
Do phytoliths play an antiherbivory role in southwest Asian Asteraceae species and to what extent?

Root and tuber foods are typically underrep-resented or absent in assemblages of charred ar-chaeobotanical macroremains for a variety ofreasons. Consumption may leave no remainsbut thin peels or inedible fibers. Roots or tuberscharred during roasting readily break up in ar-chaeological deposits and can be difficult toidentify in fragmented form. Pollen and phy-toliths (microscopic siliceous remains of plants)produced by leaf and flowering tissues provideonly indirect evidence of root or tuber utiliza-tion, and the likelihood of their deposition is re-duced because root and tuber crops are oftenpropagated vegetatively, flower infrequently,and contain lower concentrations of silica.
By contrast, starch grains extracted fromstone pounding tools are good direct sources ofevidence for root and tuber use (Barton et al.1998; Fullagar et al. 1996; Perry 2001; Pipernoand Holst 1998; Piperno et al. 2000; Piperno etal. 2004), and current research indicates thatstarch is more likely to survive on artifact sur-faces, though grains of different sizes may havedifferent survivorship archaeologically (Halsam2004). Starch preservation in soils and sedi-ments is still under investigation (Halsam 2004;Lentfer et al. 2002; Korstanje 2003; Lu 2003).Ideally, additional indicators should be recov-ered to provide more complete evidence of rootand tuber consumption. Fortunately, silica-based phytoliths survive in archaeological sam-ples where other organic remains decompose.
While analyzing archaeological tools from
Root and Tuber Phytoliths and Starch GrainsDocument Manioc (Manihot esculenta), Arrowroot(Maranta arundinacea), and Llerén (Calathea sp.) atthe Real Alto Site, Ecuador1
Karol Chandler-Ezell, Deborah M. Pearsall, and James A. Zeidler
Chandler-Ezell, Karol (Department of Sociology, 351 Liberal Arts, Stephen F. Austin StateUniversity, Nacogdoches, TX 75962), Deborah M. Pearsall (Department of Anthropology,107 Swallow Hall, University of Missouri, Columbia, MO 65211; e-mail: [email protected]), and James A. Zeidler (CEMML, Colorado State University, Ft. Collins, CO80523). Root and Tuber Phytoliths and Starch Grains Document Manioc (Manihotesculenta), Arrowroot (Maranta arundinacea), and Llerén (Calathea sp.) at theReal Alto Site, Ecuador. Economic Botany 60(2):103–120, 2006. Although roots and tu-bers are dietary staples in many parts of the world, their use is difficult to document archaeo-logically because their organic remains are often poorly preserved in archaeological sedi-ments. Here we describe the first diagnostic phytoliths from the underground storage organsof the important New World agricultural crops manioc or yuca (Manihot esculenta Crantz),arrowroot (Maranta arundinacea L.), and llerén (Calathea allouia [Aubl.] Lindl.) and demon-strate their usefulness for identifying prehistoric root and tuber processing with a study ofstone artifacts from a Valdivia 3 (2800–2400 B.C., calibrated) household at Real Alto,Ecuador. Gelatinized starch (heat-altered) and unaltered starch from maize (Zea mays L.),arrowroot, and manioc were also found on these stone tools. Our data document early evi-dence for manioc in Ecuador’s coastal lowlands. In combination, these phytoliths and starchresidues provide evidence that both raw and cooked foods were processed in this early mixedagricultural economy.
Key Words: Phytoliths, starch, Real Alto site, Valdivia culture, manioc, prehistoric agricul-ture.
Economic Botany, 60(2), 2006, pp. 103–120.© 2006, by The New York Botanical Garden Press, Bronx, NY 10458-5126 U.S.A.
11. Received 17 February 2006; accepted 1 March2006.

Structure 20 at the Real Alto site in Ecuador toestablish how commonly maize was used dur-ing the Middle Valdivia period (Pearsall et al.2004), we observed phytoliths that were similarto, but not quite typological matches for, theleaf and inflorescence bodies of several eco-nomically important plants, including arrowrootand llerén. Because the leaves and fruits ofthese plants are not usually eaten, these findingsdid not make sense. We reasoned that these sil-ica bodies could have arisen from similaranatomical structures in other tissues of theplants, in particular, the edible roots, tubers,and rhizomes.
Most studies of silica deposition mechanisms(Piperno 1988) and phytolith types and occur-rence (Pearsall 2000) focus on aerial portions ofplants; to date, no one has successfully de-scribed indicator types from the undergroundtissues that were useful for archaeological ap-plications. We hypothesized that useful phy-toliths from subterranean tissues could be iden-tified by focusing on the epidermis, processinglarger sample sizes to compensate for lower sil-ica concentrations, using gentler and adaptiveprocessing methods to cope with the roots’different chemical compositions, and identify-ing new suites of diagnostic types.
In addition to the unknown phytoliths, wealso observed what appeared to be damagedstarch grains and other unknown organicresidues on the Structure 20 tools (Pearsall etal. 2004). We designed a series of processingexperiments building upon Babot’s (2003)starch alteration research that used replicatestone tools and different methods of cookingmaize and manioc to determine how theseresidues were created, and if possible, to iden-tify them (see below, starch experiment). Wepresent here new tissue- or organ-specific diag-nostic phytolith types discovered during this re-search and apply the new identification proto-cols to the reanalysis of the Structure 20 stonetool residues. In combination, the starchresidues and phytolith results provide new in-sight into prehistoric subsistence and food pro-cessing at this early village site.
MethodsComparative Plant Tissue SamplingComparative samples came from the MU lab
collection, market samples, and the Missouri
104 ECONOMIC BOTANY [VOL. 60
Botanical Garden Herbarium. Where possible,we dissected samples into separate tissues toidentify phytoliths’ anatomical origin and tookparticular care separating root and tuber epider-mis (Table 1, Table 2). We dissected fresh sam-ples first, then dried them as separate tissuesamples. We increased sample sizes from thestandard 0.1g to increase silica yield, some-times including all of a fruit, seed, or peel. Anote explaining our use of phytolith nomencla-ture is found in Appendix 1.
Phytolith Extraction and AnalysisPhytoliths were isolated from the modern
comparative plant tissues using standard MUlab protocols (Pearsall 2000) for sample pre-treatment and chemical oxidation with Schulzesolution (3:1 ratio of nitric acid to potassiumchlorate) with a few modifications: sonicationand oven-drying were omitted from pretreat-ment to avoid physical damage to more delicatesilicified tissues; 50mL centrifuge tubes and500mL beakers were used to accommodatelarger samples; and tough, hard tissues werestarted in beakers on hot plates instead of in thewaterbath. Samples that were resistant to theSchulze solution were rinsed and switched to a27% hydrogen peroxide solution (Pearsall et al.2004) for further oxidation. In a few samples,an oily, waxy residue formed which was resist-ant to both oxidizers. These samples wererinsed and returned to the hot water bath withan acetonitrile (Methyl cyanide) treatment todissolve the complex organic compounds (plantwaxes and oils); then the dissolved organicssuspended in the acetonitrile were removed bydecanting off the supernatant. After evaporatingresidual acetonitrile, processing was completedby rinsing all samples with hot (>80 degrees C)distilled water and drying them in a low tem-perature oven.
Extracts were weighed, stored, and mounted(~0.001g/slide, in immersion oil) (Pearsall2000). Each slide was scanned in its entiretyusing a Nikon light microscope at 400x, and thecontents recorded using an abundance code sys-tem.
Comparative Phytolith StudyA total of 62 comparative plant taxa were
sampled, with successful silica extraction from58, and identification of diagnostic phytoliths in45 (Table 1, Table 2). A major aim of the study

2006] CHANDLER-EZELL, ET AL.: ROOT AND TUBER PHYTOLITHS FROM REAL ALTO 105
Table 1. Diagnostic phytoliths observed in comparative taxa and tissues studied.1
Plant Family Name MU Lab Number Tissues Sampled Types Observed
Anacardiaceae Spondias purpurea 2624 F 140I, 140IIBAnnonaceae Annona squamosa 2630 F 80IBAraceae *Colocasia esculenta 2567/2564 TEP/TF 23, 110/23, 100ICa, 130I,
130IIA, 140IIAAraceae Xanthosoma 2633 T 130IIABixaceae Bixa orelliana 2637 SD 80IIBb, 150I, 150IICannaceae Canna crotalifera 2704 R 80IB, 130ICannaceae Canna edulis 2591/2639 L/TEP 80IB, 80IE/80IECannaceae Canna glauca 2707 R 110Cannaceae Canna paniculata 2705 R 80IE, 140IIAChrysobalanaceae Chrysobalanus icaco 2628 F 80IAa1, 80IIBb, 140I, 150IIConvolvulaceae Ipomoea batatas 2555/2553/2554 TEP/TT/TEY N/130I/130IDioscoreaceae Dioscorea 2577/2576 TTEP/TC 26IIA, 100ICa, 140IIB/110,
130IDioscoreaceae Dioscorea 2574/2578 TEP/TC 130I, 140IIB/ 130I, 130IIA,
140IIAEbenaceae Diospyros ebenaster 2641 SD 23, 80IIBb, 110Euphorbiaceae Manihot brachyloba 2713 ST 110, 130IEuphorbiaceae Manihot esculenta N/2539/2552 RC/RC/RC 80IIBb/130I/80IIBb, 130I,
130IIAEuphorbiaceae Manihot esculenta 254/635/637 160I/160I/160IEuphorbiaceae Manihot esculenta 2541/2540 RR/RR 24IC, 160I/110, 130I, 160IEuphorbiaceae Manihot esculenta 2547/2548 RRI/RRE 110, 160I/110, 130I, 160IEuphorbiaceae Manihot esculenta 2532/67/ 69 L/L/L 40IVF/110, 160I/160IEuphorbiaceae Manihot esculenta 1117/2531/2708 L, ST/ST/ST N/40IVB, 110, 160I/130I,
160I, 110Euphorbiaceae Manihot esculenta 2534/2533 F/F 40IVF/110, 160IEuphorbiaceae Manihot esculenta 2538/2542 REP/R 160I/130I, 140IIBEuphorbiaceae Manihot hunzikerii 2711/2718 R/R 160I/130IEuphorbiaceae Manihot leptophylla 2712 ST 140IEuphorbiaceae Manihot cf. tristis 2716 R 130IFabaceae Acacia macrocantha 2660 SD 150I, 150IIFabaceae Arachis hypogaea 2669 P 150IIFabaceae Canavalia 2655 SD 150IFabaceae Inga spectabilis 2654/2653 P/SD N/80IB, 80IIBb, 130IIB, 140I
140IIBFabaceae Pachyrhizus erosus 2569/2572 STT/RRE 130I, 130IIA/130I, 140IIAFabaceae Phaseolus vulgaris 2635 SD 150IMarantaceae Calathea allouia 2672 T 26IAa, 26IAb, 26IB, 26IIBMarantaceae Calathea allouia 2672 T 80ICa, 80IE, 80IIIB, 110Marantaceae Calathea inocephala 2720 ST 26IB, 80IB, 80ICa, 80IE,
80IIIBMarantaceae Calathea inocephala 2721/2722 R/R 80IB, 80ICa, 80IE/80IB,
80ICa, 110, 130IMarantaceae Calathea macrosepalia 2723/2724 R/RZ 80ICa, 140IIA/80ICaMarantaceae Calathea marantifolia 2725 ST 26IIB, 80ICa, 80IEMarantaceae Ctenanthe amphiandina 2726 R 130IMarantaceae Ischnosiphon leucophaeus 2728 R 80ICaMarantaceae Maranta amazonica 2729/2730 R/RZ 80IB, 110/80IE, 80IIIB, 110,
130I

was to investigate and identify new types anddistributions of phytolith production in eco-nomically important edible plant tissues, so weincluded domesticated lowland root and tubercrops as well as cultivated, domesticated, andwild fruit trees, sampling specific tissues fromthese plants’ edible portions. Taxa selected rep-resent food resources that may have been avail-able to the Real Alto population 4,400–4,800years ago.
We tested the domesticated species manioc(Manihot esculenta), arrowroot (Maranta arun-dinacea), and llerén (Calathea allouia) as wellas a selection of wild taxa so we could deter-mine the taxonomic level of any new diagnos-tics (i.e., whether a diagnostic phytolith identi-fied a plant to species, genus, family, or order).We tested a selection of wild Manihot,Maranta, and Calathea species to determinewhether diagnostics found in the domesticatesalso occurred in wild species in the same genus,
106 ECONOMIC BOTANY [VOL. 60
and we examined all New World genera andfamilies in the Zingiberales (the order that in-cludes the Marantaceae) to be sure that therewere no phytolith “confusers” in this group ofclosely-related plants. We had already con-ducted a broad overview of the Euphorbiaceae(family that includes Manihot) prior to thisproject. Wild species that occur on the coast ofEcuador were selected, with the exception ofManihot, which does not occur on the coast(Jørgensen and León-Yánez 1999).
A variety of diagnostic phytoliths were ob-served in root, tuber, and inflorescence tissues(Table 1). Some are new types; others aretypes previously defined at MU or by Piperno(1988, 1989) found to occur in new tissues.Only selected types can be described and illus-trated here (Fig. 1); see Appendix 2 for de-scriptions of other types and the MU Phytolithwebsite (http://www.missouri.edu/~phyto) forphotographs of types. Numbers follow the MU
Table 1. Continued
Plant Family Name MU Lab Number Tissues Sampled Types Observed
Marantaceae Maranta arundinacea 2673 R, RH 26IB, 80IB, 80ICa, 80IE,80IIIB
Marantaceae Maranta arundinacea 2674 RZ 26IB, 80IB, 80ICa, 80IE,80IIIB
Marantaceae Maranta gibba 2732/2733 STT/R 80IB, 80ICd, 80IIIB/80ICa,110
Marantaceae Maranta sobilifera 2736/2737 STT/R 80IB, 80ICa, 80IIIB, 110/80I110
Marantaceae Stromanthe jacquinii 2738 R 80ICa, 80IIIBMarantaceae Thalia geniculata 2739/2740 STT/R 80IB, 80ICa, 80IIIB/80IE,
80IIIBMusaceae Heliconia latispatha 2741 R 80IB, 80IIAb, 80IIAc, 110,
130IIA, 140I, 140IIAMusaceae Heliconia latispatha 2742 RZ 80IIAb, 80IIAcMyrtaceae Psidium guajava 2636 SD 110Solanaceae Capsium 2646 SD 130IVerbenaceae Vitex gigantea 2627 F 80IB, 140IZingiberaceae Costus guanaiensis var.
guanaiensis2746 RZ 80IE
Zingiberaceae Costus scaber 2744 R 80IB, 80ICa, 80IEZingiberaceae Renealmia alpinia 2748 RZ 140IIAZingiberaceae Renealmia cernua 2745 REP 140I
*Not a New World native1 See MU Phytolith website (http://www.missouri.edu/�phyto) for a listing of all phytoliths observed in samples and
tissues. Table key: Codes for tissues sampled: F fruit, FR fruit rind, L leaf, P pod, R root, RC root cortex, REP rootepidermis, RH root hairs, RR root rind, RRE root rind exterior, RRI root rind interior, RZ rhizome, SD seed, ST stem,STT stem tip, T tuber, TC tuber cortex, TEP tuber epidermis, TEY tuber eyes, TF tuber fibers, TT tuber tips, TTEP tubertip epidermis. N�none.

phytolith classification system, which is basedupon phytolith origin in tissue (Pearsall andDinan 1992). The taxonomic level of the diag-nostic is indicated in the type name. The fol-lowing phytoliths are helpful for identifyingthe presence of root, tuber, or rhizome tissuefrom Manihot, Calathea, and Calathea/Maranta:
26IAa Calathea (Marantaceae) irregular rhi-zome cylinder: Small cylindrical bodies withundulating, beaded, nodular, or spiked surfacedecoration. Abruptly narrowed tip or head. Wasnot observed in other genera.
26IAb Calathea (Marantaceae) flat domedrhizome cylinder: Cylindrical body with ciliateor beaded decoration along cylinder and a dis-tinctive, smooth head. Overall shape a cylinderthat is polygonal in cross section. Body size issmaller than related Marantaceae inflorescencetypes. Was not observed in other genera.
2006] CHANDLER-EZELL, ET AL.: ROOT AND TUBER PHYTOLITHS FROM REAL ALTO 107
26IB Maranta/Calathea (Marantaceae) rhi-zome spindle: Cylindrical body that tapersgradually to a pointed tip (lacks distinctive headarea or distinctly separate cylinder and headareas). Nodular surface decoration. Body isround in cross section. Not observed in othergenera.
26IIB Calathea (Marantaceae) flat rhizometype: Flattened, somewhat elongated body witha decorated, beaded edge, and an elongate, ir-regularly stellate center. Center appears some-what elevated above the ends (platform appear-ance). Was observed in C. allouia and C.marantifolia.
160I Manihot (Euphorbiaceae) secretorybody: Manihot esculenta (yuca, manioc) rootrind, leaf, stem, and fruit silicify distinctivecells resembling pores or nectaries. These bod-ies also occur rarely in one wild species tested(M. hunzikerii). Bodies are small (5–12 microns
Table 2. Plant specimens and tissues studied that did not produce diagnostic phytoliths.
I. Silica present, but no diagnostic phytoliths produced:Anacardiaceae, Anacardium occidentale, blossom, fruit husk*Araceae, Colocasia esculenta, stem end of tubers, roots, cor-
texCannaceae, Canna indica, stem, seedClusiaceae, Mammea americana, fruitDioscoreaceae, Dioscorea, tuber cortex, tip epidermisEuphorbiaceae
Manihot esculenta, seed; stem end of root; fibers fromstem tip, central root, and root bottom tip; exterior cor-tex adjacent to root rind
M. brachyloba, stem, rootFabaceae
Acacia macracantha, seedArachis hypogaea, seedPachyrhizus erosus, interior fibrous epidermisPhaseolus lunatus, podProsopis juliflora, seed
Malpighiaceae, Bunchosia armeniaca, fruitMalvaceae, Gossypium barbadense, fruit, seedMarantaceae
Calathea allouia., inflorescence, leafIschnosiphon leucophacus, stem baseMaranta orbiculata root, stem base
Passifloraceae, Passiflora, seedRutaceae, Casimiroa edulis, seedSapotaceae, Mastichodendron foetidissimum, seedSolanaceae, Nicotiana rustica, seed, fruitSterculiaceae, Theobroma cacao, fruit*Not a New World native; studied to increase Araceae
sample.
II. Silica absent:Anacardiaceae, Anacardium occidentale,
fruit fleshBixaceae, Bixa orellana, fruitConvolvulaceae, Ipomoea batatas, vascu-
lar tissue, tuber tip flesh, cortex, coreof tuber
Elaeocarpaceae, Muntingia calabura, fruitEuphorbiaceae, Manihot esculenta, cortex
from stem tip, root tip, root epider-mis fibers
FabaceaeAcacia macracantha, podCassia, pod, seedPachyrrhizus erosus, root tip, cortexPachyrrhizus ahipa, root rindPhaseolus lunatus, seedAlbizia multiflora, podProsopis juliflora, seed
Malvaceae, Gossypium barbadense, fiberSapotaceae, Manilkara zapota, fruit flesh,
fruit rindSolanaceae, Capsicum, fruit

108 ECONOMIC BOTANY [VOL. 60
Fig. 1. Selected new diagnostic phytoliths. See MU Phytolith website (http://www.missouri.edu/~phyto)for photographs of all types. a. 26IAa Calathea (Marantaceae) irregular rhizome cylinder. b, c. 26IAbCalathea (Marantaceae) flat domed rhizome cylinder. d. 26IB Maranta/Calathea (Marantaceae) rhizomespindle. e, f. 26IIB Calathea (Marantaceae) flat rhizome type. g, h. 160I Manihot (Euphorbiacae) secretorycell.

in diameter), heart-shaped, with a raised exte-rior and indented/open interior. A thin, flat mar-ginal flange is sometimes present.
Phytoliths observed in samples we studiedincrease our range of diagnostic tools for futureinvestigations by documenting the presence ofknown diagnostics in new tissues and discover-ing new types (Table 1). Three classes of diag-nostics resulted from the study: overlappingbodies, functional tissue-level bodies, and taxa-and tissue-specific bodies.
Overlapping bodies are those phytoliths thatoccur in underground tissues as well as leafand/or inflorescence: for example, 160I Mani-hot secretory body (root, leaf, fruit); 80Aa1Chrysobalanaceae small smooth sphere (leaf,fruit); 80IIAb,c Heliconia trough bodies (leaf,root, rhizome); 80ICa Marantaceae nodularsphere and 80IIIB Marantaceae conical body(leaf, rhizome); and 80IE Zingiberales irregu-larly angled/folded sphere. Overlapping bodiescan provide supporting archaeological evidencefor the presence of the underground portion, in-stead of only leaves or inflorescences of maniocand the Marantaceae.
Functional tissue-level bodies are those phy-toliths indicative of “rootiness” or “fruitiness”in general, such as 130I, blocky parenchyma,130IIA, multilobed parenchyma, 140IIA,straight transport tissue (roots, tubers), and140I, undulating transport elements (fruits,seeds, predominantly) (Table 1, Appendix 2).Functional tissue-level “rooty” bodies can beused archaeologically in combination withstarch grains and diagnostic phytoliths as sup-port for interpretations that other, overlappingbodies came from root or rhizome tissue.
Taxa- and tissue-specific phytoliths includetypes useful for documenting the economicallyimportant rhizome/tuber crops Maranta arundi-nacea and Calathea allouia (Marantaceae). Wefound one rhizome-specific type (26IB) pro-duced by Maranta and Calathea species, andthree types (26IAa, 26IAb, 26IIB) producedonly by Calathea. These phytoliths are morpho-logically similar to, but distinctive from, Ca-lathea and Maranta seed phytoliths first identi-fied by Piperno (1989). Presence of the newrhizome-specific phytoliths archaeologicallywould demonstrate use of specific tissues ofwild or domesticated plants in these genera. Incases in which wild taxa can be eliminated onecological or contextual grounds, these types
2006] CHANDLER-EZELL, ET AL.: ROOT AND TUBER PHYTOLITHS FROM REAL ALTO 109
can provide a line of evidence for use of do-mesticated species.
It should be noted that in our study of wildZingiberales taxa, we found no phytoliths thatcould be confused with the new Calathea orCalathea/Maranta rhizome types. Further, a re-view of all data sheets from the comprehensivePhytoliths in the Flora of Ecuador project(http://www.missouri.edu/~phyto) showed nooccurrence of 160I, Manihot secretory body, inany other genus, including genera in the Eu-phorbiaceae. Manioc, the most important rootcrop of the lowland Neotropics, has been elu-sive in the archaeological record. The Manihotsecretory body documented in our research pro-vides another avenue for its identification.
Starch Processing ExperimentsA number of researchers have proven the
usefulness of starch residues extracted fromstone pounding tools (Barton et al. 1998; Fulla-gar and Field 1996; Fullagar et al. 1997; Keal-hofer et al. 1999; Lentfer et al. 2002; Loy 1994;Loy et al. 1992; Perry 2001; Piperno and Holst1998; Piperno et al. 2000; Therin et al. 1999).As mentioned earlier, we observed what ap-peared to be damaged starch on tools studiedfor maize occurrence (Pearsall et al. 2004). Toinvestigate the source of this starch and otherorganic residues, we designed a series of exper-iments building upon Babot’s (2003) experi-mental work on how food processing affectsstarch. Babot’s work suggests that variousforms of traditional food processing techniquesalter starch in predictable ways. We conductedprocessing and cooking experiments usingmanioc and maize to determine the extent towhich we could infer food processing from theresidue assemblages on the Structure 20 tools.We cut and pounded fresh sweet manioc rootsand ground dried flour maize kernels with stone“artifakes”; we processed manioc roots bysoaking, fermenting, toasting on a griddle,oven-roasting, and boiling; we processed sweetcorn kernels by oven-roasting and toasting on agriddle. The results are summarized here; pho-tographs of the residues are presented on theMU Phytolith website (http://www.missouri.edu/~phyto).
Five minutes of pounding peeled maniocroots with a stone “mano” on a stone “metate”produced abundant starch residues on both im-plements. Granules from the mano typically

showed enlarged fissures centered on the hilumthat were circular to star-shaped, sometimesproducing an indented area. Most granules al-tered in this way lost the interference cross. An-other pattern was splitting of the surface of thegranule; this did not always result in loss of theinterference cross. Most manioc compoundgrains disarticulated into component granules.Interestingly, the pattern of granule alterationon the flake tool used to peel the roots was sim-ilar to that observed on the mano, but there wasmuch less damage to granules adhering to themetate. In the latter case, while some granulesshowed wider and longer fissures, the area ofthe hilum was usually not altered to the pointthat the interference cross was lost.
Grinding of maize for 10 minutes resulted inabundant starch assemblages on mano andmetate tools and similar patterns of alteration.Relatively few granules lost the interferencecross, but granule surfaces were “roughened” inappearance and fissures were wider and longerin comparison to unprocessed maize. Undam-aged granules were also observed, as were torntransparent tissues from the kernel. Babot’s(2003) tests of milling (grinding) maize, beans,and freeze-dried potatoes demonstrated similarstructural damage to granules: incomplete andfractured granules and granules with enlargedfissures (some showing darkened centers, as inroasted maize; see below). Damaged granulesalso showed defects in the shape of the interfer-ence cross. According to Babot (2003), millingdid not produce gelatinization or aggregation;this was our observation as well.
By contrast, soaking manioc led to someswelling of granules and increased fissuring andvacuole formation, but retention of the interfer-ence cross. Compound grains disarticulated intoindividual granules. Boiling and oven-roasting(high temperature heat) completely gelatinizedthe starch in manioc roots. No unaltered gran-ules remained. Roasting also largely gelatinizedstarch in sweet corn, but left some granules un-altered (still showing hilum and interferencecross) and others very translucent, two-dimensional, and lacking the interference cross.Babot (2003) observed similar results forroasted flour maize with pronounced rounded,irregular, or star-shaped projections appearingat the hilum (creating a dark center of the gran-ule). Toasting and fermentation (low-tempera-ture heat) of manioc each resulted in a combi-
110 ECONOMIC BOTANY [VOL. 60
nation of unaltered, aggregated, and gelatinizedstarch in samples. Toasting corn kernels (lowtemperature) produced masses of droplets (cornoil) of widely varying diameters, and left mostof the starch unaltered in appearance but largelylacking extinction crosses. A few granules withextinction crosses were found, however.
As a result of our experiments and literaturereview, we suggest that processing of raw maize(e.g., kernel grinding) or raw manioc (e.g.,pounding or cutting of peeled roots) should bothleave abundant starch residues on tools. In thecase of maize, both unaltered starch (unswollengranules with good interference crosses and fewfissures) and starch showing milling effects(roughened surfaces, enlarged fissures) would beexpected, and a high proportion of granuleswould retain the interference cross. Maniocstarch is more easily damaged by pounding, butstructurally intact granules should be present ifraw manioc is processed on tools, and some mayretain the interference cross, depending on thekind of force that was applied to the tissues. Pro-cessing of soaked, fermented, or toasted/parchedmanioc or maize on tools would result in in-creasing proportions of swollen and damagedgranules, granule aggregates, and gelatinizedstarch on tools. Processing fully cooked maniocand maize on tools would result in deposition ofgelatinized starch and other organic tissues fromboth foods, and some recognizable maize starchgranules. One would expect little recognizablemanioc starch in this case.
Root and Fruit Processing atthe Real Alto Site
We applied the new phytolith types and in-sights from starch processing to interpretingresidues from stone tools previously investi-gated for maize use (Pearsall et al. 2004). Thetools came from the floor deposits of Structure20, a domestic structure at the Real Alto site,Ecuador (Fig. 2, Fig 3). Real Alto is an EarlyFormative Valdivia Period site (Damp 1979;Lathrap et al. 1977; Marcos 1978; Marcos1998; Pearsall 1979; Pearsall et al. 2004; Zei-dler 1984; Zeidler 2003) occupied from Val-divia Phase 1 through Phase 7 (4400–1800B.C., calibrated) (Marcos 1998; Zeidler 2003)located in southwest Guayas province at 80° 43'W longitude 2° 22' S latitude near the modernport of Chanduy. The site is important in NewWorld prehistory because it arguably represents
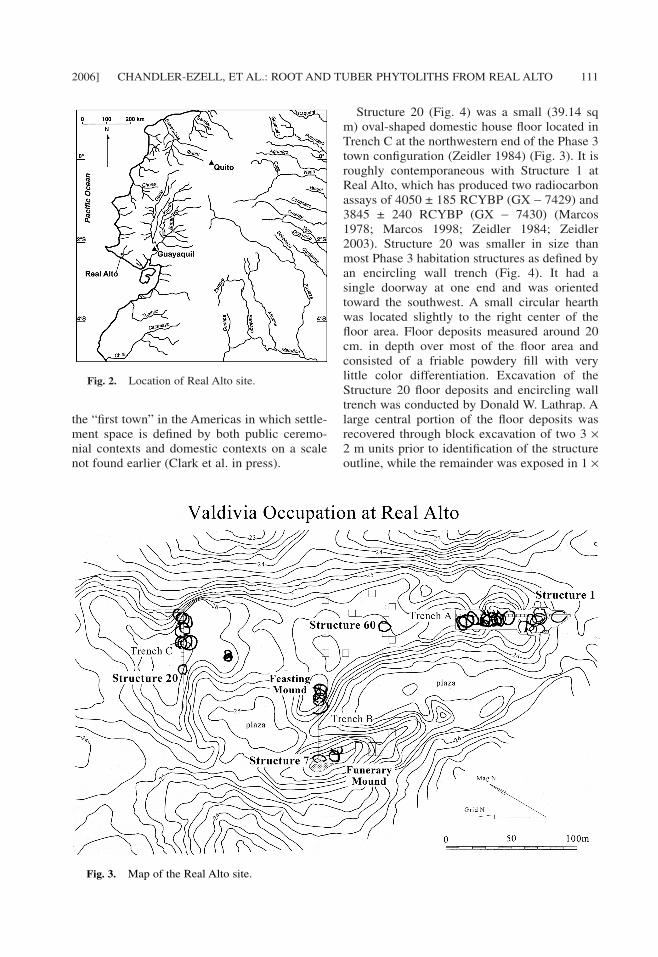
the “first town” in the Americas in which settle-ment space is defined by both public ceremo-nial contexts and domestic contexts on a scalenot found earlier (Clark et al. in press).
2006] CHANDLER-EZELL, ET AL.: ROOT AND TUBER PHYTOLITHS FROM REAL ALTO 111
Structure 20 (Fig. 4) was a small (39.14 sqm) oval-shaped domestic house floor located inTrench C at the northwestern end of the Phase 3town configuration (Zeidler 1984) (Fig. 3). It isroughly contemporaneous with Structure 1 atReal Alto, which has produced two radiocarbonassays of 4050 ± 185 RCYBP (GX − 7429) and3845 ± 240 RCYBP (GX − 7430) (Marcos1978; Marcos 1998; Zeidler 1984; Zeidler2003). Structure 20 was smaller in size thanmost Phase 3 habitation structures as defined byan encircling wall trench (Fig. 4). It had asingle doorway at one end and was orientedtoward the southwest. A small circular hearthwas located slightly to the right center of thefloor area. Floor deposits measured around 20cm. in depth over most of the floor area andconsisted of a friable powdery fill with verylittle color differentiation. Excavation of theStructure 20 floor deposits and encircling walltrench was conducted by Donald W. Lathrap. Alarge central portion of the floor deposits wasrecovered through block excavation of two 3 ×2 m units prior to identification of the structureoutline, while the remainder was exposed in 1 ×
Fig. 2. Location of Real Alto site.
Fig. 3. Map of the Real Alto site.

1 m units at four successive 5-cm levels (Zei-dler 1984; see Fig. 4 herein). All artifacts fromeach level were piece-plotted in situ prior to re-moval and bagging (see Zeidler 1984: Maps44–47, 91–116; see also Stahl and Zeidler1988).
We selected 17 stone tool fragments (two percontext, with one exception) from nine prove-niences in the bottom two levels (Levels 3 and4) of the Structure 20 floor deposits (Fig. 4).The tools were still stored in their original arti-fact bags and unwashed. (Tools are illustratedand described in Pearsall et al. 2004). In addi-tion to the tools, which were the primary focusof the case study, we reexamined and comparedphytolith extracts from Structure 20 floor sedi-ments originally studied by Pearsall (1979) andsediments from Valdivia 1-2 contexts. Recoveryof starch and phytoliths from tools is describedin Pearsall et al. (2004) and Chandler-Ezell andPearsall (2003).
Diagnostic phytoliths were recovered from16 of 17 tools but were not highly concentrated.Nondiagnostic silica bodies also occurred onsome of the tools. Manihot, Calathea, arrow-root family (Marantaceae), squash (Cucurbitaesubfamily), and bean family (Fabaceae) phy-toliths occurred on the tools in addition to themaize cob phytolith residues reported in
112 ECONOMIC BOTANY [VOL. 60
Pearsall et al. (2004) (Table 3, Fig. 5; seehttp://www.missouri.edu/~phyto for completeresults by sample and more illustrations). Thestrongest phytolith evidence for use of the toolsfor manioc root processing is presence of 160IManihot secretory body on five tools and in twofloor sediments from the structure. No wildManihot species occur on the coast of Ecuador(Jørgensen and León-Yánez 1999), making cul-tivated manioc the likely source. In addition, weidentified 24IC Euphorbiaceae epidermal cellon four other tools, including three cobbles.This phytolith occurs in manioc root and leafepidermis.
Phytolith type 26IIB, Calathea flat rhizomebodies, provide direct evidence for Calatheatuber processing on two tools (Table 3). In ad-dition Calathea/Maranta rhizome phytolithsand/or the conical and spherical phytoliths pro-duced in leaves and roots of Marantaceae oc-curred on six tools. We observed masses ofephemeral, perfect non-silica spheres of diversesizes on some tools that disappeared from slide-mounts over time. Similar spheres interpretedas oil droplets were observed on slides madefrom comparative samples of some modernmaize varieties, peanut, cashew, and chili pep-per. Other silicified residues found on tools included rooty transport elements, rooty
Fig. 4. Structure 20 house floor.

parenchyma, and fruity transport tissues (seeAppendix 2 for descriptions).
Phytolith production and concentration differin plant tissues. The roots, tubers, and rhizomeswe studied produced much lower concentra-tions of silica than leaves and inflorescences.Because of this, lower numbers or concentra-tions of phytoliths from underground tissuescan be expected in archaeological samples, andthe presence of even a few may indicate moreplant matter than an equal number of leaf phy-toliths. Though we found low concentrations ofroot-type phytoliths, they were diverse and sup-ported by other phytoliths and starch residues.
Starch residues were observed on all 17 tools(Table 3). We identified a total of 12 arrowrootstarch (Maranta arundinacea) granules fromeight tools. One tool, 3290-A, produced a frag-ment identifiable as Canna starch, which islarge and fragile. Three tools (3293-A, 3294-A,3331-A) produced small, hemispherical starchgrains. Though produced in manioc, thesegrains are not considered diagnostic, since othercrops, including maize, also produce them. Onediagnostic manioc starch granule occurred on3294-B in conjunction with the manioc 160Iphytolith type. As reported in Pearsall et al.(2004), maize starch granules occurred on alltools.
2006] CHANDLER-EZELL, ET AL.: ROOT AND TUBER PHYTOLITHS FROM REAL ALTO 113
A number of Structure 20 tools contained or-ganic material other than identifiable starchgrains, including bodies consistent in appear-ance with starch (yet lacking the interferencecross) and transparent and grainy materials re-sembling gelatinized starch. When heated,starch granules swell and lose the interferencecross under polarized light. If heating contin-ues, starch granules continue to swell. Thisgelatinization process is common to allstarches, although the temperature at whichgelatinization occurs varies somewhat amongplant species (Hancock and Tarbet 2000; Ship-man 1967). Even low temperature heating cancause slight surface gelatinization, with gran-ules clumping together into large aggregateswhich retain their diagnostic features.
By contrast, phytoliths preserve in eithercooked or raw tissues. The more common oc-currence of Manihot and Calathea phytolithsthan starch on tools suggests these plants wereprocessed cooked rather than raw (compareManihot phytoliths on five tools, manioc starchon one; Calathea phytoliths on two tools, starchon none). Undamaged arrowroot and maizestarch recovered on tools suggests that theseplants were processed raw, or in the case ofmaize perhaps after low-temperature processing(e.g., parching). While we cannot definitively
Table 3. Summary of food residues on Structure 20 tools.1
Tool Manioc Arrowroot Calathea CannaArrowroot
FamilyRooty
TransportRooty Pa-renchyma
FruityTransport
AlteredStarch Maize
3290-A S P P P P G, M, T S3290-B P P P G, T S3292-A T P, S3292-B P D, G, T S3293-A S P P M, T S3293-B S P P P M, T S3294-A S P P P D, T P, S3294-B P, S S P P P P G, T P, S3331-A P P P G, T P, S3331-B P S P T P, S3383-A P S P P P M S3383-B S M, T S3386 P P P P G, T P, S3415-A P P P P G, T S3415-B P P G, T S, P3417-A S P G, M, T S3417-B P S
1 See MU Phytolith website (http://www.missouri.edu/�phyto) for occurrence of residues by sediment. Table key: Pphytolith, S starch, D damaged starch granule, G granular tissue, M damaged maize granule, T transparent tissue.

Fig. 5. Selected phytoliths and starch granules recovered from tools. See MU Phytolith website (http://www.missouri.edu/~phyto) for other photographs. a. 26IAa Calathea irregular rhizome cylinder, PS1749 FS 3290-A. b.26IAb Calathea flat domed rhizome cylinder, PS1749 FS 3290-A. c. 26IAb Calathea flat domed rhizome cylinder,PS1765 FS 3293-B. d. 26IIB Calathea flat rhizome type, PS1804 FS3294-B. e. 160I Manihotsecretory cell,PS1804FS 3294-B. f. 160I Manihot secretory cell, PS1804FS 3294-B. g. 160I Manihot secretory cell, PS3,Structure 20 floor sediment. h. 160I Manihot secretory cell, PS1804 FS3294-B. i. Manihot esculenta starchgranule, SS42 FS3294-B. j. Broken Canna starch granule, SS15 FS3290-A. k. Maranta arundinacea starchgranule, SS42 FS3294-B l altered starch, SS85 FS3417-A.

identify the transparent and grainy organicresidues on tools as gelatinized starch, the ap-pearance of these residues is consistent withthis source based on our experiments. Possiblemechanisms for deposition of gelatinized starchinclude grinding or mashing of cooked foods onthe tools.
Alternative explanations for the observedresidue patterns must be considered. We did nottest arrowroot for the effects of cooking; it maybe that enough granules survive unaltered aftercooking to negate our interpretation of raw pro-cessing of this plant. Arrowroot granules arequite fragile, however, even damaged by ex-tended exposure to light microscopy (personalobservation; see also Korstanje 2003), makingit unlikely that they would survive cooking. It ispossible that manioc, llerén, and Canna starchare more subject to long-term post-depositionaldamage than maize, and thus less likely to sur-vive on tools, creating the observed pattern. Ourexperiments show that manioc is more easilydamaged by food processing than is maize, butlong-term survivorship is unknown. Researchby Lu (2003) comparing survivorship of taro,yam (roots/tubers), millet, and rice (grains)starch on milling stones buried 71 days, how-ever, found similar survival rates (roots/tubers:76%, 65% survival; grains: 80%, 77% sur-vival). It is possible that starch became gelati-nized after deposition on the tools, rather thanas a result of processing cooked foods. Thetools do show the effects of heating. We feelthis scenario is less likely than processing ofcooked foods on the tools, however, since bothunaltered and altered starch granules are pres-ent. The presence of charcoal particles on toolsalso suggests that toasted or roasted foods wereprocessed on the tools (Pearsall et al. 2004).There is no evidence that the patterns of starchpreservation and alteration observed in theStructure 20 tool assemblage were caused bylaboratory processing: we used low tempera-tures during processing, minimized use of caus-tic chemicals (rarely using a flotation medium),used glycerol rather than water-mounts, andminimized exposure of slides to high intensitylight (Korstanje 2003).
To summarize, there are a number of waysmanioc could be processed on a tool that wouldrarely leave identifiable starch, but could resultin deposition of phytoliths or amorphous or-ganic tissues. Lack (or rare occurrence) of man-
2006] CHANDLER-EZELL, ET AL.: ROOT AND TUBER PHYTOLITHS FROM REAL ALTO 115
ioc starch—but presence of manioc phytolithsor root-type tissues—implies that the tools wereused on already cooked or fermented roots,rather than on raw or merely soaked roots.Maize processing in any form—raw, parched,roasted, popped—would result in deposition oflarge to small amounts of undamaged starchgranules on tools.
There are differences in the numbers andkinds of plants represented on the tools. Tool3290-A, for example, appears to be the multi-purpose tool of the Structure 20 household,used for grinding maize, crushing roots and tu-bers, and processing sapote, squash, bean, orpossibly other fruits. Tool 3290-B, from thesame context, is very similar in its functions.By contrast, tools 3292-A, 3292-B, and 3417-Bhave many fewer kinds of residues, suggestinga more specialized function as maize process-ing tools (with some fruit use). The other toolsshow a consistent combination of root/tuber andmaize processing, with and without use forfruits. These results suggest that stone “grind-ing” and “pounding” tools are typically multi-purpose and more rarely specialized. It is pos-sible, however, that a few tools look morespecialized because starch has been degraded orthe fragile phytoliths produced by fruits de-stroyed. Several tools with few kinds ofresidues have little residue overall. However,the few remains preserved on tool 3290-B givealmost as diverse an assemblage as the muchbetter preserved 3290-A.
The results of this study provide insights intothe prehistoric distribution of manioc, and itsrole in early tropical forest agriculture. Re-search suggests that manioc originated either inthe seasonal forests of inland Guianas/Venezuela or the cerrados of central Brazil(Piperno and Pearsall 1998). Perry’s (2002) re-search into size and morphology of starch frommodern and archaeological manioc from coastalPeru reveals a distinctive “coastal” maniocstarch morphology: hemispherical granuleswith basal faceting that are less “inflated” thanAmazonian or Panamanian manioc, and havelinear central fissures, rather than stellate fis-sures. The Real Alto manioc granule is of this“coastal” type, as is manioc grown today incoastal Ecuador. Coastal and lowland Neotropi-cal manioc types have been separated for sev-eral millennia (Perry 2002).
The archaeological record of manioc is

scanty (Piperno and Pearsall 1998; Pearsall1992). Early finds come from the Peruvianhighlands, representing introduction from theeast: at Tres Ventanas Cave (8000–6000 B.C.)and at sites in Quebrada de Las Pircas (6200–5500 B.C.). The earliest manioc on the Peru-vian coast comes from Los Gavilanes (2700–2200 B.C.); other finds date to the Initial period(after 1800 B.C.) and after. Occurrence of man-ioc at Real Alto in southwest Ecuador at 2800–2400 cal. B.C. is thus roughly contemporarywith the earliest coastal Peruvian record, andextends the known distribution of the crop inSouth America northward into Ecuador.
Manioc was not the first root crop grown incoastal Ecuador. Llerén inflorescence phytolithsoccur at the Vegas site by 7000 B.C. (Pipernoand Pearsall 1998), and as documented in our re-search, use of both Calathea and arrowroot tu-bers occurs along with manioc at Real Alto. Pro-ductive and undemanding, manioc producesmore carbohydrates per edible portion than otherlowland root/tubers (Piperno and Pearsall 1998).The introduction of manioc to coastal Ecuador,into a subsistence system based on maize andMarantaceae tubers, may have supported thepopulation expansions out of southwest Ecuadorthat are documented in late Valdivia times(Pearsall 1999; Staller 1994; Zeidler 1991).
ConclusionsIn a study that began with our puzzling over
phytoliths found on stone tools that were simi-lar to, but not quite typological matches for, theleaf and inflorescence bodies of twoeconomically-important plants, we have identi-fied phytoliths produced in roots, tubers, andrhizomes and demonstrated their potential foridentifying a new category of plant tissue ar-chaeologically. This research is particularly rel-evant for economically important root and tubercrops. For a variety of botanical, preservation,and cultural reasons, such dietary staples havebeen elusive in the archaeological record. Manyof these plants are propagated vegetatively, re-ducing the chances of finding inflorescence re-mains. In other cases, harvesting and process-ing of many food crops may leave most of thenon-edible portions (leaves and inflorescences)of these plants in the field. The ability to iden-tify the edible portion of the plant in a contextsuch as on tools, hearths, dental calculus, ves-sels, and structures provides more direct evi-
116 ECONOMIC BOTANY [VOL. 60
dence for plant consumption as food. The phy-toliths observed in these plants increase therange of diagnostic tools for future investiga-tions. We suggest the time has come to investi-gate phytolith production in edible undergroundstorage organs such as roots, tubers, and rhi-zomes to shed new light on the ways theseplants were used in the past.
Phytoliths observed in plants we studied in-crease our range of diagnostic tools for futureinvestigations by documenting the presence ofknown diagnostics in new tissues and discover-ing new types. Of particular importance is iden-tification of 160I Manihot secretory body, andfour types that enhance our ability to identifyCalathea and Maranta. The discovery that sil-ica casts of parenchyma and transport elementscan indicate the “rooty” or “fruity” origin ofresidues provides us with another line of evi-dence for understanding food use. Our findingsdemonstrate the utility of studying plants orplant tissues that produce low concentrations ofphytoliths, and enhance our understanding ofphytolith formation in plants.
In the late 1980s Piperno revolutionized phy-tolith analysis by demonstrating that distinctlydifferent diagnostic phytoliths occur in inflores-cences than in leaves (Piperno 1988; Piperno1989). We suggest the time has come to investi-gate phytolith production in underground stor-age organs, focusing on the epidermis, process-ing larger samples to compensate for lowersilica concentrations, and adapting processingmethods to cope with the different chemicalconstituents of roots and tubers.
Application of our findings, including newinsights into alteration of starch residues duringfood processing, provided insight into EarlyFormative food-ways and tool use at Real Altoduring its Phase 3 town configuration. Structure20’s inhabitants routinely processed maize(Pearsall et al. 2004) as well as a suite of rootand tuber foods that contributed to a high diver-sity diet. These findings constitute the earliest,clearest record for use of manioc, Calathea, andarrowroot roots and tubers in coastal Ecuador(2800–2400 B.C. calibrated) and provide addi-tional support for the agricultural basis of Val-divia society.
AcknowledgmentsFinancial support by MU Research Board
grant to Pearsall. Thomas E. Emerson for ar-

ranging loan of Real Alto stone tools curated atUniversity of Illinois. Marie J. Zeidler for Fig.3 and 4; William Grimm and Neil Duncan forthe other figures. Dolores Piperno and LindaPerry for assistance with manioc starch identifi-cation. Tom Ezell for assistance with acetoni-trile. César Veintimilla, Centro de Estudios Arqueológicos y Antropológicos, Escuela Supe-rior Politécnica del Ecuador, and Karen Stothertfor assistance locating Calathea rhizomes.Robin Kennedy for discussion of anatomicalorigins of root/tuber phytoliths.
Literature CitedBabot, M. del P. 2003. Starch grain damage as an in-
dicator of food processing. Pages 69–81 in D. M.Hart and L. A. Wallis, eds. Phytolith and starch re-search in the Australian-Pacific-Asian regions:The state of the art. Vol. 19. Pandanus Books,Australian National University, Canberra.
Barton, H., R. Torrence, and F. Fullagar. 1998. Cluesto stone tool function re-examined: Comparingstarch grain frequencies on used and unused ob-sidian artefacts. Journal of Archaeological Sci-ence 25:1231–1238.
Bowdery, D., D. M. Hart, C. Lentfer, and Lynley A.Wallis. 2001. A universal phytolith key. Pages267–278 in Jean Dominique Meunier and FabriceColin, eds. Phytoliths: Applications in earth sci-ences and human history. A. A. Balkema, Lisse,The Netherlands.
Chandler-Ezell, K., and D. M. Pearsall. 2003. “Piggy-back” microfossil processing: Joint starch andphytolith sampling from stone tools. The Phy-tolitharien 15:2–8.
Clark, J. E., J. L. Gibson, and J. A. Zeidler. n.d. Firsttowns in the Americas: Searching for agricultureand other enabling conditions. In William Pren-tiss and Ian Kuijt, eds. The archaeology of sub-sistence and social change in intermediate scalesocieties. University of Arizona Press, Tucson (inpress).
Damp, J. 1979. Better homes and gardens: The lifeand death of the early Valdivia community. Ph.D.thesis. Archaeology Department, University ofCalgary.
Fullagar, R., F. Furby, and B. Hardy. 1996. Residueson stone artifacts: State of a scientific art. Antiq-uity 70:740–745.
———, and J. Field. 1997. Pleistocene seed-grinding implements from the Australian aridzone. Antiquity 71:300–307.
Haslam, M. 2004. The decomposition of starchgrains in soils: Implications for archaeologicalresidue analysis. Journal of Archaeological Sci-ence (in press, corrected proof).
2006] CHANDLER-EZELL, ET AL.: ROOT AND TUBER PHYTOLITHS FROM REAL ALTO 117
Hancock, R., and B. Tarbet. 2000. The other doublehelix: The fascinating chemistry of starch. Journalof Chemical Education 77:988–992.
Jørgensen, P. M., and S. León-Yánez. 1999. Cata-logue of the vascular plants of Ecuador. MissouriBotanical Garden Press, St. Louis.
Kealhofer, L., R. Torrence, and R. Fullagar, 1999. In-tegrating phytoliths within use-wear/residue stud-ies of stone tools. Journal of Archaeological Sci-ence 26:527–546.
Korstanje, M. 2003. Taphonomy in the laboratory:Starch damage and multiple microfossil recoveryfrom sediments. Pages 105–118 in D. M. Hart andL. A. Wallis, eds. Phytolith and starch research inthe Australian-Pacific-Asian regions: The state ofthe art. Vol. 19. Pandanus, Australian NationalUniversity, Canberra.
Lathrap, D., J. G. Marcos, and J. A. Zeidler. 1977.Real Alto: An ancient ceremonial center. Archae-ology 30(1):2–13.
Lentfer, C., M. Therin, and R. Torrence. 2002. Starchgrains and environmental reconstruction: A mod-ern test case from West New Britain, Papua NewGuinea. Journal of Archaeological Science29:687–698.
Loy, T. H. 1994. Methods in the analysis of starchresidues on prehistoric stone tools. Pages 86–114in J. G. Hather, ed. Tropical archaeobotany: Appli-cations and new developments. Routledge, Lon-don.
———, M. Spriggs, and S. Wickler. 1992. Direct ev-idence for human use of plants 28,000 years ago:Starch residues on stone artefacts from the north-ern Solomon Islands. Antiquity 66:898–912.
Lu, T. 2003. The survival of starch residue in a sub-tropical environment. Pages 119–126 in D. M.Hart and L. A. Wallis, eds. Phytolith and starch re-search in the Australian-Pacific-Asian regions:The state of the art. Vol. 19. Pandanus, AustralianNational University, Canberra.
Madella, M., A. Alexandre, and T. Ball. 2005. Inter-national code for phytolith nomenclature 1.0. An-nals of Botany 96:253–260.
Marcos, J. G. 1978. The ceremonial precinct at RealAlto: Organization of time and space in Valdiviasociety. Ph.D. thesis. Anthropology Department,University of Illinois, Urbana.
———. 1998. A reassessment of the chronology ofthe Ecuadorian Formative. Pages 277–324 in M.Guinea, J. G. Marcos, and J-F. Bouchard, eds. ElÁrea Septentrional Andina: Arqueología y etno-historia. Ediciones Abya-Yala, Quito.
Pearsall, D. M. 1979. The application of ethnobotan-ical techniques to the problem of subsistence inthe Ecuadorian Formative. Ph.D. thesis. Anthro-pology Department, University of Illinois, Ur-bana.
———. 1992. The origins of plant cultivation in

South America. Pages 173–205 in C. W. Cowanand P. J. Watson, eds. Origins of agriculture: Aninternational perspective. Smithsonian InstitutionPress, Washington, D.C.
———. 1999. Agricultural evolution and the emer-gence of Formative societies in Ecuador. Pages161–170 in M. Blake, ed. Pacific Latin America inprehistory: The evolution of Archaic and Forma-tive cultures. Washington State University Press,Pullman.
———. 2000. Paleoethnobotany. A handbook ofprocedures. 2nd ed. Academic Press, San Diego.
———, K. A. Chandler-Ezell, and J. A. Zeidler.2004. Maize in ancient Ecuador: Results ofresidue analysis of stone tools from the Real Altosite. Journal of Archaeological Science 31:423–442.
———, and R. Dinan. 1992. University of Missouriphytolith classification system. Pages 37–64 in G.Rapp, Jr., and S. Mulholland, eds. Studies in phy-tolith systematics. Plenum, New York.
Perry, L. 2001. Prehispanic subsistence in the MiddleOrinoco Basin: Starch analyses yield new evi-dence. Ph.D. thesis. Anthropology Department,Southern Illinois University, Carbondale.
———. 2002. Starch granule size and the domesti-cation of manioc (Manihot esculenta) and sweetpotato (Ipomoea batatas). Economic Botany56:335–349.
Piperno, D. R. 1988. Phytolith analysis: An archaeo-logical and geological perspective. AcademicPress, San Diego.
———. 1989. The occurrence of phytoliths in repro-ductive structures of selected tropical an-giosperms and their significance in tropical pale-oecology, paleoethnobotany, and systematics.Rev. Paleobot. Palynol. 61:147–173.
———, and I. Holst. 1998. The presence of starchgrains on prehistoric tools from the humidNeotropics: Indications of early tuber use andagriculture in Panama. Journal of ArchaeologicalScience 25:765–776.
———, and D. M. Pearsall. 1998. Origins of agricul-ture in the lowland Neotropics. Academic Press,San Diego.
———, A. J. Ranere, I. Holst, and P. Hansell. 2000.Starch grains reveal early root crop horticulture in
118 ECONOMIC BOTANY [VOL. 60
the Panamanian tropical forest. Nature 407:894–897.
———, E. Weiss, I. Holst, and D. Nadel. 2004. Pro-cessing of wild cereal grains in the Upper Pale-olithic revealed by starch grain analysis. Nature430:670–673.
Shipman, L. 1967. Manufacture of tapioca, arrow-root, and sago starches. Pages 103–119 in R.Whistler and E. Paschall, eds. Starch: Chemistryand technology. Vol. II, Industrial aspects. Aca-demic Press, New York.
Stahl, P. W., and J. A. Zeidler. 1988. The spatial cor-respondence of selected bone properties and in-ferred activity areas in an Early Formativedwelling structure (S 20) at Real Alto, Ecuador.Pages 275–298 in Nicholas J. Saunders andOlivier de Montmollin, eds. Recent studies in Pre-Columbian archaeology. BAR International Series421, Oxford.
Staller, J. E. 1994. Late Valdivia occupation in El OroProvince, Ecuador: Excavations at the Early Forma-tive period (3500–1500 B.C.) site of La Emeren-ciana. Ph.D. thesis. Anthropology Department,Southern Methodist University, Dallas, Texas.
Therin, M., R. Fullagar, and R. Torrence. 1999.Starch in sediments: A new approach to the studyof subsistence and land use in Papua New Guinea.Pages 438–462 in C. Gosden and J. Hather, eds.The prehistory of food: Appetites for change.Routledge, London.
Zeidler, J. A. 1984. Social space in Valdivia society:Community patterning and domestic structures atReal Alto, 3000–2000 B.C. Ph.D. thesis. Anthro-pology Department, University of Illinois, Ur-bana.
———. 1991. Maritime exchange in the Early For-mative period of coastal Ecuador: Geopoliticalorigins of uneven development. Pages 247–268 inB. L. Isaac, ed. Research in economic anthropol-ogy. Volume 13. JAI Press, Greenwich, Connecti-cut.
———. 2003. Formative Period chronology for thecoast and western lowlands of Ecuador. Pages487–527 in J. Raymond and R. Burger, eds. Ar-chaeology in Formative Ecuador, A symposium atDumbarton Oaks. Dumbarton Oaks Research Li-brary and Collection, Washington, D.C.
Appendix 1. Phytolith Nomenclature
We conducted the research reported here during the period when the ICPN (International Code for PhytolithNomenclature) working group, among others (e.g., Bowdery et al. 2001), was developing an international code forphytolith nomenclature. The types described here were finalized within the MU phytolith classification system priorto publication of the ICPN code by Madella et al. (2005). The ICPN code formalizes phytolith naming based onthree descriptors: shape, texture or ornamentation, and anatomical origin (at tissue structure or cell level), if known.
We support the efforts of the ICPN working group and contributed ideas to the code. Below are names for themajor types reported here (Fig. 1) that conform, as much as possible, to the code. A few additions to the descriptorspublished by Madella et al. (2005) are necessary to describe the types:

2006] CHANDLER-EZELL, ET AL.: ROOT AND TUBER PHYTOLITHS FROM REAL ALTO 119
Undulating: corresponds to ICPN sinuate, but used as a surface feature, not an edge feature as in the code.Gemmate: a more formal term for beaded: having rounded processes that are equal in diameter and height.Nodular: having unevenly distributed processes with obtuse (> 90 degree edge junctions) or rounded tips.Ciliate: having fine rod or pillar-like processes that are longer than wide, and are not straight.Stellate, as an ornamentation descriptor: having raised linear features in a star-shaped pattern.Our spiked corresponds to ICPN echinate; smooth to ICPN psilate.The ICPN shape categories can be applied to some of the root/tuber types if modifiers referring to cross-section are
added. There is no ICPN shape category that fits the manioc secretory cell, which is heart-shaped. Using theLatin root word pectus, this shape would be pectate. The flattened, somewhat elongated, platform shape of 26IIBcould be glossed as platform, or pulpitate (Latin root word pulpitum).
The major types, following the ICPN code are:26IAa: cylindric (irregular cross section) undulating/gemmate/nodular/echinate26IAb: cylindric (polygonal cross section) psilate (head)/ciliate/gemmate (body)26IB: acicular nodular26IB: pulpitate (irregular cross section) gemmate (edge)/stellate (center)160I: pectate psilate secretory cell
Appendix 2. Description of other distinctive phytoliths observed in plant specimens stud-ied during the project. * marks new types. Phytolith photographs and a table documentingthe occurrence of types by specimen and tissue are posted on the MU Phytolith website(http://www.missouri.edu/~phyto).
23 Occluded plates, cell walls: Blackened bodies derived from epidermis or possibly parenchyma cell walls.24IC Euphorbiaceae epidermal cell: Blocky epidermal cells with multiple facets, previously identified in Eu-
phorbiaceae leaves, now also found in manioc (Manihot esculenta) root rind.*26IIA Dioscorea (yam) elongate, undulating root body: Non-quadrilateral body with flat, undulating appear-
ance, overall ovate outline, and a central ridge.40IVB Hair base composed of rounded cells: General class of hair bases documented in a variety of Neotropical
taxa leaf tissues (see MU website, http://www.missouri.edu/~phyto). Here documented in manioc stem.40IVF Hair base with stellate center: Broad class of hair bases, occurring in leaf tissues, observed here in man-
ioc fruits.80IAa1 Chrysobalanaceae small smooth sphere: Previously identified (and described) leaf type now documented
in Chrysobalanus icaco fruits.80IB Rugulose sphere: Observed in leaves of numerous taxa; also occurs in root/tuber and seed tissue, including
Canna and Marantaceae.80ICa Marantaceae nodular sphere: Spherical phytolith with irregularly spaced nodules on surface. Previously
documented in Marantaceae leaves, also occurs in Maranta and Calathea rhizomes, and very rarely in Is-chnosiphon leucophaeus root. A similar sphere occurs in the root of Costus scaber (Zingiberaceae); we plan towork further on this type.
80ICd Marantaceae (Calathea/Maranta) flattened nodular body: Flattened to spheroidal with linear ridges,nodular projections, and occasional spinules. Originally observed in Calathea allouia inflorescence, this typealso occurs in Maranta gibba stem base.
80IE Zingiberales irregularly angled and folded sphere: This type, defined early in Neotropical phytolith re-search (Pearsall 2000), occurs widely in leaves of Zingiberales taxa, including Canna, Calathea, and Maranta.This study documents these spheres in underground tissues of these taxa, as well as in Thalia geniculata rootand Costus root and rhizome.
80IIAb Heliconia trough body, pointed ends: Elongated trough body with pointed ends, a deep, compact trough,and small edge projections. Originally observed in Heliconia leaf; also occurs in root and rhizome.
80IIAc Heliconia trough body, blunt ends: Elongated trough body with blunt ends, a deep, compact trough, andsmall edge projections. Originally observed in Heliconia leaf; also occurs in root and rhizome.
*80IIBb Verrucate trough body: Decorated, textured verrucate platform with a multi-outlined pit in the center ofone side. Seeds, fruit, and root cortex.
80IIIB Marantaceae conical body: Body has a single conical projection, nodular and rugulose surface decoration.Marantaceae leaf type also occurs in Maranta and Calathea rhizomes, as well as in Stromathe jacquinii andThalia geniculata root and stem base.
100ICa Smooth cystolith, sculptured: Cystoliths are found in leaves of numerous taxa. We observed thesesmooth, sculpted cystoliths in Dioscorea and Araceae tubers.

120 ECONOMIC BOTANY [VOL. 60
110 Schlerids: Commonly silicified in leaf tissue, especially in woody plants. We observed schlerids in seeds, tu-bers, and roots.
*130I Blocky parenchyma: Large, rounded blocks of silica which are casts of parenchyma cells formed from sil-ica sequestered there. Bodies are very translucent, with flattened or undulating surfaces, and occur in root andtuber cortex tissue.
*130IIA Multiple-lobed parenchyma: Silica casts of parenchyma with multiple rounded lobes, such as thoseformed in rapidly growing tissues, observed in roots and tubers.
*130IIB Two-lobed parenchyma: Silica casts of parenchyma limited to two rounded lobes observed in Ingaseeds. Further comparative research needed to determine the distribution of this type in other Fabaceae taxa.
*140I Undulating transport elements: Silica casts of transport elements (xylem and phloem) observed in a num-ber of taxa, predominantly in fruits and seeds. Type includes thin-walled, smooth, undulating tubes without endplates.
*140IIA Straight transport elements: Straight, silicified transport elements arranged in bundles. Found in rootsand tubers.
*140IIB Striated, wide transport elements: Open-ended transport elements with striations running perpendicularto the long dimension. Found in roots, fruits, and seeds.
*150I Fiber bundles: Groupings of long, small diameter fibers, packed closely in off-set ranks.*150II Fibrous mesh: We observed silicified fibers in irregular masses, mesh-like in appearance, in Fabaceae
seeds and pod.
Copyright © 2022 FDOKUMEN




















