate adults. About 20% of the surviving flieshad distorted wings, moved uncoordinately, and died within 2–3 days after eclosion. To assess the efficiency of the UAS-reaper-in-duced cell death, we made immuno-staining of late larval braincomplexes with antibodies specific for the GFP and/or the cleaved(activated) caspase 3 protease, a signal of the advanced apoptotic
process. As the microscopic examination of the samples showed,the UAS-rpr induced cell death was rather effective: the numberof the GFP-positive neurons dropped down to about 30% and 3%in the UAS-rpr/+; RS-11/UAS-GFP and UAS-rpr/+; RS-8/UAS-GFP com-binations, respectively, while they disappeared from the UAS-rpr/+;RS-17/UAS-GFP sample (Fig. S3 and Table S10).
The flies transheterozygous for the Gal4 drivers and UAS-rprwere subjected to the ReSH test. The results are shown in Fig. 3for the females and Fig. S2 for the males. Of the UAS-rpr/+; RS8-Gal4/+ females, only flies with normal wings were selected for thetest. As Fig. 3A shows, these flies had a low pre-puff activity whichwas only slightly elevated by the air puffs. Interestingly, the usualMVM peak representing the puffs was missing in this case. Examin-ing the video-recordings showed that the flies mostly flocked to-gether at the rear end of the tube, and did not move much uponand following the air puffs. On the contrary, the activity of theUAS-rpr/Y;RS8-Gal4/+; males was close to the control (Fig. S2A). Thiswould mean that the RS8-Gal4-induced neuronal ablation inter-fered with the regulation of the startle-induced locomotor responsein a sex-specific way. Among the female combinations, the activityof the UAS-rpr/+; RS8-Gal4/+ and UAS-rpr/+; RS17-Gal4/+ flies wasmuch lower than that of UAS-rpr/+; RS11-Gal4/+ (compare Fig. 3Aand C to B). This is in accordance with the frequency of the residualRS-specific cells shown in Table S10. In the case of the female flies

Fig. 4. RS8-Gal4-induced ablation of specific FMRF-producing neurons does notaffect the negative geotaxis of female flies. The bars represent the average numberof flies climbed up above the 95 mm threshold level. Blue and green: controls; red:ablation effect. Fly genotypes are indicated in the upper right corner. Abbreviations:RS8-Gal4 = Gal4 driven by a regulatory sequence from the 50 region of the Fmrf gene;rpr = UAS-reaper transgene.
144 B. Kiss et al. / General and Comparative Endocrinology 191 (2013) 137–145
(Fig. 3), the MVM differences were significant at the p < 0.01 level(Student t-test, Tables S5, S7).
We further examined the locomotor activity of the ‘‘sluggish’’UAS-rpr/+; RS8-Gal4/+ females in the negative geotaxis test (Sec-tion 2.6). Interestingly, the flies were just as active as the controlsin the geotaxis test (Fig. 4). This would mean that the startle-in-duced locomotor activity and the negative geotaxis are differentlyregulated in the UAS-rpr/+; RS8-Gal4/+ female flies. As for theremaining two combinations, the MVM curves of UAS-rpr/+;RS11-Gal4/+ and UAS-rpr/+; Fmrf-Gal4/+ practically coincided withthe control activities (Fig. 3B and D, respectively).
In the male combinations, the activity of the UAS-rpr/Y; RS8-Gal4/+ and UAS-rpr/Y; RS11-Gal4/+ flies was not significantly differ-ent from the controls (Fig. S2A and B). The UAS-rpr/Y;RS17-Gal4/+combination only moderately decreased the MVM (Fig. S2C) whilethe behavior of UAS-rpr/Y; Fmrf-Gal4/+ flies was indistinguishablefrom the controls (Fig. S2D).
The above experiments resulted in several unexpected observa-tions. First of all, the completely normal activity and behavior ofthe flies carrying the UAS-rpr and Fmrf-Gal4 transgenes was a sur-prise. In these flies most of the FMRF-producing neurons are likely
missing, consequently the level of the FMRF peptides should bevery low. (It has to be noted, however, that in this case the activityof the FR receptor is still normal.) On the other hand, when the RS8,RS11 and RS17 drivers were used to drive the expression of UAS-rpr,only specific parts of the FMRF-expressing neuronal pattern wereablated, and yet it influenced the flies’ locomotor activity (see Sec-tion 3.4. above as well as Figs. 3 and S2). Also, when the genes cod-ing for the FMRF peptides and the specific FR receptor weresilenced together, the MVM of the flies became much lower thanthat of the control (see Section 3.3. and Fig. 2). In the light of thesefindings, the ineffectiveness of the UAS-rpr/+; Fmrf-Gal4/+ combina-tion seems to be enigmatic. However, we recently found that theoriginal Fmrf-Gal4 stock we used, contained Fmrf-Gal4 transgenesinserted in the 2nd and 3rd, and possibly in the X chromosomes,and the individual flies carried accidental combinations of them.As we believed that the Fmrf-Gal4 was on the 2nd chromosome,the X and 3rd could be exchanged for ‘‘wild’’ chromosomes duringthe crosses. This means that the number of the Fmrf-Gal4 transgeneand its activity could unexpectedly be low in the flies, resulting in aweak effect. Otherwise one can speculate that the neurons acti-vated in the Fmrf-Gal4 expression pattern are in some kind of func-tional balance, so when the whole pattern is missing, the lackingparts could neutralize each other’s effect. This notion may be sup-ported by the observation that in the RS8, RS11 and RS17 driversthe Gal4 is expressed in extra neurons outside of the FMRF pattern,and might interfere with this hypothetical balance leading to thelow MVM phenotype. In this respect, the RS8 driver activates mostof the extra neurons (Fig. 1D). Furthermore, in the UAS-rpr; RS8-Gal4 combination the high incidence of pharate adult lethalityand the distorted wing phenotype suggests that the RS8 drivercan express the Gal4 in other tissues as well outside of the CNS,e.g. in the wing imaginal disc epithelium. All these may mean thatif a partial sequence is taken out of the original context of a regu-latory region, ectopic new activities may arise.
Acknowledgments
This work was supported by the Hungarian grants OTKAT75774 and partially by OTKA K104011 as well as by TAMOP-4.2.2/B-10/1-2010-0012. The support for travel and subsidy ex-penses (2010–2012) by the Bilateral Research Agreement betweenthe Hungarian and Czech Academy of Sciences is gratefullyacknowledged. Special thanks are due to Drs. Frantisek Sehnaland Dalibor Kodrik (Biological Centre, CAS, Ceské Budejovice) forscientific and mentoring support, Dr. Ferhan Ayaydin for his helpin confocal microscopy as well as Dr. Paul H. Taghert for providingthe Fmrf-Gal4 stock.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.ygcen.2013.05.023.
References
Adams, M.D. et al., 2000. The genome sequence of Drosophila melanogaster. Science287, 2185–2195.
Baumgardt, M., Miguel-Aliaga, I., Karlsson, D., Ekman, H., Thor, S., 2007.Specification of neuronal identities by feedforward combinatorial coding.PLoS Biol. 5, e37.
Bendena, W.G., Campbell, J., Zara, L., Tobe, S.S., Chin-Sang, I.D., 2012. Selectneuropeptides and their G-protein coupled receptors in Caenorhabditis elegansand Drosophila melanogaster. Front. Endocrinol. 3, 1–12.
Benveniste, R.J., Taghert, P.H., 1999. Cell type-specific regulatory sequences controlexpression of the Drosophila FMRF-NH2 neuropeptide gene. J. Neurobiol. 38,507–520.

B. Kiss et al. / General and Comparative Endocrinology 191 (2013) 137–145 145
Brody, T., Cravchik, A., 2000. Drosophila melanogaster G protein-coupled receptors. J.Cell Biol. 150, 83–88.
Cazzamali, G., Grimmelikhuijzen, C.J.P., 2002. Molecular cloning and functionalexpression of the first insect FMRFamide receptor. Proc. Natl. Acad. Sci. USA 99,12073–12078.
Chen, X., Ganetzky, B., 2012. A neuropeptide signaling pathway regulates synapticgrowth in Drosophila. J. Cell Biol. 196, 529–543.
Chen, X., Peterson, J., Nachman, R.J., Ganetzky, B., 2012. Drosulfakinin activatesCCKLR-17D1 and promotes larval locomotion and escape response inDrosophila. Fly (Austin) 6, 290–297.
Clark, J., Milakovic, M., Cull, A., Klose, M.K., Mercier, A.J., 2008. Evidence forpostsynaptic modulation of muscle contraction by a Drosophila neuropeptide.Peptides 29, 1140–1149.
Clynen, E., Reumer, A., Baggerman, G., Mertens, I., Schoofs, L., 2010. Neuropeptidebiology in Drosophila. Adv. Exp. Med. Biol. 692, 192–210.
Duttlinger, A., Berry, K., Nichols, R., 2002. The different effects of three Drosophilamelanogaster dFMRFamide-containing peptides on crop contractions suggestthese structurally related peptides do not play redundant functions in gut.Peptides 23, 1953–1957.
Echeverri, C.J., Beachy, P.A., Baum, B., Boutros, M., Buchholz, F., Chanda, S.K.,Downward, J., Ellenberg, J., Fraser, A.G., Hacohen, N., Hahn, W.C., Jackson, A.L.,Kiger, A., Linsley, P.S., Lum, L., Ma, Y., Mathey-Prévôt, B., Root, D.E., Sabatini,D.M., Taipale, J., Perrimon, N., Bernards, R., 2006. Minimizing the risk ofreporting false positives in large-scale RNAi screens. Nat. Methods 3, 777–779.
Hauser, F., Williamson, M., Cazzamali, G., Grimmelikhuijzen, C.J.P., 2006a.Identifying neuropeptide and protein hormone receptors in Drosophilamelanogaster by exploiting genomic data. Brief. Funct. Genomic Proteomic 4,321–330.
Hauser, F., Cazzamali, G., Williamson, M., Blenau, W., Grimmelikhuijzen, C.J.P.,2006b. A review of neurohormone GPCRs present in the fruitfly Drosophilamelanogaster and the honey bee Apis mellifera. Prog. Neurobiol. 80, 1–19.
Hewes, R.S., Taghert, P.H., 2007. Neuropeptides and neuropeptide receptors in theDrosophila melanogaster genome. Genome Res. 11, 1126–1142.
Kahsai, L., Martin, J.-R., Winther, Å.M.E., 2010. Neuropeptides in the Drosophilacentral complex in modulation of locomotor behavior. J. Exp. Biol. 213, 2256–2265.
Kaminski, S., Orlowski, E., Berry, K., Nichols, R., 2002. The effects of three Drosophilamelanogaster myotropins on the frequency of foregut contractions differ. JNeurogenet. 16, 125–134.
Kulkarni, M.M., Booker, M., Silver, S.J., Friedman, A., Hong, P., Perrimon, N., Mathey-Prevot, B., 2006. Evidence of off-target effects associated with long dsRNAs inDrosophila melanogaster cell-based assays. Nat. Methods 3, 833–838.
Lebestky, T., Chang, J.S., Dankert, H., Zelnik, L., Kim, Y.C., Han, K.A., Wolf, F.W.,Perona, P., Anderson, D.J., 2009. Two different forms of arousal in Drosophila areoppositely regulated by the dopamine D1 receptor ortholog DopR via distinctneural circuits. Neuron 64, 522–536.
Liu, Y., 2008. Genetic Dissection of the Drosophila Fmrf gene. (Ph.D. dissertation),The University of Wisconsin-Milwaukee.
McCormick, J., Nichols, R., 1993. Spatial and temporal expression identifydromyosuppressin as a brain-gut peptide in Drosophila melanogaster. J. Comp.Neurol. 338, 279–288.
Merte, J., Nichols, R., 2002. Drosophila melanogaster myotropins have uniquefunctions and signaling pathways. Peptides 23, 757–763.
Nässel, D.R., Wegener, C., 2011. A comparative review of short and longneuropeptide F signaling in invertebrates: any similarities to vertebrateneuropeptide Y signaling? Peptides 32, 1335–1355.
Nässel, D.R., Winther, A.M., 2010. Drosophila neuropeptides in regulation ofphysiology and behavior. Prog. Neurobiol. 92, 42–104.
Nässel, D.R., Enell, L.E., Santos, J.G., Wegener, C., Johard, H.A., 2008. A largepopulation of diverse neurons in the Drosophila central nervous systemexpresses short neuropeptide F suggesting multiple distributed peptidefunctions. BMC Neurosci. 9, 90.
Ni, J.-Q., Markstein, M., Binari, R., Pfeiffer, B., Liu, L.-P., Villalta, C., Booker, M.,Perkins, L., Perrimon, N., 2008. Vector and parameters for targeted transgenicRNA interference in Drosophila melanogaster. Nat. Methods 5, 49–51.
Nichols, R., 2003. Signaling pathways and physiological functions of Drosophilamelanogaster FMRFamide-related peptides. Annu. Rev. Entomol. 48, 485–503.
Nichols, R., 2006. FMRFamide-related peptides and serotonin regulate Drosophilamelanogaster heart rate: mechanisms and structure requirements. Peptides 27,1130–1137.
Nichols, R., Lim, I.A., 1996. Spatial and temporal immunocytochemical analysis ofdrosulfakinin (Dsk) gene products in the Drosophila melanogaster centralnervous system. Cell Tissue Res. 283, 107–116.
Nichols, R., McCormick, J., Lim, I., 1999. Regulation of Drosophila FMRFamideneuropeptide gene expression. J. Neurobiol. 39, 347–358.
Nichols, R., Bendena, W.G., Tobe, S.S., 2002. Myotropic peptides in Drosophilamelanogaster and the genes that encode them. J. Neurogenet. 16, 1–28.
Nichols, R., Manoogian, B., Walling, E., Mispelon, M., 2009. Plasticity in the effects ofsulfated and nonsulfated sulfakinin on heart contractions. Front. Biosci. 1,4035–4043.
Palmer, G.C., Tran, T., Duttlinger, A., Nichols, R., 2007. The drosulfakinin 0 (DSK 0)peptide encoded in the conserved Dsk gene affects adult Drosophilamelanogaster crop contractions. J. Insect Physiol. 53, 1125–1133.
Park, D., Veenstra, J.A., Park, J.H., Taghert, P.H., 2008. Mapping peptidergic cells inDrosophila: where DIMM fits in. PLoS ONE 3, e1896.
Sambrook, J., Russell, D.W., 2001. Molecular Cloning: A Laboratory Manual. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY.
Santos, J.G., Vömel, M., Struck, R., Homberg, U., Nässel, D.R., Wegener, C., 2007.Neuroarchitecture of peptidergic systems in the larval ventral ganglion ofDrosophila melanogaster. PLoS ONE 2, e695.
Schneider, L.E., O’Brien, M.A., Taghert, P.H., 1991. In situ hybridization analysis ofthe FMRFamide neuropeptide gene in Drosophila. I. Restricted expression inembryonic and larval stages. J. Comp. Neurol. 304, 608–622.
Schneider, L.E., Roberts, M.S., Taghert, P.H., 1993. Cell type-specific transcriptionalregulation of the Drosophila FMRFamide neuropeptide gene. Neuron 10, 279–291.
Söderberg, J.A., Carlsson, M.A., Nässel, D.R., 2012. Insulin-producing cells in thedrosophila brain also express satiety-inducing cholecystokinin-like peptide,Drosulfakinin. Front. Endocrinol. 3, 109.
Vanden Broeck, J., 2001. Neuropeptides and their precursors in the fruit fly,Drosophila melanogaster. Peptides 22, 241–254.
Veenstra, J.A., 2009. Peptidergic paracrine and endocrine cells in the midgut of thefruit fly maggot. Cell Tissue Res. 336, 309–323.
Wegener, C., Gorbashov, A., 2008. Molecular evolution of neuropeptides in thegenus Drosophila. Genome Biol. 9, R131.
Copyright © 2022 FDOKUMEN




















