
Do phytoliths play an antiherbivory role in southwest Asian Asteraceae species and to what extent?
11
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights
Transcript of Do phytoliths play an antiherbivory role in southwest Asian Asteraceae species and to what extent?

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Do phytoliths play an antiherbivory role in southwest Asian Asteraceaespecies and to what extent?
Ofir Katz a,1,*, Simcha Lev-Yadun b, Pua Bar a
aDepartment of Geography and Environmental Development, Ben-Gurion University of the Negev, Be’er-Sheva 84105, IsraelbDepartment of Biology & Environment, Faculty of Natural Sciences, University of Haifa – Oranim, Tivon 36006, Israel
A R T I C L E I N F O
Article history:Received 12 November 2013Accepted 20 March 2014Edited by R. LöschAvailable online 9 May 2014
Keywords:AsteraceaeDefenceHerbivoryPhytolithPoaceaeSilica
A B S T R A C T
Phytoliths (silica bodies) occur in Poaceae species in large numbers and have been shown to haveantiherbivory roles. However, phytoliths occur also in many other taxa in much smaller numbers, whichraises the question of the extent of both their potential and actual antiherbivory role in these taxa. Inorder to address the question of their potential antiherbivory role, we sampled 20 wild-growingsouthwest Asian species of the family Asteraceae, species of which have a much lower phytolithconcentrations than Poaceae taxa. We studied the potential positive effect of grazing on phytolithformation and the possible tendency of plants to have higher concentrations of such defence structures intheir reproductive organs. We sampled plants from populations of 12 non-spiny and eight spiny speciesgrowing in un-grazed and grazed plots in seven sites along a large rainfall gradient (80–900 mm meanannual) in Israel, a region known for its long and intensive grazing history. The study included 21 pairs ofun-grazed and grazed plants from 16 of these 20 species. In addition, ten populations of eight specieswere sampled in order to examine whether phytolith concentrations in the reproductive organs arehigher than in vegetative organs. We did not find consistently higher phytolith concentrations in grazedplants compared to un-grazed plants of the same species and habitat (15 species), and in 12 out of 21 pairsof un-grazed and grazed plants (from 15 species) we even found higher phytolith concentrations in un-grazed plants, a phenomenon which was more common in the more arid sites. Phytolith concentrationsin inflorescences are commonly (6 out of the 8 species) lower than in the rest of the shoot. We concludethat the antiherbivory potential of phytoliths in the southwest Asian Asteraceae as a group is muchsmaller than in the Poaceae.
ã 2014 Elsevier GmbH. All rights reserved.
Introduction
Phytoliths are tiny bodies of amorphous silica, detectable bylight microscopy, deposited either inside or in association withplant cells, thus shaped with some or even strong similarity to theshapes of their cells of origin. Phytoliths in Poaceae species occur inlarge numbers (hundreds of thousands to tens of millions per gramplant dry matter) and are the main form of silica in most plantspecies in which they occur (Piperno, 2006). Silica and phytolithsplay multiple roles in Poaceae species, alleviating both abiotic andbiotic stresses, such as water deficit, extreme high and lowtemperatures, heavy metal toxicity, salinity and herbivory (Cookeand Leishman, 2011; Epstein, 1999; Guntzer et al., 2012; Ma, 2004;
Ma and Yamaji, 2008; Piperno, 2006). Invertebrate (Reynolds et al.,2009) and small vertebrate (Massey and Hartley, 2006) herbivoreshave been shown to be negatively affected by plant silica. Theseeffects include abrasion of mouth parts of invertebrates and teethof mammals (e.g., Kaiser et al., 2013; Massey and Hartley, 2006;Mihlbachler et al., 2011), reduced digestibility caused by phytoliths(e.g., Hunt et al., 2008), nutritional effects of both phytoliths andsoluble silica (e.g., Massey and Hartley, 2009) and attractingnatural enemies of herbivorous insects induced by soluble silica(Kvedaras et al., 2010). Accordingly, many herbivores prefer plantswith lower silica and phytolith concentrations when given thechoice between plants from the same or closely related species ofthe Poaceae under controlled conditions (Massey et al., 2007a,b;Van-Soest, 2006). The little available evidence gathered so farindicates that phytoliths have similar although less prominenteffects on larger herbivores such as deer and cattle (Hunt et al.,2008; Soininen et al., 2013).
In contrast to Poaceae, species of many other plant familiesusually have much lower phytolith concentrations, usually not
* Corresponding author at: Ben-Gurion University of the Negev, POB 653, Be’er-Sheva 84105, Israel. Tel.: +972 8 6477379.
E-mail address: [email protected] (O. Katz).1 In partial fulfilment of requirements for a Ph.D. degree.
http://dx.doi.org/10.1016/j.flora.2014.03.0100367-2530/ã 2014 Elsevier GmbH. All rights reserved.
Flora 209 (2014) 349–358
Contents lists available at ScienceDirect
Flora
journal homepage: www.elsevier .com/ locate /flora

Author's personal copy
exceeding tens of thousands of phytoliths per gram dry matter(e.g., Piperno, 2006; Tsartsidou et al., 2007), and it is thusuncertain whether phytoliths play a significant antiherbivory rolein these taxa as well. One key issue is that phytoliths are most likelya quantitative defence mechanism, and are therefore expected tohave a lower antiherbivory function as phytolith concentrationsdecrease. Nonetheless, since studies of the defensive role ofphytoliths so far have focused on the Poaceae (to the best of ourknowledge, there is no study of non-Poaceae species), only alimited range of phytolith and silica concentrations has beendiscussed so far. Moreover, the range of phytolith concentrations inPoaceae species is closer to the higher end of the phytolith/silicaconcentrations spectrum found in plants, and there are practicallyno data about the defensive potential of phytoliths along most ofthis spectrum. We therefore do not know whether the anti-herbivory function of phytoliths simply declines with decreasingphytolith concentrations, or whether a threshold exists underwhich phytoliths have no defensive function. In order toinvestigate this question, we have studied whether Asteraceaespecies from a region with a very long history of grazing and withvarying phytolith concentrations follow defence-related patternsas those previously observed in the more phytolith-rich Poaceae.
The Asteraceae is one of the largest and most diverse plantfamily (Bremer, 1994), consisting in the flora of southwest Asia ofhundreds of both spiny and non-spiny species (Feinbrun-Dothanand Danin, 1991; Ronel and Lev-Yadun, 2012; Ronel et al., 2009).Phytolith concentrations in Asteraceae are low compared toPoaceae, as it is the case also in many other phytolith-poor plantfamilies (Katz et al., 2013; Piperno, 2006; Tsartsidou et al., 2007;Wallis, 2003). In a previous study, we found that phytolithconcentrations in the Asteraceae range between almost zero andseveral tens of thousands of phytoliths per g dry matter (Katz et al.,2013), compared to hundreds of thousands to tens of millions per gdry matter in the Poaceae. Therefore, the Asteraceae complementthe lower range of phytolith concentrations in higher plants andare suitable for our study.
We focused our study on 20 wild Asteraceae species (about 7%of the Asteraceae species in the flora of Israel), providing one of thelargest studies of potential phytolith functions in terms of speciesnumber. Our study is composed of two parts, as detailed below. We
studied the potential positive effect of grazing on phytolithformation and the possible tendency of plants to have higherconcentrations in their reproductive organs of these structuresthat potentially function as defence against herbivory. Bothphenomena were previously observed with respect to phytolithsof Poaceae. We therefore suggest that if these phenomena are alsofound to characterise the Asteraceae species, then phytoliths havean antiherbivory function in these species.
The effect of grazing on phytolith formation in Asteraceae
Brizuela et al. (1986) observed that silica concentrations innaturally growing Poaceae species are negatively correlated to thedistance from colonies of prairie dogs (Cynomys ludovicianus;Rodentia), the main small mammalian herbivore species in theirstudy area. Cid et al. (1989) suggested that selection against silica-low plants on one hand and induced response to herbivore damageon the other are the main causes of this phenomenon. Massey et al.(2008) showed a cyclic reciprocal relationship between grass silicaconcentration and vole (Microtus agrestis L., Rodentia) populationsize, which implies negative effects of high silica concentrations onvoles and grazing-induced natural selection for silica-rich grasses.Induction of increased silicification by herbivory in the Poaceae iswell documented (Massey et al., 2007b, 2008, 2009; McNaughtonand Tarrants, 1983; McNaughton et al., 1985), but the level ofinduced silicification varies according to the identity of theherbivores (Massey et al., 2007b; Soininen et al., 2013) andplant genotypes (Garbuzov et al., 2011; Melzer et al., 2010;Soininen et al., 2013).
Recently, Soininen et al. (2013) studied the differences inphytolith concentrations in eight naturally growing Poaceaespecies with and without the presence of a large mammalianherbivore (Rangifer tarandus L., Artiodactyla). They found that thegrazed plants did not always have higher silica concentrations thanthe un-grazed plants, and even found lower silica concentrations ingrazed compared to ungrazed Avenella flexuosa L., one of thespecies with relatively lower silica concentrations in their study.These results further imply that the effect of herbivory on phytolithformation is more common in species with relatively greater silicaaccumulation capabilities. We therefore hypothesise that if
Table 1Site descriptions (see map in Fig. 1). (a) Principal study sites. Mean annual rainfall was interpolated from nearby Israeli Meteorological Service stations (http://www.ims.gov.il): for Har Tsiv’on from Har-Kna’anand Mitspe-Harashim stations and for Sayeret Shaked Park from Be’er-Sheva station. Ramat-Hanadiv has its own station (http://www.ramatahanadiv-edu.org.il/weather). (b) Soil type and silica availability in the four principal sites, as well as in the sites in which plants were sampled to compare reproductiveand vegetative parts. The number of samples per soil type is five.
(a)
Site Year Har Tsiv’on Ramat-Hanadiv Sayeret Shaked Mitspe-Ramon
Location 33�100 N, 35�150 E 32�300 N, 34�570 E 31�150 N, 34�390 E 30�360 N, 34�450 EElevation (m above sea level) 850 120 200 800Mean annual rainfall (mm) �900 533 206 �80Rainfall in sampling year (mm) 2010 180 <80
2011 1402012 740 530 1402013 510 180
Studied grazing herbivores Cattle Goat/sheep Goat/sheep Goat/sheep
(b)
Site Soil type FAO classification Soil silica availability (mg Si g�1 soil, �SD)
Har Tsiv’on Terra Rossa Eutric/Vetric Cambisol 0.557 (�0.063)Ramat-Hanadiv Terra Rossa Eutric/Vetric Cambisol 0.701 (�0.090)Neve-Yam Nazaz Eutric Planosol 0.253 (�0.120)Binyamina Hamra Chromic Luvisol 0.116 (�0.015)Caesarea Sand dune Albic Arenosol 0.026 (�0.005)Sayeret Shaked Loess Calcaric Fluvisol 0.162 (�0.055)Mitspe-Ramon Loess Calcaric Fluvisol 0.179 (�0.038)
350 O. Katz et al. / Flora 209 (2014) 349–358

Author's personal copy
phytoliths play a significant antiherbivory role in Asteraceaespecies, grazed plants will have higher phytolith concentrationsthan un-grazed plants of the same species growing underotherwise similar natural conditions.
Furthermore, we consider the possibility that under resource-limited conditions plants allocate more resources to antiherbivorydefences and have more constitutive defences than inducible ones(Coley et al., 1985; Endara and Coley, 2011). The rationale is thatunder these conditions plants are limited in their response todefoliation (by re-growth, production of defences, etc.) and shouldthus be better defended in advance. We therefore hypothesise thatif phytoliths play an antiherbivory role in the Asteraceae, phytolithconcentrations in plants growing in grazed plots will be higherthan in un-grazed plots mostly in more humid sites, while in morearid sites the difference between grazed and un-grazed plots willbe smaller because under limited resources defences should bemore constitutive than inducible (e.g., Endara and Coley, 2011).
Increased defence of reproductive organs
In accordance with optimal defence hypotheses, plants, especial-ly herbaceous ones, are expected to allocate more defences toreproductive organs, which are of greater value to them than otherorgans (e.g., Meldau et al., 2012; but see McCall and Fordyce, 2010).Grubb (1992) noted that many tropical species with a single apicalmeristem are spiny. Ronel and Lev-Yadun (2012) found that in theflora of Israel, the reproductive organs of herbaceous species are themost frequent spiny or prickly plant parts. This phenomenon is alsoprominent in the southwest Asian Asteraceae (Ronel and Lev-Yadun,2012; Ronel et al., 2009). Phytoliths are thus thought to follow thesame patterns. Tsartsidou et al. (2007) found higher phytolithconcentrations in the inflorescences (compared to leaves and stems)of Poaceae and other monocotyledonous species in Greece. Somespecies, mostly those with relatively low phytolith concentrationsand including three out of the four studied woody species, did notfollow this trend (Tsartsidou et al., 2007). Therefore, we hypothesisethat if phytoliths play an antiherbivory role in the Asteraceae,phytolith concentrations and absolute phytolith numbers will behigher in inflorescences than in the rest of the shoot.
Materials and methods
Study areas
Israel, like most countries of southwest Asia, is characterised bya very long history of intense grazing (Tchernov, 1979; Zohary,1983) and by large rainfall gradients (Joseph and Ganor, 1986), andis thus a most suitable natural laboratory for this kind of study. Wefocused on seven sites which represent four spots along the largerainfall gradient of Israel. The four principal study sites in whichplants were collected in both un-grazed and grazed plots wereMitspe-Ramon and Sayeret Shaked Park in the arid southern NegevDesert and Ramat-Hanadiv and Har Tsiv’on in the northern andmore humid Mediterranean district (Table 1; Fig. 1). For thepurpose of studying differences between reproductive andvegetative organs, plants were also collected in three additionalsites near Ramat-Hanadiv in the north (Neve-Yam, Binyamina andCaesarea), which differ from Ramat-Hanadiv in soil type and inwhich herbivory is not controlled (Table 1; Fig. 1).
The southern and northern sites represent (respectively) theBShs and Csa climate regions, according to Köppen’s classification,and the desert and humid subtropical climates in Israel, accordingto a modified Budyko-Lettau dryness index (Joseph and Ganor,1986). Since silica availability in the soil (Katz et al., 2013) and itsuptake by plants (e.g., Rafi and Epstein, 1999; Morikawa andSaigusa, 2004) vary during the year, sampling was conducted in the
spring (March–May) of four consecutive years (2010–2013), in eachsite and year according to the specific seasonal rainfall and plantgrowth and flowering, in order to sample all plants in as mature a
Fig. 1. A map of Israel, indicating the locations of Har Tsiv’on (1), Neve-Yam (2),Ramat-Hanadiv (3), Binyamina (4), Caesarea (5), Sayeret Shaked Park (6), andMitspe-Ramon (7). Isohyets represent 1981–2010 means from the IsraeliMeteorological Service website (http://www.ims.gov.il).
O. Katz et al. / Flora 209 (2014) 349–358 351

Author's personal copy
state as possible (i.e., after being given enough time to form asmany phytoliths as possible).
The southernmost and most arid (80 mm mean annual rainfall)site is located near Mitspe-Ramon (Table 1), consisting of anecotourism site and a nearby grazed, wide, dry ephemeralstreambed (wadi). We acknowledge that the choice of wadi plotsmay suggest higher water availability compared to typicalmoderate slopes, but the vegetation cover and assemblage onthe slopes was too poor to allow adequate sample size. Plants inthis site were sampled in 2010, following a winter with less than80 mm rainfall, which was, however, the only year during the studyperiod (four years) with sufficient germination and plant growththere. The second southern site is the long-term ecologicalresearch (LTER) site Sayeret Shaked Park near Be’er-Sheva(206 mm mean annual rainfall; Table 1). Plants were sampled inSayeret Shaked Park in all four study years (2010–2013).
The principal Mediterranean district sites were the LTER sitesRamat-Hanadiv (530 mm mean annual rainfall) and Har Tsiv’on(900 mm mean annual rainfall; Table 1). Plants in Har Tsiv’on weresampled in 2012 following 740 mm of rainfall. Plants in Ramat-Hanadiv were sampled in 2012; and 2013 following 530 and510 mm of rainfall, respectively. Plants in the three additional sites(Neve-Yam, Binyamina and Caesarea) were sampled in 2013. Theydo not differ from Ramat-Hanadiv in terms of rainfall, but havedifferent soil types (Table 1b) and elevation (10–30 m above meansea level compared to 120 m in Ramat-Hanadiv) and were thusincluded only in that part of study which discusses the differencesin phytolith concentrations between reproductive and vegetativeplant parts, in which specific site traits are of lesser importance andno comparison between sites was involved.
Since the vegetation in the two southern arid (desert) sites is amixture of herbaceous species and dwarf shrubs, while in thenorthern (more humid Mediterranean) sites it is composed of alllife forms, dominated by shrubs and trees, we were concerned thatany comparison between the two site types may be biased.Therefore, plant sampling in the more humid northern Mediterra-nean sites was conducted in deforested plots which are nowdominated by forbs and dwarf shrubs. For the same reason, wehave also put more emphasis on sampling herbaceous species(Table 2). Soil silica availability varies largely between sites,depending mostly on soil type rather than rainfall (Table 1b; Katzet al., 2013), but non-Poaceae species are less sensitive than
Poaceae species to silica availability (Cooke and Leishman, 2012;Katz et al., 2013).
The effect of grazing
We focussed on grazing by medium and large mammalianherbivores (mainly goats and cattle), which are known to benegatively affected by high silica concentrations (Massey andHartley, 2006) and to prefer silica-low plants (Hunt et al., 2008;Massey et al., 2009). We preferred to focus on these herbivoressince they are more easily controlled in the field (i.e., by simplefencing), while excluding smaller herbivores (such as birds,rodents, insects or molluscs) from large areas is more complicatedtechnically and the means used to exclude them (such as nets) mayhave other more profound influences on the ecosystem (e.g., onsun irradiation, temperature, evaporation, pollination and decom-position). This was especially important in the southern sites,where vegetation cover is low and a large area was needed tocollect enough biomass.
In these two desert sites (Mitspe-Ramon and Sayeret ShakedPark), grazing is performed mostly by herds of goats and sheep,commonly accompanied by a few donkeys and camels. Wildgazelles also occasionally graze near the two sites, as well as ibexesnear Mitspe-Ramon. Accurate data concerning grazing intensity inboth sites are unavailable, due to the opportunistic nature oftraditional Bedouin herding and the sporadic activity of the wildgazelles and ibexes. However, the un-grazed and grazed plots usedin this study represent two extremes, and the effects of grazing onphytolith formation are probably independent of herbivore density(Soininen et al., 2013). Grazed plots were designated in locations inwhich herding was observed, usually within a few hundred metersof ephemeral encampments of the Bedouin herdsmen, and weretypically littered by large amounts of dung pellets. The un-grazedplots were fenced so the local herds did not enter them, and dungpellets were not observed there. In Ramat-Hanadiv we preferred tosample plants in plots grazed by goats rather than by cattle sincegoats are common in the two southern desert sites, but in andaround Har Tsiv’on cattle are the primary medium/large mamma-lian herbivores.
Inaddition, anannual grassspecies (Avena sterilis L.) was sampledin order to mediate between our naturally growing Asteraceae andthe various previously studied grasses. This species is not found
Table 2The studied species: life forms (Ann: Annual, Hem- Hemicryptophyte, Dsh: Dwarf shrub) and chorotypes (MT: Mediterranean; IT: Irano-Turanian; SA: Saharo-Arabian).
Species Life form Spinescence Chorotype
Anthemis spp. (including pseudocotula) Ann � MT-IT-SAAtractylis serratuloides Boiss. DSh + SACalendula arvensis L. Ann � MT-ITCarthamus tenuis (Boiss. et Blanche) Bornm. Ann + MTCentaurea hyalolepis Boiss. Ann + MT-ITCentaurea iberica Spreng. Ann + MT-ITChrysanthemum coronarium L. Ann � MTCrepis reuteriana Boiss. Hem � MTCrupina crupinastrum (Moris.) Vis. Ann � MT-ITEchinops adenocaulos Boiss. Hem + MTEchinops polyceras Boiss. Hem + ITHelichrysum sanguineum (L.) Kostel. Hem � MTLeontodon laciniatum (Bertol.) Widd. Ann � IT-SANotobasis syriaca (L.) Cass. Ann + MTPallenis spinosa (L.) Cass. Hem + MTPicris damascena Boiss. et Gaill Ann � ITScolymus maculatus L. Ann + MTScorzonera papposa DC. Hem � ITSenecio glaucus L. Ann � IT-SASilybum marianum (L.) Gaertn. Ann + MT-ITUrospermum picroides (L.) F.W. Schmidt Ann � MT-IT
352 O. Katz et al. / Flora 209 (2014) 349–358

Author's personal copy
naturally inside the Sayeret Shaked Park site and was thereforesampled in a nearby un-grazed area. It also occurs in Mitspe-Ramonbut was not sampled there due to its very small numbers.
Increased defence of reproductive organs
In order to test our hypothesis of formation of more phytolithsin reproductive organs, we sampled mature plants from eightspecies (Anthemis pseudocotula, Carthamus tenuis, Chrysanthemumcoronarium, Echinops adenocaulos, E. polyceras, Notobasis syriaca,Pallenis spinosa and Scolymus maculatus; Table 2) in order toanalyse inflorescences separately from the rest of the shoot. Plantswere sampled in both Sayeret Shaked Park and Ramat-Hanadiv, butsince sampling was conducted relatively late in the season (May2013) in order to have seed-carrying plants, some species were notfound in sufficient amounts in Ramat-Hanadiv and were thussampled in nearby Caesarea, Neve-Yam and Binyamina sites, whichdiffer in soil types (Table 1b). We acknowledge that there aremorphological differences between stems and leaves, which alsocause differences in phytolith concentrations between these twovegetative organs (e.g., Tsartsidou et al., 2007). However, as thisstudy focuses on difference between reproductive and vegetativeorgans, we did not separate leaves from stems.
Sampling procedures
Whenever possible, the study focused on annual species, whoseturnover is faster and therefore population-level processes are
expected to be more rapid. Following previous studies (Albert et al.,1999, 2008; Tsartsidou et al., 2007), each sample consisted of aminimum dry weight of 3 g. Since in most cases the weight of asingle plant is lower, phytolith extraction and quantification fromeach individual separately was in most cases undesired or evenimpossible. Instead of having an inaccurate measurement fromeach individual, we preferred to group all individuals of a similarcontext (species, site, plot and year) into a single sample whichallowed a more accurate measurement (although losing data onintra-population variability).
Plots in the different sites were notof similar sizes, andtherewerealso great differences in vegetation cover between arid andMediterranean sites. Therefore, the numbers and sizes of sampledplots varied. In each site and plot type, the total area in which plantswere sampled was at least 4000 m2. This had little effect on the studydesign and statistical analyses, since the criteria for sample size werethe minimal requirementsof plant weight and numberof individuals(as described above) and hence independent of plot size.
Plants which were representative of the average and range ofsizes within the population were cut in their bases, 0–1 cm abovethe soil surface, so only above-ground parts (including flowers andseeds) were analysed. For the study of the grazing effects, care wastaken to sample species which are known to be grazed by sheep,goats and cattle. Twenty-one pairs of un-grazed and grazed plantsamples from the same species and site, belonging to 16 species,were sampled (Table 3; Fig. 2). Field observations were madeeither directly (i.e., observing actual grazing) or indirectly (i.e.,observing bite marks) to ascertain that these species are in fact
Table 3Phytolith concentrations (phytoliths g�1 dry matter) in various species under un-grazed and grazed conditions, the grazing effect (Eg) and its significance (P). Number of sub-replicates is four, unless stated otherwise.
Species Year Spinescence Phytolith g�1 dry matter Eg (%) P
Un-grazed plants Grazed plants
(a) Har Tsiv’onAvena sterilis (Poaceae) � 910,000 (�24.8%) 740,000 (�15.5%) �18.5 0.4857Centaurea iberica + 940 (�29.9%) 1200 (�32.8%) +24.9 0.4857Crepis reuteriana � 12 (�24.6%) 32 (�40.6%) +158.2 0.0286Helichrysum sanguineum � 26 (�80.2%) 150 (�44.5%) +476.7 0.0286
(b) Ramat-HanadivAvena sterilis (Poaceae) � 840,000 (�4.2%) 940,000 (�6.9%) +11.8 0.1143Anthemis spp. � 19,000 (�16.4%) 20,000 (�11.5%) +8.1 0.8857Crupina crupinastrum � 25,000 (�12.8%) 6100 (�38.1%) �75.7 0.0286Pallenis spinosa + 3800 (�8.8%) 2000 (�10.2%) �48.5 0.3429Silybum marianum + 20 (�35.7%) 1600 (�17.8%) +7379.3 0.0286Urospermum picroides � 7900 (�62.1%) 8100 (�14.8%) +1.8 1.0000
(c) Sayeret Shaked ParkAnthemis spp. 2010 � 6200 (�7.9%)a 5900 (�47.0%) �6.0 0.8571Anthemis spp. 2011 � 890 (�89.7%) 400 (�49.2%) �55.5 1.0000Atractylis serratuloides 2012 + 22,000 (�16.3%) 30 (�46.7%) �99.9 0.0286Centaurea hyalolepis 2011 � 800 (�22.8%) 720 (�9.6%) �9.5 0.8857Chrysanthemum coronarium 2011 � 1100 (�45.6%) 220 (�14.2%) �79.9 0.0286Chrysanthemum coronarium 2012 � 240 (�39.5%) 60 (�82.5%) �73.5 0.0571Leontodon laciniatum 2011 � 40 (�84.8%) 70 (�47.0%) +60.8 0.3429Senecio glaucus 2010 � 30 (�44.8%) 4 (�41.9%) �86.2 0.0286
(d) Mitspe-RamonAnthemis spp. – 10,000 (�24.6%) 4800 (�24.3%) �52.3 0.0286Picris damascena – 1400 (�15.0%) 970 (�29.9%) �28.3 0.2Senecio glaucus – 90 (�28.4%) 100 (�63.4%) +7.6 0.6875Scorzonera papposa – 470 (�50.3%) 1100 (�13.3%) +124.8 0.0606Calendula arvensis – 120 (�23.9%) 60(�14.2%) �51.0 0.0286
a Only three microscope slides were available.
O. Katz et al. / Flora 209 (2014) 349–358 353
Author's personal copy
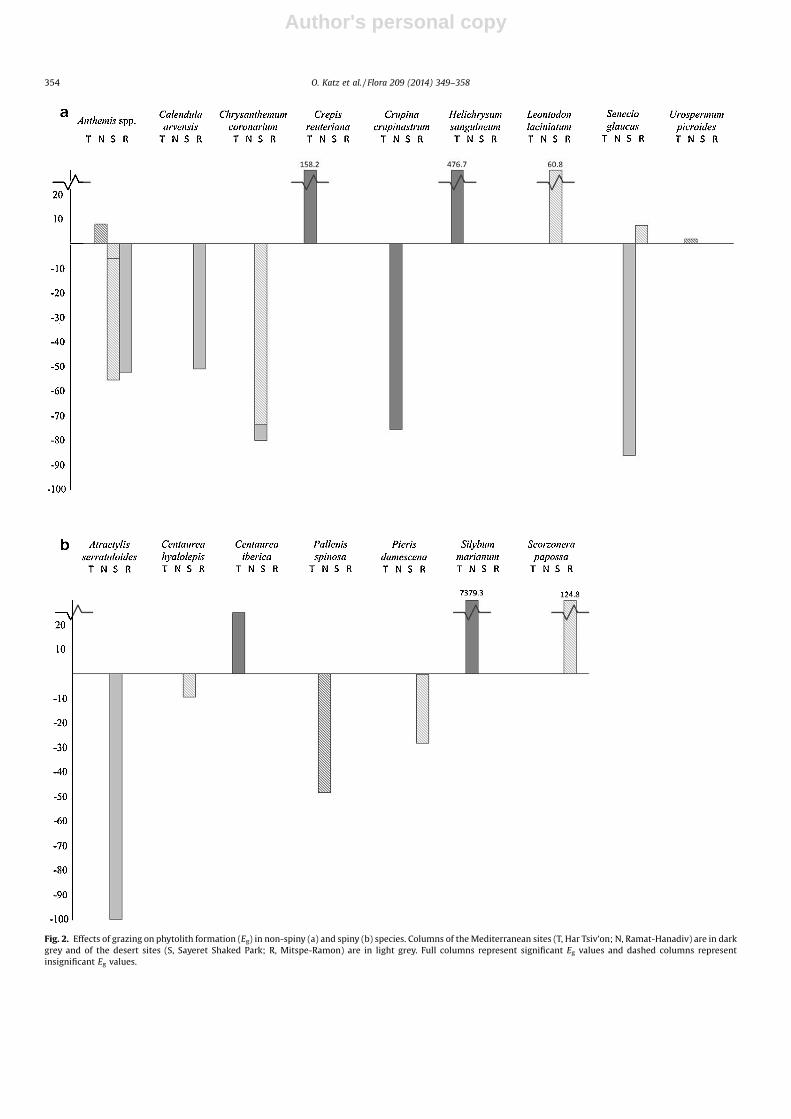
Fig. 2. Effects of grazing on phytolith formation (Eg) in non-spiny (a) and spiny (b) species. Columns of the Mediterranean sites (T, Har Tsiv’on; N, Ramat-Hanadiv) are in darkgrey and of the desert sites (S, Sayeret Shaked Park; R, Mitspe-Ramon) are in light grey. Full columns represent significant Eg values and dashed columns representinsignificant Eg values.
354 O. Katz et al. / Flora 209 (2014) 349–358

Author's personal copy
grazed in the study sites. For the study of the differences betweenreproductive and vegetative parts, four individuals were sampledfrom each species and placed in four separate bags, to insure thatinflorescences of different individuals are not mixed.
Laboratory procedures and data analysis
Phytolith extraction and quantification followed Albert et al.(1999) procedure with some modifications following its more recentapplications (Albert et al., 2008; Katz et al., 2007, 2010; Shahack-Gross et al., 2003; Tsartsidou et al., 2007). This method has beenshown to best preserve phytolith assemblages (Jenkins, 2009; Katzet al., 2010). All samples were initially washed in distilled water untilall apparentsoilwasremovedanddriedat105 �Cforat least12 hours.In the case of the samples used for the study of differences betweenreproductive and vegetative parts, the dried plants were cut toseparate the inflorescences from the rest of the shoot. Then, thebiomass of the inflorescences and shoots of each individual wereweighedinordertocalculatethe inflorescenceweightpercentageforeach individual. We then grouped all inflorescences of the samespecies into one sample, and did the same for the rest of the shoot, toconform to our minimum weight criterion. The samples used for thegrazing effect study were simply dried and weighed as a single unit,without regard to different plant parts or different individuals.
The weighed dry matter was burnt at 500 �C for 4 h to removeorganicmatter.Theashwasthendissolvedin3 NHCl + HNO3solutiontoremovecarbonate andphosphatecompounds,rinsedand dried(asdescribed above). Remnant organic and/or charred matter wasremovedbyburningat500 �Cforanadditionalhour,andsoilmineralswere removed by separation in a 2.3 g ml�1 sodium-polytungstate(Na6(H2W12O40)�H2O) solution (these last two steps were notrequired for A. sterilis). The dried final extract was also weighed.
Four aliquots of 1–2 mg each were weighed from the finalextract of each sample and mounted on four separate microscopeslides, taking care to mix the phytoliths in the mounting medium(Entellan New, Merck) to produce a slide as homogenous aspossible. The use of four slides has proved important since manyslides contained much less than 500 phytoliths, which isconsidered a minimum number to arrive at an error level lowerthan 30% (Katz et al., 2010). However, when the number ofphytoliths on the slide exceeded 500 by much, only 20% of the slidewas used (10 random lines of view out of a total of 50 on the slide,at �400 magnification), with the total number of countedphytoliths multiplied by five. This does not increase errorscompared to counting phytoliths on whole slides (Katz et al., 2010).
For each sample, the four slides were averaged and the coefficientof variation (CV) was calculated. Therefore, CVs reflect analyticalerrors rather than sampling errors or variation within each plot. Inthe study of phytolith concentrations in inflorescences compared toother plant parts, we have calculated the percentage of phytoliths inthe inflorescences (relative to the whole plant) as follows:
pf ¼ðPfwfÞ
ðPfwf þ Psð100% � wfÞÞ� 100%
where pf is the percentage of phytoliths in inflorescences, Pf and Psare the phytolith concentration in the inflorescences and the rest ofthe shoot (respectively) and wf is the inflorescences weightpercentage.
The grazing effect for each species in a given site and year (Eg)was calculated as:
Eg ¼ ðPg � PcÞPc
� 100%
where Pc and Pg are phytolith concentrations in un-grazed andgrazed plants, respectively. Thus, positive Eg values represent
higher phytolith concentrations under grazing (as observed inmany grasses) while negative Eg values represent lower phytolithconcentrations under grazing.
As mentioned above, sites differed in plot size and number andin vegetation cover, and the minimum weight requirementsfurther limited the number of individuals per sample. Small-sizedand abundant species required a very large number of individuals,which could not have been replicated for larger-sized and lessabundant species. The dry weight of samples ranged between 3and 30 g, with the lower end more typical of small-sized species (aminimum weight constraint) and the higher end more typical oflarge-sized species (a minimal number of individuals constraint).Our data thus reflect phytolith concentrations per biomass, ratherthan per individual, population or area. We therefore employed thenon-parametric Mann–Whitney test, and the reported P values forcomparisons between samples refer to four slides (sub-replicates)per sample, unless stated otherwise. All the statistical work wasconducted using PAST software (version 2.12; Hammer et al.,2001).
Soil silica availability
Five soil samples were taken in each site to determine silicaavailability (following Jenkins et al., 2011, with some modifica-tions). Samples were taken in spring at depths of 1–15 cm. The top0.5–1 cm was removed to avoid misrepresentations due toaccumulations of organic matter and litter. For each sample, 3 gof 2 mm-sieved soil were dissolved in 30 ml of 25 mM citric acid.The solution was shaken at 100 rpm for 6 h at 30 �C and centrifugedat 3000 rpm for 15 min. The supernatant was then filtered(Whatman paper No. 1) and stored at 4 �C till analysis. Wanget al. (2004) have demonstrated that citric acid is one of the mostefficient extractants of plant-available silica from soils. Determi-nation of silicon availability in the soil extracts was donecolorimetrically, using a standard kit (Merck #100,857: molybdateions in sulfuric acid) and a Merck Spectroquant Pharo100spectrophotometer. Previous experience shows that there is agood correlation between ICP (used by Jenkins et al., 2011) andcolorimetric determinations of citric acid-extracted silica (Wanget al., 2004).
Results
Comparisons of pairs of un-grazed and grazed plants of thesame species in the same site gave mixed results (Table 3; Fig. 2).Within the non-spiny species (16 pairs from 11 species), cases ofpositive Eg (i.e., higher phytolith concentrations in grazedcompared to un-grazed plants of the same species) were foundin seven pairs (two of which were statistically significant using theMann–Whitney test, four pseudo-replications per sample) andcases of negative Eg were found in nine pairs (five cases weresignificant). Within pairs of un-grazed and grazed plants fromspiny species (five pairs from five species), positive Eg was foundtwice (one case was significant) and negative Eg three times (onecase was significant). In total (21 pairs from 15 species) there werenine cases of positive Eg (three cases were significant) compared to12 cases of negative Eg (six cases were significant).
As a whole, the Asteraceae species which we studied tended notto have either positive or negative Eg values (P = 0.2157 usingWilcoxon’s signed-ranks test), but the annual non-spiny species asa group showed a marginally significant tendency towardsnegative Eg values (nine out of 13 pairs, P = 0.0804). Eg valuesabove +20% (or otherwise significantly positive) were found only inspecies in which the phytolith concentrations were relatively lowcompared to many other Asteraceae species. Eg was not clearlydependent on spinescence, although significant and/or negative Eg
O. Katz et al. / Flora 209 (2014) 349–358 355

Author's personal copy
values were more common in non-spiny species. Therefore,although evidence for a positive effect of grazing on phytolithformation exists, in 12 out of 21 pairs of un-grazed and grazedplants occurring in nine of 16 species (six of which are non-spinyspecies) we found that grazed plants had lower phytolithconcentrations than their un-grazed counterparts.
Cases of negative Eg were more common in the southern aridsites (Table 3). Out of 12 cases of both significant and insignificantnegative Eg, 10 occurred in plants from Mitspe-Ramon and SayeretShaked Park, the two arid southern sites (Anthemis spp., Crysan-themum coronarium, Calendula arvensis, Senecio glaucus, Centaureahyalolepis, Atractylis serratuloides and Picris damascena), comparedto only two cases in the Mediterranean northern Har Tsiv’on andRamat-Hanadiv (Crupina crupinastrum and Pallenis spinosa). Out ofthe six cases of significantly negative Eg, three were in the aridsouthern Mitspe-Ramon and Sayeret Shaked Park (C. coronarium in2011, but not in 2012, in Sayeret Shaked Park, Anthemis spp. and C.arvensis in Mitspe-Ramon). Out of the nine cases of significant andinsignificant positive Eg, five were in the Mediterranean northernHar Tsiv’on and Ramat-Hanadiv (Anthemis spp., Centaurea iberica,Crepis reuteriana, Helichrysum sanguineum and Silybum marianum),including all three significant cases (C. reuteriana, H. sanguineum, S.marianum). The reference grass species A. sterilis from the twoMediterranean sites had a positive Eg in Ramat-Hanadiv and anegative Eg in Har Tsiv’on, both insignificant.
Contingency tables for the four sites were analysed in two sets.First, we divided the cases into four categories (significant positiveEg, insignificant positive Eg, insignificant negative Eg and significantnegative Eg) and found insignificant (P = 0.1730 using a x2 test) andsignificant (P = 0.0184) tendencies towards more significantlynegative Eg values as sites were more arid, for all species andnon-spiny species only, respectively. We then used only twocategories (positive vs. negative Eg) and found a marginallysignificant (P = 0.0544) and insignificant (P = 0.1697) tendenciestowards having more cases of negative Eg values as sites were morearid, for all species and non-spiny species only, respectively. Thus,Asteraceae plants in the more arid southern sites tend to formfewer phytoliths when exposed to grazing compared to plants fromthe same species from Mediterranean northern sites.
Six species (the non-spiny A. pseudocotula and the spiny C.tenuis, E. adenocaulos, E. polyceras, P. spinosa and S. maculatus) werefound to have lower phytolith concentrations in their inflores-cences than their vegetative parts (Table 4). For instance, theinflorescences of C. tenuis comprised 30–40% of the dry plantbiomass, but less than 5% of the plant’s phytoliths. Two species (thenon-spiny C. coronarium and the very spiny N. syriaca) had higherphytolith concentrations in their inflorescences (Table 4).
Discussion
There seems to be no clear general correlation between grazingand higher phytolith concentrations in the studied Asteraceaespecies, as both significantly negative and positive Eg were found.Negative Eg values (i.e., higher phytolith concentrations in un-grazed plants) were found in a total of 12 out of 21 pairs of un-grazedand grazed plants from 15 out of 16 species (nine pairs were fromeight out of the 12 non-spiny species). Positive Eg values were foundin nine out of 21 pairs from eight out of 16 species (five pairs werefrom seven out of the 12 non-spiny species) and were significantthree times (all in spiny species). Positive Eg values (i.e., higherphytolith concentrations in grazed plants) were common in specieswith relatively low phytolith concentrations, and these smallnumbers of phytoliths suggest that they have little if any actualantiherbivory value for the plant. Within the annual non-spinyspecies, it is evident that there were more negative Eg (nine out of 13pairs; P = 0.0804). Therefore, the studied Asteraceae species do nottend as a group to form more phytoliths when exposed to grazing innature, in contrast to previous finds from various Poaceae species inboth laboratory experiments and field studies (Brizuela et al.,1986;Cid et al.,1989; Masseyet al., 2008; McNaughton and Tarrants,1983;McNaughton et al., 1985). Thus, our hypothesis of higher phytolithconcentrations in grazed Asteraceae plants should be rejected forthe studied species as a group.
According to our results, herbivory by medium and largemammalian herbivores does not affect phytolith formation intypical southwest Asian members of the Asteraceae and also doesnot select for phytolith-rich individuals in the grass A. sterilis inthe Mediterranean sites. It is already known that the effects ofgrazing on Poaceae species vary among species (Garbuzov et al.,2011; Melzer et al., 2010; Soininen et al., 2013) and that mediumand large mammalian herbivores may have a smaller effect onphytolith formation compared to smaller mammalian andinvertebrate herbivores (Massey et al., 2007b; Soininen et al.,2013). However, while no or negative effect of herbivory onphytolith formation is relatively uncommon in the previousstudies of Poaceae species, it is very common in our study ofAsteraceae species. Alternatively, the long and intensive grazinghistory of southwest Asia, as opposed to northern Europe, wheresome of the previous studies (Garbuzov et al., 2011; Massey et al.,2007b; Soininen et al., 2013) were conducted, may have providedsufficient evolutionary time and selection pressure for naturalselection to operate in favour of relatively silica-rich Poaceaeindividuals, thus maximising phytolith concentrations in variousPoaceae species regardless of the actual grazing history of currentindividuals. Notwithstanding, it is clear that Eg values in many of
Table 4Phytolith concentrations in inflorescences compared to the rest of the shoot in selected species, and the percentages of inflorescence biomass and phytoliths relative to thewhole shoot. Number of sub-replicates is four in all samples. All differences in phytolith concentrations between inflorescences and the rest of the shoot are significant(P = 0.0286).
Species Site Phytolith g�1 dry matter % contribution of inflorescences
Inflorescences Rest of shoot Plant biomass Plant phytoliths
Anthemis pseudocotula Binyamina 8 (�105.2%) 5000 (�22.0%) 38.6 <0.1Carthamus tenuis Ramat-Hanadiv 750 (�14.7%) 7300 (�7.7%) 36.2 4.7Carthamus tenuis Sayeret Shaked 120 (�25.0%) 3500 (�23.1%) 30.7 1.4Chrysanthemum coronarium Caesarea 3500 (�37.1%) 1300 (�15.4%) 44.8 71.9Chrysanthemum coronarium Sayeret Shaked 3100 (�18.4%) 220 (�72.1%) 36.0 98.2Echinops adenocaulos Ramat-Hanadiv 7 (�129.0%) 22,000 (�16.0%) 40.3 <0.1Echinops polyceras Sayeret Shaked 860 (�20.7%) 9800 (�22.7%) 19.0 1.9Notobasis syriaca Neve-Yam 940 (�20.2%) 150 (�44.3%) 32.7 74.5Pallenis spinosa Ramat-Hanadiv 710 (�21.2%) 3200 (�9.2%) 47.8 16.1Scolymus maculatus Ramat-Hanadiv 41,000 (�13.3%) 88,000 (�17.6%) 50.9 31.7
356 O. Katz et al. / Flora 209 (2014) 349–358

Author's personal copy
the studied Asteraceae species are more negative and moresignificant than in A. sterilis.
Significant negative Eg values are more common in theplants from arid southern sites in which we sampled and arealso more negative when compared to plants growing in theMediterranean northern sites we studied. There is also noevidence for a general pattern of higher phytolith concentrationsin plants growing in the arid southern sites (Katz et al., 2013).The lack of tendencies towards having higher phytolithconcentrations and stronger induction of phytolith formationin more arid sites does not fit the predictions of the resourceavailability hypothesis (Coley et al., 1985; Endara and Coley,2011), and further does not support a defensive function ofphytoliths in the Asteraceae.
Significant negative Eg values occur in Asteraceae species withboth relatively high and relatively low phytolith concentrations,but significant positive Eg values were found only in species withvery low phytolith concentrations. These results suggest that thereis little correlation between phytolith concentrations and grazingeffect in the southwest Asian Asteraceae species. Nevertheless, ourfindings suggest that there is a difference in the grazing effectbetween Poaceae and Asteraceae species, the former probablytending towards a positive effect and the latter towards a negativeeffect. Therefore, if a threshold for the effect of grazing onphytolith formation exists, then it possibly lies somewherebetween the phytolith concentration ranges of these two plantfamilies.
Only two species had higher phytolith concentrations andabsolute phytolith numbers in the inflorescences compared to therest of the shoot (C. coronarium and N. syriaca), while in the othersix species which were studied concerning this characterA. pseudocotula, C. tenuis, E. adenocaulos, E. polyceras, P. spinosaand S. maculatus) there were much lower phytolith concentrationsand absolute phytolith numbers in the inflorescences compared tothe rest of the shoot (Table 4). Notably, C. coronarium and C. tenuisare not phyletically close to each other within the Asteraceae, buteach of them is closer to some of the other six species (Bremer,2004). Therefore, we reject the hypothesis that Asteraceae speciesas a group produce more phytoliths in association with theirreproductive organs.
We did not find that formation of phytoliths in reproductiveorgans is dependent on whole-plant phytolith concentrations.This does not fit previous observations in Poaceae and variousother monocotyledonous species (Tsartsidou et al., 2007), furtherdemonstrating the difference between the Poaceae and Aster-aceae families. This could suggest that Poaceae species have amore pronounced tendency to form more phytoliths in theirreproductive organs, if it were not for the two Asteraceae specieswhich clearly also form more phytoliths in their reproductiveorgans.
In conclusion, we find that the southwest Asian Asteraceae as agroup do not form more phytoliths when exposed to grazing,when they grow under more resource-limited conditions or inassociation with their reproductive organs. Hence, variability inphytolith concentrations in the studied southwest Asian Aster-aceae species cannot be unequivocally related to defensive roles,except for a few specific species, and point to a smaller defensivepotential of phytoliths in the Asteraceae compared to thesituation in Poaceae. The differences in the defensive role ofphytoliths between Poaceae and Asteraceae species can probablybe explained by the better chemical (Zdero and Bohlmann, 1990)and physical (Ronel et al., 2009) defences of the regionalmembers of the Asteraceae compared to the Poaceae (e.g.,Prychid et al., 2004). It is therefore difficult to determine at whatphytolith concentration ranges the antiherbivory function orresponse to grazing becomes significant.
Acknowledgments
Dr Itzhak Katra and Ester Shani (BGU) permitted the use oflaboratory facilities. Prof. Roni Shneck (BGU) generously allowedthe use of his furnace and Yehoshua Ratzon (BGU) suppliedtechnical assistance. Permissions to sample were granted by DrBertrand Boeken (BGU) in Sayeret Shaked Park, Dr Liat Hadar(Ramat-Hanadiv) in Ramat-Hanadiv, and Prof. Gidi Ne’eman andHar’el Agra (University of Haifa) in Har Tsiv’on. Ben Drori (Natureand Parks Authority) assisted in locating sampling sites. Dr RachelSchwartz-Tzachor (Ramat-Hanadiv), Alvit Nachshon and GiliShimerling assisted in the fieldwork. Maoz Dor assisted in soilsampling and analyses. This study is supported by a grant from theIsrael Science Foundation (103/11).
References
Albert, R.M., Lavi, O., Estroff, L., Weiner, S., Tsatskin, A., Ronen, A., Lev-Yadun, S.,1999. Mode of occupation of Tabun Cave, Mt Carmel, Israel during theMousterian Period: a study of the sediments and phytoliths. J. Archaeol. Sci. 26,1249–1260.
Albert, R.M., 2008. Phytolith-rich layers from the Late Bronze and Iron Ages at TelDor (Israel): mode of formation and archaeological significance. J. Archaeol. Sci.35, 57–75.
Bremer, K.,1994. Asteraceae: Cladistics and Classification. Timber Press, Portland, USA.Brizuela, M.A., Detling, J.K., Cid, M.S.,1986. Silicon concentration of grasses growing in
sites with different grazing histories. Ecology 67, 1098–1101.Cid, M.S., Detling, J.K., Brizuela, M.A., Whicker, A.D., 1989. Patterns in grass
silicification: responses to grazing history and defoliation. Oecologia 80,268–271.
Coley, P.D., Bryant, J.P., Chapin III, F.S., 1985. Resource availability and plantantiherbivore defense. Science 230, 895–899.
Cooke, J., Leishman, M.R., 2011. Is plant ecology more siliceous than we realize?Trends Plant Sci. 16, 61–68.
Cooke, J., Leishman, M.R., 2012. Tradeoffs between foliar silicon and carbon-baseddefences: evidence from vegetation communities of contrasting soil types.Oikos 121, 2052–2060.
Endara, M.J., Coley, P.D., 2011. The resource availability hypothesis revisited: a meta-analysis. Funct. Ecol. 25, 389–398.
Epstein, E., 1999. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50, 641–664.Feinbrun-Dothan, N., Danin, A., 1991. Analytical Flora of Eretz-Israel. Cana,
Jerusalem [in Hebrew].Garbuzov, M., Reidinger, S., Hartley, S.E., 2011. Interactive effects of plant-available
soil silicon and herbivory on competition between two grass species. Ann. Bot.108, 1355–1363.
Guntzer, F., Keller, C., Meunier, J.D., 2012. Benefits of silicon for crops: a review.Agron. Sustain Dev. 32, 201–213.
Grubb, P.J., 1992. A positive distrust in simplicity – lessons from plant defences andfrom competition among plants and among animals. J. Ecol. 80, 585–610.
Hammer, O., Harper, D.A.T., Ryan, P.D., 2001. PAST: paleontological statistics softwarepackage for education and data analysis. Palaeontol. Electron. 4 (1) 9 pp..
Hunt, J.W., Dean, A.P., Webster, R.E., Johnson, G.N., Ennos, A.R., 2008. A novelmechanism by which silica defends grasses against herbivory. Ann. Bot. 102,653–656.
Jenkins, E., 2009. Phytolith taphonomy: a comparison of dry ashing and acidextraction on the breakdown of conjoined phytoliths formed in Triticum durum.J. Archaeol. Sci. 36, 2402–2407.
Jenkins, E., Jamjoum, K., Nuimat, S., 2011. Irrigation and phytolith formation: anexperimental study. In: Mithen, S., Black, R. (Eds.), Water, Life and Civilisation:Climate, Environment and Society in the Jordan Valley. Cambridge UniversityPress, Cambridge, pp. 347–372.
Joseph, J.H., Ganor, E., 1986. Variability of climatic boundaries in Israel – use ofmodified Budyko–Lettau aridity index. J. Climatol. 6, 69–82.
Kaiser, T.M., Müller, D.W.H., Fortelius, M., Schulz, E., Cordon, D., Clauss, M., 2013.Hypsodonty and tooth facet development in relation to diet and habitat inherbivorous ungulates: implications for understanding tooth wear. MammalRev. 43, 34–46.
Katz, O., Gilead, I., Bar (Kutiel), P., Shahack-Gross, R., et al., 2007. et al., Chalcolithicagricultural life at the site of Grar, northern Negev, Israel: dry farmed cerealsand dung-fueled hearths. Paléorient 33, 101–116.
Katz, O., Cabanes, D., Weiner, S., Maeir, A., Boaretto, E., Shahack-Gross, R., 2010.Rapid phytolith extraction for analysis of phytolith concentrations andassemblages during an excavation: an application at Tell es-Safi/Gath.J. Archaeol. Sci. 37, 1557–1563.
Katz, O., Lev-Yadun, S., Bar (Kutiel), P., 2013. Plasticity and variability in the patternsof phytolith formation in Asteraceae species along a large rainfall gradient inIsrael. Flora 208, 438–444.
Kvedaras, O.L., An, M., Choi, Y.S., Gurr, G.M., 2010. Silicon enhances natural enemyattraction and biological control through induced plant defences. Bull. Entomol.Res. 100, 367–371.
O. Katz et al. / Flora 209 (2014) 349–358 357

Author's personal copy
Ma, J.F., 2004. Role of silicon in enhancing the resistance of plants to biotic andabiotic stresses. Soil Sci. Plant Nutr. 50, 11–18.
Ma, J.F., Yamaji, N., 2008. Function and transport of silicon in plants. Cell. Mol. LifeSci. 11, 392–397.
Massey, F.P., Hartley, S.E., 2006. Experimental demonstration of the antiherbivoreeffects of silica in grasses: impacts on foliage digestibility and vole growth rates.Proc. R. Soc. London B 273, 2299–2304.
Massey, F.P., Hartley, S.E., 2009. Physical defences wear you down: progressive andirreversible impacts of silica on insect herbivores. J. Anim. Ecol. 78, 281–291.
Massey, F.P., Ennos, A.R., Hartley, S.E., 2007a. Grasses and the resource availabilityhypothesis: the importance of silica-based defences. J. Ecol. 95, 414–424.
Massey, F.P., Ennos, A.R., Hartley, S.E., 2007b. Herbivore specific induction of silica-based defences. Oecologia 152, 677–683.
Massey, F.P., Smith, M.J., Lambin, X., Hartley, S.E., 2008. Are silica defences in grassesdriving vole population cycles? Biol. Lett. 4, 419–422.
Massey, F.P., Massey, K., Ennos, A.R., Hartley, S.E., 2009. Impacts of silica-baseddefenses in grasses on the feeding preferences of sheep. Basic Appl. Ecol. 10,622–630.
McCall, A.C., Fordyce, J.A., 2010. Can optimal defence theory be used to predict thedistribution of plant chemical defences? J. Ecol. 98, 985–992.
McNaughton, S.J., Tarrants, J.L., 1983. Grass leaf silicification: natural selection foran inducible defense against herbivory. Proc. Natl. Acad. Sci. U. S. A. 80,790–791.
McNaughton, S.J., Tarrants, J.L., McNaughton, M.M., Davis, R.H., 1985. Silica asdefense against herbivory and a growth promoter in African grasses. Ecology 66,528–535.
Meldau, S., Erb, M., Baldwin, I.T., 2012. Defence on demand: mechanisms behindoptimal defence patterns. Ann. Bot. 110, 1503–1514.
Melzer, S.E., Knapp, A.K., Kirkman, K.P., Smith, M.D., Blair, J.M., Kelly, E.F., 2010. Fireand grazing impacts on silica production and storage in grass dominatedecosystems. Biogeochemistry 97, 263–278.
Mihlbachler, M.C., Rivals, F., Solounias, N., Semprebon, G.M., 2011. Dietary changeand evolution of horses in North America. Science 331, 1178–1181.
Morikawa, C.K., Saigusa, M., 2004. Mineral composition and accumulation of siliconin tissues of blueberry (Vaccinum corymbosus cv. Bluecrop) cuttings. Plant Soil258, 1–8.
Piperno, D.R., 2006. Phytoliths: A Comprehensive Guide for Archaeologists andPaleoecologists. AltaMira Press, Oxford.
Prychid, C.J., Rudall, P.J., Gregory, M., 2004. Systematics and biology of silica inmonocotyledons. Bot. Rev. 69, 377–440.
Rafi, M.M., Epstein, E., 1999. Silicon absorption by wheat (Triticum aestivum L.). PlantSoil 211, 223–230.
Reynolds, O.L., Keeping, M.G., Meyer, J.H., 2009. Silicon-augmented resistance ofplants to herbivorous insects: a review. Ann. Appl. Biol. 155, 171–186.
Ronel, M., Lev-Yadun, S., 2012. The spiny, thorny and prickly plants in the flora ofIsrael. Bot. J. Linn. Soc. 168, 344–352.
Ronel, M., Khateeb, S., Lev-Yadun, S., 2009. Protective spiny modules in thistles ofthe Asteraceae in Israel. J. Torrey Bot. Soc. 136, 46–56.
Shahack-Gross, R., Marshall, F., Weiner, S., 2003. Geo-ethnoarchaeology of pastoralsites: the identification of livestock enclosures in abandoned Maasai settle-ments. J. Archaeol. Sci. 30, 439–459.
Soininen, E.M., Bråthen, K.A., Herranz Jusdado, J.G., Reidinger, S., Hartley, S.E., 2013.More than herbivory: levels of silica-based defences in grasses vary with plantspecies, genotype and location. Oikos 122, 30–41.
Tchernov, E.,1979. Quaternary fauna. In: Horowitz, A. (Ed.), The Quaternary of Israel.Academic Press, New York, pp. 257–290.
Tsartsidou, G., Lev-Yadun, S., Albert, R.M., Rosen, A.M., Efstratiou, N., Weiner, S.,2007. The phytolith archaeological record: strengths and weaknesses evaluatedbased on a quantitative modern reference collection from Greece. J. Archaeol.Sci. 34, 1262–1275.
Van-Soest, P.J., 2006. Rice straw, the role of silica and treatments to improve quality.Anim. Feed Sci. Technol. 130, 137–171.
Wallis, L., 2003. An overview of leaf phytolith production patterns in selectednorthwest Australian flora. Rev. Palaeobot. Palynol. 125, 201–248.
Wang, J.J., Dodla, S.K., Henderson, R.E., 2004. Soil silicon extractability with sevenselected extractants in relation to colorimetric and ICP determination. Soil Sci.169, 861–870.
Zdero, C., Bohlmann, F., 1990. Systematics and evolution within the Compositae,seen with the eyes of a chemist. Plant Syst. Evol. 171, 1–14.
Zohary, M., 1983. Man and vegetation in the Middle East. In: Holzner, W., Werger, M.J.A., Ikusima, I. (Eds.), Man’s Impact on Vegetation. Dr. W. Junk Publisher, TheHague, pp. 287–295.
358 O. Katz et al. / Flora 209 (2014) 349–358




















