The diversity of harmful algal blooms: a challenge for science and management

Estuarine, Coastal and Shelf Science 63 (2005) 285–300
www.elsevier.com/locate/ECSS
Seasonal variability of plankton blooms in the Ria de Ferrol(NW Spain): II. Plankton abundance, composition and biomass
Antonio Bode*, M. Teresa Alvarez-Ossorio, Nicolas Gonzalez,Jorge Lorenzo, Cristina Rodrıguez, Manuel Varela, Marta M. Varela
Instituto Espanol de Oceanografıa, Centro Oceanografico de A Coruna, Apdo. 130, E-15080 A Coruna, Spain
Received 16 August 2004; accepted 22 November 2004
Abstract
The abundance, taxonomic composition and biomass of plankton components were studied in the mostly eutrophic waters of the
Ria de Ferrol (Galicia, NW Spain) in contrasting seasons. Three stations arranged in a transect along the main ria axis were sampledduring cruises in February, May, July and September 2000. Phytoplankton, bacteria, micro- (40–200 mm) and mesozooplankton(O200 mm) compartments were considered. Phytoplankton blooms (O103 cel ml�1) and high total plankton biomass (up to 44 g Cm�2) was found at all seasons, except in winter when values were !1 g C m�2. Phytoplankton generally accounted for most of total
plankton biomass, particularly in late summer, thus driving most of plankton dynamics. The blooming species were always diatoms,either fast-growing, chain-forming species, well adapted to relatively turbulent conditions (e.g. Chaetoceros socialis), or disturbance-tolerant, estuarine adapted species (e.g. Skeletonema costatum). In addition, microflagellates (!10 mm) reached high abundances,
particularly during summer. The influence of shelf waters, where coastal upwelling events are frequent for most of the spring andsummer, prevents the establishment of a marked pycnocline and the dominance of dinoflagellates. Microheterotrophs (bacteria,protozoa and larval stages of metazoa) increased their abundance and biomass from winter to late summer, while mesozooplankton
peaked in spring and summer. Zooplankton dynamics were characterised by the presence of large numbers of larvae of bothplanktonic copepods and benthic metazoans, the latter mainly cirripeds and bivalve molluscs. The absence of a definite successionpattern in the mesozooplankton species abundance data, in contrast with phytoplankton data, along with the dominance of estuarinespecies (e.g.Acartia margalefi), suggest that mesozooplankton communities inside the ria behave differently from communities in shelf
waters. Despite its small size and reduced influence of upwelling compared with other Galician rias, the input of shelf waters in the Riade Ferrol drives massive phytoplankton blooms supporting a rich heterotrophic community.� 2004 Elsevier Ltd. All rights reserved.
Keywords: phytoplankton; zooplankton; bacteria; biomass; abundance; bloom; rias; NW Spain
1. Introduction
High biological production characterised the rias ofGalicia (NW Spain), coastal inlets under the influence ofa seasonal upwelling from March to October (Woosteret al., 1976; Fraga, 1981). Biogeochemical and hydro-dynamic processes explain the effective use of inorganic
* Corresponding author.
E-mail address: [email protected] (A. Bode).
0272-7714/$ - see front matter � 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.ecss.2004.11.021
nutrients, firstly as primary agents of phytoplanktongrowth inside the rias and secondly as regeneratednutrients from organic matter in shelf waters that are re-entered in the rias (Alvarez-Salgado et al., 1997, 2000).These characteristics contrast with those found in otherestuarine ecosystems with a larger influence of conti-nental waters (e.g. McClelland and Valiela, 1998). Theexcess of primary production supports both massivemussel and shellfish harvest inside the rias (Tenore et al.,1982) and fisheries over the shelf (Tenore et al., 1995;

286 A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
Porteiro et al., 1996), the latter driven by organic matterexported from the rias (Lopez-Jamar et al., 1992).
Most studies of plankton communities in the riaswere made in the large and highly productive RiasBaixas (e.g. Alvarez-Ossorio, 1977; Corral et al., 1981;Campos and Marino, 1984; Figueiras and Niell, 1987;Figueiras and Pazos, 1991; Figueiras and Rios, 1993;Nogueira et al., 2000; Tilstone et al., 2000), locatedsouth of Cape Finisterre (43 � N), while studies on thesmaller Rias Altas were scarce (e.g. Marino et al., 1985;Bode and Varela, 1998; Varela et al., 2001; Varela andPrego, 2003). Despite the lower nutrient inputs of RiasAltas compared with those of the Rias Baixas (Pregoet al., 1999), primary production rates are of similarmagnitude in both regions (Bode et al., 1996; Bode andVarela, 1998). Total primary production of the RiasBaixas is higher than in the Rias Altas mostly due to thelarger surface and volume of the former (Fraga, 1996).Previous studies in the Ria de Ferrol, one of the RiasAltas, showed phytoplankton biomass and primaryproduction rates in excess of values measured in thenearby shelf, thus suggesting enhanced productivity(Bode and Varela, 1998). Nevertheless, and in contrastto the Rias Baixas, upwelling waters were not detectedinside the Ria de Ferrol (Prego and Varela, 1998).
The objective of this paper is to characterise thedifferences in plankton abundance, taxonomic compo-sition and biomass in relation to seasonal successionpatterns typical of the Galician Rias. This study is partof a series of cruises made during the year 2000 tocharacterise plankton blooms in the Ria de Ferrol.Nutrient concentrations and plankton uptake rates wereanalysed in a companion paper (Bode et al., 2005), whileVarela et al. (2003) analysed the differential role ofbacteria and in situ sedimentation in the removal oforganic matter during the cruises.
2. Methods
Plankton samples were collected at three stationsrepresentative of outer, middle and inner ria zones(Fig. 1) during February, May, July and September.Hydrographic characteristics and in situ fluorescenceprofiles were measured with a CTD SeaBird SBE-25.Additional CTD observations were also taken at otherfour stations (Bode et al., 2005). In this paper, onlymeasurements of phytoplankton, bacteria and zoo-plankton abundance will be discussed.
2.1. Phytoplankton
Samples were collected with Niskin-type bottles at5–6 depths immediately after CTD casts. Chlorophyll-a(Chl-a) concentration was determined in acetonicextracts of plankton retained by Albet type-F filters.Extracts were kept cold (�20 �C) and dark overnightbefore measurement in a Turner Designs A-10 fluorom-eter (UNESCO, 1994). The values of in situ fluorescencemeasured along with CTD casts were converted to Chl-aby calibration with the acetonic extracts made atdiscrete depth levels. Phytoplankton species composi-tion was determined in Lugol’s preserved samplesobserved with an inverted microscope at magnificationpowers up to !1000 (Casas et al., 1999; Varela et al.,2004). Organisms too small for accurate identification atspecies or genus level by this method were grouped insize categories. In addition, naked ciliates were recorded.Phytoplankton species names were given accordingto Thomas (1997). Phytoplankton carbon biomass wasestimated from the biovolumes of individual cellsmeasured in an image analysis system and using theconversion equations in Strathman (1967), as described
8° 10' W
N
43° 30'study site
8° 4° 0°
36°
40°
44°
48°
Iberian Peninsula
St. 6
St. 4 St. 2
Fig. 1. Map of stations sampled for the study of plankton variables.

287A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
in Varela and Prego (2003) and Varela et al. (2004).Only autotrophic species of dinoflagellates (according toLarsen and Sournia, 1991 and references therein) wereincluded. The biomass of small flagellates was parti-tioned in autotrophic and heterotrophic components byassuming that heterotrophs were 10% of total carbonbiomass (Linley et al., 1983). In addition, protozoanbiomass was estimated in the same samples as the sumof the carbon biomass of heterotrophic flagellates(including heterotrophic dinoflagellates) plus ciliates.
2.2. Bacteria
Abundance of heterotrophic bacteria was determinedin samples preserved with glutaraldehyde (5% finalconcentration), stained with DAPI (4#-6 diamidino-2-phenylindole) and observed using an epifluorescencemicroscope, following the method of Porter and Freig(1980) as described in Varela et al. (2003). Bacterialcarbon was estimated from biovolumes measured withan image analysis system and the equations in Norlandet al. (1987).
2.3. Microzooplankton
Water column samples of microzooplankton (40–200 mm) were obtained with vertical hauls of a Bongo-type net of 40 mm mesh-size (20 cm diameter) equippedwith a flow meter from near the bottom to the surface.In addition, 10 L of water collected with a Niskin-typebottle at the depth of the chlorophyll maximum werefiltered through a 40 mm mesh. All samples werescreened through a nylon mesh of 200 mm and preservedwith borate-buffered formalin (5% final concentration)until examination under the stereoscopic microscope.Specimens were grouped in broad taxonomic categoriesas described in Valdes et al. (1991). Microzooplanktoncarbon biomass was estimated by multiplying theabundance of each group by the factors given in Beersand Stewart (1970).
2.4. Mesozooplankton
Samples were collected by means of double-obliquetows from the bottom to the surface of a Juday–Bogorov type net of 200 mm mesh-size (50 cm diameter)and equipped with flow and depth meters. At eachstation, samples were collected during the morning(before 12:00 GMT) and in the evening (approx. at17:00 GMT). Specimens for taxonomic analyses werepreserved as described for microzooplankton andexamined under the stereoscopic microscope. Identifi-cation was made at species level where possible. Inaddition, subsamples for biomass determinations werefiltered through glass-fibre Millipore Type-A filters and
stored frozen (�20 �C). In the laboratory, biomasssamples were dried (60 �C) and weighted to obtain thedry weight (DW). Subsamples of dry mesozooplanktonwere either calcinated in a furnace (450 �C) to obtain theash-free dry weight (AFDW) or fed into a Perkin ElmerCNH analyser to directly measure the carbon content(Bode et al., 1998).
2.5. Statistical analysis
The variability of species composition of phyto- andmesozooplankton was summarised by means of princi-pal component analysis (PCA) performed on the log-transformed abundance data (e.g. Fernandez and Bode,1994). Separate PCA were made for phytoplankton andfor mesozooplankton species after selection of the mostabundant taxa (i.e. those with frequencies higher than10% and 2% of total sample abundance for phyto- andmesozooplankton, respectively). For these analyses thestatistical software SPSS was employed.
3. Results
3.1. Water column structure and phytoplankton blooms
Two different types of vertical structure in the watercolumn were found (Fig. 2). In February, the watercolumn was thoroughly mixed at all stations, asevidenced by the homogeneity in the profiles oftemperature and salinity. The influence of water ofcontinental origin was minimal and the thermohalinecharacteristics of the water were those typical of shelfwater. The concentration of Chl-a reached the minimumvalues of the study (!2 mg Chl-a m�3). In all othercruises, a surface layer with water of lower salinity andhigher temperature than shelf water developed, beingmore marked at the innermost station (St. 2). Suchstructure caused a pycnocline associated with highchlorophyll concentrations forming a distinct subsurfacemaximum. The position of the Chl-a maximum deep-ened from the inner to the outer station and from Mayto September. In addition, the highest Chl-a values (upto 19 mg m�3) were always found at the innermoststation and increased from May to September (Fig. 2).In all cruises the influence of shelf water was dominantover continental sources at all stations sampled,therefore most of the nutrient inputs for phytoplanktongrowth came from the shelf (Bode et al., 2005).
3.2. Phytoplankton
Except in February, phytoplankton abundance washigh at all stations, with average values O105 cel ml�1
in May and September and O3!103 cel ml�1 in July.Small flagellates were the most abundant cells but

288 A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
February
May
July
September
Temperature (°C)
St. 2
St. 4St. 6
10 12 14 16 18 20 24 28 32 36 0 5 10 15 20
10 12 14 16 18 20 24 28 32 36 0 5 10 15 20
10 12 14 16 18 20 24 28 32 36 0 5 10 15 20
10 12 14 16 18 20 24 28 32 36 0 5 10 15 20
Dep
th (m
)D
epth
(m)
Dep
th (m
)D
epth
(m)
Salinity Chlorophyll-a (mg m-3)
Fig. 2. Vertical profiles of temperature ( �C), salinity and chlorophyll-a (mg m�3) measured by a CTD profiler with attached fluorometer at the
sampling stations near high tide in February, May, July and September 2000.
chain-forming diatoms of genera Chaetoceros, Pseudo-nitzschia, Leptocylindrus and Thalassiosira generallyexceeded 103 cel ml�1 (Table 1). During summer, therewas a relative increase of dinoflagellates, as Heterocapsaniei and species of Gymmnodinium and Gyrodinium buttheir abundances were generally!102 cel ml�1.
The PCA on the species with abundance valuesO10% of total sample abundance produced two firstfactors accounting for ca. 50% of variance (Fig. 3). Thefirst factor separated species typical of the July bloom(e.g. Thalassionema nitzschioides, Asterionellopsis glacia-lis and Protoperidinium steinii) from those characteristic

289A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
Table 1
Mean (GSE) abundance (cel ml�1) of phytoplankton species representing O10% of total abundance in the samples
Group Code n Mean SE
Dinophyceae (dinoflagellates)
Heterocapsa niei (Loeblich) Morril and Loeblich III HETNIE 29 23.0 8.4
Ceratium lineatum (Ehrenberg) Cleve CERLIN 17 4.1 1.5
Dinophysis acuminata Claparede and Lachmann DINACU 14 0.9 0.5
Gyrodinium spirale (Bergh) Kofoid and Swezy GYRSPI 26 1.0 0.2
Protoperidinium bipes (Paulsen) Balech PROBIP 15 1.0 0.3
Protoperidinium diabolus (Paulsen) Balech PRODIA 15 0.4 0.1
Protoperidinium steinii (Jorgensen) Balech PROSTE 16 0.8 0.2
Scrippsiella trochoidea (Stein) Loeblich SCRTRO 18 1.4 0.3
Bacillarophycea (diatoms)
Asterionellopsis glacialis (Castracane) Round ASTGLA 18 34.4 11.2
Chaetoceros danicus Cleve CHADAN 17 1.4 0.3
Chaetoceros didymus Ehrenberg CHADID 21 50.8 37.2
Chaetoceros socialis Lauder CHASOC 21 3297.0 1388.7
Leptocylindrus danicus Cleve LEPDAN 33 2293.6 656.7
Leptocylindrus minimus Gran LEPMIN 13 3.7 1.6
Nitzschia longissima (Brebisson in Kutzing) Grunow NITLON 53 16.2 2.6
Nitzschia longissima (small forms) NITLONP 34 1873.0 640.6
Pseudo-nitzschia cf delicatissima (Cleve) Heiden PSEDEL 21 8.6 2.0
Pseudo-nitzschia cf pungens (Grunow ex Cleve) Hasle PSEPUN 32 3.4 0.9
Proboscia alata (Brightwell) Sundstrom RHIALA 17 1.9 0.6
Guinardia delicatula (Cleve) Hasle RHIDEL 30 2.6 0.4
Rhizosolenia setigera Brightwell RHISET 25 1.4 0.3
Guinardia striata (Stolterforth) Hasle RHISTO 18 1.2 0.4
Skeletonema costatum (Greville) Cleve SKECOSG 27 53.1 15.9
Skeletonema costatum (small forms) SKECOSP 13 278.0 97.9
Thalassionema nitzschioides (Grunow) Van Heurck THANIT 35 27.9 7.4
Other groups
Solenicola setigera Pavillard SOLSET 11 3.7 1.7
Dictyocha speculum Ehrenberg DICSPE 17 0.8 0.2
Cryptomonas spp. CRYPTO 54 201.6 23.9
Other flagellates
Monads O10 mm 58 146.4 15.5
Monads 5–8 mm 30 477.3 47.7
Monads 3–5 mm 30 3875.9 455.1
Monads !3 mm 30 6340.3 650.4
Ciliates
Ciliates O100 mm 37 0.4 0.1
Ciliates 30–100 mm 51 5.0 0.7
Ciliates !30 mm 36 7.1 1.2
These species were employed in the PCA on phytoplankton species and the codes employed in Fig. 3 are listed. n, number of samples where each
species was found. The abundance values of unidentified flagellates and ciliates (not used in the PCA) are also given.
of May and September (e.g. Ceratium lineatum, Nitz-schia longissima and Leptocylindrus danicus). Betweenboth groups of species were other present in both Mayand July samples, as Heterocapsa niei, Chaetocerossocialis and Scrippsiella trochoidea. Therefore, the firstPCA factor can be interpreted as a measure of thespecies that dominate phytoplankton abundance inblooms during spring and summer (Fig. 4a). The secondfactor allows for the separation of samples obtained inSeptember, characterised by relatively high abundancesof species like Nitszchia longissima, Leptocylindrusdanicus and Skeletonema costatum, as shown by theplot of samples in the space of the two factors (Fig. 4b).
The species composition of samples from the cruise inFebruary was not very different from those of May,being the main difference their absolute abundancevalues. In contrast, the bloom observed in July hada phytoplankton composition clearly different fromthose observed in May and September (Fig. 4a), whichhad relatively higher cell abundances. Thus, the seasonalcycle of phytoplankton was characterised by the differ-ences in the relative dominance of diatom species in theassemblages, while dinoflagellates and other phyto-plankton had relatively low importance. Most of thevariability in phytoplankton composition was due to thevertical stratification observed at stations 2 and 4,

290 A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
particularly in July, which translated in a large disper-sion of samples in the factorial space (Fig. 4b). Samplesfrom the outer ria St. 6 were comparatively less variable.
3.3. Bacteria
The variations in the abundance of heterotrophicbacteria were lower than those in Chl-a and phyto-plankton abundance. Values ranged from 0.3 to1.8!106 cel ml�1, being the lowest values fromFebruary and the highest from September samples.Their vertical distribution generally peaked at thesurface and below the subsurface Chl-a maximum(Fig. 5) except at the inner ria station in July, whenthe vertical distribution of abundance clearly followedthe Chl-a profile. In contrast, bacterial abundanceprofiles at the outer ria station were more homogeneousthan those measured at the other stations, except inSeptember (Fig. 5), when maximum differences betweensubsurface (5 m depth) and deep samples (25 m) werefound.
3.4. Microzooplankton
The abundance of microzooplankton was generallyhigher inside the ria compared to the outer ria station(Fig. 6). Both water column integrated (net) samples andsamples from the subsurface Chl-a maximum (bottle)were similar. The only exceptions were the samples fromSeptember, when there were differences between theabundance recorded at the Chl-a maximum and theabundance averaged in the water column, notably atthe middle ria station. These differences were due to the
-1.0
-0.6
-0.2
0.2
0.6
1.0
Factor 1
Fact
or 2
NITLONGP
LEPDANSKECOSP
SOLSET
SKECOSG
PROSTE
THANIT
PRODIA
CHASOCSCRTRO
ASTGLA
DICSPE
CERLIN
DINACU
CHADID
-1.0 -0.6 -0.2 0.2 0.6 1.0
Fig. 3. Distribution of phytoplankton species along the first two
factors of the PCA performed on phytoplankton species abundances.
The species showing the highest correlation values with both factors
are indicated (see species codes in Table 1).
accumulation of microzooplankton organisms near theChl-amaximum at the inner and outer ria stations but atother depths levels in the case of the middle ria station(St. 4).
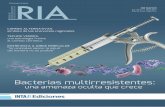
Copepods, mostly nauplii and juvenile stages, dom-inated the abundance distributions (Fig. 7), reaching upto 94% of total microzooplankton abundance inFebruary. The contribution of meroplankton increasedduring May and July, especially at the inner ria station,and in September this group was dominant at the outerria station. Larvae of benthic molluscs and polychaetsformed the bulk of meroplankton in May and Septem-ber, while in July nauplii of cirripeds reached mean
-3
-2
-1
0
1
2
3
February
May
July
September
-3
-2
-1
0
1
2
3
Fact
or 2
Fact
or 2
St. 2
St. 4St. 6
a
b
Mid-summer
Winter & Spring
Late-summer
Factor 1-3 -2 -1 0 1 2 3
-3 -2 -1 0 1 2 3Factor 1
Fig. 4. Distribution of samples in the space of the two first factors of
the PCA performed on phytoplankton species abundance values. The
samples are coded according to sampling date (a) or station (b). The
seasonal succession of phytoplankton assemblages is indicated by
arrows linking the main seasons.

291A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
0
5
10
15
20
25
0.0
0
5
10
15
20
25
0
5
10
15
20
25
0
5
10
15
20
25
Bacterial abundance (x106 cel ml-1)
February
May
July
September
St. 2St. 4
St. 6
0.5 1.0 1.5 2.0
0.0 0.5 1.0 1.5 2.0
0.0 0.5 1.0 1.5 2.0
0.0 0.5 1.0 1.5 2.0
Dep
th (m
)D
epth
(m)
Dep
th (m
)D
epth
(m)
Fig. 5. Vertical distribution of bacterial abundance (!106 cel ml�1) at
the sampling stations in February, May, July and September.
Fig. 6. Abundance of microzooplankton (40–200 mm, indiv. L�1) in net
(water column integrated) and bottle samples (collected at the
chlorophyll maximum) at the sampling stations in February, May,
July and September. Net samples were not collected for the February
cruise.
292 A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
abundances O40 indiv. L�1, notably at the innermoststation. Cirriped nauplii also dominated microzoo-plankton abundance at the outer ria station inSeptember. Other noticeable microzooplankton taxawere appendicularia, always present at all stationsbut notably in July at St. 2 with peak abundances of35 indiv. L�1.
3.5. Mesozooplankton
Mesozooplankton biomass increased from Februaryto July cruises, but decreased in September (Table 2).The largest range was observed at the deepest station(St. 6). Samples collected in the evening generallyshowed higher biomass values than those collected inthe morning, except in February. The mean differencebetween morning and evening biomass values was27.1% (GSE 7.1%) of their average in the case ofDW, and 28.6% (G6.7%) in the case of AFDW. Suchdifference, however, amounted only 5.6% (G1.1%) inthe case of the AFDW/DW ratio. None of these dif-ferences was significant (Mann–Whitney test, PO0:5,
Table 2
Mesozooplankton (O200 mm) biomass (mg m�3) at the sampling
stations measured in the morning (before 12:00 GMT) and in the
evening (near 17:00 GMT) during the study
Cruise Station Sample Depth DW AFDW AFDW/DW
February 2 Morning 7 26.9 13.2 49.0
Evening 8 25.7 11.2 43.6
4 Morning 11 43.7 16.2 37.2
Evening 17 31.1 10.9 35.2
6 Morning 22 12.9 6.5 50.0
Evening 22 11.7 6.1 52.0
May 2 Morning 7 58.3 45.8 78.7
Evening 7 66.5 51.0 76.6
4 Morning 23 23.9 19.3 80.6
Evening 27 47.2 36.4 77.1
6 Morning 17 49.5 35.9 72.5
Evening 23 61.4 49.9 81.3
July 2 Morning 7 11.8 8.5 72.6
Evening 7 13.0 9.3 71.6
4 Morning 7 21.4 14.9 69.8
Evening 13 43.1 26.9 62.3
6 Morning 20 57.3 38.9 68.0
Evening 20 107.3 76.4 71.2
September 2 Morning 9 15.7 12.6 80.4
Evening 10 21.3 18.1 85.1
4 Morning 19 10.8 8.5 78.7
Evening 14 10.7 8.7 81.3
6 Morning 27 22.0 17.7 80.8
Evening 20 23.7 19.3 81.4
DW, dry weight; AFDW, ash-free dry weight. Depth: sampling depth
in metres. The relative content of ash-free dry matter (AFDW/DW,%)
is also given.
Fig. 7. Taxonomic group composition of microzooplankton (40–
200 mm) net samples at stations 6, 4 and 2 in February, May, July and
September.
293A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
nZ24), indicating that, on average, both morning andevening samples contained the same type of organisms.This was further confirmed by the taxonomic determi-nation of mesozooplankton samples.
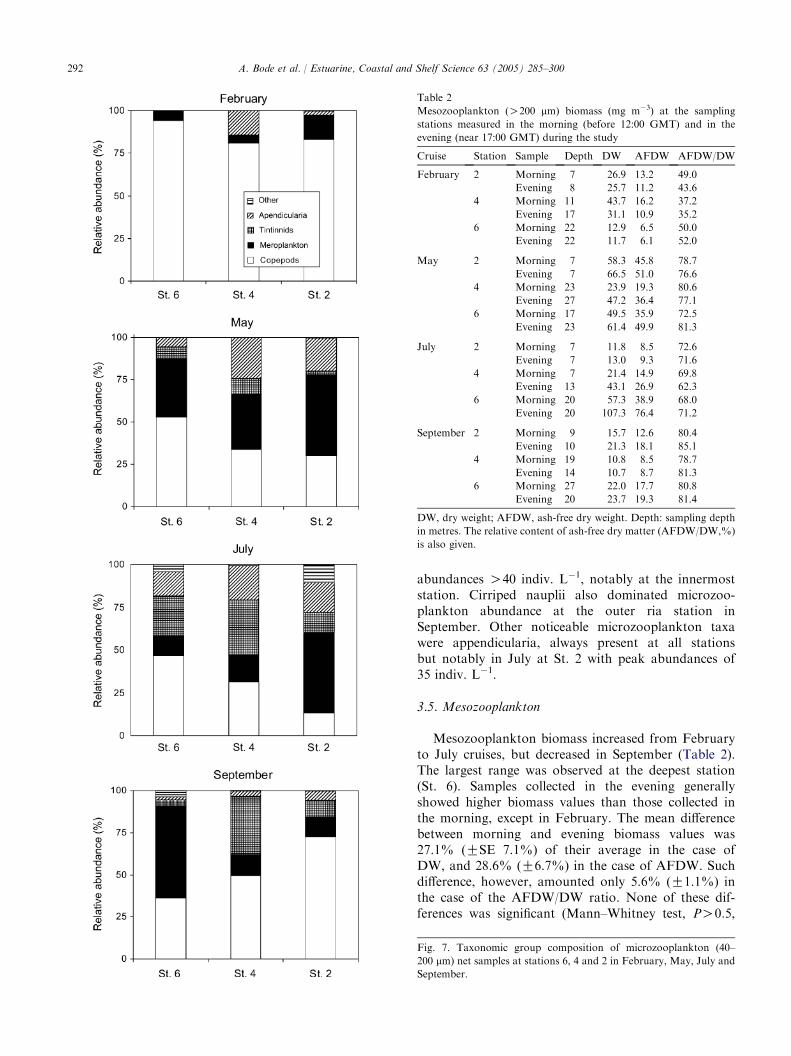
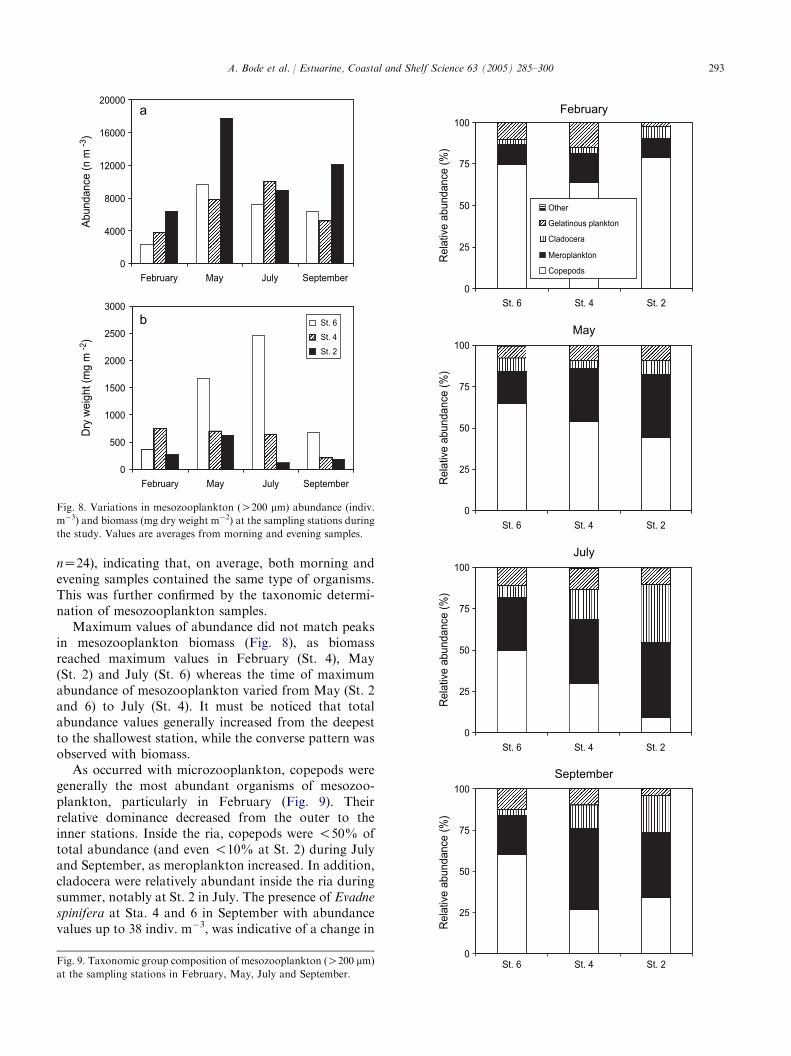
Maximum values of abundance did not match peaksin mesozooplankton biomass (Fig. 8), as biomassreached maximum values in February (St. 4), May(St. 2) and July (St. 6) whereas the time of maximumabundance of mesozooplankton varied from May (St. 2and 6) to July (St. 4). It must be noticed that totalabundance values generally increased from the deepestto the shallowest station, while the converse pattern wasobserved with biomass.
As occurred with microzooplankton, copepods weregenerally the most abundant organisms of mesozoo-plankton, particularly in February (Fig. 9). Theirrelative dominance decreased from the outer to theinner stations. Inside the ria, copepods were !50% oftotal abundance (and even !10% at St. 2) during Julyand September, as meroplankton increased. In addition,cladocera were relatively abundant inside the ria duringsummer, notably at St. 2 in July. The presence of Evadnespinifera at Sta. 4 and 6 in September with abundancevalues up to 38 indiv. m�3, was indicative of a change in
4000
8000
12000
16000
20000
500
1000
1500
2000
2500
3000St. 6St. 4St. 2
a
0
Abun
danc
e (n
m -3
)
0February May July September
February May July September
Dry
weig
ht (m
g m
-2)
b
Fig. 8. Variations in mesozooplankton (O200 mm) abundance (indiv.
m�3) and biomass (mg dry weight m�2) at the sampling stations during
the study. Values are averages from morning and evening samples.
Fig. 9. Taxonomic group composition of mesozooplankton (O200 mm)
at the sampling stations in February, May, July and September.
0
0
25
50
75
100
0
25
50
75
100
25
50
September
St. 6 St. 4 St. 2
St. 6 St. 4 St. 2
St. 6 St. 4 St. 2
St. 6 St. 4 St. 2
Rela
tive
abun
danc
e (%
)Re
lativ
e ab
unda
nce
(%)
Rela
tive
abun
danc
e (%
)
0
25
50
75
100
Rela
tive
abun
danc
e (%
)
Other
Gelatinous plankton
Cladocera
Meroplankton
Copepods
75
100
February
May
July

294 A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
the type of shelf waters entering the ria (Bode et al.,2005), as this cladoceran is characteristic of warm waters(Fernandez de Puelles et al., 2004). Appendicularia werealways present (with abundance values of up to 1697indiv. m�3) and other gelatinous taxa, as medusae andsalps, showed up occasionally.
The PCA on mesozooplankton abundance of groupscontributing O2% to total sample abundance producedthree main factors contributing to 76% of total variance(33, 29 and 14% for factors 1, 2 and 3, respectively). Forthis analysis, both morning and evening samples wereemployed. The first factor was related to high abun-dances of the cladoceran Evadne nordmanni, meroplank-tonic larvae (mostly gastropods and nauplii ofcirripeds), and to the copepod Pseudocalanus elongatus(Fig. 10). The second factor was positively correlated to
-0.5
0.0
0.5
1.0
0.0 0.5 1.0
COPDI ZPISI
-1.0
-0.5
0.0
0.5
1.0
-1.0-1.0 -0.5
-0.5Factor 1
Fact
or 2
Fact
or 3
PSEEL PARPA
ACACA
LLAME
ACACJAPEND
LGAST
NCIRR
POPOLACAMA
0.0 0.5 1.0Factor 2
PSEEL
COPDIZPISI
PARPA
ACACA
LLAME
ACACJ
APEND
LGAST
EVNOR
EVNOR
NCIRR
POPOL
ACAMA
a
b
Fig. 10. Distribution of mesozooplankton species in the spaces defined
by the first three factors of the PCA performed on mesozooplankton
species abundance values. The species codes appear in Table 3.
the abundance of mainly copepodites and zoean larvaeof the benthic decapod Pisidia longicornis, and nega-tively correlated with typical estuarine species, such asthe cladoceran Podon polyphemoides and the copepodAcartia margalefi. The third factor allowed for theseparation of samples with estuarine species from thosewith high abundance of mollusc larvae (mostly frombenthic bivalves). The distribution of samples in thefactorial spaces showed a temporal pattern characterisedby the end members in February, with low abundance ofall groups, and in May, with maximum abundances ofcopepodites and small adult copepods and meroplank-tonic larvae (Fig. 11). Still, the reconstruction ofa seasonal succession was not so evident as in the caseof phytoplankton composition. According to the valuesof the second and third factors, the influence ofcontinental water on mesozooplankton compositionwas maximal at the innermost station, where peaksin the abundance of estuarine species (up to 2375indiv. m�3 of Acartia margalefi in February) alternatedwith peaks in meroplanktonic larvae (up to 818 indiv.m�3 of bivalve larvae in September). Such alternationcaused that, as for the PCA on phytoplankton compo-sition (Fig. 4), the largest range of variability of meso-zooplankton composition occurred at St. 2 (Fig. 11).
Table 3
Mean, maximum and minimum abundance (indiv. m�3) of the
mesozooplankton (O200 mm) taxa contributing O2% to total sample
abundance
Taxa Code n Mean Min. Max.
Copepoda
Paracalanus parvus
(Claus, 1863)
PARPA 24 217 13 645
Pseudocalanus elongatus
(Boeck, 1864)
PSEEL 17 335 4 1241
Acartia clausi (Giesbretch,
1889)
ACACA 24 649 10 2310
Acartia clausi (young) ACACJ 24 689 31 2263
Acartia margalefi Alcaraz,
1976
ACAMA 15 583 4 2375
Copepodites COPDI 24 754 56 6585
Cladocera
Evadne nordmanni
Loven, 1835
EVNOR 24 510 21 2057
Podon polyphemoides
(Leuckart, 1859)
POPOL 7 1471 189 3354
Apendicularia APEND 24 507 59 1697
Meroplankton
Gastropod larvae LGAST 24 250 19 853
Bivalve larvae LLAME 22 293 5 1194
Cirriped nauplii NCIRR 24 1616 84 4387
Pisidia longicornis zoeae ZPISI 23 215 3 875
These taxa were employed in the PCA on mesozooplankton and the
codes employed in Fig. 10 are listed. n: number of samples where each
taxon was found.

295A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
-3
-2
-1
0
1
2
3
-3 -2 -1 0 1 2
a
Factor 1
Fact
or 2
FebruaryMay
JulySeptember
b
-3 -2 -1 0 1 2Factor 2
-3
-2
-1
0
1
2
3
Fact
or 3
-3 -2 -1 0 1 2Factor 1
-3
-2
-1
0
1
2
3
Fact
or 2
c
-3 -2 -1 0 1 2Factor 2
-3
-2
-1
0
1
2
3
Fact
or 3
d
St. 2St. 4St. 6
Fig. 11. Distribution of samples in the spaces defined by the first three factors of the PCA performed on mesozooplankton species abundance values.
The samples are coded according to sampling date (a,b) or station (c,d).
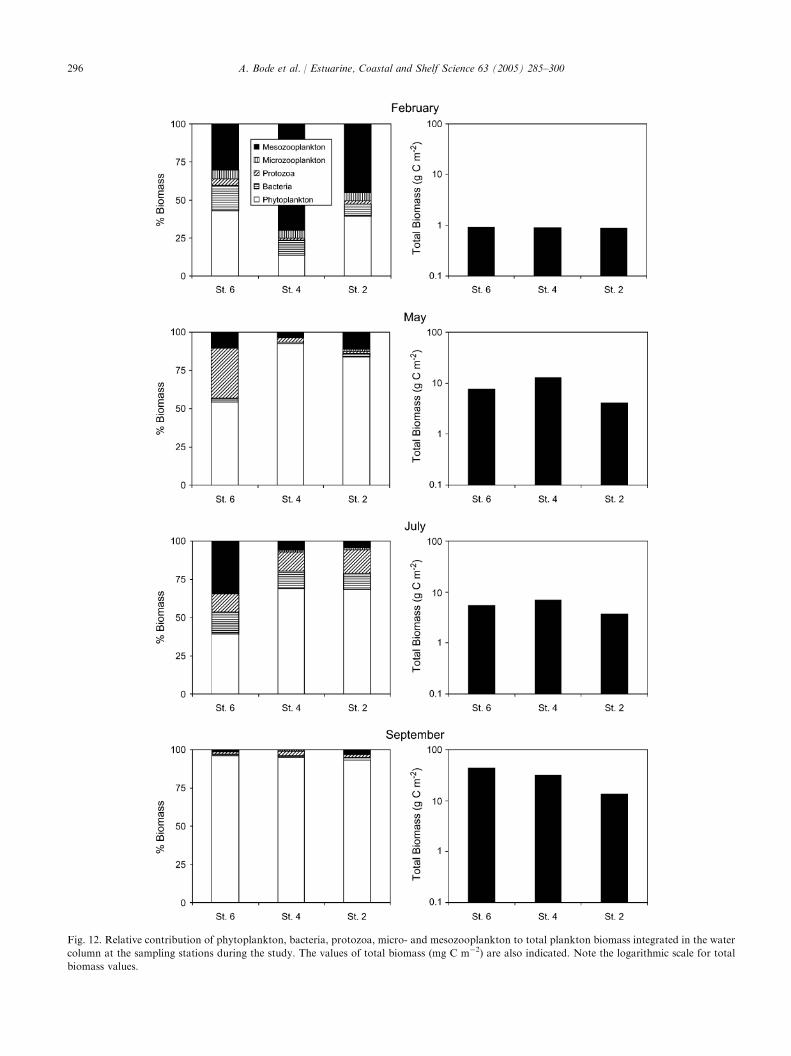
3.6. Total plankton biomass
Total plankton biomass was computed as the sum ofthe estimates made for the plankton compartmentsconsidered (Fig. 12). Large seasonal changes in plank-ton biomass were evident when comparing the differentcruises, with values ranging from !1 g C m�2 inFebruary to O10 g C m�2 (with a maximum of 44 g Cm�2 at St. 6) in September. Compared to such temporalchanges the variability in biomass between stations wasrelatively low, despite the differences in depth. Only inSeptember a clear trend of biomass decrease with watercolumn depth was found. Phytoplankton generallyformed the bulk of plankton biomass during blooms,particularly inside the ria (Fig. 12). The dominance ofphytoplankton biomass was maximal in Septemberwhen it was O90% of total plankton carbon, butheterotrophic carbon exceeded phytoplankton carbon inFebruary at all stations and at St. 6 in July. In addition,mesozooplankton was the main fraction of planktoncarbon in February at St. 4.
4. Discussion
4.1. Seasonal phytoplankton succession
The present study shows that massive phytoplanktonblooms (i.e. O104 cel ml�1 and O10 mg Chl-a m�3) are
a typical characteristic of the Ria de Ferrol during mostof the year, thus confirming previous observations(Bode and Varela, 1998). Only during winter phyto-plankton abundance and biomass reached low values,although the species composition at that time of the yearwas similar to that producing blooms in spring. Theseresults contrast with the observations of phytoplanktonin other rias of Galicia and adjacent shelf waters, wherethe dynamics of phytoplankton were characterised by analternation of periods of high and low abundance andbiomass (e.g. Casas et al., 1999 and references therein).In any case, and despite the reduced number ofsituations along the annual cycle considered in thisstudy, a seasonal succession of phytoplankton commu-nities in the Ria de Ferrol can be reconstructed.
Spring blooms were formed by typically chain-forming, but of small individual cell-size diatom species,as Chaetoceros socialis. Other species, as the dinofla-gellates Heterocapsa niei and Scripsiella trochoidea alsoreached relatively high abundances. The former speciesare typical competitors that grow easily even inrelatively well-mixed waters (e.g. Casas et al., 1999;Nogueira et al., 2000) showing high abundance andbiomass values. The latter species can be considered asruderal or disturbance-tolerant and typical of conditionswhere physical disturbances (i.e. upwelling or runoff)were moderate and allowed for phytoplankton growth(Nogueira et al., 2000). This would explain theirpersistence in the community for most of spring and
296 A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
Fig. 12. Relative contribution of phytoplankton, bacteria, protozoa, micro- and mesozooplankton to total plankton biomass integrated in the water
column at the sampling stations during the study. The values of total biomass (mg C m�2) are also indicated. Note the logarithmic scale for total
biomass values.

297A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
summer, since the main physical factor driving phyto-plankton succession in the Galician rias is the seasonalupwelling, although its influence is more marked in thesouthern Rias Baixas (Figueiras and Niell, 1987;Figueiras and Pazos, 1991; Figueiras and Rios, 1993;Nogueira et al., 2000; Tilstone et al., 2000; Varela et al.,2004) than in the northern Rias Altas (Casas et al., 1999;Varela et al., 2001; Varela and Prego, 2003). In the caseof the Ria de Ferrol, ruderal phytoplankton specieswould benefit from the large influence of shelf water,even when the direct influence of the upwelling was low(Bode et al., 2005). In this way, upwelling events werefrequent in the nearby shelf during spring and summer(Prego and Varela, 1998; Casas et al., 1999; Prego et al.,1999; Varela et al., 2001). Ruderal species, bothdinoflagellates and diatoms (e.g. Thalassionema nitz-schioides), would dominate mid-summer blooms, as theone observed in July, because of the disturbanceproduced by upwelling events in the shelf. Thesedisturbances would also explain the absence of thetypical phytoplankton community with a significantdominance of dinoflagellates characteristic of stratifiedwaters during summer in other rias and shelf waters(Figueiras and Pazos, 1991; Fernandez and Bode, 1994;Casas et al., 1999). As nutrients were never completelyexhausted, even in the shallow, stratified surface layer(Bode et al., 2005), severe stress caused by nutrientlimitation was not likely, in contrast to the situationfound in other rias (Nogueira et al., 2000). Stress-tolerant species were noticeable during summer, butsome of them, as those of the genus Protoperidinium,were reported as heterotrophic (e.g. Olseng et al., 2002).The resulting community would be well adapted torelatively high nutrient concentrations and significantlydependent on nitrate, as indicated by the values of the fratio O0.5 (Bode et al., 2005).
Late-summer blooms, as the one observed inSeptember, were also characterised by competitor fast-growing diatoms, but in this case the dominant specieswere Leptocylindrus danicus and Skeletonema costatum,virtually absent in winter and spring but appearing insummer samples. Almost the same phytoplanktoncommunity was found in a previous study of the Ria deFerrol in August 1993 (Bode and Varela, 1998).Nogueira et al. (2000) described a similar communityas responsible of late-summer and autumn blooms in theRia de Vigo (the southernmost Ria Baixa), which wereattributed to the high population growth rate of thesmall-size cells, typical r-strategists, and to the existenceof an initial inoculum of these species from previousblooms in the ria. In this way, Tilstone et al. (2000)showed a succession of spring blooms of contrastingcommunities formed either by Skeletonema–Thalassios-sira or by Chaetoceros–Cerataulina assemblages. Thedominance of one or another community was explainedby their different sensitivity to hydrographic versus
biogeochemical factors, as the former community wasdependent on the availability of silicate whereas thelatter was well adapted to strong upwelling conditions.In the Ria de Ferrol, however, Skeletonema andLeptocylindrus were almost absent of winter and springsamples, and only increased during summer until theybloom. This pattern was also found in the nearby Bay ofA Coruna and adjacent shelf waters (Casas et al., 1999;Varela et al., 2001; Varela and Prego, 2003). Inputs ofsilicate were not noticeable during summer (Bode et al.,2005) but the low direct influence of upwelling inside theRia de Ferrol may favour the growth of these speciesthat will outcompete the fast-growing Chaetocerosspecies, that are adapted to more turbulent conditions,as those found in spring.
4.2. Heterotrophic community
Phytoplankton blooms appear as the main drivingforce for changes in the whole planktonic communityin the Ria de Ferrol. Abundance and production ofheterotrophic bacteria were significantly correlated withChl-a and temperature (Varela et al., 2003). Thecoupling between bacteria and phytoplankton was moreevident inside the ria, where there was a higher matchbetween vertical profiles of bacterial abundance andChl-a than at the outer ria station. In addition, bacterialproduction measured concurrently with the presentstudy was high inside the ria (Varela et al., 2003) butit only consumed !10% of the available dissolvedorganic carbon, and most of the organic carbonproduced by the phytoplankton blooms was lost assedimented particles. Other studies in Galician riasshowed a rapid, but generally delayed, response ofplanktonic bacteria to phytoplankton blooms caused bythe upwelling (Hanson et al., 1986; Zdanowski andFigueiras, 1997). In the case of the Ria de Ferrol, therelatively high temperature of the water inside the riaduring summer would explain that the response ofbacteria to phytoplankton blooms during this seasonwould be higher that the response in spring (Varelaet al., 2003). Their main biogeochemical role, however,would be through the remineralisation of nutrients, asammonium (Bode et al., 2005), through microbial foodwebs. Thus bacteria would contribute to the support ofphytoplankton blooms inside the ria when the nitratestock, originated from upwelling in the shelf, is reduced.
Other heterotrophs also displayed a seasonal re-lationship with phytoplankton blooms in the Ria deFerrol. Protozoa (including heterotrophic dinoflagel-lates) increased during spring and summer, althoughtheir contribution to total plankton biomass wasgenerally small. There are relatively few reports of theabundance of this group in Galician waters. Most of thepublished information refers to unidentified flagellatesand ciliates observed with the inverted microscope along

298 A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
with phytoplankton (e.g. Figueiras and Pazos, 1991;Bode and Varela, 1994; Bode et al., 1996; Casas et al.,1999; Fileman and Burkill, 2001; Varela et al., 2001;Varela and Prego, 2003). Bode et al. (2004) showedthe increase in the abundance of heterotrophic flag-ellates (i.e. non pigmented when observed with anepifluorescence microscope) and planktonic ciliates fromspring to summer in shelf waters near the Ria de Ferrol.Despite its low biomass, relative to other components ofplankton in the ria, this group deserves future detailedstudies because of their likely participation in theremineralisation of the organic matter produced by thephytoplankton (Nagata, 2000). Fileman and Burkill(2001) noted that estimates of ciliate carbon biomassmade for the shelf waters off the Rias Baixas duringoligotrophic conditions in summer were almost tenfoldlower than those made for the shelf off the Rias Altas byBode and Varela (1994), although estimates for upwell-ing conditions near the coast were similar in bothstudies. A high biomass of ciliates is indicative ofa significant activity of microbial food webs, as ciliatesgenerally dominate the biomass distribution of micro-heterotrophs in coastal waters (Dolan, 1991; Bode andVarela, 1994; Quevedo and Anadon, 2000; Fileman andBurkill, 2001; Bode et al., 2004). In this way, Filemanand Burkill (2001) showed a rapid increase of ciliatestocks in 3 days during an upwelling-relaxation cycle.
The microzooplankton component considered in thisstudy (O40 mm) was mostly composed of metazoa,larval stages of copepods and molluscs, but ciliatetintinnids constituted up to 30% of total abundanceduring summer. Despite some studies concluded thatthis component could account for most of zooplanktongrazing on phytoplankton in Galician shelf waters(Braun et al., 1990), only a few studies described itstaxonomic composition (Valdes et al., 1991, 1992). Thepresent results show that the variability in compositionand biomass of microzooplankton in the Ria de Ferrolwere related to the reproduction of both planktonic andbenthic species, the latter affecting mainly to the stationsinside the ria. These results agree with the seasonalobservations in shelf waters reporting maximum abun-dance values in spring and summer, especially at coastalstations (Valdes et al., 1991, 1992).
Mesozooplankton biomass in the Ria de Ferrol wasgenerally equivalent to previous reports in other Galicianrias, particularly when dry weight values were compared(Alvarez-Ossorio, 1977; Corral et al., 1981; Valdes et al.,1990, 1991), and higher than those reported for openshelf areas (Valdes et al., 1990, 1991; Halvorsen et al.,2001; Teira et al., 2003). Only maximum values (up to60 mg C m�3 at St. 6) exceeded those reported for thehighly productive Rias Baixas (Corral et al., 1981). Thenumerical dominance of copepods observed was char-acteristic of shelf waters, although meroplanktoniclarvae increased during spring and summer. The absence
of mussel rafts in the Ria de Ferrol, an excellent habitatfor the decapod Pisidia longicornis, would explain thelower abundance of zoeae of this species compared tovalues measured in the Ria de Arousa (Corral et al.,1981). Minimum biomass and abundance values ofmesozooplankton were observed in winter only at theouter ria station, whereas inside the ria the lowestbiomass values were measured in summer. This feature,along with the dominance of small sized copepodssuggests the existence of different mesozooplanktoncommunities inside the ria. The presence of typicalestuarine species, as Acartia margalefi (Alcaraz, 1979),further supports the differential dynamics of mesozoo-plankton inside and outside the ria. Furthermore, a clearidentification of seasonal succession of mesozooplank-ton, as those described in other studies in the nearbyshelf (Valdes et al., 1991; Bode and Alvarez-Ossorio,2004), was not possible in the present study. Thiscontrasts with the results found with phytoplanktonspecies (Fig. 4), which follow a succession patternsimilar to those described for Galician rias, and suggesta rapid response of mesozooplankton to phytoplanktonblooms in this ria. In this way, Hanson et al. (1986)showed a close coupling between phytoplankton andheterotrophs after an upwelling-induced phytoplanktonbloom in the Ria de Arousa. Therefore, zooplanktonspecies succession inside the rias would be different fromthe succession occurring in open shelf waters, becauseof the higher frequency of phytoplankton blooms insidethe rias.
4.3. Plankton biomass structure
The pelagic ecosystem in the Rıa de Ferrol appears asan eutrophic ecosystem, where phytoplankton generallysupports heterotrophic components. In this way, phyto-plankton biomass accounted for a large fraction of totalplankton biomass, except in winter when heterotrophsdominated. Maximum phytoplankton stocks were foundin late summer, thus appearing as the result ofaccumulation during the season of high growth. Theseresults agree with reports of phytoplankton carbondominance in situations of coastal upwelling in the shelf(Varela et al., 1988; Bode and Varela, 1994; Teira et al.,2003), while mesozooplankton excess over phytoplank-ton was characteristic of open shelf, oligotrophic waters(Varela et al., 1988). Microheterotrophs (includingbacteria, protozoa and microzooplankton) reachedhigher biomass values than mesozooplankton in latesummer, thus supporting their participation in theremineralisation of organic matter required to supportprimary production (Bode et al., 2005). This result agreeswith carbon budgets for the nearby shelf, which indicatesthat most of the organic matter produced photosynthet-ically was channelled through the microbial food web

299A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
(Bode and Varela, 1994; Teira et al., 2003; Bode et al.,2004).
The high biomass of phytoplankton recorded in thisand previous studies of the Ria de Ferrol, along with thecorrespondence of heterotrophs in terms of abundanceand biomass, suggests that most of the biogeochemicalprocesses in this ria are driven by phytoplankton bloomsoriginated from nutrients provided by shelf waters. Thisresults in a different behaviour of the Ria de Ferrolwhen compared with other Galician rias, mostly withthe highly productive Rias Baixas, where upwellingevents directly affect productivity inside the ria (e.g.Alvarez-Salgado et al., 2000). The low flow of itstributary rivers causes a low influence of nutrients andorganic matter from continental sources (Bode et al.,2005). Furthermore, its relatively small size and theabsence of a large tidal marsh surface reduces theinfluence of benthic vegetation in primary production,which in other rias can account for a significant fractionof organic matter input (Varela et al., 1984).
Acknowledgements
We acknowledge the collaboration of the crew of R/VLura during sampling. We are indebted to BegonaCastro and Marıa Sanchez for phytoplankton andbacterial sampling, to Isabel Gonzalez and Elena Reyfor zooplankton analyses, and to Marisa Fermoso forCHN analysis. This research was funded in part byproject 1FD97-0479-C03-03 of the Plan Nacional deICD (Spain) and by funds of the Instituto Espanol deOceanografıa. M.M.V. was supported by a PFPI Ph.D.studentship of the Ministerio de Ciencia y Tecnologıa(Spain).
References
Alcaraz, M., 1979. Ecologıa, competencia y segregacion de grupos de
especies asociadas de copepodos del genero Acartia en la Ria de
Vigo. Investigacion Pesquera 43, 689–707.
Alvarez-Ossorio, M.T., 1977. Un estudio de la comunidad de
copepodos de la Rıa de Muros en noviembre de 1975. Boletın del
Instituto Espanol de Oceanografıa 233, 75–108.
Alvarez-Salgado, X.A., Castro, C.G., Perez, F.F., Fraga, F., 1997.
Nutrient mineralization patterns in shelf waters of the Western
Iberian upwelling. Continental Shelf Research 17, 1247–1270.
Alvarez-Salgado, X.A., Gago, J., Mıguez, B.M., Gilcoto, M., Perez,
F.F., 2000. Surface waters of the NW Iberian margin: upwelling on
the shelf versus outwelling of upwelled waters from the Rias
Baixas. Estuarine Coastal and Shelf Science 51, 821–837.
Beers, J.R., Stewart, G.L., 1970. Numerical abundance and estimated
biomass of microzooplankton. Bulletin of the Scripps Institute of
Oceanography 17, 67–87.
Bode, A., Alvarez-Ossorio, M.T., 2004. Taxonomic versus trophic
structure of mesozooplankton: a seasonal study of species
succession and stable carbon and nitrogen isotopes in a coastal
upwelling ecosystem. ICES Journal of Marine Science 61, 563–571.
Bode, A., Varela, M., 1994. Planktonic carbon and nitrogen budgets
for the N-NW Spanish shelf: The role of pelagic nutrient re-
generation during upwelling events. Scientia Marina 58, 221–231.
Bode, A., Varela, M., 1998. Primary production and phytoplankton in
three Galician Rias Altas (NW Spain): seasonal and spatial
variability. Scientia Marina 62, 319–330.
Bode, A., Casas, B., Fernandez, E., Maranon, E., Serret, P., Varela,
M., 1996. Phytoplankton biomass and production in shelf waters
off NW Spain: spatial and seasonal variability in relation to
upwelling. Hydrobiologia 341, 225–234.
Bode, A., Alvarez-Ossorio, M.T., Gonzalez, N., 1998. Estimations of
mesozooplankton biomass in a coastal upwelling area off NW
Spain. Journal of Plankton Research 20, 1005–1014.
Bode, A., Barquero, S., Gonzalez, N., Alvarez-Ossorio, M.T., Varela,
M., 2004. Contribution of heterotrophic plankton to nitrogen
regeneration in the upwelling ecosystem of A Coruna (NW Spain).
Journal of Plankton Research 26, 1–18.
Bode, A., Gonzalez, N., Rodrıguez, C., Varela, M., Varela, M.M.,
2005. Seasonal variability of plankton blooms in the Rıa de Ferrol
(NW Spain): I. Nutrient concentrations and nitrogen uptake rates.
Estuarine Coastal and Shelf Science, doi:10.1016/j.ecss.2004.11.020.
Braun, J.G., Alvarez-Ossorio, M.T., Fernandez-Puelles, M.L., Cami-
nas, J.A., 1990. Estudio sobre la estructura, biomasa y produccion
del meso y microzooplancton en la plataforma de Galicia en abril
de 1982. Boletın del Instituto Espanol de Oceanografıa 6, 81–
96.
Campos, M.J., Marino, J., 1984. El ciclo anual del fitoplancton en la
Ria de Arosa (1978–1979). Boletın del Instituto Espanol de
Oceanografıa 1, 20–30.
Casas, B., Varela, M., Bode, A., 1999. Seasonal succession of
phytoplankton species on the coast of A Coruna (Galicia,
northwest Spain). Boletın del Instituto Espanol de Oceanografıa
15, 413–429.
Corral, J., Masso, C., Alvarez-Ossorio, M.T., 1981. Un estudio
comparado preliminar de la biomasa seca, materia organica,
carbono, nitrogeno, relacion C/N, y del contenido energetico del
zooplancton de las rıas de Arosa y Muros. Boletın del Instituto
Espanol de Oceanografıa 330, 222–240.
Dolan, J., 1991. Microphagous ciliates in mesohaline Chesapeake Bay
waters: estimates of growth rates and consumption by copepods.
Marine Biology 111, 303–309.
Fernandez, E., Bode, A., 1994. Succession of phytoplankton assemb-
lages in relation to the hydrography in the southern Bay of Biscay:
a multivariate approach. Scientia Marina 58, 191–205.
Fernandez de Puelles, M.L., Valencia, J., Jansa, J., Morillas, A., 2004.
Hydrographical characteristics and zooplankton distribution in the
Mallorca channel (Western Mediterranean): spring 2001. ICES
Journal of Marine Science 61, 654–666.
Figueiras, F.G., Niell, F.X., 1987. Composicion del fitoplancton en la
Ria de Pontevedra. Investigacion Pesquera 51, 371–409.
Figueiras, F.G., Pazos, Y., 1991. Microplankton assemblages in three
Rias Baixas (Vigo, Arosa and Muros, Spain) with a subsurface
chlorophyll maximum: their relationships to hydrography. Marine
Ecology Progress Series 76, 219–233.
Figueiras, F.G., Rios, A.F., 1993. Phytoplankton succession, red tides
and the hydrographic regime in the Rıas Bajas of Galicia. In:
Smayda, T.J., Shimizu, Y. (Eds.), Toxic Phytoplankton Blooms in
the Sea. Elsevier Science Publishers, New York, pp. 239–244.
Fileman, E., Burkill, P., 2001. The herbivorous impact of micro-
zooplankton during two short-term Lagrangian experiments off the
NW coast of Galicia in summer 1998. Progress in Oceanography
51, 361–383.
Fraga, F., 1981. Upwelling off the Galician coast, Northwest Spain. In:
Richards, F.A. (Ed.), Upwelling Ecosystems. American Geo-
physical Union, Washington, pp. 176–182.
Fraga, F., 1996. As rıas. In: As Augas de Galicia. Consello da Cultura
Galega, Santiago de Compostela, pp. 215–280.

300 A. Bode et al. / Estuarine, Coastal and Shelf Science 63 (2005) 285–300
Halvorsen, E., Hirst, A.G., Batten, S.D., Tande, K.S., Lampitt, R.S.,
2001. Diet and community grazing by copepods in an upwelled
filament off the NW coast of Spain. Progress in Oceanography 51,
399–421.
Hanson, R.B., Alvarez-Ossorio, M.T., Cal, R., Campos, M.J., Roman,
M., Santiago, G., Varela, M., Yoder, J.A., 1986. Plankton response
following a spring upwelling event in the Ria de Arosa, Spain.
Marine Ecology Progress Series 32, 101–113.
Larsen, J., Sournia, A., 1991. The diversity of heterotrophic
dinoflagellates. In: Patterson, D.J., Larsen, J. (Eds.), The Biology
of Free-living Heterotrophic Flagellates. Oxford University Press,
Oxford, pp. 313–332.
Linley, E.A.S., Newell, R.C., Lucas, M.I., 1983. Quantitative relation-
ships between phytoplankton, bacteria and heterotrophic micro-
flagellates in shelf waters. Marine Ecology Progress Series 12,
77–89.
Lopez-Jamar, E., Cal, R.M., Gonzalez, G., Hanson, R.B., Rey, J.,
Santiago, G., Tenore, K.R., 1992. Upwelling and outwelling effects
on the benthic regime of the continental shelf off Galicia, NW
Spain. Journal of Marine Research 50, 465–488.
Marino, J., Campos, M.J., Nunes, M.T., 1985. Variacion estacional de
los factores ambientales y del fitoplancton en la zona de Lorbe (Rıa
de Ares y Betanzos). Boletın del Instituto Espanol de Oceanografıa
2, 88–99.
McClelland, J.W., Valiela, I., 1998. Changes in food web structure
under the influence of increased anthropogenic nitrogen inputs to
estuaries. Marine Ecology Progress Series 168, 259–271.
Nagata, T., 2000. Production mechanisms of dissolved organic matter.
In: Kirchman, D.L. (Ed.), Microbial Ecology of the Oceans. Wiley-
Liss Inc., New York, pp. 121–152.
Nogueira, E., Ibanez, F., Figueiras, F.G., 2000. Effect of meteorolog-
ical and hydrographic disturbances on the microplankton commu-
nity structure in the Ria de Vigo (NW Spain). Marine Ecology
Progress Series 203, 23–45.
Norland, S., Heldal, M., Tumyr, O., 1987. On the relation between dry
matter and volume of bacteria. Microbial Ecology 13, 95–101.
Olseng, C.D., Naustvoll, L.-J., Paasche, E., 2002. Grazing by the
heterotrophic dinoflagellate Protoperidinium steinii on a Ceratium
bloom. Marine Ecology Progress Series 225, 161–167.
Porteiro, C., Carrera, P., Miquel, J., 1996. Analysis of Spanish acoustic
surveys for sardine 1991–1993: abundance estimates and inter-
annual variability. ICES Journal of Marine Science 53, 429–433.
Porter, K.G., Freig, Y.S., 1980. The use of DAPI for identifying and
counting aquatic microflora. Limnology and Oceanography 25,
943–948.
Prego, R., Varela, M., 1998. Hydrography of the Artabro Gulf in
summer: western coastal limit of Cantabrian seawater and wind-
induced upwelling at Cape Prior. Oceanologica Acta 21, 145–155.
Prego, R., Barciela, M.C., Varela, M., 1999. Nutrient dynamics in the
Galician coastal area (Northwestern Iberian Peninsula): Do the
Rias Bajas receive more nutrient salts than the Rias Altas?
Continental Shelf Research 19, 317–334.
Quevedo, M., Anadon, R., 2000. Spring microzooplankton composi-
tion, biomass and potential grazing in the Central Cantabrian coast
(southern Bay of Biscay). Oceanologica Acta 23, 297–309.
Strathman, R.R., 1967. Estimating the organic carbon content of
phytoplankton from cell volume or plasma volume. Limnology and
Oceanography 12, 411–418.
Teira, E., Abalde, J., Alvarez-Ossorio, M.T., Bode, A., Carino, C.,
Cid, A., Fernandez, E., Gonzalez, N., Lorenzo, J., Valencia, J.,
Varela, M., 2003. Plankton carbon budget in a coastal wind-driven
upwelling station off A Coruna (NW Iberian Peninsula). Marine
Ecology Progress Series 265, 31–43.
Tenore, K.R., Boyer, L.F., Cal, R.M., Corral, J., Garcıa-Fernandez,
C., Gonzalez, N., Gonzalez-Gurriaran, E., Hanson, R.B., Iglesias,
J., Krom, M., Lopez-Jamar, E., McClain, J., Pamatmat, M.M.,
Perez, A., Rhoads, D.C., de Santiago, G., Tiejten, J., Westrich, J.,
Windom, H.L., 1982. Coastal upwelling in the Rias Bajas, NW
Spain: Contrasting the benthic regimes of the Rias de Arosa and de
Muros. Journal of Marine Research 40, 701–772.
Tenore, K.R., Alonso-Noval, M., Alvarez-Ossorio, M., Atkinson,
L.P., Cabanas, J.M., Cal, R.M., Campos, H.J., Castillejo, F.,
Chesney, E.J., Gonzalez, N., Hanson, R.B., Mcclain, C.R.,
Miranda, A., Roman, M.R., Sanchez, J., Santiago, G., Valdes,
L., Varela, M., Yoder, J., 1995. Fisheries and Oceanography off
Galicia, NW Spain (FOG): mesoscale spatial and temporal changes
in physical processes and resultant patterns of biological pro-
ductivity. Journal of Geophysical Research 100, 10943–10966.
Thomas, C.R., 1997. Identifying Marine Phytoplankton. Academic
Press, San Diego, CA, 858 pp.
Tilstone, G.H., Mıguez, B.M., Figueiras, F.G., Fermın, E.G., 2000.
Diatom dynamics in a coastal ecosystem affected by upwelling:
coupling between species, succession, estuarine circulation and
biogeochemical processes. Marine Ecology Progress Series 205, 23–
41.
UNESCO, 1994. Protocols for the Joint Global Ocean Flux Study
(JGOFS) core measurements. UNESCO/IOC Manuals and Guides
29, 1–170.
Valdes, L., Alvarez-Ossorio, M.T., Varela, M., 1990. Relacion entre la
estructura de tallas y la biomasa del zooplancton nerıtico en la
plataforma de La Coruna (NO de Espana). Boletın del Instituto
Espanol de Oceanografıa 6, 97–106.
Valdes, L., Alvarez-Ossorio, M.T., Lavin, A., Varela, M., Carballo,
R., 1991. Ciclo anual de parametros hidrograficos, nutrientes y
plancton en la plataforma continental de La Coruna (NO, Espana).
Boletın del Instituto Espanol de Oceanografıa 7, 91–138.
Valdes, L., Alvarez-Ossorio, M.T., Miranda, A., 1992. Composicion y
abundancia del microzooplancton en la plataforma continental
gallega. Thalassas 10, 107–114.
Varela, M., Prego, R., 2003. Hydrography and phytoplankton in an
isolated and non-pristine ria area: the A Coruna Harbour (NW
Spain). Acta Oecologica 24, 113–124.
Varela, M., Fuentes, J.M., Penas, E., Cabanas, J.M., 1984. Produccion
primaria de las Rıas Baixas de Galicia. In: As Rias Galegas.
Seminario de Estudos Galegos, Santiago de Compostela, pp. 173–
182.
Varela, M., Alvarez-Ossorio, M.T., Valdes, L., Cal, R., Miranda, A.,
de Santiago, G., Costas, E., 1988. Particion de la materia organica
particulada en el area de afloramiento de la plataforma de Galicia
(NO de Espana) durante la campana BREOGAN 684. Boletın del
Instituto Espanol de Oceanografıa 5, 97–108.
Varela, M., Prego, R., Belzunce, M.J., Martın Salas, F., 2001. Inshore-
offshore differences in seasonal variations of phytoplankton
assemblages: the case of a Galician Ria Alta (Ria de A Coruna)
and its adjacent shelf (NW of Spain). Continental Shelf Research
21, 1815–1838.
Varela, M.M., Bode, A., Gonzalez, N., Rodrıguez, C., Varela, M.,
2003. Fate of organic matter in the Rıa de Ferrol (Galicia, NW
Spain): uptake by pelagic bacteria vs. particle sedimentation. Acta
Oecologica 24, S77–S86.
Varela, M., Prego, R., Pazos, Y., 2004. Vertical biogenic particle flux
in a western Galician ria (NW Iberian Peninsula). Marine Ecology
Progress Series 269, 17–32.
Wooster, W.S., Bakun, A., McLain, D.R., 1976. The seasonal
upwelling cycle along the eastern boundary of the North Atlantic.
Journal of Marine Research 34, 131–141.
Zdanowski, M.K., Figueiras, F.G., 1997. Relationships between the
abundance of bacteria and other biota and the hydrographic
variability in the Ria de Vigo, Spain. Marine Ecology Progress
Series 147, 257–267.
Copyright © 2022 FDOKUMEN