
Seasonal Isolation of Microalgae from Municipal Wastewater for Remediation and Biofuel Applications
22
Accepted Article This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process which may lead to differences between this version and the Version of Record. Please cite this article as an 'Accepted Article', doi: 10.1111/jam.12818 This article is protected by copyright. All rights reserved. Article Type: Original Article Seasonal Isolation of Microalgae from Municipal Wastewater for Remediation and Biofuel Applications Kyoung C. Park, Crystal E.G. Whitney, Catherine Kozera, Stephen J.B. O’Leary, Patrick J. McGinn* Aquatic and Crop Resources Development, National Research Council of Canada, 1411 Oxford St., Halifax, N.S., Canada B3H 3Z1 *Corresponding author: Patrick J. McGinn Aquatic and Crop Resources Development, National Research Council of Canada, 1411 Oxford St., Halifax, N.S., Canada B3H 3Z1 Tel: 1 902 293 2746 Email: [email protected] Abstract Aims: The objective of the study was to isolate microalgae strains from treated municipal wastewater in both summer and winter seasons in order to identify strains better suited for nutrient remediation and biofuel production under either cooler or warmer temperatures. Methods and Results: Fifty-six strains in total were isolated and identified by DNA sequencing from effluent samples collected from a local wastewater treatment plant during the summer and winter of 2011. Screening of 41 isolates based on fatty acid productivity at either 22 or 10 °C resulted in the selection of 12 strains organized into two groups of 6 - the M (mild) and C (cool) groups, respectively. Four of the C- group strains were isolated from the winter sample, while four of the M-group isolates were isolated from
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Seasonal Isolation of Microalgae from Municipal Wastewater for Remediation and Biofuel Applications

Acc
epte
d A
rtic
le
This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process which may lead to differences between this version and the Version of Record. Please cite this article as an 'Accepted Article', doi: 10.1111/jam.12818 This article is protected by copyright. All rights reserved.
Article Type: Original Article
Seasonal Isolation of Microalgae from Municipal Wastewater for Remediation and Biofuel
Applications
Kyoung C. Park, Crystal E.G. Whitney, Catherine Kozera, Stephen J.B. O’Leary, Patrick J. McGinn*
Aquatic and Crop Resources Development, National Research Council of Canada,
1411 Oxford St., Halifax, N.S., Canada B3H 3Z1
*Corresponding author: Patrick J. McGinn
Aquatic and Crop Resources Development, National Research Council of Canada,
1411 Oxford St., Halifax, N.S., Canada B3H 3Z1
Tel: 1 902 293 2746
Email: [email protected]
Abstract
Aims: The objective of the study was to isolate microalgae strains from treated municipal wastewater in
both summer and winter seasons in order to identify strains better suited for nutrient remediation and
biofuel production under either cooler or warmer temperatures.
Methods and Results: Fifty-six strains in total were isolated and identified by DNA sequencing from
effluent samples collected from a local wastewater treatment plant during the summer and winter of 2011.
Screening of 41 isolates based on fatty acid productivity at either 22 or 10 °C resulted in the selection of
12 strains organized into two groups of 6 - the M (mild) and C (cool) groups, respectively. Four of the C-
group strains were isolated from the winter sample, while four of the M-group isolates were isolated from

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
the summer sample. Fatty acid pools in M-group strains were heavily regulated in response to growth
temperature while C-group strains were more insensitive. In three of the six C-group strains, the rates of
biomass and fatty acid productivity at 10 °C exceeded the corresponding rates at 22 °C. Conversely, M-
group were always more productive at 22 compared to 10 °C. Mixotrophic strategies to enhance
productivity were generally unsuccessful in M-group strains at 22°C but proved more effective in C-
group cultures at 10 °C.
Conclusions: In general, C group strains appeared better suited for growth in municipal wastewater at 10
°C, while M group strains were better suited at 22 °C. On balance, C-group isolates were more likely to
come from winter wastewater samples while M-group strains were more likely to come from the summer
sample.
Significance and Impact: Our results demonstrate that the effects of temperature on microalgal growth
for wastewater remediation can be mitigated somewhat by isolation and careful selection of strains
adapted to seasonal wastewater conditions.
Keywords: Wastewater, bioremediation, biomass productivity, microalgae, CO2 capture, biofuel, fatty
acids, temperature
Introduction
In recent years, tremendous efforts, both public and private, have been undertaken to produce
microalgae-based biofuels. However, comparatively little effort has being conducted in Northern regions
due to strong seasonal effects on climate and solar insolation levels, which are likely to negatively affect
feasibility and production costs. However, these disadvantages can be potentially mitigated by careful
selection of strains that can be grown in free and abundant wastewater and at lower temperatures. It has
been shown that some freshwater strains grow and accumulate lipid in municipal wastewater media as
efficiently as in traditional laboratory growth media (Li et al. 2011; Zhou et al. 2011; McGinn et al. 2012;
Park et al. 2012).

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Biofuel production using wastewater-grown microalgae has attracted considerable attention due
to its environmental and economic benefits (Li et al. 2011; Pittman et al. 2011; McGinn et al. 2012; Park
et al. 2012). Large-scale microalgae production using municipal wastewater is especially attractive in
northern regions, since treated wastewater contains abundant nutrients and has a relatively more stable
annual temperature range of 10-25 °C compared to ambient seasonal temperatures in Northern locations
such as Canada (Talbot and de la Noüe 1993; McGinn et al. 2011).
It has been shown that some strains show better performance under mixotrophic culture
conditions compared to phototrophic or heterotrophic culture modes (Li et al. 2011). In a previous study
we reported that microalgae grown in secondary municipal wastewater (SMW) had characteristics more
similar to those grown in inorganic media such as B3NV (Park et al. 2012). This is due to the low organic
carbon concentration (low BOD) in SMW. Biomass and lipid production were limited by the
concentration of dissolved inorganic/organic nutrients in SMW. However, recent studies (Li et al. 2011;
Zhou et al. 2011; Park et al. 2012) have shown that SMW combined with other external carbon sources
such as glycerol or centrate (a highly concentrated municipal wastewater) can boost biomass and lipid
production. The centrate, generated from centrifugal dewatering of waste solids and sludges, has some
advantages when used as a cultivation media for microalgae production. These include a high
concentration of nutrients such as carbon, nitrogen, phosphorous and essential minerals, and the
availability of large production volumes (Li et al. 2011; Zhou et al. 2011). Biodiesel production from
waste fats and greases currently generates large amounts of glycerol as a by-product (Perez-Garcia et al.
2011). Although other carbon sources, such as glucose and acetate, have been tested to boost microalgae
biomass and lipid production, glycerol has some advantages including lower cost and abundant supply
(Cerón-Garcia et al. 2006, Park et al. 2012). Strains of microalgae commonly isolated from wastewater,
including species of the genera Actinastrum, Chlamydomonas, Chlorella, Micractinium and Scenedesmus
are known to grow well mixotrophically (Park et al. 2011; Zhou et al. 2011; Park et al. 2012).
Much of the previous research exploring the potential for integration of microalgal biofuel
production and municipal wastewater treatment (Zhou et al. 2011; Park et al. 2012) has been conducted at
relatively high temperatures (22-25 °C) and therefore is less relevant to the cold-weather conditions such
as the winter season in Northern regions. In Nova Scotia, Canada, SMW outflow typically has a
temperature range between 10-25 °C annually (Dereck Avery, Mill Cove Wastewater Treatment Plant,
personal communication). This provides Northern regions with a promising possibility for year-round

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
microalgal biofuel production using municipal wastewater, provided suitable strains with acceptable
growth rates at the lower temperature ranges can be isolated and successfully cultivated.
Even though there are some reports of the effects of low temperatures on fatty acid profiles in
microalgae (Joh et al. 1993; Thompson 1996; McLarnon-Riches et al. 1998; Jiang and Gao 2004),
information related to the potential for microalgae to remediate wastewater under such conditions is still
rather limited. However, it has been generally established that membrane lipids are strongly influenced by
temperature changes. When temperature is decreased, more unsaturated fatty acids are produced in
microalgae membranes; while more saturated fatty acids are produced when temperature is increased
(Guschina and Harwood 2006).
To overcome cold-weather disadvantages in Northern regions, we isolated 56 strains of
microalgae that were tolerant to cold-temperatures (5°C) from a secondary municipal wastewater, from
which we further down-selected to a group of 19 isolates. We compared their biomass and lipid
production at two temperatures, 10 and 22°C, which represented the approximate range of temperatures
expected in wastewater effluent in Northern latitudes over an annual cycle. We also explored the
bioremediation efficiency of these strains and their potential to grow mixotrophically as a strategy to
augment growth and biomass production.
Materials and Methods
Microalgae strain isolation
Municipal wastewater was obtained from the Mill Cove Waste Water treatment plant (Bedford,
Nova Scotia) in June, 2010 (summer sample) and January, 2011 (winter sample). Secondary wastewater
effluent was collected in carboys (15 l) and maintained in an incubator for one month at 5°C under
continuous illumination of 50 µmol photons m-2 s-1. Carboys were passively aerated using 0.2 µm PTFE
membrane filter (Pall Life Sciences, Port Washington). After a month of incubation in these conditions, a
small volume (100 μl) of wastewater was transferred onto ten solidified agar plates containing modified
Bold’s basal medium (Bold 1949), as described elsewhere (Park et al.2012). A total of 56 microalgae
isolates (36 and 20 from the summer and winter samples, respectively) were eventually obtained after
serial plating on agar containing ampicillin/chloramphenicol/streptomycin (100 μg ml-1 for each). Strains
sensitive to antibiotics were transferred without them until no bacterial colonies developed.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
PCR and DNA sequencing of rDNA for strain identification
Fifty six individual strains were identified by sequencing the internal transcribed spacer regions
(ITS1 and ITS2) of the 18S ribosomal DNA gene. Detailed methods were as described in Park et al.
(2012). Briefly, PCR products were cloned into sequencing vectors (pGEM T-Easy, Mississauga) and
propagated in E. coli. Cloned plasmids were extracted (Qiagen, Toronto) and sequenced. Sequence results
were aligned and compared with known sequences using the National Center for Biotechnology and
Information BLAST server (Benson et al. 2000) optimized for highly similar sequences (megablast).
Putative identities were determined based on a minimum of 96% sequence identity over a coverage of at
least 600 base pairs with sequences in GenBank. All sequences of 56 microalgae isolates have been
deposited in the GenBank under accession numbers KP204548-KP204598. The ITS sequencing results of
the 56 strains resulted in the putative identification of 11 different species from 6 genera, including
Chlorella, Dictyosphaerium, Micractinium, Mucidosphaerium, Pseudotetracystis and Tetracystis. From
the original 56 strains, 15 were not considered for further study due to slow growth or redundancy,
leaving 41 (Table 1).
Initial Screen of 41 Isolates at 22 °C
For cultivation experiments, UV-treated municipal wastewater effluent was obtained from the
Mill Cove Waste Water (MCWW) secondary treatment plant in Bedford, Nova Scotia. The MCWW was
filtered again using Stericup & Steritop filtration with a 0.2μm pore size (Millipore, USA). Two replicates
of each of the 41 MCWW isolates were grown photoautotrophically in 250-ml Erlenmeyer flasks (150 ml
of culture) for 8 days. Growth performance of each strain was assessed by measuring the specific growth
rate (μ) in the exponential phase, as well as final biomass and fatty acid production. Each culture was
inoculated with the same bio-volume (2x106 μm3, equivalent to 100,000 cells ml-1 Chlorella sp.) in
duplicate 250-ml Erlenmeyer flasks (150 ml of culture). Duplicate cultures were shaken at 175 rpm
(Innova 2100, New Brunswick Scientific, Edison, USA) at 22 °C under continuous illumination of 75
µmol photons m-2 s-1 white light in a plant growth chamber (Conviron, Winnipeg, Canada). No CO2
supplementation in the air was provided to these cultures. The cells were counted daily using a Multisizer
3 particle sizer (Beckman Coulter, Canada) for 8 days. On the final day, cells were harvested and freeze
dried for biomass and fatty acid analyses.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Down-Selection to 19 Isolates and Cultivation at 10 °C
Isolates which demonstrated superior fatty acid production with a minimum 2.2 mg l-1d-1in the
initial screen of 41 strains were selected for further study at 10 °C. In addition, at least one strain from
each species was also selected for evaluation of growth at low temperature. The combination of these two
criteria resulted in the down-selection to 19 isolates from 41 (Table 2). With the exception of the lower
cultivation temperature, all other conditions were the same as those used for the initial screen at 22 °C.
For the growth experiments at 10 °C, the selected strains were sub-cultured at the lower temperature for
two weeks before inoculation (Table 2).
Mixotrophic cultivation of top 6 selected strains with MCWW at 10 °C or 22 °C
Based on the results of the screen of 19 isolates grown at 10 and 22 °C (cool and mild
temperature, respectively), 12 strains which showed better growth characteristics under either cool (6
isolates) or mild (6 isolates) temperatures were selected for further study under mixotrophic cultivation
conditions. Two replicate batch cultures of each isolate were grown in 250-ml Erlenmeyer flasks
containing 150 ml of MCWW media for either 10 days (10°C) or 8 days (22°C). The initial light intensity
was approximately 75 µmol photons m-2 s-1 but was increased to 125 µmol photons m-2 s-1 at day 3. During
growth, CO2 was gradually supplemented to the air-stream to a final concentration of 1.5% (v/v) by day 4.
The pH of the culture at the final day was around 8. For mixotrophic cultivation, MCWW was
supplemented with 3 different sources of carbon: glycerol (50 mmol); acetate (14 mmol) or centrate (1%;
centrate was obtained from the Annacis Island Wastewater Treatment Plant, Vancouver, B.C.).
MCWW-nutrient level and biochemical information
Wastewater samples were taken at day 0 and the final day of cultivation to assess the extent of
microalgal mediated N and P removal. Ten mL of culture were filtered using 0.2 μm filter and stored until
analysis. Residual nutrients in the sample were measured using a DR 2800 spectrophotometer (Hach,
Loveland, CO, U.S.A.) with the following commercial assay kits supplied by Hach Co. (CO, USA) :
ammonia-N by the salicylate method with a detection range of 2-47 mg N /L (supplier # TNT832),
phosphorus-P by the ascorbic acid method with a detection range of 0.15-4.5 mg P/L (supplier #
TNT843), total nitrogen by the Kjeldahl method (supplier # TNT880) and low range total organic carbon
(TOC) by direct digestion and oxidation to CO2 with a detection range of 0.3-20 mg/L (supplier #
2760345)The initial nutrient concentrations in MCWW were 20.8 mg l-1 and 2.15 mg l-1 for ammonium

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
and phosphate, respectively. Centrate was diluted to 1% (v/v) in MCWW to give 32.8 mg l-1, 4.46 mg l-1
and 300 mg l-1 for ammonium, phosphate, and total organic carbon, respectively.
Specific growth rate, biomass and lipid-FAMEs production measurement
Specific growth rate (μ, units of d-1) was measured by cell counts from day 1 to day 4, using the
equation:
μ (d-1) = (1)
where N2 and N1 are the cell numbers sampled at times T2 (day 4) and T1 (day 1), respectively.
The biomass productivity (BP) and fatty acid productivity (FAP) were calculated using the equations, as
described by Griffiths and Harrison (2009)
BP (mg l-1d-1) = (2)
FAP (mg l-1d-1) = (3)
where B2 and B1 are the biomass at times T2 and T1, respectively and FA2 and FA1 are the fatty acid
contents of the biomass (in % dry biomass wt.) at times T2 and T1, respectively.
Cells were harvested by filtration using 25 mm Whatman GF/F filters and were freeze dried using
a Freezone 4.5 l Benchtop Freeze Dryer ( Labconco, USA). Mean biomass (n=2) were determined by
gravimetric analysis (Zhu and Lee 1997) and the filtered biomass was used for the analysis of fatty acids.
Fatty acid content in the biomass was determined by direct transesterification to methyl esters (FAMEs)
as previously described (McGinn et al. 2012). The filtered biomass was placed directly into tubes using
1ml of anhydrous toluene (Sigma-aldrich) to extract lipid along with 2 ml of 5% acetyl chloride
(Fluka)/anhydrous methanol (Sigma-Aldrich), heated for 1h at 100°C. Derivatized FAME was washed
with 5 ml of milli-Q water with 5% NaCl and 4 ml of milli-Q water with 2% NaHCO3. The samples were
then made anhydrous with sodium sulfate (Caledon) and evaporated at 37°C under a stream of nitrogen.
Total dried derivatized FAME was diluted to 5 mg ml-1 with hexane and analyzed (Ehimen et al. 2010)
using an Agilent 7890 gas chromatograph with an Omegawax 250 column.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Statistical analysis
For the statistical analysis of each results, we performed a t-test using the GraphPad (GraphPad
Software, Inc. San Diego, CA). Results at the P<0.05 significance level were regarded as statistically
significant.
Results
Strain identification
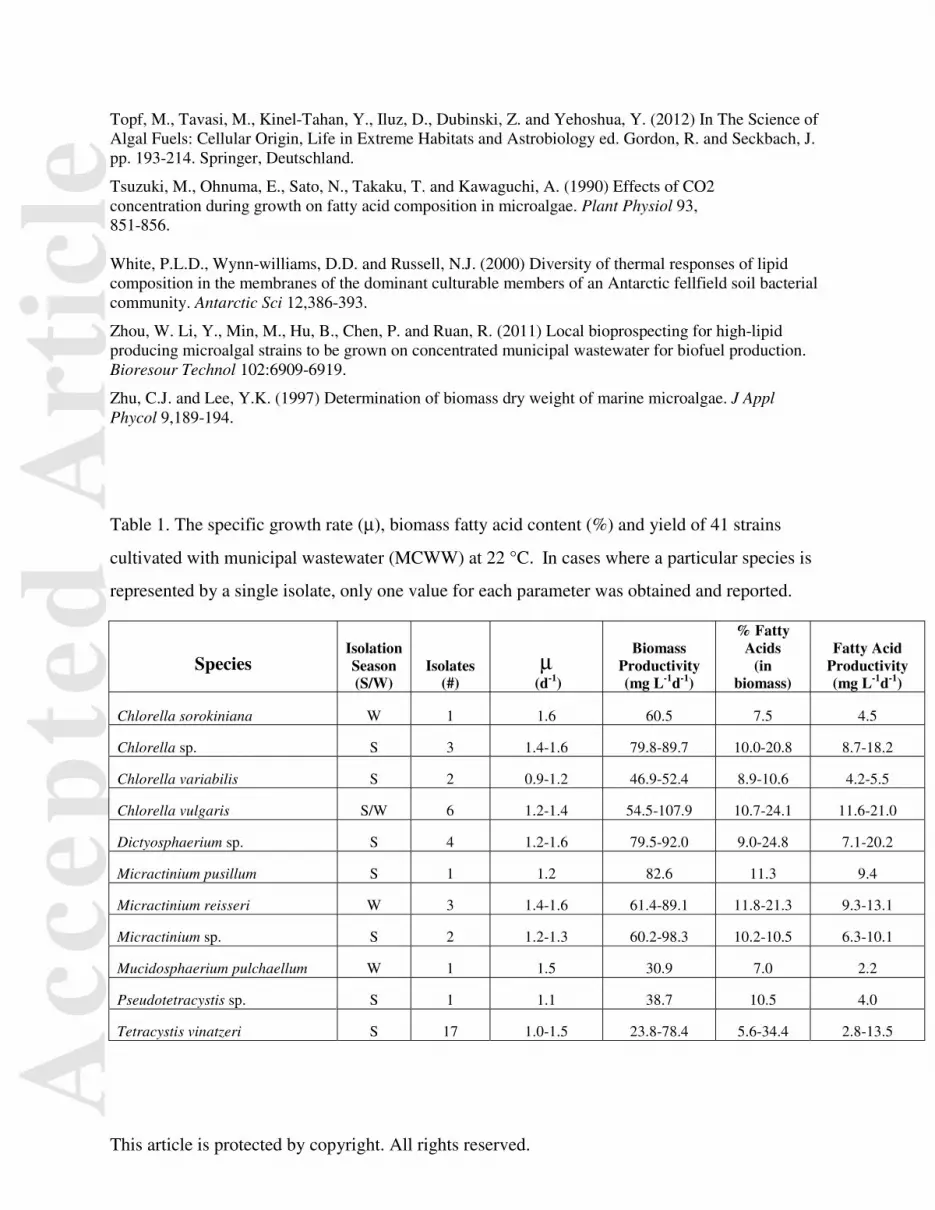
Of the 41 isolates initially screened, a total of 11 microalgae species were identified. Seven of
these identified strains were found in the summer sample only, while three (Chlorella sorokiniania,
Micractinium reisseri and Mucidosphaerium pulchaellum) were found in the winter sample and one
(Chlorella vulgaris) in both samples (Table 1). All of the isolates are freshwater chlorophytes.
Initial screen of 41 strains at 22°C and down-selection of 19 strains at 10°C in MCWW media
Initially, 41 individual isolates were tested for their capacity to grow in MCWW. The specific
growth rates (μ), biomass and fatty acid production during 8 days of cultivation in MCWW at 22°C were
used as the bases of comparison (Table 1). These averaged 1.4±0.2 d-1, 72±22 mg l-1d-1, and 11±6 mg l-1d-
1, respectively (Table 2). Based on the rankings of the final fatty acid yields at day 8, 19 strains were
selected from the 41 and cultivated at 10°C with the same culture conditions used at 22°C.
Based on results after growth at the two temperatures, the 19 strains were organized into three
groups (M, C and N). The six strains having the highest rates of fatty acid productivity at either 22 °C
(p<0.05, six highest strains at 22 °C in comparison with the 13 others) or 10 °C (p<0.05, six highest at 10
°C in comparison with the 13 others) were collected into the M group (for ‘Mild’ temp.) or C group (for
‘Cool’ temp.), respectively (Table 2). The mean rates of fatty acid productivity among the M group were
16 ± 5 and 4 ± 2 mg l-1 d-1 at 22 and 10 °C, respectively, equivalent to a 75% decrease at the lower
temperature. Four of the 6 strains within the M group were isolated from the summer wastewater sample.
The six strains in the C group showed essentially no difference in fatty acid productivity between the two
temperatures. The mean rates among the C group were 9 ± 6 and 8 ± 2 mg l-1 d-1 at 22 and 10 °C,
respectively. Interestingly, four of the 6 strains within the C group were isolated from the winter
wastewater sample. In half of the C-group strains, the rates of biomass and lipid productivity at 10 °C

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
exceeded the corresponding rates at 22 °C (Table 2). This differed from the M-group where biomass and
productivities were always higher at 22 °C. A third group of 7 isolates were identified in which the rates
of fatty acid productivity were generally lower (p=0.06) than the M-group at 22 °C and were significantly
lower (p<0.05) than the C-group at 10 °C (Table 2, N-group for ‘Neither’).
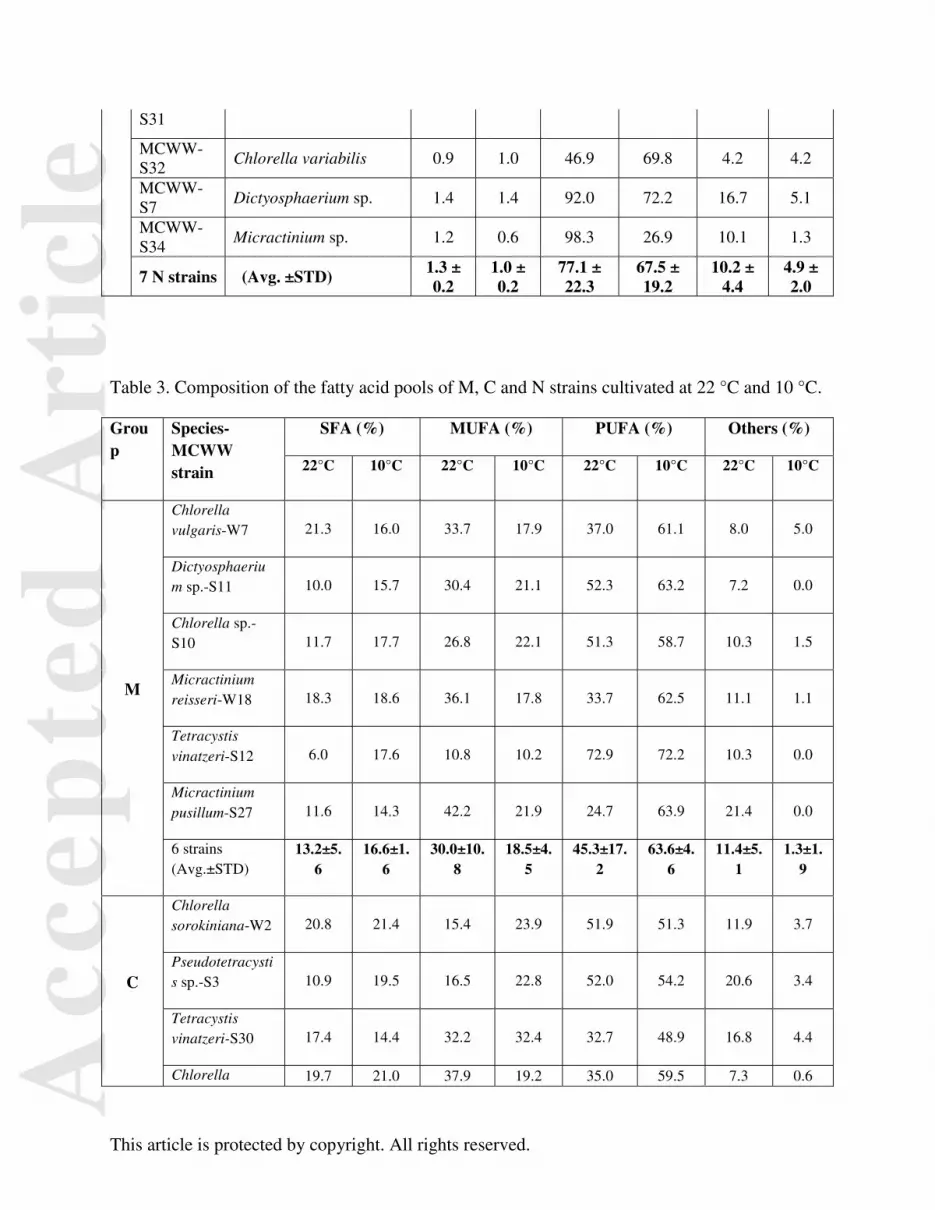
Effect of Growth Temperature on Algal Fatty Acid Profiles
The saturation state of the fatty acid pool appeared to be influenced by growth temperature (Table
3). Irrespective of grouping or growth temperature, the proportion of saturated fatty acids (SFA) was
similar in all isolates and ranged between 13 and 19% of the total pool, on average. Within the M and N
groups, the proportion of MUFAs averaged around 30% when grown at 22°C but significantly decreased
to averages of 19% (M group, p<0.05) and 21% (N group, p<0.05) when grown at 10 °C (Table 3). Most
of this low temperature decrease in MUFAs in M and N groups was due to decreases in C18:1n9 (oleic
acid). In contrast, the proportion of MUFAs in C group isolates was relatively insensitive to growth
temperature, averaging 22% under both conditions. In terms of PUFAs, the proportion in M and N group
isolates increased from averages of 45% and 41% at 22°C to 64% and 60%, respectively, when grown at
10°C. Within the M group, the large relative increase in PUFA at 10°C was mainly due to increases in
two fatty acids, C16:3n3 (hexadecatrienoic acid) and C18:3n3 (α-linolenic acid). The proportion of
PUFAs increased with the shift to low temperature in the C group also, but to a lesser extent, increasing
from 45% to 56%. In general, M group strains responded to the 12 °C drop in growth temperature by re-
organizing the composition of the fatty acid pool to contain fewer MUFAs but more PUFAs. The
composition of fatty acids in C group isolates was largely insensitive to temperature overall, with an
average increase in PUFAs of about 25% at low growth temperature the only difference observed.
Mixotrophic Cultivation of M and C Group Isolates
The M and C group isolates were screened for biomass and fatty acid productivity at 22 and
10°C, respectively, under mixotrophic conditions achieved by supplementing municipal wastewater with
various sources of exogenous organic carbon including glycerol (50 mM), acetate (14 mM) and centrate
wastewater (1% v/v) (Table 4). Unlike previous experiments, the mixotrophic cultures were also
supplemented with high CO2 in the air stream (1.5 % v/v) to maximize that the rate of carbon supply. In
wastewater aerated with high CO2 but no organic C supplementation, mean biomass and fatty acid
productivities from M-group isolates were 326±92 and 104±57 mg l-1 d-1 , respectively, and from the C-

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
group 168±47 and 44±9 mg l-1 d-1 (Table 4). Surprisingly, we observed generally negative effects on
biomass and fatty acid productivity of mixotrophic supplementation on M-group strains grown at 22 °C.
In particular, the addition of glycerol to M group cultures caused a significant decrease (p < 0.5) in
biomass productivity compared to the autotrophic control (Table 4). Problems with pH control after
addition of acetate to M-group isolates appeared to inhibit growth thereby preventing any opportunity to
obtain data from that treatment. Mixotrophic supplementation did not significantly increase the mean
biomass or fatty acid productivities in C group cultures (Table 4). However, there appeared to be strain
dependent effects. Addition of glycerol, acetate or centrate to C group cultures grown at 10 °C improved
biomass and lipid productivities in two, two and 5 strains, respectively, compared to the autotrophic
control. The biomass productivities of two C group strains in particular, Mucidosphaerium pulchaellum
and Pseudotetracyctis sp. were always increased under mixotrophic conditions compared to control
(Table 4). Biomass productivity in Mucidosphaerium pulchaellum increased by 63%, 66% and 57% under
supplementation with glycerol, acetate or centrate, respectively, compared to control. In Pseudotetracyctis
sp., the increases were 91%, 11% and 24%, respectively.
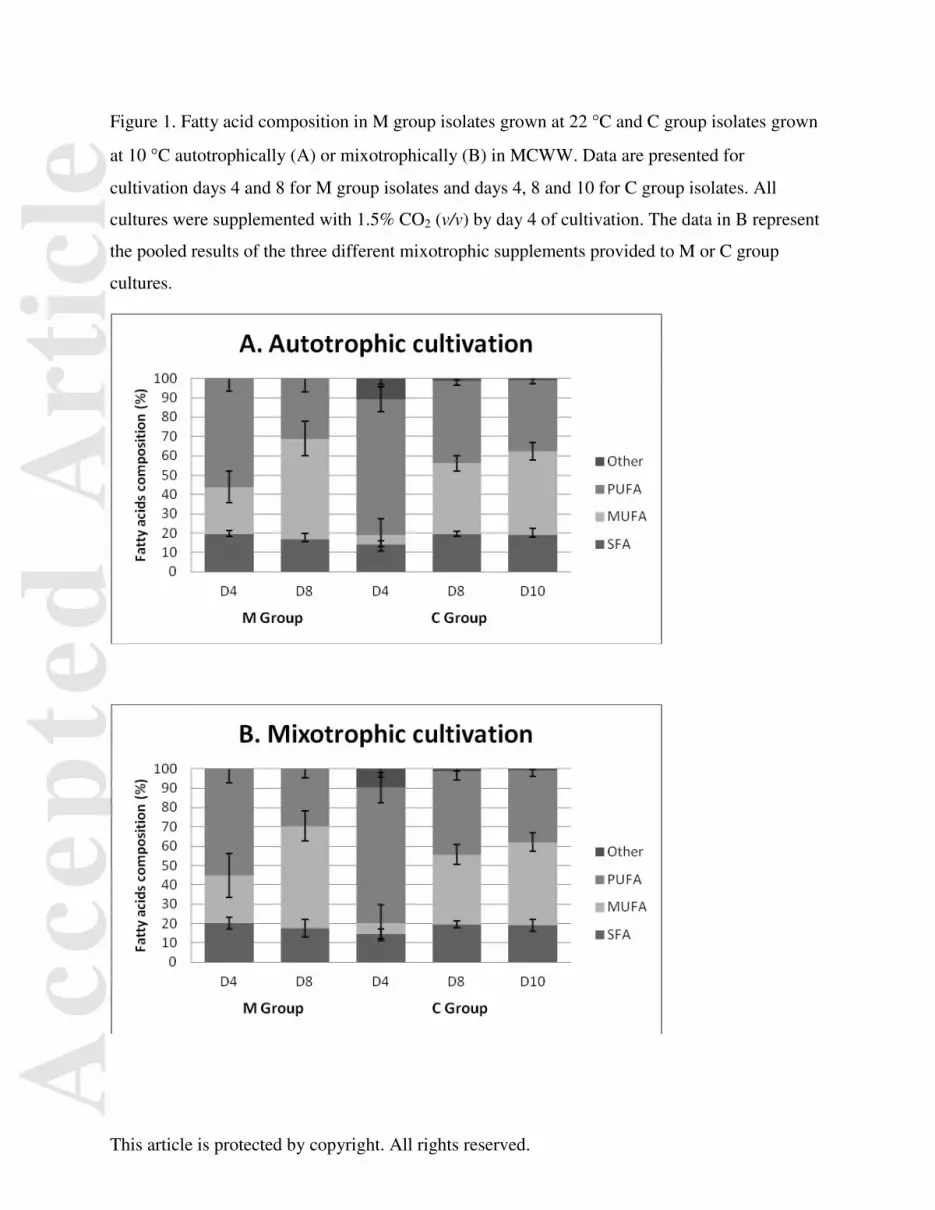
Effects of Culture Age on Fatty Acid Pools in M and C Group Isolates
The saturation states of the fatty acid pools were also examined in M and C group isolates grown
mixotrophically (Fig. 1). In both M and C groups, the fatty acid profiles were very similar in
mixotrophically grown cells compared to controls grown autotrophically in unamended wastewater (Fig
1.A and B). In all treatments and groups, there was an apparent effect of culture age on the fatty acid
profiles which was clearly more significant than culture mode. As the cultures grew, the proportion of
MUFAs increased and PUFAs decreased while saturated fatty acids remained virtually unchanged. In M-
group isolates, the fatty acid profiles changed significantly between days 4 and 8 of cultivation, regardless
of culture mode, becoming more enriched in MUFAs and less in PUFAs by day 8 (Fig. 1A and B). A
similar trend was observed in C-group isolates in both auto- and mixotrophic modes between days 4, 8
and 10 of cultivation, although by day 4, C-group strains were significantly more enriched in PUFAs
compared to the M-group (p<0.05).
Discussion
The objective of this study was to exploit the presumed seasonal variability in natural microalgae
assemblages in municipal wastewater in order to isolate strains best suited for remediation and biofuel
production in either warmer or cooler seasons. Through a down-selection process conducted with 19

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
strains at 22 °C and 10°C, twelve strains with the highest rates of fatty acid productivity at either the
higher or lower temperature were collected into two groups of six isolates designated the M-group and C-
group, respectively (Table 2). Of the C-group strains, five strains had biomass productivity rates which
exceeded 70 mg l-1 d-1 and all six displayed rates of fatty acid productivity of at least 6 mg l-1 d-1 at 10°C.
Of the M-group strains grown at 10 °C, only three strains exceeded 70 mg l-1 d-1 with two strains at < 40
mg l-1 d-1 of biomass productivity with only two strains exceeding 6 mg l-1 d-1 for fatty acid productivity.
Furthermore, in three of the six C-group strains, biomass and fatty acid productivity were actually higher
at 10°C than at 22°C (Table 2). All of the isolates in the M-group were more productive, both in terms of
biomass and fatty acids, at the higher temperature. These data suggested that the C-group strains were
better suited than M-group strains for growth and wastewater remediation at 10°C compared to 22°C and
that the M-group strains were better suited for 22 °C. These suppositions make sense in light of the fact
that of the six C-group strains, four were isolated from the winter wastewater sample, while of the six M-
group strains, four were isolated from the summer sample. Consistent with this, the most productive M-
group strain grown at 10 °C was Chlorella vulgaris-W7, an isolate from the winter sample (Table 2).
Upon supplementation with up to 1.5 (v/v) % CO2 to the M-group cultures at 22 °C, there was an increase
in biomass and fatty acid productivity of 4.3-fold and 6.5-fold on average, respectively (Table 4). In
contrast, addition of CO2 to C-group cultures at 10 °C increased biomass and fatty acid productivity by
only 2.2-fold and 5.5-fold, on average. The biomass productivities in M and C group isolates at their
respective temperatures were similar to those reported by others from strains grown in similar
wastewaters (Li et al. 2011, Zhou et al. 2011, Song et al. 2013). Supplementation with organic carbon for
mixotrophic growth did not generally enhance productivities in M-group cultures at 22°C. Mixotrophy
proved to be a more effective strategy in C-group cultures at 10°C for enhancing biomass and fatty acid
productivity. The highest rates of biomass and fatty acid productivity among M-group strains were 408
and 163 mg l-1 d-1, respectively in Dictyosphaerium sp. S-11 grown in unamended MCWW supplemented
with 1.5% CO2 (Table 4). Amongst C-group strains, Micractinium reisseri-W12 displayed the highest rate
of biomass productivity of 282 mg l-1 d-1 while Chlorella vulgaris-W8 had the highest rate of fatty acid
productivity at 75 mg l-1 d-1, both from MCWW supplemented with centrate and high CO2 (Table 4).
Previous research has demonstrated that microalgae isolated from wastewater tend to grow well
in it (Li et al. 2011; Park et al. 2011; Zhou et al. 2011). Microalgae strains most commonly isolated and
studied for wastewater research typically come from the following genera: Chlamydomonas, Chlorella,
Chlorococcum, Micractinium and Scenedesmus. Not surprisingly, much of the research looking at
applications of microalgae to wastewater was conducted using species of the Chlorella clade (Li et al.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
2011, Park et al. 2012). In this study, as in other reports, we also isolated members of the Chlorella
clade, such as Chlorella and Micractinium species. In addition to these strains however, we also isolated
several strains belonging to the Parachlorella clade such as Dictyosphaerium sp. and Mucidosphaerium
pulchaellum as well as two unusual strains, Pseudotetracystis sp. and Tetracystis vinatzeri, that are not
commonly associated with wastewater treatment and biofuel research. In fact, T. vinatzeri was the most
common strain of microalgae isolated from the summer wastewater sample.
Temperature is one of the most important factors affecting algal lipid production and composition
as well as growth rate and biomass production (Topf et al. 2012). The main outcome of this study is the
finding that metabolic flexibility of microalgae enables isolation of apparently cold-adapted strains of
microalgae that are better suited to remediation of wastewater at 10 °C than strains isolated from warmer
wastewaters. In the strains under investigation here, cold-temperature adapted strains showed an overall
greater insensitivity to growth temperature in their fatty acid profiles, compared to warm-temperature
adapted strains, suggesting a causal link between temperature acclimation and lipid metabolism in
microalgae.
There was a significant effect of CO2 supplementation on biomass (p<0.05) and fatty acid
productivity (p<0.05) in the M-group strains (compare Table 2 to Table 4). This was not so apparent from
the growth rates, which were only slightly greater in CO2-supplemented cultures. However, exponential
growth rates were obtained from days 1-3 of growth, where CO2 was unlikely to have been limiting. The
effect of high CO2 was much more apparent on biomass productivity rates (estimated as the rate of
biomass accumulation between days 4-7), which suggests a likely carbon limitation of growth from day 4
onwards in cultures without CO2 supplementation. The average pH of cultures collected on the final day
(day 8) were significantly different than the pH of cultures without CO2 supplementation (p<0.05) . At the
final day of cultivation with CO2 supplementation, average pH of M group was 7.08 ± 0.29; while, it was
9.37 ± 0.53 when they were cultivated without CO2 supplementation. A similar effect of high CO2 was
also observed in C-group strains. The composition of the fatty acid pool was noticeably affected by CO2
supplementation and affected M and C group strains in the same way. Under high CO2 supplementation,
the fatty acid pools in both groups were relatively more enriched in MUFA and depleted in PUFA,
compared to cultures of the same age and growth temperature yet with no CO2 supplementation (Fig. 1A
compared to Table 3). This is consistent with other reports which have suggested that CO2 levels
experienced during growth can affect the saturation state of the fatty acid pool in microalgae. Tsuzuki et
al. (1990) showed a higher proportion of saturated fatty acids upon a shift from low to high CO2 in

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Chlorella vulgaris and Chlamydomonas reinhardtii cultures. Sato (1989) reported a similar effect in C.
reinhardtii when shifted from low to high CO2. Riebesell et al. (2000) reported a higher proportion of
saturated fatty acids and MUFAs, relative to PUFAs, in the marine coccolithophore Emiliania huxleyi
grown at high CO2 under N-replete conditions.
From the results of our previous study (Park et al., 2012) we found that microalgae grow
photoautotrophically in MCWW, due to the near absence of organic carbon in secondary wastewater
effluent. Recent studies (Zhou et al. 2011; Li et al. 2012; Park et al. 2012) proved that SMW
supplemented with organic carbon sources could increase biomass and lipid production for some
microalgae strains. In this study, we had mixed success with mixotrophic strategies aimed at enhancing
biomass and fatty acid productivity. The benefit was more apparent in C-group strains cultivated at 10 °C,
in which 5 of 6 strains responded with either increased biomass production or fatty acid production or
both; only one M-group showed any kind of positive effect (Table 4). In particular, the C-group strain
Mucidospaerium pulchaellum-W1 showed a significant increase in fatty acid productivity (53-73%) from
all three external carbon sources (Table 4). Grown with acetate supplementation, this strain had a rate of
fatty acid productivity of 68 mg l-1d-1, just behind C. vulgaris-W8 in MCWW supplemented with centrate
at 75 mg l-1d-1 (Table 4). However, this increase and the increases seen with supplementation with other
carbon sources were not correlated with any alterations in fatty acid profiles. In our previous study (Park
et al. 2012), we found that microalgae grown in a mixotrophic TAP medium were more productive and
showed different fatty acid profiles compared to the autotrophic growth media. Thus, merely adding
organic carbon to MCWW may not constitute a true mixotrophic growth mode, as it may not fully engage
heterotrophic assimilatory pathways as it appears to with growth on TAP media. Mixotrophic growth is
typically conducted under low growth light intensities (Read et al. 1989), but light intensities in our
experiments were 125 μE m-2 s-1, which may have been too high for effective mixotrophic assimilation of
carbon substrates. In addition, CO2 supplementation may have masked any mixotrophic responses. It has
been shown, for instance, that some microalgae can only assimilate glycerol as a carbon source in the
presence of light and air levels of CO2 for growth (Ingram et al. 1973; Kaplan et al. 1986).
Returning to the effects of experimental treatments on fatty acids, under no conditions did
mixotrophic enhancement noticeably alter the fatty acid profile (Fig. 1 A compared to B). Fatty acid
profiles were more directly influenced by culture age rather than external supplementation with organic
carbon substrates. In general, the proportion of SFAs changed little during growth; MUFA and PUFA
composition were opposite each other. In M and C group strains, PUFA composition during exponential

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
growth was maximum and MUFA composition was minimum, while the opposite was true at stationary
phase. This result is consistent with other studies that reported low-temperature adapted or acclimated
microorganisms accumulate PUFA in membrane lipids to maintain membrane fluidity during periods of
rapid growth (White et al. 2000; Dalsgaard et al. 2003).
The saturation state of triacylglycerol in microalgae is important because it influences the
properties of the biodiesel fuel which could potentially be produced from it. Indeed, the presence of
saturated and monounsaturated fatty acids is considered an important factor when screening strains for
biodiesel production (Ramos et al. 2009; Song et al. 2013; Talebi et al. 2013). Fatty acid compositions of
microalgae differ from species to species, and also depend on growth conditions and culture age.
Generally, C14:0, C16:0, C16:1n-7, C18:1n-9, C18:2n-6, C18:3n-3, C20:5n-3 constitute the major fatty
acid components of microalgal lipids (Pratoomyot et al. 2005; Song et al. 2013; Talebi et al. 2013).
Palmitic (C16:0) and oleic (C18:1n9) acids, are major fatty acid components of vegetable (Ramos et al.
2009) and microalgal triacylglycerides (Danielewicz et al. 2011; Talebi et al. 2013). We also found that
palmitic and oleic acids were major components in SFA and MUFA, respectively, of both M- and C-
group isolates (data not shown). Palmitic acid was very conspicuous across strains and persisted,
irrespective of culture age or growth temperature. On the other hand, oleic acid was highly inducible in
response to culture conditions (data not shown).
For large-scale wastewater treatment and biofuel production scenarios, microalgae need to be
able to grow despite the presence of bacteria and fungi. The strains of microalgae studied here were
isolated from wastewater and, therefore, we expect that these candidate strains are capable of growing
under such conditions. However, there has been concern that using a mixotrophic strategy to boost
production in wastewater is not a practical process, even though algal wastewater treatment itself is
considered a quite worthwhile endeavor (Benemann et al. 2012). In this study, we experienced fungal
contamination in the Pseudotetracyctis sp-S3 strain cultivated in MCWW supplemented with glycerol. It
is because of these findings that further study of the interactions between selected microalgae strains and
endogenous microbial contaminants present in wastewater system is warranted.
Acknowledgements
We would like to thank staff members Dereck Avery, Shawn Taylor and Nigel Crouse of Mill
Cove Wastewater Treatment Plant, Bedford, Nova Scotia, for their kind support and access to secondary
effluent. We would also like to thanks Kelly Soanes (NRC-Halifax) for the helpful comments which

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
improved the manuscript. This work was supported by NRC Canada’s flagship program in Algal Carbon
Conversion.
Conflict of Interest
The authors declare that there are no conflicts of interest.
References
Benemann, J., Woertz, I. and Lundquist, T. (2012) Life cycle assessment for microalgae oil production. Disruptive Science and Technology 1,68-78.
Benson, D.A., Karsch-Mizrachi, I., Lipman, D.J., Ostell, J., Rapp, B.A. and Wheeler, D.L. (2000) GenBank. Nucleic Acids Res 28,15-18.
Bold, H.C. (1949) The morphology of Chlamydomonas chlamydogama, sp. nov. Bull. Torrey Bot. Club 76,101-108.
Cerón-García, M.C., García Camacho, F., Sánchez Mirón, A., Fernández, J.M., Chisti, Y. and Molina Grima, E. (2006) Mixotrophic production of marine microalga Phaeodactylum tricornutum on various carbon sources. J Microbiol Biotechnol 16,689-694.
Danielewicz, M.A., Anderson, L.A. and Franz, A.K. (2011) Triglycerol profiling of marine microalgae by mass spectrometry. J Lipid Res 52,2101-2108.
Dalsgaard, J., St John, M., Kattner, G., Muller-Navarra, D. and Hage, W. (2003) Fatty acid trophic markers in the pelagic marine environment. Adv Mar Biol 46,225-340.
Ehimen, E.A., Sun, Z.F. and Carrington, C.G. (2010) Variables affecting the in situ transesterification of microalgae lipids. Fuel 89,677-684.
Guschina, I.A. and Harwood, J.L. (2006) Lipids and lipid metabolism in eukaryotic algae. Prog Lipid Res 45,160-186.
Griffiths, M.J. and Harrison, S.T.L. (2009) Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J Appl Phycol 21,493-507.
Ingram, L.O., Van Baalen, C. and Calder, J.A. (1973) Role of reduced exogenous organic compounds in the physiology of the blue-greeen bacteria (algae): photoheterotrophic growth of an “autotrophic” blue-green bacterium. J Bacteriol 114,701-705.
Jiang, H. and Gao, K. (2004) Effects of lowering temperature during culture on the production of polyunsaturated fatty acids in the marine diatom Phaeodactylum tricornutum (bacillariophyceae). J Phycol 40,651-654.
Joh, T., Yoshida, T., Yoshimoto, M., Miyamoto, T. and Hatano, S. (1993) Composition and positional distribution of fatty acids in polar lipids from Chlorella ellipsoidea differing in chilling susceptibility and frost hardiness. Physiol Plant 89,285-290.
Kaplan, D., Richmond, A.E., Dubinsky, Z. and Aaronson, S. (1986) In Handbook for Microalgal Mass Culture ed. Richmond, A. pp. 147-198. CRC Press, Boch Raton, FL., USA.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Li, Y., Zhou, W., Hu, B., Min, M., Chen, P. and Ruan, R.R. (2011) Integration of algae cultivation as biodiesel production feedstock with municipal wastewater treatment: strains screening and significance evaluation of environmental factors. Biores Technol 102,10861-10867.
McGinn, P.J., Dickinson, K.E., Bhatti, S., Frigon, J.C., Guiot, S.R., O'Leary, S.J.B. (2011) Integration of microalgae cultivation with industrial waste remediation for biofuel and bioenergy production: opportunities and limitations. Photosyn Res 109, 231–247. McGinn, P.J., Dickinson, K.E., Park, K.C., Whitney, C.G., MacQuarrie, S.P., Black, F.J., Frigon, J.C., Guiot, S.R. and O'Leary SJ (2012) Assessment of the bioenergy and bioremediation potentials of the microalga Scenedesmus sp. AMDD cultivated in municipal wastewater effluent in batch and continuous mode. Algal Research 1, 155-165.
McLarnon-Riches, C.J., Rolph, C.E., Greenway, D.L.A. and Robinson, P.K. (1998) Effects of environmental factors and metals on Selenastrum capriornutum lipids. Phytochemistry 49,1241-1247.
Park JBK, Craggs RJ, Shilton AN (2011) Wastewater treatment high rate algal ponds for biofuel production. Biores Technol 102:35-42
Park, K.C., Whitney, C., McNichol, J., Dickinson, K., MacQuarrie, S., Skrupski, B.P., Zhou, J., Wilson, K., O’Leary, S.J.B. and McGinn, P.J. (2012) Mixotrophic and photoautotrophic cultivation of fourteen microalgae isolates from Saskatchewan, Canada: potential applications for wastewater remediation for biofuel production. J Appl Phycol 24,339-348.
Perez-Garcia, O., Escalante, F.M.E., de-Bashan, L.E. and Bashan, Y. (2011) Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res 45,11-36.
Pittman, J.K., Dean, A.P. and Osundeko, O. (2011) The potential of sustainable algal biofuel production using wastewater resources. Biores Technol 102,17-25.
Pratoomyot, J., Srivilas, P. and Noiraksar, T. (2005) Fatty acids composition of 10 microalgal species. Songklanakarin J Sci Technol 27,1179-1187.
Ramos, M.J., Fernández, C.M., Casas, A., Rodríguez, L. and Pérez, Á. (2009) Influence of fatty acid composition of raw materials on biodiesel properties. Biores Technol 100,261-268.
Read, H.S., Reads, S. and Park, B. (1989) The estimation of algal yield parameters associated with mixotrophic and photoheterotrophic growth under batch cultivation. Biomass 18,153-160.
Riebesell, U., Revill, A.T., Holdsworth, D.G. and Volkman, J. (2000) The effects of varying CO2 concentration on lipid composition and carbon isotope fractionation in Emiliania huxleyi. Geochim Cosmochim Acta 64, 4179–4192.
Sato, N. (1989) Modulation of lipid and fatty acid content by carbon dioxide in Chlamydomonas reinhardtii. Plant Sci 61,17-21.
Song, M., Pei, H., Hu, W. and Ma, G. (2013) Evaluation of the potential of 10 microalgal strains for biodiesel production. Bioresour Technol 141,245-251.
Talbot, P. and de la Noue, J. (1993) Tertiary treatment of wastewater with Phormidium bohneri (Schmidle) under various light and temperature condition. Water Res 27,153-159.
Talebi, A.F., Mohtashami, S.K., Tabatabaei, M., Tohidfar, M., Bagheri, A., Zeinalabedini, M., Mirzaei, H.H., Mirzajanzadeh, M., Shafarroudi, S.M. and Bakhtiari, S. (2013) Fatty acids profiling: A selective criterion for screening microalgae strains for biodiesel production. Algal Research 2,258-267.
Thompson, G.A. (1996) Lipids abd membrane function in green algae. Biochim Biophys Acta 1302,17-45.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Topf, M., Tavasi, M., Kinel-Tahan, Y., Iluz, D., Dubinski, Z. and Yehoshua, Y. (2012) In The Science of Algal Fuels: Cellular Origin, Life in Extreme Habitats and Astrobiology ed. Gordon, R. and Seckbach, J. pp. 193-214. Springer, Deutschland.
Tsuzuki, M., Ohnuma, E., Sato, N., Takaku, T. and Kawaguchi, A. (1990) Effects of CO2 concentration during growth on fatty acid composition in microalgae. Plant Physiol 93, 851-856. White, P.L.D., Wynn-williams, D.D. and Russell, N.J. (2000) Diversity of thermal responses of lipid composition in the membranes of the dominant culturable members of an Antarctic fellfield soil bacterial community. Antarctic Sci 12,386-393.
Zhou, W. Li, Y., Min, M., Hu, B., Chen, P. and Ruan, R. (2011) Local bioprospecting for high-lipid producing microalgal strains to be grown on concentrated municipal wastewater for biofuel production. Bioresour Technol 102:6909-6919.
Zhu, C.J. and Lee, Y.K. (1997) Determination of biomass dry weight of marine microalgae. J Appl Phycol 9,189-194.
Table 1. The specific growth rate (μ), biomass fatty acid content (%) and yield of 41 strains
cultivated with municipal wastewater (MCWW) at 22 °C. In cases where a particular species is
represented by a single isolate, only one value for each parameter was obtained and reported.
Species
Isolation Season (S/W)
Isolates (#)
μ (d-1)
Biomass Productivity (mg L-1d-1)
% Fatty Acids
(in biomass)
Fatty Acid Productivity (mg L-1d-1)
Chlorella sorokiniana W 1 1.6 60.5 7.5 4.5
Chlorella sp. S 3 1.4-1.6 79.8-89.7 10.0-20.8 8.7-18.2
Chlorella variabilis S 2 0.9-1.2 46.9-52.4 8.9-10.6 4.2-5.5
Chlorella vulgaris S/W 6 1.2-1.4 54.5-107.9 10.7-24.1 11.6-21.0
Dictyosphaerium sp. S 4 1.2-1.6 79.5-92.0 9.0-24.8 7.1-20.2
Micractinium pusillum S 1 1.2 82.6 11.3 9.4
Micractinium reisseri W 3 1.4-1.6 61.4-89.1 11.8-21.3 9.3-13.1
Micractinium sp. S 2 1.2-1.3 60.2-98.3 10.2-10.5 6.3-10.1
Mucidosphaerium pulchaellum W 1 1.5 30.9 7.0 2.2
Pseudotetracystis sp. S 1 1.1 38.7 10.5 4.0
Tetracystis vinatzeri S 17 1.0-1.5 23.8-78.4 5.6-34.4 2.8-13.5

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Table 2. Specific growth rate (μ), biomass and fatty acid productivities at 22 °C and 10 °C for 19
microalgae strains isolated from MCWW. The abbreviations M and C indicate selection for mild
(M) and cool (C) temperature cultivation while N represents strains not selected for further
consideration.
Strain Designation
Species
μ (d-1)
Biomass Productivity (mg L-1 d-1)
Fatty Acid Productivity (mg L-1 d-1)
22 °C 10 °C 22 °C 10 °C 22 °C 10 °C
M
MCWW-W7
Chlorella vulgaris 1.4 1.1 92.9 83.5 21.0 6.1
MCWW-S11
Dictyosphaerium sp. 1.6 0.9 81.8 65.7 20.2 4.7
MCWW-S10
Chlorella sp. 1.6 1.2 88.2 74.6 18.2 6.5
MCWW-W18
Micractinium reisseri 1.6 0.9 61.4 37.9 13.1 2.5
MCWW-S12
Tetracystis vinatzeri 1.5 0.8 39.3 33.6 13.5 2.4
MCWW-S27
Micractinium pusillum 1.2 0.9 82.6 70.8 9.3 4.7
6 M strains (Avg. ±STD) 1.5 ±
0.1 1.0 ± 0.2
74.3 ± 20.3
61.0 ± 20.5
15.9 ± 4.6
4.5 ± 1.7
C
MCWW-W2
Chlorella sorokiniana 1.6 1.1 60.5 73.7 4.5 11.5
MCWW-S3
Pseudotetracystis sp. 1.1 1.2 38.7 79.9 4.0 9.2
MCWW-S30
Tetracystis vinatzeri 1.3 1.0 78.4 73.9 11.3 7.3
MCWW-W8
Chlorella vulgaris 1.4 1.2 83.9 81.6 18.6 6.6
MCWW-W12
Micractinium reisseri 1.5 1.1 89.1 85.6 10.4 6.5
MCWW-W1
Mucidosphaerium pulchaellum
1.5 1.1 30.9 67.8 2.2 6.3
6 C strains (Avg. ±STD) 1.4 ± 0.2
1.1 ± 0.1
63.6 ± 24.4
77.1 ± 6.5
8.5 ± 6.1
7.9 ± 2.1
N
MCWW-S24
Chlorella sp. 1.5 1.1 89.7 87.5 9.0 7.3
MCWW-S2
Chlorella vulgaris 1.2 1.1 62.6 75.6 13.2 6.3
MCWW-S28
Chlorella vulgaris 1.4 1.0 97.5 64.9 12.4 3.9
MCWW- Chlorella variabilis 1.2 1.1 52.4 75.5 5.5 6.3
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
S31
MCWW-S32
Chlorella variabilis 0.9 1.0 46.9 69.8 4.2 4.2
MCWW-S7
Dictyosphaerium sp. 1.4 1.4 92.0 72.2 16.7 5.1
MCWW-S34
Micractinium sp. 1.2 0.6 98.3 26.9 10.1 1.3
7 N strains (Avg. ±STD) 1.3 ± 0.2
1.0 ± 0.2
77.1 ± 22.3
67.5 ± 19.2
10.2 ± 4.4
4.9 ± 2.0
Table 3. Composition of the fatty acid pools of M, C and N strains cultivated at 22 °C and 10 °C.
Group
Species-MCWW strain
SFA (%) MUFA (%) PUFA (%) Others (%)
22°C 10°C 22°C 10°C 22°C 10°C 22°C 10°C
M
Chlorella vulgaris-W7 21.3 16.0 33.7 17.9 37.0 61.1 8.0 5.0
Dictyosphaerium sp.-S11 10.0 15.7 30.4 21.1 52.3 63.2 7.2 0.0
Chlorella sp.-S10 11.7 17.7 26.8 22.1 51.3 58.7 10.3 1.5
Micractinium reisseri-W18 18.3 18.6 36.1 17.8 33.7 62.5 11.1 1.1
Tetracystis vinatzeri-S12 6.0 17.6 10.8 10.2 72.9 72.2 10.3 0.0
Micractinium pusillum-S27 11.6 14.3 42.2 21.9 24.7 63.9 21.4 0.0
6 strains (Avg.±STD)
13.2±5.6
16.6±1.6
30.0±10.8
18.5±4.5
45.3±17.2
63.6±4.6
11.4±5.1
1.3±1.9
C
Chlorella sorokiniana-W2 20.8 21.4 15.4 23.9 51.9 51.3 11.9 3.7
Pseudotetracystis sp.-S3 10.9 19.5 16.5 22.8 52.0 54.2 20.6 3.4
Tetracystis vinatzeri-S30 17.4 14.4 32.2 32.4 32.7 48.9 16.8 4.4
Chlorella 19.7 21.0 37.9 19.2 35.0 59.5 7.3 0.6

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
vulgaris-W8
Micractinium reisseri-W12 21.8 16.4 18.6 21.4 44.0 56.3 13.8 6.0
Mucidosphaerium pulchaellum-W1
21.0 18.4 15.2 13.8 51.8 63.2 12.9 4.6
6 strains (Avg.±STD)
18.6±4.1
18.5±2.7
22.7±9.9 22.3±6.
1 44.6±8.9
55.6±5.3
13.9±4.5
3.8±1.8
N
Chlorella sp.-S24 15.5 12.8 25.8 25.6 44.0 54.4 14.7 7.1
Chlorella vulgaris-S2 17.8 15.9 35.5 22.0 37.6 61.2 10.5 1.8
Chlorella vulgaris-S28 22.1 14.4 20.7 17.9 40.4 67.1 16.8 1.1
Chlorella variabilis-S31 18.3 13.7 30.4 22.4 44.9 55.1 7.0 8.8
Chlorella variabilis-S32 17.8 14.2 23.6 21.7 51.0 56.5 7.6 8.4
Dictyosphaerium sp.-S7 12.7 15.6 31.9 19.1 44.4 64.4 11.0 0.8
Micractinium sp.-S34 12.9 19.0 54.5 19.3 23.1 58.9 9.6 5.6
7 strains (Avg.±STD)
16.7±3.3
15.1±2.0
31.8±11.2
21.1±2.6
40.8±8.8 59.7±4.
8 11.0±3.
6 4.8±3.
5

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Table 4. Specific growth rates (μ, d-1), biomass (BAP) and fatty acid (FAP) productivities of M
and C group isolates grown under mixotrophic conditions: MCWW (control); MCWW-G
(MCWW with 50 mM glycerol); MCWW-A (MCWW with 15 mM acetate); MCWW-C
(MCWW with 1% centrate wastewater).
M Group MCWW MCWW-G MCWW-A MCWW-C
μ BP FAP μ BP FAP μ BP FAP μ BP FAP
Dictyosphaerium sp.-S11 1.7 407.8 162.5 1.7 219.0 91.9 n.d. n.d. n.d. 1.7 344.7 133.8
Chlorella sp.-S10 1.7 391.2 146.7 1.7 276.9 108.8 n.d. n.d. n.d. 1.7 341.8 119.1 Chlorella vulgaris-
W7 1.5 280.3 129.0 1.5 176.5 68.8 n.d. n.d. n.d. 1.5 288.0 118.1 Micractinium reisseri-W18 1.7 324.6 113.9 1.8 109.0 55.9 n.d. n.d. n.d. 1.8 226.9 97.0 Micractinium pusillum-S27 1.6 195.5 50.5 1.7 227.4 55.6 n.d. n.d. n.d. 1.4 199.9 44.3 Tetracystis
vinatzeri-S12 1.4 310.9 19.1 1.6 146.9 20.9 n.d. n.d. n.d. 1.4 305.3 12.2 6 strains
(Avg±STD) 1.6±0.1 318.4±77.4 103.6±56.6 1.7±0.1
192.6±60.6
67.0±30.8 n.d. n.d. n.d. 1.6±0.2 284.3±59.9 87.4±48.4
C Group MCWW MCWW-G MCWW-A MCWW-C
μ BP FAP μ BP FAP μ BP FAP μ BP FAP
Chlorella vulgaris-W8 1.4 145.1 59.2 1.4 144.6 62.3 1.4 139.3 57.6 1.3 207.3 74.7
Tetracystis vinatzeri-S30 1.3 228.8 44.4 1.3 207.8 45.6 1.4 176.5 38.4 1.3 218.5 45.1 Micractinium reisseri-W12 1.3 216.5 43.9 1.4 204.7 40.8 1.3 209.2 44.1 1.3 281.7 55.3
Chlorella sorokiniana-W2 1.2 110.5 42.5 1.2 113.9 39.6 1.2 104.6 40.9 1.3 140.2 53.0
Mucidosphaerium pulchaellum-W1 1.2 135.8 39.0 1.2 220.6 59.7 1.2 226.4 67.5 1.2 213.4 60.7 Pseudotetracystis
sp.-S3 1.4 169.8 33.5 1.3 324.6 49.6 1.3 187.7 39.5 1.3 210.7 31.1 6 strains
(Avg±STD) 1.3±0.1 167.8±46.8 43.7±8.6 1.3±0.1 202.7±72.8 49.6±9.6 1.3±0.1 173.9±45.2 48.0±11.9 1.3±0.0 212±44.9 53.3±14.7
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Figure 1. Fatty acid composition in M group isolates grown at 22 °C and C group isolates grown
at 10 °C autotrophically (A) or mixotrophically (B) in MCWW. Data are presented for
cultivation days 4 and 8 for M group isolates and days 4, 8 and 10 for C group isolates. All
cultures were supplemented with 1.5% CO2 (v/v) by day 4 of cultivation. The data in B represent
the pooled results of the three different mixotrophic supplements provided to M or C group
cultures.




















