
Evolution of cassava ( Manihot esculenta Crantz) after recent introduction into a South Pacific...
10
Evolution of cassava (Manihot esculenta Crantz) after recent introduction into a South Pacific Island system: the contribution of sex to the diversification of a clonally propagated crop J. Sardos, D. McKey, M.F. Duval, R. Malapa, J.L. Noyer, and V. Lebot Abstract: Cassava (Manihot esculenta Crantz) is a clonally propagated crop that was introduced into the South Pacific archipelago of Vanuatu in the 1850s. Based on a survey conducted in 10 different villages throughout the archipelago, we present here a study of its diversity. Farmers’ knowledge about cultivation cycle and sexual reproduction of cassava was recorded during group interviews in each village. Using a set of 11 SSR markers, we genotyped the 104 landraces col- lected and 60 supplementary accessions from a within-landrace study (12 landraces  5 plants). Out of the 104 landraces collected, we discovered 77 different multilocus genotypes and the within-landrace study identified several polyclonal landraces. Our data suggest a number of hypotheses about the dynamics of diversity of cassava in Vanuatu. Key words: cassava, clonally propagated crop, farmers’ practices, genetic diversity, landraces, Manihot esculenta Crantz, sexual reproduction. Re ´sume ´: Le manioc (Manihot esculenta Crantz), plante a ` multiplication ve ´ge ´tative, a e ´te ´ introduit au Vanouatou (Paci- fique Sud) dans les anne ´es 1850. Dans cet archipel, il a e ´te ´ bien adopte ´ par la population mais sa diversite ´ est peu connue. Une campagne de collecte a e ´te ´ re ´alise ´e dans 10 villages puis comple ´te ´e par l’enregistrement des savoirs paysans sur les cycles de culture et sur la biologie de la reproduction du manioc. Nous avons ensuite ge ´notype ´ les 104 varie ´te ´s tradition- nelles collecte ´es et 60 accessions d’une e ´tude de variabilite ´ intra-varie ´tale (12 varie ´te ´s paysannes  5 plants) a ` l’aide de 11 marqueurs microsatellites. Sur les 104 varie ´te ´s collecte ´es, 77 pre ´sentent des ge ´notypes multilocus distincts. De plus, l’e ´tude de la variabilite ´ intra-varie ´tale a mis en lumie `re plusieurs varie ´te ´s paysannes polyclonales. Nos donne ´es nous per- mettent de formuler plusieurs hypothe `ses concernant la dynamique de la diversite ´ du manioc dans ce pays. Mots-cle ´s : diversite ´ ge ´ne ´tique, Manihot esculenta Crantz, manioc, pratiques paysannes, propagation clonale, reproduction sexue ´e, varie ´te ´s paysannes. Introduction In Vanuatu, a volcanic archipelago of the southwest Pacific, root crops are staples. Among the many cultivated species in the country, taro (Colocasia esculenta (L.) Schott) and yams (Dioscorea spp.) are strongly entwined in social life and deeply anchored in farmers’ lives through the me- dium of ‘‘custom’’, the traditional way of life. New species are gaining in importance in diets, particularly cassava (Manihot esculenta Crantz), the cultivation of which is increasing almost everywhere in the country. The last agri- culture census (National Statistics Office 2000) reported that in some islands, such as Tanna in the south, its produc- tion outranks that of all other crops. Although cassava is free of serious pests and diseases in Vanuatu, it is vulnerable to cyclones and so remains generally a supplementary food, except in areas where soil fertility is low. In Vanuatu, exclu- sively sweet varieties of cassava are planted; bitter varieties are absent. Cassava is eaten boiled or roasted or made into ‘‘laplap’’, the traditional pudding made from freshly ground roots cooked in folded laplap (Heliconia indica Lam.) leaves. Cassava leaves are not consumed in Vanuatu. Current information suggests that the probable direct wild ancestor of cassava is Manihot esculenta subsp. flabellifolia (Pohl) Ciperri and that the plant was domesticated along the southern border of the Amazon basin (Olsen and Schaal 1999, 2001). Initially grown by Amerindians, it was intro- duced into Africa by Portuguese slave ships from Brazil beginning in the 1500s, with multiple introductions until the 1800s (Fregene et al. 2003), and was first introduced into Asia in the 1700s. In the South Pacific, cassava was intro- duced into New Caledonia by missionaries at the beginning of the 1850s; James Paddon, the first European to buy land Received 10 July 2008. Accepted 28 July 2008. Published on the NRC Research Press Web site at genome.nrc.ca on 3 October 2008. Corresponding Editor: B. Golding. J. Sardos 1 and J.L. Noyer. CIRAD, UMR DAP 1098, Montpellier, F-34398 France. D. McKey. CEFE, CNRS, UMR 5175, Montpellier, F-34293 France. M.F. Duval. CIRAD, UPR Multiplication Ve ´ge ´tative, Montpellier, F-34398 France. R. Malapa. VARTC, Espiritu Santo, POB 231 Vanuatu. V. Lebot. CIRAD, UPR Multiplication Ve ´ge ´tative, Port-Vila, POB 946 Vanuatu. 1 Corresponding author (e-mail: [email protected]). 912 Genome 51: 912–921 (2008) doi:10.1139/G08-080 # 2008 NRC Canada Genome Downloaded from www.nrcresearchpress.com by 192.64.10.76 on 06/05/13 For personal use only.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Evolution of cassava ( Manihot esculenta Crantz) after recent introduction into a South Pacific...

Evolution of cassava (Manihot esculenta Crantz)after recent introduction into a South PacificIsland system: the contribution of sex to thediversification of a clonally propagated crop
J. Sardos, D. McKey, M.F. Duval, R. Malapa, J.L. Noyer, and V. Lebot
Abstract: Cassava (Manihot esculenta Crantz) is a clonally propagated crop that was introduced into the South Pacificarchipelago of Vanuatu in the 1850s. Based on a survey conducted in 10 different villages throughout the archipelago, wepresent here a study of its diversity. Farmers’ knowledge about cultivation cycle and sexual reproduction of cassava wasrecorded during group interviews in each village. Using a set of 11 SSR markers, we genotyped the 104 landraces col-lected and 60 supplementary accessions from a within-landrace study (12 landraces � 5 plants). Out of the 104 landracescollected, we discovered 77 different multilocus genotypes and the within-landrace study identified several polyclonallandraces. Our data suggest a number of hypotheses about the dynamics of diversity of cassava in Vanuatu.
Key words: cassava, clonally propagated crop, farmers’ practices, genetic diversity, landraces, Manihot esculenta Crantz,sexual reproduction.
Resume : Le manioc (Manihot esculenta Crantz), plante a multiplication vegetative, a ete introduit au Vanouatou (Paci-fique Sud) dans les annees 1850. Dans cet archipel, il a ete bien adopte par la population mais sa diversite est peu connue.Une campagne de collecte a ete realisee dans 10 villages puis completee par l’enregistrement des savoirs paysans sur lescycles de culture et sur la biologie de la reproduction du manioc. Nous avons ensuite genotype les 104 varietes tradition-nelles collectees et 60 accessions d’une etude de variabilite intra-varietale (12 varietes paysannes � 5 plants) a l’aide de11 marqueurs microsatellites. Sur les 104 varietes collectees, 77 presentent des genotypes multilocus distincts. De plus,l’etude de la variabilite intra-varietale a mis en lumiere plusieurs varietes paysannes polyclonales. Nos donnees nous per-mettent de formuler plusieurs hypotheses concernant la dynamique de la diversite du manioc dans ce pays.
Mots-cles : diversite genetique, Manihot esculenta Crantz, manioc, pratiques paysannes, propagation clonale, reproductionsexuee, varietes paysannes.
Introduction
In Vanuatu, a volcanic archipelago of the southwestPacific, root crops are staples. Among the many cultivatedspecies in the country, taro (Colocasia esculenta (L.) Schott)and yams (Dioscorea spp.) are strongly entwined in sociallife and deeply anchored in farmers’ lives through the me-dium of ‘‘custom’’, the traditional way of life. New speciesare gaining in importance in diets, particularly cassava
(Manihot esculenta Crantz), the cultivation of which isincreasing almost everywhere in the country. The last agri-culture census (National Statistics Office 2000) reportedthat in some islands, such as Tanna in the south, its produc-tion outranks that of all other crops. Although cassava isfree of serious pests and diseases in Vanuatu, it is vulnerableto cyclones and so remains generally a supplementary food,except in areas where soil fertility is low. In Vanuatu, exclu-sively sweet varieties of cassava are planted; bitter varietiesare absent. Cassava is eaten boiled or roasted or made into‘‘laplap’’, the traditional pudding made from freshly groundroots cooked in folded laplap (Heliconia indica Lam.)leaves. Cassava leaves are not consumed in Vanuatu.
Current information suggests that the probable direct wildancestor of cassava is Manihot esculenta subsp. flabellifolia(Pohl) Ciperri and that the plant was domesticated along thesouthern border of the Amazon basin (Olsen and Schaal1999, 2001). Initially grown by Amerindians, it was intro-duced into Africa by Portuguese slave ships from Brazilbeginning in the 1500s, with multiple introductions until the1800s (Fregene et al. 2003), and was first introduced intoAsia in the 1700s. In the South Pacific, cassava was intro-duced into New Caledonia by missionaries at the beginningof the 1850s; James Paddon, the first European to buy land
Received 10 July 2008. Accepted 28 July 2008. Published on theNRC Research Press Web site at genome.nrc.ca on 3 October2008.
Corresponding Editor: B. Golding.
J. Sardos1 and J.L. Noyer. CIRAD, UMR DAP 1098,Montpellier, F-34398 France.D. McKey. CEFE, CNRS, UMR 5175, Montpellier, F-34293France.M.F. Duval. CIRAD, UPR Multiplication Vegetative,Montpellier, F-34398 France.R. Malapa. VARTC, Espiritu Santo, POB 231 Vanuatu.V. Lebot. CIRAD, UPR Multiplication Vegetative, Port-Vila,POB 946 Vanuatu.
1Corresponding author (e-mail: [email protected]).
912
Genome 51: 912–921 (2008) doi:10.1139/G08-080 # 2008 NRC Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
192.
64.1
0.76
on
06/0
5/13
For
pers
onal
use
onl
y.

and settle in Vanuatu, is believed to have brought a singleclone to the island of Anatom (South Vanuatu) before 1859(Weightman 1989). Now, 150 years after its introduction,cassava is cultivated throughout the country and is availablein every market of the archipelago. Cassava is vegetativelypropagated by farmers: stem cuttings are planted andquickly take root. Despite clonal propagation, which doesnot create diversity, the morphological variation exhibitedby local landraces is remarkable.
Does the morphological variation reflect substantial ge-netic diversity? If so, how did this diversity arise? Cassavais diploid and allogamous. In its Amazonian homeland,Amerindian farmers regularly incorporate ‘‘volunteer’’ seed-lings into their stocks of clones. This is known to be an im-portant source of diversity in cassava populations managedunder traditional farming practices in Amazonia (Elias andMcKey 2000; Elias et al. 2001; Pujol et al. 2005). The in-corporation of plants resulting from seeds into farmers’stocks of vegetatively propagated clones has also been dem-onstrated for other clonally propagated crops, such as potatoin Bolivia (Johns and Keen 1986), ensete in Ethiopia (Shi-geta 1996), and yam in Benin (Scarcelli et al. 2006a,2006b). Is sexual reproduction an important source of cassa-va’s diversity in Vanuatu?
Diversity of cassava has been studied in South Americausing isozymes (Cabral et al. 2002), AFLP markers (Elias etal. 2000), and microsatellites (Carvalho and Schaal 2001; El-ias et al. 2004) and in Africa using RAPD markers (Asanteand Offei 2003; Zacarias et al. 2004) and SSRs (Fregene etal. 2003). Although cassava is also widely cultivated insoutheast Asia and the Pacific, its genetic diversity has notbeen thoroughly investigated in these areas. In this study,we evaluate the extent of cassava’s genetic diversity throughthe description and the SSR genotyping of plants from 10villages, each on a different island of the archipelago. For12 landraces, we also estimate the within-landrace variabilityby the genotyping five individuals from each one of them.
Materials and methods
Plant materialsVanuatu is an archipelago stretching about 850 km from
north to south. Its climate thus varies from humid tropicalin the northern islands from the Torres group to seasonallydry tropical in the islands of Tanna and Erromango in thesouth (Fig. 1). We conducted a survey of cassava landracesin 10 villages of the country between June and November2005. Here, the term ‘‘landrace’’ refers to a group of cassavaplants given the same name by farmers. Each village beinglocated on a different island, natural (climatic and geo-graphic) and human (sociological, religious, and economic)factors were taken into consideration with the aim of study-ing a wide range of situations.
In each village, a list of all of the different named landra-ces was established in close collaboration with the commun-ities. We collected both vegetative propagules (stemcuttings) and leaf samples of each landrace. Cuttings andleaves were taken from a single individual plant. Leaves
from a final set of 104 samples (Table 1) were then storedin filter paper and conserved under silica gel until our returnto the Vanuatu Agricultural Research and Technical Centerstation on the island of Espiritu Santo close to Luganville(Fig. 1) where they were dried in an incubator at 40 8C for48 h. Cuttings were planted at the station to constitute an exsitu collection. In some cases, initial leaf samples that hadnot been well conserved were replaced by leaves from thecorresponding planted cuttings and were then dried in thesame conditions.
In January 2007, we returned to two villages, on Santoand Epi islands, to study within-landrace variation. In eachvillage, we chose five landraces and then collected fiveplants for each. The five plants collected for a landracewere located in five different fields, each belonging to a dif-ferent household. Then, with the aim of evaluating intrafieldvariation, we chose two landraces in a single farmer’s field,located not in a village but close to the Vanuatu AgriculturalResearch and Technical Center, and collected five differentsamples of each. The 60 samples collected during the 2007study (see Table 4) were then processed like those collectedin 2005.
Analysis of data on folk knowledgeStudying how farmers name landraces can give insight
into how they manage cassava diversity. In each village, 5–10 farmers were interviewed about cassava. Landrace names(when these existed) and their translations were recorded.We also asked for the main morphological characteristics ofthe landraces collected (i.e., colours of the root flesh and ofaerial parts). During these interviews, we also gained infor-mation on cultivation cycles and the place of cassava withinthem and recorded farmers’ observations about the sexualreproduction of cassava (i.e., their knowledge about flowers,fruits, seeds, and seedlings).
DNA markersDNA was extracted from 150 mg of dried leaves for each
accession. Leaves were ground with sand and polyvinylpoly-pyrrolidone in liquid nitrogen then 5 mL of lysis bufferwarmed at 37 8C was added following the method describedby Risterucci et al. (2000).
Accessions were genotyped for 11 polymorphic SSR(simple sequence repeat) loci. Primers developed at CentroInternacional de Agricultura Tropical (Mba et al. 2001; Hur-tado et al., in preparation2) (Table 2) were selected for theirhigh polymorphism. Primers were radiolabelled with 33Pbefore PCR amplifications. PCR products were visualizedby running 4 mL of each sample on 5% polyacrylamide gelsthat were then dried and exposed to autoradiographic films.To minimize genotyping errors, we used two controlindividuals and a ladder as size standard on each run. Posi-tions of bands on the gel were scored as allelic data. Theallelic pattern of each accession was scored by a first readerand then checked independently by a second reader.
Analysis of genetic dataAn advantage of using SSR markers in genetic population
2 P. Hurtado, L. Montes, C. Buitrago, C. de Vicente, and M. Fregene. Development of a reference microsatellite marker kit to analyze ge-netic diversity in cassava (Manihot esculenta Crantz).
Sardos et al. 913
# 2008 NRC Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
192.
64.1
0.76
on
06/0
5/13
For
pers
onal
use
onl
y.

studies is their codominant nature: heterozygote and homo-zygote individuals can be strictly distinguished. They thusallow the strict assessment of parameters of genetic varia-tion. Furthermore, they are highly variable, maximizing thepotential for detecting diversity in populations with a sup-posedly narrow genetic base. Molecular data from collec-tions made in 2005 and 2007 were analyzed separately tostudy variation within landraces.
Accessions collected in 2005 were analyzed using thecomputer software DARwin version 5.0 (Perrier et al. 2003;Perrier and Jacquemoud-Collet 2006) to construct aweighted neighbour-joining tree (NJ tree) (Saitou and Nei1987) based on simple matching dissimilarity measures.
One thousand bootstraps were run to determine how wellthe tree was supported by data. Thus, we evaluated G, thenumber of distinct multilocus genotypes (MLGs) present ineach sample, and identified genetic groups. The index ofclonal diversity G/N was computed, where G is the numberof MLGs and N is the total number of genotyped individu-als, to evaluate the global clonality rate of the sample (Hal-kett et al. 2005; Arnaud-Haond et al. 2007). The diversitywas then described using the software GENETIX 4.02 (Bel-khir et al. 2002) by calculating the mean number of alleles,allele frequencies, observed and expected mean heterozygos-ities (Hobs and Hexp, respectively), and Wright’s fixationindex (Fis) at each locus under the null hypothesis of
Fig. 1. Map of Vanuatu.
914 Genome Vol. 51, 2008
# 2008 NRC Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
192.
64.1
0.76
on
06/0
5/13
For
pers
onal
use
onl
y.

Hardy–Weinberg equilibrium. The Hexp was determinedusing the unbiased estimate method of Nei (1978); Fisvalues were calculated following Weir and Cockerham(1984).
Accessions collected in 2007 were also analyzed using anNJ tree based on simple matching dissimilarity measures(data not shown). We performed 100 bootstrap runs andthen compared the different within-landrace MLGs to ex-plore the causes of variation.
Results
Diversity and naming of landracesResults of the survey for each village are presented in Ta-
ble 1. Out of the 104 cassava landraces collected in 2005, 92were given names by farmers. The remaining 12 were rec-ognized as different but farmers could not name them. Fourof the latter were collected in Lolosori (Aoba) (‘‘unnamed 7to 10’’), three in Metaruk (Pentecost) (‘‘unnamed 4 to 6’’),two in Endu (Ambrym) (‘‘unnamed 2 and 3’’), two in Ipota(Erromango) (‘‘unnamed 11 and 12’’), and one in Pesena(Santo) (‘‘unnamed 1’’). The average number of landracesfound per village was 10.4, ranging from 5 in Avunamalaı(Malo) and Metaruk (Pentecost) to 18 in Pesena (EspirituSanto).
Twenty-six landraces were named in bislama, the nationalpidgin English used as a vehicular language, 41 were namedin a local vernacular language, and 25 were named after aperson or a place.
We elucidated the meaning of 72 of the names, whichcould be divided into three thematic groups. The first themewas the origin of landraces: where they originated, how theycame to the village, or who brought them back or foundthem (34 landraces). Among these 34, 19 landraces werenamed after places and three after people. Names of theother 12 evoked anecdotes about the ways by which thelandraces were introduced. The second theme, representing20 names, was the description of aerial organs or of tuber-ous roots of landraces. The third theme, representing 18names, was the specific culinary or agronomical propertiesexhibited by the landraces. Homonyms occurred: sevenwere named ‘‘rice’’ and eight were named ‘‘kari’’. Thename ‘‘rice’’ refers to the cooking properties of the landraces(which farmers considered similar to those of rice), while
‘‘kari’’ refers to the yellow colour of the root flesh of certainlandraces.
Cultivation of cassava and its sexual reproduction infields
Farmers practice slash-and-burn agriculture. After a 3- to4-year fallow period, old fields are reopened and cultivatedfor 2 years. Each farmer has several fields of different ageunder cultivation and he rotates among them. Generally, thecultivation cycle has three phases. The first one is dedicatedto the cultivation, often exclusive, of the culturally dominantcrop, yam or taro. In the second phase, numerous crops aregrown (e.g., maize, yam, sweet potato, cassava, papaya,banana, and sugarcane). In the third phase, solely cassava isgrown. It is then left in the field as ‘‘live food storage’’ andis never cut. Thus, most cassava plants have ample time toflower and fruit. Slashing and burning of vegetation at theend of the fallow period, in the first phase of the cultivationcycle, favors the temperature-dependent germination of cas-sava seeds (Pujol et al. 2002). All farmers reported that theyknew the flowers, fruits, and seeds of cassava. Many ofthem had already observed ‘‘volunteer’’ plants issued fromseeds growing in their new fields. Their attitude regardingthese seedlings is variable, but we recorded numerous inter-views in which farmers stated that they let these volunteerplants grow. We directly observed, in Ambrym, a cassavaplant issued from seed that the farmer allowed to grow totest it.
Analyses of genetic dataThe NJ tree calculated on molecular data issued from the
104 landraces collected in 2005 (one individual per land-race) is presented in Fig. 2. Examination of the tree showsthat five main genetic groups (A to E) can be distinguishedand are, respectively, composed of 35, 22, 18, 25, and 4individuals. These groupings were not supported by highbootstrap values, but this is partly due to the low number ofSSR markers used in this study. Bootstrapping creates datasets randomly resampled from the initial data set; with asmall number of markers, groups are not stable. The internalbranches of the tree were short, while the external brancheswere long, indicating that within-group variability washigher than that between groups. However, some clusterswere supported by high bootstrap values and exhibited shortbranches, showing that within these groups, variabilityamong individuals was low. No geographical organizationof the genetic diversity of cassava (i.e., among islands) wasevident (data not shown).
The sample was composed of 77 distinct MLGs at the 11loci studied, giving a G/N ratio of 0.74. Despite the largenumber of different MLGs in the sample, repetitions oc-curred; we considered that they correspond to a set of clone-mates, i.e., units of the same genetic individual. Forexample, clone R1 is composed of five landraces with vari-ous names and origins: ‘‘vila’’ (Ambrym), ‘‘koten manioko’’(Epi), ‘‘unnamed 12’’ (Erromango), ‘‘tawera mwele’’(Santo), and ‘‘nivila gbaou gbaou’’ (Vanua Lava).
In the total data set, 14 landraces were yellow-fleshed butthese represented only five different MLGs (Table 3) withtwo main sets of clonemates (R2 and R3). They all clusteredin group D of the NJ tree. The largest set of clonemates
Table 1. Number of landraces collected ineach village.
Island Village NVanua Lava Lalngetak 13Espiritu Santo Pesena 18Malo Avunamalaı 5Aoba Lolosori 10Pentecost Metaruk 5Malekula Brenwe 7Epi Burumba 10Ambrym Endu 11Erromango Ipota 14Tanna Lamlu 11Total 104
Sardos et al. 915
# 2008 NRC Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
192.
64.1
0.76
on
06/0
5/13
For
pers
onal
use
onl
y.

(R2) constitutes nine landraces. Two of these landraces arewhite-fleshed, ‘‘naı tom poun’’, ‘‘the king of cassava’’, and‘‘maıok awat’’, ‘‘Awat’s cassava’’, where Awat is the nameof a person. The seven others, the five ‘‘kari’’ (two fromAmbrym, one from Aoba, one from Erromango, and onefrom Malo), ‘‘napound wes’’ (Malekula), and ‘‘maıok ngoıngoı’’ (Vanua Lava), both of the latter names meaning‘‘yellow cassava’’ in local languages, are yellow-fleshed.
We collected in Epi a variegated landrace named ‘‘flowermaniok’’. Interestingly, another landrace named ‘‘salomon’’,collected in the same village but not variegated, exhibited anidentical MLG. We also noticed that within the ‘‘unnamed’’landraces, nine exhibited unique MLGs, while three be-longed to sets of clonemates, i.e., they exhibited MLGsidentical to those of individuals from other landraces.
The number of alleles per locus in our sample rangedfrom two to seven, with an average of 5.36 (Table 2). Forloci NS 189, SSRY 100, SSRY 101, SSRY 103, SSRY 179,SSRY 182, SSRY 20, and SSRY 51, values for Hobs werehigher than those expected under Hardy–Weinberg equili-brium. The negative values of Fis at these loci confirm theheterozygote excess. Overall, Hobs at all loci (0.7195) wasalso higher than Hexp (0.6485) and the multilocus Fis valuewas negative (–0.105) but ranged from 0.076 at NS 376to –0.2100 at SSRY 100 and SSRY 101. Thus, cassava inVanuatu was not under Hardy–Weinberg equilibrium,presenting an excess of heterozygote individuals.
Within-landrace variationA weighted NJ tree based on dissimilarities (100 boot-
straps) revealed two groups among the 12 landracescollected for this analysis (data not shown). Six landraceswere genetically homogeneous but six were heterogeneous,i.e., composed of at least two different MLGs. These resultsare summarized in Table 4. We identified low (£0.05) andhigh (>0.20) dissimilarity values within the landraces. Ex-amination of the different MLGs present within the landra-ces (data not shown) showed that the low dissimilarityvalues corresponded to a variation at one allele over the11 loci genotyped. This was observed twice at the within-field scale and once at the village scale. The high dissimilar-ity values corresponded to variation at several alleles andwere revealed at the village scale only.
Discussion
A comparative survey conducted in Ghana, where cassavawas imported in the 16th century, revealed an average num-ber of 3.5 landraces per village (Manu-Aduening et al.2005), while we collected in Vanuatu an average of 10.4landraces per village. This is a high number, especially fora country where the crop was introduced recently. Manu-Aduening et al. (2005) suggested that environmental con-straints limit the evolution of cassava in Ghana. In Vanuatu,in contrast, cassava is free of serious pests and diseases;environmental constraints are thus low. Also, the relation-ship between named diversity and underlying genetic diver-sity may be different in the two countries. Ghanaian farmersgrow polyclonal landraces composed of plants issued fromdifferent seeds (Manu-Aduening et al. 2005). Vanuatu farm-ers have long and diverse experience in observing thewithin-species diversity of vegetatively propagated crops;their naming of landraces might be more precise than inGhana, thus increasing the number of named landraces. Fur-thermore, at the country scale, 109 different vernacularlanguages are spoken in Vanuatu and the main naming proc-ess is linked to the origin of landraces: most landraces arerenamed by farmers when they arrive in a village. For thesereasons, the names are not by themselves a reliable indica-tion of diversity.
The genetic study gave insights on the nature and the ori-gin of this diversity. As the number of alleles obtained withour set of SSR markers ranged from two to seven, it is ob-vious that, in contrast with statements from the literature,more than a single clone was introduced into the country.The high number of MLGs that we determined, 77, wasalso surprising, given the hypothesis of a single initial intro-duction. The control individuals and the ladder included inthe study minimized scoring errors and we consider thatthis result is consistent. More than a single introduction ofcassava has occurred, but it is highly improbable that eachof these MLGs, or the uncollected ones that certainly occurelsewhere in the country, were all issued from independentintroduction events. Thus, it is likely that much of the diver-sity has appeared in situ, i.e., after the introduction of arelatively small number of clones (at unknown times andplaces). For example, the yellow-fleshed MLGs, which all
Table 2. Number of individuals genotyped, number of alleles, expected and observedheterozygosity, and fixation indices calculated at each locus.
Locus No. of individuals No. of alleles Hexp Hobs Fis
NS 189 104 2 0.2400 0.2788 –0.1570NS 376 104 5 0.7664 0.7115 0.0760SSRY 19 99 6 0.6130 0.6162 0.0000SSRY 20 103 6 0.6960 0.7670 –0.0970SSRY 51 104 5 0.7414 0.8846 –0.1890SSRY 100 104 6 0.7598 0.9231 –0.2100SSRY 101 104 5 0.7048 0.8558 –0.2100SSRY 103 104 7 0.7452 0.8173 –0.0920SSRY 161 104 4 0.4375 0.4231 0.0380SSRY 179 102 5 0.6940 0.8333 –0.1960SSRY 182 102 5 0.7359 0.8039 –0.0880Multilocus 104 5.36 0.6485 0.7195 –0.1050SD . . 0.1649 0.2030 .
916 Genome Vol. 51, 2008
# 2008 NRC Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
192.
64.1
0.76
on
06/0
5/13
For
pers
onal
use
onl
y.
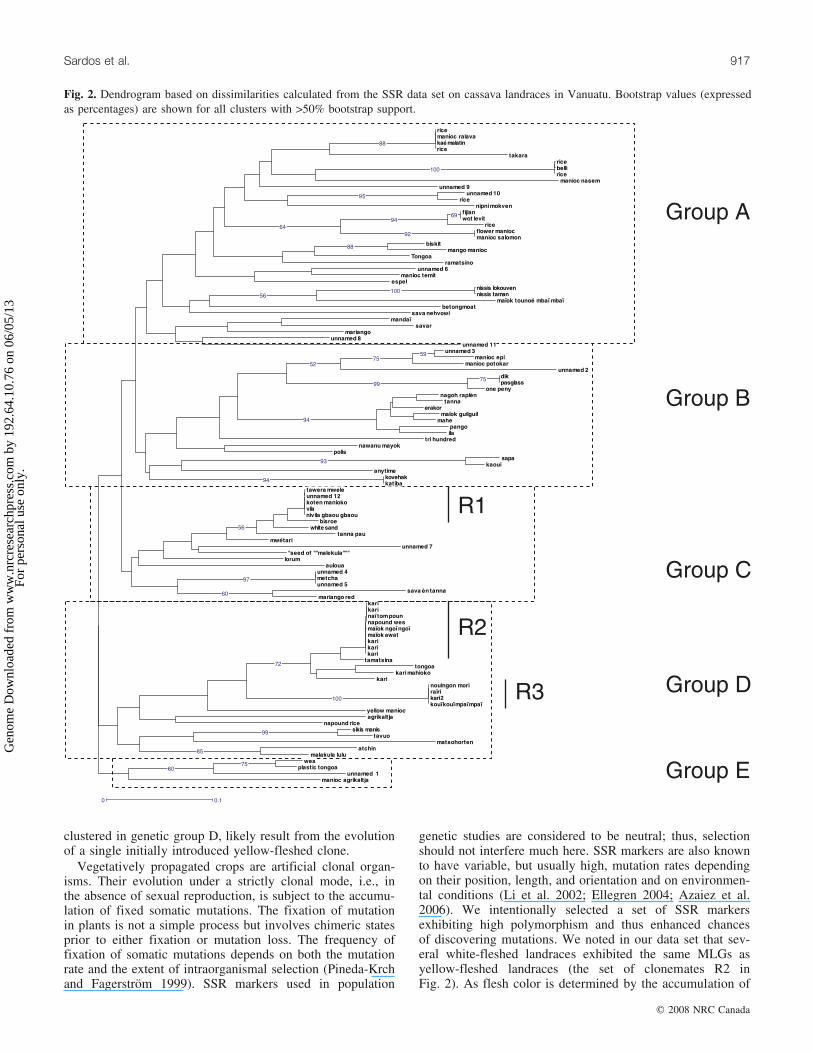
clustered in genetic group D, likely result from the evolutionof a single initially introduced yellow-fleshed clone.
Vegetatively propagated crops are artificial clonal organ-isms. Their evolution under a strictly clonal mode, i.e., inthe absence of sexual reproduction, is subject to the accumu-lation of fixed somatic mutations. The fixation of mutationin plants is not a simple process but involves chimeric statesprior to either fixation or mutation loss. The frequency offixation of somatic mutations depends on both the mutationrate and the extent of intraorganismal selection (Pineda-Krchand Fagerstrom 1999). SSR markers used in population
genetic studies are considered to be neutral; thus, selectionshould not interfere much here. SSR markers are also knownto have variable, but usually high, mutation rates dependingon their position, length, and orientation and on environmen-tal conditions (Li et al. 2002; Ellegren 2004; Azaiez et al.2006). We intentionally selected a set of SSR markersexhibiting high polymorphism and thus enhanced chancesof discovering mutations. We noted in our data set that sev-eral white-fleshed landraces exhibited the same MLGs asyellow-fleshed landraces (the set of clonemates R2 inFig. 2). As flesh color is determined by the accumulation of
Fig. 2. Dendrogram based on dissimilarities calculated from the SSR data set on cassava landraces in Vanuatu. Bootstrap values (expressedas percentages) are shown for all clusters with >50% bootstrap support.
Sardos et al. 917
# 2008 NRC Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
192.
64.1
0.76
on
06/0
5/13
For
pers
onal
use
onl
y.

carotene in roots of cassava, a trait believed to be deter-mined by a few major genes (Iglesias et al. 1997), a muta-tion at one of these loci could be responsible for theabsence of the coloring in these landraces but would be un-detectable with our set of neutral markers. The variegatedlandrace ‘‘Flower maniok’’, is also a product of a somaticmutation: variegation is due to the partial fixation of a muta-tion occurring on the chlorophyll-synthesis pathway (Marco-trigiano 1997). It is thus likely that ‘‘Salomon’’, whichexhibits an identical MLG, is the nonmutated morphotypeof ‘‘Flower maniok’’, which was probably intentionally se-lected by farmers who strive to obtain diversity (Lebot etal. 2005) and capture all variations from an initial phenotype(Caillon and Lanouguere-Bruneau 2005), independently ofwhether these variations might be agronomically useful. Asthe sample comprises landraces that must have issued frommutations on coding sequences of the genome, it is likelythat some of the genotyped MLGs are issued from mutationon the SSR loci that we used in this study. The contributionof somatic mutations to the evolution of other vegetativelypropagated crops, such as grapes (Franks et al. 2002; Cre-span 2004; Hocquigny et al. 2004), olives (Cipriani et al.2002; Lopes et al. 2004), and yams (Scarcelli 2005), has al-ready been demonstrated. Despite its recent introduction intothe country, cassava also seems to have undergone this typeof event. It is likely that the MLGs with very low dissimilar-ity values, i.e., a one-motif variation between alleles at onelocus observed within landraces, are issued from the fixationof somatic mutations on the SSR loci.
According to farmers’ remarks during interviews, they ob-serve cassava seeds and volunteer seedlings in their fields.Currently available data are insufficient to determine if theyare consciously or unconsciously incorporated into landra-ces. However, MLGs with high dissimilarity values (i.e.,diverging at several loci) are unlikely to have resulted fromsomatic mutations and are much more likely to have re-sulted from recombination, i.e., from seeds. Furthermore,Fis values (Table 2) exhibited high variance over loci, whichreflects the occurrence of rare events of sexual recombina-tion in a clonal population (Balloux et al. 2003). Moleculardata, combined with interviews of farmers and field observa-
tions, support the assumption that plants issued from seedsare incorporated into farmers’ stocks of clonally propagatedplants. Thus, both clonal and sexual processes contribute tothe evolution of cassava’s diversity in Vanuatu.
The mixed clonal–sexual reproductive system of cassavaprovides two explanations, not mutually exclusive, for heter-ozygote excess (i.e., negative multilocus Fis value). In astrictly clonal population, heterozygote excess is expectedfrom the independent accumulation of fixed mutations oneach haplotype (Balloux et al. 2003; Halkett et al. 2005). Ina sexual population, heterozygote excess could be due to se-lective advantages enjoyed by heterozygote individuals, par-ticularly in outcrossing plants such as cassava. Worldwide,numerous studies of local cassava landraces have shown anexcess of heterozygote individuals (Pujol et al. 2005; Balye-jusa Kizito et al. 2007; Rocha et al. 2008). In Amazonia, se-lection favoring heterozygote individuals has beendemonstrated (Elias et al. 2004; Pujol 2004; Pujol et al.2005; Pujol and McKey 2006), and heterozygote excess isgenerally imputed to selection among recombinant individu-als. Are the same mechanisms also involved in the adoptionof volunteer plants by farmers in Vanuatu? Further study offarmers’ practices, combined with molecular analysis ofplants at the field scale, could elucidate the role of similarprocesses in Vanuatu.
In Vanuatu, a mixed clonal–sexual system was previouslydescribed for another vegetatively propagated crop, taro(Caillon et al. 2006; Quero-Garcıa et al. 2006). Such a sys-tem could partly explain the weak genetic structure of thesample: clusters with very high bootstrap values and shortbranches could be issued from fixed somaclonal variation ina lineage originated from a single initial MLG, while indi-viduals placed on long branches could be issued from seeds.The lack of geographical structure in genetic diversity at thecountry scale seems to be due to farmers’ practices of culti-var exchange: the finding of identical MLGs collected indifferent islands illustrates cases where clones have beenwidely exchanged throughout the country.
ConclusionThe genetic diversity of cassava in Vanuatu that we have
revealed in this study is much higher than expected: morethan a single clone was introduced into the country and veg-etative propagation is not the only process involved in itscultivation and diversification. Since the crop is clonallypropagated, many authors have in the past argued that sexcould not contribute to the genetic diversification of cassava.However, our results show that although landraces are in-deed clonally propagated, this mode of reproduction cannotaccount for all of the diversity observed. Our unpublishedobservations show that Vanuatu farmers do sometimes clo-nally propagate volunteer seedlings; the results of this studysuggest that this practice has had an important impact on thedynamics of diversity. With such a mixed clonal–sexualsystem, genetic diversification can be extremely rapid infarmers’ fields (Elias et al. 2001). The selection and clonalpropagation of volunteer seedlings appears to be the mainfactor contributing to the rapid diversification of cassava inVanuatu. The dynamics of this process should thus bestudied in detail. Vanuatu’s socioeconomic system ensurescirculation of clones, including new ones propagated from
Table 3. Yellow-fleshed landraces of cassava, their islands ofcollection, and their location in the NJ tree based on moleculardata.
Landrace Island of collection Genetic groupKari Erromango D (R2)Kari Malo D (R2)Napound wes Malekula D (R2)Maıok nogoI ngoı Vanua Lava D (R2)Kari Aoba D (R2)Kari Ambrym D (R2)Kari Ambrym D (R2)Kari mahioko Epi DKari Tanna DNoulgon mori Erromango D (R3)Kari 2 Aoba D (R3)Kouı kouı mpaı mpaı Vanua Lava D (R3)Tamatsina Santo DYellow maniok Pentecost D
918 Genome Vol. 51, 2008
# 2008 NRC Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
192.
64.1
0.76
on
06/0
5/13
For
pers
onal
use
onl
y.

volunteer seedlings, throughout the islands and therebyhomogenizes the genetic diversity at the country scale.Meanwhile, fixation of somatic mutations and reshufflingby recombination continue to generate genotypic diversity.
In 1993, the tragic consequences of the introduction of thetaro leaf blight fungus into Samoa (Naidu and Umar 2003)abruptly revealed the need to enhance the genetic base ofvegetatively propagated crops in the Pacific region. In thecase of cassava in Vanuatu, we recommend the use of thegeographic distribution of allelic diversity, already proposedfor the conservation of minor root crops (Lebot et al. 2005),to assess diversity and enhance it where needed. Introduc-tions of new clones in strategic places would be a meansfor the introduction of alleles of interest (e.g., for synthesisof carotene). Considering the dynamics in place in Vanuatu,these alleles would spread throughout the country by meansof cuttings and naturally enter the preexisting system, favor-ing the recombination and capture of new genotypes pos-sessing the targeted attribute. In which time periodrecombination and capture of such genotypes can occur isstill a point to clarify. This study might be the first step forfurther investigations on this topic. In the 10 villages, thedistribution of clones holding drastically different alleles onthis set of SSR loci followed by cassava’s surveys and gen-otyping after different time periods would help to determinethe speed of the process.
AcknowledgementsThis study would not have been possible without the
financial support of the Agrobiodiversity of Root CropsProject in Vanuatu funded by the FFEM (Fonds Francaispour l’Environnement Mondial’’), CIRAD, and the Ministryof Agriculture in Vanuatu. We thank Marie Melteras, TariMolisale, and the Root Crops Team of the Vanuatu Agricul-tural Research and Technical Center for their support in thefield. We also thank farmers for their patience and hospital-
ity. We also are grateful to Paula Hurtado from CentroInternacional de Agricultura Tropical, Colombia, for provid-ing SSR markers.
ReferencesArnaud-Haond, S., Duarte, C.M., Alberto, F., and Serrao, E.A.
2007. Standardizing methods to address clonality in populationstudies. Mol. Ecol. 16: 5115–5139. doi:10.1111/j.1365-294X.2007.03535.x. PMID:17944846.
Asante, I.K., and Offei, S.K. 2003. RAPD-based genetic diversitystudy of fifty cassava (Manihot esculenta Crantz) genotypes. Eu-phytica, 131: 113–119. doi:10.1023/A:1023056313776.
Azaiez, A., Bouchard, E.F., Jean, M., and Belzile, F.J. 2006.Length, orientation, and plant host influence the mutation fre-quency in microsatellites. Genome, 49: 1366–1373. doi:10.1139/G06-099. PMID:17426751.
Balloux, F., Lehmann, L., and de Meeus, T. 2003. The populationgenetics of clonal and partially clonal diploids. Genetics, 164:1635–1644. PMID:12930767.
Balyejusa Kizito, E., Chiwona-Karltun, L., Egwang, T., Fregene,M., and Westerbergh, A. 2007. Genetic diversity and varietycomposition of cassava on small-scale farms in Uganda: aninterdisciplinary study using genetic markers and farmer inter-views. Genetica, 130: 301–318. doi:10.1007/s10709-006-9107-4.PMID:17082904.
Belkhir, K., Borsa, P., Chikhi, L., Raufaste, N., and Bonhomme, F.2002. GENETIX 4.04, logiciel sous Windows TM pour la genet-ique des populations. Laboratoire Genome, Populations, Inter-actions CNRS UMR 5000, Universite de Montpellier II, France.
Cabral, B.L.R., Souza, J.A.D., Ando, A., Veasey, E.A., and Car-doso, E.M.R. 2002. Isoenzymatic variability of cassava acces-sions from different regions in Brazil. Sci. Agric. 59: 521–527.doi:10.1590/S0103-90162002000300017.
Caillon, S., and Lanouguere-Bruneau, V. 2005. Ethnoecologie enOceanie: gestion de l’agrobiodiversite dans un village de VanuaLava (Vanuatu): strategies de selection et enjeux sociaux. J. Soc.Ocean. 120: 129–148.
Caillon, S., Quero-Garcia, J., Lescure, J.-P., and Lebot, V. 2006.
Table 4. Number of MLGs within selected landraces at the village level and dissimilarityvalues calculated on these MLGs.
Landrace GDissimilarityvalue
Pesena (Espiritu Santo) Kari 1Grindo 1Bik lif 1Raıri 1Sikis manis 2 0.409Total 6
Burumba (Epi) Tongoa 1Blak hand 1Kari 2 0.364Kotten 2 0.227Salomon 4 0.545, 0.045
. 0.500, 0.500
. 0.409, 0.545Total 10
Intra-field (near the Vanuatu Agricultural Researchand Technical Center, Espiritu Santo)
Kari 2 0.050
Tri manis 2 0.045Total 4
Sardos et al. 919
# 2008 NRC Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
192.
64.1
0.76
on
06/0
5/13
For
pers
onal
use
onl
y.

Nature of taro (Colocasia esculenta (L.) Schott) genetic diver-sity prevalent in a Pacific Ocean Island, Vanua Lava, Vanuatu.Genet. Resour. Crop Evol. 53: 1273–1289. doi:10.1007/s10722-005-3877-x.
Carvalho, L.J.C.B., and Schaal, B.A. 2001. Assessing genetic di-versity in the cassava (Manihot esculenta Crantz) germplasmcollection in Brazil using PCR-based markers. Euphytica, 120:133–142. doi:10.1023/A:1017548930235.
Cipriani, G., Marrazzo, M.T., Marconi, R., and Cimato, A. 2002.Microsatellite markers isolated in Olive (Olea europaea L.) aresuitable for individual fingerprinting and reveal polymorphismwithin ancient cultivars. Theor. Appl. Genet. 104: 223–228.doi:10.1007/s001220100685. PMID:12582690.
Crespan, M. 2004. Evidence of the evolution of polymorphism ofmicrosatellite markers in varieties of Vitis vinifera L. Theor.Appl. Genet. 108: 231–237. doi:10.1007/s00122-003-1419-5.PMID:13679982.
Elias, M., and McKey, D. 2000. The unmanaged reproductive ecol-ogy of domesticated plants in traditional agroecosystems: an ex-ample involving cassava and a call for data. Acta Oecol. 21:223–230. doi:10.1016/S1146-609X(00)00053-9.
Elias, M., Panaud, O., and Robert, T. 2000. Assessment of geneticvariability in a traditional cassava (Manihot esculenta Crantz)farming system, using AFLP markers. Heredity, 85: 219–230.doi:10.1046/j.1365-2540.2000.00749.x. PMID:11012725.
Elias, M., Penet, P., Vindry, P., McKey, D., Panaud, O., and Ro-bert, T. 2001. Unmanaged sexual reproduction and the dynamicsof genetic diversity of a vegetatively propagated crop plant, cas-sava (Manihot esculenta Crantz), in a traditional farming sys-tem. Mol. Ecol. 10: 1895–1907. doi:10.1046/j.0962-1083.2001.01331.x. PMID:11555234.
Elias, M., Muhlen, G.S., McKey, D., Roa, A.C., and Tohme, J.2004. Genetic diversity of traditional South American landracesof cassava (Manihot esculenta Crantz): an analysis using micro-satellites. Econ. Bot. 58: 242–256. doi:10.1663/0013-0001(2004)058[0242:GDOTSA]2.0.CO;2.
Ellegren, H. 2004. Microsatellites: simple sequences with complexevolution. Nat. Rev. Genet. 5: 435–445. doi:10.1038/nrg1348.PMID:15153996.
Franks, T., Botta, R., and Thomas, M.R. 2002. Chimerism ingrapevines: implications for cultivars identity, ancestry and ge-netic improvement. Theor. Appl. Genet. 104: 192–199. doi:10.1007/s001220100683. PMID:12582686.
Fregene, M.A., Suarez, M., Mkumbira, J., Kulembeka, H., Ndedya,E., Kulaya, A., et al. 2003. Simple Sequence Repeat markerdiversity in cassava landraces: genetic diversity and differentia-tion in an asexualy propagated crop. Theor. Appl. Genet. 107:1083–1093. doi:10.1007/s00122-003-1348-3. PMID:12856084.
Halkett, F., Simon, J.C., and Balloux, F. 2005. Tackling the popu-lation genetics of clonal and partially clonal organisms. TrendsEcol. Evol. 20: 194–201. doi:10.1016/j.tree.2005.01.001.PMID:16701368.
Hocquigny, S., Pelsy, F., Dumas, V., Kindt, S., Heloir, M.C., andMerdinoglu, D. 2004. Diversification within grapevine cultivarsgoes through chimeric states. Genome, 47: 579–589. doi:10.1139/g04-006. PMID:15190375.
Iglesias, C., Mayer, J., Chavez, L., and Calle, F. 1997. Geneticpotential and stability of carotene content in cassava roots.Euphytica, 94: 367–373. doi:10.1023/A:1002962108315.
Johns, T., and Keen, S.L. 1986. Ongoing evolution of the potato(Solanum sp.) on the altiplano of Western Bolivia. Econ. Bot.40: 409–424.
Lebot, V., Ivancic, A., and Abraham, K. 2005. The geographicaldistribution of allelic diversity, a practical means of preserving
and using minor root crop genetic resources. Exp. Agric. 41:475–489. doi:10.1017/S0014479705002875.
Li, Y.-C., Korol, A.B., Fahima, T., Beiles, A., and And Nevo, E.2002. Microsatellites: genomic distribution, putative functionsand mutational mechanisms: a review. Mol. Ecol. 11: 2453–2465. doi:10.1046/j.1365-294X.2002.01643.x. PMID:12453231.
Lopes, M.S., Mendonca, D., Sefc, K.M., Gil, F.S., and Machado,A. 2004. Genetic evidence of intra-cultivar variability withinIberian olive cultivars. HortScience, 39: 1562–1565.
Manu-Aduening, J.A., Lamboll, R.I., Dankyi, A.A., and Gibson,R.W. 2005. Cassava diversity in Ghanaian farming systems. Eu-phytica, 144: 331–340. doi:10.1007/s10681-005-8004-8.
Marcotrigiano, M. 1997. Chimeras and variegation: pattern ofdeceit. HortScience, 32: 773–784.
Mba, R.E.C., Stephenson, P., Edwards, K., Melzer, S., Nkumbira,J., Gullberg, U., et al. 2001. Simple sequence repeat (SSR) mar-kers survey of the cassava (Manihot esculenta Crantz) genome:towards an SSR-based molecular genetic map of cassava. Theor.Appl. Genet. 102: 21–31. doi:10.1007/s001220051614.
Naidu, V., and Umar M. 2003. Surviving the blight: socio-economic consequences of taro leaf blight (TLB) disease inSamoa. J. South Pac. Agric. 10: 1–9.
Nei, M. 1978. Estimation of average heterozygosity and geneticdistance from a small number of individuals. Genetics, 89: 583–590. PMID:17248844.
National Statistics Office. 2000. The 1999 Vanuatu national popu-lation and housing census, main report. National StatisticsOffice, Port-Vila.
Olsen, K.M., and Schaal, B. 1999. Evidence on the origin ofcassava: phylogeography of Manihot esculenta. Proc. Natl.Acad. Sci. U.S.A. 96: 5586–5591. doi:10.1073/pnas.96.10.5586.PMID:10318928.
Olsen, K.M., and Schaal, B. 2001. Microsatellite variation incassava (Manihot esculenta, Euphorbiaceae) and its wildrelatives: further evidence for southern Amazonian origin ofdomestication. Am. J. Bot. 88: 131–142. doi:10.2307/2657133.PMID:11159133.
Perrier, X., and Jacquemoud-Collet, J.P. 2006. DARwin software.Available from http://www.darwin.cirad.fr/darwin/Home.php[accessed 17 September 2008].
Perrier, X., Flori, A., and Bonnot, F. 2003. Data analysis methods.In Genetic diversity of cultivated tropical plants. Edited byP. Hamon, M. Seguin, X. Perrier, and J.C. Glaszmann. EnfieldSciences Publisher, Montpellier, France. pp. 43–76.
Pineda-Krch, M., and Fagerstrom, T. 1999. On the potential forevolutionary change in meristematic cell through intraorganis-mal selection. J. Evol. Biol. 12: 681–688. doi:10.1046/j.1420-9101.1999.00066.x.
Pujol, B. 2004. Dynamique evolutive de la diversite morphologiqueet genetique d’une plante domestiquee a « propagationvegetative »: le manioc (Manihot esculenta Crantz). Ph.D. thesis,Universite Montpellier II, Montpellier, France.
Pujol, B., and McKey, D. 2006. Size asymmetry in intraspecificcompetition and the density-dependence of inbreeding depres-sion in a natural plant population: a case study in cassava (Man-ihot esculenta Crantz, Euphorbiaceae). J. Evol. Biol. 19: 85–96.doi:10.1111/j.1420-9101.2005.00990.x. PMID:16405580.
Pujol, B., Gigot, G., Laurent, G., Pinheiro-Kluppel, M., Elias, M.,Hossaert-McKey, M., and McKey, D. 2002. Germination ecol-ogy of cassava (Manihot esculenta Crantz, Euphorbiaceae) intraditional agroecosystems: seed and seedling biology of a vege-tatively propagated domesticated plant. Econ. Bot. 56: 366–379.doi:10.1663/0013-0001(2002)056[0366:GEOCME]2.0.CO;2.
Pujol, B., David, P., and McKey, D. 2005. Microevolution in agri-
920 Genome Vol. 51, 2008
# 2008 NRC Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
192.
64.1
0.76
on
06/0
5/13
For
pers
onal
use
onl
y.

cultural environments: how a traditional Amerindian farmingpractice favours heterozygosity in cassava (Manihot esculentaCrantz, Euphorbiaceae). Ecol. Lett. 8: 138–147. doi:10.1111/j.1461-0248.2004.00708.x.
Quero-Garcıa, J., Noyer, J.-L., Weber, A., Perrier, X., McKey, D.,and Lebot, V. 2006. Recombination and clonality in taro (Colo-casia esculenta (L.) Schott): implications for the evolution ofcultivar diversity. Paper presented at the 14th Triennial Sympo-sium of the International Society for Tropical Root Crops(ISTRC), Thiruvananthapuram, India, 21–26 November 2006.
Risterucci, A.M., Grivet, L., N’Goran, J.A.K., Pieretti, I., Flament,M.H., and Lanaud, C. 2000. A high density linkage map ofTheobroma cacao L. Theor. Appl. Genet. 101: 948–955. doi:10.1007/s001220051566.
Rocha, O.J., Zaldivar, M.E., Castro, L., Castro, E., and Barrantes,R. 2008. Microsatellite variation of cassava (Manihot esculentaCrantz) in home gardens of Chibchan Amerindians from CostaRica. Conserv. Genet. 9: 107–118. doi:10.1007/s10592-007-9312-4.
Saitou, N., and Nei, M. 1987. The neighbor-joining tree method: anew method for reconstructing phylogenetic trees. Mol. Biol.Evol. 4: 406–425. PMID:3447015.
Scarcelli, N. 2005. Structure et dynamique de la diversite d’uneplante cultivee a multiplication vegetative: le cas des ignamesau Benin (Dioscorea sp.). Ph.D. thesis, Universite MontpellierII, Montpellier, France.
Scarcelli, N., Tostain, S., Marioc, C., Agbangla, C., Daınou, O.,Berthaud, L., and Pham, J.L. 2006a. Genetic nature of yams(Dioscorea sp.) domesticated by farmers in Benin (West Africa).Genet. Resour. Crop Evol. 53: 121–130. doi:10.1007/s10722-004-1950-5.
Scarcelli, N., Tostain, S., Vigouroux, Y., Agbangla, C., Daınou, O.,and Pham, J.-L. 2006b. Farmers’ use of wild relative and sexualreproduction in a vegetatively propagated crop. The case of yamin Benin. Mol. Ecol. 15: 2421–2431. doi:10.1111/j.1365-294X.2006.02958.x. PMID:16842416.
Shigeta, M. 1996. Creating landrace diversity: the case of the Aripeople and ensete, Ensete ventricosum, in Ethiopia. In Redefin-ing nature: ecology, culture and domestication. Edited by R. El-len and K. Fukui. Oxford University Press, Oxford, U.K.pp. 233–229.
Weightman, B. 1989. Agriculture in Vanuatu: a historical review.British Friends of Vanuatu, Surrey.
Weir, B.S., and Cockerham, C.C. 1984. Estimating F-statistics forthe analysis of population structure. Evolution, 38: 1358–1370.doi:10.2307/2408641.
Zacarias, A.M., Botha, A.M., Labuschagne, M.T., and Benesi,I.R.M. 2004. Characterization and genetic distance analysis ofcassava (Manihot esculenta Crantz) germplasm from Mozambi-que using RAPD fingerprinting. Euphytica, 138: 49–53. doi:10.1023/B:EUPH.0000047057.05642.74.
Sardos et al. 921
# 2008 NRC Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
192.
64.1
0.76
on
06/0
5/13
For
pers
onal
use
onl
y.




















