
Selection and use of autochthonous multiple strain cultures for the manufacture of high-moisture...
10
Selection and use of autochthonous multiple strain cultures for the manufacture of high-moisture traditional Mozzarella cheese Maria De Angelis a, ⁎, Silvia de Candia a , Maria Piera Calasso a , Michele Faccia b , Timothy P. Guinee c , Maria C. Simonetti d , Marco Gobbetti a a Dipartimento di Protezione delle Piante e Microbiologia Applicata, Università degli Studi di Bari, Bari 70126, Italy b Dipartimento di Progettazione e Gestione dei Sistemi Agro-zootecnici e Forestali, Università degli Studi di Bari, Italy c Dairy Products Research Centre, Teagasc, Moorepark, Fermoy, Co. Cork, Ireland d Labo-Biotech, Faculty of Biotechnology, University of Bari, 70126 Bari, Italy ABSTRACT ARTICLE INFO Article history: Received 15 September 2007 Received in revised form 14 February 2008 Accepted 24 March 2008 Keywords: Starter cultures Italian Mozzarella cheeses Lactobacillus plantarum 18A, Lactobacillus helveticus 2B, Lactobacillus delbrueckii subsp. lactis 20F, Strepto- coccus thermophilus 22C, Enterococcus faecalis 32C and Enterococcus durans 16E were the most acidifying strains within 146 isolates for natural whey starters. The effect of media and temperature on 2 autochthonous multiple strain cultures (AMSI: 18A, 2B, 20F and 22C, 32C and 16E and AMSII: 18A, 2B, 20F and 22C) was studied. Genomic analysis showed a constant cell numbers for AMSII during 16 days of propagation in whey milk. Mozzarella cheese was made by using AMSII, commercial starter (CS) or citric acid (DA). Compared to other cheeses, the DA had a lower level of protein, ash, Ca, free amino acids and a higher level of moisture. Based on confocal laser scanning microscopy analysis, AMSII cheese showed the lowest microstructural variations during the period of storage compared to other cheeses. All the sensory attributes were scored highest for AMSII cheese. ASMII extend the shelf-life to ca. 12−15 days instead of the 5−7 days of traditional high-moisture Mozzarella cheese. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Starters cultures are mainly species of lactic acid bacteria that are deliberately added to milk where their primary role is to start the production of lactic acid for the cheese manufacturing process (Broome, 2007). It is well known that starter cultures fulfil other important functions: inhibition of spoilage and pathogenic micro- organisms, improvement of cheese keeping quality, direct and indirect contributions to flavour and aroma. Starter cultures may be classified into two large groups: traditional (including artisanal, natural starters and multiple strain culture starters) and defined or commercial starters (Limsowtin et al., 1995). A high-moisture cows' Mozzarella cheese (also defined as “Fior di Latte”) is traditionally manufactured in the Southern Italy, mainly in the Apulia region. Recently, its production has been estimated ca. 220,400 tonnes per year (www.databank.it). During the last decade, the increasing consumption of cows' Mozzarella cheese favoured the differentiation of several manufacturing processes, which mainly depended on the size of the cheese factories. Natural whey or milk starter cultures represent one of the technological options and they are daily produced and propagated at the cheese plant. Overall, the composition of natural starter cultures is complex, variable and subjected to unpredictable performance (Coppola et al., 2001; de Candia et al., 2007). In most of the cases, natural starters have been replaced by (i) commercial starters, usually consisting of Streptococcus thermophilus alone or mixtures of S. thermophilus and Lactobacillus helveticus or Lactobacillus delbrueckii subsp. bulgaricus (Coppola et al., 2001; Coppola et al., 2006; de Candia et al., 2007) or by (ii) direct acidification with lactic acid, citric acid or glucono-δ-lactone (Parente and Moschetti, 1997). Overall, the choice between biological and chemical acidification and between natural and commercial starter cultures may have important repercussion on several features of cows' Mozzarella cheese. The effect of defined starter cultures was studied on physical, melt and rheological characteristics (Dave et al., 2003), microstructure (Merrill et al., 1996), and composition, proteolysis and functional properties of Mozzarella cheese (Yun et al., 1995). The thermophilic lactic acid bacteria used to make Mozzarella cheese are particularly important in determining the physical properties (Oberg et al., 1991a,b). Besides, Yun et al. (1995) reported that a balanced rod- to-coccus ratio influenced proteolysis, springiness and viscosity of Mozzarella cheese. Overall, the use of commercially available starter cultures seemed to cause a less characteristic flavour of the cheese (Broome, 2007). Several studies have characterized the microbial biodiversity which occurs in natural whey starter cultures for cows' Mozzarella cheese. Molecular, physiological and technological International Journal of Food Microbiology 125 (2008) 123–132 ⁎ Corresponding author. Dipartimento di Protezione delle Piante e Microbiologia Applicata, Via G. Amendola 165/a, Università degli Studi di Bari, Bari 70126, Italy. Tel.: +39 080 5442949; fax: +39 080 5442911. E-mail address: [email protected] (M. De Angelis). 0168-1605/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.ijfoodmicro.2008.03.043 Contents lists available at ScienceDirect International Journal of Food Microbiology journal homepage: www.elsevier.com/locate/ijfoodmicro
Transcript of Selection and use of autochthonous multiple strain cultures for the manufacture of high-moisture...

International Journal of Food Microbiology 125 (2008) 123–132
Contents lists available at ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r.com/ locate / i j foodmicro
Selection and use of autochthonous multiple strain cultures for the manufacture ofhigh-moisture traditional Mozzarella cheese
Maria De Angelis a,⁎, Silvia de Candia a, Maria Piera Calasso a, Michele Faccia b, Timothy P. Guinee c,Maria C. Simonetti d, Marco Gobbetti a
a Dipartimento di Protezione delle Piante e Microbiologia Applicata, Università degli Studi di Bari, Bari 70126, Italyb Dipartimento di Progettazione e Gestione dei Sistemi Agro-zootecnici e Forestali, Università degli Studi di Bari, Italyc Dairy Products Research Centre, Teagasc, Moorepark, Fermoy, Co. Cork, Irelandd Labo-Biotech, Faculty of Biotechnology, University of Bari, 70126 Bari, Italy
⁎ Corresponding author. Dipartimento di ProtezioneApplicata, Via G. Amendola 165/a, Università degli StTel.: +39 080 5442949; fax: +39 080 5442911.
E-mail address: [email protected] (M. De Ang
0168-1605/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.ijfoodmicro.2008.03.043
A B S T R A C T
A R T I C L E I N F OArticle history:
Lactobacillus plantarum 18A Received 15 September 2007Received in revised form 14 February 2008Accepted 24 March 2008Keywords:Starter culturesItalian Mozzarella cheeses
, Lactobacillus helveticus 2B, Lactobacillus delbrueckii subsp. lactis 20F, Strepto-coccus thermophilus 22C, Enterococcus faecalis 32C and Enterococcus durans 16E were the most acidifyingstrains within 146 isolates for natural whey starters. The effect of media and temperature on 2autochthonous multiple strain cultures (AMSI: 18A, 2B, 20F and 22C, 32C and 16E and AMSII: 18A, 2B, 20Fand 22C) was studied. Genomic analysis showed a constant cell numbers for AMSII during 16 days ofpropagation in whey milk. Mozzarella cheese was made by using AMSII, commercial starter (CS) or citric acid(DA). Compared to other cheeses, the DA had a lower level of protein, ash, Ca, free amino acids and a higherlevel of moisture. Based on confocal laser scanning microscopy analysis, AMSII cheese showed the lowestmicrostructural variations during the period of storage compared to other cheeses. All the sensory attributeswere scored highest for AMSII cheese. ASMII extend the shelf-life to ca. 12−15 days instead of the 5−7 days oftraditional high-moisture Mozzarella cheese.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Starters cultures are mainly species of lactic acid bacteria that aredeliberately added to milk where their primary role is to start theproduction of lactic acid for the cheese manufacturing process(Broome, 2007). It is well known that starter cultures fulfil otherimportant functions: inhibition of spoilage and pathogenic micro-organisms, improvement of cheese keeping quality, direct and indirectcontributions to flavour and aroma. Starter cultures may be classifiedinto two large groups: traditional (including artisanal, natural startersand multiple strain culture starters) and defined or commercialstarters (Limsowtin et al., 1995).
A high-moisture cows' Mozzarella cheese (also defined as “Fior diLatte”) is traditionally manufactured in the Southern Italy, mainly inthe Apulia region. Recently, its production has been estimated ca.220,400 tonnes per year (www.databank.it). During the last decade,the increasing consumption of cows' Mozzarella cheese favoured thedifferentiation of several manufacturing processes, which mainlydepended on the size of the cheese factories. Natural whey or milkstarter cultures represent one of the technological options and they
delle Piante e Microbiologiaudi di Bari, Bari 70126, Italy.
elis).
l rights reserved.
are daily produced and propagated at the cheese plant. Overall, thecomposition of natural starter cultures is complex, variable andsubjected to unpredictable performance (Coppola et al., 2001; deCandia et al., 2007). In most of the cases, natural starters have beenreplaced by (i) commercial starters, usually consisting of Streptococcusthermophilus alone or mixtures of S. thermophilus and Lactobacillushelveticus or Lactobacillus delbrueckii subsp. bulgaricus (Coppola et al.,2001; Coppola et al., 2006; de Candia et al., 2007) or by (ii) directacidification with lactic acid, citric acid or glucono-δ-lactone (Parenteand Moschetti, 1997). Overall, the choice between biological andchemical acidification and between natural and commercial startercultures may have important repercussion on several features of cows'Mozzarella cheese. The effect of defined starter cultures was studiedon physical, melt and rheological characteristics (Dave et al., 2003),microstructure (Merrill et al., 1996), and composition, proteolysis andfunctional properties of Mozzarella cheese (Yun et al., 1995). Thethermophilic lactic acid bacteria used to make Mozzarella cheese areparticularly important in determining the physical properties (Oberget al., 1991a,b). Besides, Yun et al. (1995) reported that a balanced rod-to-coccus ratio influenced proteolysis, springiness and viscosity ofMozzarella cheese. Overall, the use of commercially available startercultures seemed to cause a less characteristic flavour of the cheese(Broome, 2007). Several studies have characterized the microbialbiodiversity which occurs in natural whey starter cultures for cows'Mozzarella cheese. Molecular, physiological and technological

124 M. De Angelis et al. / International Journal of Food Microbiology 125 (2008) 123–132
analyses were carried out to select lactic acid bacteria to be eventuallyincluded in the starter formula for cheese manufacture (Morea et al.,1999; Bianchi et al., 2000). In a previous work (de Candia et al., 2007),the dominantmicrobial species of 7 natural whey starter cultures usedfor the manufacture of traditional cows' milk high-moisture Mozzar-ella cheese of the Apulia region were identified. A large biodiversitywas found at species level, which included mesophilic (Lactobacillusplantarum and/or Lactobacillus casei subsp. casei) and thermophilic(L. helveticus, L. delbrueckii subsp. lactis, L. delbrueckii subsp. bulgari-cus and Lactobacillus fermentum) lactobacilli, streptococci (S. thermo-philus) and enterococci (Enterococcus durans, Enterococcus faecium,Enterococcus faecalis and Enterococcus sp.). However, none of abovestudies has developed new autochthonous multiple strain cultures(AMS) which would be representative of the microbial composition ofthe natural whey cultures and would ensure high performance duringmaking of high-moisture traditional cows' Mozzarella cheese. Strainselection and optimization of a protocol for producing AMS to be usedby dairy factories appear to be pre-requisites to standardize the majortraits distinguishing this cheese variety.
This paper aimed at selecting AMS and at setting up the protocolfor the use of AMS to guarantee improved texture and sensoryproperties, and shelf-life of high-moisture traditional cows' Mozzar-ella cheese.
2. Materials and methods
2.1. Lactic acid bacteria and culture conditions
Hundred forty-six strains of lactic acid bacteria belonging to theCulture Collection of the Department of Plant Protection and AppliedMicrobiology, University of Bari, Italy, and previously isolated fromnaturalwhey starter cultures for ItalianMozzarella cheeses (deCandia et al., 2007)were used in this study. The following species were used: L. helveticus (30strains); L. fermentum (25); L. plantarum (13); L. delbrueckii subsp. lactis(8); L. casei subsp. casei (7); L. delbrueckii subsp. bulgaricus (7); Lactococcuslactis (16); Lactococcus garvieae (10); S. thermophilus (10); Ent. durans(5); Ent. faecium (3); Ent. faecalis (6); and Enterococcus sp. (6).
Strains were routinely cultivated at 30 °C (L. plantarum, L. caseisubsp. casei, Lc. lactis, Lc. garvieae); or 37 °C (L. helveticus, L. fermentum,L. delbrueckii subsp. lactis, L. delbrueckii subsp. bulgaricus, S. thermo-philus, Ent. durans, Ent. faecium, Ent. faecalis, and Enterococcus sp.) for24 h on the following broth media: MRS (Oxoid, Basingstoke,Hampshire, England) for lactobacilli; M17 broth (Oxoid) for lactococciand streptococci; and Slanetz & Bartley (Oxoid) for enterococci.
2.2. Kinetics of acidification during milk fermentation
Twenty-four-hours-old cells of 146 lactic acid bacteria strains wereharvested by centrifugation (9000 ×g for 15min at 4 °C), washed twicewith sterile 0.05 M potassium phosphate buffer, pH 7.0, re-suspendedin the sterile reconstituted skimmed milk (RSM) (Oxoid) at a celldensity of ca. 7.0 log10 CFU/ml. All cellular suspensions were incubatedat 30 °C or 37 °C for 18 h, and the pH was recorded on line. Cellnumbers were determined by plating on MRS, M17 or Slanetz &Bartley agar medium at 30 °C or 42 °C for 48 h.
Acidification data were modelled according to the Gompertzequation as modified by Zwietering et al. (1990). Titratable acidity(expressed as °SH/50) was determined by using 50 ml of fermentedRSM and 0.1 M of NaOH.
2.3. Autochthonous multiple strain cultures (AMS)
Cells of the most acidifying strains (L. helveticus 2B, L. delbrueckiisubsp. lactis 20F, L. plantarum 18A, S. thermophilus 22C, Ent. faecalis32C and Ent. durans 16E) were cultivated on RSM at 30 °C or 37 °C for18 h, and treated as reported above. The thermophilic and mesophilic
strains were composed to produce two autochthonous multiple strainculture starters (AMS). Based on previous works (Parente et al., 1989;Coppola et al., 1990; Oberg et al., 1991a,b; Yun et al., 1995; Savoie et al.,2007) two rod:coccus ratios were used. The first AMS (AMSI) consistedof L. helveticus 2B, L. delbrueckii subsp. lactis 20F, L. plantarum 18A,S. thermophilus 22C, Ent. faecalis 32C and Ent. durans 16E at the ratio1:1:1:1:0.5:0.5, respectively. The second AMS (AMSII) consisted ofL. helveticus 2B, L. delbrueckii subsp. lactis 20F, L. plantarum 18A, andS. thermophilus 22C at the ratio 1:1:1:4, respectively. Each type of AMSwas inoculated in commercial whey (Sigma Chemical Co. Milan, Italy)at a final cell density of ca. 7.0 log10 CFU/ml and incubated at 30 and37 °C for 18 h. To simulate the manufacture of Mozzarella cheese,cows' pasteurized milk was also inoculated with AMSI or AMSII (finalcell density of ca. 7.0 log10 CFU/ml) and incubated at 37 °C. After30 min, cows' liquid calf rennet (ca. 0.3 ml/kg) was added. At pH 5.1,the coagulum was cut and the whey milk was further incubated at37 °C for 18 h. Back-slopping propagation (10%, v/v) of both AMSI andAMSII in fresh commercial whey or whey milk was lasting 16 daysaccording to the protocol described above. During daily propagation,pH was recorded on line. At the end of daily propagation cell numbersand titratable acidity were determined also. Propagations werecarried out in triplicate from independent batches and data for thekinetics of acidification and growth were averaged.
Monitoring of strains during AMS propagation was by usingRandom Amplified Polymorphic DNA-Polymerase Chain Reaction(RAPD-PCR) analysis (Giraffa et al., 2000; De Angelis et al., 2006a).The molecular weight of the amplified DNA fragments was estimatedby comparison with a 1 kb plus ladder DNA (Invitrogen) ranging from100 to 12,000 bp.
2.4. Production of cows' Mozzarella cheese
Three types of cows' Mozzarella cheeses were manufactured usingselected AMSII, commercial starter culture (S. thermophilus, Sacco s.r.l.,Como, Italy) (CS) or direct acidification with citric acid (DA).Mozzarella cheeses were made in triplicate at the factory Gioiella-Capurso Azienda Casearia S.p.A. (Bari, Italy) and stored at 4 °C for15 days.
2.5. Lactic acid bacteria enumeration
Samples (10 g) of Mozzarella cheeses were diluted in 90ml sodiumcitrate (2% w/v) solution and homogenized. Serial dilutions weremade in quarter-strength Ringer's solution and plated on thefollowing agar media (Oxoid) for viable counts: MRS for mesophilicand thermophilic lactobacilli incubated at 30 and 42 °C for 48 h,respectively; M17 for lactococci and streptococci incubated at 30 and42 °C for 48 h, respectively, and Slanetz & Bartley for enterococciincubated at 37 °C for 48 h.
2.6. Compositional analysis
Mozzarella cheese samples were analysed for moisture (IDF, 1982),protein by macro-Kjeldahl method (IDF, 1964), fat by Gerber method(IIRS, 1955), ash and calcium by Guinee et al. (2000a). Salt in cheesewas determined potentiometrically (Fox, 1963), and pHwas measuredon a cheese: water slurry (1:2). All compositional analyses wereperformed in triplicate. Analysis of variancewas performed using SPSSstatistical package (Version 11.0, SPSS Inc, Chicago, USA) with Tukey'spair wise comparisons at the 95% confidence level.
2.7. Assessment of proteolysis
The pH 4.6-soluble and-insoluble extracts of theMozzarella cheesesamples were obtained as described by Kuchroo and Fox (1982). Thenitrogen content of these extracts was determined by the macro-

125M. De Angelis et al. / International Journal of Food Microbiology 125 (2008) 123–132
Kjeldahl method (IDF, 1964). The pH 4.6-insoluble fractions wereanalysed by urea-polyacrylamide gel electrophoresis (urea-PAGE)using a Protean IIXI vertical slab gel unit (Bio-Rad Laboratories Ltd,Watford, UK) and the stacking gel system described by Andrews(1983). Gels were stained by the method of Blakesley and Boezi (1977)with Coomassie Brilliant Blue G250, distained in distilled water andscanned on a flatbed scanner (Epson Perfection 4180 Photo, SeikoEpson Corporation, Nagano, Japan). Densitometric analysis wasperformed on the scanned image using gel analysis software (TotalLab 1D, Nonlinear Dynamix, Newcastle-Upon-Tyne, UK).
The peptide profiles of the pH 4.6-soluble fractions weredetermined by Reverse-Phase Fast Protein Liquid Chromatography(RP-FPLC) with a Resource RPC column as reported by Gobbetti et al.(1999), using an ÄKTA FPLC equipment with a UV detector operatingat 214 nm (Amersham Biosciences, Piscataway, NJ, USA). Peptideprofiles were analyzed by multivariate statistical techniques. The datafor factor reduction analysis were obtained by visually recognizing thepeaks and taking peak heights as variables (Pripp et al., 1999). Totaland individual free amino acids (FAA) of the pH 4.6-soluble fractionwere analyzed by a Biochrom 30 series Amino Acid Analyzer(Biochrom Ltd., Cambridge Science Park, England) with a Na-cation-exchange column (20 by 0.46 cm inner diameter). A mixture of aminoacids at known concentration (Sigma Chemical Co.) was added withcysteic acid, methionine sulphoxide, methionine sulphone, trypto-phan and ornithine, and used as standard. Amino acids were post-column derivatised with ninhydrin reagent and detected by absor-bance at 440 (proline and hydroxyproline) or 570 nm (all the otheramino acids). Data from FAA were subjected to Principal ComponentAnalysis (PCA). Statistical analyses were performed by using thesoftware Statistica for Windows (Statistica 6.0 per Windows 1998).
2.8. Confocal laser scanning microscopy (CLSM) and cheese firmness
Small pieces of cheese were taken from the whole Mozzarellacheese and put in isopentane cooled by liquid nitrogen at −150 °C, inorder to freeze the samples without formation of ice crystals. Twenty-μm-thick sections were achieved at −27 °C using a cryotome. Afterdrying on a slide, sections were stained by a few drops of a solutioncontaining Fluorescein Iso Thyo-Cyanate (FITC), labeling the proteins,and Nile Red, for the labeling of lipids. After 30 min, sections wererinsed using distilled water, and an upper lip was sealed on the sectionusing nail polish. Microscopic observations were performed using aLEICA Confocal microscope (LEICA Microsystems Heidelberg GmbH,Germany) used in epi mode (Herbert et al., 1999). Fluorescence imagesobtained for each component can be transferred into differentchannels, thus attributing a colour to each component. The super-imposition of the pictures shows the relative distribution of thecomponents in the system, protein appearing in green and lipids in red.
Four to six cylindrical samples were taken from each cheese, placedin anairtight plastic bag, held at8 °Covernight and compressed to 30%oforiginal height on a model 112 Universal Instron Testing Machine(Instron, Ltd., HighWycomb, UK), as described by Guinee et al. (2000b).Firmness was the force required to compress the samples of cheese to30% of the original height.
2.9. Sensory analysis
The sensory analysis of Mozzarella cheeses was determined by usingthe descriptive model of Coppola et al. (1990) with a few modifications.Intensity, firmness, sour and sweet were the sensory attributesconsidered by using a scale 0–5. A non-trained panel consisting of 10judges was used. Cheeses were cut into slices (approximately2.5×2.5 cm) at 1 h prior to serving and placed in plastic cups; cheeseswere randomized to avoid positional bias. Duplicate trays of sampleswere presented at10min intervals to thepanel. Resultswere classified aspreferable (total points: 13.9–20), barely acceptable (total points: 11–
12.9), needs modification (total points: 8–10.9) and not acceptable (totalpoints: 0–7.9).
3. Results
3.1. Screening based on the kinetics of acidification
Preliminarily, 146 strains of lactic acid bacteria, isolated fromnatural whey starter cultures (de Candia et al., 2007), were screenedbased on their kinetics of acidification during RSM fermentation(Parente, and Moschetti, 1997). After fermentation, all strains reacheda cell density which ranged from 7.9 to 9.2 log10 CFU/g. The stationaryphase of acidification was reached in 10 to 18 h depending on thestrain. The median values for ΔpH were found to be 0.88 and 2.15 at 4and 18 h, respectively. However, the ability of the strains to cause adecrease of the pH of RSM varied markedly. After 4 h of fermentation,the values corresponding to the 5th and 95th percentile of theaggregate strain data were 0.02 and 1.66, respectively. After 4 h, thehighest values of ΔpH for each species were the following: Lc. lactissubsp. lactis 10C (ΔpH 1.0; median value 0.45); Lc. garvieae 12C (ΔpH0.8; median value 0.40); L. plantarum 18A (ΔpH 1.19; median value1.10); L. helveticus 27D (ΔpH 1.58; median value 1.10); L. delbrueckiisubsp. lactis 20F (ΔpH 1.66; median value 1.15); L. delbrueckii subsp.bulgaricus 7E (ΔpH 0.84; median value 0.72); L. fermentum 17A (ΔpH1.28; median value 1.06); L. casei subsp. casei (ΔpH 0.64; median value0.40); Ent. durans 16E (ΔpH of 1.34; median value 0.89); Ent. faecium25D (ΔpH of 1.25; median value 0.47); Enterococcus sp. 18F (ΔpH 1.19;median value 1.16) and S. thermophilus 22C (ΔpH 0.81; median value0.09). After 18 h, the values corresponding to the 5th and 95thpercentile of the aggregate strain data were 0.70 and 3.56, respec-tively. The major part of the strains showed values of ΔpH whichranged from 1.5 to 2.7. L. plantarum 18A, L. helveticus 2B, L. delbrueckiisubsp. lactis 20F, S. thermophilus 22C, Ent. faecalis 32C and Ent. durans16E were the highest acidifying strains (ΔpH 3.0–3.1).
3.2. Selection of autochthonous multiple strain cultures
L. plantarum 18A, L. helveticus 2B, L. delbrueckii subsp. lactis 20F, S.thermophilus 22C, Ent. faecalis 32C and Ent. durans 16E were selectedand differently used in the AMSI and AMSII formula. Some of themajor pre-requisites that had to be reached for setting up the starterculture for Mozzarella cheesewere: (i) elevated initial cell density (9.0log10 CFU/g) to overcome delay in the acidification rate; (ii) rapidgrowth during propagation; and (iii) maintaining of the acidifyingactivity during prolonged propagation.
AMSI andAMSIIwere daily propagated in commercialwhey orwheymilk for 16 days at 30 or 37 °C. After 18 h of fermentation at 30or 37 °C incommercial whey, presumptive thermophilic and mesophilic lactoba-cilli, streptococci and enterococci of AMSI grew up to the 9.2±0.1–8.86±0.15 log10 CFU/ml. After 16 days of propagation at 30 °C, thermophiliclactobacilli, streptococci and enterococci decreased to 8.0±0.1, 8.1±0.2and 6.0±0.1 log10 CFU/ml, respectively. Cell numbers of mesophiliclactobacilli remained constant throughout 16 days. For AMSI, after16 days of propagation at 37 °C, the cell numbers of thermophilic andmesophilic lactobacilli and enterococci also decreased to 8.5±0.34, 7.5±0.37 and 7.0±0.21, respectively. Cell numbers of streptococci remainedconstant throughout 16 days. During 16 days of propagation incommercial whey at 30 or 37 °C, cell numbers of AMSII showed similarperformance as those found for AMSI.
After 18 h of fermentation at 37 °C in whey milk, cell numbers ofAMSI was similar as those found during propagation in commercialwhey. Under this condition, cell numbers of presumptive thermophilicand mesophilic lactobacilli and streptococci remained constantthroughout 16 days. After 16 days, only enterococci decreased to 7.0±0.28 log10 CFU/ml. Cell numbers of AMSII in whey milk remainedalmost constant throughout 16 days.

126 M. De Angelis et al. / International Journal of Food Microbiology 125 (2008) 123–132
Strains of AMSI and AMSII were monitored by RAPD-PCR analysisalso (Fig. 1A). After 18 h of propagation at 30 °C or 37 °C in commercialwhey or whey milk, all lactic acid bacteria included in the AMSI orAMSII formula were detected. Nevertheless, the ratios betweenspecies varied during 16 days. Only the propagation of AMSII on
Fig. 1. Random Amplified Polymorphic DNA-Polymerase Chain Reaction (RAPD-PCR)profiles of strains used for the production of autochthonous multiple strain cultures(AMSI and AMSII) (A) and strain fractions (%) of the lactic acid bacteria grown incommercial whey or whey milk started with AMSI (B) or AMSII (C) after 16 days ofcultures propagation. The initial microbial composition of AMSI and AMSII is reportedonMaterials and methods. The values were the average of three replicates and standarddeviation was indicated by vertical bars. S, DNA molecular size standards, 32C, Enter-ococcus faecalis 32C; 16E, Enterococcus durans 16E; 22C, Streptococcus thermophilus 22C;2B, Lactobacillus helveticus 2B; 20F, Lactobacillus delbrueckii subsp. lactis 20F; and 18A,Lactobacillus plantarum 18A.
whey milk at 37 °C allowed almost constant ratios between strainswhich approached those found initially (L. plantarum 18A, 14.28%;L. helveticus 2B, 14.28%; L. delbrueckii subsp. lactis 20F, 14.28% andS. thermophilus 22C, 57.12%) (Fig. 1B and C). After 16 days ofpropagation, L. delbrueckii subsp. lactis 20F was found only in AMSIIand Ent. durans 16E was not found in AMSI.
The acidification of AMSI and AMSII during 16 days of propagationin commercial whey at 30 °C or 37 °C showed median values of pH inthe range of 4.49–4.44 (Fig. 2A). Lower median values (4.08–3.9) werefound during propagation in whey milk at 37 °C. Apart from the typeof AMS and temperature, daily propagation in commercial whey wasalso characterized by large variation in the final values of pH. On thecontrary, a limited variation of pH was found when AMSI and,especially, AMSII were propagated at 37 °C inwheymilk. The values oftitratable acidity mirrored those found for pH (Fig. 2B). In particular,AMSII propagated in whey milk in a 37 °C showed almost uniformvalues of titratable acidity.
Based on the above results, AMSII to be propagated inwheymilkwasselected and used for the manufacture of high-moisture Mozzarellacheese.
4. Microbiological profile of Mozzarella cheese
Table 1 shows that the microbial counts for Mozzarella cheesesstartedwith AMSII, CS or DA. Numbers of thermophilic andmesophiliclactobacilli in CS cheese were low (5.50±0.34 and 5.74±0.24 log10CFU/g), while the cell density of streptococci and enterococci wassimilar to that found for AMSII cheese. DA cheese had a cell numbersof lactic acid bacteria lower than 6.0 log10 CFU/g which, except formesophilic lactobacilli, did not vary throughout storage at 4 °C. DAcheese showed an increase of coliforms of ca. 2.5 log cycles duringstorage at 4 °C. Apart from the type of starter (CS or AMSII), the cellnumbers of thermophilic lactobacilli and streptococci decreased of ca.1 log cycle during 15 days of storage.
4.1. Compositional analysis of Mozzarella cheese
The gross composition of Mozzarella cheeses varied markedly(Table 2). After 1 day of manufacture, all Mozzarella cheeses hadvalues of moisture which ranged from 57.50±1.25 (AMSII) to 65.41±1.20 (DA) (w/w). Based on these values, all cheese were considered ashigh-moisture Mozzarella. The concentrations of protein, fat andcalcium in DA cheese were significantly (P≤0.05) lower than in thecheeses started with CS and, especially, AMSII. After 1 day ofmanufacture, the pH of CS and AMSII cheeses was significantly(P≤0.05) lower than that of DA cheese. During storage, the pH of thecheeses started with CS and AMSII remained constant or slightlyincreased. The pH of DA cheese decreased, and after 15 days of storagethe value of pH of all the cheeses was ca. 5.25.
4.2. Proteolysis of Mozzarella cheese
After 1 day of manufacture, the level of pH 4.6-soluble nitrogen,expressed as % of total nitrogen in cheese, was in the range of 4.01–4.89% (Table 2). Compared to DA cheese, the CS and AMSII cheeses hadhigher levels of pH 4.6-soluble nitrogen. During refrigerated storagethe concentration of the pH 4.6-soluble nitrogen increased in allcheeses (5.0–5.5%).
Urea-PAGE electrophoregrams of the pH 4.6-insoluble nitrogen ofDA and AMSII cheeses during storage, showed for all samples thehydrolysis of β-casein (CN) to γ-CN, and αS-CN degradation to itsbreakdown products was also evident after 1 day of manufacture. Aprogressive hydrolysis of αS-CN occurred throughout storage espe-cially for AMSII cheese. The electrophoregrams of the pH 4.6-insolublenitrogen fraction of CS cheese did not significantly differ from that ofAMSII cheese (data not shown). The pH 4.6-soluble nitrogen fractions
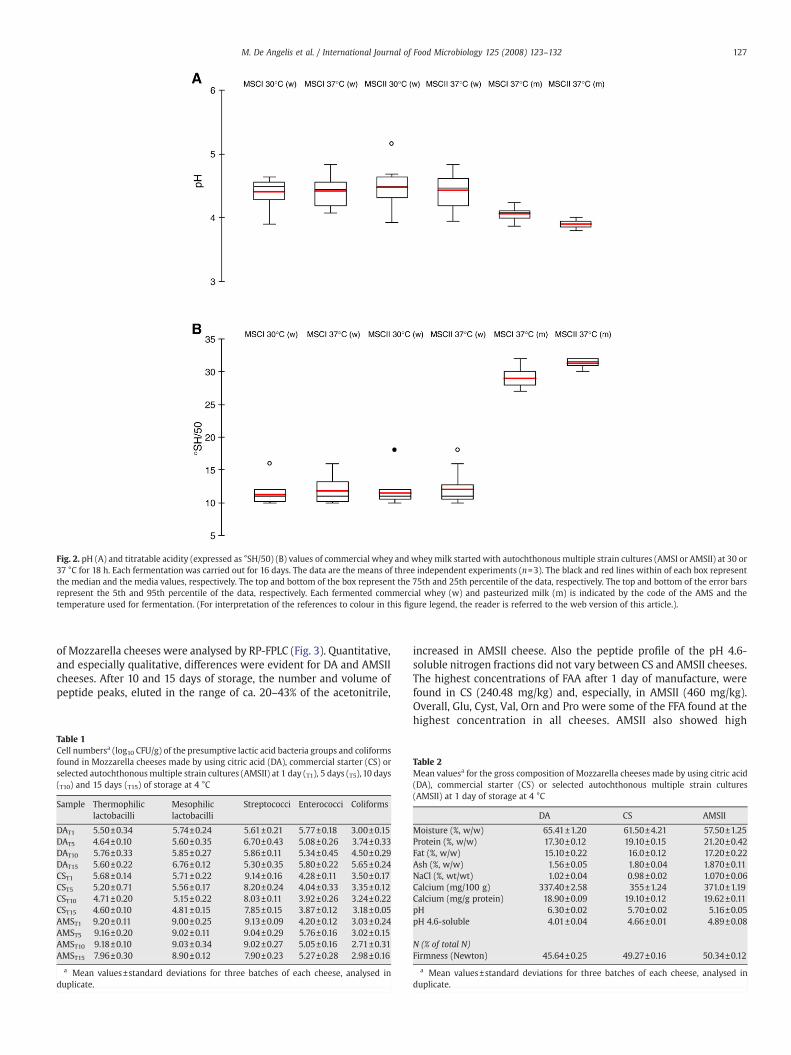
Fig. 2. pH (A) and titratable acidity (expressed as °SH/50) (B) values of commercial whey and wheymilk started with autochthonous multiple strain cultures (AMSI or AMSII) at 30 or37 °C for 18 h. Each fermentation was carried out for 16 days. The data are the means of three independent experiments (n=3). The black and red lines within of each box representthe median and the media values, respectively. The top and bottom of the box represent the 75th and 25th percentile of the data, respectively. The top and bottom of the error barsrepresent the 5th and 95th percentile of the data, respectively. Each fermented commercial whey (w) and pasteurized milk (m) is indicated by the code of the AMS and thetemperature used for fermentation. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.).
127M. De Angelis et al. / International Journal of Food Microbiology 125 (2008) 123–132
of Mozzarella cheeses were analysed by RP-FPLC (Fig. 3). Quantitative,and especially qualitative, differences were evident for DA and AMSIIcheeses. After 10 and 15 days of storage, the number and volume ofpeptide peaks, eluted in the range of ca. 20–43% of the acetonitrile,
Table 1Cell numbersa (log10 CFU/g) of the presumptive lactic acid bacteria groups and coliformsfound in Mozzarella cheeses made by using citric acid (DA), commercial starter (CS) orselected autochthonousmultiple strain cultures (AMSII) at 1 day (T1), 5 days (T5), 10 days(T10) and 15 days (T15) of storage at 4 °C
Sample Thermophiliclactobacilli
Mesophiliclactobacilli
Streptococci Enterococci Coliforms
DAT1 5.50±0.34 5.74±0.24 5.61±0.21 5.77±0.18 3.00±0.15DAT5 4.64±0.10 5.60±0.35 6.70±0.43 5.08±0.26 3.74±0.33DAT10 5.76±0.33 5.85±0.27 5.86±0.11 5.34±0.45 4.50±0.29DAT15 5.60±0.22 6.76±0.12 5.30±0.35 5.80±0.22 5.65±0.24CST1 5.68±0.14 5.71±0.22 9.14±0.16 4.28±0.11 3.50±0.17CST5 5.20±0.71 5.56±0.17 8.20±0.24 4.04±0.33 3.35±0.12CST10 4.71±0.20 5.15±0.22 8.03±0.11 3.92±0.26 3.24±0.22CST15 4.60±0.10 4.81±0.15 7.85±0.15 3.87±0.12 3.18±0.05AMST1 9.20±0.11 9.00±0.25 9.13±0.09 4.20±0.12 3.03±0.24AMST5 9.16±0.20 9.02±0.11 9.04±0.29 5.76±0.16 3.02±0.15AMST10 9.18±0.10 9.03±0.34 9.02±0.27 5.05±0.16 2.71±0.31AMST15 7.96±0.30 8.90±0.12 7.90±0.23 5.27±0.28 2.98±0.16
a Mean values±standard deviations for three batches of each cheese, analysed induplicate.
increased in AMSII cheese. Also the peptide profile of the pH 4.6-soluble nitrogen fractions did not vary between CS and AMSII cheeses.The highest concentrations of FAA after 1 day of manufacture, werefound in CS (240.48 mg/kg) and, especially, in AMSII (460 mg/kg).Overall, Glu, Cyst, Val, Orn and Pro were some of the FFA found at thehighest concentration in all cheeses. AMSII also showed high
Table 2Mean valuesa for the gross composition of Mozzarella cheeses made by using citric acid(DA), commercial starter (CS) or selected autochthonous multiple strain cultures(AMSII) at 1 day of storage at 4 °C
DA CS AMSII
Moisture (%, w/w) 65.41±1.20 61.50±4.21 57.50±1.25Protein (%, w/w) 17.30±0.12 19.10±0.15 21.20±0.42Fat (%, w/w) 15.10±0.22 16.0±0.12 17.20±0.22Ash (%, w/w) 1.56±0.05 1.80±0.04 1.870±0.11NaCl (%, wt/wt) 1.02±0.04 0.98±0.02 1.070±0.06Calcium (mg/100 g) 337.40±2.58 355±1.24 371.0±1.19Calcium (mg/g protein) 18.90±0.09 19.10±0.12 19.62±0.11pH 6.30±0.02 5.70±0.02 5.16±0.05pH 4.6-soluble 4.01±0.04 4.66±0.01 4.89±0.08
N (% of total N)Firmness (Newton) 45.64±0.25 49.27±0.16 50.34±0.12
a Mean values±standard deviations for three batches of each cheese, analysed induplicate.

Fig. 3. Reversed-Phase Fast-Protein Liquid Chromatography (RP-FPLC) of the pH 4.6-soluble fractions of the Mozzarella cheeses made by using citric acid (DA) (A) or selectedautochthonous multiple strain cultures (AMSII) (B) at 1 day (T1), 5 days (T5), 10 days (T10) and 15 days (T15) of storage at 4 °C. Three batches of DA and AMSII cheeses were analysed.mAU, milli-arbitrary units; ——, gradient of acetonitrile.
128 M. De Angelis et al. / International Journal of Food Microbiology 125 (2008) 123–132
concentrations of Lys and Ala (84.55–46.40 mg/kg, respectively). Thetotal concentration of FAA increased progressively throughout storageat 4 °C for all cheeses. After 15 days, the following concentrations ofFAA were found: 701.65, 920.24 and 1232.0 mg/kg for DA, CS andAMSII cheeses, respectively. FAA data were subjected to PrincipalComponent Analysis (PCA) using a covariance matrix (Fig. 4A and B).PC1 showed the distribution of the cheeses according to the totalconcentration of FAA, while PC2 differentiated samples based on therelative differences among individual amino acids. Cheese samplesmade with AMSII were clearly separated from the others, meaning ahigher concentration of FAA.
4.3. Confocal laser scanning microscopy (CLSM) and cheese firmness
Cheese microstructure and the relationship between its proteinmatrix and fat globules in all cheeses are depicted in Fig. 5 (A–F). At1 day, CLSM analyses revealed a protein network organized into afilamentous structure, and whey channels and fat globule clusterswere arranged along the protein fibers. Individual fat globules werealso observed residing within the protein matrix. DA cheese (Fig. 5A)showed a combination of many larger channels and few smaller poresgiving a very porous appearance to the overall protein network. CSand, especially, AMSII cheeses (Fig. 5C and E) appeared to have many

Fig. 4. Score plot (A) and loading plot (B) of the first and second principal components (PC) after Principal Component Analysis (PCA) based on free amino acids (FAA) of Mozzarellacheeses made by using citric acid (DA), commercial starter (CS) or selected autochthonousmultiple strain cultures (AMSII) at 1 day (T1), 5 days (T5), 10 days (T10) and 15 days (T15) ofstorage at 4 °C. Cheeses were carried out in triplicate from independent batches of cheese-making and data were averaged.
129M. De Angelis et al. / International Journal of Food Microbiology 125 (2008) 123–132
smaller channels. Following storage, the protein phase of theMozzarella cheeses became more continuous, entrapping and isolat-ing fat globules (Figs. 5B, 6D and F). Whey channels diminished in size,
Fig. 5. Confocal laser scanning micrographs of milk fat particles at the centre of Mozzarella chand D) or selected multiple strain cultures (AMSII) (panels E and F) at 1 day (panels A, C andgreen. As example, in panel A the protein network ( ), fat globule ( ) and channel (reader is referred to the web version of this article.).
indicating swelling of the casein and binding of the moisture to theprotein structure. CS and AMSII cheeses showed a characteristicdevelopment of the microstructure. The fat globules in the curd
eeses produced using citric acid (DA) (panels A and B), commercial starter (CS) (panels CE) and 15 days (panels B, D and F) of storage at 4 °C. Fat is stained in red and proteins in
➙) were indicated. (For interpretation of the references to colour in this figure legend, the

Fig. 6. Sensory analysis of Mozzarella cheeses produced using citric acid (DA) (♦), commercial starter (CS) (▲) or selected autochthonous multiple strain cultures (AMSII) (■) at 1 day(T1), 5 days (T5), 10 days (T10) and 15 days (T15) of storage at 4 °C. Four sensory attributes were used: flavour, sweet, sour and body/texture. The data are the means of threeindependent experiments.
130 M. De Angelis et al. / International Journal of Food Microbiology 125 (2008) 123–132
retained a similar appearance throughout storage, and individual fatglobules could still be clearly distinguished within the fat clusters in15 days old cheeses. DA developed a very dense protein network afterhydration of moisture and lacked this structure. AMSII was the onlycheeses that showed small channels until 15 days of storage.
The values of firmness decreased during storage, being influencedby the protocol of manufacture. The highest decreased of firmness wasfound for DA cheese (data not shown).
4.4. Sensory evaluation
DA Mozzarella cheese received the lowest score (Fig. 6). The maindefects noted for this cheese were body/texture (gummines until5 days and lack of structure after 10 days of storage at 4 °C) and flavour(flatness). After 10 days of storage, the samples were unsuitable formarket and the DA samples were excluded for further sensoryanalyses. About the started Mozzarella cheeses, all the sensoryattributes were significantly (Pb0.05) scored the highest for AMSIIsamples.
5. Discussion
Natural starter cultures used in the manufacture of Mozzarellacheese in the Apulia regionwere extremely variable inmicrobiologicalcomposition and performance (Morea et al., 1998, 1999; BianchiSalvatori et al., 2000; Coppola et al., 2001; De Angelis et al., 2006b; deCandia et al., 2007). Starter cultures with desirable properties are ofparticular interest to get reproducible and improved quality ofMozzarella cheeses. This paper aimed at selecting and using amultiple strain cultures consisting of autochthonous strains toguarantee optimal characteristics also suitable for reducing thevariability among Mozzarella cheeses. Preliminarily, strains (146)isolated from seven natural whey cultures, used as starters in theApulia region for the manufacture of Mozzarella cheese (de Candia etal., 2007), were screened. The growth and acidification activity in RSMmarkedly varied at species and strains level. Starter bacteria have to
produce lactic acid rapidly and in sufficient concentration to permit aneasy plasticization and stretching of the curd in hot water. Rapidacidification (ca. 2–4 h) allows the total time of cheese manufacturingto be shortened, which reduces the total amount of syneresis duringcheese-making and enables a higher moisture content to be achievedin the cheese (Barbano et al., 1994). Mesophilic lactococci (e.g., Lc.lactis) (Kindstedt et al., 2004), mesophilic lactobacilli (e.g., L. caseisubsp. casei) (Merrill et al., 1996) or thermophilic lactic acid bacteria(e.g., S. thermophilus, L. delbrueckii subsp. bulgaricus and L. helveticus)(Bianchi Salvatori et al., 2000; Kindstedt et al., 2004) may be usedalone or in mixture but the acidification rate and the concentration oflactic acid synthesized (optimum value of pH 5.1−5.4) may varymarkedly depending on the species. L. plantarum 18A, L. helveticus 2B,L. delbrueckii subsp. lactis 20F, S. thermophilus 22C, Ent. faecalis 32Cand Ent. durans 16E were selected as the most acidifying strains. Thesestrains correspond to the species that aremost frequently identified asthe dominant lactic acid bacteria of natural whey starter cultures usedfor the manufacture of Italian Mozzarella cheeses (Morea et al., 1998;Morea et al., 1999; Bianchi Salvatori et al., 2000; Coppola et al., 2001)and may guarantee optimal acidification and sensory properties(Gobbetti et al., 2007). Selection of fast-acidifying strains is only thefirst step of the work since the acidification properties must beperformed under very different and sub-optimal environmentalconditions (Savoie et al., 2007). A major problem when multiplestrain started cultures are used would be tomaintain the optimal ratiobetween strains (Coppola et al., 1990). Although natural startercultures are produced daily at the cheese plant by incubating someof the whey drained from the cheese vat under more or less selectiveenvironmental conditions, very often an extreme variability of themicrobial composition at strains and species level is found. A protocolwas developed which allowed the propagation and use of a definedAMS for the manufacture of Mozzarella cheese. The mixture of strainscomposed by L. plantarum 18A, L. helveticus 2B, L. delbrueckii subsp.lactis 20F and S. thermophilus 22C showed rapid and constantacidification capacity and maintained almost constant the ratiobetween strains under propagation in whey milk (Parente et al.,

131M. De Angelis et al. / International Journal of Food Microbiology 125 (2008) 123–132
1989; Coppola et al., 1990; Morea et al., 1998). Propagation underconditions which mimicked those used at dairy plant were preferredto the use of commercial whey as the growth medium. Interactionsbetween microorganisms, e.g. microbial co-operation and antagonis-tic effects, markedly impact on the survival, growth and activity ofmicroorganisms in multiple strain cultures and cheeses (Peláez andRequena, 2005). This study indicates that some phenotypic (growthand acidification) and genetic (RAPR-PCR) analyses could be sufficientto monitoring the microbial survival of multiple strain cultures.Overall, the usefulness of Enterococcus strains could not be shown, asthese organisms had poor competitiveness. They would also require athorough evaluation of their safety for the consumer. AMSII was usedas starter for the manufacture of high-moisture Mozzarella cheese atindustrial level and compared with Mozzarella cheeses made by usingthermophilic commercial starter (CS) or direct acidificationwith citricacid (DA). Overall, cell numbers of lactic acid bacteria were higher inAMSII cheese than in the CS and, especially, DA cheeses. Compared tothe other cheeses produced by natural whey starter cultures, theAMSII cheese had a significantly higher cell densities of presumptivelactic acid bacteria (de Candia et al., 2007). During 15 days of storage at4 °C, coliforms were shown to be negatively affected by lactic acidbacteria. This result may have repercussion on the shelf-life ofMozzarella cheese since coliforms are considered one of the bacterialgroups responsible of the most important spoilage phenomena onMozzarella balls (Altieri et al., 2005). Compared to other cheeses, theDA cheese had a significantly lower level of protein, ash, and calcium,and a higher level of moisture. The increased level of moisture in DAcheese was in agreement with other studies (Metzger et al., 2001;Guinee et al., 2002; Sheehan and Guinee, 2004) and it is consistentwith the concomitant reduction of the calcium content (Sheehan andGuinee, 2004). During storage, the pH of AMII and CS cheesesremained stable or slightly increase, while that of DA cheesedecreased. These differences may be related to variation of the cheesebuffering capacities as a consequence of the concentration of calcium(Metzger et al., 2001). Primary and secondary proteolysis affectstretch, melt, flavour and cook colour of non-ripened Mozzarellacheese (Oberg et al., 1991b) as well as the curd structure duringrefrigerated storage (Yun et al., 1993). Although with differencesbetween samples, the Mozzarella cheeses showed hydrolysis of bothαS- and β-CN due to the activity of chymosin and plasmin,respectively. Overall, secondary proteolysis as affected by startercultures may markedly vary depending on the strain. Thermophiliclactobacilli have a more intense proteolytic activity than S. thermo-philus. Among lactobacilli, L. helveticus ismore active than L. delbrueckiisubsp. bulgaricus or L. delbrueckii subsp. lactis, and, in general,mesophilic lactobacilli such as L. plantarum are characterized by anelevated peptidase activity (Oberg et al., 1991a; Yun et al.,1995;Merrillet al., 1996). AMSII showed a higher concentration of pH 4.6-solublenitrogen than CS and, especially, DA cheeses. The level of FAA in AMSIIwas significantly higher than that of the CS and, especially, in DA. Thiswas in agreement with previous findings on low moisture Mozzarellacheese made using starter culture or direct acidification (Feeney et al.,2002; Sheehan and Guinee, 2004). Cheese microstructure wasmonitored during storage to evaluate the protein matrix of thecheeses, the distribution of fat within the protein matrix and thehydration of moisture within the fat/serum channels. The protein andfat components are responsible for the resulting texture andfunctionality of cheeses (Zisu and Shah, 2005). The microstructure ofthe DA cheese differed markedly from the other cheeses. The proteinphase of the former was less fibrous and more swollen with largepores. These microstructural observations are consistent with thosefound previously for Mozzarella and string cheeses using scanningelectron microscopy (McMahon et al., 1993) or by using CLSM (Autyet al., 2001; Guinee et al., 2002; Zisu and Shah, 2005). The greaterdegree of protein swelling during the storage of CS and, especially, DAcheeses suggests an increase in casein hydration (Kindstedt and Guo,
1997). The age-related transition to a more uniform distribution of fatsuggests the swelling of the protein phase into regions previouslyoccupied by fat and serum (Kindstedt and Guo, 1997; McMahon et al.,1993; Guinee et al., 2002). Compared to other cheeses, AMSII cheeseshowed the lowest microstructural variations during the period ofstorage. The low firmness of the DA cheese may be associated with itsrelatively low level of intact casein (Guinee et al., 2000a), high-moisture content (Visser, 1991), low calcium content (Lawrence et al.,1984), and high degree of casein hydration. Overall, compared to theother cheeses produced by natural whey starter cultures, the AMSIIcheese had a significantly highest reproducible quality of Mozzarellacheeses (data not shown). By determining the sums of the scores forsensory properties, all DA samples were classified as barely acceptableuntil 5 days and needs modification at 10 days of storage. The CScheeses were barely acceptable and needs modifications until 10 and15 days, respectively. On the contrary, all AMSII cheeses were definedas preferable and barely acceptable until 10 and 15 days, respectively.CS cheese obtained consistently low scores for body/texture andflavour. This confirms the claim that using CS cultures for themanufacture of Mozzarella cheese causes a severe impoverishmentin organoleptic characteristics (Coppola et al., 1990).
6. Conclusions
This study proposes a protocol for propagation and use of anautochthonous multiple strain cultures to satisfy industrial require-ments. The starter consisted of four strong acidifying lactic acidbacteria strains (AMSII) which were selected from a large number ofnatural whey starter cultures and showed adaptation to severaltechnological factors. It was used as starter to acidify pasteurized milkfor 15 dayswithout loosing its acidifying properties and the stability ofthe microbiota. When selected AMSII was used for the manufacture ofhigh-moisture Mozzarella cheese, improved texture and sensoryproperties, and shelf-life were found. This protocol proposed couldbe also useful to extend the period of shelf-life to ca. 12−15 daysinstead of the 5−7 days of traditional high-moistureMozzarella cheese(Altieri et al., 2005; Brody, 2001; Laurienzo et al., 2006).
Acknowledgments
This work was supported by the Italian Ministry of the Universityand Research (M.I.U.R.) “Programmi di Ricerca di Interesse NazionaleAnno 2005. Functional activity of Streptococcus thermophilus underosmotic stress and microbial competition: a proteomic signature”.
References
Altieri, C., Scrocco, C., Sinigaglia, M., De Nobile, M.A., 2005. Use of chitosan to prolongMozzarella cheese shelf-life. Journal of Dairy Science 88, 2683–2688.
Auty, M.A.E., Twomey, M., Guinee, T.P., Mulvihill, D.M., 2001. Development andapplication of confocal scanning laser microscopy methods for studying thedistribution of fat and protein in selected dairy products. Journal of Dairy Research68, 417–427.
Andrews, A.T., 1983. Proteinases in normal bovine milk and their action on caseins.Journal of Dairy Research 50, 45–55.
Barbano, D.M., Yun, J.J., Kindstedt, P.S., 1994. Mozzarella cheese making by a stirred-curd, no brine procedure. Journal of Dairy Science 77, 2687–2694.
Bianchi Salvatori, B., Rottigli, C., Guerriero, P., 2000. Il formaggio Mozzarella Fiordilatte.L' industria del Latte XXXVI (3–4), 3–18.
Blakesley, R.W., Boezi, J.A., 1977. A new staining technique for proteins in polyacryla-mide gels using Coomassie Brilliant Blue G250. Analytical Biochemistry 82,580–582.
Brody, A.L., 2001. Say cheese and package it, please! Food Technology 55, 76–77.Broome,M.C., 2007. Starter culture development for improved cheeseflavour. InB.C.Weimer
CRC Press (Eds.), Improving the Flavour of Cheese Washington, DC, pp. 157–176.Coppola, S., Villani, F., Coppola, R., Parente, E., 1990. Comparison of different starter
systems for water-buffalo Mozzarella cheese manufacture. Lait 70, 411–423.Coppola, S., Blaiotta, G., Ercolini, D., Moschetti, G., 2001. Molecular evaluation of
microbial diversity occurring in different types of Mozzarella cheese. Journal ofApplied Microbiology 90, 414–420.
Coppola, S., Fusco, V., Andolfi, R., Aponte, M., Blaiotta, G., Ercolini, D., Moschetti, G.,2006. Evaluation of microbial diversity during the manufacture of Fior di Latte di

132 M. De Angelis et al. / International Journal of Food Microbiology 125 (2008) 123–132
Agerola, a traditional raw milk pasta-filata cheese of the Naples area. Journal ofDairy Research 73, 264–272.
Dave, I.R., Sharma, P., McMahon, J., 2003. Melt and rheological properties of Mozzarellacheese as affected by starter culture and coagulating enzymes. Lait 83, 61–77.
De Angelis, M., Siragusa, S., Berloco, M., Caputo, L., Settanni, L., Alfonsi, G., Amerio, M.,Grandi, A., Ragni, A., Gobbetti, M., 2006a. Isolation, identification and selection ofpotential probiotic lactobacilli from pig faeces to be used as additives in pelletedfeeding. Research in Microbiology 157, 792–801.
De Angelis, M., de Candia, S., Di Cagno, R., McSweeney, P.L.H., Gobbetti, M., 2006b.Microbiology and biochemistry of traditional mozzarella cheese. Australian Journalof Dairy Technology 61, 128–131.
de Candia, S., De Angelis, M., Dunlea, E., Minervini, F., McSweeney, P.L.H., Faccia, M.,Gobbetti, M., 2007. Molecular identification and typing of natural whey startercultures and microbiological and compositional properties of related traditionalMozzarella cheeses. International Journal of Food Microbiology 119, 182–191.
Feeney, E.P., Guinee, T.P., Fox, P.F., 2002. Effect of pH and calcium concentration onproteolysis in Mozzarella cheese. Journal of Dairy Science 85, 1646–1654.
Fox, P.F., 1963. Potentiometric determination of salt in cheese. Journal of Dairy Science23, 597–602.
Giraffa, G., Rossetti, L., Neviani, E., 2000. An evaluation of selex-based DNA purificationprotocols for the typing of lactic acid bacteria. Journal of Microbiology Methods 42,175–184.
Gobbetti, M., Folkertsma, B., Fox, P.F., Corsetti, A., Smacchi, E., De Angelis, M., Rossi, J.,Kilcawley, K., Cortini, M., 1999. Microbiology and biochemistry of Fossa (pit) cheese.International Dairy Journal 9, 763–773.
Gobbetti, M., De Angelis, R., Di Cagno, R., Rizzello, C.G., 2007. The relative contributionsof starter cultures and non-starter bacteria to the flavour of cheese. In B.C. WeimerCRC Press (Eds.), Improving the Flavour of Cheese Washington, DC, pp. 121–156.
Guinee, T.P., Mulholland, E.O., Mullins, C., Corcoran, M.O., 2000a. Effect of saltingmethod on the composition, yield, and functionality of low-moisture Mozzarellacheese. Milchwissenschaft 55, 135–138.
Guinee, T.P., Auty, M.A.E., Fenelon, M.A., 2000b. The effect of fat on the rheology,microstructure, and heat-induced functional characteristics of Cheddar cheese.International Dairy Journal 10, 277–288.
Guinee, T.P., Feeney, E.P., Auty, M.A.E., Fox, P.F., 2002. Effect of pH and calciumconcentration on some textural and functional properties of Mozzarella cheese.Journal of Dairy Science 85, 1655–1669.
Herbert, S., Bouchet, B., Riaublanc, A., Dufour, E., Gallant, D.J., 1999. Multiplefluorescence labelling of proteins, lipids and whey in dairy products using confocalmicroscopy. Lait, 79, 567–575.
IDF 1964. Determination of the protein content of processed cheeses products. IDFStandard 25.
IDF 1982. Cheese and processed cheese: Determination of the total solids content. IDFReferences method 4A.
IIRS 1955. Determination of the percentage of fat in cheese. IS Standard 69.Kindstedt, P.S., Guo, M.R., 1997. Recent developments in the science and technology of
pizza cheese. Australian Journal of Dairy Technology 52, 41–43.Kindstedt, P., Caric, M., Milanovic, S., 2004. Pasta-filata cheeses. In: Fox, P.F., Beresford,
Cogan H. (Eds.), Cheese: Chemistry, Physics and Microbiology, vol. 2. Elsevier Ltd.,pp. 251–277.
Kuchroo, C.N., Fox, P.F., 1982. Soluble nitrogen in Cheddar cheese: comparison ofextraction procedures. Milchwissenschaft 37, 331–335.
Laurienzo, P., Malinconico, M., Pizzano, R., Manzo, C., Piciocchi, N., Sorrentino, A., Volpe,M.G., 2006. Natural polysaccharide-based gels for dairy food preservation. Journalof Dairy Science 89, 2856–2864.
Lawrence, R.C., Heap, H.A., Gilles, J., 1984. A controlled approach to cheese technology.Journal of Dairy Science 67, 1642–1645.
Limsowtin, G.K.Y., Powel, I.B., Parente, E., 1995. Types of starters. In T. Cogan and J.P.Accolas VCH Publishers (Eds.), Dairy Starter Cultures. New York, NY, pp. 101–129.
McMahon, D., Oberg, C.J., McManus, W., 1993. Functionality of Mozzarella cheese.Australian Journal of Dairy Technology 48, 99–104.
Merrill, K.R., Oberg, J.O., McManus, R.W., Kalab, M., McManus, J., 1996. Microstructureand physical properties of a reduced fat Mozzarella cheese made using Lactobacilluscasei ssp. casei adjunct culture. Lebensmittel Wissenschaft and Technologie 29,721–728.
Metzger, L.E., Barbano, D.M., Kindstedt, P.S., Guo, M.R., 2001. Effect of milk preacidifica-tion on low fat Mozzarella cheese: II. Chemical and functional properties duringstorage. Journal of Dairy Science 84, 1348–1356.
Morea, M., Baruzzi, F., Cappa, F., Cocconcelli, P.S., 1998. Molecular characterization of theLactobacillus community in traditional processing of Mozzarella cheese. Interna-tional Journal of Food Microbiology 43, 53–60.
Morea, M., Baruzzi, F., Cocconcelli, P.S., 1999. Molecular and physiological characteriza-tion of dominant bacterial populations in traditional Mozzarella cheese processing.Journal of Applied Microbiology 87, 574–582.
Oberg, J.C., Merrill, K.R., Moyes, V.L., Brown, J.R., Richardson, H.G., 1991a. Effects ofLactobacillus helveticus on physical properties of mozzarella cheese. Journal ofDairy Science 74, 4101–4107.
Oberg, J.C., Wang, L., Moyes, V.L., Brown, J.R., Richardson, H.G., 1991b. Effects ofproteolytic activity of thermolactic cultures on physical properties of mozzarellacheese. Journal of Dairy Science 74, 389–397.
Parente, E., Moschetti, G., 1997. Starters for Mozzarella cheese. In Teagash Publushing(Eds.), Dublin, Fifth Cheese Symposium, Cork, Ireland, pp. 31–41.
Parente, E., Villani, F., Coppola, R., Coppola, S., 1989. A multiple strani starter for water-buffalo Mozzarella cheese manufacture. Lait 69, 271–279.
Peláez, C., Requena, T., 2005. Exploiting the potential of bacteria in the cheeseecosystem. International of Dairy Journal 15, 831–844.
Pripp, A., Shakeel-Ur-Rehman, H., McSweeney, P.L.H., Fox, P.F., 1999. Multivariatestatistical analysis of peptide profiles and free amino acids to evaluate effects ofsingle strain starters on proteolysis in miniature Cheddar-type cheeses. Interna-tional of Dairy Journal 9, 473–479.
Savoie, S., Champagne, C.P., Chiasson, S., Audet, P., 2007. Media and process parametersaffecting the growth, strain ratios and specific acidifying activities of a mixed lacticstarter containing aroma-producing and probiotic strains. Journal of AppliedMicrobiology 103, 163–174.
Sheehan, J.J., Guinee, T.P., 2004. Effect of pH and calcium level on the biochemical,textural and functional properties of reduced-fat Mozzarella cheese. Internationalof Dairy Journal 14, 161–172.
Visser, J., 1991. Factors affecting the rheological and fracture properties of hard andsemi-hard cheese. IDF Standard 268.
Yun, J.J., Barbano, D.M., Kiely, L.J., Kindstedt, P.S., Barbano, D.M., 1993. Mozzarellacheese: impact of coagulant type on functional properties. Journal of Dairy Science76, 3657–3663.
Yun, J.J., Kiely, L.J., Kindstedt, P.S., 1995. Mozzarella cheese: impact of rod:coccus ratio oncomposition, proteolysis and functional properties. Journal of Dairy Science 78,751–760.
Zisu, B., Shah, N.P., 2005. Textural and functional changes in low-fat Mozzarella cheesesin relation to proteolysis and microstructure as influenced by the use of fatreplacers, pre-acidification and EPS starter. International Dairy Journal 15, 957–972.
Zwietering, M.H., Jongeberger, I., Roumbouts, F.M., van't Riet, K., 1990. Modelling ofbacterial growth curve. Applied Environmental and Microbiology 56, 1875–1881.




















