
Schistosomamansoni gene GP22 encodes the tegumental antigen Sm25: (1) antibodies to a predicted...
15
Schistosoma mansoni gene GP22 encodes the tegumental antigen Sm25: (1) antibodies to a predicted B-cell epitope of Sm25 cross-react with other candidate vaccine worm antigens; (2) characterization of a recombinant product containing tandem-repeats of this peptide as a vaccine MARY M. PETZKE 1 , PARMJEET K. SURI 1 , RICHARD BUNGIRO 1 , MIRIAM GOLDBERG 1 , SARAH F. TAYLOR 1 , SUMANT RANJI 1 , HARRY TAYLOR 2 , JOSEPH W.MCCRAY JR 2 & PAUL M. KNOPF 1 1 Department of Molecular Microbiology and Immunology, Brown University, Providence, RI 02912, 2 Department of Biology, Morehouse College, Atlanta, GA, USA SUMMARY Monospecific antibodies against two putative epitopes of schistosome protein encoded by gene GP22 (182 codons, no introns) were used to probe worm extracts fractionated by lentil-lectin affinity chromatography or by electrophoresis. Anti-peptide-a (codons 70–84) exclusively identifies the N- glycanase-sensitive, 25 kDa tegumental glycoprotein Sm25 in the lectin-bound fraction of detergent-solubilized adult worm extract S3. In contrast, antipeptide-d (codons 151– 162) does not react with Sm25 but cross-reacts with other schistosome proteins, including candidate vaccine antigens paramyosin (Sm97) and glutathione-S-transferases (Sm26, Sm28, Sj26). Recombinant protein r4 · 47, constructed to express multiple copies of codon sequence 117–163 (con- taining d), reacts with anti-d and is uniquely recognized by protective Fischer twice-infected (F-2x) rat antiserum. Immunization with r4 · 47 induces antibodies with cross reactivities similar to anti-d, but which also recognize Sm25. Despite these cross-reactivities with protective antigens, rodents vaccinated with r4 · 47 were not protected against cercarial infection. On the basis of these data, two hypoth- eses are proposed: (1) antigenic epitopes other than d are present within the r4 · 47 sequence which induce antibodies reactive with Sm25 and/or (2) peptide-d assumes alternative antigenic conformations, dependent upon the context of neighbouring sequences, some of which mimic epitopes of proteins encoded by other schistosome genes. These mimo- topes are not targets of protective antibodies. Keywords Schistosoma mansoni, recombinant antigen, cross-reacting antigens, Sm25, expression vector pET15b, vaccine INTRODUCTION S. mansoni gene GP22 was identified in our laboratory (El-Sherbeini et al. 1991) as an intronless genomic DNA sequence which, with the exception of a single amino acid difference (one nucleotide position 1 ), was identical to the cDNA sequence of the previously described tegumental glycoprotein, Sm25 (Omer-Ali et al. 1991). Each laboratory independently isolated cDNA clones (from different lgt11 expression libraries), containing the same 47-amino acid insert, subsequently shown to be amino acid positions 117– 163 in the translated sequence. The two laboratories used different screening strategies to identify candidate vaccine antigens of the parasite. Our procedure involved screening the cDNA expression library by the ‘contrasting antiserum’ protocol. The 47-amino acid schistosome sequence within the b-galactosidase fusion protein reacted with antibodies uniquely present in protective F-2x antiserum (from twice- infected Fischer rats) but not with antibodies in nonprotective immune serum W-2x (from twice-infected Wistar–Furth rats) (El-Sherbeini et al. 1990). Parasite Immunology, 2000: 22: 381–395 q 2000 Blackwell Science Ltd 381 Correspondence: Paul M.Knopf Received: 13 July 1999 Accepted for publication: 23 March 2000 1 Nucleotide in position 143 was G in our sequence and C in the Mill Hill sequence, leading to an ATG (met) versus ATC (ile) in amino acid position 47. We have since learned of the sequence of another independently isolated cDNA of this gene, which confirms the G in position 143 (Mangold B.L. & Ricardoni M., NAMRU-3, Cairo, Egypt). However, different parasite isolates may be allelic in gene GP22.
Transcript of Schistosomamansoni gene GP22 encodes the tegumental antigen Sm25: (1) antibodies to a predicted...

Schistosoma mansoni gene GP22 encodes the tegumental antigen
Sm25: (1) antibodies to a predicted B-cell epitope of Sm25
cross-react with other candidate vaccine worm antigens;
(2) characterization of a recombinant product containing
tandem-repeats of this peptide as a vaccine
MARY M. PETZKE1, PARMJEET K. SURI1, RICHARD BUNGIRO1, MIRIAM GOLDBERG1, SARAH F. TAYLOR1,
SUMANT RANJI1, HARRY TAYLOR2, JOSEPH W.MCCRAY JR2 & PAUL M. KNOPF1
1Department of Molecular Microbiology and Immunology, Brown University, Providence, RI 02912, 2Department of Biology,
Morehouse College, Atlanta, GA, USA
SUMMARY
Monospeci®c antibodies against two putative epitopes of
schistosome protein encoded by gene GP22 (182 codons, no
introns) were used to probe worm extracts fractionated by
lentil-lectin af®nity chromatography or by electrophoresis.
Anti-peptide-a (codons 70±84) exclusively identi®es the N-
glycanase-sensitive, 25 kDa tegumental glycoprotein Sm25
in the lectin-bound fraction of detergent-solubilized adult
worm extract S3. In contrast, antipeptide-d (codons 151±
162) does not react with Sm25 but cross-reacts with other
schistosome proteins, including candidate vaccine antigens
paramyosin (Sm97) and glutathione-S-transferases (Sm26,
Sm28, Sj26). Recombinant protein r4 ´ 47, constructed to
express multiple copies of codon sequence 117±163 (con-
taining d), reacts with anti-d and is uniquely recognized by
protective Fischer twice-infected (F-2x) rat antiserum.
Immunization with r4 ´ 47 induces antibodies with cross
reactivities similar to anti-d, but which also recognize Sm25.
Despite these cross-reactivities with protective antigens,
rodents vaccinated with r4 ´ 47 were not protected against
cercarial infection. On the basis of these data, two hypoth-
eses are proposed: (1) antigenic epitopes other than d are
present within the r4 ´ 47 sequence which induce antibodies
reactive with Sm25 and/or (2) peptide-d assumes alternative
antigenic conformations, dependent upon the context of
neighbouring sequences, some of which mimic epitopes of
proteins encoded by other schistosome genes. These mimo-
topes are not targets of protective antibodies.
Keywords Schistosoma mansoni, recombinant antigen,
cross-reacting antigens, Sm25, expression vector pET15b,
vaccine
INTRODUCTION
S. mansoni gene GP22 was identi®ed in our laboratory
(El-Sherbeini et al. 1991) as an intronless genomic DNA
sequence which, with the exception of a single amino acid
difference (one nucleotide position1), was identical to the
cDNA sequence of the previously described tegumental
glycoprotein, Sm25 (Omer-Ali et al. 1991). Each laboratory
independently isolated cDNA clones (from different lgt11
expression libraries), containing the same 47-amino acid
insert, subsequently shown to be amino acid positions 117±
163 in the translated sequence. The two laboratories used
different screening strategies to identify candidate vaccine
antigens of the parasite. Our procedure involved screening the
cDNA expression library by the `contrasting antiserum'
protocol. The 47-amino acid schistosome sequence within
the b-galactosidase fusion protein reacted with antibodies
uniquely present in protective F-2x antiserum (from twice-
infected Fischer rats) but not with antibodies in nonprotective
immune serum W-2x (from twice-infected Wistar±Furth
rats) (El-Sherbeini et al. 1990).
Parasite Immunology, 2000: 22: 381±395
q 2000 Blackwell Science Ltd 381
Correspondence: Paul M.Knopf
Received: 13 July 1999
Accepted for publication: 23 March 2000
1 Nucleotide in position 143 was G in our sequence and C in the Mill
Hill sequence, leading to an ATG (met) versus ATC (ile) in amino acid
position 47. We have since learned of the sequence of another
independently isolated cDNA of this gene, which con®rms the G in
position 143 (Mangold B.L. & Ricardoni M., NAMRU-3, Cairo, Egypt).
However, different parasite isolates may be allelic in gene GP22.

Because of its unique immunoreactivity with antibodies
in protective serum F-2x, we decided to test the possibility
that epitopes in this sequence were targets of the protective
humoral immune response in rats. We also tested the vacci-
nation strategy in mice since Smithers et al. (1989), using
tegumental fractions of the worms, showed a correlation of
humoral anti-Sm25 activity with protection in this model.
Moreover, to improve on the immunogenicity of this
47-amino acid segment, we engineered the construction of
a tandem display of this sequence. As a screening reagent to
identify the expressed product, we developed a rabbit
polyclonal, monospeci®c antipeptide antibody against the
predicted B-cell epitope delta (d, amino acids 151±162)
present in the 47-amino acid sequence.
In these early vector expression experiments, utilizing
the pGEX vector to express gene GP22 sequence as fusion
protein with Sj26 (glutathione-S-transferase, GST), an
unexpected result emerged which stimulated our interest in
pursuing this direction of vaccine development. Anti-d anti-
bodies reacted very strongly by enzyme-linked immunosor-
bant assay (ELISA) with the puri®ed GST vector-only
control product. Not only was this cross-reactivity with
GST revealed, but immunoblotting assays revealed a strik-
ing reactivity of the anti-d with other S. mansoni macro-
molecules, the strongest of which was con®rmed to be Sm97
(paramyosin). Since both GST and paramyosin were con-
sidered prominent candidate vaccine antigens (Riveau &
Capron 1997), we proposed the following hypothesis: Gene
GP22-encoded glycoprotein products contain a short amino
acid segment capable of inducing humoral immune responses
which are cross-reactive with other antigens of the parasite.
This cross-reaction thereby expands the range of available
targets for the parasite-induced immune response, which
protects the host from becoming heavily reinfected and
vulnerable to the severe pathology of this disease.
The cross-reactivities of anti-d antibodies and of anti-
bodies to the tandem construct of the 47 amino acid sequence,
r4 ´ 47, are described, together with the results of experi-
ments using r4 ´ 47 as a vaccine in rats and mice. In addi-
tion, we report on the use of anti-d antibodies and antibodies
to alpha (a, amino acids 70±84), another predicted peptide
epitope for B cells, to characterize the expressed products of
gene GP22 in adult worms.
MATERIALS AND METHODS
Parasites
Biomphalaria glabrata snails infected with the Puerto Rican
strain of S. mansoni were provided under an NIAID supply
contract (AI052590) by Biomedical Research Institute,
Rockville, MD, USA. Juvenile or adult worms were isolated
from outbred mice exposed via the tail, 4 or 7 weeks earlier,
to 600 or 200 cercariae, respectively (Knopf et al. 1977).
Animals
Male Fischer rats (CD-F, 80±100 g) were obtained from
Charles River Breeding Laboratories (Wilmington, MA,
USA); male Wistar±Furth rats (80±100 g) were from M.
A. Laboratories (Bethesda, MD, USA). Female Balb/c mice
(23±25 g) were from Charles River. Female New Zealand
White rabbits were from Millbrook Farm (Amherst, MA,
USA).
Cloning and expression of recombinant proteins
r4 ´ 47 and r140
Restriction enzymes (New England Biolabs, Beverly, MA,
USA) were used according to manufacturer's instructions.
pET(4 ´ 47) was constructed by a series of steps utilizing
two vector systems. The 141 bp insert (codons 117±163 of
S. mansoni gene GP22, see Figure 4) was excised from the
pUC-19 cDNA clone (El-Sherbeini et al. 1991) by EcoRI
digestion and puri®ed (Maniatis et al. 1982). The insert, in
10-fold molar excess, was ligated into the EcoRI site of
pMAL-c2 vector (New England Biolabs) and ampicillin-
resistant transformants selected. Orientation of inserts in
transformants containing double inserts was con®rmed by
a mini-expression analysis of fusion proteins by sodium
dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-
PAGE) (Protein Fusion and Puri®cation System, New
England Biolabs). A clone, termed pMAL(2 ´ 47), contain-
ing two 141 bp inserts ligated in tandem (con®rmed by
sequencing), was identi®ed. Note that the EcoRI site joining
the inserts is a shared region, with a resulting sequence of
276 bp instead of 282 bp. Translation of this tandem insert
yields 92 amino acids instead of 94 a.a. (the C-terminal
glu-phe of the 50-insert is also the N-terminal glu-phe of the
30-insert). However, as a convenience of nomenclature, we
refer to this tandem amino acid sequence as 2 ´ 47.
pMAL(4 ´ 47), containing two copies of the tandem
insert, was then constructed. It consists of two 276 bp
sequences separated by ®ve codons of linker sequence
(veri®ed by DNA sequencing). This double tandem insert
was excised by cleavage with XmnI and BamHI and cloned,
utilizing a BglII linker sequence to maintain the correct
reading frame, into the pET-15b vector (Novagen, Madison,
WI, USA) to generate pET(4 ´ 47). The translated product
of pET(4 ´ 47), termed r4 ´ 47, is composed of two worm-
derived tandem sequences separated by a 5-amino acid linker,
joined at the amino terminal to a 23-amino acid `His.Tag'
vector sequence (containing six histidine residues) and a
4-amino acid linker, and followed by a 5-residue vector
M.M.Petzke et al. Parasite Immunology
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395382

sequence terminating with cysteine (see Figure 4). Recom-
binant protein r4 ´ 47 was expressed and puri®ed from the
soluble fraction of sonicated BL21(DE3)pLysS cells
(Novagen) by nickel-af®nity chromatography on `His.Bind'
resin, according to manufacturer's protocol. Eluted protein
was dialysed overnight at 48C against phosphate-buffered
saline (PBS)/10% glycerol/0´01% Tween-20 using a 3´5 kDa
molecular weight cutoff dialysis tubing. The solution was
concentrated using polyethylene glycol (15 000±20 000 kDa;
Sigma Chemical Co., St Louis, MO, USA) and protein
concentration established by OD at 280 nm. In one study,
r4 ´ 47 was further puri®ed by size-fractionation chromato-
graphy over Sephacryl-200 (Sigma) and called sp-r4 ´ 47.
Vector pGEX-3X (Pharmacia Biotech, Uppsala, Sweden)
was expressed in DH5a cells. Construction of pET(140) and
expression/puri®cation of r140 (GP22 codons 43±182) is
described elsewhere (Suri et al. 1997).
Synthetic peptides
The sequences are as follows (see also Figure 4): Peptide
alpha (a): (70)NSEENSNSIITDEDY(84)GGC; peptide delta
(d): (151)TPNSANKYMKNE(162)GGC.
Peptides corresponding to codons 70±84 (a sequence)
and 151±162 (d sequence), plus a linker sequence of three
residues at the carboxy termini (GGC, single letter amino
acid code) were synthesized by Multiple Peptide Systems
(San Diego, CA, USA). For immunizations, the peptides
were each coupled to an immunogenic protein carrier via
b-maleimidopropionic acid-N-hydroxysuccinimide ester
(Sigma) linkage (McCray & Werner 1989). Further details
are given below for the peptide-carrier constructs used in the
immunization of rabbits and the puri®cation of antipeptide
antibodies by af®nity chromatography.
Other antigens and worm fractions
Recombinant S. japonicum 26 kDa GST (rSj26-GST) was
puri®ed by glutathione agarose chromatography (Torian
et al. 1990) from DH5a Escherichia coli transformed with
pGEX-3X vector. Recombinant S. mansoni 26-kDa GST
(rSm26-GST) and rSm28-GST were kindly provided by
Dr B. Doughty of Texas A & M University. Recombinant
and worm-puri®ed Sm97-paramyosin were gifts from Dr
C. Shoemaker of Harvard University and S. Heiny in Dr A.
Sher's laboratory at the NIH, respectively.
The aqueous-soluble worm fraction (S2) and the deter-
gent-solubilized worm fraction (S3) extracted from the
residual aqueous-insoluble pellet were prepared as described
(Suri et al. 1997). Protein concentrations were determined
against a bovine serum albumin standard using Bradford's
Reagent (BioRad Laboratories, Hercules, CA, USA).
Aliquots containing 2 mM phenylmethylsulphonyl ¯uoride
(PMSF) (ICN Pharmaceuticals, Inc., Costa Mesa, CA, USA)
were stored at ÿ808C.
Biosynthetically radiolabelled S2 and S3 fractions were
prepared from 4-week worms incubated in culture medium
containing [35S]-methionine (Mark et al. 1991).
Sera and antibodies
Fischer twice-infected rat serum (F-2x) was obtained
3 weeks after the second infection with S. mansoni cercariae
(Barker et al. 1985). Wistar-Furth twice-infected rat serum
(W-2x) was raised following a similar protocol.
Antisera to recombinant proteins (anti-r4 ´ 47 and anti-
r140) were prepared by immunizing Fischer rats subcuta-
neously (s.c.) with nickle-af®nity puri®ed r4 ´ 47 or r140,
emulsi®ed 1 : 1 in alum (aluminium hydroxide adsorptive
gel, Rehsorptar, Intergen Company, Purchase, NY, USA).
Rabbits were injected intradermally at intervals of 4±6
weeks with 1 mg antigen mixed 1 : 1 with alum in 1 ml.
Blood was collected 2 weeks after each injection by retro-
orbital bleeding (rats) or puncture of the ear vein (rabbits).
Serum samples with the highest ELISA antibody titres were
used.
Monospeci®c antipeptide antibodies were puri®ed from
rabbit antisera by peptide-af®nity chromatography. Rabbits
were injected s.c. at multiple sites on the back with 0´2 mg of
peptide-carrier protein conjugate mixed 1 : 1 with Freund's
complete adjuvant, in a total volume of 1 ml. A booster
injection was given s.c. 3 weeks later in Freund's incom-
plete adjuvant and repeated at intervals. For immunization
with peptide-d-conjugate, the rabbits were initially immun-
ized with an ovalbumin (OVA) conjugate, four times. We
then switched to a keyhole limpet haemocyanin (KLH)
conjugate of peptide-d when the last two boosters with
OVA±conjugate failed to increase the antipeptide titre. No
differences in the speci®cities to the antigens tested were
detectable using peptide-af®nity-puri®ed antibodies from
early or later sera from these rabbits (see Discussion).
For immunization with peptide-a-conjugate, the rabbits
were always immunized with a KLH conjugate. Serum
was prepared from blood collected 7±21 days after boost-
ing. To purify antipeptide antibodies, the peptides were
coupled to Af®-Gel 102 (BioRad) by a previously published
method (McCray & Werner 1989). Sera (5 ml) were diluted
1 : 1 with PBS and loaded onto small (3±4 ml) columns of
the peptide-Af®-Gel 102 conjugate. The columns were
washed with PBS and the bound antipeptide antibodies
were eluted with 0´1 M glycine-HCl/0´15 M NaCl buffer,
pH 2´8. Eluted fractions were collected in tubes containing
2 M Tris to raise the pH to approximately 7´0 and were tested
in an ELISA for antipeptide antibody activity. Active
Volume 22, Number 8, August 2000 Properties of antibodies to gene GP22-encoded sequences
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395 383

fractions were dialysed extensively against PBS. The anti-
peptide antibodies were speci®c for the peptide used for
immunization (Petzke 1996).
Immunoassays
ELISA
This assay was utilized for preliminary determination of
the antibody-binding capacity of proteins and synthetic
peptides. Protein antigens were immobilized and antibody
titres determined as described (Suri et al. 1997). For assay-
ing peptides, each well was coated with 1 mg of synthetic
peptide, and a 1 : 5000 dilution of secondary antibody
conjugate was used (Petzke 1996).
Western immunoblots
Proteins were separated by SDS-PAGE using a Bio-Rad
Protean II vertical minigel apparatus. The gel was electro-
blotted onto nitrocellulose membrane (Hybond-ECL nitro-
cellulose, Amersham, Arlington Heights, IL, USA) using a
semidry transfer cell (BioRad) for 25 min at 10 mV. The
membrane was dried, blocked with nonfat dry milk and
incubated with primary antibody. Peroxidase-conjugated
goat anti-rat or anti-rabbit immunoglobulin heavy and
light chains was used to detect primary antibody. The
immunoblot was developed using enhanced chemilumines-
cence (ECL reagents: Amersham). Further details are pro-
vided elsewhere (Suri et al. 1997).
Competition Western immunoblots
An identical amount of protein was loaded into each well
and separated by SDS-PAGE. Following transfer to nitro-
cellulose, membranes were ®xed and proteins visualized by
Ponceau S (Sigma) staining. The membrane was cut into
strips, transferred to the wells of a mini-incubation tray
(BioRad), destained with deionized water, and blocked
overnight. Primary antibody was incubated with varying
concentrations of competitor for 1´5 h at room temperature,
transferred to a well of the mini-incubation tray and main-
tained at 378C for 1´5 h. Addition of secondary antibody
conjugate and developing are described above.
Lentil lectin sepharose chromatography of S2 and S3
Af®nity chromatography utilizing lentil lectin coupled to
beads of Sepharose 4B (Sigma) was used to adsorb and
isolate glycoproteins containing glucosyl and/or mannosyl
residues. Approximately 20 ml of lentil-lectin Sepharose 4B
suspension was packed into a glass column (2 cm ´ 22 cm,
Pharmacia) and washed with 10 bed volumes of running
buffer [9% NaCl, 10 mM CaCl2, 10 mM MnCl2, 0´1% Renex
(a nonionic detergent, Ruger Chemical Co., Irvington, NJ,
USA)]. A 2-ml volume of S2 or S3 (approximately 1´3±
1´5 mg protein/ml) was applied to the column and fractions
collected by washing with running buffer (60±80 ml) to
remove unbound sample components. The lectin-bound
components were eluted with the addition of 2´5% methyl-
a-D-mannoside (Sigma) to the running buffer. Column
¯owthrough and eluate fractions were collected assessed
for protein content by OD at 280 nm.
N-glycanase treatment of antigens
Frozen aliquots of S2 and S3 were thawed and subjected to
sonication for 10 s to resuspend particulate matter. Approxi-
mately 200 mg of each fraction containing 0´5% SDS
and 50 mM b-mercaptoethanol was denatured by boiling
5 min before addition of enzyme (3 U recombinant N-gly-
canase, Genzyme, Cambridge, MA, USA), 5 mM PMSF, and
1´1% 3-(3-cholamino-propyl)dimethylammonio-1-propane-
sulphonate (CHAPS). Reactions were for 18 h at 378C,
terminated by the addition of 5X PAGE sample buffer
followed by boiling for 5 min. Recombinant antigens r140
and r4 ´ 47 were treated in the same manner.
Immunization protocols
Three different antigens emulsi®ed 1 : 1 (v:v) in alum were
used in three separate vaccine trials with CD-F Fischer rats
(50±70 g) or female Balb/c mice (23±25 g) as indicated.
Control animals in each vaccine trial were either uninjected
or were injected with PBS emulsi®ed 1 : 1 (v:v) in alum.
Vaccine trial 1
On days 0, 24 and 51, rats received injections of 100 mg or
200 mg of nickel-af®nity-puri®ed r4 ´ 47 in a total volume of
0´2 or 0´4 ml, respectively, and mice received 75 mg in a
volume of 0´15 ml. Rats were challenged percutaneously
on day 75 by exposure of shaved abdominal skin to 520
S. mansoni cercariae (Knopf et al. 1977). Mice were
infected on the same day via tail immersion in water
containing 150 cercariae. Fluids contained < 5% residual
cercariae after 30 min exposures. Worms were collected by
portal perfusion (described below) of rats on day 103
(4 weeks postinfection) and of mice on day 124 (7 weeks
postinfection).
Vaccine trial 2
Rats were immunized s.c. on day 0 with 150 mg sp-r4 ´ 47
(Sephacryl-puri®ed r4 ´ 47), emulsi®ed in total volume
0´3 ml and were boosted on day 21 (75 mg antigen,
0´15 ml) and days 45 and 66 (150 mg antigen, 0´3 ml).
Rats were challenged percutaneously on day 76 with 500
cercariae. On day 79, one group of antigen-injected rats and
one group of PBS-injected rats (approximately 265 g)
M.M.Petzke et al. Parasite Immunology
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395384
received F-2x antiserum (0´75 ml) passively transferred by
s.c. injection. The dose of F-2x (0´25 ml/90 g rat) was
predetermined to be protective by titration but less than
the maximal amount of serum was used. Worms were
recovered by portal perfusion 4 weeks postinfection.
Vaccine trial 3
The lentil lectin-bound (LLB) material from S3 fraction of
adult worms was eluted, concentrated and used to immunize
four rats after emulsi®cation in alum. Two immuni-zations,
2 weeks apart, of 145 mg protein each of freshly prepared
lectin-bound material, was injected intramuscularly (half
into each hind leg). Two rats which yielded anti-S3 response
equivalent in titre to F-2x (positive control serum) 3 weeks
after the second injection were exsanguinated and serum
was prepared (stored at ÿ808C). The remaining two rats
were boosted again (120 mg each) and serum prepared
2 weeks later. The anti-S3 titres are given in the Results.
Rats were exposed percutaneously to 220 cercariae and on
day 3 postinfection, they received 1 ml of anti-LLB or NRS,
or 1 ml F-2x s.c. (under the scruff of the neck). On day 28
postinfection, the worm burdens in these different groups of
rats were determined after portal perfusion.
Protection assays
Worms were collected on 30 mm (rats) or 45 mm (mice)
nylon screens by perfusion of the portal-mesenteric veins
with saline (Knopf et al. 1977) and counted independently
by two researchers using a dissecting microscope and
averaged, variation < 10%. Worm pairs obtained from
mice were allowed to separate overnight, and were enum-
erated separately. The liver egg burdens were measured in
mice (Cioli et al. (1977).
RESULTS
GP22-encoded product found in S3 fraction is Sm25
glycoprotein
The amino acid sequence of gene GP22 contained two
putative N-linked glycosylation sites which could mediate
binding of the gene product(s) to lentil lectin. To character-
ize the protein product(s) of gene GP22 in adult worms, the
aqueous-soluble S2 and detergent-soluble S3 fractions were
mixed with trace amounts of the corresponding fraction
from radiolabelled worms and subjected to lentil lectin-
af®nity chromatography. The resultant chromatographic
fractions were probed for immunoreactivity by ELISA
using antiserum from twice-infected Fischer rats, F-2x (Man-
gold & Knopf 1981, Barker et al. 1985) or antiserum raised
by vaccination with GP22-encoded recombinant protein, r140
(codons 43±182) (Suri et al. 1997). In addition, radioactivity
in each column fraction was measured to monitor distribution
of proteins and glycoproteins.
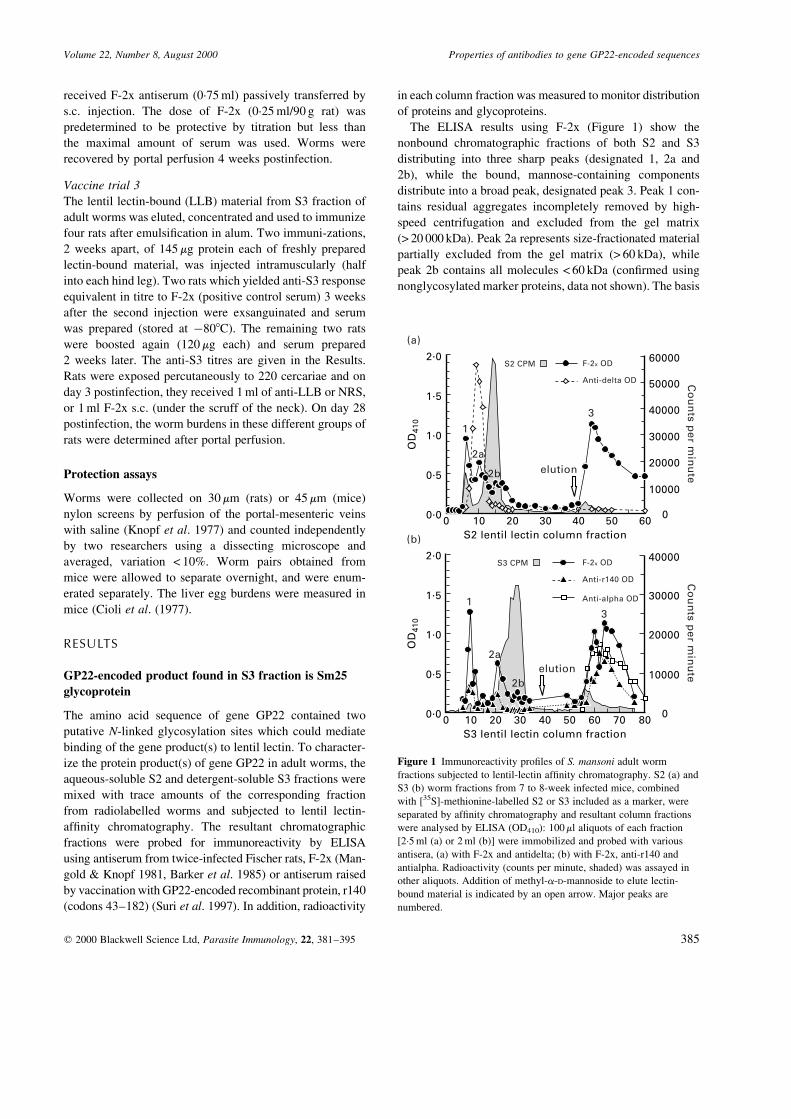
The ELISA results using F-2x (Figure 1) show the
nonbound chromatographic fractions of both S2 and S3
distributing into three sharp peaks (designated 1, 2a and
2b), while the bound, mannose-containing components
distribute into a broad peak, designated peak 3. Peak 1 con-
tains residual aggregates incompletely removed by high-
speed centrifugation and excluded from the gel matrix
(> 20 000 kDa). Peak 2a represents size-fractionated material
partially excluded from the gel matrix (> 60 kDa), while
peak 2b contains all molecules < 60 kDa (con®rmed using
nonglycosylated marker proteins, data not shown). The basis
Volume 22, Number 8, August 2000 Properties of antibodies to gene GP22-encoded sequences
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395 385
5020
2.0
1.5
60000
50000
40000
30000
20000
10000
0
1.0
0.5
0.0
OD
410
S2 lentil lectin column fraction604030100
elution
elution
13
F-2x OD
Anti-delta OD
2a
1
2a
3
2b
2b
S2 CPM
(a)
Co
un
ts per m
inu
te
50 60 7020
2.0
1.5
40000
30000
20000
10000
0
1.0
0.5
0.0
OD
410
S3 lentil lectin column fraction804030100
F-2x OD
Anti-r140 OD
Anti-alpha OD
S3 CPM
(b)
Co
un
ts per m
inu
te
Figure 1 Immunoreactivity pro®les of S. mansoni adult worm
fractions subjected to lentil-lectin af®nity chromatography. S2 (a) and
S3 (b) worm fractions from 7 to 8-week infected mice, combined
with [35S]-methionine-labelled S2 or S3 included as a marker, were
separated by af®nity chromatography and resultant column fractions
were analysed by ELISA (OD410): 100 ml aliquots of each fraction
[2´5 ml (a) or 2 ml (b)] were immobilized and probed with various
antisera, (a) with F-2x and antidelta; (b) with F-2x, anti-r140 and
antialpha. Radioactivity (counts per minute, shaded) was assayed in
other aliquots. Addition of methyl-a-D-mannoside to elute lectin-
bound material is indicated by an open arrow. Major peaks are
numbered.
for the irregular pro®le of peak 3 was not established, but
may represent differences in binding af®nity for lentil lectin
(e.g. variability in mannose residues per molecule) and/or
size-fractionation. The 35S-methionine-labelled proteins in
both fractions distributed upon chromatography (Figure 1)
into a major peak (peak 2b) and two minor peaks (peaks 1
and 3). Peak 2a contained only minimal amounts of labelled
product. For these radiolabelled products,< 5% of S2 and
approximately 10% of S3 proteins bind to the lectin; in
contrast, the relative amounts of antigen detected by F-2x
hyperinfected rat serum in the lectin-bound fractions of
S2 and S3 (Figure 1) are strikingly higher (approximately
66% and 80%, respectively). Earlier studies established
that total adult worm proteins were distributed similar to
the 35S-methionine-labelled products. We conclude that the
antigenicity of both S2 and S3 is disproportionately higher
in the lectin-bound (mannosylated) components than in the
lectin-nonbound components, when probed by antibodies in
protective F-2x.
To assess the presence of gene GP22-encoded products,
column fractions were also probed with rabbit anti-r140, the
recombinant protein containing all predicted B-cell epi-
topes. None of the chromatographic fractions of S2 showed
signi®cant reactivity with anti-r140 (data not shown), but
lectin-bound peak 3 of S3 does show strong reactivity (Figure
1b). These results establish the presence of a signi®cant
amount of GP22-encoded glycoproteins in the membrane-
associated S3 fraction.
Monospeci®c antibodies to GP22-encoded peptides
a and d identify different worm components
Computer predictions identi®ed four distinct segments of
the GP22 amino acid sequence as putative B-cell epitopes
(El-Sherbeini et al. 1991). Two of these peptide sequences,
corresponding to codons 70±84 (a sequence) and 151±162
(d sequence), were synthesized, and antisera was prepared to
protein conjugates of each in rabbits. The speci®c antipep-
tide antibodies were puri®ed by af®nity chromatography
and used to probe the S3 lectin-bound fractions. Anti-a,
similar to anti-r140, detected antigen only in the S3 bound
fraction (Figure 1b). No anti-a reactivity was observed in
the S2 bound fraction (data not shown). Surprisingly,
reactivity with anti-d was not detected with the S3 lectin-
bound fraction. Instead, its reactivity was con®ned to lectin-
nonbound peak 2, primarily peak 2a (> 60 kDa), of the S2
fraction (Figure 1a). Thus, antibodies directed against two
different peptide sequences of GP22-encoded protein
identify antigen fractions with very different characteristics
both in terms of localization in worm extracts and in the
possession of lectin-binding sugars. Anti-a, similar to anti-
r140, detects antigens in the bound glycosylated components
of S3; anti-d primarily detects antigen of > 60 kDa in the
nonbound components of S2.
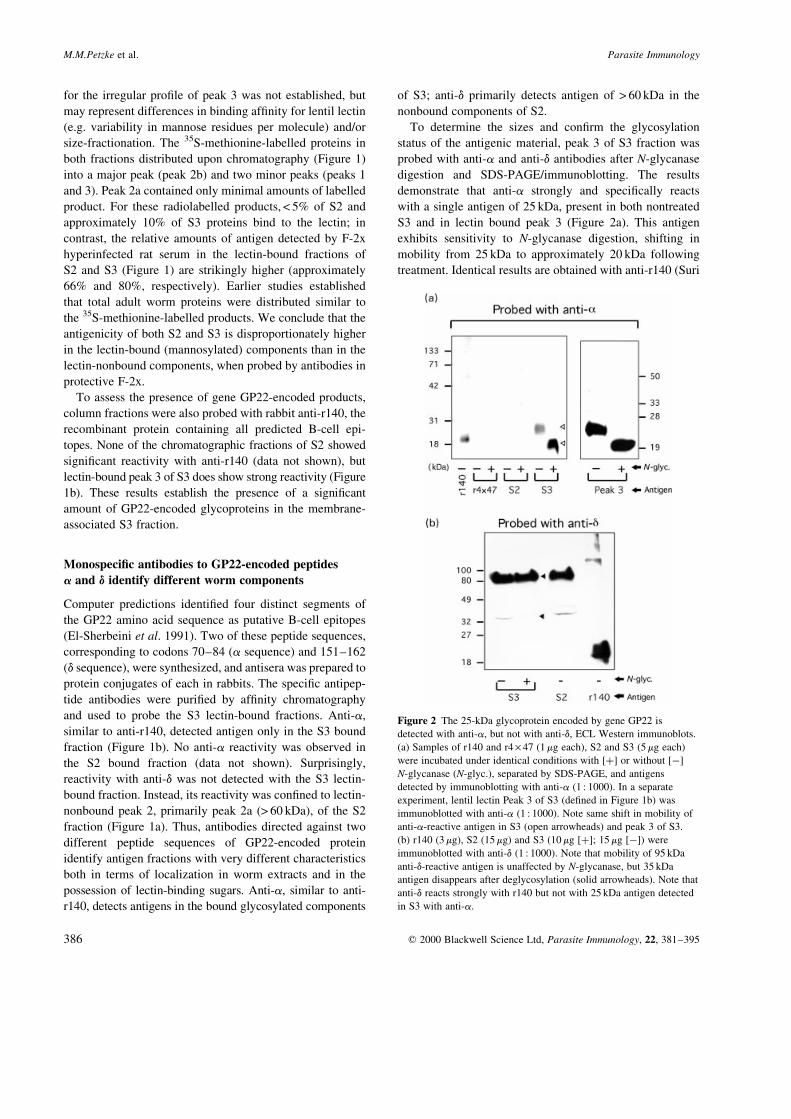
To determine the sizes and con®rm the glycosylation
status of the antigenic material, peak 3 of S3 fraction was
probed with anti-a and anti-d antibodies after N-glycanase
digestion and SDS-PAGE/immunoblotting. The results
demonstrate that anti-a strongly and speci®cally reacts
with a single antigen of 25 kDa, present in both nontreated
S3 and in lectin bound peak 3 (Figure 2a). This antigen
exhibits sensitivity to N-glycanase digestion, shifting in
mobility from 25 kDa to approximately 20 kDa following
treatment. Identical results are obtained with anti-r140 (Suri
M.M.Petzke et al. Parasite Immunology
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395386
Figure 2 The 25-kDa glycoprotein encoded by gene GP22 is
detected with anti-a, but not with anti-d, ECL Western immunoblots.
(a) Samples of r140 and r4 ´ 47 (1 mg each), S2 and S3 (5 mg each)
were incubated under identical conditions with [�] or without [ÿ]
N-glycanase (N-glyc.), separated by SDS-PAGE, and antigens
detected by immunoblotting with anti-a (1 : 1000). In a separate
experiment, lentil lectin Peak 3 of S3 (de®ned in Figure 1b) was
immunoblotted with anti-a (1 : 1000). Note same shift in mobility of
anti-a-reactive antigen in S3 (open arrowheads) and peak 3 of S3.
(b) r140 (3 mg), S2 (15 mg) and S3 (10 mg [�]; 15 mg [ÿ]) were
immunoblotted with anti-d (1 : 1000). Note that mobility of 95 kDa
anti-d-reactive antigen is unaffected by N-glycanase, but 35 kDa
antigen disappears after deglycosylation (solid arrowheads). Note that
anti-d reacts strongly with r140 but not with 25 kDa antigen detected
in S3 with anti-a.
et al. 1997). These properties identify this S3 component as
Sm25, as described by Knight et al. (1989). In contrast to
anti-a, anti-d does not react with this 25 kDa protein nor
with its deglycosylated product (Figure 2b). Instead, anti-d
cross-reacts with several schistosomal proteins, present in
both S2 and S3, of approximate sizes of 35, 50 (S2 only) and
95 kDa, which are insensitive to N-glycanase digestion.
Reactivity with the 95 kDa antigen is the strongest. While
not reacting with Sm25 or its deglycosylated product, anti-d,
similar to anti-a, does react with recombinant protein r140
(Figure 2).
Worm fractions S2 and S3 each consist of a complex
mixture of proteins, when visualized by Coomassie-blue
staining of the gel (Petzke 1996), whereas lentil lectin peak
3 consists of a subset of these proteins. In contrast, only a
fraction of these proteins are antigens reacting with anti-a
and anti-d, con®rming the speci®city of these antibodies.
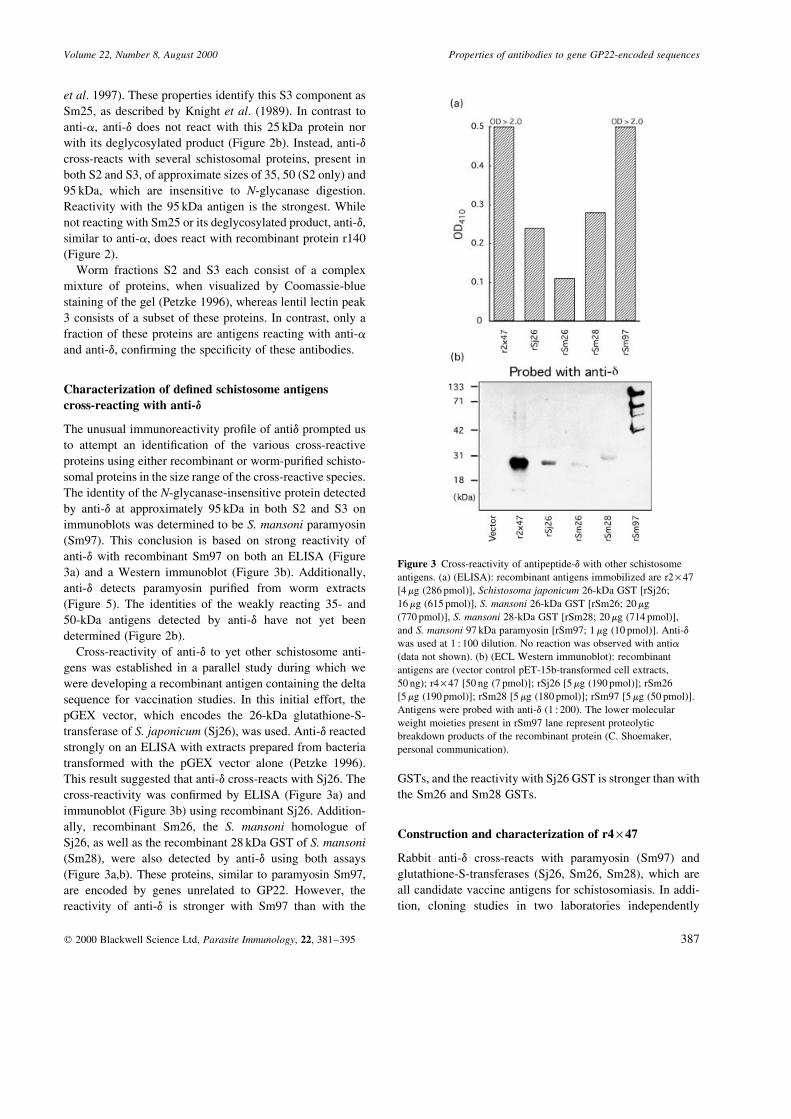
Characterization of de®ned schistosome antigens
cross-reacting with anti-d
The unusual immunoreactivity pro®le of antid prompted us
to attempt an identi®cation of the various cross-reactive
proteins using either recombinant or worm-puri®ed schisto-
somal proteins in the size range of the cross-reactive species.
The identity of the N-glycanase-insensitive protein detected
by anti-d at approximately 95 kDa in both S2 and S3 on
immunoblots was determined to be S. mansoni paramyosin
(Sm97). This conclusion is based on strong reactivity of
anti-d with recombinant Sm97 on both an ELISA (Figure
3a) and a Western immunoblot (Figure 3b). Additionally,
anti-d detects paramyosin puri®ed from worm extracts
(Figure 5). The identities of the weakly reacting 35- and
50-kDa antigens detected by anti-d have not yet been
determined (Figure 2b).
Cross-reactivity of anti-d to yet other schistosome anti-
gens was established in a parallel study during which we
were developing a recombinant antigen containing the delta
sequence for vaccination studies. In this initial effort, the
pGEX vector, which encodes the 26-kDa glutathione-S-
transferase of S. japonicum (Sj26), was used. Anti-d reacted
strongly on an ELISA with extracts prepared from bacteria
transformed with the pGEX vector alone (Petzke 1996).
This result suggested that anti-d cross-reacts with Sj26. The
cross-reactivity was con®rmed by ELISA (Figure 3a) and
immunoblot (Figure 3b) using recombinant Sj26. Addition-
ally, recombinant Sm26, the S. mansoni homologue of
Sj26, as well as the recombinant 28 kDa GST of S. mansoni
(Sm28), were also detected by anti-d using both assays
(Figure 3a,b). These proteins, similar to paramyosin Sm97,
are encoded by genes unrelated to GP22. However, the
reactivity of anti-d is stronger with Sm97 than with the
GSTs, and the reactivity with Sj26 GST is stronger than with
the Sm26 and Sm28 GSTs.
Construction and characterization of r4 ´ 47
Rabbit anti-d cross-reacts with paramyosin (Sm97) and
glutathione-S-transferases (Sj26, Sm26, Sm28), which are
all candidate vaccine antigens for schistosomiasis. In addi-
tion, cloning studies in two laboratories independently
Volume 22, Number 8, August 2000 Properties of antibodies to gene GP22-encoded sequences
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395 387
Figure 3 Cross-reactivity of antipeptide-d with other schistosome
antigens. (a) (ELISA): recombinant antigens immobilized are r2 ´ 47
[4 mg (286 pmol)], Schistosoma japonicum 26-kDa GST [rSj26;
16 mg (615 pmol)], S. mansoni 26-kDa GST [rSm26; 20 mg
(770 pmol)], S. mansoni 28-kDa GST [rSm28; 20 mg (714 pmol)],
and S. mansoni 97 kDa paramyosin [rSm97; 1 mg (10 pmol)]. Anti-d
was used at 1 : 100 dilution. No reaction was observed with antia
(data not shown). (b) (ECL Western immunoblot): recombinant
antigens are (vector control pET-15b-transformed cell extracts,
50 ng); r4 ´ 47 [50 ng (7 pmol)]; rSj26 [5 mg (190 pmol)]; rSm26
[5 mg (190 pmol)]; rSm28 [5 mg (180 pmol)]; rSm97 [5 mg (50 pmol)].
Antigens were probed with anti-d (1 : 200). The lower molecular
weight moieties present in rSm97 lane represent proteolytic
breakdown products of the recombinant protein (C. Shoemaker,
personal communication).

identi®ed the same 47-amino acid peptide, containing the
delta sequence, in their screening of cDNA expression
libraries using sera from resistant mice (Knight et al. 1989)
and rats (El-Sherbeini et al. 1991). These coincidences
were suf®ciently compelling to attempt a vaccine trial
with this region of the Sm25 protein. We used the strategy
of enhancing the immunogenicity and protective capacity of
a peptide by incorporating multiple units of the peptide onto
a polymeric core (Wolowczuk et al. 1991). Multiple copies
of the 141 bp EcoR1 restriction fragment of gene GP22
(codons 117±163) were cloned by a series of steps into
the pET-15b expression vector. The predicted recombinant
protein r4 ´ 47 consists of four contiguous copies of the 47-
amino acid sequence joined to an amino terminal 2 kDa
`His.Tag' vector sequence (Figure 4). This plasmid,
pET(4 ´ 47), was constructed to include a terminal cysteine
residue at the carboxyl end of the insert sequence, allowing
the formation of a disulphide bond between two molecules
of r4 ´ 47. Analysis of r4 ´ 47 by SDS-PAGE followed by
immunoblotting with anti-d detects a major species at
approximately 26 kDa (Figure 3b; the similar MWr to
Sm25 is coincidence) which shifts to approximately
50 kDa under nonreducing conditions (Petzke 1996).
The recombinant protein r4 ´ 47 reacts strongly with anti-
d and, as expected, is not detected by anti-a (a is codons
70±84, see Figure 4). The speci®city of this reaction is
demonstrated by a competition study shown in Figure 5, in
which soluble peptide delta completely competes the bind-
ing of anti-d to r4 ´ 47, as well as to worm-puri®ed Sm97,
the cross-reacting antigen displaying the most intense reac-
tivity with anti-d.
The recombinant protein r4 ´ 47 is recognized by F-2x,
but not by W-2x serum, in a Western immunoblot analysis
(Figure 6). This unique property of F-2x is expected, since
M.M.Petzke et al. Parasite Immunology
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395388
Figure 4 Diagramatic representation of Sm25 and r4 ´ 47. Upper: Sm25 (decoded from gene GP22) consists of 182 amino acids (open
arrowheads). The locations of peptides a (amino acids 70±84) and d (amino acids 151±162) are shown in striped bars. The segment expressed
in r140 (amino acids 43±182; recombinant protein constructed by Suri et al. (1997) and in r4 ´ 47 (amino acids 117±163; this study) are shown
(solid arrowheads). Lower: Recombinant protein r4 ´ 47 consists of (B), two tandem pairs of 47 amino acid repeats (note that the last two amino
acids of the ®rst repeat are also the ®rst two amino acids of the second repeat), each tandem pair separated by (C), a 5 amino acid linker
sequence (C�GSSIS, single-letter amino acid abbreviations), and preceded by (A), a 27 amino acid N-terminal `His.Tag' sequence
(A�MGSSHHHHHHSSGLVPRGSHMLEDLIS) encoded by the pET-15b vector. The 4 amino acid C-terminal tail (D�GSGC) encoded by the
vector contains a terminal cysteine residue, resulting in formation of a disulphide bond, under nonreducing conditions, between two r4 ´ 47
molecules, producing eight copies of the same peptide sequence per molecule of antigen.

the original 47-amino acid sequence was detected as a fusion
protein with b-galactosidase in a lgt11 phage-expression
library using the identical criterion (El-Sherbeini et al. 1990).
Anti-r4 ´ 47 detects Sm25 and cross-reacts with other
worm antigens
Anti-d antibodies cross-react with several schistosome anti-
gens not encoded by gene GP22. This cross-reactivity
phenomenon is not restricted to anti-d antibodies, but is
also observed with antibodies raised against the recom-
binant protein containing the d sequence (r4 ´ 47). The
Western immunoblot shown in Figure 7 demonstrates reac-
tivity of anti-r4 ´ 47 with these schistosome antigens. Addi-
tionally, in contrast to the inability of anti-d to detect Sm25
(Figure 2b), anti-r4 ´ 47 strongly detects Sm25 in both
N-glycanase-treated and untreated S3 extracts (Figure 8)
as well as peptide delta on ELISA (Petzke 1996). This
latter ®nding suggests that either a region of the 47-amino
acid sequence other than the d sequence contains an
epitope shared with Sm25, or that the d sequences in
r4 ´ 47 are in at least two conformations, only one of
which is present in Sm25 (the conformation not present in
`free' peptide delta). Although the binding of anti-r4 ´ 47
to r4 ´ 47 in a Western immunoblot is not blocked by
peptide-d (Petzke 1996), this result does not distinguish
between the two possibilities.
Vaccine trials with r4 ´ 47: high antibody titres but no
protection
The recombinant protein r4 ´ 47, emulsi®ed in alum, was
injected into Balb/c mice (75 mg dose) and two groups of
Fischer rats, receiving either 100 mg or 200 mg doses. Three
injections were given at two or three week intervals, and an
increase in anti-r4 ´ 47 antibody titres was observed in both
rodent models following each booster injection (Figure 9A).
The anti-r4 ´ 47 antibodies in both species were mainly of
the IgG isotype (Petzke 1996). Peak antibody titres were
reached immediately prior to infection (25 000 in mice;
9 000±11 000 in rats, with little difference between the
groups receiving 100 mg or 200 mg; Figure 9a). Cercarial
challenge, however, did not induce an anti-r4 ´ 47 response
in control mice, and titres in vaccinated animals had
dropped by the time of perfusion (to 12 000 in mice; 8000
in rats). Concurrently, no protection was observed (Table 1).
A noticeable increase in worm yields, relative to the con-
trols, was detected in the vaccinated rats, consisting of a
Volume 22, Number 8, August 2000 Properties of antibodies to gene GP22-encoded sequences
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395 389
Figure 5 Competitive Western immunoblot to de®ne anti-d
speci®city: A mixture of r4 ´ 47 [50 ng (7 pmol)/lane] and worm-
puri®ed Sm97 [2 mg (20 pmol)/lane] was separated by SDS-PAGE
and immunoblotted with anti-d (1 : 1600) which had been
preincubated with the indicated amounts of soluble peptide-d
[5 ng (2´7 pmol); 25 ng (14 pmol); 50 ng (28 pmol); 1000 ng
(560 pmol)] or no peptide [0]. Anti-d reactivity with r4 ´ 47 and
S. mansoni 97-kDa paramyosin (Sm97) is speci®c for the d epitope.
Figure 6 r4 ´ 47, but not r140, is uniquely recognized by F-2x.
Antigens, r4 ´ 47 (5 mg); extracts from pET-15b vector-transformed
cells (5 mg); r140 (3 mg) were assayed by Western immunoblotting
using F-2x (1 : 500) and W-2x (1 : 500).
Figure 7 Cross-reactivity of anti-r4 ´ 47 with schistosome antigens.
Antigens are r4 ´ 47 [50 ng (7 pmol)]; rSj26 [5 mg (190 pmol)]; rSm26
[10 mg (380 pmol)]; rSm28 [5 mg (180 pmol)]; worm-puri®ed Sm97
[1 mg (10 pmol)]. Antigens were immunoblotted with rat anti-r4 ´ 47
(1 : 200).
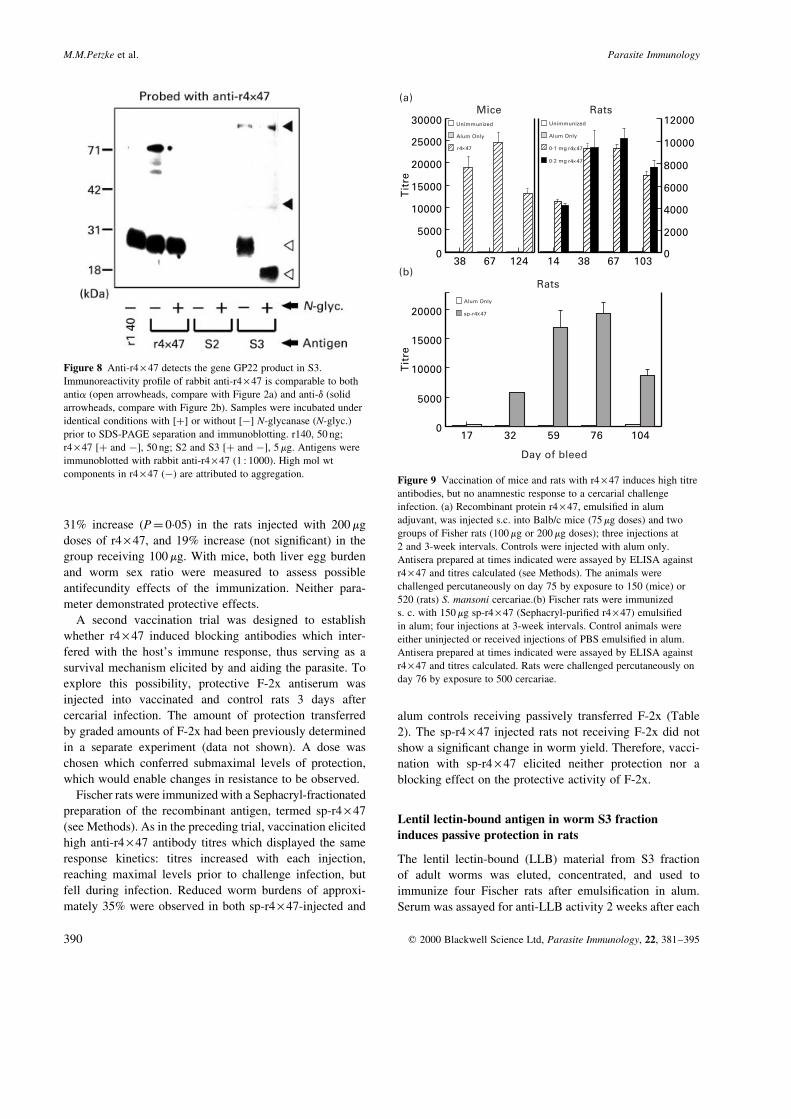
31% increase (P� 0´05) in the rats injected with 200 mg
doses of r4 ´ 47, and 19% increase (not signi®cant) in the
group receiving 100 mg. With mice, both liver egg burden
and worm sex ratio were measured to assess possible
antifecundity effects of the immunization. Neither para-
meter demonstrated protective effects.
A second vaccination trial was designed to establish
whether r4 ´ 47 induced blocking antibodies which inter-
fered with the host's immune response, thus serving as a
survival mechanism elicited by and aiding the parasite. To
explore this possibility, protective F-2x antiserum was
injected into vaccinated and control rats 3 days after
cercarial infection. The amount of protection transferred
by graded amounts of F-2x had been previously determined
in a separate experiment (data not shown). A dose was
chosen which conferred submaximal levels of protection,
which would enable changes in resistance to be observed.
Fischer rats were immunized with a Sephacryl-fractionated
preparation of the recombinant antigen, termed sp-r4 ´ 47
(see Methods). As in the preceding trial, vaccination elicited
high anti-r4 ´ 47 antibody titres which displayed the same
response kinetics: titres increased with each injection,
reaching maximal levels prior to challenge infection, but
fell during infection. Reduced worm burdens of approxi-
mately 35% were observed in both sp-r4 ´ 47-injected and
alum controls receiving passively transferred F-2x (Table
2). The sp-r4 ´ 47 injected rats not receiving F-2x did not
show a signi®cant change in worm yield. Therefore, vacci-
nation with sp-r4 ´ 47 elicited neither protection nor a
blocking effect on the protective activity of F-2x.
Lentil lectin-bound antigen in worm S3 fraction
induces passive protection in rats
The lentil lectin-bound (LLB) material from S3 fraction
of adult worms was eluted, concentrated, and used to
immunize four Fischer rats after emulsi®cation in alum.
Serum was assayed for anti-LLB activity 2 weeks after each
M.M.Petzke et al. Parasite Immunology
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395390
Figure 8 Anti-r4 ´ 47 detects the gene GP22 product in S3.
Immunoreactivity pro®le of rabbit anti-r4 ´ 47 is comparable to both
antia (open arrowheads, compare with Figure 2a) and anti-d (solid
arrowheads, compare with Figure 2b). Samples were incubated under
identical conditions with [�] or without [ÿ] N-glycanase (N-glyc.)
prior to SDS-PAGE separation and immunoblotting. r140, 50 ng;
r4 ´ 47 [� and ÿ], 50 ng; S2 and S3 [� and ÿ], 5 mg. Antigens were
immunoblotted with rabbit anti-r4 ´ 47 (1 : 1000). High mol wt
components in r4 ´ 47 (ÿ) are attributed to aggregation.
30000
25000
20000
15000
10000
5000
0
12000
10000
8000
6000
4000
2000
0
Tit
re
38 67 124 3814 67 103
(a)Mice
20000
15000
10000
5000
0
Tit
re
17 104765932
(b)Rats
Rats
Day of bleed
Unimmunized
Alum Only
Unimmunized
Alum Only
Alum Only
0.1 mg
0.2 mg
Figure 9 Vaccination of mice and rats with r4 ´ 47 induces high titre
antibodies, but no anamnestic response to a cercarial challenge
infection. (a) Recombinant protein r4 ´ 47, emulsi®ed in alum
adjuvant, was injected s.c. into Balb/c mice (75 mg doses) and two
groups of Fisher rats (100 mg or 200 mg doses); three injections at
2 and 3-week intervals. Controls were injected with alum only.
Antisera prepared at times indicated were assayed by ELISA against
r4 ´ 47 and titres calculated (see Methods). The animals were
challenged percutaneously on day 75 by exposure to 150 (mice) or
520 (rats) S. mansoni cercariae.(b) Fischer rats were immunized
s. c. with 150 mg sp-r4 ´ 47 (Sephacryl-puri®ed r4 ´ 47) emulsi®ed
in alum; four injections at 3-week intervals. Control animals were
either uninjected or received injections of PBS emulsi®ed in alum.
Antisera prepared at times indicated were assayed by ELISA against
r4 ´ 47 and titres calculated. Rats were challenged percutaneously on
day 76 by exposure to 500 cercariae.
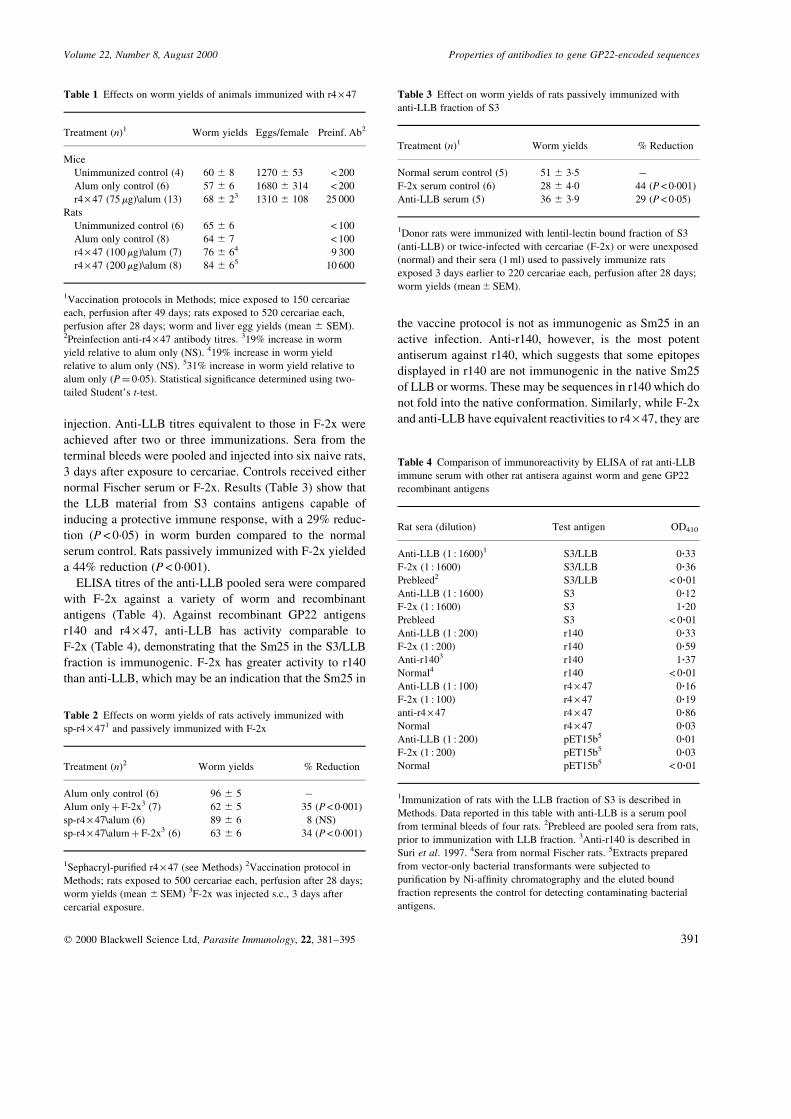
injection. Anti-LLB titres equivalent to those in F-2x were
achieved after two or three immunizations. Sera from the
terminal bleeds were pooled and injected into six naive rats,
3 days after exposure to cercariae. Controls received either
normal Fischer serum or F-2x. Results (Table 3) show that
the LLB material from S3 contains antigens capable of
inducing a protective immune response, with a 29% reduc-
tion (P < 0´05) in worm burden compared to the normal
serum control. Rats passively immunized with F-2x yielded
a 44% reduction (P< 0´001).
ELISA titres of the anti-LLB pooled sera were compared
with F-2x against a variety of worm and recombinant
antigens (Table 4). Against recombinant GP22 antigens
r140 and r4 ´ 47, anti-LLB has activity comparable to
F-2x (Table 4), demonstrating that the Sm25 in the S3/LLB
fraction is immunogenic. F-2x has greater activity to r140
than anti-LLB, which may be an indication that the Sm25 in
the vaccine protocol is not as immunogenic as Sm25 in an
active infection. Anti-r140, however, is the most potent
antiserum against r140, which suggests that some epitopes
displayed in r140 are not immunogenic in the native Sm25
of LLB or worms. These may be sequences in r140 which do
not fold into the native conformation. Similarly, while F-2x
and anti-LLB have equivalent reactivities to r4 ´ 47, they are
Volume 22, Number 8, August 2000 Properties of antibodies to gene GP22-encoded sequences
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395 391
Table 1 Effects on worm yields of animals immunized with r4 ´ 47
Treatment (n)1 Worm yields Eggs/female Preinf. Ab2
Mice
Unimmunized control (4) 60 6 8 1270 6 53 < 200
Alum only control (6) 57 6 6 1680 6 314 < 200
r4 ´ 47 (75 mg)\alum (13) 68 6 23 1310 6 108 25 000
Rats
Unimmunized control (6) 65 6 6 < 100
Alum only control (8) 64 6 7 < 100
r4 ´ 47 (100 mg)\alum (7) 76 6 64 9 300
r4 ´ 47 (200 mg)\alum (8) 84 6 65 10 600
1Vaccination protocols in Methods; mice exposed to 150 cercariae
each, perfusion after 49 days; rats exposed to 520 cercariae each,
perfusion after 28 days; worm and liver egg yields (mean 6 SEM).2Preinfection anti-r4 ´ 47 antibody titres. 319% increase in worm
yield relative to alum only (NS). 419% increase in worm yield
relative to alum only (NS). 531% increase in worm yield relative to
alum only (P� 0´05). Statistical signi®cance determined using two-
tailed Student's t-test.
Table 2 Effects on worm yields of rats actively immunized with
sp-r4 ´ 471 and passively immunized with F-2x
Treatment (n)2 Worm yields % Reduction
Alum only control (6) 96 6 5 ÿ
Alum only�F-2x3 (7) 62 6 5 35 (P< 0´001)
sp-r4 ´ 47\alum (6) 89 6 6 8 (NS)
sp-r4 ´ 47\alum�F-2x3 (6) 63 6 6 34 (P< 0´001)
1Sephacryl-puri®ed r4 ´ 47 (see Methods) 2Vaccination protocol in
Methods; rats exposed to 500 cercariae each, perfusion after 28 days;
worm yields (mean 6 SEM) 3F-2x was injected s.c., 3 days after
cercarial exposure.
Table 3 Effect on worm yields of rats passively immunized with
anti-LLB fraction of S3
Treatment (n)1 Worm yields % Reduction
Normal serum control (5) 51 6 3´5 ÿ
F-2x serum control (6) 28 6 4´0 44 (P< 0´001)
Anti-LLB serum (5) 36 6 3´9 29 (P< 0´05)
1Donor rats were immunized with lentil-lectin bound fraction of S3
(anti-LLB) or twice-infected with cercariae (F-2x) or were unexposed
(normal) and their sera (1 ml) used to passively immunize rats
exposed 3 days earlier to 220 cercariae each, perfusion after 28 days;
worm yields (mean 6 SEM).
Table 4 Comparison of immunoreactivity by ELISA of rat anti-LLB
immune serum with other rat antisera against worm and gene GP22
recombinant antigens
Rat sera (dilution) Test antigen OD410
Anti-LLB (1 : 1600)1 S3/LLB 0?33
F-2x (1 : 1600) S3/LLB 0?36
Prebleed2 S3/LLB < 0?01
Anti-LLB (1 : 1600) S3 0?12
F-2x (1 : 1600) S3 1?20
Prebleed S3 < 0?01
Anti-LLB (1 : 200) r140 0?33
F-2x (1 : 200) r140 0?59
Anti-r1403 r140 1?37
Normal4 r140 < 0?01
Anti-LLB (1 : 100) r4 ´ 47 0?16
F-2x (1 : 100) r4 ´ 47 0?19
anti-r4 ´ 47 r4 ´ 47 0?86
Normal r4 ´ 47 0?03
Anti-LLB (1 : 200) pET15b5 0?01
F-2x (1 : 200) pET15b5 0?03
Normal pET15b5 < 0?01
1Immunization of rats with the LLB fraction of S3 is described in
Methods. Data reported in this table with anti-LLB is a serum pool
from terminal bleeds of four rats. 2Prebleed are pooled sera from rats,
prior to immunization with LLB fraction. 3Anti-r140 is described in
Suri et al. 1997. 4Sera from normal Fischer rats. 5Extracts prepared
from vector-only bacterial transformants were subjected to
puri®cation by Ni-af®nity chromatography and the eluted bound
fraction represents the control for detecting contaminating bacterial
antigens.

less potent than anti-r4 ´ 47, for possibly the same reasons as
suggested for potency of anti-r140 to r140.
DISCUSSION
Antibodies to a predicted B-cell epitope of Sm25
cross-react with other candidate vaccine worm
antigens
Data presented here con®rm the identity of a predicted
glycoprotein product encoded by S. mansoni gene GP22,
cloned by our laboratory (El-Sherbeini et al. 1991], as the
Sm25 present in the tegument of adult worms, previously
described by Knight et al. (1989) and Omer-Ali et al.
(1991). Monospeci®c antibodies raised against a synthetic
peptide corresponding to a 15-codon segment termed alpha
(a) of GP22 were used to identify a glycoprotein which
possesses the following characteristics that correlate with
the properties of Sm25: (i) It is detected on Western immuno-
blots with anti-a as a 25 kDa product in a detergent-solubil-
ized fraction of worm proteins (S3, Figure 2a), consistent
with the size and localization of Sm25, which distributes
into the detergent (Triton X-114) phase of tegumental mem-
branes. (ii) It is a glycoprotein, as evidenced both by lentil-
lectin binding capacity (Figure 1b), and by mobility shift to
approximately 20 kDa following N-glycanase digestion
(Figure 2a). (iii) It reacts with anti-r4 ´ 47 and anti-r140
(Figure 8), polyspeci®c antibodies to recombinant products
of gene GP22 sequence.
Previous attempts in our laboratory to identify products of
gene GP22 had utilized an immunoprecipitation protocol
with rabbit antiserum against b-galactosidase fusion protein,
FP40, which contained one copy of the 47-amino acid
(141 bp cDNA) segment shown to be codons 117±163 of
GP22 (El-Sherbeini et al. 1991). Surprisingly, monospeci®c
antibodies to a 13-codon peptide of this segment encoding
putative B-cell epitope delta (d), do not detect Sm25 on
Western immunoblots (Figure 2b). Anti-d antibodies, how-
ever, react on Western immunoblots with proteins of approxi-
mately 35, 50 and 95 kDa in S2 and S3 (Figure 2b), in
addition to recombinant S. mansoni 26 and 28 kDa
glutathione-S-transferases (rSm26, rSm28), recombinant
26-kDa GST (rSj26) of S. japonicum (Figure 3), and
recombinant protein encoded by gene GP22, r140 (Figure
2b). Curiously, many of these antigens cross-reactive with
antipeptide-d are themselves candidate vaccine antigens
identi®ed in other studies, as follows. The most strongly
reacting antigen in worm extracts, the 95 kDa protein, was
identi®ed as S. mansoni paramyosin (Sm97): a soluble,
97 kDa schistosome protein capable of eliciting protective
immunity against S. mansoni in mice (Pearce et al. 1986,
Pearce et al. 1988). Sj26, which has been implicated in
mediating protective immunity against S. japonicum in mice,
is 70% identical in amino acid sequence (Panaccio et al.
1992) to the best characterized potential vaccine antigen of
S. mansoni, Sm28 (Auriault et al. 1988, Auriault et al.
1991), which elicits protective immunity in mice (Balloul
et al. 1987) and rats (Auriault et al. 1987, Wolowczuk et al.
1991). Genes unrelated to GP22 encode these antigens
(Sj26, Sm26, Sm28, and Sm97). The cross-reactivity of
anti-d is speci®c, as demonstrated with Sm97, and is
competed with soluble peptide-d (Figure 5).
Attempts to locate common amino acid sequences of
Sm97 and GSTs recognized by anti-d through scanning of
these schistosome proteins (performed by D. Cioli), identi-
®ed a short region of homology in peptide-d and Sm97, but
failed to detect sequence similarity between peptide-d and
GSTs. The homologous sequence consists of ®ve amino
acids, three identical and two similar: Asn±Ser±Ala±Asn±
Lys (NSANK) in peptide-d, and Asp±Thr±Ala±Asn±Lys
(DTANK) in Sm97. We attempted to determine the rele-
vance of these homologous sequences by a competition
ELISA using soluble synthetic pentapeptides NSANK and
DTANK. The pentapeptides completely failed to compete
the binding of anti-d to immobilized peptide-d, nor did anti-d
detect either of the immobilized pentapeptides (Petzke
1996).
While anti-d in Western immunoblotting does not detect
Sm25, these antibodies do recognize two recombinant
products of Sm25-encoding gene GP22 by immunoblotting:
r4 ´ 47 and r140, the latter also containing peptide-a
(Figures 3b and 2b, respectively). This result is not attribu-
table to lower af®nity of anti-d antibodies compared to anti-a
antibodies for r140, the protein which both antisera detect by
immunoblotting (Petzke 1996). However, antiserum against
recombinant protein r4 ´ 47 does detect Sm25 in addition
to r4 ´ 47 and r140 (Figure 8). Anti-r4 ´ 47 may contain
antibodies of similar speci®city as anti-d in addition to
antibodies which anti-d lacks, either to epitopes outside
of the d sequence or to alternative conformations of that
sequence within r4 ´ 47. That peptide-d does not compete
binding of anti-r4 ´ 47 to r4 ´ 47 protein (Petzke 1996) does
not resolve these possibilities. We conclude from these
results that the amino acid sequence de®ned as d can
possibly assume various conformations, some resembling
epitopes in schistosome proteins encoded by unrelated
genes (`mimotopes'), which are dependent upon the three-
dimensional context of neighbouring residues in the GP22
sequence. Alternately, multiple epitopes in addition to d are
present within the 47-amino acid segment.
Cross-reactive antibodies to the other schistosome anti-
gens (Sm97, etc.) are present in peptide-af®nity puri®ed
anti-d but are not present in peptide-af®nity puri®ed anti-a.
The carrier protein used for initial immunization of the
M.M.Petzke et al. Parasite Immunology
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395392

rabbits with peptide-d conjugate was OVA (see Methods).
Later experiments involved boosting the same rabbits with
peptide-d conjugated to KLH. The carrier protein used for
immunization of the rabbits with peptide-a conjugate was
always KLH. Since antibodies to KLH also cross-react with
schistosomal antigens (Ko & Harn 1987, Bushara et al.
1993, Thors & Linder 1998, Hamilton et al. 1999), it is
important to exclude the possibility that anti-KLH anti-
bodies were responsible for the cross reactivity observed.
Five reasons can be given which exclude a contribution
from anti-KLH in the interpretation of the results: (i)
Speci®c peptide-af®nity chromatography on a solid-phase
peptide matrix that does not include carrier protein was used
for the independent puri®cations of both anti-d and anti-a
peptide antibodies. If anticarrier antibodies were a contami-
nation and the sole source of the observed cross-reactions,
then both antipeptide antibody preparations should have
produced the observed cross-reactivity. This was obviously
not the case; as only af®nity-puri®ed antibodies to peptide-d
showed the cross reactivity (compare Figures 2 and 3). (ii)
Recombinant protein r4 ´ 47, which contains the peptide-d
sequence in four tandem copies but no KLH carrier, induced
antibodies that showed the same pattern of cross-reactivity
as antipeptide-d. Thus, con®rmation of the cross-reactivity
with a different anti-d preparation was observed indepen-
dently of KLH (see Figure 7). (iii) A competition experi-
ment was conducted using r4 ´ 47 and Sm97 as immobilized
antigens, af®nity-puri®ed anti-d antibodies, and peptide-d
(not conjugated to carrier) as the competing antigen. The
binding of anti-d antibodies to each of these antigens was
completely inhibited by addition of the free peptide only
(see Figure 5). If anti-KLH antibodies (putatively contami-
nating the af®nity puri®ed anti-d antibodies) were respon-
sible for the cross-reaction with Sm97, then peptide-d alone
should not have abolished the binding of antibodies to that
antigen. (iv) Anti-d antibodies af®nity-puri®ed from sera of
rabbits immunized with peptide-d conjugated to OVA were
used in our earliest studies. These antipeptide-d preparations
were the ones which initially provoked our excitement in
that they showed cross-reactions with a high molecular
weight antigen in S2 (see Figure 1a, peak 2a, contains
Sm97) and with sj26 GST encoded in the pGEX plasmid
vector (see Figure 3b). Thus, cross-reactivity with GST was
observed using af®nity puri®ed antipeptide-d independently
of the carrier protein used in the immunization. (v) None of
the reports involving antigens cross-reactive with anti±KLH
cite interaction with Sm97 (paramyosin) that we have
reported.
These observations eliminate the concern over anti-KLH
and clearly support the conclusion that anti-d antibodies are
responsible for the cross-reactivity. Moreover, a pilot study
with af®nity-puri®ed anti-d from a KLH-boosted rabbit
con®rms that anti-KLH is not detectable in an ELISA at
the dilutions of anti-d employed in these studies, con®rming
the success of the af®nity chromatography protocol.
Characterization of a recombinant product containing
tandem-repeats of this peptide as a vaccine
Immunization of rats and mice with r4 ´ 47 induces high
antibody titres, yet not only fails to protect against challenge
infection, but also can result in signi®cantly higher worm
burdens (Table 1). One possible explanation for this result
involves the isotype of the induced antibodies. For example,
the rat IgG2a isotype has also been associated with resist-
ance to infection, possibly via complement ®xation or
ADCC. The protective ability of IgG2a can be inhibited
by the presence of blocking antibodies of the IgG2c isotype
for a glycoprotein antigen (Grzych et al. 1982). Isotyping
studies of antigen-speci®c antibodies in r4 ´ 47 detected
IgG2a, but not IgG2c (Petzke 1996). Another approach to
testing the blocking concept was to vaccinate with the
putative inducer of blocking antibodies (r4 ´ 47) and then
to passively immunize with protective antiserum F-2x
(Table 2). Anti-r4 ´ 47 antibodies did not interfere with
the protective activity of F-2x. However, this does not
exclude the presence of blocking antibodies, if antibodies
to other antigens are also contributing to protection.
An alternative explanation to lack of protection by r4 ´ 47
and by the other recombinant product we tested, r140
(Suri et al. 1997), concerns the protective capacity of the
epitopes in these gene GP22 products. Sm25 may not be the
immunogen used by worms to produce the protective
immune response. To address this possibility, we passively
vaccinated rats with antiserum to the lentil-lectin bound S3
fraction (S3/LLB) of worms, which contains the authentic
Sm25 glycoprotein, and successfully conferred protection
(Table 3). Anti-S3/LLB and F-2x had comparable ELISA
antibody activity to the S3/LLB and to recombinant antigens
r4 ´ 47 and r140 (Table 4). While this does not demonstrate
that the anti-GP22 antibodies in either serum contribute to
the protective activity, this possibility cannot be excluded.
Since the lentil-lectin bound S3 is a mixture of products,
however, one also cannot exclude the possibilty that it was
a combination of antibodies to these other S3 antigens
(without anti-Sm25), or perhaps even anticarbohydrate anti-
bodies, which were responsible for protection. The prokary-
otic recombinant products tested in our study lacked
glycosylation.
To conclude, antibodies to peptide-a (amino acids 70±84
of Sm25) identify Sm25 in the detergent-soluble (but not the
aqueous-soluble) fraction of juvenile and adult schisto-
somes. Antibodies to peptide-d (amino acids 151±162 of
Sm25) do not react with Sm25 or its deglycosylated product
Volume 22, Number 8, August 2000 Properties of antibodies to gene GP22-encoded sequences
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395 393

but do react with r4 ´ 47 and r140, recombinant proteins
with peptide-d as a portion of their amino acid sequence.
Anti-a reacts with the recombinant protein r140, which has
peptide-a as a portion of its amino acid sequence. Anti-d
(but not anti-a) cross-reacts with other schistosome anti-
gens: Sm97, GSTs of S. mansoni (Sm26; Sm28) and S.
japonicum (Sj26) and a 50 kDa and a 35 kDa antigen of
S. mansoni yet to be de®ned. Anti-r4 ´ 47 antibodies react
with Sm25 and both recombinant antigens, and also cross-
react with other schistosome antigens detected by anti-d.
Thus, recombinant protein r4 ´ 47 possesses the `F-2x-unique
epitope(s)' and the epitope(s)/mimotope(s) which induce
cross-reactivity. The d sequence may have multiple con-
formations dependent upon neighbouring sequences; in
r4 ´ 47, the d sequence may assume some of these conforma-
tions and possibly others which are infrequent in peptide-d.
We hypothesize that the infrequent conformations are
present in Sm25.
The lentil lectin bound fraction of detergent-solubilized
adult worms induces protective antibodies in rats, including
antibodies to r4 ´ 47. However, vaccination with r4 ´ 47 is
not protective in mice and rats, despite inducing high titres
of antibodies to r4 ´ 47. Nor do anti-r4 ´ 47 antibodies inter-
fere with protection transferred by F-2x. Two hypotheses to
account for this disparity include the requirement of antipoly-
saccharide antibodies not induced by the E. coli-expressed
fusion proteins, and the possibility that the mimotopes against
which the cross-reactive antibodies were produced are not the
target epitopes of protective antibodies.
ACKNOWLEDGEMENTS
The authors wish to thank the following individuals for their
contributions to this research: Isla Garraway, whose under-
graduate honors project generated preliminary data on the
lentil lectin fractionation of worm extracts; Frank DeSilva
and Jonathan Kurtis, who assisted with several experiments
and provided technical advice; Ken Nguyen, whose assis-
tance with immunizations and perfusions is greatly appre-
ciated; Carol Macomber and Linda Fitzgibbons for technical
assistance; and Dr Andrew Campbell, whose suggestions
regarding the competition Western immunoblotting proce-
dure and critique of the results are greatly appreciated. This
project was supported by grants to P.M.K from the UNDP/
World Bank/WHO Special Program for Research and Training
in Tropical Diseases (TDR-900293) and the US Public Health
Service, NIH (AI-31224). M.M.P. received support from an
NIH Predoctoral Training Grant (5T32 GM07601±17).
REFERENCES
Auriault C., Balloul J.-M., Pierce R.J. et al. (1987) Helper T cells
induced by a puri®ed 28-kilodalton antigen of Schistosoma
mansoni protect rats against infection. Infection and Immunity
55, 1163±1169
Auriault C., Gras-Masse H., Wolowczuk I. et al. (1988) Analysis of
T and B cell epitopes of the Schistosoma mansoni P28 antigen in the
rat model by using synthetic peptides. Journal of Immunology 141,
1687±1694
Auriault C., Wolowczuk I., Gras-Masse H. et al. (1991) Review: epitopic
characterization and vaccinal potential of peptides derived from a
major antigen of Schistosoma mansoni (Sm28 GST). Peptide
Research 4, 6±11
Balloul J.-M., Grzych J.-M., Pierce R. & Capron A. (1987) A puri®ed
28 000 dalton protein from Schistosoma mansoni adult worms
protects rats and mice against experimental schistosomiasis. Journal
of Immunology 138, 3448±3453
Barker R.H. Jr, Srivastava B.S., Suri P., Goldberg M. & Knopf P.M.
(1985) Immunoprecipitation analysis of radiolabelled protein anti-
gens biosynthesized in vitro by S. mansoni. I. Identi®cation of
antigens uniquely recognized by protective antibodies. Journal of
Immunology 134, 1192±1201
Bushara H.O., Bashir M.E., Malik K.H. et al. (1993) Suppression of
Schistosoma bovis egg production with either glutathione S-transferase
or keyhole limpet hemocyanin. Parasite Immunology 15, 383±390
Cioli D., Knopf P.M. & Senft A.W. (1977) A study of Schistosoma
mansoni transferred into permissive and nonpermissive hosts. Inter-
national Journal for Parasitology 7, 293±297
El-Sherbeini M., Bostian K.A. & Knopf P.M. (1990) Schistosoma
mansoni: cloning of antigen gene sequences in Escherichia Coli.
Experimental Parasitology 70, 72±84
El-Sherbeini M., Ramadan N., Bostian K.A. & Knopf P.M. (1991)
Cloning and sequence analysis of the Schistosoma mansoni mem-
brane glycoprotein antigen gene GP22. Molecular and Biomedical
Parasitology 49, 83±98
Grzych J.M., Capron M., Bazin H. & Capron A. (1982) In vitro and in
vivo effector function of rat IgG2a monoclonal anti-S. mansoni
antibodies. Journal of Immunology 129, 2739±2743
Hamilton J.V., Chiodini P.L., Fallon P.G. & Doenhoff M.J. (1999)
Periodate-sensitive immunological cross-reactivity between keyhole
limpet hemocyanin (KLH) and serodiagnostic Schistosoma mansoni
egg antigens. Parasitology 118, 83±89
Knight M., Kelly C., Rodrigues V. et al. (1989) A cDNA clone encoding
part of the major 25000-dalton surface membrane antigen of adult
Schistosoma mansoni. Parasitology Research 75, 280±286
Knopf P.M., Nutman T.B. & Reasoner J.A. (1977) Schistosoma
mansoni: resistance to reinfection in the rat. Experimental Parasitol-
ogy 41, 74±82
Ko A.I. & Harn D.A. Jr (1987) Characterization of protective and non-
protective surface membrane carbohydrate epitopes of Schistosoma
mansoni. Memorias Do Instituto Oswald Cruz 82 (Suppl. 4), 115±
119
Mangold B.L. & Knopf P.M. (1981) Host-protective humoral immune
response to Schistosoma mansoni infections in the rat: kinetics
of hyperimmune serum-dependent sensitivity and elimination of
schistosomes in a passive transfer system. Parasitology 83, 559±574
Maniatis T., Fritsch E.F. & Sambrook J. (1982) Molecular Cloning: a
Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring,
Harbor, NY
Mark H.F.L., El-Sherbeini M., Goldberg M. et al. (1991) Schistosoma
mansoni: two-dimensional gel electrophoretic analysis of antigens
uniquely recognized by protective antibodies. Experimental Para-
sitology 72, 294±305
McCray J.W. Jr & Werner G. (1989) Production and properties of site-
speci®c antibodies to synthetic peptide antigens related to potential
M.M.Petzke et al. Parasite Immunology
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395394

cell surface receptor sites for rhinovirus. Methods in Enzymology
178, 676±692
Omer-Ali P., Jeffs S.A., Meadows H. et al. (1991) Structure of
Sm25, an antigenic integral membrane glycoprotein of adult
Schistosoma mansoni. Molecular and Biochemical Parasitology
45, 215±222
Panaccio M., Wilson L.R., Crameri S.L., Wijffels G.L. & Spithill T.W.
(1992) Molecular characterization of cDNA sequences encoding
glutathione S-transferases of Fasciola hepatica. Experimental Para-
sitology 74, 232±237
Pearce E.J., James S.L., Dalton J. et al. (1986) Immunochemical
characterization and puri®cation of Sm-97, a Schistosoma mansoni
antigen monospeci®cally recognized by antibodies from mice
protectively immunized with a nonliving vaccine. Journal of Immu-
nology 137, 3593±3600
Pearce E.J., James S.L., Hieny S., Lanar D.E. & Sher A. (1988)
Induction of protective immunity against Schistosoma mansoni by
vaccination with schistosome paramyosin (Sm97), a nonsurface
parasite antigen. Proceedings of the National Academy of Science
USA 85, 5678±5682
Petzke M.M. (1996) Immunological Characterization of a 47-amino
acid Segment of a Candidate Vaccine Schistosome Antigen, PhD
Thesis, Brown University
Riveau, G.J. & Capron, A (1997) In New Generatrion Vaccines, 2
edn, eds Levine, M.M., Woodrow, G.C., Kaper, J.B. & Cobon, G.S.,
pp. 1081±1093, Marcel Dekker, Inc., New York
Smithers S.R., Hackett F., Omer-Ali P. & Simpson A.J.G. (1989)
Protective immunization of mice against Schistosoma mansoni with
puri®ed adult worm surface membranes. Parasite Immunology 11,
301±318
Suri P.K., Goldberg M., Madikizela M. et al. (1997) Evaluation of
recombinant protein r140, a polypeptide segment of tegumental
glycoprotein Sm25, as a de®ned antigen vaccine against Schistosoma
mansoni. Parasite Immunology 19, 515±529
Thors C. & Linder E. (1998) Cross-reacting antibodies against keyhole
limpet hemocyanin may interfere with the diagnostics of acute
schistosomiasis. Parasite Immunology 20, 489±496
Torian B.E., Flores B.M., Stroeher V.L., Hagen F.S. & Stamm W.E.
(1990) cDNA sequence analysis of a 29-kD cysteine-rich surface
antigen of pathogenic Entamoeba histolytica. Proceedings of the
National Academy of Science USA 87, 6358±6362
Wolowczuk I., Auriault C., Bossus M. et al. (1991) Antigenicity and
immunogenicity of a multiple peptidic construction of the Schisto-
soma mansoni Sm-28 GST antigen in rat, mouse and monkey. 1.
Partial protection of Fischer rat after active immunization. Journal of
Immunology 146, 1987±1995
Volume 22, Number 8, August 2000 Properties of antibodies to gene GP22-encoded sequences
q 2000 Blackwell Science Ltd, Parasite Immunology, 22, 381±395 395




















