
PEMENUHAN PRINSIP ETIKA LINGKUNGAN DALAM RENCANA PEMBANGUNAN PROYEK GIANT SEA WALL DI TELUK JAKARTA
Upload
darussalamCategory
view
0download
0
Pertemuan Ilmiah Nasional Tahunan X ISOI 2013
Jakarta, 11-12 November 2014
Sebaran Spasio-Temporal Komunitas Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania...
280
SEBARAN SPASIO-TEMPORAL KOMUNITAS IKAN PADANG LAMUN
PERAIRAN PULAU BUNTAL-TELUK KOTANIA, SERAM BARAT
SPACIO-TEMPORAL DISTRIBUTION OF SEAGRASS FISH COMMUNITY
IN THE BUNTAL ISLAND-KOTANIA BAY, WEST SERAM
Husain Latuconsina, Abdul Rahim Lestaluhu dan Maulana Abas Al’aidi
Fakultas Perikanan dan Ilmu Kelautan Universitas Darussalam,
Jln. Raya Tulehu Km. 24 Ambon 97582
Email: [email protected]
Abstrak
Penelitian dilaksanakan pada bulan Juni 2013 di ekosistem padang lamun perairan pulau Buntal Teluk
Kotania, Seram Barat, Maluku. Penelitian ini bertujuan untuk mengetahui sebaran spasio-temporal
komunitas ikan padang lamun. Data komunitas ikan dikoleksi menggunakan jaring insang dasar
dengan ukuran mata jaring 2 inci dan panjang 300m yang diletakkan sejajar garis pantai pada hamparan
padang lamun. Koleksi, dilakukan 10 kali ulangan pada siang dan malam hari di dua stasiun dengan
karakteristik fisik habitat lamun berbeda. Hasil penelitian ditemukan ikan dengan total 2.678 individu,
65 spesies, dan 33 famili. Pada stasiun I dikoleksi 1.224 individu, 54 spesies, dan 29 famili, yang
terdistribusi pada siang hari sebanyak 488 individu, 43 spesies, 23 famili, dan pada malam hari
sebanyak 736 individu, 30 spesies, 16 famili. Pada stasiun II dikoleksi 1.456 individu, 23 spesies, 18
famili, yang tersebar pada siang hari sebanyak 602 individu, 19 spesies, 13 famili dan pada malam hari
sebanyak 852 individu, 18 spesies, 13 famili. Hasil uji Uji-t mendapatkan perbedaan signifikan secara
spasial indeks keanekaragaman komunitas ikan dengan nilai tertinggi pada habitat lamun yang diapit
habitat mangrove dan terumbu karang, dan secara temporal berbeda signifikan dengan nilai tertinggi
pada siang hari. Sebaliknya indeks dominansi tertinggi pada malam hari, namun tidak berbeda
signifikan antar stasiun pengamatan. Hasil penelitian membuktikan peranan ekologi padang lamun
sebagai habitat ikan ekonomis penting khususnya Siganus canaliculatus. Adanya konektivitas antara
mangrove, padang lamun dan terumbu karang terkait distribusi komunitas ikan, sehingga diperlukan
upaya konservasi ketiga habitat tersebut untuk keberlanjutan pemanfaatan sumberdaya ikan.
Kata kunci: Padang Lamun, Komunitas ikan, Pulau Buntal, Teluk Kotania-Seram Barat
Abstract
This research was conducted on June 2013 in the seagrass beds of the Buntal Island-Kotania Bay of
West Seram-Mollucas, for knowing the spasio-temporal distribution of seagrass fish community. Data
of fish community was collected using Gill nets with mesh size 2 inches and 300 m of length are placed
parallel to the coastline at the expanse of the seagrass beds, done 10 times repeat day and night time
on two stations with different physical characteristics seagrass habitat. Results of the study found fish
with total 2,678 individuals, 65 species, 33 family. At the station I were 1224 individual, 54 species and
29 family, which during the day as much as 488 individuals, 43 species 23 family, and at night as
many as 736 individual, 30 species, 16 family. On The Stastion II were 1.456 individuals, 23 species,
18 family, which spread during the day as much as 602 individual, 19 species in the 13 family and at
night as 852 individuals, 18 species, 13 families. The results of the test comparative get distinction
spatial diversity index of with the highest value in habitats that are enclosed in mangrove habitats
seagrass beds and coral reefs, and a temporal different significant with the highest value during the
daytime. Contrarily index dominance highest at night, but it is not different significant between an
observation station. Research results prove role ecology seagrass beds puffer waters off as the habitat
resources economically important fishes especially Siganus canaliculatus. The connectivity between
mangrove, seagrass beds and coral reefs related distribution of fish community, so efforts to
conservation of third habitats was required for sustainability resources utilization of fish.
Keywords: Seagrass beds, fish community, Buntal Island, Kotania Bay West Seram

Latuconsina et al.
281 Pertemuan Ilmiah Nasional Tahunan X ISOI 2013 Jakarta, 11-12 November 2014
I. PENDAHULUAN
Peranan ekosistem padang lamun (seagrass beds) salah satunya bagi
kehidupan komunitas ikan yaitu sebagai daerah asuhan (nursery ground), mencari
makan (feeding ground) dan perlindungan (Latuconsina et al 2012 dan 2013;
Latuconsina & Ambo-Rappe 2013; Ambo-Rappe et al 2013), serta alur migrasi antara
habitat yang memanfaatkan mekanisme pasang surut untuk terdistribusi pada habitat
lamun dan mangrove (Unsworth, 2007; Latuconsina et al 2012).
Supriadi et al (2004) menemukan perbedaan kehadiran spesies ikan antara
periode siang dan malam hari pada ekosistem padang lamun perairan pulau Barang
Lompo-Makassar. Penelitiannya menemukan bahwa spesies ikan yang tertangkap
pada malam hari tidak ditemukan pada siang hari yang menunjukkan sifat nokturnal
dan diurnal komunitas ikan penghuni padang lamun.Sementara Latuconsina & Ambo-
Rappe (2013) menemukan perbedaan struktur komunitas ikan antara siang dan malam
hari, dengan kelimpahan dan indeks dominansi lebih tinggi pada malam hari,
sementara indeks keanekaragaman lebih tinggi pada siang hari. Perbedaan ini diduga
terkait orientasi keberadaan ikan yang berbeda, baik untuk mencari makan maupun
untuk berlindung.Santos et al (2002 dalam Pereira et al, 2010) menemukan ikan
bervariasi sepanjang hari yang minimum pada malam hari dan maksimum pada siang
hari, dengan aktifitas migrasi kebayakan terjadi pada pagi dan sore hari.
Perairan pantai pulau Buntal-Teluk Kotania memiliki ekosistem yang unik
karena dijumpai ekosistem mangrove, padang lamun dan terumbu karang yang saling
berdekatan. Ekosistem padang lamun ditemukan mengelilingi pulau Buntal dengan
substrat yang bervariasi meliputi:pasir berlumpur, pasir kasar, dan patahan karang,
sehingga mempengaruhi sebaran spasial vegetasi lamun dengan kerapatan yang
berbeda, dengan membentuk vegetasi multispesifik.
Adanya kerapatan vegetasi lamun yang berbeda dan kedekatannya dengan
ekosistem mangrove dan terumbu karang akan turut mempengaruhi struktur
komunitas ikan secara spasial (Ambo-Rappe et al 2013), dan adanya sifat nokturnal
dan diurnal ikan padang lamun mempengaruhi struktur komunitas ikan secara
temporal (Latuconsina & Ambo-Rappe, 2013). Untuk membuktikan hal ini maka
penelitian distribusi spasial temporal komunitas ikan padang lamun di perairan Pulau
Buntal,Teluk Kotania perlu dilakukan. Hasil penelitian ini diharapkan sebagai
informasi ilmiah dalam mengetahui peranan ekologi ekosistem padang lamun bagi
komunitas ikan untuk kepentingan konservasi dan pemanfaatan sumberdaya ikan
secara berkelanjutan..
II. METODE PENELITIAN
2.1. Waktu dan Tempat
Penelitian dilakukan pada bulan Juni 2013 di ekosistem padang lamun
perairan pulau Buntal-Teluk Kotania, Kabupaten Seram Barat. Lokasi penelitian
dibagi menjadi dua stasiun yang ditentukan berdasarkan perbedaan karakteristik
habitat lamun meliputi kerapatan vegetasi dan kedekatannya dengan ekosistem
mangrove maupun terumbu karang, yaitu :
Stasiun I : terletak pada posisi 3° 3'19.15" LS - 128° 4'56.99" BT, dengan
karakteristik fisik sedimen didominasi pasir halus bercampur pasir kasar dengan
panjang garis pantai 300m dan lebar 100m yang masih ditumbuhi vegetasi lamun,
ditemukan juga vegetasi mangrove yang cukup padat dan terumbu karang yang
mengapit padang lamun.

Pertemuan Ilmiah Nasional Tahunan X ISOI 2013
Jakarta, 11-12 November 2014
Sebaran Spasio-Temporal Komunitas Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania...
282
Satsiun II : terletak pada posisi 3° 3'19.58" LS - 128° 4'44.80" BT dengan
karakteristik fisik sedimen didominasi pasir halus berlumpur dengan panjang garis
pantai 400m dan lebar 200m yang ditumbuhi vegetasi lamun, dan ditemukan
vegetasi mangrove namun tidak terlalu padat.
2.2. Teknik Sampling Komunitas Ikan
Ikan dikoleksi mengunakan jaring insang dengan ukuran mata jaring 2inci
diletakkan pada hamparan padang lamun. Pengamatan dilakukan sebanyak 20 kali (10
kali masing-masing mewakili siang dan malam) selama pasang bergerak surut pada
setiap stasiun pengamatan. Ikan yang tertangkap ditempatkan pada kantong plastik
berlabel kemudian dilakukan identifikasi jenis (spesies) dan dihitung jumlah dan
diukur panjangnya (TL). Identifikasi spesies ikan menggunakan acuan dari Allen
(1999), Carpenter & Niem (1999& 2001), Kuiter & Tonozuka (2001), Peristiwady
(2006), dan Allen & Erdmann (2012). Parameter fisika-kimia lingkungan perairan
diamati setiap kali penangkapan ikan, meliputi : kedalaman, kecerahan, kecepatan
arus, suhu dan salinitas yang diukur secara insitu.
2.3. Analisa Data
Struktur kominitas ikan dianalisis meliputi: indeks dominasi dan
keanekaragaman.Nilai indeks dominasi memberikan gambaran dominasi ikan dalam
komunitas ekologi, yang menerangkan suatu spesies lebih banyak selama
pengambilan data. Formula Margalef (1958 dalam Odum 1983) :
C =
2
Dimana : C = Indeks Dominasi Simpson
N = Jumlah Individu Seluruh Spesies
ni = Jumlah Individu Dari Spesies ke-i
Indeks keanekaragaman adalah nilai yang menunjukkan keseimbangan dalam
suatu pembagian jumlah individu tiap spesies. Indeks keanekaragaman mempunyai
nilai terbesar jika semua individu berasal dari spesies yang berbeda. Sedangkan nilai
terkecil didapat jika semua individu berasal dari satu satu spesies saja (Odum, 1983).
Nilai indeks keanekaragaman Shannon (H’) menurut Shannon and Wiener (1949
dalam Odum 1983) dihitung menggunakan formula :
H’ = -∑ Pi In (Pi)
Dimana : H’ = Indeks Keanekaragaman.
Pi = Proporsi Jumlah Individu (ni/N).
Tabel 1. Kriteria Nilai Struktur Komunitas (Setyobudiandy et al,2009)
Indeks Kisaran Katagori
Dominasi (C) 0,00 < C ≤ 0,50
0.50 < C ≤ 0,75
0,75 < C ≤ 1.00
Rendah
Sedang
Tinggi
Keanekaragaman (H’) H’ ≤ 2
2,0 < H’ ≤ 3
H’ ≥ 3.0
Rendah
Sedang
Tinggi
Perbandingan variasi spasial-temporal jumlah individu, spesies dan struktur
komunitas ikan menggunakan uji-t (independent smple test) dengan bantuan program
SPSS vs 17 (Priyatno, 2009).Tingkat pengelompokan kesamaan spesies ikan padang
lamun berdasarkan kehadiran spesies secara spasial antara stasiun dan secara temporal

Latuconsina et al.
283 Pertemuan Ilmiah Nasional Tahunan X ISOI 2013 Jakarta, 11-12 November 2014
antara siang dan malam hari menggunakan indeks kesamaan Bray-Curtis yang
ditampilkan dalam bentuk dendrogram. Pengolahan data menggunakan program
PRIMER vs. 5.
III. HASIL DAN PEMBAHASAN
3.1. Gambaran Umum Perairan Pantai Pulau Buntal
Pulau Buntal terletak dalam kawasan Teluk Kotania, sebelah Timur berbatasan
dengan pulau Tatumbu, Selatan berbatasan dengan Dusun Wael (daratan pulau
Seram) dan sebelah Utara berbatasan dengan pulau Osi (Gambar 1).
Gambar 1. Lokasi penelitian pada ekosistem padang lamun perairan Pulau Buntal-
Teluk Kotania, Seram Barat.
3.2. Kerapatan Jenis lamun
Vegetasi lamun pada perairan pulau Buntal-Teluk Kotania tersebar luas
dengan membentuk padang lamun multispesifik yang tersusun atas lima spesies
dengan kerapatan berbeda (Tabel 2).
Tabel 2. Kerapatan spesies lamun (ind/m2)
No Jenis Lamun Kerapatan vegetasi Lamun
Stasiun I Stasiun II
1. Cymodocea serrulata 4.7 3.3
2. Cymodocea rotundata 7.7 6.4
3. Thalasia hemprichii 43.6 31
4. Enhalus acoroides 29.6 22.2
5. Halophila ovalis 4 4.7
Berdasarkan Tabel 2, menunjukkan kerapatan vegetasi lamun tertinggi
ditemukan pada stasiun I untuk semua spesies lamun. Thalassia hemprichii memiliki
kerapatan tertinggi dan terendah adalah Halophila ovalis dan Cymodocea serrulata.
Tingginya kerapatan dan keragaman vegetasi lamun berpotensi mendukung tingginya
kelimpahan dan keragaman ikan pada perairan pulau Buntal yang dapat berfungsi
sebagai tempat mencari makan, asuhan dan pembesaran bagi ikan-ikan ekonomis
penting, karena terdapat hubungan erat antara karakteristik vegetasi lamun dengan
struktur komunitas ikan yang berasosiasi di dalamnya (Ambo-Rappe et al, 2013),
dimana kerapatan dan keragaman vegetasi lamun yang tinggi mendukung tingginya
kelimpahan ikan (Ambo-Rappe, 2010; Latuconsina et al, 2012).
Pertemuan Ilmiah Nasional Tahunan X ISOI 2013
Jakarta, 11-12 November 2014
Sebaran Spasio-Temporal Komunitas Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania...
284
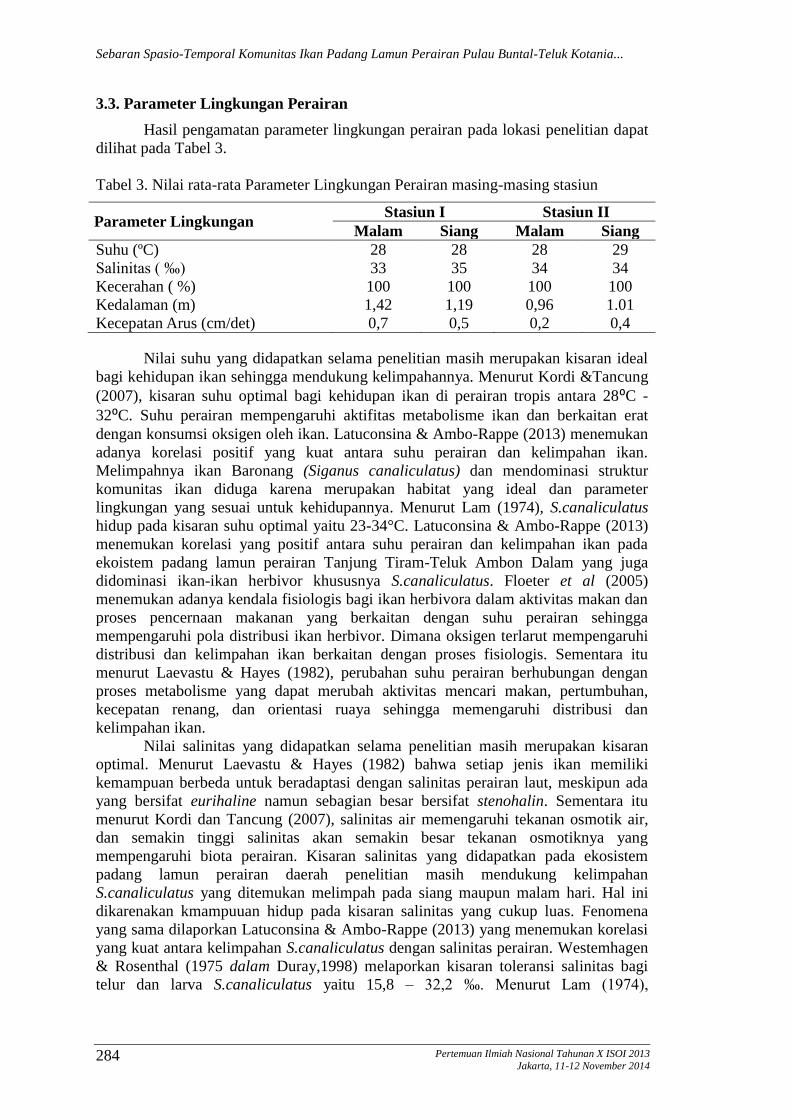
3.3. Parameter Lingkungan Perairan
Hasil pengamatan parameter lingkungan perairan pada lokasi penelitian dapat
dilihat pada Tabel 3.
Tabel 3. Nilai rata-rata Parameter Lingkungan Perairan masing-masing stasiun
Parameter Lingkungan Stasiun I Stasiun II
Malam Siang Malam Siang
Suhu (ºC) 28 28 28 29
Salinitas ( ‰) 33 35 34 34
Kecerahan ( %) 100 100 100 100
Kedalaman (m) 1,42 1,19 0,96 1.01
Kecepatan Arus (cm/det) 0,7 0,5 0,2 0,4
Nilai suhu yang didapatkan selama penelitian masih merupakan kisaran ideal
bagi kehidupan ikan sehingga mendukung kelimpahannya. Menurut Kordi &Tancung
(2007), kisaran suhu optimal bagi kehidupan ikan di perairan tropis antara 28⁰C -
32⁰C. Suhu perairan mempengaruhi aktifitas metabolisme ikan dan berkaitan erat
dengan konsumsi oksigen oleh ikan. Latuconsina & Ambo-Rappe (2013) menemukan
adanya korelasi positif yang kuat antara suhu perairan dan kelimpahan ikan.
Melimpahnya ikan Baronang (Siganus canaliculatus) dan mendominasi struktur
komunitas ikan diduga karena merupakan habitat yang ideal dan parameter
lingkungan yang sesuai untuk kehidupannya. Menurut Lam (1974), S.canaliculatus
hidup pada kisaran suhu optimal yaitu 23-34°C. Latuconsina & Ambo-Rappe (2013)
menemukan korelasi yang positif antara suhu perairan dan kelimpahan ikan pada
ekoistem padang lamun perairan Tanjung Tiram-Teluk Ambon Dalam yang juga
didominasi ikan-ikan herbivor khususnya S.canaliculatus. Floeter et al (2005)
menemukan adanya kendala fisiologis bagi ikan herbivora dalam aktivitas makan dan
proses pencernaan makanan yang berkaitan dengan suhu perairan sehingga
mempengaruhi pola distribusi ikan herbivor. Dimana oksigen terlarut mempengaruhi
distribusi dan kelimpahan ikan berkaitan dengan proses fisiologis. Sementara itu
menurut Laevastu & Hayes (1982), perubahan suhu perairan berhubungan dengan
proses metabolisme yang dapat merubah aktivitas mencari makan, pertumbuhan,
kecepatan renang, dan orientasi ruaya sehingga memengaruhi distribusi dan
kelimpahan ikan.
Nilai salinitas yang didapatkan selama penelitian masih merupakan kisaran
optimal. Menurut Laevastu & Hayes (1982) bahwa setiap jenis ikan memiliki
kemampuan berbeda untuk beradaptasi dengan salinitas perairan laut, meskipun ada
yang bersifat eurihaline namun sebagian besar bersifat stenohalin. Sementara itu
menurut Kordi dan Tancung (2007), salinitas air memengaruhi tekanan osmotik air,
dan semakin tinggi salinitas akan semakin besar tekanan osmotiknya yang
mempengaruhi biota perairan. Kisaran salinitas yang didapatkan pada ekosistem
padang lamun perairan daerah penelitian masih mendukung kelimpahan
S.canaliculatus yang ditemukan melimpah pada siang maupun malam hari. Hal ini
dikarenakan kmampuuan hidup pada kisaran salinitas yang cukup luas. Fenomena
yang sama dilaporkan Latuconsina & Ambo-Rappe (2013) yang menemukan korelasi
yang kuat antara kelimpahan S.canaliculatus dengan salinitas perairan. Westemhagen
& Rosenthal (1975 dalam Duray,1998) melaporkan kisaran toleransi salinitas bagi
telur dan larva S.canaliculatus yaitu 15,8 – 32,2 ‰. Menurut Lam (1974),

Latuconsina et al.
285 Pertemuan Ilmiah Nasional Tahunan X ISOI 2013 Jakarta, 11-12 November 2014
S.canaliculatus secara bertahap melalui proses aklimatisasi dapat berkembang pada
nilai salinitas 5 ‰ di bawah kisaran salinitas optimum.
Kecepatan arus selama pengamatan pada periode malam hari stasiun I sebesar
0,7 cm/det dan siang hari sebesar 0,5 cm/det, sementara pada periode malam hari
stasiun II sebesar 0,2 cm/det dan siang hari sebesar 0,4 cm/det. Menurut Laevastu &
Hayes (1981), arus mempengaruhi transportasi telur, larva, ikan-ikan kecil, dan
berperan dalam menentukan orientasi migrasi.
Kecerahan perairan selama pengamatan pada periode baik malam hari maupun
siang hari dan pada semua stasiun pengamatan mendapatkan nilai sebesar100%.
Tingginya kecerahan pada lokasi penelitian disebabkan tidak ada pengaruh sungai
yang membawa partikel tersuspensi maupun aktivitas manusia di daratan. Menurut
Effendi (2003) kekeruhan yang tinggi dapat menyebabkan terganggunya sistem
osmoregulasi, pernafasan dan daya lihat organisme akuatik. Kecerahan berkaitan
dengan tingkat kekeruhan pada kolom perairan, dimana kecerahan yang tinggi
menunjukkan tingkat kekeruhan perairan yang rendah.
Kedalaman perairan selama pengamatan pada periode malam hari stasiun
Isebesar 1,43 m dan siang hari sebesar 1,19 m.Sementara pada periode malam hari
stasiun II sebesar 0,97 m dan siang hari sebesar 1.01 m. Tinggi rendahnya kedalaman
perairan sangat dipengaruhi oleh ritme pasang surutyang memberikan perbedaan
ruang gerak dan pendistribusian sumber makanan serta parameter fisika-kimia
perairan. Latuconsina et al (2012) menemukan perbedaan kelimpahan ikan antara
periode pasang purnama dan pasang perbadi di perairan Tanjung Tiram-Teluk Ambon
Dalam, diduga disebabkan tinggi rendahnya permukaan air dan arus pasang surut
sehingga memengaruhi distribusi ikan. Menurut Romimohtarto & Juana (2004) gejala
pasang surut mempengaruhi tinggi rendahnya permukaan laut, sehingga secara
biologis menstimulasi biota laut dalam hal penyebaran, pemangsaan dan pemijahan.
3.4. Distirbusi Spasio-Temporal Komunitas Ikan
Adanya perbedaan komposisi spesies baik spasial maupun temporal (Tabel 4)
pada daerah penelitian membuktikan bahwa distribusi ikan dipengaruhi oleh
perbedaan karakteristik habitat dan sifat nokturnal dan diurnal ikan yang diduga
terkait dengan fluktuasi parameter fisik-kimia dan biologi lingkungan perairan.
Berdasarkan Tabel 4, jumlah individu yang lebih tinggi pada periode malam
hari, dan pada stasiun II, sedangkan jumlah spesies tertinggi ditemukan pada periode
siang hari dan pada stasiun I.Tingginya kelimpahan ikan pada malam hari karena
beberapa spesies ditemukan sangat dominan seperti S.canaliculatus yang
mendominasi pemanfaatan ruang, oksigen terlarut dan transfer energi melalui proses
rantai makanan, sehingga secara alamiah menurunkan jumlah dan komposisi spesies
lainnya secara merata. Lebih melimpahnya ikan pada malam hari diduga terkait sifat
nokturnal ikan yang lebih aktif pada malam hari (Supriadi et al. 2004; Latuconsina &
Ambo-Rappe, 2013), dan diduga akibat pengaruh ritme pasang yang merangsang ikan
dengan tingkatan trofik yang berbeda untuk terdistribusi pada ekosistem padang
lamun dari ekosistem terdekat seperti mangrove dan terumbu karang (Hindell et al.
2000, Unsworth et al. 2009, dan Latuconsina et al. 2012).
Sementara itu tingginya jumlah spesies ikan pada stasiun I diduga terkait
lokasinya yang berdekatan dengan eksosistem mangrove dan terumbu karang.
Menurut Bell & Pollard (1989), bahwa hubungan yang kuat terjadi antara padang
lamun dan habitat yang terdekat, dimana kelimpahan relatif dan komposisi spesies
ikan di padang lamun menjadi tergantung pada tipe (terumbu karang, estuaria,
mangrove) dan jarak dari habitat yang terdekat.
Pertemuan Ilmiah Nasional Tahunan X ISOI 2013 Jakarta, 11-12 November 2014
Sebaran Spasio-Temporal Komunitas Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania...
286
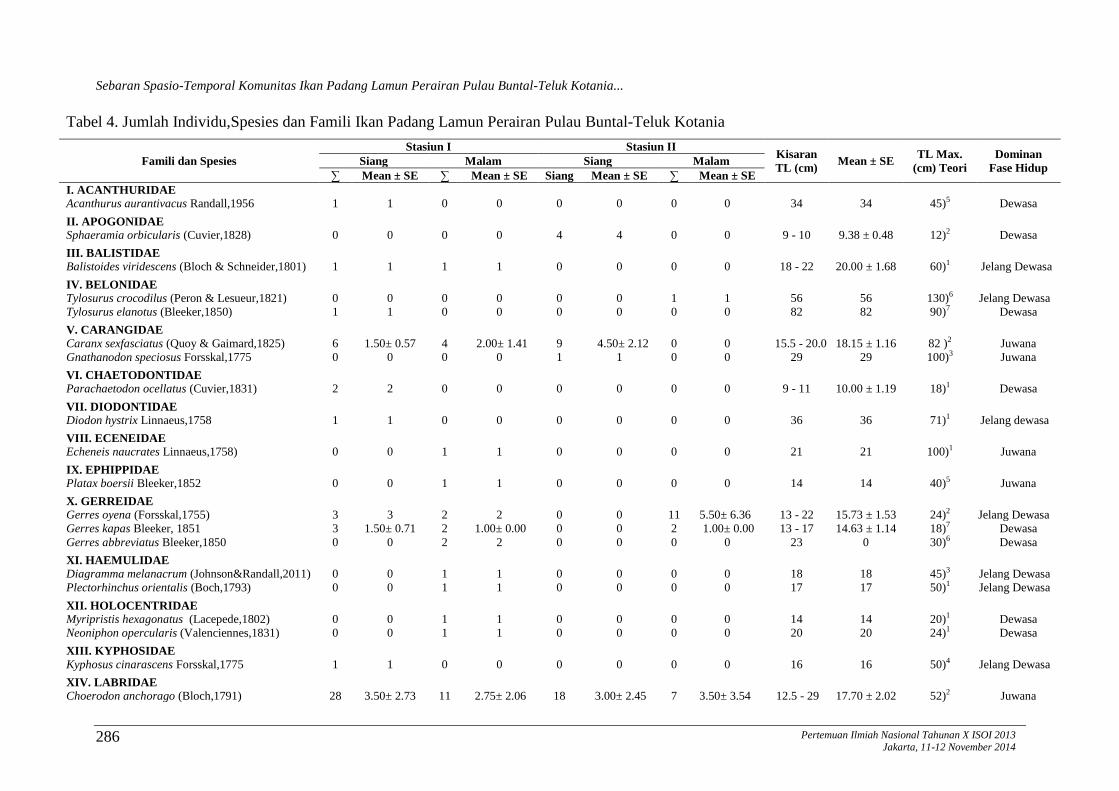
Tabel 4. Jumlah Individu,Spesies dan Famili Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania
Famili dan Spesies
Stasiun I Stasiun II Kisaran
TL (cm) Mean ± SE
TL Max.
(cm) Teori
Dominan
Fase Hidup Siang Malam Siang Malam
∑ Mean ± SE ∑ Mean ± SE Siang Mean ± SE ∑ Mean ± SE
I. ACANTHURIDAE
Acanthurus aurantivacus Randall,1956 1 1 0 0 0 0 0 0 34 34 45)5 Dewasa
II. APOGONIDAE
Sphaeramia orbicularis (Cuvier,1828) 0 0 0 0 4 4 0 0 9 - 10 9.38 ± 0.48 12)2 Dewasa
III. BALISTIDAE
Balistoides viridescens (Bloch & Schneider,1801) 1 1 1 1 0 0 0 0 18 - 22 20.00 ± 1.68 60)1 Jelang Dewasa
IV. BELONIDAE
Tylosurus crocodilus (Peron & Lesueur,1821) 0 0 0 0 0 0 1 1 56 56 130)6 Jelang Dewasa
Tylosurus elanotus (Bleeker,1850) 1 1 0 0 0 0 0 0 82 82 90)7 Dewasa
V. CARANGIDAE
Caranx sexfasciatus (Quoy & Gaimard,1825) 6 1.50± 0.57 4 2.00± 1.41 9 4.50± 2.12 0 0 15.5 - 20.0 18.15 ± 1.16 82 )2 Juwana
Gnathanodon speciosus Forsskal,1775 0 0 0 0 1 1 0 0 29 29 100)3 Juwana
VI. CHAETODONTIDAE
Parachaetodon ocellatus (Cuvier,1831) 2 2 0 0 0 0 0 0 9 - 11 10.00 ± 1.19 18)1 Dewasa
VII. DIODONTIDAE
Diodon hystrix Linnaeus,1758 1 1 0 0 0 0 0 0 36 36 71)1 Jelang dewasa
VIII. ECENEIDAE
Echeneis naucrates Linnaeus,1758) 0 0 1 1 0 0 0 0 21 21 100)1 Juwana
IX. EPHIPPIDAE
Platax boersii Bleeker,1852 0 0 1 1 0 0 0 0 14 14 40)5 Juwana
X. GERREIDAE
Gerres oyena (Forsskal,1755) 3 3 2 2 0 0 11 5.50± 6.36 13 - 22 15.73 ± 1.53 24)2 Jelang Dewasa
Gerres kapas Bleeker, 1851 3 1.50± 0.71 2 1.00± 0.00 0 0 2 1.00± 0.00 13 - 17 14.63 ± 1.14 18)7 Dewasa
Gerres abbreviatus Bleeker,1850 0 0 2 2 0 0 0 0 23 0 30)6 Dewasa
XI. HAEMULIDAE
Diagramma melanacrum (Johnson&Randall,2011) 0 0 1 1 0 0 0 0 18 18 45)3 Jelang Dewasa
Plectorhinchus orientalis (Boch,1793) 0 0 1 1 0 0 0 0 17 17 50)1 Jelang Dewasa
XII. HOLOCENTRIDAE
Myripristis hexagonatus (Lacepede,1802) 0 0 1 1 0 0 0 0 14 14 20)1 Dewasa
Neoniphon opercularis (Valenciennes,1831) 0 0 1 1 0 0 0 0 20 20 24)1 Dewasa
XIII. KYPHOSIDAE
Kyphosus cinarascens Forsskal,1775 1 1 0 0 0 0 0 0 16 16 50)4 Jelang Dewasa
XIV. LABRIDAE
Choerodon anchorago (Bloch,1791) 28 3.50± 2.73 11 2.75± 2.06 18 3.00± 2.45 7 3.50± 3.54 12.5 - 29 17.70 ± 2.02 52)2 Juwana

Latuconsina et al.
287 Pertemuan Ilmiah Nasional Tahunan X ISOI 2013
Jakarta, 11-12 November 2014
Tabel 4. Jumlah Individu,Spesies dan Famili Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania
Famili dan Spesies
Stasiun I Stasiun II Kisaran
TL (cm) Mean ± SE
TL Max.
(cm) Teori
Dominan
Fase Hidup Siang Malam Siang Malam
∑ Mean ± SE ∑ Mean ± SE Siang Mean ± SE ∑ Mean ± SE
Cheilinius chlorurus 4 1.33± 0.57 2 2 0 0 0 0 14 0 36)2 Juwana
Hemigymnus melapterus (Bloch,1791) 3 3 0 0 0 0 0 0 18 - 21 19.67 ± 1.24 90)1 Juwana
Halichoeres cloropterus (Bloch, 1791) 1 1 0 0 0 0 0 0 14 14 19)2 Dewasa
Cheilio inermis (Forsskal,1775) 1 1 2 2 0 0 0 0 30 - 32 31.33 ± 1.07 50)1 Jelang Dewasa
XV. LETHRINIDAE
Lethrinus lentjan (Lacepede,1802) 3 3 4 2.00± 1.41 0 0 0 0 16 - 20 18.33 ± 1.44 50)2 Jelang Dewasa
Lethrinus erythropterus Valenciennes,1830 10 3.33± 4.04 2 1.00± 0.00 1 1 1 1 14 - 21 17.71 ± 1.53 45)1 Jelang Dewasa
Lethrinus laticaudis (Alleyne & Macleay,1877) 0 0 0 0 1 1 0 0 17 0 80)1 Juwana
Lethrinus reticulatus Valenciennes, 1830 1 3.00± 1.58 1 1 13 3.25± 2.87 7 2.33± 1.53 14 - 22 19.03 ± 1.29 40)1 Jelang Dewasa
Psammoperca waigiensis (Cuvier, 1828) 0 0 1 1 0 0 0 0 25 0 47)1 Jelang Dewasa
XVI. LUTJANIDAE
Lutjanus carponotatus (Richardson,1842) 1 1 16 4.00 ± 5.36 6 3.00± 1.41 11 5.50 ± 2.95 18 - 26 21.00 ± 1.53 40)1 Jelang Dewasa
Lutjanus fulviflamma (Forsskal,1775) 2 1.00± 0.00 1 1 0 0 0 0 14 - 17 16.63 ± 1.26 35)2 Dewasa
Lutjanus decussatus (Cuvier,1828) 1 1 0 0 0 0 0 0 13 0 30)1 Jelang Dewasa
XVII. MONACANTHIDAE
Acriecthys tomentosus (Linaeus, 1758) 0 0 0 0 1 1 0 0 9 9 11.5)2 Dewasa
XVIII. MUGILIDAE
Mugil cephalus Linneaus,1758 1 1 5 5 0 0 0 0 9.5 - 23.0 18.58 ± 2.22 79)1 Juwana
XIX. MURAENIDAE
Gymnothorax pseudothyrsoideus (Bleeker,1852) 1 1 0 0 0 0 0 0 63 63 80)1 Dewasa
XX. MULLIDAE
Parupeneus indicus (Shaw,1903) 4 4 0 0 0 0 0 0 15 - 18 16.50 ± 1.46 35)2 Jelang Dewasa
Parupeneus barberinus (Lacepede,1801) 4 2.00± 0.00 0 0 0 0 0 0 15 - 17 16.00 ± 1.07 30)1 Jelang Dewasa
XXI. NEMIPTERIDAE
Scolopsis ciliatus (Lacepede,1802) 1 1 2 1.00± 0.00 0 0 0 0 14 - 17 15.50 ± 1.46 25)2 Jelang Dewasa
Pentapodus trivittatus (Bloch,1791) 9 1.28± 0.75 8 1.14± 0.37 2 2 5 1.25± 0.50 12 - 22 17.93 ± 1.71 28)2 Jelang Dewasa
Pentapodus setosus (Valenciennes,1830) 0 0 0 0 0 0 1 1 20 20 20)1 Dewasa
XXII. OSTRACIIDAE
Lactoria cornuta (Linnaeus, 1758) 0 0 1 1 0 0 0 0 9 9 46)2 Juwana
XXIII. PLOTOSIDAE
Plotosus anguilaris (Bloch,1794) 23 7.66 ± 10.69 0 0 19 19 2 2 19 - 26 21.07 ± 1.26 - -
XXIV. PLATYCEPHALIDAE
Papilloculiceps nematophthalmus (Gunther,1860) 2 2 0 0 0 0 2 1.00± 0.00 26 - 28 27.25 ± 0.98 58)1 Jelang Dewasa
XXV. POMACENTRIDAE
Dischistodus chrysopoecilus Schiegel&Muller,1839 0 0 0 0 1 1 0 0 15 15 15)7 Dewasa
(lanjutan....)

Pertemuan Ilmiah Nasional Tahunan X ISOI 2013 Jakarta, 11-12 November 2014
Sebaran Spasio-Temporal Komunitas Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania...
288
Tabel 4. Jumlah Individu,Spesies dan Famili Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania
Famili dan Spesies
Stasiun I Stasiun II Kisaran
TL (cm) Mean ± SE
TL Max.
(cm) Teori
Dominan
Fase Hidup Siang Malam Siang Malam
∑ Mean ± SE ∑ Mean ± SE Siang Mean ± SE ∑ Mean ± SE
Dichistodus prosopotaenia Bleeker,1852 3 1.50± 0.71 0 0 0 0 0 0 9 - 17 12.67 ± 2.01 17) Dewasa
Dischistodus perspicillatus Cuvier,1830 2 2 0 0 0 0 0 0 14 - 17 15.50 ± 1.46 18) Dewasa
Hemiglyphidodon plagiometopon (Bleeker,1852) 2 1.00± 0.00 0 0 0 0 0 0 12 - 17 14.50 ± 1.88 18) Dewasa
Pomacentrus sp 2 2 0 0 0 0 0 0 13 - 19 16.00 ± 2.06 - Dewasa
XXVI. SCARIDAE
Scarus gobban (Forsskal,1775) 4 2 ± 1.41 1 1 0 0 0 0 18 - 23 20.50 ± 1.54 75)2 Jelang Dewasa
Leptoscarus vaigiensis (Quoy & Gaimard, 1824) 1 1 0 0 0 0 0 0 13 13 35)1 Jelang Dewasa
Hyposcarus longiceps (Bleeker,1862) 0 0 0 0 0 0 2 2 16 16 45)1 Jelang Dewasa
XXVII. SCOMBRIDAE
Rastrelliger kanagurta Cuvier,1817 0 0 8 0 0 0 0 0 23 - 26 24.33 ± 1.17 35)7 Dewasa
XXVIII. SERRANIDAE
Epinephelus ongus (Bloch,1790) 1 1 0 0 2 2 0 0 16 - 19 17.50 ± 1.46 40)1 Jelang Dewasa
Epinephelus fuscoguttatus (Forsskal,1775) 1 1 0 0 0 0 0 0 29 29 90)1 Juwana
Epinephelus corallicola (Valenciennes,1828) 1 1 0 0 0 0 0 0 15 15 31)1 Jelang Dewasa
Cephalopholis microprion (Bleeker, 1852) 1 1 0 0 0 0 0 0 13 13 23)1 Jelang Dewasa
XXIX. SCORPAENIDAE
Synanceja horrida (Linnaeus,1766) 1 1 0 0 0 0 0 0 28 28 30)2 Dewasa
XXX. SIGANIDAE
Siganus canaliculatus (Park,1797) 341 34.10 ± 23.83 643 64.30 ± 30.43 495 49.50 ± 60.22 784 78.40±52.91 11 - 24 16.87 ± 1.40 29)2 Jelang Dewasa
Siganus doliatus (Cuvier,1830) 0 0 0 0 3 1.50 ± 0.84 0 0 9.5 - 18 14.17 ± 2.08 30)1 Jelang Dewasa
Siganus lineatus (Linnaeus,1835) 7 2.33 ± 1.15 6 1.20 ± 0.45 2 1.00± 0.00 9 2.25 ± 0.50 10 - 23 14.68 ± 1.94 43 )2 Jelang Dewasa
Siganus punctatus (Forster,1801) 1 1 2 1.00± 0.00 8 2.00± 1.41 3 1.50 ± 0.71 13 - 26 19.11 ± 2.08 40)1 Jelang Dewasa
XXXI. SPYRAENIDAE
Sphyraena pinguis Gunther, 1874 0 0 1 1 0 0 0 0 21 21 35)3 Dewasa
Sphyraena barracuda (Walbaum,1792) 1 1 2 1.00± 0.00 7 7 2 2 29 - 38 34.00 ± 1.67 180)1 Dewasa
XXXII. TERAPONTIDAE
Pelates quadrilineatus (Bloch,1790) 0 0 0 0 9 9 1 1 13 - 17 13.80 ± 1.15 20)1 Dewasa
XXXIII. TETRAODONTIDAE
Arothron hispidus (Linnaeus,1758) 0 0 0 0 0 0 1 1 53 53 51)1 Dewasa
Jumlah Individu 488 736 602 853
Jumlah Spesies 43 30 19 18
Jumlah Famili 23 16 13 13
Total Jumlah Individu Tiap Stasiun 1224 1454
Total Jumlah Spesies Tiap Stasiun 54 23
Total Jumlah Famili Tiap Satsiun 29 18 Sumber: )1 Allen (1999), )2 Allen & Erdmann (2012), )3Kuiter & Tonozuka Part 1, )4 Kuiter & Tonozuka Part 2, )5 Kuiter & Tonozuka Part 3, )6 Peristiwadi, )7Carpenter & Niem.
(lanjutan....)

Latuconsina et al.
289 Pertemuan Ilmiah Nasional Tahunan X ISOI 2013
Jakarta, 11-12 November 2014
Kelimpahan ikan yang lebih tinggi pada malam hari membuktikan adanya
distribusi harian ikan antara ekosistem padang lamun dan habitat terdekat seperti
mangrove dan terumbu karang (Nagelkerken et al. 2000 dan Pereira et al. (2010).
Banyak spesies ikan menggunakan lingkungan terumbu karang sebagai termpat
berlindung selama siang hari. Namun memiliki hubungan erat dan bergantung pada
ekosistem padang lamun dan habitat lainnya untuk mencari makan pada malam dan
sore hari (Pereira et al. 2010).Tingginya kelimpahan ikan dominan di daerah
penelitian mempengaruhi struktur komunitas ikan yang menyebabkan dominansi lebih
tinggi pada malam hari, dan keanekaragaman lebih tinggi pada siang hari. Menurut
Soegianto (1995), struktur suatu komunitas tidak hanya dipengaruhi oleh hubungan
antar spesies, tetapi juga oleh jumlah relatif organisme dari spesies-spesies tersebut,
sehingga kelimpahan suatu spesies dapat mempengaruhi fungsi suatu komunitas,
distribusi individu, dan memengaruhi keseimbangan stabilitas komunitas.
Berdasarkan kisaran panjang total, secara umum ikan yang ditemukan pada
ekosistem padang lamun selama penelitian termasuk dalam fase juwana dan jelang
dewasa. Fenomena ini membuktikan peran ekologis ekosistem padang lamun pulau
Buntal sebagai daerah asuhan dan pembesaran ikan-ikan komersial yang selalu
menjadi target penangkapan nelayan lokal setempat. Fenomena yang sama ditemukan
oleh Nagelkerken et al. (2000); Nagelkerken et al. (2002), Gell & Whittington (2002),
Arifin et al (2004), Marasabessy (2010), Unsworth et al. (2010), Latuconsina et al.
(2012 dan 2013), Latuconsina & Ambo-Rappe (2013), dan Ambo-Rappe et al (2013).
Fungsi potensial daerah penelitian sebagai daerah pembesaran terbukti dengan
ditemukannya spesies yang mendominasi seperti S.canaliculatus dengan kriteria
jelang dewasa dengan kelimpahan tinggi dibandingkan berukuran dewasa.
Latuconsina et al (2013) menemukan kelimpahan dan dominasi juwana
S.canaliculatus dengan ukuran panjang rata-rata 3,70 cm pada perairan Tanjung
Tiram selama periode pasang purnama dan pasang perbani. Sementara itu. Munira et
al (2010) juga menemukan sebagian besar S.canaliculatus yang hidup di padang
lamun Selat Lonthoir berukuran juwana dan umumnya dijumpai dalam satu kelompok
umur. Fenomena ini membuktikan peranan ekologi padang lamun di daerah penelitian
sebagai tempat pembesaran ikan-ikan ekonomis penting seperti famili Siganidae,
Lethrinidae, Lutjanidae, Serranidae, Carangidae, Scaridae, Labridae, Mullidae, dan
Nemipteridae. Sehingga keberadaan ekosistem padang lamun memberikan manfaat
ekonomi bagi masyarakat nelayan setempat.
Ditemukannya ikan-ikan penghuni padang lamun yang memiliki nilai ekonomis
penting yang sebagian besar berukuran juwana, membuktikan ekosistem tersebut
memiliki potensi ekonomi sebagai penyedia stok alamiah ikan ekonomis. Fenomena
yang sama dilaporkan Latuconsina & Ambo-Rappe (2013) pada perairan Tanjung
Tiram Teluk Ambon Dalam. Arifin et al. (2004) di perairan pulau Barrang Lompo-
Makassar menemukan ikan baronang (Siganus), kerapu (Epinephelus), kakap
(Lutjanus), dan ikan lencam (Lethrinus), sehingga keberadaan ekosistem padang
lamun berperan sebagai penyedia stok dan penentu keberadaan ikan-ikan berukuran
juwana.
3.5. Struktur Komunitas Ikan
Berdasarkan Gambar 2, memperlihatkan adanya fluktuasi struktur komunitas
ikan antara periode siang dan malam hari, dimana stasiun I dan II indeks dominansi
lebih rendah pada siang hari dan tinggi pada malam hari, sedangkan indeks
kenakeragaman lebih tinggi pada siang hari dan rendah pada malam hari. Meskipun
demikian nilai rata-rata indeks dominansi pada malam hari di stasiun I dan II masuk

Pertemuan Ilmiah Nasional Tahunan X ISOI 2013 Jakarta, 11-12 November 2014
Sebaran Spasio-Temporal Komunitas Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania...
290
kategori tinggi dan masuk kategori rendah pada siang hari. Sementara Indeks
keanekaragaman pada stasiun I dan II di siang maupun malam hari masuk kategori
rendah yang menunjukkan secara ekologi tidak terdpat spesies yang sangat
mendominasi dalam pemanfaatan ruang dan sumberdaya yang ada pada ekosistem
padang lamun di lokasi penelitian.
Gambar 2. Struktur Komunitas Ikan padang lamun Perairan Pulau Buntal-Teluk
Kotania.
Menurut Hemingga & Duarte (2000), terdapat empat faktor yang relevan
terkait dengan variabilitas komunitas ikan padang lamun , yaitu : (1) struktur vegetasi
lamun, (2) tingkat larva dan ikan juvenil yang menghuni padang lamun, mortalitas
dan proses migrasi, (3) lokasi vegetasi lamun terhadap habitat lainnya, dan (4)
parameter fisika kimia pada habitat lamun.
Hasil uji komparatif (Uji t) keanekaragaman spesies ikan padang lamun antara
siang dan malam pada stasiun I mendapatkan thitung > ttabel (4,396 >2,101), sedangkan
pada stasiun II didapatkan nilai thitung > ttabel (2,178 >2,101) artinya terdapat perbedaan
keanakeragaman komunitas ikan antara siang dan malam hari dengan keragaman
tertinggi pada siang hari. Sementara perbandingan keanekaragaman antar stasiun
pengamatan mendapatkan nilai thitung > ttabel (2,244 >2,024) artinya terdapat perbedaan
keragaman komunitas ikan antara stasiun I dan II dengan keragaman tertinggi
ditmukan pada stasiun I. Menurut Brower et al (1990), karagaman jenis adalah suatu
ekspresi dari struktur komunitas, dimana suatu komunitas dikatakan memiliki
0,38
0,68
0,47
0,64 0,68
0,9
0,51
0,8
0,66 0,64 0,62 0,62
0,18
0,91
0,28
0,82
0,44
0,81
0,46
0,9
1,38
0,74
1,04 0,95
0,76
0,27
1,07
0,44
0,87 0,95
0,79 0,79
1,89
0,22
1,8
0,45
1,41
0,46
1,3
0,25
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
Stru
ktu
r ko
mu
nit
as
Stasiun I Dominansi Keragaman
0,37
0,52 0,57
0,77
0,22
0,94
0,6
0,94
0,7
0,9 0,98
0,57
0,81
0,93
0,54
0,85
0,65
0,91
0,76
0,91
1,04
0,92
0,63 0,54
1,63
0,17
0,72
0,13
0,78
0,26
0,1
0,97
0,4
0,17
1,04
0,37
0,72
0,23
0,49
0,19
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
Stru
ktu
r K
om
un
itas
Stasiun II
Dominansi Keragaman

Latuconsina et al.
291 Pertemuan Ilmiah Nasional Tahunan X ISOI 2013
Jakarta, 11-12 November 2014
karagaman jenis tinggi, jika proporsi antar jenis secarakeseluruhan sama banyak,
sehingga jika ada beberapa jenis dalamkomunitas memiliki kelimpahan (dominansi
yang besar) makakeanekaragaman dan keseragamannya rendah.
Hasil uji komparatif (Uji t) dominansi spesies ikan padang lamun antara siang
dan malam pada stasiun I mendapatkan -thitung < -ttabel (-4,831 < -2,101), sedangkan
pada stasiun II didapatkan nilai -thitung < -ttabel (-2,412 < -2,101) artinya terdapat
perbedaan dominansi komunitas ikan antara siang dan malam hari dengan dominansi
tertinggi pada malam hari. Sementara perbandingan dominansi komunitas ikan
mendapatkan nilai -thitung > -ttabel (-1,539 > -2,024) yang menenujukkan tidak terdapat
perbedaan dominansi komunitas ikan antara stasiun I dan II. Menurut Soegianto
(1995), struktur suatu komunitas tidak hanya dipengaruhi oleh hubungan antar
sepsies, tetapi juga jumlah relatif organisme dari spesies-sepsies tersebut, sehingga
kelimpahan relatif suatu spesies mempengaruhi fungsi suatu komunitas, distribusi
individu dalam komunitas, dan dapat mempengaruhi keseimbangan komunitas dan
akhirnya pada stabilitas komunitas tersebut.
3.6. Kesamaan Spesies Ikan
Indeks kesamaan jenis Bray – Curtis secara spasial antar stasiun pengamatan
dengan karaktersitik fisik habitat yang berbeda dan secara temporal pada periode
siang dan malam hari memperlihatkan adanya perbedaan (Gambar 3).
Gambar 3. Dendrogram Pengelompokan Kesamaan Spesies Ikan Padang Lamun
Berdasarkan Kehadirannya Antar Stasiun Pengamatan di Lokasi Penelitian Antara
Siang dan Malam Hari
Terdapat pengelompokan spesies ikan berdasarkan kehadirannya secara
spasial maupun temporal. Kesamaan spesies tertinggi ditemukan pada stasiun II
antara siang dan malam hari sebesar 59.46 %. Sementara kesamaan spesies antara
stasiun I dan II sebesar 38,71 %. Fenomena ini menunjukkan bahwa kesamaan spesies
secara spasial antar stasiun pengamatan maupun secara temporal antara siang dan
malam hari tidak terlalu tinggi. Kesamaan spesies ikan tertinggi antara siang dan
malam pada Stasiun II diduga berkaitan dengan kedekatan ekosistem padang lamun
hanya dengan ekosistem mangrove, sementara pada stasiun I kesamaan spesies
sebesar 54,05 %. Hal ini diduga berkaitan dengan spesies-spesies ikan yang
mendiamai mangrove dan terumbu karang yang bersifat diurnal maupun noktrunal
Sia
ng (S
tasiu
n II)
Mala
m (S
tasiu
n II)
Sia
ng (S
tasiu
n I)
Mala
m (S
tasiu
n I)100
80
60
40
Sim
ilari
ty
Kesaman Spesies Ikan padang lamun

Pertemuan Ilmiah Nasional Tahunan X ISOI 2013 Jakarta, 11-12 November 2014
Sebaran Spasio-Temporal Komunitas Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania...
292
yang terdistribusi mengikuti ritme pasang surut baik untuk mencari makan maupun
aktivitas biologis lainnya.
Spesies ikan yang ditemukan melimpah pada ekosistem padang lamun dan
selalu tersebar pada ekosistem mangrove adalah Caranx sexfasciatus (Carangidae),
Plotosus anguilaris (Plotosidae), Gerres oyena dan Gerres kapas (Gerreidae),
Sphyraena pinguis dan Sphyraena barracuda (Spyraenidae), Synanceja horrida
(Scorpaenidae), dan Siganus canaliculatus (Siganidae), sedangkan spesies ikan yang
berasosiasi dengan terumbu karang seperti :Parachaetodon ocellatus
(Cahetodontidae), Scarus ghobban (Scaridae),dan ikan dari famili Serranidae dan
Holocentridae membuktikan bahwa tingginya kelimpahan ikan pada eksosistem
padang lamun perairan pulau Buntak-Teluk Kotania turut dipengaruhi oleh
kedekatannya dengan ekosistem mangrove dan terumbu karang. Fenomena
keterkaitan komunitas ikan padang lamun dengan ekosistem mangrove diperkuat
temuan Unsworth et al. (2009) terkait kontribusi ekosistem mangrove sebagai habitat
ikan karena terkait ruaya pasang, di mana pasang tertinggi mendukung kelimpahan
ikan yang lebih besar dari habitat lamun dan saat surut akan terdistribusi pada
ekosistem padang lamun. Menurut Verweij et al. (2006), ikan-ikan dari famili
Carangidae dan Sphyraenidae lebih cenderung menggunakan struktur fisik lamun
untuk meningkatkan efisiensi pemangsaan daripada sebagai tempat perlindungan.
Fenomena ini membuktikan bahwa ekosistem padang lamun perairan pulau Buntal
dijadikan sebagai areal mencari makan bagi ikan-ikan mangrove yang memanfaatkan
mekanisme pasang. Dengan demikian ada konektivitas yang tinggi antara ekosistem
mangrove, padang lamun, dan terumbu karang terkait distribusi spasio-temporal
komunitas ikan. Menurut Adrim (2006), keragaman spesies ikan pada eksosistem
padang lamun sangat didukung oleh konektivitas ekosistem lamun dengan ekosistem
di sekitarnya seperti mangrove dan terumbu karang.
Adanya konektivitas dan pentingnya fungsi ekosistem padang lamun sebagai
tempat asuhan dan pembesaran ikan-ikan khas mangrove dan terumbu karang,
dibuktikan oleh Nakamura (2010) yang menemukan hilangnya padang lamun di
selatan kepulauan Ryukyu berdampak negatif terhadap penurunan jumlah spesies
ikan komersial penghuni terumbu karang yang memanfaatkan padang lamun sebagai
tempat pembesaran. Nagelkerken et al(2002) juga membuktikan bahwa kelimpahan
ikan di terumbu karang merupakan fungsi keberadaan mangrove dan padang lamun
sebagai areal asuhan dan pembesaran ikan. Degradasi habitat mangrove dan padang
lamun dapat menimbulkan dampak signifikan pada persediaan stok ikan karang di
Karibia. Hal yang sama dikemukakan Chittaro et al. (2005) yang menemukan
vegetasi mangrove dan lamun memberikan fungsi yang lebih besar bagi komunitas
ikan sebagai daerah asuhan dan pembesaran dibandingkan dengan terumbu karang.
IV. KESIMPULAN
1. Jumlah spesies dan famili ikan tertinggi ditemukan pada habitat lamun yang
diapit mangrove dan terumbu karang, sedangkan kelimpahan ikan tertinggi pada
habitat lamun yang hanya berdekatan dengan mangrove.
2. Terdapat perbedaan signifikan kenekaragaman komunitas ikan, secara temporal
lebih tinggi pada siang hari dan secara spasial lebih tinggi pada habitat lamun
yang diapit mangrove dan terumbu karang. Sementara itu terdapat perbedaan
indeks dominansi, secara temporal lebih tinggi pada malam hari, dan secara
spasial lebih tinggi pada habitat lamun yang hanya diapit mangrove namun tidak
berbeda signifikan antar habitat lamun

Latuconsina et al.
293 Pertemuan Ilmiah Nasional Tahunan X ISOI 2013
Jakarta, 11-12 November 2014
3. Perbedaan struktur komunitas ikan padang lamun secara spasial menunjukkan
keberadaan ikan pada habitat padang lamun sangat dipengaruhi kedekatannya
dengan ekosistem disekitarnya, dan perbedaan struktur komunitas ikan secara
temporal menunjukkan adanya sifat nokturnal maupun diurnal ikan yang
mempengaruhi orientasi ikan yang berbeda pada habitat lamun.
DAFTAR PUSTAKA
Adrim M. 2006. Asosiasi ikan di padang lamun. Oseana. 31(4):1-7.
Allen, G. 1999. Marine Fishes of South-East Asia; A Guide For Anglers And Divers.
Periplus edition. Singapore. 292 pp.
Allen GR & Erdmann MV. 2012. Reef fishes of the East Indies. Volume I-III.
Tropical Reef Research, Perth, Australia. 1292 pp.
Ambo-Rappe, R. 2010. Sturktur Komunitas Ikan padang Lamun yang berbeda di
Pulau Barrang Lompo. Jurnal Ilmu dan Teknologi Kelautan Tropis, 2(2):62-
73.
Ambo-Rappe, R., M.N.Nessa., H. Latuconsina & D.L. Lajus. 2013. Relationship
between the tropical seagrass bedcharacteristics and the structure of the
associated fish community. Open Journal of Ecology. Vol.3(5):331-342.
Arifin, La Nafie YA, &Supriadi. 2004. Studi kondisi dan potensi ekosistem padang
lamun sebagai daerah asuhan berbagai jenis biota laut di perairan pulau
Barrang Lompo, Makassar. Torani, 14(5): 241-250.
Bell, J.D., & Pollard. 1989. Ecologi Of Fish Assemblages And Fisheries Associated
With Seagres. In Lerkum et al, (eds). Biologi of siagress. Aquatic Plant Studies
2. Elsevier Science Pub.B.V. Amsterdam. pp.565-609.
Carpenter K.E & Niem V.H (eds). 1999. The living marine resources of the Western
Central Pacific. Bony fishes part 2 (Mugilidae to Carangidae. FAO species
identification.Vol.4. Rome. pp. 2069-2790.
Carpenter K.E & Niem V.H (eds). 2001. The living marine resources of the Western
Central Pacific. (Labridae to Latimeriidae), estuarine crocodiles, sea turtles,
sea snakes and marine mammals. FAO species identification.Vol.6. Bony
fishes part 4. Rome. pp. 3381-4218.
Chittaro, P.M., P. Usseglio, &P.F., Sale, 2005. Variation in fish density, assemblage
composition and relative rates of predation among mangrove, seagrass and
coral reef habitats. Environmental Biology of Fishes, 72: 175–187.
Effendi, H. 2003. Telaah kualitas air bagi pengelolaan sumberdaya dan lingkungan
perairan. Kanisius. Yogyakarta. 112 pp.
Floeter, S.R, Behrens MD, Ferreira CEL, Paddack MJ, Horn MH. 2005.
Geographical gradients of marine herbivorous fishes: patterns and processes.
Marine Biology, 146: 1435-1447.
Gell FR & Whittington MW. 2002. Diversity of fishes in seagrass beds in the
Quirimba Archipelago, Northern Mozambique. Marine and Freshwater
Research 53: 115-121.
Hemingga, AM. & CM Duerte. 2000.Seagrass Ecology. Candbridge University Press.
New York. 322 pp.

Pertemuan Ilmiah Nasional Tahunan X ISOI 2013 Jakarta, 11-12 November 2014
Sebaran Spasio-Temporal Komunitas Ikan Padang Lamun Perairan Pulau Buntal-Teluk Kotania...
294
Kordi M.G.H &A.B., Tancung, 2007. Pengelolaan kualitas air dalam budidaya
perairan. Rineka Cipta. Jakarta. 208 pp.
Kuiter, R.H. & T. Tonozuka. 2001. Indonesia Reef Fishes. Psrt 3. Jawfishes-
Sunfishes. Zoonetic, Melbourne. Australia. 123 pp.
Kuiter, R.H. & T. Tonozuka. 2001.Indonesia Reef Fishes. Part 2. Fusiliert to
Dragonets.: Caesionedea to Callyonimidea. Zoonetic, Melbourne. Australia.
161 pp.
Laevastu T & Hayes M. 1982. Fisheries oceanography and ecology. Fishing News
Book, Ltd. Farnham. Surrey. England. 199 pp.
Lam, TJ. 1974. Siganids; their biology and mariculture potential. Aquaculture, 3: 325-
354.
Latuconsina H, M.N., Nessa & R. Ambo-Rappe,2012. Komposisi spesies dan struktur
komunitas ikan padang lamun perairan Tanjung Tiram-Teluk Ambon Dalam.
Ilmu&Teknologi Kelautan Tropis4(1): 35-46.
Latuconsina H, R. Ambo-Rappe, &M.N. Nessa, 2013. Asosiasi ikan baronang
(Siganus canaliculatus Park, 1797) pada ekosistem padang lamun perairan
Teluk Ambon Dalam. In: Simanjuntak CPH (eds.). Prosiding Seminar
Nasional Ikan VII. Masyarakat Iktiologi Indonesia. pp. 123-137.
Latuconsina, H dan R. Ambo-Rappe. 2013. Variabilitas Harian Komunitas Ikan
padang Lamun Perairan Tanjung Tiram-Teluk Ambon Dalam.Jurnal Iktiologi
Indonesia. Vol.13(1):35-53.
Marasabessy, M.D. 2010. Sumber daya ikan di perairan padang lamun pulau-pulau
Derawan Kalimantan Timur. Oseanologi dan Limnologi Indonesia 36(2): 193-
210.
Munira, Sulistiono, &Zairion. 2010. Hubungan panjang-bobot dan pertumbuhan ikan
beronang, Siganus canaliculatus (Park, 1797) di padang lamun Selat Lonthoir,
Kepulauan Banda, Maluku. Jurnal Iktiologi Indonesia. 10(2): 153-163.
Nagelkerken I, G. van der Velde, G.W. Gorissen,G.J. Meijer, T. van’t Hof, C.den
Hartog. 2000. Importance of mangroves, seagrass beds and the shallow coral
reef as nursery for importance reef fishes, using a visual cencus technique.
Estuarine, Coastal and Shelf Science 51:31-44.
Nagelkerken I, C.M. Roberts, G. van der Velde,M. Dorenbosch, M.C. van Riel,
Cocheret de la Morinière E,& P.H. Nienhuis, 2002. How important are
mangroves and seagrass beds for coral-reef fish? The nursery hypothesis tested
on an island scale. Marine Ecology Progress Series, 244: 299–305.
Nakamura Y. 2010. Patterns in fish response to seagrass beds loss at the southern
Ryukyu Island, Japan. Marine Biology, 157: 2397-2406.
Odum, E.P.. 1983. Basic Ecology. Saunders College Publishing, New York. 612 pp.
Pereira PHC, B.P., Ferreira, & S.M., Rezende. 2010. Community structure of the
ichthyofauna associated with seagrass beds (Halodule wrightii) in Formoso
River estuary – Pernambuco, Brazil. Anais da Academia Brasileira de
Ciências, 82(3): 617-628.
Peristiwady, 2006. Ikan-ikan Laut Ekonomis Penting di Indonesia. LIPI Press.
Jakarta.

Latuconsina et al.
295 Pertemuan Ilmiah Nasional Tahunan X ISOI 2013
Jakarta, 11-12 November 2014
Priyatno, D. 2009. 5 jam belajar olah data SPSS 17. Penerbit Andi. Yogyakarta.
Romimohtarto, K. dan S. Juwana. 2004. Meroplankton Laut: Larva laut yang menjadi
plankton. Djambatan. Jakarta. 214 pp.
Pereira PHC, Ferreira BP, Rezende SM. 2010. Community structure of the
ichthyofauna associated with seagrass beds (Halodule wrightii) in Formoso
River estuary – Pernambuco, Brazil. Anais da Academia Brasileira de
Ciências, 82(3): 617-628.
Supriadi., Y.A.La Nafie dan A.I. Burhanuddin. 2004. Inventarisasi Jenis, Kelimpahan
dan Biomas Ikan di Padang Lamun Pulau Barranglompo Makassar.
Torani,Vol.14 (5):288-295.
Unsworth, 2007. Fish assemblages in seagress beds are infulenced by the proximity of
mangrove forests. Marine Biology.150:993-1002.
Unsworth RFK, Garrard SL, de León PS, Cullen LC, Smith DJ, SlomanKA, &Bell JJ.
2009. Structuring of Indo-Pacific fish assemblages along the mangrove–
seagrass continuum. Aquatic Biology, 5: 85–95.
Unsworth RKF, Cullen LC, Pretty JN, Smith DJ, &Bell JJ. 2010. Economic and
subsistence values of the standing stocks of seagrass fisheries: Potential
benefits of no-fishing marine protected area management. Ocean and Coastal
Management 30: 1-7.
Verweij MC, Nagelkerken I, de Graaff D, Peeters M, Bakker EJ, &van der Velde G.
2006. Structure, food and shade attract juvenile coral reef fish to mangrove
and seagrass habitats: a field experiment. Marine Ecology Progress Series,
306: 257–268.
Copyright © 2022 FDOKUMEN




















