
Seasoned adaptive antibody immunity for highly pathogenic pandemic influenza in humans
10
REVIEW Seasoned adaptive antibody immunity for highly pathogenic pandemic influenza in humans Garry W Lynch 1,2,3 , Paul Selleck 4 , W Bret Church 5 and John S Sullivan 1,6 Fundamentally new approaches are required for the development of vaccines to pre-empt and protect against emerging and pandemic influenzas. Current strategies involve post-emergent homotypic vaccines that are modelled upon select circulating ‘seasonal’ influenzas, but cannot induce cross-strain protection against newly evolved or zoonotically introduced highly pathogenic influenza (HPI). Avian H5N1 and the less-lethal 2009 H1N1 and their reassortants loom as candidates to seed a future HPI pandemic. Therefore, more universal ‘seasoned’ vaccine approaches are urgently needed for heterotypic protection ahead of time. Pivotal to this is the need to understand mechanisms that can deliver broad strain protection. Heterotypic and heterosubtypic humoral immunities have largely been overlooked for influenza cross-protection, with most ‘seasoned’ vaccine efforts for humans focussed on heterotypic cellular immunity. However, 5 years ago we began to identify direct and indirect indicators of humoral-herd immunity to protein sites preserved among H1N1, H3N2 and H5N1 influenzas. Since then the evidence for cross-protective antibodies in humans has been accumulating. Now proposed is a rationale to stimulate and enhance pre-existing heterotypic humoral responses that, together with cell-mediated initiatives, will deliver pre-emptive and universal human protection against emerging epidemic and pandemic influenzas. Immunology and Cell Biology (2012) 90, 149–158; doi:10.1038/icb.2011.38; published online 7 June 2011 Keywords: antibody; H5N1; heterosubtypic; influenza; pandemic; vaccine The rapid global circulation of ‘swine flu’ in 2009 delivered an emphatic wake-up call to the world. That North American H1N1 hybrid of porcine, avian and human-like influenza signatures had, it appears, been brewing in the background for some time. But within days of bursting onto the international radar, it had crossed from Mexico to the US and quickly distributed itself across the globe, warranting a rapid escalation to World Health Organization (WHO) pre-pandemic level-5 alert classification and ultimately full Pandemic level-6 status as the first pandemic of the twenty-first century (http:\\www.who.int/csr/disease/avian_ influenza/phase/en/). 1–3 A hallmark of that new virus was its high human-to-human transmissibility. Fortuitously however, the 2009 H1N1 has revealed itself to be a mild pathogen. But if it were to become as lethal as the currently circulating avian H5N1 influenza, that has an overall mortality of B60% among humans, we would have a highly pathogenic influenza (HPI) pandemic of unprecedented proportions. That the two strains might mix to give more human adaptive reassortments, either together or with other circulating subtypes, remains a distinct possibility, especially with the spread of H1N1 within regions endemic for H5N1. If both the types co-infect inter- mediate hosts with receptors for both avian and human/pig-like influenzas, such as chickens and pigs, new, human-transmissible and highly lethal hybrid strains could emerge. In a study assessing the conservation of human specificity and mortality, 34 biomarkers were identified to be conserved among past pandemic influenzas, in most (that is, 9) of the influenza proteins. 4 Surprisingly, 25 of the 34 biomarkers detected were among the non-surface influenza proteins, NP, PA, PB1, PB2, M1, NS1, NS2, and only 3 and 6 in the envelope HA and NA proteins, respectively. In demonstration of the variety of mixed strains that can result from co-infection, particularly sobering are studies that reveal important molecular changes that govern influenza transmission in exhaled respiratory droplets. 5 Revealing, in a ferret model of human infection (Supplementary Information S1), the relative ease with which following co-infection of highly patho- genic avian influenza (that is, HPAI) H9N2 with human H3N2, the H9N2 was able to acquire the outside shell proteins of the H3N2 strain, while maintaining its complement of internal virion (intra- Received 11 November 2009; revised 31 March 2011; accepted 4 April 2011; published online 7 June 2011 1 Biosafety, Immunobiology, Global Health and Pandemic Infections Research, Central Clinical School, Faculties of Medicine, The University of Sydney, Camperdown, New South Wales, Australia; 2 Veterinary Science, University of Sydney, Camperdown, New South Wales, Australia; 3 Cellular & Molecular Pathology Research Unit, Oral Pathology and Oral Medicine, Westmead Centre for Oral Health, Westmead Hospital and Faculty of Dentistry, University of Sydney, Westmead, New South Wales, Australia; 4 The Australian Animal Health Laboratories, CSIRO, Geelong, Victoria, Australia; 5 Group in Biomolecular Structure and Informatics, Pharmaceutical Chemistry, Faculty of Pharmacy, University of Sydney, Camperdown, New South Wales, Australia and 6 Children’s Cancer Institute Australia for Medical Research, Randwick, New South Wales, Australia Correspondence: Dr GW Lynch, Veterinary Science, University of Sydney, Room 551, Gunn Building, Camperdown, New South Wales 2006, Australia. E-mail: [email protected] Immunology and Cell Biology (2012) 90, 149–158 & 2012 Australasian Society for Immunology Inc. All rights reserved 0818-9641/12 www.nature.com/icb
Transcript of Seasoned adaptive antibody immunity for highly pathogenic pandemic influenza in humans

REVIEW
Seasoned adaptive antibody immunity for highlypathogenic pandemic influenza in humans
Garry W Lynch1,2,3, Paul Selleck4, W Bret Church5 and John S Sullivan1,6
Fundamentally new approaches are required for the development of vaccines to pre-empt and protect against emerging and
pandemic influenzas. Current strategies involve post-emergent homotypic vaccines that are modelled upon select circulating
‘seasonal’ influenzas, but cannot induce cross-strain protection against newly evolved or zoonotically introduced highly
pathogenic influenza (HPI). Avian H5N1 and the less-lethal 2009 H1N1 and their reassortants loom as candidates to seed a
future HPI pandemic. Therefore, more universal ‘seasoned’ vaccine approaches are urgently needed for heterotypic protection
ahead of time. Pivotal to this is the need to understand mechanisms that can deliver broad strain protection. Heterotypic and
heterosubtypic humoral immunities have largely been overlooked for influenza cross-protection, with most ‘seasoned’ vaccine
efforts for humans focussed on heterotypic cellular immunity. However, 5 years ago we began to identify direct and indirect
indicators of humoral-herd immunity to protein sites preserved among H1N1, H3N2 and H5N1 influenzas. Since then the
evidence for cross-protective antibodies in humans has been accumulating. Now proposed is a rationale to stimulate and
enhance pre-existing heterotypic humoral responses that, together with cell-mediated initiatives, will deliver pre-emptive
and universal human protection against emerging epidemic and pandemic influenzas.
Immunology and Cell Biology (2012) 90, 149–158; doi:10.1038/icb.2011.38; published online 7 June 2011
Keywords: antibody; H5N1; heterosubtypic; influenza; pandemic; vaccine
The rapid global circulation of ‘swine flu’ in 2009 delivered anemphatic wake-up call to the world. That North American H1N1hybrid of porcine, avian and human-like influenza signatures had,it appears, been brewing in the background for some time. Butwithin days of bursting onto the international radar, it hadcrossed from Mexico to the US and quickly distributed itselfacross the globe, warranting a rapid escalation to World HealthOrganization (WHO) pre-pandemic level-5 alert classificationand ultimately full Pandemic level-6 status as the first pandemicof the twenty-first century (http:\\www.who.int/csr/disease/avian_influenza/phase/en/).1–3 A hallmark of that new virus was its highhuman-to-human transmissibility. Fortuitously however, the 2009H1N1 has revealed itself to be a mild pathogen. But if it were tobecome as lethal as the currently circulating avian H5N1 influenza,that has an overall mortality of B60% among humans, we would havea highly pathogenic influenza (HPI) pandemic of unprecedentedproportions.
That the two strains might mix to give more human adaptivereassortments, either together or with other circulating subtypes,
remains a distinct possibility, especially with the spread of H1N1within regions endemic for H5N1. If both the types co-infect inter-mediate hosts with receptors for both avian and human/pig-likeinfluenzas, such as chickens and pigs, new, human-transmissible andhighly lethal hybrid strains could emerge. In a study assessing theconservation of human specificity and mortality, 34 biomarkers wereidentified to be conserved among past pandemic influenzas, in most(that is, 9) of the influenza proteins.4 Surprisingly, 25 of the 34biomarkers detected were among the non-surface influenza proteins,NP, PA, PB1, PB2, M1, NS1, NS2, and only 3 and 6 in the envelopeHA and NA proteins, respectively. In demonstration of the variety ofmixed strains that can result from co-infection, particularly soberingare studies that reveal important molecular changes that governinfluenza transmission in exhaled respiratory droplets.5 Revealing, ina ferret model of human infection (Supplementary Information S1),the relative ease with which following co-infection of highly patho-genic avian influenza (that is, HPAI) H9N2 with human H3N2, theH9N2 was able to acquire the outside shell proteins of the H3N2strain, while maintaining its complement of internal virion (intra-
Received 11 November 2009; revised 31 March 2011; accepted 4 April 2011; published online 7 June 2011
1Biosafety, Immunobiology, Global Health and Pandemic Infections Research, Central Clinical School, Faculties of Medicine, The University of Sydney, Camperdown, New SouthWales, Australia; 2Veterinary Science, University of Sydney, Camperdown, New South Wales, Australia; 3Cellular & Molecular Pathology Research Unit, Oral Pathology and OralMedicine, Westmead Centre for Oral Health, Westmead Hospital and Faculty of Dentistry, University of Sydney, Westmead, New South Wales, Australia; 4The Australian AnimalHealth Laboratories, CSIRO, Geelong, Victoria, Australia; 5Group in Biomolecular Structure and Informatics, Pharmaceutical Chemistry, Faculty of Pharmacy, University of Sydney,Camperdown, New South Wales, Australia and 6Children’s Cancer Institute Australia for Medical Research, Randwick, New South Wales, AustraliaCorrespondence: Dr GW Lynch, Veterinary Science, University of Sydney, Room 551, Gunn Building, Camperdown, New South Wales 2006, Australia.E-mail: [email protected]
Immunology and Cell Biology (2012) 90, 149–158& 2012 Australasian Society for Immunology Inc. All rights reserved 0818-9641/12
www.nature.com/icb

virion) proteins, and thereby achieve the aerosol transmissibility of thehuman strain.5 Similarly, a genetic ‘shift’ that combines avian H5N1and porcine H1N1 may enable the swift evolution of a deadly aero-solized H5N1 hybrid capable of rapid spread. The potential for viablelethal reassortants of H5N1 with H3N2 co-infection has also beenshown in the laboratory, with the generation of highly transmissible6
and virulent7 H5N1/H3N2 reassortants. Thus, various lethal hybridcombinations might assort in the future, despite earlier studies arguingagainst such a possibility.8 Consequently, pertinent questions remain,as to where and when this might occur, and the possible degree oftransmissibility or pathogenicity that might be expected in such anoutbreak.
Media-generated alarm is not warranted, of course, but equally wecannot afford not to be prepared for the worst-case scenario. Detailedpredictive analyses have forecast that a mild influenza pandemic wouldcost 1.4 million lives and US$330 billion, and a severe pandemic 142.2million lives and US$4.4 trillion.8 This modelling was based on the‘mild’ 1968–69 pandemic, the ‘moderate’ 1957 pandemic, and the‘severe’ to ‘ultra severe’ pandemic of 1918–19. None of the influenzastrains causing pandemics in the twentieth century were as lethal asthe currently circulating H5N1 strains that have a mortality rate morethan an order greater than any previous human-infecting HPI, raisingconcerns that a rapidly circulating new strain of similar lethality couldmathematically bring about a billion deaths and huge economic losseson a global scale.
Irrespective of whether caused by an avian-derived H5N1 or astrain phylogenetically related to H1N1 or H3N2 influenzas thatreadily transmit among humans, or by other avian-derived strains(for example, H7N7, H7N2, H9N2 (Supplementary Information S1)),or other novel re-assortment combinations, it was generally agreedthat another influenza pandemic was overdue. With past frequenciesof 11- to 40-year cycles between pandemics uncannily after 41 years,a HINI pandemic was delivered. Pandemic influenzas developover a period while they acquire mutations and traits for humaninfection leading ultimately to an explosive emergence, and thisraises pressing questions of what our pre-existing defences are andhow we can provide or stimulate adequate protection for populationsat large.
CURRENT ANTI-VIRAL APPROACHES AND THEIR
LIMITATIONS FOR PANDEMIC INFLUENZA
Our proven best defence against influenza is vaccination. Against mildseasonal epidemics, vaccines are designed to stimulate the immunesystem to target highly immunogenic epitopes of the predominantcirculating influenzas. These ‘seasonal’ vaccines all work by developinghigh-titre, neutralizing antibodies to the major strain-specific, immuno-dominant antigen (Ag) determinants in the vaccine.9 Characteristicallythese antibodies target various highly antigenic determinants of hemag-glutinin’s (HA) globular head and neutralize infection by blocking ordisrupting cell receptor binding. This is schematically represented inFigure 1 to show separate immunodominant hot spots in the globularhead of HAs (that is, marked by translucent blue patches).10–14 A highrate of antigenic drift (mutation) of this domain of HA generatesvaried primary sequences and secondary structural conformations thatalter the antigenic determinants that define influenza strains. Alsoschematically illustrated in Figure 1 revealing the structural proteomeof influenza, is the dominance of antibody responses to these Ags ofthe HA globular head, compared with Ags expressed by the otherenvelope proteins NA and M2, and even to sites on the stalk of HAitself. For effect this has been represented by size and colour differ-ences. Despite being shadowed by competing HA Ags, major antigenic
determinants in the globular-head domain of neuraminidase (NA)of the intact virus or representative vaccines can additionallypromote non-neutralizing antibodies that protect against influenzaby interfering with the release of progeny virus from infected cells. Asmentioned for the prominent antigenic regions of HA, those of NAare also mutation prone and consequently highly variable, such thatslight amino-acid alteration can affect relatively subtle changes instructure, but dramatic changes in immune recognition, and permitthe evolving virus to escape the immune response and render strain-specific vaccines ineffective. To keep in step with these amino-acidadaptations, a constant updating and remodelling of HA- and NA-focussed vaccines is necessary on an annual or even biannual basis.15,16
Broader-acting, small-molecule antiviral drugs that target the func-tional sites of NA and surface ion channel protein M2 of influenzaprovide an additional line of defence.17–19 Vaccination is effectively apre-infection strategy designed currently to block the spread of analready known and characterized circulating strain, while small-molecule antivirals in clinical use are post-infection strategies, capableof broader strain inhibition. Both of these, vaccine and antiviralapproaches, are undeniably successful against seasonal epidemics,saving countless lives and billions of dollars in healthcare costs andlost productivity.
Against highly lethal pandemics, current approaches are essentiallythe same and deploy vaccines specifically modelled on the newlyemergent pandemic strain to provide homotypic neutralizing anti-body immunity to its major HA and NA Ags. But peak antibodyresponses optimally take 4–7 weeks to develop20 and are therefore toolate to prevent primary infection and development of symptoms. Bythe time such ‘seasonal’-type vaccines have been prepared in sufficientquantities for mass inoculation, for many people it may be too late.Furthermore, the efficacy of these vaccines may be short-lived. Aspandemics tend to break in multiple waves, with the strain continuallyevolving, it is likely that a vaccine-evading variant may eventuallyemerge, to once again leave people defenceless. Further to this it hasbeen questioned as to whether our current stocks of antivirals aresufficient to defend against a global pandemic, especially if alreadycompromised by an increasingly widespread emergence of antiviralresistance.21
All our present vaccination strategies are by definition post-emergent: they take considerable time, technology, materials andcost to develop, and therefore are ill-equipped to globally combata new, rapidly spreading, HPI. To minimize our vulnerability toanother pandemic, we need to develop vaccines and anti-virals thatcan be pre-emptively deployed, and provide universal rather thansubtype-restricted protection. Admittedly, this ‘ holy grail’ posesconsiderable challenges. We suggest, however, that a better under-standing of the broad-acting immune antibody defences that naturallyprotect us against all sorts of environmental pathogens can helpus identify new molecular sites for protective targeting and enableanti-viral strategies with more universal protection than currentlyexists.
NATURAL BROAD-ACTING ADAPTIVE ANTIBODY IMMUNITY
We began by asking whether unexposed humans really are antibody-defenceless against a pandemic influenza. Although the lethality of the1918 pandemic is frequently reported,22,23 what is seldom appreciatedis the fact that the majority (497%) of those infected survived. Giventhat strain-specific ‘seasonal’ neutralizing antibodies to highly variablebut dominant antigenic sites can take weeks to months to develop,24
what major immune mechanisms were immediately capable and availableto protect such a large proportion of the population? This question led
Broad human antibody protection for pandemic influenzaGW Lynch et al
150
Immunology and Cell Biology
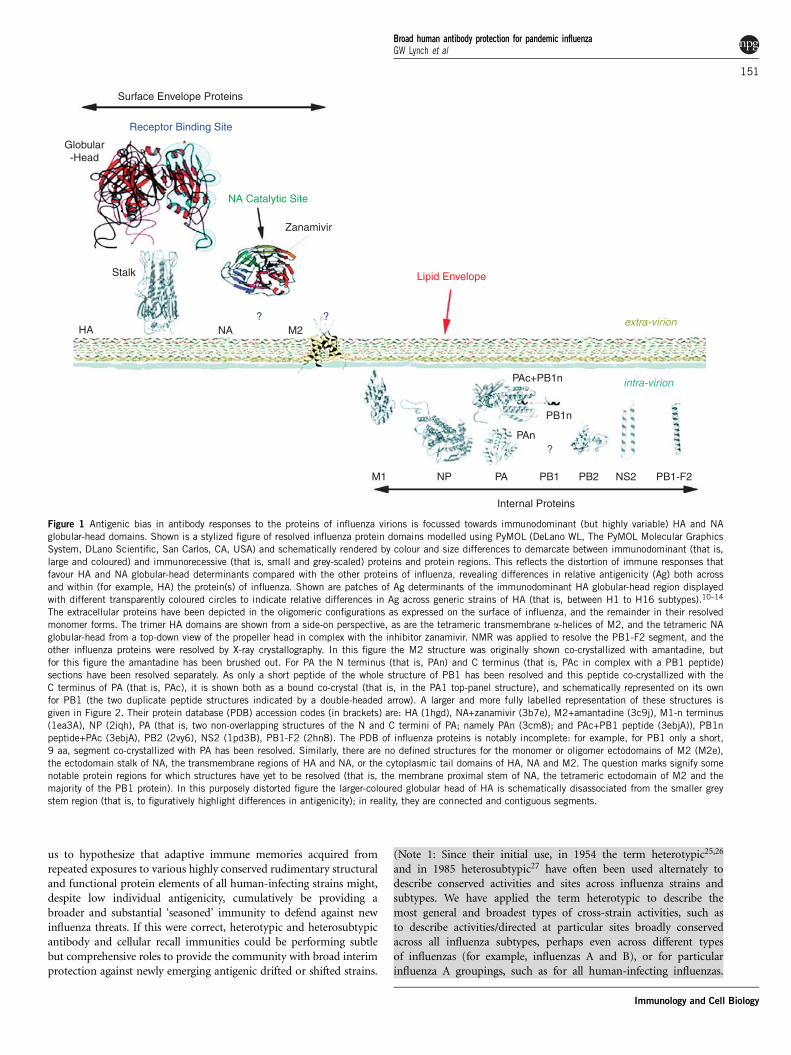
us to hypothesize that adaptive immune memories acquired fromrepeated exposures to various highly conserved rudimentary structuraland functional protein elements of all human-infecting strains might,despite low individual antigenicity, cumulatively be providing abroader and substantial ‘seasoned’ immunity to defend against newinfluenza threats. If this were correct, heterotypic and heterosubtypicantibody and cellular recall immunities could be performing subtlebut comprehensive roles to provide the community with broad interimprotection against newly emerging antigenic drifted or shifted strains.
(Note 1: Since their initial use, in 1954 the term heterotypic25,26
and in 1985 heterosubtypic27 have often been used alternately todescribe conserved activities and sites across influenza strains andsubtypes. We have applied the term heterotypic to describe themost general and broadest types of cross-strain activities, such asto describe activities/directed at particular sites broadly conservedacross all influenza subtypes, perhaps even across different typesof influenzas (for example, influenzas A and B), or for particularinfluenza A groupings, such as for all human-infecting influenzas.
Surface Envelope Proteins
Globular-Head
Zanamivir
extra-virion
intra-virion
Stalk
HA NA M2
M1 NP PA PB1
Internal Proteins
PB2 NS2 PB1-F2
PAc+PB1n
PB1n
PAn
Lipid Envelope
?
?
?
*
Receptor Binding Site
NA Catalytic Site
Figure 1 Antigenic bias in antibody responses to the proteins of influenza virions is focussed towards immunodominant (but highly variable) HA and NA
globular-head domains. Shown is a stylized figure of resolved influenza protein domains modelled using PyMOL (DeLano WL, The PyMOL Molecular Graphics
System, DLano Scientific, San Carlos, CA, USA) and schematically rendered by colour and size differences to demarcate between immunodominant (that is,
large and coloured) and immunorecessive (that is, small and grey-scaled) proteins and protein regions. This reflects the distortion of immune responses that
favour HA and NA globular-head determinants compared with the other proteins of influenza, revealing differences in relative antigenicity (Ag) both across
and within (for example, HA) the protein(s) of influenza. Shown are patches of Ag determinants of the immunodominant HA globular-head region displayed
with different transparently coloured circles to indicate relative differences in Ag across generic strains of HA (that is, between H1 to H16 subtypes).10–14
The extracellular proteins have been depicted in the oligomeric configurations as expressed on the surface of influenza, and the remainder in their resolved
monomer forms. The trimer HA domains are shown from a side-on perspective, as are the tetrameric transmembrane a-helices of M2, and the tetrameric NA
globular-head from a top-down view of the propeller head in complex with the inhibitor zanamivir. NMR was applied to resolve the PB1-F2 segment, and the
other influenza proteins were resolved by X-ray crystallography. In this figure the M2 structure was originally shown co-crystallized with amantadine, but
for this figure the amantadine has been brushed out. For PA the N terminus (that is, PAn) and C terminus (that is, PAc in complex with a PB1 peptide)
sections have been resolved separately. As only a short peptide of the whole structure of PB1 has been resolved and this peptide co-crystallized with the
C terminus of PA (that is, PAc), it is shown both as a bound co-crystal (that is, in the PA1 top-panel structure), and schematically represented on its own
for PB1 (the two duplicate peptide structures indicated by a double-headed arrow). A larger and more fully labelled representation of these structures is
given in Figure 2. Their protein database (PDB) accession codes (in brackets) are: HA (1hgd), NA+zanamivir (3b7e), M2+amantadine (3c9j), M1-n terminus
(1ea3A), NP (2iqh), PA (that is, two non-overlapping structures of the N and C termini of PA; namely PAn (3cm8); and PAc+PB1 peptide (3ebjA)), PB1n
peptide+PAc (3ebjA), PB2 (2vy6), NS2 (1pd3B), PB1-F2 (2hn8). The PDB of influenza proteins is notably incomplete: for example, for PB1 only a short,
9 aa, segment co-crystallized with PA has been resolved. Similarly, there are no defined structures for the monomer or oligomer ectodomains of M2 (M2e),
the ectodomain stalk of NA, the transmembrane regions of HA and NA, or the cytoplasmic tail domains of HA, NA and M2. The question marks signify some
notable protein regions for which structures have yet to be resolved (that is, the membrane proximal stem of NA, the tetrameric ectodomain of M2 and the
majority of the PB1 protein). In this purposely distorted figure the larger-coloured globular head of HA is schematically disassociated from the smaller grey
stem region (that is, to figuratively highlight differences in antigenicity); in reality, they are connected and contiguous segments.
Broad human antibody protection for pandemic influenzaGW Lynch et al
151
Immunology and Cell Biology

In contrast, the term heterosubtypic is used to describe a morelimited conservation of sites or activities that are also maintainedacross some select subtypes, but not all the subtypes, such as thatmight include activities/epitopes conserved between human-infectingH1N1 and H5N1 strains, but not shared with H3N2 influenzas.)Conceivably, the net effect of this would dampen downstreamcytokine-storm and cellular pathogenic responses and secondaryinfections during a pandemic until such time as a ‘seasonal’ high-titre strain-specific neutralizing host response could be mounted.There is, however, limited information or understanding of protectiveheterotypic antibody immunity to defend humans against emerginginfluenzas and especially novel zoonotic strains.
The concept of broad humoral and cellular heterotypic and hetero-subtypic immunity against influenza has been appreciated for manydecades,25 and in principle substantiated in the interim from a largenumber of mouse, rat, chicken and ferret model studies of influenzainfection (for example, reviews Epstein28 and Grebe et al.29 andSupplementary Information S1). But surprisingly, equivalent evidencethat such protective mechanisms could also be demonstrated in humaninfections has until now been limited to select conserved epitopes, andalmost exclusively involved in cell-mediated heterotypic protection ofinfluenza pathogenicity. Impeding progress and compounding thelack of evidence for heterotypic antibodies in humans has been theabsence of a suitable and generally available in vitro assay to dis-criminate, detect and specifically measure heterotypic antibodies ortheir responses. Recently an immunoblotting procedure has been usedby the authors30,32 and an Ag-specific B-cell ELISPOT assay byothers,33–35 and these are capable of discriminating heterotypic anti-body activities, but both require specialized reagents (for example,foreign (avian) virus or recombinant constructs). Hence, until nowthere has been an exclusive reliance on in vivo animal experimentation,and for the most part these have been from studies in mice. But asexpanded upon below and in the supplement, mouse influenzainfections provide a relatively poor animal model for human influenzainfection (Supplementary Table S1). Importantly, further support hasbeen demonstrated in ferrets, the most suitable animal model for thestudy of human influenza infection and heterotypic responses.36,37
Ferret infection studies of heterotypic protection are performed byfirst infecting or vaccinating with a representative construct of oneinfluenza subtype and then challenging with a more lethal subtypevirus.33,38–41 Alternately, heterotypic antibody responses generated toone subtype can be passively transferred to infection-naive animals forevaluation of protection against challenge by a different subtype.40
Early studies in ferrets, however, indicated that although beneficial forlimiting viral shedding, prior exposure to an earlier different virus,despite showing some protection, did not block infection. Further-more, because this activity was without in vitro anti-HA antibodyinhibition, the likely involvement of antibodies for heterotypic protec-tion was generally discounted.38 In contrast, a clearer and moredefinitive role for T-cell immunity in heterotypic protection wasidentified and substantiated, and has since warranted intensive hetero-typic cell-mediated vaccine efforts.38,42–45 However, more recent find-ings increasingly implicate a role for protective heterotypicantibodies,46 which can be non-neutralizing, and protect both miceand ferrets against lethal intra-nasal H5N1 infection41 (SupplementaryInformation S1).
Considering the population as a whole, in the shadow of promi-nent neutralizing homotypic antibodies that target strain-specificimmunodominant epitopes of circulating influenzas, is the possibilitythat heterotypic and heterosubtypic antibodies can be produced thattarget immunorecessive sites that are widely conserved across strains.
However, the latter possibility has until recently generally beendiscounted for humans as non-existent or irrelevant as a protectoragainst circulating or novel influenzas. Findings from our laboratoryand elsewhere raise the question as to whether this is indeed thecase. This article reviews this emerging field of heterotypic antibodiesand raises the issue of whether broad-acting heterotypic humanantibodies may cooperatively provide an important first line ofdefence against new and emerging strains of influenza. Because of apaucity of information on human heterotypic antibodies, it is there-fore appropriate to be guided from heterotypic and heterosubtypicantibody responses observed in animal models of infection36,37
(Supplementary Information S1).
‘Seasonal’ and ‘seasoned’ antibody immune responses in humansIt is proposed that protection against pandemic influenza in thepopulation at large is provided by two separate adaptive antibodyimmune responses: ‘seasonal’ and ‘seasoned’ immunities.
Seasonal immunity is well established as a strain-specific antibodyresponse directed at variable mutation-prone, immunodominantepitopes of particular influenzas and/or vaccines modelled on them.Seasonal antibodies neutralize virus and can be measured both in vitroand in vivo. Producing these antibodies is the focus of current vaccineinitiatives.
Seasoned immunity, on the other hand, would provide a broad-acting (heterotypic, heterosubtypic), adaptive antibody responsedirected at a variety of highly conserved, immunorecessive epitopes,broadly displayed in all influenzas that infect humans, and thereforemay synergistically affect the disruption of most, if not every phase ofviral infection and replication. Because those sites are of low immu-nogenicity, predictably, seasoned immunity would develop slowlyfrom repeated lifetime exposures to the same conserved sites ofmany different influenza strains, and if so, would be age-dependent.Our prediction is that seasoned responses will include both neutraliz-ing and non-neutralizing antibodies, and in general singularly be ofmodest action, and therefore are difficult to detect in vitro, butcollectively they would give significant protection in vivo, and thelatter can be measured. If proven, a stimulation of seasoned immunitymay then offer an enticing means upon which new vaccine strategiescould be developed for the targeting of epitopes conserved across all,or at least many, influenza strains. This is an area of increasing interestfor R&D and it is proposed that conserved epitopes for vaccines andsmall-molecule inhibitors for universal influenza protection can berevealed by seasoned heterotypic human antibodies sampled inintravenous immunoglobulins, prepared from tens of thousands ofnormal blood donations.30,32,47
Although not the subject of this article, help from the T-lympho-cytic arm of the immune system, via heterotypic T-cell responses,would offer an additional level of protection against HPI-inducedpathogenesis and death, by facilitating viral clearance and reducing thedisease severity and T-cell helper activities,15,48–53 to synergisticallycomplement and assist heterotypic antibody responses (see Figure 1 inLynch et al.54). Although both types of heterotypic immune defencesmay in themselves individually be modest in comparison withimmunodominant homotypic neutralizing responses, in combinationthey could conceivably provide a formidable barrier to emerging infec-tious agents. Thus, we believe that as-yet-to-be designed new-generationheterotypic antibody-focussed vaccines, along with new heteroge-neously acting, small-molecule antivirals, directed at conserved func-tional sites (for example, NA and M2 inhibitors and potentially newones to be added), will add to, and complement, the various hetero-typic T-cell vaccine and other different initiatives being separately
Broad human antibody protection for pandemic influenzaGW Lynch et al
152
Immunology and Cell Biology

pursued in the quest for the development of universal vaccines forinfluenza.55–60
THE EVIDENCE FOR AN ACQUIRED HUMAN ‘SEASONED’
ANTIBODY IMMUNITY
Both indirect and direct indicators of seasoned heterotypic humoralimmunity in humans are steadily mounting. First, in the survival datafrom the epidemics and pandemics of the twentieth century, webelieve the signs of its existence are apparent (Supplementary Infor-mation S1). Even in the case of H5N1, where mortality is far higher(60%) than that of the 1918 H1N1 influenza (o3%), again attentionis drawn to the significant rate of survival of approximately 40%.61
Epidemiological studies have also found that of the H5N1 survivorsapproximately 60% are over the age of 40, in contrast with fewerthan 25% for the 10–20-years age group.62 This suggests that protec-tion is age-dependent. Similarly, data from the 1918 H1N1 and 1957H2N2 pandemics indicate a greater level of protection in adults,63,10
and this has now also become evident for 2009 H1N1 infections.64
Fascinatingly, this appears to reduce the impact of infection bypandemic strains for the subpopulation of adults older than 65 toa level more similar to a seasonal infection.10 As more than 97% ofthe infected population have survived past pandemic influenzas, thesefindings imply a significant level of pre-existing adaptive immuneprotection beyond that afforded by innate or adaptive heterotypic cell-mediated immune processes, thus implicating that pools ofpre-existing and broad-acting antibodies, may assist CD4+ T-cellprocesses in providing comprehensive and heterotypic protectionagainst novel influenzas, including human-infecting avian and porcinestrains.
Studies of intravenous immunoglobulins sourced from differentgeneral global populations have revealed age-dependent hetero-typic antibodies that show specificity in reacting with the proteinsof both intact and solubilized H5N1, bind H5-hemagglutinin andneutralize H5N1 infection in vitro.30,65,66 In defining the typesof heterotypic antibodies that could be involved, several groupshave identified human-derived cross-protective monoclonal anti-bodies, for example, from combinatorial B cell libraries fromseasonal influenza-vaccinated individuals.67–69 These antibodies alltarget conserved sites of the stem HA2 domain of HA. This isthe most highly conserved section of HA70,71 and contains theactivation cleavage site and membrane fusion peptide. Notably,these different heterotypic antibodies provide cross-neutralizationof a wide range of influenza strains that include HA clades 1 and 2(that is, including H1, H2, H5 and H9 strains),67,68,71,72,11 and thusprotect mice69 and ferrets40 against H5N1 infection and pathogenesis.(Note 2: The HA subtypes, 1–16, have been classified into four clades,from defined or predicted HA structures, and from their respectiveamino-acid and sequence phylogenetic analyses. Namely, as Clade-173
(also termed the H1-clade69), which comprises H1, H2, H5, H6, H11,H13 and H6 subtypes; Clade-2 (H9-clade), with H8, H9 and H12;Clade-3 (H3-clade), with H3, H4 and H14; and Clade-4 (H7-clade),with H7, H10 and H15.69,73 From their phylogenetic relationships,the clades themselves have been further demarcated into group 1(that is, clades 1 and 2) and group 2 clades (that is, clades 3 and 4).)But, notably, this antibody cohort does not protect against influenzasubtypes of viruses belonging to HA clades 3 and 4 (including H3,H7 and H10 strains), thereby implicating a class of heterosubtypic(see Note 1), rather than heterotypic, antibodies that target HA stemdomains conserved in select strains for which they cross-protect, butnot for other strains. A plausible explanation for such inter-straindifferences in targeting could be the inter-HA subtype variability in
glycosylation, secondary structure or amino acids that affects theexposure or accessibility of the conserved region and/or small butsignificant amino-acid differences (for example, affecting charge) inthe site itself.67,11,74,75 As a driver of antigenic drift, the role of muta-tional change in the immunodominant hyper-variable HA1 globularhead is well documented. Notably, in a study of antibody escapemutants of the 1957 strain of H2N2, a monoclonal antibody to aunique site on the HA2 stem of HA was identified,76 and in supportof the importance of neutralizing antibodies to this region of H1and H2 subtypes.67,68,72,11 Further to cross-typic HA sites, Sandbulteet al.77 also revealed cross-reactive anti-neuraminidase antibodiesfrom unexposed humans that partially protect against H5N1 infectionin mice. These, together with heterotypic antibodies to conserved sitesof M2e, NP, M1, NS1 (below and Supplementary Information S2)and likely others yet-to-be-identified, provide a means to explain theanti-H5N1 heterotypic antibody activities of intravenous immuno-globulins30,32 and findings showing that seasonal vaccination ofindividuals naive to H5N1 can stimulate heterotypic and neutralizingantibody protection against H5N1.24,71
Why is it that evidence of seasoned immunity is only now comingto light? Influenza is a complex disease and there are many factors—viral, host, man-made and environmental—that affect infection,morbidity and mortality. Operating in the shadow of dramatic andobvious homotypic neutralizing antibodies that disrupt receptorbinding, the question of whether, all along, diverse and subtleheterotypic responses have also been providing a significant level ofimmune defence has little been considered or properly addressed.There may be several reasons for overlooking heterotypic antibodyresponses in humans: the means of assessment and the size of thecross-typic antibody response, which, if detected at all, is likely to bemeasured in the low to equivocal levels of currently deployed in vitroassays (that is, hemagglutinin inhibition and virus microneutraliza-tion), recently considered to underrate the actual in vivo protection;78
the properties of the influenza Ags used in cross-protection studies;the competitive nature of Ag responses promoted by influenza; andthe modes of delivery of vaccines with or without adjuvant. We expectthat most heterotypic antibody responses are limited to studies ofnatural infection or for in vivo animal models in laboratories that aresuitably equipped. In the light of the accruing new data, it is, however,now vital to test the seasoned immunity hypothesis in whole-bodyinfections, and to understand how select, universally conservedinfluenza protein epitopes might be optimally packaged and deliveredto ensure a maximal and broad stimulation of putative ‘background’reservoir(s) of heterotypic B-cell immunity. However, it is impor-tant to be alert to genetic and environmental factors that mayinfluence heterotypic and heterosubtypic immune responses, as wellas age-dependent response differences in children and adolescents(developing immune systems), adults (mature) and the elderly(immunosenescence)35,79,80 that clearly influence immune susceptibil-ities to epidemic and pandemic influenzas, host cytokine responsesand vulnerability to secondary infection.81–84
SEASONAL VERSUS SEASONED VACCINES
In considering the design of vaccines to generate pre-emptiveheterotypic antibody immunity against emerging influenzas, it isnecessary to take into account what appears to be a disconnectionbetween how seasonal vaccines are modelled and the requirementsneeded for seasoned vaccines. In the well-established seasonalvaccination process, present vaccines are modelled to provide HAglobular-head Ags that match the circulating virus strains and stimu-late high-titre neutralizing antibodies. Protective antibody activities
Broad human antibody protection for pandemic influenzaGW Lynch et al
153
Immunology and Cell Biology
and responses for these can be rapidly measured by hemagglutinininhibition and micro-neutralization assays in vitro. In this scheme,any collateral non-neutralizing antibody-mediated responses forinfluenza virus and infected cell clearance are not usually measured,and any such protective benefits generated en route would in effectbe an unmeasured and unrealized bonus. The challenge now presentedis how to design vaccines capable of promoting broad heterotypic (orheterosubtypic) protection against new, foreign, zoonotic and pre-pandemic strains, as well as antigenically drifted human influenzasthat cause epidemics. In this new ‘seasoned’ vaccine scenario, it ispredicted that a significant fraction of the heterotypic antibodyresponses may be non-neutralizing, but ably backed by a neutralizingheterosubtypic antibody response component (for example, to theconserved HA stem67,68). Major requirements are that such conservedsites be accessible to antibodies or designer drugs, and that, viamultiple heterotypic B-cell-dependent mechanisms, collectively becapable of disrupting all aspects of infection, replication and patho-genicity.85 For optimal protection, it would of course be prudent toultimately stimulate both antibody and T-cell responses to conserveddeterminants of all human-infecting influenzas.
REQUIREMENTS FOR SEASONED HUMORAL VACCINE
DESIGN
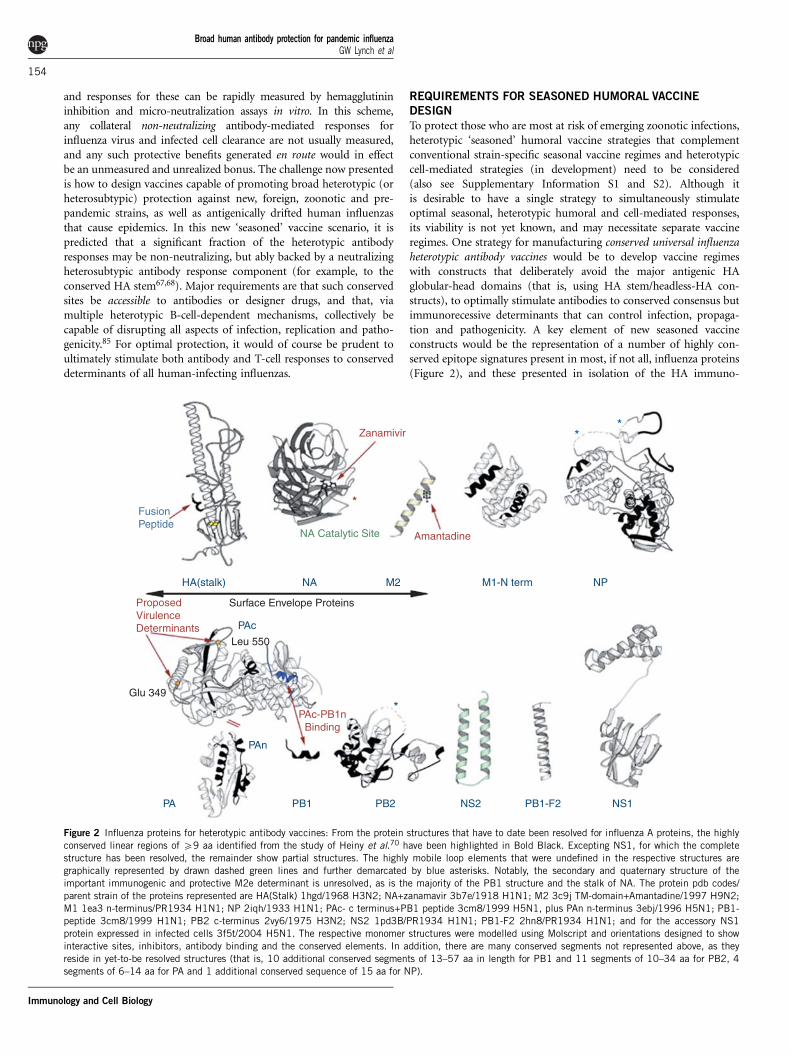
To protect those who are most at risk of emerging zoonotic infections,heterotypic ‘seasoned’ humoral vaccine strategies that complementconventional strain-specific seasonal vaccine regimes and heterotypiccell-mediated strategies (in development) need to be considered(also see Supplementary Information S1 and S2). Although itis desirable to have a single strategy to simultaneously stimulateoptimal seasonal, heterotypic humoral and cell-mediated responses,its viability is not yet known, and may necessitate separate vaccineregimes. One strategy for manufacturing conserved universal influenzaheterotypic antibody vaccines would be to develop vaccine regimeswith constructs that deliberately avoid the major antigenic HAglobular-head domains (that is, using HA stem/headless-HA con-structs), to optimally stimulate antibodies to conserved consensus butimmunorecessive determinants that can control infection, propaga-tion and pathogenicity. A key element of new seasoned vaccineconstructs would be the representation of a number of highly con-served epitope signatures present in most, if not all, influenza proteins(Figure 2), and these presented in isolation of the HA immuno-
FusionPeptide
NA Catalytic Site
HA(stalk) NA M2 M1-N term NP
PAc
PAn
PA PB1 PB2 NS2 PB1-F2 NS1
*
Leu 550
Glu 349
PAc-PB1nBinding
Surface Envelope Proteins
*
Zanamivir
Amantadine
ProposedVirulenceDeterminants
**
Figure 2 Influenza proteins for heterotypic antibody vaccines: From the protein structures that have to date been resolved for influenza A proteins, the highly
conserved linear regions of X9 aa identified from the study of Heiny et al.70 have been highlighted in Bold Black. Excepting NS1, for which the complete
structure has been resolved, the remainder show partial structures. The highly mobile loop elements that were undefined in the respective structures are
graphically represented by drawn dashed green lines and further demarcated by blue asterisks. Notably, the secondary and quaternary structure of the
important immunogenic and protective M2e determinant is unresolved, as is the majority of the PB1 structure and the stalk of NA. The protein pdb codes/
parent strain of the proteins represented are HA(Stalk) 1hgd/1968 H3N2; NA+zanamavir 3b7e/1918 H1N1; M2 3c9j TM-domain+Amantadine/1997 H9N2;
M1 1ea3 n-terminus/PR1934 H1N1; NP 2iqh/1933 H1N1; PAc- c terminus+PB1 peptide 3cm8/1999 H5N1, plus PAn n-terminus 3ebj/1996 H5N1; PB1-peptide 3cm8/1999 H1N1; PB2 c-terminus 2vy6/1975 H3N2; NS2 1pd3B/PR1934 H1N1; PB1-F2 2hn8/PR1934 H1N1; and for the accessory NS1
protein expressed in infected cells 3f5t/2004 H5N1. The respective monomer structures were modelled using Molscript and orientations designed to show
interactive sites, inhibitors, antibody binding and the conserved elements. In addition, there are many conserved segments not represented above, as they
reside in yet-to-be resolved structures (that is, 10 additional conserved segments of 13–57 aa in length for PB1 and 11 segments of 10–34 aa for PB2, 4
segments of 6–14 aa for PA and 1 additional conserved sequence of 15 aa for NP).
Broad human antibody protection for pandemic influenzaGW Lynch et al
154
Immunology and Cell Biology
dominant regions (and possibly NA-globular-head determinants)traditionally selected for seasonal vaccines. Alternatively, presentationof those conserved Ags in an environment (for example, using MF-59adjuvant) conducive to enhance the antigenicity of otherwise immu-norecessive conserved sites (Supplementary Information S3) may alsohelp overcome the immune imbalance between poorly immunogenicconserved domains39,86–88 and immunodominant epitopes of the HAand NA globular heads(s). Such approaches are necessary to provide aless-distracted immune focus for the stimulation of broadly protectiveantibodies to conserved determinants across the proteins of all (ormany) influenza subtypes, for which a summation of many differentheterotypic and heterosubtypic antibody effects could affect blanketprotection.
The already identified candidate sites for broadly protective vaccineswould include immunorecessive consensus sites of the HA stem,67,68
and the conserved and exposed ectodomain of M2 (M2e) to stimulateheterosubtypic antibody protection, as found against heterologousstrains of influenza in mice, ferret and monkey studies (see Supple-mentary Information S2).89–98 Although M2 is minimally expressedon the virion, it is highly expressed by influenza-infected cells and istherefore a suitable antibody target for the disruption of viral replica-tion and infected-cell clearance or lysis, and may involve antibody-dependent NK-cell mediated cytotoxicity.89,91,92,99 As shown for HAstem sites, it is predicted is that as-yet-to-be identified inter-subtypicconserved sites of NA, including conformational and discontinuousepitopes, will similarly be found, for which heterosubtypic antibodytargeting could disrupt sialidase activity and retard new virion bud-ding.31,77,100 It would appear that a minimal requirement for universalvaccines is that they be multivalent and represent conserved protectiveantigenic signatures representative of the respective phylogenetic; HAgroup 1 and 2 subtypes, NA group 1 and 2 subtypes, and M2esignatures of human and swine infecting strains.101–103 However, forsustained and complete protection, vaccines would also need tocomprehensively and combinatorially include conserved signatures
of the highly conserved intra-virion proteins PB2, NP, M1, and so on(Figure 1, Supplementary Information S1).104–112
Therefore, universal heterosubtypic antibody-based vaccine strate-gies would be complemented by alternate less-universal heterosubty-pic, perhaps other non-coincident, antibody-based approaches, usingpanels of less-widely conserved more subtype-selective libraries fordeterminants of neutralization and pathogenicity that would includeconstructs with conserved discontinuous epitopes (for example,globular-head HA and NA domains) displayed within conforma-tion-maintained low to neutral Ag scaffolds,113 with the latter directedat subtype-specific conserved elements, in HA and NA, to promoteneutralizing subtype-specific antibody responses. DNA and recombi-nant approaches may be best suited to deliver the complex arrayof conserved linear and discontinuous and conformational Ag signa-tures required for universal seasoned (for example, Figure 2 andSupplementary Information S2) or subtype-selective (for example,Figure 3 and Supplementary Information S2) antibody-based vaccineprotection. To support such initiatives, additional combinatorial,in silico and small-molecule drug design studies are also needed, forthe design of site-specific agents to disrupt protein functions thatengage conserved epitopes displayed in antibody-inaccessible clefts.Important conserved sites for targeting would include epitopesthat could disrupt determinants involved in transmission, virulence,human adaptation or pathogenesis4,115 (see Supplementary Table 1 ofBautista et al.116).
CONCLUSIONS
To meet the challenge of the next influenza pandemic, a morecomprehensive defence approach is needed than is currently providedby strain-specific trivalent vaccines. This approach should target asmany protective sites as possible that are broadly conserved across allthe influenza proteins and, predictably, this will promote cross-reactiveantibody protection against new and emergent influenzas, includingzoonotic strains from birds and swine. For this, two different anti-body-based protective approaches are proposed, a conserved universalvaccine strategy based on targeting many conserved but immuno-recessive elements maintained across influenza subtypes, and aconserved subtype-selective vaccine strategy to target elements morerestrictively conserved within strains or protein subgroups, whichmay include immunodominant epitopes. Underlying these strategiesis the need for concerted efforts to test the new hypotheses elucidatedhere, namely (i) that intravenous immunoglobulin antibodies canguide identification of new heterotypic protective antibody epi-topes;30,32,54 (ii) that conserved epitopes (such as those of the HAstem) presented alone without immunodominant HA globular-head(and possibly NA) domains will significantly increase heterotypicantibody responses (Supplementary Information S2); (iii) that vacci-nation with an extensive panel representing conserved epitopes inboth external and intravirion, NP, HA, NA, M2, HMWT polymerase,M1 and NS proteins, will stimulate broad (universal) protectionagainst most, if not all, influenzas (Supplementary Information S2);and (iv) that adjuvants could improve the immune visibility ofimmunorecessive conserved epitopes, for the delivery of sound (uni-versal) antibody protection (Supplementary Information S3). It isenvisaged the proposed seasoned vaccines will act in concert with, andsynergistically complement, rather than replace, pre-existing seasonalvaccine initiatives. Substantial validation has recently been providedon (ii) above, since our earlier reports and submissions,30,32,54
namely, with the demonstration that vaccination with HA-stem orheadless-HA constructs confers heterotypic neutralizing antibodyprotection across clade 1 and 2 HA subtype influenzas.117,118 A finding
EASLQIIP
SWSFLD21.140
YNNTFLA5.10
Figure 3 H5 HA globular-head epitopes for neutralizing mAbs that broadly
protect across the clades of H5N1. To visualize the binding epitopes for the
mAbs FLA5.10 and FLD21.140 identified in the Khurana et al. study,114
using Molscript these are shown as ball-and-stick sites on the pdb 2ibx
ribbon structure of the HA globular head of A/Human/VietNam/1194/2004
H5N1, and using single letter aa coding the discontinuous sites for the
protective mAb FLD21.140 demarcated in red from the mAb FLA5.10 sites
in blue.
Broad human antibody protection for pandemic influenzaGW Lynch et al
155
Immunology and Cell Biology

solidly predicted by an earlier visionary study in mice is thatheterotypic antibodies generated to the globular-head-deleted(headless) HA construct could protect mice against H1 and H2strain infections.119
It would seem that the generation of appreciable heterotypic anti-body protection in humans is a slow process that will take 30 years ormore to adequately develop in response to repeated seasonal influenzaexposures (and is enhanced by some seasonal vaccine strategies).Perhaps the prolonged, subtle and measured development of thisadaptive response has historically undermined its real value forimmune protection, especially as the early heterologous influenzaexperiments in animals involved single cycles of infection or vaccina-tion, and, although it generated some heterotypic protection against adifferent influenza subtype, owing to lack of an additive enhancementof multiple different exposures, had failed to drive visible heterotypicantibody immunity. This may have undervalued the real protectivepotential for heterotypic antibodies in vaccine design, until now.Therefore, a need for new vaccine initiatives directed at conserveddomains, and perhaps including strategies that may incorporate invivo electroporation and vaccination of synthetic HA, NA, M2 and NPconsensus DNA Ags is indicated120,121 (Supplementary InformationS2), in order to drive more efficient and rapid stimulation ofheterotypic antibody responses than can be delivered by environ-mental exposures alone. However, while advocating the developmentof heterotypic-based vaccines, it is also necessary to demarcate thoseseasonal vaccines, currently in use, that are capable of stimulatingsome heterotypic antibody co-protection, but mindful of what effectthey might have on homotypic and heterotypic antibody responses tosubsequent influenza exposures.34
Since our initial report on heterotypic human anti-influenza anti-bodies in 2006,66 interest in the subject has grown considerably, withmore than 50 articles being published over the past 2 years in supportof a protective role for heterotypic antibodies in humans. Requirednow are strategies that can overcome the major shortcoming ofcurrent antiviral regimens that can be negated by escape mutants.21
Predictably, a broad and simultaneous blanket targeting of multipleconserved influenza epitopes, to provide sustained universal influenzaprotection against antigenic drift and shift, can accomplish this. Asanti-viral resistance has been a growing trend in the evolution of otherviruses (for example, HIV, HCV, HBV), a more general application ofthe principles proposed here for influenza may also be viable forprotection against other infectious diseases. To support this sugges-tion, a somewhat parallel situation has emerged with studies revealingprotective antibodies from HIV-exposed individuals that are directedat conserved epitopes shared across the clades of HIV and directed atthe stem of HIV’s envelope, gp120.67,68,122,123 Exciting times lie aheadfor the realization of the various targets and complementary new waysthat can be marshalled to achieve universal protection of influenza andother pathogens capable of pandemic spread.
In the event of a full-blown HPI pandemic, how confident are we thatour current antiviral capabilities will contain it? Will future generationsjudge that we should have used the lull following the many warningsignals since the first H5N1 outbreak in 1997 to develop broader actinganti-virals? (Note 3: WHO H5N1 Timeline of H5N1 events: http://www.who.int/csr/disease/avian_influenza/timeline.pdf). The H1N1 pandemicin 2009 and reports of adaptive H5N1 mutations124 may be indicatorsthat this valuable window of opportunity could end abruptly.
CONFLICT OF INTEREST
The authors declare no conflict of interest.
ACKNOWLEDGEMENTSWe thank colleagues who have offered constructive and valued appraisal and
comment of various aspects of our proposals or manuscript, namely Professors
William Rawlinson (Clinical Virologist), Peter Doherty (Immunologist, Virol-
ogist), Peter Williamson (Immunologist), Hans Zoellner (Pathologist) and
Robert Booy (Clinical Epidemiologist). We also thank Mr Tim Haydon, Write
Media, for his advice on writing presentation. Drs Lynch and Sullivan are
recipients of a University of Sydney grant for the study of pandemic influenza.
Author contributions: GWL and JSS wrote the article with advice from
PS and WBC, particularly on those sections pertaining to their individual
expertise in avian influenza and molecular modelling, respectively. GWL,
PS and JSS performed the human antibody studies for reactivity with and
inhibition of avian H5N1 influenzas, as those investigations with published
and yet-to-be-published findings have underpinned the tenets of the arguments
presented and design of this article. WBC and GWL designed and analysed
the protein structures.
1 Fraser C, Donnelly CA, Cauchemez S, Hanage WP, Van Kerkhove MD, Hollingsworth TD
et al. Pandemic potential of a strain of influenza A (H1N1): early findings. Science
2009; 324: 1557–1561.2 Morens DM, Taubenberger JK, Fauci AS. The persistent legacy of the 1918 influenza
virus. New Engl J Med 2009; 361: 225–229.3 Smith GJ, Vijaykrishna D, Bahl J, Lycett SJ, Worobey M, Pybus OG et al. Origins and
evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. Nature
2009; 459: 1122–1125.4 Allen JE, Gardner SN, Vitalis EA, Slezak TR. Conserved amino acid markers from past
influenza pandemic strains. BMC Microbiol 2009; 9: 77.5 Sorrell EM, Wan H, Araya Y, Song H, Perez DR. Minimal molecular constraints for
respiratory droplet transmission of an avian-human H9N2 influenza A virus. Proc NatlAcad Sci USA 2009; 106: 7565–7570.
6 Jackson S, Van Hoeven N, Chen LM, Maines TR, Cox NJ, Katz JM et al. Reassortment
between avian H5N1 and human H3N2 influenza viruses in ferrets: a public health
risk assessment. J Virol 2009; 83: 8131–8140.7 Li C, Hatta M, Nidom CA, Muramoto Y, Watanabe S, Neumann G et al. Reassortment
between avian H5N1 and human H3N2 influenza viruses creates hybrid viruses with
substantial virulence. Proc Natl Acad Sci USA 2010; 107: 4687–4692.8 McKibben WJ, Sidorenko AA. Global Macroeconomic Consequences of Pandemic
Influenza. February Analysis. Lowy Institute for International Policy, 2006, pp 1–79.9 Adar Y, Singer Y, Levi R, Tzehoval E, Perk S, Banet-Noach C et al. A universal
epitope-based influenza vaccine and its efficacy against H5N1. Vaccine 2009; 27:
2099–2107.10 Stevens J, Corper AL, Basler CF, Taubenberger JK, Palese P, Wilson IA. Structure of
the uncleaved human H1 hemagglutinin from the extinct 1918 influenza virus.
Science 2004; 303: 1866–1870.11 Xu R, McBride R, Paulson JC, Basler CF, Wilson IA. Structure, receptor binding, and
antigenicity of influenza virus hemagglutinins from the 1957 H2N2 pandemic. J Virol
2010; 84: 1715–1721.12 Stevens J, Blixt O, Tumpey TM, Taubenberger JK, Paulson JC, Wilson IA. Structure
and receptor specificity of the hemagglutinin from an H5N1 influenza virus. Science
2006; 312: 404–410.13 Xu R, Ekiert DC, Krause JC, Hai R, Crowe Jr JE, Wilson IA. Structural basis of
preexisting immunity to the 2009 H1N1 pandemic influenza virus. Science 2010;
328: 357–360.14 Wright PF, Webster RG. Orthomyxoviruses. In: Knipe DM, Howley PM (eds). Fields
Virology, 4th edn. Lippincott: Philadelphia, 2001, pp 1533–1579.15 Kreijtz JH, de Mutsert G, van Baalen CA, Fouchier RA, Osterhaus AD, Rimmelzwaan
GF. Cross-recognition of avian H5N1 influenza virus by human cytotoxic T-lymphocyte
populations directed to human influenza A virus. J Virol 2008; 82: 5161–5166.16 Richard SA, Viboud C, Miller MA. Evaluation of Southern Hemisphere influenza
vaccine recommendations. Vaccine 2010; 28: 2693–2699.17 Gong J, Fang H, Li M, Liu Y, Yang K, Liu Y et al. Potential targets and their relevant
inhibitors in anti-influenza fields. Curr Med Chem 2009; 16: 3716–3739.18 Gong J, Xu W, Zhang J. Structure and functions of influenza virus neuraminidase. Curr
Med Chem 2007; 14: 113–122.19 Schmidt AC. Antiviral therapy for influenza: a clinical and economic comparative
review. Drugs 2004; 64: 2031–2046.20 Couch RB, Kasel JA. Immunity to influenza in man. Annu Rev Microbiol 1983; 37:
529–549.21 Layne SP, Monto AS, Taubenberger JK. Pandemic influenza: an inconvenient muta-
tion. Science 2009; 323: 1560–1561.22 Palese P, Tumpey TM, Garcia-Sastre A. What can we learn from reconstructing the
extinct 1918 pandemic influenza virus? Immunity 2006; 24: 121–124.23 Taubenberger JK, Morens DM. 1918 Influenza: the mother of all pandemics. Emerg
Infect Dis 2006; 12: 15–22.
The H1N1 pandemicanti-virals?
Broad human antibody protection for pandemic influenzaGW Lynch et al
156
Immunology and Cell Biology

24 Gioia C, Castilletti C, Tempestilli M, Piacentini P, Bordi L, Chiappini R et al. Cross-subtype immunity against avian influenza in persons recently vaccinated for influenza.Emerg Infect Dis 2008; 14: 121–128.
25 Schulman JL, Kilbourne ED. Induction of partial specific heterotypic immunity inmice by a single infection with influenza a virus. J Bacteriol 1965; 89: 170–174.
26 Howe HA. Studies of active immunogenesis in poliomyelitis. I. Persistence and recallby homotypic or heterotypic superinfection of neutralizing antibody originally inducedin chimpanzees by vaccination or infection. Am J Hyg 1954; 60: 371.
27 Burlington DB, Wright PF, van Wyke KL, Phelan MA, Mayner RE, Murphy BR.Development of subtype-specific and heterosubtypic antibodies to the influenza Avirus hemagglutinin after primary infection in children. J Clin Microbiol 1985; 21:847–849.
28 Epstein SL. Control of influenza virus infection by immunity to conserved viralfeatures. Expert Rev Anti-Infect Ther 2003; 1: 627–638.
29 Grebe KM, Yewdell JW, Bennink JR. Heterosubtypic immunity to influenza A virus:where do we stand? Microbes Infect Institut Pasteur 2008; 10: 1024–1029.
30 Lynch GW, Selleck PW, Axell A-M, Downton T, Kapitza N, Boehm I et al. Cross-reactiveanti-avian H5N1 influenza neutralizing antibodies in a normal ‘Exposure-Naı̈ve’Australian blood donor population. Open Immunol J 2008; 1: 13–19.
31 Reid AH, Fanning TG, Janczewski TA, Taubenberger JK. Characterization of the 1918‘Spanish’ influenza virus neuraminidase gene. Proc Natl Acad Sci USA 2000; 97:6785–6790.
32 Sullivan JS, Selleck PW, Downton T, Boehm I, Axell AM, Ayob Y et al. Heterosubtypicanti-avian H5N1 influenza antibodies in intravenous immunoglobulins from globallyseparate populations protect against H5N1 infection in cell culture. J Mol Genet Med2009; 3: 217–224.
33 Cheng X, Eisenbraun M, Xu Q, Zhou H, Kulkarni D, Subbarao K et al. H5N1 vaccine-specific B cell responses in ferrets primed with live attenuated seasonal influenzavaccines. PLoS One 2009; 4: e4436.
34 Sasaki S, He XS, Holmes TH, Dekker CL, Kemble GW, Arvin AM et al. Influence ofprior influenza vaccination on antibody and B-cell responses. PLoS One 2008; 3:e2975.
35 Sasaki S, Jaimes MC, Holmes TH, Dekker CL, Mahmood K, Kemble GW et al.Comparison of the influenza virus-specific effector and memory B-cell responses toimmunization of children and adults with live attenuated or inactivated influenza virusvaccines. J Virol 2007; 81: 215–228.
36 Belser JA, Szretter KJ, Katz JM, Tumpey TM. Use of animal models to understand thepandemic potential of highly pathogenic avian influenza viruses. Adv Virus Res 2009;73: 55–97.
37 Maines TR, Szretter KJ, Perrone L, Belser JA, Bright RA, Zeng H et al. Pathogenesis ofemerging avian influenza viruses in mammals and the host innate immune response.Immunol Rev 2008; 225: 68–84.
38 Yetter RA, Barber WH, Small Jr PA. Heterotypic immunity to influenza in ferrets. InfectImmun 1980; 29: 650–653.
39 Forrest HL, Khalenkov AM, Govorkova EA, Kim JK, Del Giudice G, Webster RG. Single-and multiple-clade influenza A H5N1 vaccines induce cross protection in ferrets.Vaccine 2009; 27: 4187–4195.
40 Friesen RH, Koudstaal W, Koldijk MH, Weverling GJ, Brakenhoff JP, Lenting PJ et al.New class of monoclonal antibodies against severe influenza: prophylactic andtherapeutic efficacy in ferrets. PLoS One 2010; 5: e9106.
41 Perrone LA, Ahmad A, Veguilla V, Lu X, Smith G, Katz JM et al. Intranasal vaccinationwith 1918 influenza virus-like particles protects mice and ferrets from lethal 1918and H5N1 influenza virus challenge. J Virol 2009; 83: 5726–5734.
42 Braciale TJ. Immunologic recognition of influenza virus-infected cells. I. Generation ofa virus-strain specific and a cross-reactive subpopulation of cytotoxic T cells in theresponse to type A influenza viruses of different subtypes. Cell Immunol 1977; 33:423–436.
43 Effros RB, Doherty PC, Gerhard W, Bennink J. Generation of both cross-reactive andvirus-specific T-cell populations after immunization with serologically distinct influ-enza A viruses. J Exp Med 1977; 145: 557–568.
44 Doherty PC, Kelso A. Toward a broadly protective influenza vaccine. J Clin Invest2008; 118: 3273–3275.
45 Liang S, Mozdzanowska K, Palladino G, Gerhard W. Heterosubtypic immunity toinfluenza type A virus in mice. Effector mechanisms and their longevity. J Immunol1994; 152: 1653–1661.
46 Nguyen HH, van Ginkel FW, Vu HL, McGhee JR, Mestecky J. Heterosubtypic immunityto influenza A virus infection requires B cells but not CD8+ cytotoxic T lymphocytes.J Infect Dis 2001; 183: 368–376.
47 Pichyangkul S, Jongkaewwattana A, Thitithanyanont A, Ekchariyawat P, Wiboon-ut S,Limsalakpetch A et al. Cross-reactive antibodies against avian influenza virus A(H5N1). Emerg Infect Dis 2009; 15: 1537–1539.
48 Roti M, Yang J, Berger D, Huston L, James EA, Kwok WW. Healthy human subjectshave CD4+ T cells directed against H5N1 influenza virus. J Immunol 2008; 180:1758–1768.
49 Jameson J, Cruz J, Terajima M, Ennis FA. Human CD8+ and CD4+ T lymphocytememory to influenza A viruses of swine and avian species. J Immunol 1999; 162:7578–7583.
50 Lee LY, Ha DL, Simmons CP, de Jong MD, Chau NV, Schumacher R et al.Memory T cells established by seasonal human influenza A infection cross-react with avian influenza A (H5N1) in healthy individuals. J Clin Invest 2008;118: 3478–3490.
51 McMichael AJ, Gotch FM, Noble GR, Beare PA. Cytotoxic T-cell immunity to influenza.N Engl J Med 1983; 309: 13–17.
52 Gotch F, McMichael A, Smith G, Moss B. Identification of viral molecules recog-nized by influenza-specific human cytotoxic T lymphocytes. J Exp Med 1987; 165:
408–416.53 McKinstry KK, Strutt TM, Swain SL. Hallmarks of CD4T cell immunity against
influenza. J Intern Med 2011; 269: 507–518.54 Lynch GW, Selleck P, Sullivan JS. Acquired heterosubtypic antibodies in human
immunity for avian H5N1 influenza. J Mol Genet Med 2009; 3: 205–209.55 Hampson AW. Vaccines for pandemic influenza. The history of our current vaccines,
their limitations and the requirements to deal with a pandemic threat. Ann Acad MedSingapore 2008; 37: 510–518.
56 Nabel GJ, Fauci AS. Induction of unnatural immunity: prospects for a broadly
protective universal influenza vaccine. Nat Med 2010; 16: 1389–1391.57 Epstein SL, Price GE. Cross-protective immunity to influenza A viruses. Expert Rev
Vaccines 2010; 9: 1325–1341.58 Stanekova Z, Vareckova E. Conserved epitopes of influenza A virus inducing protective
immunity and their prospects for universal vaccine development. Virol J 2010; 7: 351.59 Cassone A, Rappuoli R. Universal vaccines: shifting to one for many. MBio 2010; 1:
pii: e00042-10.60 Han T, Marasco WA. Structural basis of influenza virus neutralization. Ann NY Acad
Sci 2011; 1217: 178–190.61 WHO-Human-H5N1 Cumulative cases of H5N1 infections in humans reported to
the WHO up to the 8th April 2009. http://www.who.int/csr/disease/avian_influenza/country/en/2009.
62 Smallman-Raynor MC. A.D. avian influenza A (H5N1) age distribution in humans.
Emerg Infect Dis 2007; 13: 510–512.63 Epstein SL. Prior H1N1 influenza infection and susceptibility of Cleveland Family
Study participants during the H2N2 pandemic of 1957: an experiment of nature.J Infect Dis 2006; 193: 49–53.
64 Hancock K, Veguilla V, Lu X, Zhong W, Butler EN, Sun H et al. Cross-reactive antibody
responses to the 2009 pandemic H1N1 influenza virus. New Engl J Med 2009; 361:1945–1952.
65 Stelzer-Braid S, Wong B, Robertson P, Lynch GW, O’Brian K, Shaw R et al.
Cross-reactive influenza A H3N2 and H1N1 antibodies may give false positiveresults when using serological tests to determine human H5N1 infection. Clin J
Virol 2008; 42: 241–243.66 Lynch GW, Kapitza N, Fitzgerald A, Walsh B, Sullivan J. The anti-influenza specifi-
cities of intravenous immunoglobulins (IVIGs) for targeting human and avian influenza
antigens: comparisons of northern and Southern Hemisphere IVIGs. Austr Health MedRes Congr Proc Melbourne Victoria 2006; 5: 467.
67 Ekiert DC, Bhabha G, Elsliger MA, Friesen RH, Jongeneelen M, Throsby M et al.
Antibody recognition of a highly conserved influenza virus epitope. Science 2009;324: 246–251.
68 Sui J, Hwang WC, Perez S, Wei G, Aird D, Chen LM et al. Structural and functional
bases for broad-spectrum neutralization of avian and human influenza A viruses. NatStruct Mol Biol 2009; 16: 265–273.
69 Throsby M, van den Brink E, Jongeneelen M, Poon LL, Alard P, Cornelissen L et al.Heterosubtypic neutralizing monoclonal antibodies cross-protective against H5N1
and H1N1 recovered from human IgM+ memory B cells. PLoS One 2008; 3: e3942.70 Heiny AT, Miotto O, Srinivasan KN, Khan AM, Zhang GL, Brusic V et al. Evolutionarily
conserved protein sequences of influenza a viruses, avian and human, as vaccine
targets. PLoS ONE 2007; 2: e1190.71 Corti D, Suguitan Jr AL, Pinna D, Silacci C, Fernandez-Rodriguez BM, Vanzetta F et al.
Heterosubtypic neutralizing antibodies are produced by individuals immunized with a
seasonal influenza vaccine. J Clin Invest 2010; 120: 1663–1673.72 Okuno Y, Isegawa Y, Sasao F, Ueda S. A common neutralizing epitope conserved
between the hemagglutinins of influenza A virus H1 and H2 strains. J Virol 1993; 67:
2552–2558.73 Russell RJ, Gamblin SJ, Haire LF, Stevens DJ, Xiao B, Ha Y et al. H1 and H7 influenza
haemagglutinin structures extend a structural classification of haemagglutinin sub-
types. Virology 2004; 325: 287–296.74 Skehel JJ, Wiley DC. Receptor binding and membrane fusion in virus entry: the
influenza hemagglutinin. Annu Rev Biochem 2000; 69: 531–569.75 Wilson IA, Skehel JJ, Wiley DC. Structure of the haemagglutinin membrane glycopro-
tein of influenza virus at 3 A resolution. Nature 1981; 289: 366–373.76 Tsuchiya E, Sugawara K, Hongo S, Matsuzaki Y, Muraki Y, Li ZN et al. Antigenic
structure of the haemagglutinin of human influenza A/H2N2 virus. J Gen Virol 2001;
82: 2475–2484.77 Sandbulte MR, Jimenez GS, Boon AC, Smith LR, Treanor JJ, Webby RJ. Cross-reactive
neuraminidase antibodies afford partial protection against H5N1 in mice and are
present in unexposed humans. PLoS Med 2007; 4: e59.78 Boon AC, deBeauchamp J, Krauss S, Rubrum A, Webb AD, Webster RG et al. Cross-
reactive neutralizing antibodies directed against pandemic H1N1 2009 virus are
protective in a highly sensitive DBA/2 mouse influenza model. J Virol 2010; 84:7662–7667.
79 Cancro MP, Hao Y, Scholz JL, Riley RL, Frasca D, Dunn-Walters DK et al. B cells andaging: molecules and mechanisms. Trends Immunol 2009; 30: 313–318.
80 Panda A, Arjona A, Sapey E, Bai F, Fikrig E, Montgomery RR et al. Human innate
immunosenescence: causes and consequences for immunity in old age. TrendsImmunol 2009; 30: 325–333.
81 Canaday DH, Amponsah NA, Jones L, Tisch DJ, Hornick TR, Ramachandra L.
Influenza-induced production of interferon-alpha is defective in geriatric individuals.J Clin Immunol 2010; 30: 373–383.
Broad human antibody protection for pandemic influenzaGW Lynch et al
157
Immunology and Cell Biology

82 Rothberg MB, Haessler SD. Complications of seasonal and pandemic influenza. CritCare Med 2010; 38: e91–e97.
83 Poland GA, Ovsyannikova IG, Jacobson RM. Immunogenetics of seasonal influenzavaccine response. Vaccine 2008; 26 (Suppl 4): D35–D40.
84 Ahmed R, Oldstone MB, Palese P. Protective immunity and susceptibility toinfectious diseases: lessons from the 1918 influenza pandemic. Nat Immunol2007; 8: 1188–1193.
85 Rangel-Moreno J, Carragher DM, Misra RS, Kusser K, Hartson L, Moquin A et al. Bcells promote resistance to heterosubtypic strains of influenza via multiple mechan-isms. J Immunol 2008; 180: 454–463.
86 Ansaldi F, Zancolli M, Paolo D, Montomoli E, Sticchi L, Del Giudice G et al. Antibodyresponse against heterogeneous circulating influenza virus strains elicited by MF59-and non-adjuvanted vaccines during seasons with good or partial matching betweenvaccine strain and clinical isolates. Vaccine 2010; 28: 4123–4129.
87 Banzhoff A, Gasparini R, Laghi-Pasini F, Staniscia T, Durando P, Montomoli E et al.MF59-adjuvanted H5N1 vaccine induces immunologic memory and heterotypicantibody responses in non-elderly and elderly adults. PLoS One 2009; 4: e4384.
88 Galli G, Hancock K, Hoschler K, DeVos J, Praus M, Bardelli M et al. Fast rise of broadlycross-reactive antibodies after boosting long-lived human memory B cells primed byan MF59 adjuvanted prepandemic vaccine. Proc Natl Acad Sci USA 2009; 106:7962–7967.
89 Jegerlehner A, Schmitz N, Storni T, Bachmann MF. Influenza A vaccine based on theextracellular domain of M2: weak protection mediated via antibody-dependent NK cellactivity. J Immunol 2004; 172: 5598–5605.
90 Lamb RA, Kug RM. Orthomyxoviridae: The Viruses and Their Replication. In: KnipeDM et al. (eds). Fields Virology, 4th edn, Vol. 2. Lippincott Williams and Wilkins:Philadelphia, USA, 2002.
91 Neirynck S, Deroo T, Saelens X, Vanlandschoot P, Jou WM, Fiers W. A universalinfluenza A vaccine based on the extracellular domain of the M2 protein. Nat Med1999; 5: 1157–1163.
92 Fan J, Liang X, Horton MS, Perry HC, Citron MP, Heidecker GJ et al. Preclinical studyof influenza virus A M2 peptide conjugate vaccines in mice, ferrets, and rhesusmonkeys. Vaccine 2004; 22: 2993–3003.
93 Fu TM, Grimm KM, Citron MP, Freed DC, Fan J, Keller PM et al. Comparativeimmunogenicity evaluations of influenza A virus M2 peptide as recombinant virus likeparticle or conjugate vaccines in mice and monkeys. Vaccine 2009; 27: 1440–1447.
94 Rao SS, Kong WP, Wei CJ, Van Hoeven N, Gorres JP, Nason M et al. Comparativeefficacy of hemagglutinin, nucleoprotein, and matrix 2 protein gene-based vaccinationagainst H5N1 influenza in mouse and ferret. PLoS One 2010; 5: e9812.
95 Sui Z, Chen Q, Wu R, Zhang H, Zheng M, Wang H et al. Cross-protection againstinfluenza virus infection by intranasal administration of M2-based vaccine withchitosan as an adjuvant. Arch Virol 2010; 155: 535–544.
96 Wu F, Yuan XY, Li J, Chen YH. The co-administration of CpG-ODN influencedprotective activity of influenza M2e vaccine. Vaccine 2009; 27: 4320–4324.
97 Zhao G, Lin Y, Du L, Guan J, Sun S, Sui H et al. An M2e-based multiple antigenicpeptide vaccine protects mice from lethal challenge with divergent H5N1 influenzaviruses. Virol J 2010; 7: 9.
98 Zhao G, Sun S, Du L, Xiao W, Ru Z, Kou Z et al. An H5N1 M2e-based multipleantigenic peptide vaccine confers heterosubtypic protection from lethal infection withpandemic 2009 H1N1 virus. Virol J 2010; 7: 151.
99 Lamb RA, Zebedee SL, Richardson CD. Influenza virus M2 protein is an integralmembrane protein expressed on the infected-cell surface. Cell 1985; 40: 627–633.
100 Bui HH, Peters B, Assarsson E, Mbawuike I, Sette A. Ab and T cell epitopes ofinfluenza A virus, knowledge and opportunities. Proc Natl Acad Sci USA 2007; 104:246–251.
101 Furuse Y, Suzuki A, Kamigaki T, Oshitani H. Evolution of the M gene of the influenza Avirus in different host species: large-scale sequence analysis. Virol J 2009; 6: 67.
102 Furuse Y, Suzuki A, Oshitani H. Large-scale sequence analysis of M gene of influenzaA viruses from different species: mechanisms for emergence and spread of amanta-dine resistance. Antimicrob Agents Chemother 2009; 53: 4457–4463.
103 Gamblin SJ, Skehel JJ. Influenza haemagglutinin and neuraminidase membraneglycoproteins. J Biol Chem 2010; 285: 28403–28409.
104 Altstein AD, Gitelman AK, Smirnov YA, Piskareva LM, Zakharova LG, Pashvykina GVet al. Immunization with influenza A NP-expressing vaccinia virus recombinant
protects mice against experimental infection with human and avian influenza viruses.Arch Virol 2006; 151: 921–931.
105 Carragher DM, Kaminski DA, Moquin A, Hartson L, Randall TD. A novel role for non-neutralizing antibodies against nucleoprotein in facilitating resistance to influenzavirus. J Immunol 2008; 181: 4168–4176.
106 Epstein SL, Kong WP, Misplon JA, Lo CY, Tumpey TM, Xu L et al. Protection againstmultiple influenza A subtypes by vaccination with highly conserved nucleoprotein.Vaccine 2005; 23: 5404–5410.
107 Prokudina EN, Semenova NP. Localization of the influenza virus nucleoprotein:cell-associated and extracellular non-virion forms. J Gen Virol 1991; 72 (Pt 7):1699–1702.
108 Virelizier JL, Allison AC, Oxford JS, Schild GC. Early presence of ribonucleo-protein antigen on surface of influenza virus-infected cells. Nature 1977; 266:52–54.
109 Yewdell JW, Frank E, Gerhard W. Expression of influenza A virus internal antigens onthe surface of infected P815 cells. J Immunol 1981; 126: 1814–1819.
110 Zhirnov OP, Isaeva EI, Konakova TE, Thoidis G, Piskareva LM, Akopova II et al.Protection against mouse and avian influenza A strains via vaccination with acombination of conserved proteins NP, M1 and NS1. Influenza Other Respir Viruses2007; 1: 71–79.
111 Lamere MW, Moquin A, Lee FE, Misra RS, Blair PJ, Haynes L et al. Regulation of anti-nucleoprotein IgG by systemic vaccination and its effect on influenza virus clearance.J Virol 2011; 85: 5027–5035.
112 Lamere MW, Lam HT, Moquin A, Haynes L, Lund FE, Randall TD et al. Contributionsof antinucleoprotein IgG to heterosubtypic immunity against influenza virus. J Immunol2011; 186: 4331–4339.
113 Chen W, Zhu Z, Feng Y, Dimitrov DS. Human domain antibodies to conserved stericallyrestricted regions on gp120 as exceptionally potent cross-reactive HIV-1 neutralizers.Proc Natl Acad Sci USA 2008; 105: 17121–17126.
114 Khurana S, Suguitan Jr AL, Rivera Y, Simmons CP, Lanzavecchia A, Sallusto F et al.Antigenic fingerprinting of H5N1 avian influenza using convalescent sera and mono-clonal antibodies reveals potential vaccine and diagnostic targets. PLoS Med 2009; 6:e1000049.
115 Miotto O, Heiny A, Tan TW, August JT, Brusic V. Identification of human-to-humantransmissibility factors in PB2 proteins of influenza A by large-scale mutual informa-tion analysis. BMC Bioinformatics 2008; 9 (Suppl 1): S18.
116 Bautista E, Chotpitayasunondh T, Gao Z, Harper SA, Shaw M, Uyeki TM et al. Clinicalaspects of pandemic 2009 influenza A (H1N1) virus infection. N Engl J Med 2010;362: 1708–1719.
117 Steel J, Lowen AC, Wang T, Yondola M, Gao Q, Haye K et al. Influenza virusvaccine based on the conserved hemagglutinin stalk domain. MBio 2010; 1: pii:e00018-10.
118 Wei CJ, Boyington JC, McTamney PM, Kong WP, Pearce MB, Xu L et al. Induction ofbroadly neutralizing H1N1 influenza antibodies by vaccination. Science 2010; 329:1060–1064.
119 Sagawa H, Ohshima A, Kato I, Okuno Y, Isegawa Y. The immunological activity of adeletion mutant of influenza virus haemagglutinin lacking the globular region. J GenVirol 1996; 77: 1483–1487.
120 Laddy DJ, Yan J, Kutzler M, Kobasa D, Kobinger GP, Khan AS et al. Heterosubtypicprotection against pathogenic human and avian influenza viruses via in vivo electro-poration of synthetic consensus DNA antigens. PLoS One 2008; 3: e2517.
121 Stanekova Z, Kiraly J, Stropkovska A, Mikuskova T, Mucha V, Kostolansky F et al.Heterosubtypic protective immunity against influenza A virus induced by fusionpeptide of the hemagglutinin in comparison to ectodomain of M2 protein. Acta Virol2011; 55: 61–67.
122 Kwong PD, Wilson IA. HIV-1 and influenza antibodies: seeing antigens in new ways.Nat Immunol 2009; 10: 573–578.
123 Stamatatos L, Morris L, Burton DR, Mascola JR. Neutralizing antibodies generatedduring natural HIV-1 infection: good news for an HIV-1 vaccine? Nat Med 2009; 15:866–870.
124 Nguyen T, Davis CT, Stembridge W, Shu B, Balish A, Inui K et al. Characterization of ahighly pathogenic avian influenza H5N1 virus sublineage in poultry seized at ports ofentry into Vietnam. Virology 2009; 387: 250–256.
The Supplementary Information that accompanies this paper is available on the Immunology and Cell Biology website (http://www.nature.com/icb)
Broad human antibody protection for pandemic influenzaGW Lynch et al
158
Immunology and Cell Biology




















