
Okadaic acid accumulation in macrofilter feeders subjected to natural blooms of Dinophysis acuminata
7
Okadaic acid accumulation in macrofilter feeders subjected to natural blooms of Dinophysis acuminata Sofia Reizopoulou a, * , Evangelia Strogyloudi a , Antonia Giannakourou a , Kalliopi Pagou a , Ioannis Hatzianestis a , Christina Pyrgaki a , Edna Grane ´li b a Hellenic Centre for Marine Research, Institute of Oceanography, P.O. Box 712, 19013 Anavyssos, Greece b Marine Sciences Department, University of Kalmar, S-39182 Kalmar, Sweden Received 1 February 2007; received in revised form 7 August 2007; accepted 16 August 2007 Abstract Thermaikos Gulf is a eutrophic area located in the Northwestern part of the Aegean Sea in the Eastern Mediterranean. Interspecific differences among various filter feeders in their ability to accumulate okadaic acid, were observed during natural blooms of Dinophysis acuminata in the gulf. Okadaic acid analyses by high performance liquid chromatography (HPLC) were performed on benthic specimens and D. acuminata cell densities and cell toxin content were estimated in water samples. Seven filter feeding species were collected in the gulf during two DSP outbreaks in May 2003 and March 2004. The various species showed a different potential to accumulate okadaic acid in their tissues. The highest concentrations were found in the mussel populations (Mytilus galloprovincialis and Modiolus barbatus), while among the non-bivalve filter feeders, ascidians were the main accumulators of okadaic acid. The rest of shellfish populations (Flexopecten proteus, Chlamys varia and Venus verrucosa) were found to contain toxins only during 2004, when D. acuminata densities were found above 10000 cells l 1 . M. galloprovincialis was proved to be the most appropriate indicator for a safe warning of okadaic acid contamination in Thermaikos Gulf. # 2007 Elsevier B.V. All rights reserved. Keywords: Okadaic acid; DSP toxins; Accumulation; Benthos; Dinophysis acuminata 1. Introduction Harmful algal blooms cause severe economical losses to aquaculture and adversely impact human health. Diarrhetic Shellfish Poison (DSP) in humans causes mainly gastrointestinal symptoms as a result of eating shellfish contaminated with okadaic acid (OA) group of toxins. OA and its derivatives are powerful cytotoxins that can block dephosphorylation of proteins in mammals and plants, but very little is known regarding the effect of these compounds on marine organisms (Bauder et al., 2001). The most important DSP toxin producers belong to the genus Dinophysis, which produce toxins consisting mostly of OA and its derivatives. Thermaikos Gulf (N. Aegean Sea, Eastern Mediter- ranean) is enclosed and eutrophic, with reduced water flux, receiving high nutrient inputs from rivers, urban and industrial runoff (Gotsis-Skretas and Friligos, 1990; Balopoulos and Friligos, 1993; Moncheva et al., 2001). The shellfish farming activity in this area represents about 85% of the total production in Greece (s10 million annually; Zanou and Anagnostou, 2001), with 70–80% of the product exported (Karageorgis et al., 2005). Mussel rafts increase water stability and water residence time, and create microhabitats favouring dinoflagellate growth (Grane ´li et al., 1998). Many sources of nutrients such as sewage and animal wastes, agricultural and fertilizer www.elsevier.com/locate/hal Harmful Algae 7 (2008) 228–234 * Corresponding author. E-mail address: [email protected] (S. Reizopoulou). 1568-9883/$ – see front matter # 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.hal.2007.08.001
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Okadaic acid accumulation in macrofilter feeders subjected to natural blooms of Dinophysis acuminata

www.elsevier.com/locate/hal
Harmful Algae 7 (2008) 228–234
Okadaic acid accumulation in macrofilter feeders subjected
to natural blooms of Dinophysis acuminata
Sofia Reizopoulou a,*, Evangelia Strogyloudi a, Antonia Giannakourou a,Kalliopi Pagou a, Ioannis Hatzianestis a, Christina Pyrgaki a, Edna Graneli b
a Hellenic Centre for Marine Research, Institute of Oceanography, P.O. Box 712, 19013 Anavyssos, Greeceb Marine Sciences Department, University of Kalmar, S-39182 Kalmar, Sweden
Received 1 February 2007; received in revised form 7 August 2007; accepted 16 August 2007
Abstract
Thermaikos Gulf is a eutrophic area located in the Northwestern part of the Aegean Sea in the Eastern Mediterranean.
Interspecific differences among various filter feeders in their ability to accumulate okadaic acid, were observed during natural
blooms of Dinophysis acuminata in the gulf. Okadaic acid analyses by high performance liquid chromatography (HPLC) were
performed on benthic specimens and D. acuminata cell densities and cell toxin content were estimated in water samples. Seven filter
feeding species were collected in the gulf during two DSP outbreaks in May 2003 and March 2004. The various species showed a
different potential to accumulate okadaic acid in their tissues. The highest concentrations were found in the mussel populations
(Mytilus galloprovincialis and Modiolus barbatus), while among the non-bivalve filter feeders, ascidians were the main
accumulators of okadaic acid. The rest of shellfish populations (Flexopecten proteus, Chlamys varia and Venus verrucosa) were
found to contain toxins only during 2004, when D. acuminata densities were found above 10000 cells l�1. M. galloprovincialis was
proved to be the most appropriate indicator for a safe warning of okadaic acid contamination in Thermaikos Gulf.
# 2007 Elsevier B.V. All rights reserved.
Keywords: Okadaic acid; DSP toxins; Accumulation; Benthos; Dinophysis acuminata
1. Introduction
Harmful algal blooms cause severe economical
losses to aquaculture and adversely impact human
health. Diarrhetic Shellfish Poison (DSP) in humans
causes mainly gastrointestinal symptoms as a result of
eating shellfish contaminated with okadaic acid (OA)
group of toxins. OA and its derivatives are powerful
cytotoxins that can block dephosphorylation of proteins
in mammals and plants, but very little is known
regarding the effect of these compounds on marine
organisms (Bauder et al., 2001). The most important
* Corresponding author.
E-mail address: [email protected] (S. Reizopoulou).
1568-9883/$ – see front matter # 2007 Elsevier B.V. All rights reserved.
doi:10.1016/j.hal.2007.08.001
DSP toxin producers belong to the genus Dinophysis,
which produce toxins consisting mostly of OA and its
derivatives.
Thermaikos Gulf (N. Aegean Sea, Eastern Mediter-
ranean) is enclosed and eutrophic, with reduced water
flux, receiving high nutrient inputs from rivers, urban and
industrial runoff (Gotsis-Skretas and Friligos, 1990;
Balopoulos and Friligos, 1993; Moncheva et al., 2001).
The shellfish farming activity in this area represents
about 85% of the total production in Greece (s10 million
annually; Zanou and Anagnostou, 2001), with 70–80% of
the product exported (Karageorgis et al., 2005). Mussel
rafts increase water stability and water residence time,
and create microhabitats favouring dinoflagellate growth
(Graneli et al., 1998). Many sources of nutrients such as
sewage and animal wastes, agricultural and fertilizer

S. Reizopoulou et al. / Harmful Algae 7 (2008) 228–234 229
runoff, as well as the growing aquaculture industry in
coastal areas, can also contribute to stimulate harmful
algal blooms (Anderson et al., 2002).
In Thermaikos Gulf since 2000, DSP outbreaks have
been confirmed and the causative organism has been
identified as Dinophysis acuminata. The most serious
HAB event was recorded during the winter of 2000,
when D. acuminata reached 8.5 � 104 cells l�1 at the
end of January (Koukaras and Nikolaidis, 2004). During
each bloom period (usually early spring), mussel
cultures remain closed to harvest, resulting in a
substantial socio-economic impact in the area (eco-
nomic losses of �s3 million annually) (Karageorgis
et al., 2005).
As filter feeder animals gather food by filtering the
water, contamination with DSP toxins occurs after their
feeding on toxic Dinophysis cells. The filtering capacity
of the filter feeding communities may play an important
role on the control of phytoplankton blooms (Graneli
et al., 1998; Cloern, 2001). The accumulation of OA,
which D. acuminata mainly produces (Vale et al.,
1998), has been until now studied in shellfish
populations, while for other benthic groups (ascidians
and polychaetes), to our knowledge there is no available
information in the published literature.
Two successive annual DSP toxin outbreaks in
Thermaikos Gulf presented an opportunity to (1) assess,
determine and compare the amounts of free okadaic
acid in different filter feeding species naturally
contaminated by D. acuminata; and (2) identify the
most appropriate species to be used as an indicator of
Fig. 1. Sampling stations
DSP contamination in the Gulf, as a tool in the shellfish
stocks management in DSP-affected areas, such as
Thermaikos Gulf.
2. Material and methods
2.1. Description of the study area
The investigation area, Thermaikos Gulf, is located
in the nortwestern part of the Aegean Sea (Fig. 1). Five
major and several minor rivers flow into the gulf,
introducing significant amounts of particulate matter
and nutrient loads (Lykousis and Chronis, 1989;
Friligos et al., 1997; Karageorgis et al., 2005). The
city of Thessaloniki, with its partly untreated domestic
sewage effluents and adjacent industrial zone, con-
tributes to the eutrophication of the Gulf (Anagnostou
et al., 1997). The bottom relief in the area is smooth and
covered by muddy sediments (Lykousis and Chronis,
1989).
2.2. Sampling methods
Field surveys were performed during two successive
D. acuminata blooms in the Thermaikos Gulf. The first
bloom occurred in May 2003 (D. acuminata >2000 cells l�1) and the second and more prominent
bloom occurred in March 2004 (D. acuminata >10,000 cells l�1).
Phytoplankton samples and benthic macroinverte-
brates were collected from four stations, outside (TP7,
in Thermaikos Gulf.

S. Reizopoulou et al. / Harmful Algae 7 (2008) 228–234230
TP2 and TP6, �5.5, 19 and 8.5 m depth, respectively)
and inside (DA3, 8.5 m depth) the mussel culture area
(Fig. 1). Station DA3 is located between mussel rafts in
the area of Chalastra, near the mouth of Axios River.
Seawater samples, for enumeration of Dinophysis cells
in the water column, were taken from different depths
(2 m, 5 m and also from 10 m in TP2), using Niskin
bottles with teflon-coated springs and O-rings. Samples
were fixed with Lugol solution. Species identification
and cell counts were done with an inverted microscope
(Olympus CK2) after 24 h of sedimentation in 25 ml
settling chambers (Utermohl, 1958). Dinophysis cells
were also collected on GFC glass-fibre filters after
filtering 1 l of seawater. Filters were stored at �18 8Cuntil extraction for toxin analysis.
During the two bloom periods, cultured and natural
filter feeding populations were collected. Cultured
mussels (M. galloprovincialis) from the mussel rafts
(Fig. 1, station DA3) were collected from two different
depths (2 and 5 m). Benthic populations were sampled
from stations TP7, TP2 and TP6 (Fig. 1) by SCUBA
diving and using a rectangular dredge appropriate to
catch large animals. The most abundant filter feeders
collected were bivalves, as the mussel Modiolus
barbatus, scallops (Flexopecten proteus and Chlamys
varia) and clams (Venus verrucosa). The sabellid
polychaete Sabella spallanzanii was mainly found at
station TP7 (Fig. 1).
During the second sampling survey in 2004, solitary
ascidians associated with the mussel cultures and
Mytilus galloprovincialis from the sediments below the
cultures also were collected. M. galloprovincialis is
usually found under the mussel rafts (station DA3), as
animals detach from the rafts and develop benthic
populations.
Pooled samples of 3–5 individuals of each species
with similar body size were prepared, weighed and
stored at �20 8C for whole tissue toxin analyses.
2.3. Okadaic acid determination
Okadaic acid analysis was performed with high
performance liquid chromatography (HPLC)-fluores-
cence detection after pre-column derivatization with
3-bromomethyl-7-methoxy-1,4-benzoxazin-2-one
(BrMB) (Zhou et al., 1999). Fresh homogenized animal
tissue (1.0 g) or a filter containing the phytoplankton
cells was extracted with 4 ml of aqueous 80% methanol.
After adding 60 ml of water the methanolic extract was
further extracted three times with 10 ml of dichlor-
omethane. The extracts were combined, evaporated to
dryness and the residue was redissolved in 0.5 ml of
dichloromethane and cleaned up by solid phase
extraction (SPE). A SPE cartridge (Sep-Pak) containing
1 g of silica, washed with 8.0 ml acetone/water (97:3)
and 10 ml dichloromethane and the extract was then
added. The cartridge was purged with 10 ml of
dichloromethane and 5 ml methanol/acetone (3:97)
and the toxins were eluted with 10 ml of methanol/
acetone (30:70). The eluate was transferred to a glass
vial and derivatized with BrMB (Fluka). The eluate was
dried under nitrogen gas, 50 ml of N-ethyl-diisopropy-
lamine solution (0.1% in acetone), and 50 ml of BrMB
solution (0.1% in acetone) were added, and the mixture
was heated at 70 8C for 20 min. The derivatized mixture
was immediately evaporated to dryness under nitrogen
gas and the residue was redissolved in 100 ml of
acetonitrile for HPLC analysis.
The OA determination was performed with a
Shimadzu HPL chromatograph equipped with a LC-
10AD ternary pump and a RF-10Axl fluorescence
detector. The LC column was a Spherisorb ODS2,
25 cm � 4.6 mm, 5 mm and maintained at 28 8C.
Isocratic conditions were used; the mobile phase was
acetonitrile/water 65:35, and the flow rate was
1.0 ml min�1, and the injection volume was 20 ml.
The fluorescence detector was set to 345 nm excitation
and 440 nm emission. OA concentrations were calcu-
lated using a calibration curve that was based on
injections of standard OA (Calbiochem) solutions
derivatized under the same conditions. The detection
limit (S/N 3:1) for the standard OA solutions was
0.05 ng/injection, corresponding to 0.25 ng filter�1 for
the phytoplankton samples, whereas for tissue samples
the detection limit ranged from 0.8 to 5 ng/g depending
on the substrate. The protocol was tested by analyzing
certified reference material MUS-2, from the Institute
for Marine Biosciences in Halifax, Canada.
3. Results
Fig. 2 illustrates the spatial and vertical distribution
of D. acuminata cell densities and their cellular toxin
content during the blooms of May 2003 and March
2004.
In May 2003, D. acuminata cell densities varied
from 500 cells l�1 in stations DA3 and TP7 to
2200 cells l�1 at station TP2 and the average OA
intracelullar concentrations in D. acuminata varied
from 3.7 to 8.6 pg cell�1 (Fig. 2).
The highest D. acuminata cell densities were
recorded during March 2004. Cell densities ranged
from 600 cells l�1 at station DA3 (mussel culture area)
to 10,700 cells l�1 at station TP2. OA in D. acuminata
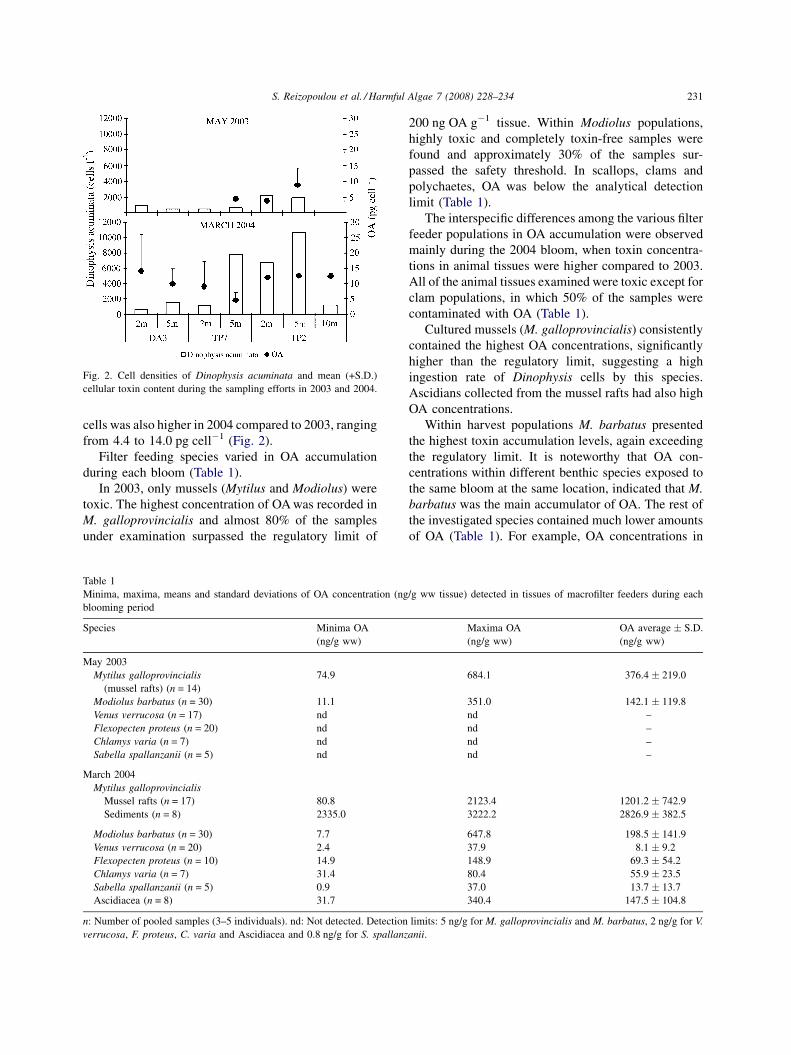
S. Reizopoulou et al. / Harmful Algae 7 (2008) 228–234 231
Fig. 2. Cell densities of Dinophysis acuminata and mean (+S.D.)
cellular toxin content during the sampling efforts in 2003 and 2004.
cells was also higher in 2004 compared to 2003, ranging
from 4.4 to 14.0 pg cell�1 (Fig. 2).
Filter feeding species varied in OA accumulation
during each bloom (Table 1).
In 2003, only mussels (Mytilus and Modiolus) were
toxic. The highest concentration of OA was recorded in
M. galloprovincialis and almost 80% of the samples
under examination surpassed the regulatory limit of
Table 1
Minima, maxima, means and standard deviations of OA concentration (ng
blooming period
Species Minima OA
(ng/g ww)
May 2003
Mytilus galloprovincialis
(mussel rafts) (n = 14)
74.9
Modiolus barbatus (n = 30) 11.1
Venus verrucosa (n = 17) nd
Flexopecten proteus (n = 20) nd
Chlamys varia (n = 7) nd
Sabella spallanzanii (n = 5) nd
March 2004
Mytilus galloprovincialis
Mussel rafts (n = 17) 80.8
Sediments (n = 8) 2335.0
Modiolus barbatus (n = 30) 7.7
Venus verrucosa (n = 20) 2.4
Flexopecten proteus (n = 10) 14.9
Chlamys varia (n = 7) 31.4
Sabella spallanzanii (n = 5) 0.9
Ascidiacea (n = 8) 31.7
n: Number of pooled samples (3–5 individuals). nd: Not detected. Detection
verrucosa, F. proteus, C. varia and Ascidiacea and 0.8 ng/g for S. spallanz
200 ng OA g�1 tissue. Within Modiolus populations,
highly toxic and completely toxin-free samples were
found and approximately 30% of the samples sur-
passed the safety threshold. In scallops, clams and
polychaetes, OA was below the analytical detection
limit (Table 1).
The interspecific differences among the various filter
feeder populations in OA accumulation were observed
mainly during the 2004 bloom, when toxin concentra-
tions in animal tissues were higher compared to 2003.
All of the animal tissues examined were toxic except for
clam populations, in which 50% of the samples were
contaminated with OA (Table 1).
Cultured mussels (M. galloprovincialis) consistently
contained the highest OA concentrations, significantly
higher than the regulatory limit, suggesting a high
ingestion rate of Dinophysis cells by this species.
Ascidians collected from the mussel rafts had also high
OA concentrations.
Within harvest populations M. barbatus presented
the highest toxin accumulation levels, again exceeding
the regulatory limit. It is noteworthy that OA con-
centrations within different benthic species exposed to
the same bloom at the same location, indicated that M.
barbatus was the main accumulator of OA. The rest of
the investigated species contained much lower amounts
of OA (Table 1). For example, OA concentrations in
/g ww tissue) detected in tissues of macrofilter feeders during each
Maxima OA
(ng/g ww)
OA average � S.D.
(ng/g ww)
684.1 376.4 � 219.0
351.0 142.1 � 119.8
nd –
nd –
nd –
nd –
2123.4 1201.2 � 742.9
3222.2 2826.9 � 382.5
647.8 198.5 � 141.9
37.9 8.1 � 9.2
148.9 69.3 � 54.2
80.4 55.9 � 23.5
37.0 13.7 � 13.7
340.4 147.5 � 104.8
limits: 5 ng/g for M. galloprovincialis and M. barbatus, 2 ng/g for V.
anii.

S. Reizopoulou et al. / Harmful Algae 7 (2008) 228–234232
scallops consistently were lower than the safety limit for
human consumption.
In each bloom period a high spatial (vertical and
horizontal) intraspecific variation of OA accumulation
was observed, both for natural and cultured populations.
Variations in toxin concentration levels were found in
M. galloprovincialis collected from different depths,
although not statistically significant. Also within the
natural populations there was a spatial variation in toxin
accumulation among the different sampling locations.
However no statistically significant correlation between
animal toxin accumulation levels and Dinophysis cell
abundances was found.
It is important to note that OA levels in suspended
mussels were significantly lower than the levels in
mussels collected from the sediments below the rafts
(t-test, p = 0.0003). OA concentrations were maximal in
the bottom mussel populations exceeding the public
health safety threshold.
4. Discussion
The intense toxic bloom of March 2004 in
Thermaikos Gulf was associated with an increased
OA accumulation in animal tissues and also a higher
number of filter feeding species found to be con-
taminated, in comparison to the toxic bloom of 2003.
During both toxic blooms, D. acuminata cell densities
in the mussel culture area (st. DA3) were generally
comparable, indicating that mussels are effective filter
feeders and their dense populations, are able to reduce/
or control Dinophysis cell densities. The OA concen-
tration in D. acuminata cells found in the present study
was comparable with those of other studies (Lee et al.,
1989; Maneiro et al., 2000; Morono et al., 2003).
Within the filter feeders functional group there was a
marked interspecific variation in the OA accumulation,
indicating that not all species concentrated OA to the
same extent. M. galloprovincialis had the highest OA
accumulation compared to the other filter feeding
species. Sidari et al. (1998) suggested that the mussel is
preferentially selecting dinoflagellates rather than
diatoms and described a selective preference of this
species for the genus Dinophysis. Also the ascidians
associated with cultured mussels contained high OA
concentrations. Knowledge of phytoplankton species
that filter feeders can preferentially remove from the
water column is important for a better understanding of
the degree to which these species can control the toxic
blooms.
Interspecific differences in toxin accumulation can
also be attributed to species-specific metabolism of OA.
Suzuki and Mitsuya (2001) found that mussels have a
higher potential to accumulate OA homologues than
scallops. Differences in animal toxin accumulation may
arise from selective retention or elimination of
individual toxins or from other transformation pro-
cesses (Brijeli and Shumway, 1998). Taleb et al. (2001)
reported that clams accumulate PSP toxins to a lesser
degree than other species; however, members of the
family Veneridae can become very toxic by accumulat-
ing other toxins such as domoic acid (Vale and
Sampayo, 2001). In the present investigation only the
form of free OA was quantified, however, further
research is needed to investigate the presence of
conjugated forms of OA within the various species.
Variations in OA accumulation could also reflect the
different exposure of each species to toxin amounts. The
benthic populations collected outside the mussel culture
area were exposed to particulate material such as
sedimenting toxic algae and algae derived detritus,
depending mainly on the water mass circulation and
particle sinking rates. Among these populations, M.
barbatus was the most contaminated species while the
other species accumulated much less OA, and only
during the toxic bloom of 2004. Scallops (F. proteus and
C. varia) were less contaminated with OA, well below
the regulatory limit and similarly the clam V. verrucosa
presented very low OA levels. Clams can minimize their
exposure to toxins by reducing their feeding activity and
rejecting cells as pseudofaeces (Wikfors, 2005).
Another efficient filter feeding species, such as the
polychaete S. spallanzanii, had only traces of OA.
OA concentrations were maximal in Mytilus
collected from the sediments (>2 mg g�1) under the
mussel rafts, suggesting that these populations were
subjected to a major source of toxins. In addition to
settling phytoplankton and decaying cells, the benthic
mussels were also exposed to high amounts of toxins in
faecal material from the overlying suspended cultured
mussels. Faeces and pseudofaeces may contain high
amounts of toxins, contributing to the transfer of toxins
from the water phase to the sediment (Svensen et al.,
2005). Moreover, high ingestion rates in suspended
mussels would increase digestive activity with faster
elimination of toxins through faecal deposition (Blanco
et al., 1999).
The variation of toxin accumulation could therefore
be attributed to different selection efficiencies among
the various filter feeding species (Shumway and Cucci,
1987), to different species-specific OA metabolism
(Brijeli and Shumway, 1998), but also to the degree of
exposure of each species to different amounts of toxins
within the various habitats.

S. Reizopoulou et al. / Harmful Algae 7 (2008) 228–234 233
Intraspecific spatial variability of OA accumulation
in macrofilter feeding populations reflected the
horizontal and vertical patchiness of the toxic blooms.
The abundance of toxic cells and toxin concentrations in
animal tissues are sometimes not significantly related
(Lassus et al., 1991; Dahl and Johannessen, 2001), as
organisms can be found contaminated without the toxic
algae being detected (MacKenzie et al., 2004). Toxic
cell concentrations can decline while the toxins still
remain in the water column (MacKenzie et al., 2004), or
the toxin effect may be diluted in the presence of
alternative food sources (Dahl and Johannessen, 2001).
In conclusion, among shellfish, mussels (M. gallo-
provincialis and M. barbatus) were the main accumu-
lators of OA and their tissue concentrations exceeded
the levels recommended for consumption. The highest
amounts of OA were detected in M. galloprovincialis,
suggesting this species as the most appropriate
candidate for use as an indicator for DSP toxins in
Thermaikos Gulf. Further investigation on shellfish
toxin accumulation has important implications for
managing bivalve populations for human consumption.
Monitoring of macroinvertebrates is crucial to deter-
mine toxin accumulation in various benthic organisms
that could potentially act as vectors for toxin transfer in
the food web and to explore the dynamics of filtering
capacity of the system, since the dense filter feeding
populations in Thermaikos Gulf strongly influence
matter and energy flow in the coastal ecosystem.
Acknowledgements
We thank I. Varkintzi, A. Gremmenas and S.
Mitsoudi for their help during the field surveys in
Thermaikos Gulf. Financial support was provided by
the European Commission through the FATE project
‘‘Transfer and Fate of Harmful Algal Bloom (HAB)
Toxins in European Marine Waters’’ (contract EVK3-
CT01-00055) as part of the EC-EUROHAB cluster.[SS]
References
Anagnostou, Ch., Kaberi, H., Karageorgis, A., 1997. Environmental
impact on the surface sediments of the bay and the gulf of
Thessaloniki according to the Geoaccumulation Index classifica-
tion. In: Rajar, J., Brebbia, C.A. (Eds.), Water Pollution IV,
Measuring Modelling and Prediction. Computational Mechanics
Publications, Southampton.
Anderson, D.M., Glibert, P.M., Burkholder, J.M., 2002. Harmful algal
blooms and eutrophication: nutrient sources, composition and
consequences. Estuaries 25, 704–726.
Balopoulos, E.T., Friligos, N.C., 1993. Water circulation and eutro-
phication in the North-West Aegean Sea: Thermaikos Gulf. J.
Environ. Sci. Health A 28, 1311–1329.
Bauder, A.G., Cembella, A.D., Brijeli, V.M., Quilliam, M.A., 2001.
Uptake and fate of diarrhetic shellfish poisoning toxins from the
dinoflagellate Prorocentrum lima in the bay scallop Argopecten
irradians. Mar. Ecol. Prog. Ser. 213, 39–52.
Blanco, J., Fernandez, M.L., Mıguez, A., Morono, A., 1999. Okadaic
acid depuration in the mussel Mytilus galloprovincialis: one- and
two-compartment models and the effect of environmental condi-
tions. Mar. Ecol. Prog. Ser. 176, 153–163.
Brijeli, V.M., Shumway, S.E., 1998. Paralytic shellfish toxins in
bivalve mollusks: occurrence, transfer kinetics, and bio-transfor-
mation. Rev. Fish Sci. 6, 315–383.
Cloern, J.E., 2001. Our evolving conceptual model of the coastal
eutrophication problem. Mar. Ecol. Prog. Ser. 210, 223–253.
Dahl, E., Johannessen, T., 2001. Relationship between occurrence of
Dinophysis species (Dinophyceae) and shellfish toxicity. Phyco-
logia 40, 223–227.
Friligos, N., Kondylakis, J.C., Psyllidou-Giouranovits, R., Georgako-
poulou-Gregoriadou, E., 1997. Eutrophication and phytoplankton
abundance in the Thermaikos Gulf, Greece. Fresenius Environ.
Bull. 6, 27–31.
Gotsis-Skretas, O., Friligos, N., 1990. Contribution to eutrophication
and phytoplankton ecology in the Thermaikos Gulf. Thalassogra-
phica 13, 1–12.
Graneli, E., Sakshang, E., Pallon, J., Maestrini, S., Luckas, B.,
Martinez, R., 1998. Effect of nutrient ratios on harmful phyto-
plankton and their toxin production (NUTOX). In: Third European
Marine Science and Technology Conference (MAST Conference),
Project Synopses vol. 1, Marine Systems, Lisbon, May 23–27, pp.
99–103.
Karageorgis, A.P., Skourtos, M.S., Kapsimalis, V., Kontogianni, A.D.,
Skoulikidis, N.Th., Pagou, K., Nikolaidis, N.P., Drakopoulou, P.,
Zanou, B., Karamanos, H., Levkov, Z., Anagnostou, Ch., 2005. An
integrated approach to watershed management within the DPSIR
framework: Axios River Catchment and Thermaikos Gulf. Reg.
Environ. Change 5, 138–160.
Koukaras, K., Nikolaidis, G., 2004. Dinophysis blooms in Greek
coastal waters (Thermaikos Gulf, NW Aegean Sea). J. Plankton
Res. 26, 445–457.
Lassus, P., Herbland, A., Lebaut, C., 1991. Dinophysis blooms and
toxic effects along the French coast. World Aquacult. 22, 49–54.
Lee, J.S., Igarashi, T., Fraga, S., Dahl, E., Hovgaard, P., Yasumoto, T.,
1989. Determination of diarrhetic shellfish toxins in various
dinoflagellates species. J. Appl. Phycol. 1, 147–152.
Lykousis, V., Chronis, G., 1989. Mechanisms of sediment transport
and deposition: sediment sequences and accumulation during the
Holocene on the Thermaikos Plateau, the continental slope and
Basin (Sporadhes basin), NWAegean Sea, Greece. Mar. Geol. 87,
15–26.
MacKenzie, L., Beuzenberg, V., Holland, P., McNabb, P., Selwood,
A., 2004. Solid phase adsorption toxin tracking (SPATT): a new
monitoring tool that simulates the biotoxin contamination of filter
feeding bivalves. Toxicon 44, 901–918.
Maneiro, I., Frangopoulos, M., Guisande, C., Fernadez, M.L., Reg-
uera, B., Riveiro, I., 2000. Zooplankton as a potential vector of
diarrhetic shellfish poisoning toxins through the food web. Mar.
Ecol. Prog. Ser. 201, 155–163.
Moncheva, S., Gotsis-Skretas, O., Pagou, K., Krastev, A., 2001.
Phytoplankton blooms in Black Sea and Mediterranean ecosys-
tems subjected to anthropogenic eutrophication: similarities and
differences. Estuar. Coast. Shelf Sci. 53, 281–295.
Morono, A., Arevalo, F., Fernandez, M.L., Maneiro, J., Pazos, Y.,
Salgado, C., Blanco, J., 2003. Accumulation and transformation of

S. Reizopoulou et al. / Harmful Algae 7 (2008) 228–234234
DSP toxins in mussels Mytilus galoprovincialis during a toxic
episode caused by Dinophysis acuminata. Aquat. Toxicol. 62,
269–280.
Sidari, L., Nichetto, P., Cok, S., Sosa, S., Tubaro, A., Honsell, G.,
Della Loggia, R., 1998. Phytoplankton selection by mussels, and
diarrhetic shellfish poisoning. Mar. Biol. 131, 103–111.
Shumway, S.E., Cucci, T.L., 1987. The effects of the toxic dinofla-
gellate Protogonyaulax tamarensis on the feeding behavior of
bivalve mollusks. Aquat. Toxicol. 10, 9–27.
Suzuki, T., Mitsuya, T., 2001. Comparison of dinophysistoxin-1 and
esterified dinophysistoxin-1 (dinophysistoxin-3) contents in the
scallop Patinopecten yessoensis and the mussel Mytilus gallopro-
vincialis. Toxicon 39, 905–908.
Svensen, C., Strogyloudi, E., Riser, C.W., Dadlmann, J., Legrand, C.,
Wassmann, P., Graneli, E., Pagou, K., 2005. Reduction of cya-
nobacterial toxins through coprofagy in Mytilus edulis. Harmful
Algae 4, 329–336.
Taleb, H., Vale, P., Jaime, E., Blaghen, M., 2001. Study of paralytic
shellfish poisoning toxin profile in shellfish from the Mediterra-
nean shore of Morocco. Toxicon 39, 1855–1861.
Utermohl, H., 1958. Zur vervollkommnung der quantitativen Phyto-
plankton Methodik. Mitt. Int. Ver. Theor. Angew Limnol. 9, 1–38.
Vale, P., Sampayo, M.A.M., Quilliam, M.A., 1998. DSP complex
toxin profiles relation with Dinophysis spp. occurrence and
domoic acid confirmation by LC–MS in Portuguese bivalves.
In: Reguera, B., Blanco, J., Fernandez, M.L., Wyatt, T. (Eds.),
Harmful Algae. Xunta de Galicia and IOC of UNESCO Spain, pp.
503–542.
Vale, P., Sampayo, M.A.M., 2001. Domoic acid in Portuguese shell-
fish and fish. Toxicon 39, 893–904.
Wikfors, G.H., 2005. A review and new analysis of trophic interac-
tions between Prorocentrum minimum and clams, scallops, and
oysters. Harmful Algae 4, 585–592.
Zanou, B., Anagnostou, Ch., 2001. Integrated management in the
drainage basin of the Thermaikos Gulf (NW-Aegean Sea): meth-
odological steps and proposed measures, according to the win-win
policy. Eur. Water Manage. 4, 33–42.
Zhou, M., Li, J., Luckas, B., Yu, R., Yan, T., Hummert, Ch., Kastrup,
S., 1999. A recent shellfish toxin investigation in China. Mar.
Pollut. Bull. 39, 331–334.


![High Efficiency Transformation of Banana [Musa acuminata L. cv. Matti (AA)] for Enhanced Tolerance to Salt and Drought Stress Through Overexpression of a Peanut Salinity-Induced Pathogenesis-Related](https://static.fdokumen.com/doc/165x107/6332b7765f7e75f94e093b00/high-efficiency-transformation-of-banana-musa-acuminata-l-cv-matti-aa-for.jpg)











