
RT-PCR cloning, characterization and mRNA expression analysis of a cDNA encoding a type II...
11
RT-PCR cloning, characterization and mRNA expression analysis of a cDNA encoding a type II asparagine synthetase in common bean Daniel Osuna, Gregorio Ga ¤lvez, Manuel Pineda*, Miguel Aguilar Departamento de Bioqu| ¤mica y Biolog| ¤a Molecular, e Instituto Andaluz de Biotecnolog| ¤a, Facultad de Ciencias, Universidad de Co ¤rdoba, Avda. San Alberto Magno, s/n. 14071 Co ¤rdoba, Spain Received 14 September 1998; received in revised form 21 December 1998; accepted 28 January 1999 Abstract Following a RT-PCR strategy based on the design of degenerate oligonucleotides resembling conserved domains of asparagine synthetase (AS; EC 6.3.5.4), we isolated a 2 kb cDNA clone (PVAS2) from root tissue of the common bean (Phaseolus vulgaris). PVAS2 encodes a protein of 584 amino acids with a predicted relative molecular mass of 65 810 Da, an isoelectric point of 6.4, and a net charge of 37.2 at pH 7.0. The amino acid sequence of the protein encoded by PVAS2 is very similar to that encoded by the soybean SAS2 asparagine synthetase gene. The amino-terminal residues of the predicted PVAS2 protein are identical to the amino acids that constitute the glutamine-binding (GAT) domain of AS from other plant species, which suggests that the PVAS2 cDNA encodes a type II glutamine-dependent form of asparagine synthetase. Southern blot analysis indicates that the common bean AS is part of a small family composed of at least two genes. Expression analysis by Northern blot revealed that the PVAS2 transcript accumulates to a high level in roots and, to a lesser extent, in nodules and developing pods. Accumulation of the PVAS2 transcript in the root seems to be negatively regulated by light and sucrose, and positively regulated by nitrate. ß 1999 Elsevier Science B.V. All rights reserved. Keywords : Asparagine synthetase ; cDNA clone ; Reverse transcriptase-polymerase chain reaction ; (Phaseolus) 1. Introduction Atmospheric nitrogen is biologically ¢xed in le- gume radical nodules. Although glutamine is the ini- tial product of ammonia assimilation during nitrogen ¢xation, this amino acid is rarely transported from the nodule into the xylem. Most nodulated legumes tend to export the ureides allantoin and allantoic acid (tropical species, i.e. cowpea, soybean, mung bean, common bean) [1], or the amide asparagine (mainly temperate species, i.e. lupin, pea, clovers) via xylem to other plant organs [2,3]. The relative distribution of ureides and amides in xylem is also determined by a number of developmental, nutrition- al and environmental factors [4,5]. Ureides, having four atoms of nitrogen per mole- cule (C:N, 1:1), have an ‘advantage’ over asparagine with only two atoms of nitrogen per molecule (C:N, 2:1) in terms of nitrogen export and e/cient carbon use. On the other hand, the low solubility of ureides may limit the amount of nitrogen that can be trans- ported under this form in temperate conditions. In ureide-exporting species, the ureides dominate xylem composition only while the plant satis¢es its nitrogen requirements mainly from the atmospheric nitrogen. In amide-producing legumes, asparagine is always 0167-4781 / 99 / $ ^ see front matter ß 1999 Elsevier Science B.V. All rights reserved. PII:S0167-4781(99)00016-0 * Corresponding author. Fax: +34 (957) 218606; E-mail : [email protected] Biochimica et Biophysica Acta 1445 (1999) 75^85
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of RT-PCR cloning, characterization and mRNA expression analysis of a cDNA encoding a type II...

RT-PCR cloning, characterization and mRNA expression analysis of acDNA encoding a type II asparagine synthetase in common bean
Daniel Osuna, Gregorio Galvez, Manuel Pineda*, Miguel AguilarDepartamento de Bioqu|mica y Biolog|a Molecular, e Instituto Andaluz de Biotecnolog|a, Facultad de Ciencias, Universidad de Cordoba,
Avda. San Alberto Magno, s/n. 14071 Cordoba, Spain
Received 14 September 1998; received in revised form 21 December 1998; accepted 28 January 1999
Abstract
Following a RT-PCR strategy based on the design of degenerate oligonucleotides resembling conserved domains ofasparagine synthetase (AS; EC 6.3.5.4), we isolated a 2 kb cDNA clone (PVAS2) from root tissue of the common bean(Phaseolus vulgaris). PVAS2 encodes a protein of 584 amino acids with a predicted relative molecular mass of 65 810 Da, anisoelectric point of 6.4, and a net charge of 37.2 at pH 7.0. The amino acid sequence of the protein encoded by PVAS2 is verysimilar to that encoded by the soybean SAS2 asparagine synthetase gene. The amino-terminal residues of the predictedPVAS2 protein are identical to the amino acids that constitute the glutamine-binding (GAT) domain of AS from other plantspecies, which suggests that the PVAS2 cDNA encodes a type II glutamine-dependent form of asparagine synthetase.Southern blot analysis indicates that the common bean AS is part of a small family composed of at least two genes.Expression analysis by Northern blot revealed that the PVAS2 transcript accumulates to a high level in roots and, to a lesserextent, in nodules and developing pods. Accumulation of the PVAS2 transcript in the root seems to be negatively regulatedby light and sucrose, and positively regulated by nitrate. ß 1999 Elsevier Science B.V. All rights reserved.
Keywords: Asparagine synthetase; cDNA clone; Reverse transcriptase-polymerase chain reaction; (Phaseolus)
1. Introduction
Atmospheric nitrogen is biologically ¢xed in le-gume radical nodules. Although glutamine is the ini-tial product of ammonia assimilation during nitrogen¢xation, this amino acid is rarely transported fromthe nodule into the xylem. Most nodulated legumestend to export the ureides allantoin and allantoicacid (tropical species, i.e. cowpea, soybean, mungbean, common bean) [1], or the amide asparagine(mainly temperate species, i.e. lupin, pea, clovers)
via xylem to other plant organs [2,3]. The relativedistribution of ureides and amides in xylem is alsodetermined by a number of developmental, nutrition-al and environmental factors [4,5].
Ureides, having four atoms of nitrogen per mole-cule (C:N, 1:1), have an `advantage' over asparaginewith only two atoms of nitrogen per molecule (C:N,2:1) in terms of nitrogen export and e¤cient carbonuse. On the other hand, the low solubility of ureidesmay limit the amount of nitrogen that can be trans-ported under this form in temperate conditions. Inureide-exporting species, the ureides dominate xylemcomposition only while the plant satis¢es its nitrogenrequirements mainly from the atmospheric nitrogen.In amide-producing legumes, asparagine is always
0167-4781 / 99 / $ ^ see front matter ß 1999 Elsevier Science B.V. All rights reserved.PII: S 0 1 6 7 - 4 7 8 1 ( 9 9 ) 0 0 0 1 6 - 0
* Corresponding author. Fax: +34 (957) 218606;E-mail : [email protected]
BBAEXP 93250 2-4-99
Biochimica et Biophysica Acta 1445 (1999) 75^85

the main form of reduced nitrogen despite changes ofnitrogen nutritional status. In spite of these estab-lished experimental facts, the mechanisms by whichplants are determined to produce ureides or amidesremain elusive [2,4,6].
Asparagine biosynthesis in plants is mediatedmainly by the action of asparagine synthetase (AS;EC 6.3.5.4). There are two types of asparagine syn-thetases: a glutamine-dependent form [7^9], and anammonia-dependent one [10]. In plants, the mostcommon is the glutamine-dependent form, which cat-alyzes the ATP-dependent transfer of the amidegroup from glutamine to aspartate, and yields gluta-mate and asparagine [11]. The biochemical studies ofplant asparagine synthetases have encountered majordi¤culties due to enzyme instability [12], the pres-ence of heat stable inhibitors of AS in crude extracts[13], and the presence of contaminating asparaginaseactivity [14]. As a result, only partial puri¢cations ofasparagine synthetase have been achieved so far[3,12,15,16]. A molecular approach has been usedwith more success, and several AS have been clonedfrom di¡erent sources: Pisum sativum [14,16], Aspa-ragus o¤cinalis [17], Zea mays [18], Arabidopsis thali-ana [19], Glycine max [20], among others. Still, reg-ulation of the di¡erent isoforms of asparaginesynthetase is poorly understood.
We have undertaken the cloning of the asparaginesynthetase gene(s) from a ureide legume as Phaseolusvulgaris, because of our idea that some molecularfactors might be responsible for the integration andco-regulation of ureide and amide biosynthesis in le-gume roots. In the present paper, we report thecDNA cloning and expression analysis of PVAS2, atype II asparagine synthetase gene from Ph. vulgaris.
2. Materials and methods
2.1. Materials
All plant tissues were isolated from common bean(Ph. vulgaris) cv. Creat Northern (Eurosemillas,Spain). Seeds were imbibed and germinated in asand/vermiculite medium (1:3). After germination,plants were inoculated with a fresh suspension ofRhizobium leguminosarum (ISC14 strain) which hadbeen cultured at low temperatures (below 30³C) to
prevent the loss of plasmidic ¢xation genes. Plantswere cultured in growth chambers under a long dayphotoperiod (16 h light, 8 h dark), 70% relative hu-midity and 22³C. Escherichia coli DH5KFP [21] wasused in all the cloning experiments.
Restriction enzymes, T4 ligase, DNA polymeraseKlenow fragment, DNA and RNA markers, alkalinephosphatase from calf thymus, Taq DNA polymer-ase and pBluescript SK/KS were acquired fromBoehringer Mannheim, Perkin Elmer, Stratagene orAmersham. All enzymes and reagents for RACE(rapid ampli¢cation of cDNA ends) were from theMarathon cDNA ampli¢cation system (Clontech).Oligonucleotides were purchased from MWG-Bio-tech. All other common reagents were from Sigmaor Amersham.
2.2. Isolation of Phaseolus AS cDNA clones
Highly conserved regions of the AS gene wereidenti¢ed by comparing AS sequences from Z.mays [18], Asp. o¤cinalis [17], P. sativum [14] andA. thaliana [19]. Degenerate oligonucleotide primerswere designed to resemble conserved regions: ASP2,5P-ATG AC(A, C, T) GA(C, T) GT(G, C, T) CC(C,T) TT(C, T) GG-3P ; ASP3, 5P-TA(A, T) CC(A, T)CC(A, G) AA(A, G, T) AT(C, T) TC(A, G) TC-3P).A minimal degeneracy was introduced taking intoaccount the codon usage for Phaseolus. Theseprimers were used in a polymerase chain reaction(PCR) with Phaseolus cDNA synthesized with Super-script II (Gibco BRL) from root mRNA of lightgrown plants to amplify a fragment of the ASgene. The following PCR conditions were used: aninitial denaturation step at 94³C, followed by 94³Cfor 1 min, 50³C for 1 min, and 72³C for 1 min (40cycles), with a 7 min ¢nal extension. A 388 bp PCRproduct was generated by this procedure. After clon-ing and sequencing this internal AS cDNA fragment(p380PVAS2), two gene-speci¢c primers were de-signed so that the ampli¢ed cDNA ends wouldhave a 300 nucleotide overlap: ASN11 (5P-CTCCGG AGA TAA CCC ATT TGA CAC C-3P),ASN10 (5P-GGC ATC CAT CAC TTC TCG CTACTT G-3P). AS11 and AS10 were used for 5P- and 3P-RACE, respectively, as gene-speci¢c primers with aMarathon cDNA ampli¢cation system. The ampli¢-cation generated a 1.1 kb 5P-end cDNA fragment
BBAEXP 93250 2-4-99
D. Osuna et al. / Biochimica et Biophysica Acta 1445 (1999) 75^8576

and a 1.3 kb 3P-end cDNA fragment. Both partialcDNAs were cloned in pBluescript (p5PVAS2 andp3PVAS2) for nucleotide sequencing. A full-lengthcDNA was ampli¢ed and cloned in pBluescript(pPVAS2), after a fusion reaction of the overlapping5P-end and 3P-end fragments following the protocolsof the Marathon system. In addition, a 1.9 kb cDNAwas ampli¢ed directly using the two gene-speci¢c pri-mers AS20 (5P-TCT GAT CAC TCT CGA GCCAAA AG-3P) and AS21 (5P-TGC GTG CAT AACTGA GAA AAT AA-3P). This cDNA was ¢nallycloned in pBluescript (p2PVAS2) for sequencing.
Plasmid DNA was isolated by the alkaline lysismethod [22] in Qiagen plasmid minipreps. DNAwas double strand sequenced using the dye termina-tor method [23] in an ABI 310 automatic sequencer(Perkin Elmer).
Protein and DNA databases were searched for se-quence similarities using the BLAST algorithm [24].For multiple sequence comparisons, the Lipman-Pearson method of the Lasergene software package(DNASTAR) was used. A dendrogram was con-structed using the Kimura distance algorithm [25]to unveil evolutionary relationships among severalAS amino acid sequences from di¡erent species.
2.3. DNA and RNA analyses
For Southern blot analyses, total genomic DNAwas prepared from common bean young leaves.Fresh material was ground with pestle and mortarin the presence of glass beads and proteinase Kbu¡er (0.2 M Tris-HCl (pH 8)/0.1 M EDTA/1%(w/v) N-lauroylsarcosine/100 Wg/ml proteinase K),transferred to Corex tubes and incubated at 50³Cfor 1 h. After low speed centrifugation, the super-natant was ethanol precipitated. The pellet of nucleicacids was dissolved in TE and ethanol precipitatedonce more. DNA was further puri¢ed by ultracentri-fugation in a CsCl gradient, subjected to two morerounds of ethanol precipitation and ¢nally dissolvedin TE. DNA samples were digested with restrictionenzymes, separated by electrophoresis in 0.8% agar-ose gels, and blotted onto Hybond N� membranesby capillary blotting using 0.4 N NaOH as transfersolution. Nylon membranes were hybridized with the388 bp AS cDNA fragment from clone p380PVAS2.Hybridization probes were labeled with the oligola-
beling kit (Pharmacia) following the manufacturer'sinstructions. Prehybridizations and hybridizationswere performed according to Church and Gilbert[26] at 65³C for 6 and 18 h, respectively. Blots werewashed twice at a stringency of 1USSC/0.1% SDS atroom temperature, twice at a stringency of 2USSC/0.1% SDS at 42³C, and twice more with 0.1USSC/0.1% SDS at 42³C. X-Ray ¢lms were then exposed tothe blots for 72 h at 380³C. Afterwards, the blotswere washed twice at a stringency of 2USSC/0.1%SDS at 65³C and twice more with 0.1USSC/0.1%SDS at 65³C. Finally, new X-ray ¢lms were exposedto the blots for 72 h at 380³C.
For Northern blot analyses, total RNA was iso-lated from several common bean tissues (nodules,developing pods, £owers, roots, stems and leaves)grown under di¡erent conditions. Frozen samplesof the di¡erent organs and tissues were groundedwith mortar and pestle and homogenized at 4³C inextraction bu¡er: 200 mM Tris-borate (pH 7.6), sup-plemented with 10 mM EDTA, 0.5% SDS and 0.5%L-mercaptoethanol (3 ml bu¡er per gram of tissue).The homogenate was mixed with an equal volume ofa solution of phenol, chloroform and isoamyl alcohol(50:50:1, v/v/v), vigorously shaken, equilibrated toroom temperature, and centrifuged at 10 000Ug for10 min at 15³C. The supernatant was diluted with2 vols. of water, and sodium acetate was added toa ¢nal concentration of 80 mM. Additional L-mer-captoethanol (0.4 vol.) was added to the extract, in-cubated at 4³C for 30 min, and centrifuged at10 000Ug for 20 min. The supernatant was supple-mented with 0.6 vol. of 0.5% L-mercaptoethanol, in-cubated at 4³C for 30 min, and centrifuged at10 000Ug for 20 min. The nucleic acids pellet waswashed with 70% ethanol, 100 mM potassium ace-tate, and centrifuged at 10 000Ug for 5 min. After anadditional wash with 100% ethanol, the pellet wasdissolved in water. Finally, 1 vol. of 4 M LiCl wasadded and centrifuged at 10 000Ug for 30 min. Thetotal RNA pellet was washed as before, and dis-solved in water. Total RNA was size-fractionatedon a denaturing formaldehyde agarose gel and trans-ferred onto nylon membranes by capillary blotting.Membranes were hybridized with a probe con-structed with a 866 bp fragment including the 3P-un-translated region and the carboxyl terminus of thePVAS2 cDNA (nucleotides 1089^1955). This probe
BBAEXP 93250 2-4-99
D. Osuna et al. / Biochimica et Biophysica Acta 1445 (1999) 75^85 77

was constructed by PCR using the followingprimers: AS15 (5P-TTT CAC AAG GCA CCCAAC AAG-3P), AS21 (5P-TGC GTG CAT AACTGA GAA AAT AA-3P). Prehybridization and hy-bridization were carried out at 65³C in Church bu¡er[26]. The ¢nal wash for the Northern blots was at42³C with a stringency of 0.1USSC/0.1% SDS.X-Ray ¢lms were exposed to the membranes for3 days at 380³C.
3. Results
3.1. Isolation and characterization of a PVAS2 cDNAclone
A 380 bp internal fragment of a putative AScDNA (p380PVAS2) was ¢rst isolated from totalroot RNA by RT-PCR using degenerate primersASP2/ASP3. The sequence of this internal cDNAfragment allowed the design of gene-speci¢c primersthat were used in 5P- and 3P-RACE PCR experi-ments. Clones p5PVAS2 and p3PVAS2 were isolatedand sequenced. This allowed the reconstruction of analmost full-length cDNA of 2044 bp (pPVAS2). Con-sidering the existence of at least two AS genes in thecommon bean, and to discard the possibility that thereconstructed full-length AS cDNA (pPVAS2) couldbe a hybrid of the two genes, we performed a ¢nalPCR with two gene-speci¢c primers AS20 and AS21.The 1.9 kb PCR product was cloned in pBluescript.Both ends of this cDNA were sequenced, and thesequences compared to those of the 5P- and 3P-RACE-PCR products previously isolated. We couldconclude that PVAS2 cDNA is not a hybrid, but itreally represents a single gene. The nucleotide anddeduced amino acid sequence corresponding to thelongest open reading frame (ORF) of PVAS2 cDNAis shown in Fig. 1. The putative ATG start codon islocated at nucleotides 1^3. A 5P in-frame stop codon(TAA) is located 12 nucleotides upstream from thesuggested start codon, which indicates that the trans-lation of this ORF could not start further upstream.A putative polyadenylation signal (AATAAGA) islocated at nucleotides 1941^1947. This signal ishighly homologous to the consensus sequence ofplant polyadenylation signal (AAUAAAA) [27].The open reading frame from the putative start co-
don to the stop codon (TAA) encodes a 584 aminoacid peptide with a calculated relative molecularmass of 65 810 Da, an isoelectric point of 6.4, anda net charge of 37.2 at pH 7.0.
The deduced amino acid sequence of PVAS2 wasaligned with the sequences of soybean, pea, lotus,faba bean, Arabidopsis, E. coli and human aspara-gine synthetases to determine conserved regions with-in these sequences (Fig. 2). PVAS2 shares highly ho-mologous regions with all asparagine synthetases,beginning with the proposed glutamine-binding do-main (GAT), which is very speci¢c to AS and otherNtn amide transferases [28]. All the essential residuesof this domain (Cys1-Arg26-Gly27-Gly32-Arg73-Pro86-Asn101-Gly102-Asp127, PRPP amidotransferase nota-tion) are conserved in the AS from soybean [20],pea [14], lotus [29], faba bean [30], Arabidopsis [19],human [31], E. coli [32], and also in the commonbean PVAS2 (Cys1-Arg30-Gly31-Gly36-P60-Asn74-Gly75-Asp97, PVAS2 notation after removal of initialmethionine).
Essential residues for the binding of aspartate andATP are also conserved in PVAS2. Three amino acidresidues (Thr315, Thr316 and Arg318) were previouslyproved to take part in aspartate binding or stabiliza-tion of intermediates in the formation of L-aspartyl-AMP in E. coli ASNB [33]. Cys522, which is alsoconserved in all asparagine synthetases except humanAS1, was suggested to be part of the aspartate bind-ing site of E. coli ASNB [34]. Another amino acidstretch, located between Ser233 and Ser237 (PVAS2notation) is conserved throughout all AS and otherATP-hydrolyzing enzymes like GMP synthetase [35]and ATP sulfurylase [36]. Recent computer analysisof E. coli ASNB protein sequence suggests that theseresidues might be involved in pyrophosphate binding[37,38].
As shown in Fig. 2, the predicted amino acid se-quence of PVAS2 displayed signi¢cant similarity toother asparagine synthetases. It showed the highesthomology to the AS from other legumes like soybeanAS2 (95.9%), lotus AS1 (90.9%), pea AS2 (90.6%),and fava bean AS (86.1%), while it had a much lowersimilarity to E. coli ASNB (52.3%) and human AS1(31.9%). In addition, the percentage of amino acidsimilarity and identity, together with the dendrogramshown in Fig. 3, indicates that PVAS2 is more closelyrelated to other type II asparagine synthetases.
BBAEXP 93250 2-4-99
D. Osuna et al. / Biochimica et Biophysica Acta 1445 (1999) 75^8578

Southern blot analysis was used to analyze thecomplexity of the asparagine synthetase gene family.Total genomic DNA was digested with several re-striction enzymes (EcoRI, BamHI, HindIII andPstI), run on a 0.8% agarose gel, and Southern blots
were probed with a 388 32P-labeled cDNA fragment(nucleotides 688^1075) of PVAS2 (Fig. 4). Underhigh stringency hybridization conditions, and lowstringency washes, multiple fragments of genomicDNA hybridized to this PVAS2 fragment at low
Fig. 1. Nucleotide sequence of the cDNA encoding common bean AS2. The nucleotide sequence of PVAS2 is shown in the mRNAsense. The deduced amino acid sequence is denoted below in the standard one-letter code. The translation stop codon is designatedwith an asterisk (*). A putative polyadenylation signal is underlined.
BBAEXP 93250 2-4-99
D. Osuna et al. / Biochimica et Biophysica Acta 1445 (1999) 75^85 79

stringency. Some of the fragments produced by agiven restriction enzyme hybridized more weaklythan the other fragments, and they practically disap-peared after higher stringency washes. This hybrid-
ization pattern is consistent with the existence of atleast two asparagine synthetase genes in commonbean.
Fig. 2. Amino acid sequence alignment of the protein encoded by PVAS2 with several deduced asparagine synthetases. The Clustalmethod of the Lasergene software package of DNASTAR was used to perform the alignment. Dots indicate identical amino acids,and dashes represent the gaps introduced to maximize homology. The AS polypeptides used in the comparison are common beanAS2 (PVAS2), soybean AS2 (GMAS2 [20]), pea AS2 (PSAS2 [14]), lotus AS1 (LJAS1 [29]), fava bean AS1 (VFAS1 [30]), ArabidopsisAS2 (ATAS2 [19]), E. coli ASNB (ECASB [32]), and human AS1 (HSAS1 [31]). Essential residues of the GAT and the synthetase do-mains are underlined.
BBAEXP 93250 2-4-99
D. Osuna et al. / Biochimica et Biophysica Acta 1445 (1999) 75^8580
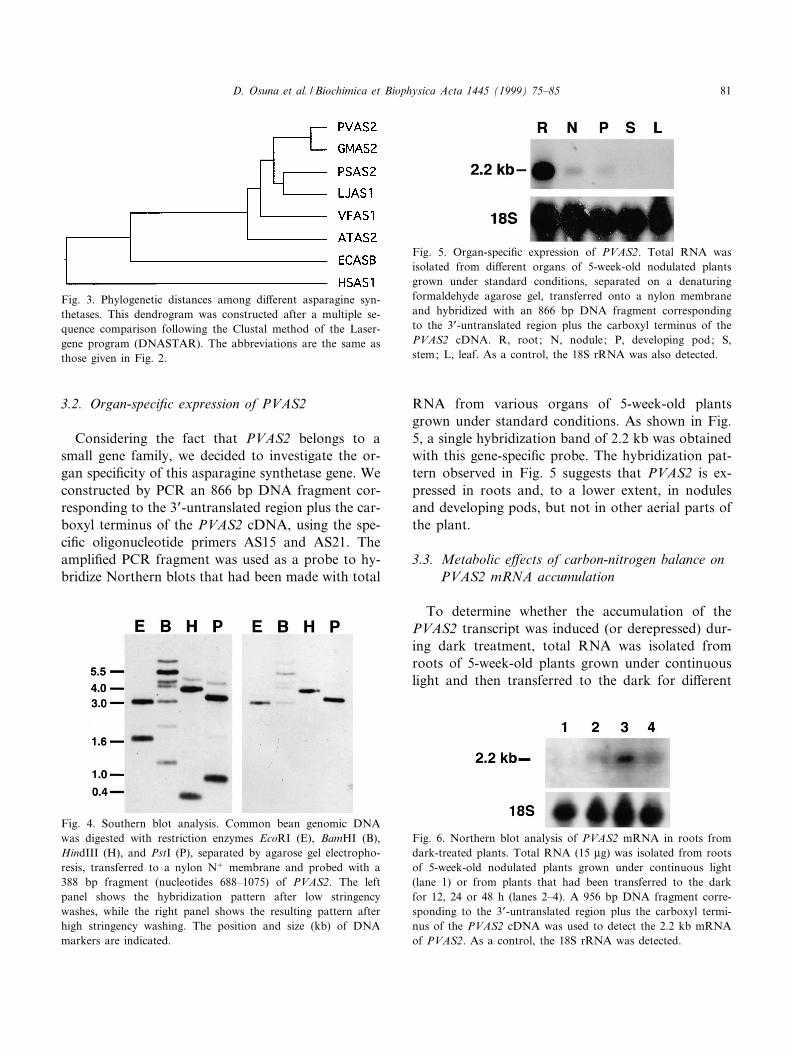
3.2. Organ-speci¢c expression of PVAS2
Considering the fact that PVAS2 belongs to asmall gene family, we decided to investigate the or-gan speci¢city of this asparagine synthetase gene. Weconstructed by PCR an 866 bp DNA fragment cor-responding to the 3P-untranslated region plus the car-boxyl terminus of the PVAS2 cDNA, using the spe-ci¢c oligonucleotide primers AS15 and AS21. Theampli¢ed PCR fragment was used as a probe to hy-bridize Northern blots that had been made with total
RNA from various organs of 5-week-old plantsgrown under standard conditions. As shown in Fig.5, a single hybridization band of 2.2 kb was obtainedwith this gene-speci¢c probe. The hybridization pat-tern observed in Fig. 5 suggests that PVAS2 is ex-pressed in roots and, to a lower extent, in nodulesand developing pods, but not in other aerial parts ofthe plant.
3.3. Metabolic e¡ects of carbon-nitrogen balance onPVAS2 mRNA accumulation
To determine whether the accumulation of thePVAS2 transcript was induced (or derepressed) dur-ing dark treatment, total RNA was isolated fromroots of 5-week-old plants grown under continuouslight and then transferred to the dark for di¡erent
Fig. 6. Northern blot analysis of PVAS2 mRNA in roots fromdark-treated plants. Total RNA (15 Wg) was isolated from rootsof 5-week-old nodulated plants grown under continuous light(lane 1) or from plants that had been transferred to the darkfor 12, 24 or 48 h (lanes 2^4). A 956 bp DNA fragment corre-sponding to the 3P-untranslated region plus the carboxyl termi-nus of the PVAS2 cDNA was used to detect the 2.2 kb mRNAof PVAS2. As a control, the 18S rRNA was detected.
Fig. 5. Organ-speci¢c expression of PVAS2. Total RNA wasisolated from di¡erent organs of 5-week-old nodulated plantsgrown under standard conditions, separated on a denaturingformaldehyde agarose gel, transferred onto a nylon membraneand hybridized with an 866 bp DNA fragment correspondingto the 3P-untranslated region plus the carboxyl terminus of thePVAS2 cDNA. R, root; N, nodule; P, developing pod; S,stem; L, leaf. As a control, the 18S rRNA was also detected.
Fig. 4. Southern blot analysis. Common bean genomic DNAwas digested with restriction enzymes EcoRI (E), BamHI (B),HindIII (H), and PstI (P), separated by agarose gel electropho-resis, transferred to a nylon N� membrane and probed with a388 bp fragment (nucleotides 688^1075) of PVAS2. The leftpanel shows the hybridization pattern after low stringencywashes, while the right panel shows the resulting pattern afterhigh stringency washing. The position and size (kb) of DNAmarkers are indicated.
Fig. 3. Phylogenetic distances among di¡erent asparagine syn-thetases. This dendrogram was constructed after a multiple se-quence comparison following the Clustal method of the Laser-gene program (DNASTAR). The abbreviations are the same asthose given in Fig. 2.
BBAEXP 93250 2-4-99
D. Osuna et al. / Biochimica et Biophysica Acta 1445 (1999) 75^85 81

periods of time. As shown in Fig. 6, the PVAS2transcript was more abundant in dark-grown plantsthan in plants that had been grown under continuouslight, conditions at which the level of PVAS2 tran-script was almost undetectable. Maximal accumula-tion of PVAS2 transcript was observed after 24 h inthe dark.
The e¡ect of an exogenous supplied metabolizablesugar on the accumulation of PVAS2 mRNA wasalso analyzed. Fig. 7A shows that the level ofPVAS2 in the common bean roots decays dramati-cally 48 h after the addition of 10 mM sucrose to thewatering medium of nodulated plants. On the con-trary, the addition of an inorganic source of nitrogen(10 mM KNO3) to the watering medium signi¢cantlyinduced the accumulation of PVAS2 mRNA in theroots of nodulated plants after 48 h (Fig. 7B).
4. Discussion
In higher plants, synthesis from aspartic acid byAS is the most important source of asparagine, amajor compound involved in nitrogen mobilization.However, AS enzyme activity has not been charac-terized adequately because of the numerous problemsderived from the high instability of the enzyme invitro [12], and the presence of AS inhibitors andcontaminating asparaginase activity in crude extracts
[13,14]. Because of these di¤culties to study AS atthe biochemical level, we decided to undertake a mo-lecular approach to the characterization of this en-zyme in common bean.
Our RT-PCR strategy led us to the isolation of afull-length cDNA clone (pPVAS2) whose longestORF encodes a putative asparagine synthetase. Thepresence of an in-frame stop codon (TAA) 12 nu-cleotides upstream from the putative translation startindicates that pPVAS2 contains the whole codingregion of an AS gene (PVAS2). The molecular pa-rameters of the predicted protein are very similar tothose predicted for other plant asparagine synthe-tases [14,19,20,29,30].
In E. coli, there are two genes encoding two ASwith di¡erent evolutionary origin: ASNA encodes anammonia-dependent AS [39], while ASNB encodes aglutamine-dependent AS enzyme [32]. In yeast, theammonia- and glutamine-dependent AS are also en-coded by two di¡erent genes [40]. In plants and ani-mals, however, all the asparagine synthetases thathave been characterized so far at the biochemicalor molecular level seem to use glutamine preferen-tially [18,41]. Peptide sequence analysis is in agree-ment with the biochemical data, since it revealed thatall the cloned plant AS are homologous to E. coliASNB. In addition, all the glutamine-dependent as-paragine synthetases seem to belong to the Ntn classof amide transferases, sharing a distinctive ¢nger-print of invariant residues with glutamine phospho-ribosyl amidotransferase (EC 2.4.2.14), the ¢rst en-zyme of purine biosynthesis [28,42]. The residuesand motifs that proved to be essential for aspartateand ATP binding in E. coli ASNB [33,34,37] areconserved also in PVAS2 and related plant AS,which adds new evidence to support their proposedfunction.
The conservation in PVAS2 of all the importantresidues with a known function in E. coli ASNB,together with the high amino acid similarity to otherAS ureide grain legumes like soybean and pea (Fig.3), lets us conclude that PVAS2 really represents aGln-dependent type II asparagine synthetase.
Southern blot analysis of genomic DNA revealedthat the AS gene family in common bean consists ofat least two genes, as in soybean [20], pea [14], andArabidopsis [19]. We are currently trying to isolatecDNAs corresponding to the other AS genes. Our
Fig. 7. Northern blot analysis of PVAS2 mRNA in roots from(A) sucrose- or (B) nitrate-treated plants. (A) Total RNA (15Wg) was extracted from roots of 5-week-old nodulated plantsgrown under standard conditions (lane 1), or from roots ofplants treated with exogenous 10 mM sucrose for 48 h (lane 2).(B) Total RNA (15 Wg) was extracted from roots of 5-week-oldnodulated plants grown under standard conditions (lane 1), ortreated with 10 mM KNO3 for 48 h (lane 2). A 956 bp DNAfragment corresponding to the 3P-untranslated region plus thecarboxyl terminus of the PVAS2 cDNA was used to detect the2.2 kb mRNA of PVAS2. As a control, the 18S rRNA wasalso detected.
BBAEXP 93250 2-4-99
D. Osuna et al. / Biochimica et Biophysica Acta 1445 (1999) 75^8582

unsuccessful attempts suggest that other AS genesare expressed at a much lower level than PVAS2.
Northern blot analysis showed that the steady-state level of PVAS2 mRNA is high in root tissue,much lower in developing pods and nodules, andcomparatively undetectable in stems, £owers andleaves of nodulated common bean plants grownunder a normal long day light cycle (Fig. 5). Thisexpression pattern resembles the pattern found inother legumes, where the highest level of expressionof AS is found in root tissue. However, in soybean,another ureide legume, both AS (SAS1 and SAS2)are expressed at a similar level in stems when com-pared to roots [20]. Unfortunately, there are noavailable data on the steady-state level of ASmRNA in soybean nodules or developing pods,though AS activity was detected and partially puri-¢ed from soybean root nodules [12]. The organ ex-pression pattern of PVAS2 in common bean is con-sistent with the putative physiological role ofasparagine synthetase, which is responsible for thesynthesis of a storage/transport amino acid. In devel-oping pods, nitrogen is reallocated under the form ofasparagine from the hull (ovary wall) into the matur-ing seeds where storage proteins are synthesized [43].It is intriguing, though, to detect the expression ofAS in the nodules of a ureide legume, which exportsthe ¢xed nitrogen mainly under the chemical form ofa ureide (allantoin or allantoate) [44]. The presenceof AS in the ¢xing nodules of a ureide plant addsnew evidence to the idea of a mixed character ofureide and amide plants. This also means that therelative abundance of ureides and amides in the xy-lem sap is in£uenced by nutritional and environmen-tal factors, besides being genetically determined.
Accumulation of PVAS2 mRNA in common beanroots seems to be repressed by light. A signi¢cantincrease in the steady-state level of PVAS2 mRNAwas observed as early as 6 h after transferring plantsto darkness (Fig. 6). Direct regulation of AS geneexpression by light was previously reported in peaand soybean aerial organs, and in Arabidopsis seed-lings, where phytochrome was shown to be mediatingthe repression process [19]. Asparagine synthetase isone of the few genes whose transcription is nega-tively regulated by light. The short list includes phy-tochrome [45], photochlorophyllide reductase [46],HMG CoA reductase [47], the putative transcription
factor Athb-2 [48] and GDH [49]. Light repression ofPVAS2 mRNA expression in roots, however, is un-likely to be a direct regulatory process. In addition tophytochrome-mediated e¡ects, light has been shownto exert indirect e¡ects on AS gene expressionthrough associated changes in carbon metabolites.Under light conditions, nitrogen is assimilated intometabolically active glutamine and glutamatethrough the GS-GOGAT cycle, and transported assuch for their use in anabolic reactions [49]. Uponthe transition to darkness, photosynthesis abortionhas a dramatic e¡ect on the availability of carbonthat extends throughout the whole plant. When car-bon skeletons become limiting, the storage of nitro-gen as asparagine via AS is favored on the basis ofthe higher content of N per molecule and the highersolubility and chemical stability of asparagine com-pared to glutamine [44]. In asparagus, AS mRNAlevels increase in harvested spears, a light-independ-ent process that runs in parallel to a decrease ofsugar content [17].
Concomitantly, under carbon-su¤cient conditions,as when plants were supplied with a reduced carbonsource, the steady-state level of PVAS2 mRNA de-cayed in common bean nodulated roots (Fig. 7A).During the last years there has been some discussionabout the possible interrelation between light controlof gene expression and metabolic regulation bysugars [50]. In fact, sucrose is known to simulatethe e¡ect of light on the expression of other genesof nitrogen metabolism like nitrate reductase (NR),glutamine synthetase (GS2), glutamate synthase(GOGAT) and glutamate dehydrogenase (GDH)[49,51,52]. The fact that nitrogen assimilation is regu-lated by the carbon status implies the existence of aclose interrelation between carbon and nitrogen me-tabolism. After several studies of the metabolic con-trol by sugars on genes involved in photosynthesisand carbon metabolism [50,53,54], the enzyme hex-ose kinase was proposed as the sensor of carbonavailability [50]. Plant homologues of the yeast cat-abolic repression trans-acting factor SNF1 have beenidenti¢ed in rye, Arabidopsis and barley [55^57].However, the possible function of SNF1 in the con-trol of C:N balance remains to be determined.
Addition of an inorganic source of nitrogen(KNO3) to the medium signi¢cantly induced the ac-cumulation of PVAS2 mRNA in the roots of nodu-
BBAEXP 93250 2-4-99
D. Osuna et al. / Biochimica et Biophysica Acta 1445 (1999) 75^85 83

lated plants after 48 h (Fig. 7B). Fertilization with aninorganic source of nitrogen is known to inhibit N2
¢xation in root nodules. Under these conditions, theconcentration of ureide in xylem parallels the declineof N2 ¢xation in ureide legumes including the com-mon bean [58]. In a ureide legume like soybean, ad-dition of inorganic nitrogen caused a 75% decrease inthe activity of PRAT, the enzyme that catalyzes thecommitted step in the biosynthesis of purines, theprecursors of ureides [59]. It is an extended observa-tion that nitrate-fed ureide legumes experience a shiftfrom ureide synthesis to amide synthesis and export[4].
Nitrate is the primary regulatory signal of nitrateassimilation, and plants respond to nitrate with aninduction of the expression of many genes includingthose encoding NR, GS and GOGAT. Though thestudy of nitrate regulation led to the identi¢cation ofseveral regulatory genes that are responsible for ni-trate induction, none of them have been found inplants [60]. So far no evidence has been found tosupport direct regulation of AS expression by nitrate.
Metabolic regulation of AS expression by C:Nbalance was previously shown in maize root tips[18], Arabidopsis seedlings [19] and harvested aspar-agus spears [17]. C:N balance is actually a key issuein the study of the joint metabolisms of carbon andnitrogen, and also in the study of the symbiotic sys-tem Rhizobium/legume. In di¡erent metabolic con-texts where carbon and nitrogen metabolisms mustbe integrated, glutamine has been proposed as a key-stone metabolic signal. In pea, the glutamine synthe-sized by GS was suggested to act as an inducer ofasparagine synthetase gene expression in cotyledonsof germinating seedlings and in nodules [14].
In our continuing work we are trying to isolate therest of the AS genes in common bean, and analyzetheir regulation particularly in the metabolic contextof an N2-¢xing nodule. This will help elucidate themolecular mechanisms responsible for the coordinateregulation of ureide and asparagine metabolism dur-ing nitrogen ¢xation and mobilization from the nod-ule to the aerial parts of the plant.
Acknowledgements
This work was supported by Direccion General de
Ensen¬anza Superior (Grant PB96-0504-C02-02) andby Plan Andaluz de Investigacion (CV-0115). D.O.thanks the Ministerio de Educacion y Cultura(Spain) for a fellowship award.
References
[1] J.S. Pate, C.A. Atkins, S.T. White, R.M. Rainbird, K.C.Woo, Plant Physiol. 65 (1980) 961^965.
[2] T.G. Patterson, T.A. LaRue, Crop Sci. 23 (1983) 819^824.[3] T.C. Ta, F.D.H. MacDowall, M.A. Faris, Biochem. Cell
Biol. 67 (1989) 455^459.[4] K.R. Schubert, Annu. Rev. Plant Physiol. 37 (1986) 539^
574.[5] A.A. Urquhart, K.W. Joy, Plant Physiol. 68 (1981) 750^754.[6] M.B. Peoples, M.N. Sudin, D.F. Herridge, J. Exp. Bot. 38
(1987) 567^579.[7] B. Horowitz, A. Meister, J. Biol. Chem. 247 (1972) 6708^
6719.[8] R. Hubert, R.D. Simoni, J. Bacteriol. 142 (1980) 212^220.[9] S.E. Rognes, FEBS Lett. 14 (1975) 1975^1982.
[10] H. Cedar, J.H. Schwartz, J. Biol. Chem. 244 (1969) 4112^4121.
[11] P.J. Lea, S.A. Robinson, G.R. Stewart, in: B.J. Mi£in, P.J.Lea (Eds.), The Biochemistry of Plants, vol. 16, AcademicPress, San Diego, CA, 1991, pp. 121^159.
[12] T.A. Huber, J.G. Streeter, Plant Sci. 42 (1985) 9^17.[13] K.W. Joy, R.J. Ireland, P.J. Lea, Plant Physiol. 73 (1983)
165^168.[14] F.Y. Tsai, G.M. Coruzzi, EMBO J. 9 (1990) 323^332.[15] P.J. Lea, L. Fowden, Proc. R. Soc. London B 192 (1975) 13^
26.[16] F.Y. Tsai, G.M. Coruzzi, Mol. Cell. Biol. 11 (1991) 4966^
4972.[17] K.M. Davies, G.A. King, Plant Physiol. 102 (1993) 1337^
1340.[18] C. Chevalier, E. Bourgeois, D. Just, P. Raymond, Plant J. 9
(1996) 1^11.[19] H.M. Lam, S.S. Peng, G.M. Coruzzi, Plant Physiol. 106
(1994) 1347^1357.[20] C.A. Hughes, S.B. Hunter, B.F. Mathews, Plant Mol. Biol.
33 (1997) 301^311.[21] D. Hannahan, J. Mol. Biol. 166 (1983) 557^580.[22] T. Maniatis, E.F. Fritsch, J. Sambrook, Molecular Cloning:
a Laboratory Manual, 2nd edn. Cold Spring Harbor Labo-ratory Press, Cold Spring Harbor, NY, 1989.
[23] C. Connell, S. Fung, C. Heiner, J. Bridgham, V. Chakerian,E. Heron, B. Jones, S. Menchen, W. Mordan, M. Ra¡, M.Recknor, L. Smith, J. Springer, S. Woo, M. Hunkapillar,Biotechniques 5 (1987) 342^348.
[24] S.F. Altschul, W. Gish, E.W. Myers, D.J. Lipman, J. Mol.Biol. 215 (1990) 403^410.
[25] M. Kimura, The Neutral Theory of Molecular Evolution,Cambridge University Press, Cambridge, 1983.
BBAEXP 93250 2-4-99
D. Osuna et al. / Biochimica et Biophysica Acta 1445 (1999) 75^8584

[26] G.M. Church, W. Gilbert, Proc. Natl. Acad. Sci. USA 81(1984) 1991^1995.
[27] C.P. Joshi, Nucleic Acids Res. 15 (1987) 9624^9640.[28] H. Zalkin, J.L. Smith, Adv. Enzymol. Relat. Areas Mol.
Biol. 72 (1998) 87^144.[29] R.N. Waterhouse, A.J. Smyth, A. Massonneau, I.M. Pross-
er, D.T. Clarkson, Plant Mol. Biol. 30, (5) (1996) 883^897.[30] H. Kuester, U. Albus, M. Fruehling, S.A. Tchetkova, I.A.
Tikhonovitch, A. Puehler, A.M. Perlick, Plant Sci. 124(1997) 89^95.
[31] I.L. Andrulis, J. Chen, P.N. Ray, Mol. Cell. Biol. 7 (1987)2435^2443.
[32] M.A. Sco¢eld, W.S. Lewis, S.M. Schuster, J. Biol. Chem.265 (1990) 12895^12902.
[33] S.K. Boehlein, E.S. Walworth, N.G.J. Richards, S.M. Schus-ter, J. Biol. Chem. 272 (1997) 12384^12392.
[34] S.K. Boehlein, E.S. Walworth, S.M. Schuster, Biochemistry36 (1997) 10168^10177.
[35] P. Ma«ntsa«la, H. Zalkin, J. Bacteriol. 174 (1992) 1883^1990.
[36] T.S. Leyh, T.F. Vogt, Y. Suo, J. Biol. Chem. 267 (1992)10405^10410.
[37] N.G.J. Richards, Adv. Enzymol. Relat. Areas Mol. Biol. 72(1998) 145^198.
[38] P. Bork, E.V. Koonin, Proteins Struct. Funct. Genet. 20(1994) 347^355.
[39] M. Nakamura, M. Yamada, Y. Hirota, K. Sugimoto, A.Oka, M. Takanami, Nucleic Acids Res. 9 (1981) 4669^4676.
[40] F. Ramos, J.M. Wiame, Eur. J. Biochem. 108 (1980) 373^378.
[41] K.A. Sieciechowicz, K.W. Joy, R.J. Ireland, Phytochemistry27 (1988) 663^671.
[42] J.Y. Tso, M.A. Hermdsen, H. Zalkin, J. Biol. Chem. 257(1982) 3532^3536.
[43] C. Rochat, J.P. Boutin, Plant Physiol. Biochem. 27 (1989)881^887.
[44] P.J. Lea, B.J. Mi£in, in: B.J. Mi£in, P.J. Lea (Eds.), TheBiochemistry of Plants, vol. 16, Academic Press, San Diego,CA, 1991, pp. 121^159.
[45] W.B. Bruce, A.H. Christensen, T. Klein, M. Fromm, P.H.Quail, Proc. Natl. Acad. Sci. USA 86 (1989) 9692^9696.
[46] E. Mosinger, A. Batschauer, E. Schafer, K. Apel, Eur. J.Biochem. 147 (1985) 137^142.
[47] R.M. Learned, Plant Physiol. 110 (1996) 645^655.[48] M. Carabelli, G. Morelli, G. Whitelam, I. Ruberti, Proc.
Natl. Acad. Sci. USA 93 (1996) 3530^3535.[49] H.M. Lam, K.T. Coshigano, I.C. Oliveira, R. Melo-Oliveira,
G. Coruzzi, Annu. Rev. Plant Physiol. Plant Mol. Biol. 47(1996) 569^593.
[50] J.C. Jang, J. Sheen, Plant Cell 6 (1994) 1665^1679.[51] N.M. Crawford, H.N.J. Arst, Annu. Rev. Genet. 27 (1993)
115^146.[52] C.L. Cheng, G.N. Acedo, M. Cristinsin, M.A. Conklin,
Proc. Natl. Acad. Sci. USA 89 (1992) 1861^1864.[53] J.Y. Sheen, Plant Cell 2 (1990) 1027^1038.[54] J.Y. Sheen, Photosynth. Res. 39 (1994) 427^438.[55] A. Alderson, P.A. Sabelli, J.R. Dickinson, D. Cole, M. Ri-
chardson, Proc. Natl. Acad. Aci. USA 88 (1991) 8602^8605.[56] L. Le Guen, M. Thomas, M. Bianchi, N.G. Halford, M.
Kreis, Gene 120 (1992) 249^254.[57] U. Hannapel, J. Vicente-Carbajosa, J.H.A. Barker, P.R.
Shewry, N.J. Halford, Plant Mol. Biol. 27 (1995) 1235^1240.[58] C.F. Rice, M. Krystina, K.M. Lukasewski, S. Walker, D.G.
Blevins, R.G. Winkler, D.D. Randall, J. Plant Nutr. 13(1990) 1539^1553.
[59] P.H.S. Reynolds, D.G. Blevins, D.D. Randall, Arch. Bio-chem. Biophys. 229 (1982) 623^631.
[60] N.M. Crawford, Plant Cell 7 (1995) 859^868.
BBAEXP 93250 2-4-99
D. Osuna et al. / Biochimica et Biophysica Acta 1445 (1999) 75^85 85




















