
Role of Toll-Like Receptor 4 in Induction of Cell-Mediated Immunity and Resistance to Brucella...
11
INFECTION AND IMMUNITY, Jan. 2004, p. 176–186 Vol. 72, No. 1 0019-9567/04/$08.000 DOI: 10.1128/IAI.72.1.176–186.2004 Copyright © 2004, American Society for Microbiology. All Rights Reserved. Role of Toll-Like Receptor 4 in Induction of Cell-Mediated Immunity and Resistance to Brucella abortus Infection in Mice Marco A. Campos, 1,2 Gracia M. S. Rosinha, 1,3 Igor C. Almeida, 4 Xirlene S. Salgueiro, 4 Bruce W. Jarvis, 5 Gary A. Splitter, 5 Nilofer Qureshi, 6 Oscar Bruna-Romero, 2 Ricardo T. Gazzinelli, 1,2 and Sergio C. Oliveira 1 * Department of Biochemistry and Immunology, Biological Sciences Institute, Federal University of Minas Gerais, 1 and Centro de Pesquisas Rene ´ Rachou, Oswaldo Cruz Foundation, 2 Belo Horizonte, EMBRAPA-CNPC, Sobral-CE, 3 and Department of Parasitology, University of Sa ˜o Paulo, Sa ˜o Paulo, 4 Brazil; Department of Animal Health & Biomedical Sciences, University of Wisconsin-Madison, Madison, Wisconsin 53706 5 ; and Department of Basic Medical Science, Medical School, University of Missouri, Kansas City, Missouri 64108 6 Received 31 July 2003/Returned for modification 20 September 2003/Accepted 3 October 2003 Initial host defense to bacterial infection is executed by innate immunity, and therefore the main goal of this study was to examine the contribution of Toll-like receptors (TLRs) during Brucella abortus infection. CHO reporter cell lines transfected with CD14 and TLRs showed that B. abortus triggers both TLR2 and TLR4. In contrast, lipopolysaccharide (LPS) and lipid A derived from Brucella rough (R) and smooth (S) strains activate CHO cells only through TLR4. Consistently, macrophages from C3H/HePas mice exposed to R and S strains and their LPS produced higher levels of tumor necrosis factor alpha (TNF-) and interleukin-12 compared to C3H/HeJ, a TLR4 mutant mouse. The essential role of TLR4 for induction of proinflammatory cytokines was confirmed with diphosphoryl lipid A from Rhodobacter sphaeroides. Furthermore, to determine the contribution of TLR2 and TLR4 in bacterial clearance, numbers of Brucella were monitored in the spleen of C3H/HeJ, C3H/HePas, TLR2 knockout, and wild-type mice at 1, 3, and 6 weeks following B. abortus infection. Interest- ingly, murine brucellosis was markedly exacerbated at weeks 3 and 6 after infection in animals that lacked functional TLR4 (C3H/HeJ) compared to C3H/HePas that paralleled the reduced gamma interferon produc- tion by this mouse strain. Finally, by mass spectrometry analysis we found dramatic differences on the lipid A profiles of R and S strains. In fact, S lipid A was shown to be more active to trigger TLR4 than R lipid A in CHO cells and more effective in inducing dendritic cell maturation. In conclusion, these results indicate that TLR4 plays a role in resistance to B. abortus infection and that S lipid A has potent adjuvant activity. Brucella is a facultative intracellular bacterium that infects humans and domestic animals (6). The pathological manifes- tations of brucellosis are diverse and include arthritis, endo- carditis, and meningitis in humans, while animal brucellosis is characterized by spontaneous abortion (58). Previously, immu- nity to intracellular bacteria was considered to be exclusively mediated by CD4 T cells (62). More recently, our studies have evoked a central role for CD8 T cells in conferring protective immunity against intracellular pathogens such as Brucella (40). In addition, initial host defense against bacterial infection is executed by innate immunity stimulated by patho- gen-associated molecular patterns, conserved molecular struc- tures common to different groups of pathogens that are rec- ognized by host receptors (33). In Drosophila melanogaster, Toll receptors play a key role in antimicrobial host defense (27). Recently, mammalian homo- logues of Toll, designated Toll-like receptors (TLRs), were also discovered (34, 45, 51). Bacterial cell wall components and lipopolysaccharide (LPS) are recognized by host macrophages and dendritic cells via TLRs (33) that results in activation of professional antigen-presenting cells, initiation of acquired im- mune responses, and further elimination of the invasive bac- teria (53, 54). Among TLR family members, both TLR2 and TLR4 have been shown to recognize bacterial components. Recent studies have demonstrated that TLR2 is largely required for signaling by numerous ligands from gram-negative and gram-positive bacteria (20, 29, 31, 48, 53, 57), mycobacteria (29, 32, 56), spirochetes (2, 19, 29), and mycoplasmas (29), such as lipotei- choic acid, peptidoglycan, lipoproteins (2, 5, 19, 33), and cer- tain types of LPS (20, 31). In contrast, TLR4 fails to confer responsiveness to gram-positive bacteria and their components and has been postulated to be the main LPS signaling receptor. LPS is a major constituent of the outer membrane of gram- negative bacteria such as Brucella and is known to activate neutrophils, monocytes, macrophages, and other cell types to upregulate expression of adhesion molecules and produce a number of pro and anti-inflammatory cytokines (37). Recog- nition of bacterial LPS is mediated by CD14 (17). However, CD14 lacks transmembrane and intracellular domains neces- sary for signal transduction and thus requires the involvement of molecules belonging to the TLR family. Recently, with flu- orescence resonance energy transfer techniques, LPS was shown to trigger a physical association between CD14 and TLR4 (25). B. abortus induces interleukin (IL)-12 production from human monocytes and this effect was blocked by anti- CD14 antibody, suggesting that the Brucella binding and/or signaling to monocytes was mediated via LPS (59). Addition- * Corresponding author. Mailing address: Departamento de Bio- quimica e Imunologia, Universidade Federal de Minas Gerais, Av. Antonio Carlos 6627, Pampulha, Belo Horizonte, MG, Brazil, 30161- 970. Phone and fax: 55-31-34992666. E-mail: [email protected]. 176
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Role of Toll-Like Receptor 4 in Induction of Cell-Mediated Immunity and Resistance to Brucella...

INFECTION AND IMMUNITY, Jan. 2004, p. 176–186 Vol. 72, No. 10019-9567/04/$08.00�0 DOI: 10.1128/IAI.72.1.176–186.2004Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Role of Toll-Like Receptor 4 in Induction of Cell-Mediated Immunityand Resistance to Brucella abortus Infection in Mice
Marco A. Campos,1,2 Gracia M. S. Rosinha,1,3 Igor C. Almeida,4 Xirlene S. Salgueiro,4Bruce W. Jarvis,5 Gary A. Splitter,5 Nilofer Qureshi,6 Oscar Bruna-Romero,2
Ricardo T. Gazzinelli,1,2 and Sergio C. Oliveira1*Department of Biochemistry and Immunology, Biological Sciences Institute, Federal University of Minas Gerais,1 and Centro de
Pesquisas Rene Rachou, Oswaldo Cruz Foundation,2 Belo Horizonte, EMBRAPA-CNPC, Sobral-CE,3 and Department ofParasitology, University of Sao Paulo, Sao Paulo,4 Brazil; Department of Animal Health & Biomedical Sciences,
University of Wisconsin-Madison, Madison, Wisconsin 537065; and Department of Basic Medical Science,Medical School, University of Missouri, Kansas City, Missouri 641086
Received 31 July 2003/Returned for modification 20 September 2003/Accepted 3 October 2003
Initial host defense to bacterial infection is executed by innate immunity, and therefore the main goal of thisstudy was to examine the contribution of Toll-like receptors (TLRs) during Brucella abortus infection. CHOreporter cell lines transfected with CD14 and TLRs showed that B. abortus triggers both TLR2 and TLR4. Incontrast, lipopolysaccharide (LPS) and lipid A derived from Brucella rough (R) and smooth (S) strains activateCHO cells only through TLR4. Consistently, macrophages from C3H/HePas mice exposed to R and S strainsand their LPS produced higher levels of tumor necrosis factor alpha (TNF-�) and interleukin-12 compared toC3H/HeJ, a TLR4 mutant mouse. The essential role of TLR4 for induction of proinflammatory cytokines wasconfirmed with diphosphoryl lipid A from Rhodobacter sphaeroides. Furthermore, to determine the contributionof TLR2 and TLR4 in bacterial clearance, numbers of Brucella were monitored in the spleen of C3H/HeJ,C3H/HePas, TLR2 knockout, and wild-type mice at 1, 3, and 6 weeks following B. abortus infection. Interest-ingly, murine brucellosis was markedly exacerbated at weeks 3 and 6 after infection in animals that lackedfunctional TLR4 (C3H/HeJ) compared to C3H/HePas that paralleled the reduced gamma interferon produc-tion by this mouse strain. Finally, by mass spectrometry analysis we found dramatic differences on the lipid Aprofiles of R and S strains. In fact, S lipid A was shown to be more active to trigger TLR4 than R lipid A inCHO cells and more effective in inducing dendritic cell maturation. In conclusion, these results indicate thatTLR4 plays a role in resistance to B. abortus infection and that S lipid A has potent adjuvant activity.
Brucella is a facultative intracellular bacterium that infectshumans and domestic animals (6). The pathological manifes-tations of brucellosis are diverse and include arthritis, endo-carditis, and meningitis in humans, while animal brucellosis ischaracterized by spontaneous abortion (58). Previously, immu-nity to intracellular bacteria was considered to be exclusivelymediated by CD4� T cells (62). More recently, our studieshave evoked a central role for CD8� T cells in conferringprotective immunity against intracellular pathogens such asBrucella (40). In addition, initial host defense against bacterialinfection is executed by innate immunity stimulated by patho-gen-associated molecular patterns, conserved molecular struc-tures common to different groups of pathogens that are rec-ognized by host receptors (33).
In Drosophila melanogaster, Toll receptors play a key role inantimicrobial host defense (27). Recently, mammalian homo-logues of Toll, designated Toll-like receptors (TLRs), werealso discovered (34, 45, 51). Bacterial cell wall components andlipopolysaccharide (LPS) are recognized by host macrophagesand dendritic cells via TLRs (33) that results in activation ofprofessional antigen-presenting cells, initiation of acquired im-
mune responses, and further elimination of the invasive bac-teria (53, 54).
Among TLR family members, both TLR2 and TLR4 havebeen shown to recognize bacterial components. Recent studieshave demonstrated that TLR2 is largely required for signalingby numerous ligands from gram-negative and gram-positivebacteria (20, 29, 31, 48, 53, 57), mycobacteria (29, 32, 56),spirochetes (2, 19, 29), and mycoplasmas (29), such as lipotei-choic acid, peptidoglycan, lipoproteins (2, 5, 19, 33), and cer-tain types of LPS (20, 31). In contrast, TLR4 fails to conferresponsiveness to gram-positive bacteria and their componentsand has been postulated to be the main LPS signaling receptor.
LPS is a major constituent of the outer membrane of gram-negative bacteria such as Brucella and is known to activateneutrophils, monocytes, macrophages, and other cell types toupregulate expression of adhesion molecules and produce anumber of pro and anti-inflammatory cytokines (37). Recog-nition of bacterial LPS is mediated by CD14 (17). However,CD14 lacks transmembrane and intracellular domains neces-sary for signal transduction and thus requires the involvementof molecules belonging to the TLR family. Recently, with flu-orescence resonance energy transfer techniques, LPS wasshown to trigger a physical association between CD14 andTLR4 (25). B. abortus induces interleukin (IL)-12 productionfrom human monocytes and this effect was blocked by anti-CD14 antibody, suggesting that the Brucella binding and/orsignaling to monocytes was mediated via LPS (59). Addition-
* Corresponding author. Mailing address: Departamento de Bio-quimica e Imunologia, Universidade Federal de Minas Gerais, Av.Antonio Carlos 6627, Pampulha, Belo Horizonte, MG, Brazil, 30161-970. Phone and fax: 55-31-34992666. E-mail: [email protected].
176

ally, Brucella’s ability to elicit IL-12 secretion enables it to driveTh0 cells to differentiate into Th1 effector and memory cellsthat is a central feature of the potential use of B. abortus as avaccine carrier and adjuvant.
B. abortus strain RB51 is a stable rough (R) mutant derivedfrom the standard smooth (S) virulent strain 2308. This roughmutant strain that does not contain the O-antigen (O polysac-charide chain of the smooth LPS) is attenuated in its virulencecompared to their smooth virulent parental strain (3). Eventhough the Brucella RB51 rough strain has been widely used asa live vaccine, it induces lower protection compared to thesmooth vaccine strain S19 (56). Similar to most intracellularbacterial infections, cell-mediated immunity plays a major rolein acquired resistance to brucellosis, although antibodies tosurface antigens, especially to the O-antigen, can confer acertain level of protection against a challenge infection (4).Therefore, it is pivotal to dissect how this difference in protec-tion engendered by these two vaccine strains could be relatedto the ability of R versus S LPS to signal through TLRs trig-gering the innate immune system.
In the present study, we investigated the ability of B. abortusR (vaccine) and S (virulent) strains and their purified LPS aswell as lipid A to trigger TLR2 and TLR4. Heat-killed bacteriaactivate both TLR2 and TLR4, whereas purified LPS and lipidA signal through TLR4. Further, we demonstrated that opti-mal induction of tumor necrosis factor alpha (TNF-�) andIL-12 by macrophages exposed to B. abortus as well as efficientbacterial clearance in mouse spleens required functionalTLR4. Finally, lipid A from a B. abortus S strain had a strongerimmunostimulatory activity than lipid A derived from the Rstrain. We believe that these findings have important implica-tions in bacterial pathogenesis and vaccine developmentagainst brucellosis.
MATERIALS AND METHODS
Mice. TLR4 mutants (C3H/HeJ) were obtained from FIOCRUZ (Rio deJaneiro, Brazil), and the control mice, C3H/HePas, were purchased from theUniversity of Sao Paulo (Sao Paulo, Brazil) and maintained at the FederalUniversity of Minas Gerais (Belo Horizonte, Brazil). TLR2 knockout mice andwild-type littermates (C57BL/6 � 129/Ola) were kindly provided by Shizuo Akirafrom Osaka University in Japan. BALB/c mice at 6 to 8 weeks of age wereobtained and maintained at CPqRR/FIOCRUZ (Belo Horizonte, Brazil).
Bacterial strains and LPS and lipid A extraction. Brucella abortus S2308 is thewild-type virulent S strain. The RB51, the current live attenuated U.S. vaccinederived from strain S2308 deficient in O-chain LPS (rough), was a gift from G. G.Schurig (Virginia-Maryland Regional College of Veterinary Medicine, VirginiaPolytechnic Institute and State University, Blacksburg, Va.). The R LPS from B.abortus strain RB51 and the S LPS from strain S2308 were purified as detailed byMoreno et al. (35). Final purity of the LPS was determined by both thin-layerchromatography and sodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE) on a 4 to 20% gradient gel that was subsequently silver-stained(Pierce Chemical Co., Rockford, Ill.) as previously described (20).
Preliminary dose-response analyses were conducted with heat-killed Brucellaand LPS concentrations as described elsewhere (13). The lipid A moiety wasisolated from R and S LPS after mild acid hydrolysis with 0.25 N HCl (Pierce) for1 h at 100°C, followed by Folch partition (12). Lipid A was recovered in theupper phase, dried under N2 and redissolved in chloroform-methanol (1:1, vol/vol). Under the conditions used here, the lipid A species from both Brucellastrains were released at high yield and purity. This was inferred from compara-tive experiments carried out with authentic LPSs from Escherichia coli, Salmo-nella minnesota, Bordetella pertussis, and Serratia marcescens, strictly under thesame conditions. Neither internal lipid A fragments, partially hydrolyzed lipid Aspecies, nor contaminant oligosaccharides were observed on the lipid A prepa-rations when analyzed by electrospray ionization time-of-flight mass spectrom-etry (ESI-TOF-MS).
Electrospray ionization mass spectrometry. Lipid A species isolated from Rand S LPSs were analyzed by electrospray ionization time-of-flight mass spec-trometry (ESI-TOF-MS) with a Q-TOF-MS instrument (Ultima, MicromassLtd., Manchester, United Kingdom). Samples, diluted in chloroform-methanol(1:1, vol/vol), were introduced into the electrospray source by infusion through a20-�l loop with the assistance of a high-pressure liquid chromatography (HPLC)pump (Shimazu, Japan), at a flow rate of 20 �l/min. Spectra were collected innegative-ion mode over 1 to 2 min at 2 s/scan with an interscan delay of 0.2 s, overa mass range of 1,000 to 2,300. Source, API probe, and desolvation temperatureswere set at 80, 100, and 150°C, respectively. Cone voltage and collision energywere 100 and �10 V, respectively. Combined raw spectra were further processedfor background subtraction and smoothing (Saviztky-Golay method) with theMassLynx V3.5 software. Multiply charged ion species were deconvoluted tosingle-ion species with the MaxEnt3 function of the same software. Authenticlipid A preparations from E. coli and S. minnesota (Sigma Chemical Co., St.Louis, Mo.) were used to calibrate the instrument.
CHO cell lines. The CHO reporter cell lines (CHO/CD14 and CHO/CD14/TLR2) (29) were derived from clone 3E10, a CHO/CD14 cell line that has beenstably transfected with a reporter construct containing the gene for CD25 underthe control of the human E-selectin promoter. This promoter contains an NF-�Bbinding site, and CD25 expression is completely dependent upon NF-�B trans-location (10). Cells expressing TLRs were obtained by stable transfection of theCHO/CD14 reporter cell line (which contains endogenous TLR4) with thecDNA for human TLR2 as described (29). In addition to the LPS-responsive celllines described above, we also tested a CHO/CD14 LPS nonresponder cell line(10) designated clone 7.19 as well as a clonal line derived from this mutant thatwas transfected with CD14 and TLR2 (7.19/CD14/TLR2). This cell line wasderived from 3E10, and reports NF-�B activation via the surface expression ofCD25, similarly to the other CHO lines described here. The LPS-nonresponsivephenotype of the 7.19 cell lines appears to be due to a mutation in the MD-2gene, and thus they are defective in signaling via TLR4 (10, 18, 55). All the celllines were maintained as adherent monolayers in Ham�s F-12/Dulbecco’s mod-ified Eagle’s medium (DMEM) supplemented with 5% fetal bovine serum, at37°C, 5% CO2, and antibiotics as described (29). Furthermore, the cell lines 7.19,7.19/CD14/TLR2, CHO/CD14, and CHO/CD14/TLR2 were also termed here asLPS nonresponder control cell lines TLR2, TLR4, and TLR2/TLR4, respec-tively, according to the TLR that they express.
Flow cytometry analysis. Reporter cells were plated at a density of 105 cells/well in 24-well tissue culture dishes. The next day, either 103 heat-killed bacteria(heat inactivation was performed at 80°C for 1 h)/cell ratio from the S and Rstrain, 1 �g/ml of purified S LPS and R LPS, and 40, 200, and 1,000 ng of purifiedS lipid A and R lipid A per ml were added as indicated, in a total volume of 250�l of medium/well, for 18 h. The cells were harvested with trypsin-EDTA treat-ment for 50 s, washed once with medium containing 5% serum and again withphosphate-buffered saline, and then 105 cells were stained with phycoerythrin-labeled anti-CD25 (mouse monoclonal antibody to human CD25, PE conjugate;CALTAG Laboratories, Burlingame, Calif.) 1:200 in phosphate-buffered saline,on ice, in the dark, for 45 min. The cells were also harvested physically withouttrypsin with similar results. After two washes, the cells were resuspended in 1mM sodium azide in phosphate-buffered saline, and 10,000 cells were examinedby flow cytometry (BD Biosciences, San Jose, Calif.). Dead cells were excludedby gating them with the forward scatter and side scatter parameters, and afterthat the average of 7,641 � 546 (Fig. 1) and 7,471 � 265 (Fig. 5A) live cells wereanalyzed for the expression of surface CD25. Analyses were performed with CellQuest software (BD Biosciences).
IL-12/TNF-� measurements. TLR2 knockout (52), C57BL/6 � 129/Ola (wild-type), C3H/HeJ (LPS hyporesponsive), and C3H/HePas mice were inoculatedintraperitoneally with 2 ml of 3% thioglycolate and, 4 days later, the elicitedperitoneal exudate cells were harvested in cold serum-free DMEM. The mediumused in the macrophage cultures (MacMed) consisted of DMEM supplementedwith 40 �g of gentamicin/ml and 5% heat-inactivated fetal bovine serum. Themacrophages were washed and resuspended in MacMed at 2 � 106/ml, and100-�l aliquots were dispensed into the wells of a 96-well plate. Cells wereallowed to adhere at 37°C and 5% CO2, for 3 h, and were then washed once withserum-free DMEM and 150 �l of MacMed was added to each well, in thepresence and absence of 20 U of IFN- per ml and incubated overnight at 37°Cand 5% CO2. The same concentration of Brucella and purified LPS used in flowcytometry was added to the macrophage cultures in a final volume of 200 �l/wellat the same time as IFN-. Aliquots of the supernatant were collected after 24 hand 48 h of culture for TNF-� and IL-12 (p70) measurements, respectively (8, 9).Levels of TNF-� and IL-12 (p70) in the supernatants were measured by acommercially available enzyme-linked immunosorbent assay (ELISA) kit (Duo-set; R&D Systems, Minneapolis, Minn.).
VOL. 72, 2004 TLR4 SIGNALING DURING BRUCELLA INFECTION 177
Macrophage cell line and DPLA competition assay. Rhodobacter sphaeroidesdiphosphoryl lipid A (DPLA) was purified as described by Qureshi et al. (43).RAW 264.7 murine macrophage-like cells were obtained from the AmericanType Culture Collection (ATCC) (cell line TIB-71), cultured as previously de-scribed (23), except that the medium was RPMI 1640 supplemented with 10%fetal bovine serum, penicillin (50 U/ml) and streptomycin (50 �g/ml). LPS wasadded to RAW cells 18 h before the culture supernatant was assayed for TNF-�.In blocking experiments, DPLA was added 30 min prior to addition of BrucellaLPS. TNF-� was assayed with duplicate determinations by means of the TNF-�Quantikine M Immunoassay (R&D Systems, Minneapolis, Minn.). Each assaywas repeated three times.
Infection and Brucella quantitation in spleens. Mice were infected intraperi-toneally with 105 CFU of B. abortus strain 2308. To count residual Brucella CFUin the spleens of mice, 10 animals from each group were examined at eachsampling period. Spleens from individual animals were homogenized in phos-phate-buffered saline, 10-fold serially diluted, and plated on Brucella Broth agar(Difco, Detroit, Mich.). Plates were incubated at 37°C in air with 5% CO2, andthe number of CFU was counted after 3 days.
Spleen cultures and IFN-� detection assay. Freshly removed spleens of miceinfected with B. abortus strain S2308 were placed in petri dishes (60 � 15 mm)containing 10 ml of phosphate-buffered saline and passed through steel mesh to
obtain single-cell suspensions. The splenocyte suspension were then isolated bydensity gradient centrifugation with Ficoll-Paque (Sigma) for 30 min in a SorvallH-1000B rotor at 2,000 rpm (800 � g) in 50-ml tubes. Spleen cells floating on topof the high-density solution were isolated by slowly moving the tip of the pipetteover the surface of the high-density layer and drawing cells up in a 5-ml pipetteand transferred to a sterile 50-ml tube. Cells were then washed twice with sterilephosphate-buffered saline containing penicillin G sodium (100 U/ml), strepto-mycin sulfate (100 �g/ml), and amphotericin B (250 ng/ml) per ml.
Afterward, splenocytes were cultured in RPMI 1640 medium supplementedwith 2 mM L-glutamine, 25 mM HEPES (N-2-hydroxyethylpiperazine-N�-2-eth-anesulfonic acid), 50 �M 2-mercaptoethanol, penicillin G sodium (100 U/ml),streptomycin sulfate (100 �g/ml), amphotericin B (250 ng/ml), and 10% heat-inactivated fetal bovine serum (Sigma), then placed at 106 cells per well in96-well tissue culture plates. Murine splenocytes from infected animals werestimulated with 1 �g of Brucella S LPS and 108 heat-inactivated Brucella abortusS2308 per ml. Unstimulated splenocytes were used as a negative control, andsplenocytes stimulated with concanavalin A (5 �g/ml) were a T-cell-activatingcontrol. After 72 h at 37°C under 5% CO2, splenocyte culture supernatants weretested for the presence of cytokine with the ELISA Duoset kit (R & D Systems)to mouse IFN-.
Immunization with lipid A-activated dendritic cells and IFN-� analysis by
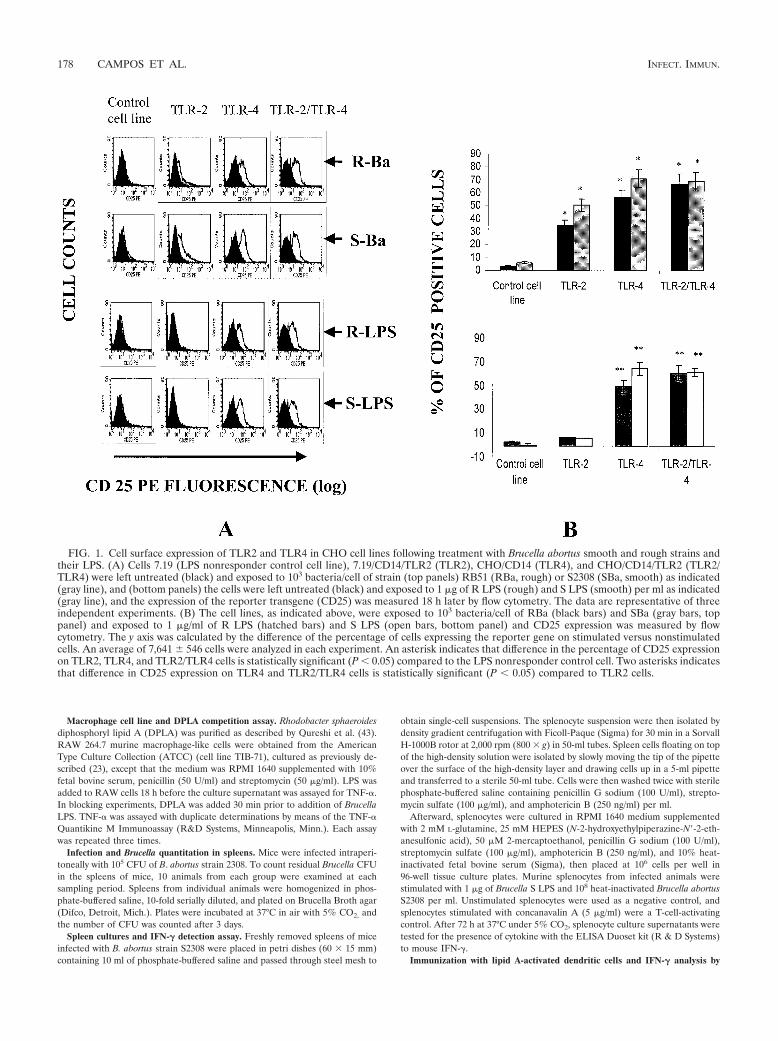
FIG. 1. Cell surface expression of TLR2 and TLR4 in CHO cell lines following treatment with Brucella abortus smooth and rough strains andtheir LPS. (A) Cells 7.19 (LPS nonresponder control cell line), 7.19/CD14/TLR2 (TLR2), CHO/CD14 (TLR4), and CHO/CD14/TLR2 (TLR2/TLR4) were left untreated (black) and exposed to 103 bacteria/cell of strain (top panels) RB51 (RBa, rough) or S2308 (SBa, smooth) as indicated(gray line), and (bottom panels) the cells were left untreated (black) and exposed to 1 �g of R LPS (rough) and S LPS (smooth) per ml as indicated(gray line), and the expression of the reporter transgene (CD25) was measured 18 h later by flow cytometry. The data are representative of threeindependent experiments. (B) The cell lines, as indicated above, were exposed to 103 bacteria/cell of RBa (black bars) and SBa (gray bars, toppanel) and exposed to 1 �g/ml of R LPS (hatched bars) and S LPS (open bars, bottom panel) and CD25 expression was measured by flowcytometry. The y axis was calculated by the difference of the percentage of cells expressing the reporter gene on stimulated versus nonstimulatedcells. An average of 7,641 � 546 cells were analyzed in each experiment. An asterisk indicates that difference in the percentage of CD25 expressionon TLR2, TLR4, and TLR2/TLR4 cells is statistically significant (P 0.05) compared to the LPS nonresponder control cell. Two asterisks indicatesthat difference in CD25 expression on TLR4 and TLR2/TLR4 cells is statistically significant (P 0.05) compared to TLR2 cells.
178 CAMPOS ET AL. INFECT. IMMUN.

ELISPOT. Immature dendritic cells were obtained by in vitro differentiation ofBALB/c mouse bone marrow-derived precursors, as previously reported (7).Differentiated dendritic cells, uninfected and infected 24 h before with AdPyCS,a recombinant adenovirus expressing the CS protein from Plasmodium yoelii,were used to immunize different groups of mice. To determine the effect of LPSon dendritic cell activation, mice received subcutaneously 750,000 dendritic cells/mouse that were treated right after adenoviral infection with B. abortus purifiedS lipid A and R lipid A (1 �g/ml). Ten days after immunization, mice weresacrificed and the presence of CS-specific IFN--producing CD8� and CD4� Tcells in their spleens was assayed by ELISPOT with CS-derived CD8� (SYVPSAEQI) and CD4� (YNRNIVNRLLGDALNGKPEEK) peptides, as previouslydescribed (7).
Statistical analysis. Statistical analyses were performed with Student’s t testand analysis of variance with computer software package MINITAB (MinitabInc., State College, Pa.).
RESULTS
LPS from B. abortus triggers cell activation through TLR4.CHO reporter cell lines that were stably transfected with CD14alone (CHO/CD14), TLR2 and CD14 (CHO/CD14/TLR2)both expressing endogenous TLR4, and the control cell lines7.19 expressing CD14 and 7.19/CD14/TLR2 expressing CD14and TLR2, but not functional TLR4 were exposed for 18 h toB. abortus S and R strains and S LPS and R LPS, and NF-�Bactivation was assessed by measuring the expression of CD25by flow cytometry (10, 29). Increased induction of CD25 ex-pression by 7.19/CD14/TLR2, CHO/CD14, and CHO/CD14/TLR2 cells exposed to R and S B. abortus was observed, indi-cating that heat-killed Brucella activates both TLR2 and TLR4pathways (Fig. 1A, top panels). Additionally, Fig. 1B (toppanel) showed that the difference in the percentage of CD25positive CHO cells stimulated with R and S B. abortus wasmuch higher (P 0.05) on those cells that express TLR2,TLR4, and TLR2/TLR4 compared to the LPS nonresponder
control cell line (7.19). In contrast, NF-�B activation was de-tected only in CHO/CD14 and CHO/CD14/TLR2 but not in7.19/CD14/TLR2 cells treated with S LPS and R LPS (Fig. 1A,bottom panels). Further, the percentage of CD25-positive cellsfollowing treatment with S LPS and R LPS was statisticallysignificant on CHO cells that express TLR4 and TLR2/TLR4compared to those cells that possess TLR2 and the LPS non-responder control cell line (Fig. 1B, bottom panel). These datademonstrate that LPS derived from either S or R strains ofBrucella triggers cell activation through TLR4 but not TLR2.
TLR4 is required for maximal proinflammatory activity ofB. abortus and its LPS on murine macrophages. Previous stud-ies have demonstrated that B. abortus induces high levels ofIFN-, TNF-�, and IL-12 (38, 39). Here, we tested the involve-ment of TLR2 and TLR4 on cytokine production by murineinflammatory macrophages stimulated with B. abortus S and Rstrains and their LPS. C3H/HePas cells treated with Brucella Sand R strains and their purified LPS produced significantlyhigher levels of TNF-� and IL-12 compared to cells fromC3H/HeJ TLR4 mutant mice (Table 1). No statistically signif-icant differences were observed in TNF-� and IL-12 produc-tion by macrophages from TLR2 knockout mice compared tothe wild-type control group (C57BL/6 � 129/Ola). Addition-ally, priming inflammatory macrophages with IFN- enhancedTNF-� and IL-12 synthesis by these cells of all mouse strainsstudied, but a significant difference was still observed only oncytokine production by C3H/HePas versus C3H/HeJ cells. Fur-thermore, IFN--primed macrophages of all mice studied stim-ulated with S LPS produced much higher levels of IL-12 com-pared to R LPS. This result corroborates the datademonstrated here showing more effective adjuvant activity ofB. abortus S LPS. Overall, these findings suggest that TLR4 has
TABLE 1. Levels of TNF-� and IL-12 (p70) in the supernatants of inflammatory macrophages derived from C3H/HeJ, C3H/HePas, TLR2knockout or wild-type mice treated with heat-inactivated B. abortus smooth or rough strains and their purified LPSa
Macrophages, cytokine,and strain
Amt (pg/ml)
C3H/HeJ C3H/HePas TLR2 knockout Wild type (for TLR2)
Unprimed macrophagesTNF-�
Ba (smooth) 550 � 39 850 � 35 810 � 41 710 � 34Ba (rough) 520 � 40 780 � 38 780 � 38 660 � 31S-LPS 10 467 � 53 470 � 45 540 � 52R-LPS 10 690 � 65 500 � 70 600 � 63
IL-12Ba (smooth) 10 45 � 6 38 � 3 35 � 4Ba (rough) 10 38 � 5 47 � 6 25 � 4S-LPS 10 60 � 7 27 � 3 25 � 5R-LPS 10 55 � 6 20 � 4 21 � 4
IFN--primed macrophagesTNF-�
Ba (smooth) 850 � 61 1,940 � 68 1,310 � 30 1,300 � 54Ba (rough) 780 � 43 1,580 � 50 1,900 � 68 1,540 � 75S-LPS 10 645 � 59 1,620 � 150 1,300 � 105R-LPS 10 806 � 78 870 � 130 800 � 130
IL-12Ba (smooth) 682 � 44 1,666 � 72 1,599 � 75 1,240 � 69Ba (rough) 58 � 4 180 � 10 150 � 13 198 � 20S-LPS 10 1643 � 67 749 � 58 950 � 81R-LPS 10 162 � 15 54 � 10 51 � 4
a Macrophages were exposed to B. abortus smooth or rough strains at 103 bacteria/macrophage or their LPS at 1 �g/ml, and supernatants were harvested 24 and 48 hlater to measure TNF-� and IL-12 (p70), respectively. The results shown here are the mean � standard deviation of duplicate measurements from two separateexperiments. Data from C3H/HePas mice were statistically different compared to C3H/HeJ mice, P 0.05.
VOL. 72, 2004 TLR4 SIGNALING DURING BRUCELLA INFECTION 179

a dominant role in macrophage signaling and production ofproinflammatory cytokines by macrophages exposed to R andS strains of B. abortus and their LPS.
DPLA inhibits B. abortus LPS activity on macrophages.Diphosphoryl lipid A from Rhodobacter sphaeroides (DPLA) isa potent LPS antagonist, and it blocks the release of TNF-�and IL-1� in both murine and human cells through CD14/TLR4 interaction (24, 30, 41, 42). The DPLA inhibitory effecton the ability of B. abortus LPS to induce the production ofTNF-� was tested. Addition of 1 �g of DPLA on BrucellaLPS-treated RAW 264.7 macrophage-like cells dramaticallyreduced TNF-� production by these cells (Fig. 2). This resultconfirmed our previous data described here regarding the roleof TLR4 on proinflammatory cytokine production induced byBrucella LPS.
Role of TLR4 in B. abortus clearance. To determine thecontribution of TLR2 and TLR4 in bacterial clearance, num-bers of Brucella were monitored in the spleens of C3H/HeJ,C3H/HePas, TLR2 knockout, and wild-type mice at 1, 3, and 6weeks following B. abortus infection. The animals were sacri-ficed weekly and the numbers of B. abortus CFU were deter-mined. Interestingly, murine brucellosis was markedly exacer-bated, at weeks 3 and 6 after infection, in animals that lackedfunctional TLR4 (C3H/HeJ) compared to C3H/HePas (Fig. 3,top panel). In contrast, TLR2 knockout mice showed no sta-tistically significant difference in bacterial load compared tocontrol animals at all intervals studied (Fig. 3, bottom panel).These in vivo findings reinforce the role of TLR4 in triggeringthe immune response against an intracellular bacterial patho-gen such as Brucella, functioning as an important molecule inthe control of infection.
Decrease in IFN-� production by spleen cells parallels en-hanced susceptibility to Brucella infection in TLR4 mutantmice. IFN- is a pivotal cytokine involved in the control ofmurine brucellosis (26). To determine the level of IFN- pro-duced by C3H/HeJ and C3H/HePas mice following infection,we measured this cytokine in the supernatant of splenocytesfrom these animals after stimulation with B. abortus and itsLPS. At 1 week postinfection, no difference was observed inthe levels of IFN- produced by TLR4 mutant and C3H/HePascells treated with B. abortus (Table 2), but a statistically signif-
icant difference in IFN- production by C3H/HePas spleno-cytes was observed compared to C3H/HeJ cells when S LPSwas used as the stimulus. Interestingly, at 3 and 6 weeks postin-fection, a major difference in IFN- production was detected inC3H/HePas splenocyte supernatants compared to C3H/HeJwhen B. abortus was tested as the antigen. These data parallelour in vivo study that shows the decrease in bacterial load inC3H/HePas spleens compared to C3H/HeJ at 3 and 6 weekspostinfection. Together, these results demonstrate that TLR4is required for inducing a robust adaptive immunity and IFN-synthesis for B. abortus infection and therefore an efficientcontrol of murine brucellosis.
FIG. 2. DPLA inhibits the activity of B. abortus LPS on RAW 264.7macrophage-like cells. B. abortus R LPS (100 ng) was added to 5 � 105
RAW cells in 1 ml of medium in a 12-well culture plate with andwithout a 30-min preincubation with 1 �g of DPLA, and TNF-� wasmeasured. Data points are the average of duplicate measurementsfrom three separate experiments � standard deviation. Differences inrelation to DPLA�BaLPS treatment is denoted by an asterisk (*) forP 0.05.
FIG. 3. Course of B. abortus infection in C3H/HeJ, C3H/HePas,TLR2 knockout and wild-type mice. The graph illustrates B. abortusCFU in the spleen from TLR4 mutant (top panel) and TLR2 knockout(bottom panel) mice and their controls determined at 1, 3, and 6 weeksafter infection. Data are expressed as means � standard deviation of10 animals per time point of two separate experiments. Differences inrelation to C3H/HeJ mice are denoted by an asterisk (*) for P 0.05.
TABLE 2. Levels of IFN- produced by splenocytes from TLR4mutant and C3H/HePas mice following infection and in vitro
stimulated with heat-inactivated B. abortus smooth strain and itspurified LPSa
Timepostinfection
(wk)Antigen
IFN- (pg/ml)
C3H/HeJ C3H/HePas
1 S-Ba 7,141 � 976 6,100 � 837S-LPS 472 � 92 2,397 � 447
3 S-Ba 1,600 � 162 4,701 � 1,043S-LPS 156 � 52 1,722 � 15
6 S-Ba 1,400 � 161 3,800 � 850S-LPS 128 � 44 1,520 � 110
a The results are the mean � standard deviation of duplicate measurementsfrom two separate experiments. Data from C3H/HePas mice were differentstatistically compared to C3H/HeJ mice, P 0.05.
180 CAMPOS ET AL. INFECT. IMMUN.

Mass spectrometry analysis of lipid A from rough andsmooth strains. Since the proinflammatory activity of LPS isdetermined by its lipid A moiety (11, 14), we decided to carryout a comparative analysis of lipid A of R and S LPS from B.abortus. The lipid A moieties were obtained by mild acid treat-ment of the R and S LPS and analyzed by negative-ion modeESI-TOF-MS. Both R and S lipid A original mass spectrashowed a complex array of singly, doubly, and triply chargedion species (data not shown). These mass spectra were decon-voluted to spectra displaying only singly charged ion species inorder to simplify the analysis. As clearly shown in Fig. 4, the Slipid A preparation exhibits a much more complex ion profilethan the R lipid A preparation.
S lipid A species form ion clusters in at least four regions ofthe spectrum, at m/z 1154 to 1350, 1432 to 1588, 1753 to 1855,and 1965 to 2107 (Fig. 4A). In contrast, the majority of R lipidA ion species were grouped in a single region of the spectrumat m/z 1965 to 2107 (Fig. 4B). Nevertheless, few minor scat-tered ion species could also be observed in the R lipid Apreparation at m/z 1154 to 1350, 1432 to 1588 and 1753 to1855. The four ion clusters observed in the mass spectra ofboth S and R lipid A preparations correspond to lipid A spe-cies with a variable degree of acylation. For instance, ion spe-cies at m/z 1154 to 1350 represent tetraacylated lipid A species,whereas ion clusters at m/z 1432 to 1588, 1753 to 1855 and 1965to 2107, correspond to penta-, hexa-, and heptaacylated lipid Aspecies, respectively. This was inferred from the observationthat several ion species along the spectrum seemed to originatefrom species at higher m/z range, by the loss of 238 Da, whichmay correspond to the loss of an ester-linked C16:0 fatty acidchain (Fig. 4A, B). Taken together, the ESI-TOF-MS dataindicate that the S lipid A preparation contains a higher pro-portion of tetra- and pentaacylated ion species than the R lipidA preparation, which by contrast displays essentially heptaacy-lated species.
This structural difference may be the key factor that contrib-utes to the enhanced activity observed when S lipid A is usedas the stimulus. Based on previous work by Qureshi et al. (44)and Moryion and Lopez-Goni (36) and our present observa-tions, we tentatively assigned the structure of lipid A seriesoriginated from m/z 2065 of R and S strains (Fig. 4C). It isnoteworthy to point out that the predicted monoisotopicmasses (pMmi) for all tentative structures are very close (inaverage 0.200 Da mass error) to the experimental monoiso-topic masses (eMmi) obtained in this study. Nevertheless, weare currently carrying out extensive GC-MS, ESI-TOF-MS/MS, and nuclear magnetic resonance analyses of R and S lipidA species in order to confirm the proposed structural assign-ments and to determine the structure of the species that wecould not determine here.
Enhanced TLR4 signaling through purified Brucella smoothstrain lipid A. To determine whether the difference in compo-sition of lipid A species from Brucella S and R strains couldactually result in different biological activity of these mole-cules, we measured cell activation through TLR4 signaling.Differently from the data shown on Fig. 1 for whole LPS (1�g/ml), purified lipid A from Brucella S strain was shown toactivate TLR4 in much lower concentrations (40 and 200 ng/ml) compared to lipid A from Brucella R strain as observed byincreased CD25 expression on CHO/CD14/TLR2 (TLR2/
TLR4) and CHO/CD14 (TLR2) cell surface (Fig. 5, top andbottom panels).
Brucella lipid A activates dendritic cells to trigger IFN-�production by malaria peptide-specific CD4� and CD8� Tcells. Bruna-Romero and Rodriguez (7) have previously shownthat dendritic cells expressing the CS protein from the rodentmalaria parasite Plasmodium yoelii with a CS recombinantadenovirus vector were able to present this malaria antigen tothe immune system upon maturation. In the present work, weused equivalent adenovirus-transduced immature dendriticcells treated with purified S lipid A and R lipid A from B.abortus to immunize mice to test the dendritic cell maturationability of these LPS molecules.
BALB/c instead of C57BL/6 mice were used in this experi-ment because of the H-2d restriction of the CS peptides. Micevaccinated with adenovirus-transduced dendritic cells treatedwith S lipid A induced a much greater antigen-specific IFN-production by either CD4� and CD8� T cells compared toanimals immunized with uninfected dendritic cells and in-fected dendritic cells not treated or treated with R lipid A (Fig.5B). Enhanced IFN- production by CD4� and CD8� T lym-phocytes of mice immunized with S lipid A-treated dendriticcells is probably related to the ability of Brucella S LPS toinduce high levels of IL-12 in primed mouse macrophagescompared to R LPS (Table 1). These results suggest that S LPSobtained from B. abortus has a great capacity to activate den-dritic cells compared to R LPS and, as a consequence, displaysa strong adjuvant effect for the induction of an adaptive im-mune response.
DISCUSSION
Brucella abortus is the causative agent of human and animalbrucellosis, and this air-borne pathogen not only resists killingby neutrophils but also replicates inside macrophages (28). Bymechanisms that are still not completely understood, Brucellapersists as a chronic infection in mammalian hosts by escapingthe microbicidal activities of host macrophages (47). The re-cent discovery of TLR proteins, a family of mammalian patternrecognition receptors, has provided new insights into our un-derstanding of the mechanisms by which Brucella can elicitcellular responses from innate immune cells. In this study, weinvestigated the ability of B. abortus and its purified LPS andlipid A to trigger TLR2 and TLR4 and the impact of thisactivation in innate recognition and elimination of invadingbacteria.
First, with CHO reporter cell lines transfected with CD14alone, TLR2 and CD14, both expressing endogenous TLR4and MD-2 and the control cell line 7.19 not expressing MD-2and therefore not possessing functional TLR4, we determinedNF-�B activation by measuring CD25 expression on the cellsurface after exposure to B. abortus R and S strains and theirLPS. The data presented here demonstrate that B. abortustriggers TLR2 and TLR4 activation, while its LPS induces cellactivation through TLR4. Recent studies have demonstratedthat TLR2 confers responsiveness to a wide variety of gram-positive bacterial cell wall components as well as to lipopro-teins that are found in gram-positive and gram-negative bac-teria (54). Further, Lorenz et al. (31) have demonstrated thatLPS from Bacteroides fragilis functions through TLR4-indepen-
VOL. 72, 2004 TLR4 SIGNALING DURING BRUCELLA INFECTION 181

FIG. 4. ESI-Q-TOF-MS profile of lipid A species from Brucella abortus smooth and rough strains. (A and B) Deconvoluted spectra of S andR lipid A species. (C) Tentative structural assignments for ion species m/z 1350, 1588, 1827, and 2065 of R and S lipid A. eMmi, experimentalmonoisotopic mass; pMmi, predicted monoisotopic mass; �M, mass difference between eMmi and pMmi. Predicted monoisotopic masses werecalculated with the ACD/ChemSketch 5.12 version (ACD Labs, Toronto). �C16:0, loss of a C16:0 fatty acid chain.
182 CAMPOS ET AL. INFECT. IMMUN.

dent mechanisms. But LPS preparations from Escherichia coliand Salmonella spp. lose their ability to induce TLR2-depen-dent responses after removal of contaminant proteins, suggest-ing that lipoproteins rather than the LPS are responsible forcellular activation via TLR2 (21).
Thus, B. abortus NF-�B activation via TLR2 in 7.19/CD14/TLR2 cells is probably due to lipoproteins and other cell wallcomponents from this organism, since LPS from R and Sstrains did not trigger TLR2. Regarding TLR4 signaling, B.abortus and its LPS from R and S strains activate NF-�B in theCHO/CD14 cell line that does not possess TLR2 on the sur-face. Thus, TLR2 expression is not required for mediatingresponses to LPS from B. abortus strains used here. The im-portance of TLR4 for responses to LPS and gram-negativebacteria has been confirmed by the observation that targeteddisruption of the tlr4 gene renders mice unresponsive to LPSand that this defect can be reversed in macrophages derived
from TLR4-deficient mice by reintroducing a wild-type copy ofthe gene (22).
As with other intracellular microbial pathogens, IL-12-de-rived IFN- production, and TNF-� are critical components ofthe immune system to control murine brucellosis (38, 39). Ourfindings revealed that TLR4 is required for B. abortus LPSsignaling to produce TNF-� and IL-12, while TLR2 is not.Additionally, when whole B. abortus was used as the stimulus,we observed a marked difference on inflammatory macrophagederived IL-12 and TNF-� production from C3H/HePas micevia TLR4 activation compared to C3H/HeJ mouse cells. Nev-ertheless, C3H/HeJ cells still produced TNF-� and IL-12 im-plying the involvement of additional TLRs besides TLR4 in B.abortus signaling.
To confirm our hypothesis that TLR4 itself is sufficient toconfer the ability to recognize Brucella LPS, we took advantageof the species-specific pharmacology of diphosphoryl lipid A
FIG. 5. Enhanced cell activation and IFN- production through TLR4 by Brucella abortus lipid A from a smooth strain compared to a roughstrain. (A) CHO/CD14/TLR2 (TLR2/TLR4, solid bars), 7.19/CD14/TLR2 (TLR2, hatched bars), CHO/CD14 (TLR4, open bars), and 7.19 (LPSnonresponder control, gray bars) reporter cells were exposed to 40, 200, and 1,000 ng/ml of purified lipid A from Brucella abortus smooth (toppanel) and rough (bottom panel) strains. After 18 h of exposure, CD25 gene expression was analyzed by flow cytometry. The y axis was calculatedby the difference of the percentage of cells expressing the reporter gene on stimulated versus nonstimulated cells. An average of 7,471 � 265 cellswere analyzed in each experiment. An asterisk indicates that difference in CD25 expression on cells stimulated with the same concentration of Slipid A versus R lipid A is statistically significant (P 0.05). The results are representative of three independent experiments. (B) Antimalaria (CSspecific) cellular immune responses induced 10 days after immunization in BALB/c mice that received immature dendritic cells, uninfected andinfected with AdPyCS. Infected dendritic cells were additionally treated with B. abortus R lipid A and S lipid A (1 �g/ml). The number ofCS-specific IFN--producing CD8� and CD4� T cells was determined by ELISPOT with splenocytes and A20.2J cell line preincubated with thecorresponding CS-derived CD8� and CD4� peptides as antigen-presenting cells. The results are representative of two independent experimentsand are shown as the mean � standard deviation. An asterisk indicates differences (P 0.05) from S lipid A-treated group in relation to R lipidA and infected and uninfected dendritic cells without lipid A treatment for CD8� (solid bars) and (open bars) for CD4� T cells. Statisticaldifferences were calculated by analysis of variance.
VOL. 72, 2004 TLR4 SIGNALING DURING BRUCELLA INFECTION 183

from Rhodobacter sphaeroides (DPLA). This molecule acts asan antagonist of LPS in mouse cells (30). Macrophage cell lineRAW 264.7 treated with DPLA dramatically reduced TNF-�production induced by B. abortus LPS. The most likely expla-nation for the mechanism of LPS inhibition by DPLA is thatthis antagonist competes with the lipid A portion of LPS for acommon binding site on TLR4, since DPLA blocks E. coliLPS-induced signaling in a TLR4-dependent fashion (30). Incontrast, DPLA fails to block Porphyromonas gingivalis LPS-induced TNF-� production because P. gingivalis LPS signalsvia TLR2 (20).
To determine the role of TLR4 and TLR2 in Brucella infec-tion, we infected C3H/HeJ, C3H/HePas, TLR2 knockout, andwild-type mice with the virulent B. abortus strain S2308. TLR4mutant mice (C3H/HeJ) demonstrated an enhanced suscepti-bility to B. abortus infection at 3 and 6 weeks postinfectioncompared to C3H/HePas mice. Recently, TLR4 mutant micewere apparently highly effective in controlling the primary My-cobacterium tuberculosis infection, but host immune protectionto M. tuberculosis was insufficient for long-term persistence invivo (1). Regarding TLR2-deficient mice, these animals aremore susceptible to infection by Staphylococcus aureus (50).But, in the case of infection with B. abortus we found thatTLR2 knockout mice controlled infection as efficiently as wild-type mice. These in vivo data confirmed the dominant role ofTLR4 versus TLR2 in triggering innate immunity in murinebrucellosis and therefore helping the host to control this bac-terial infection. Additionally, Brucella-primed splenocytesfrom C3H/HeJ exposed to heat-killed bacteria produced muchlower levels of IFN- at 3 and 6 weeks postinfection time whenthe number of bacteria was elevated compared to C3H/HePasanimals.
A recent study with IFN--deficient mice has demonstratedthat at 3 weeks postinfection with B. abortus these animals hadtwice the spleen weights, a threefold increase in the total num-ber of splenic leukocytes, and a higher number of bacteria thanthe control mice (39). A major mechanism to explain enhancedC3H/HeJ susceptibility to infection is based on limited IFN-production. This drop in IFN- synthesis probably occurreddue to decreased IL-12 and TNF-� production by C3H/HeJinflammatory macrophages exposed to Brucella. Zhan andCheers (61) previously demonstrated that TNF-� produced byBrucella acts as an autocrine factor to up-regulate IL-12 andconsequently IFN- production.
Knowing that the lipid A moiety is the major moleculeresponsible for the proinflammatory properties of LPS, wedecided to define the Brucella S and R strains lipid A profile bymass spectrometry. The mass spectra revealed very distinctprofiles between R and S lipid A (Fig. 4). These differencesseem to be a result of variations in the number and size of thefatty acid chains of the lipid A species from both strains, andperhaps it could account for some of the significant differencesin their biological activity observed here. Goldstein et al. (16)demonstrated that Brucella abortus lipid A has diaminoglu-coses instead of glucosamines and long-chain fatty acids, andtherefore it is less pyrogenic than enterobacterial LPS. Toconfirm if this difference in lipid A structure between S and Rstrains could be responsible for different biological activity ofthese molecules, we measured cell activation through TLR4signaling. Our results demonstrated that at the same concen-
tration S lipid A had a greater ability to induce NF-�B activa-tion trough TLR4 than R lipid A.
Immature dendritic cells are unable to present antigens andinduce immune responses in the absence of inflammatory stim-uli (like TNF-� and IL-1), which, acting immediately afterantigen uptake, induce dendritic cells to mature. We wanted togain more insights into the ability of Brucella S LPS and R LPSto activate dendritic cells and, as a consequence, to inducespecific adaptive immune responses. Mice immunized with ad-enovirus-transduced immature dendritic cells expressing theCS protein from the rodent malaria parasite Plasmodium yoeliiand treated with purified Brucella S lipid A, induced an en-hanced IFN- production by murine CD4� and CD8� T cellsspecific to CS-derived peptides compared to untreated den-dritic cells and treated with R lipid A. These data confirm thatBrucella S LPS is a more powerful adjuvant compared to RLPS not only to trigger innate but also adaptive immune re-sponses. Recently, B. abortus LPS has been shown to inducerelevant quantities of �-chemokines known to bind to the hu-man immunodeficiency virus type 1 coreceptor CCR5 andblock virus entry (60). Therefore, heat-inactivated B. abortusand its LPS have been proposed as carriers for therapeuticvaccines for individuals with human immunodeficiency virus(15). LPS from B. abortus differs structurally from the LPS ofE. coli and S. enterica serovar Typhimurium, is much less py-rogenic in rabbits and mice, and induces less TNF-� fromhuman monocytes (16). Thus, these properties make B. abortusLPS a safe adjuvant to be used in vaccine production.
The activation of macrophages represents one of the firstevents in the innate resistance to intracellular infection. Intra-cellular pathogens such as Brucella, phagocytosed by macro-phages and acting through surface molecules like LPS thatsignal through TLR4 activate these cells to produce a series ofcytokines that further contribute to bacterial clearance. In thisstudy, we have shown that TLR4 has a dominant effect com-pared to TLR2 on Brucella macrophage activation and hostcontrol of infection. In addition to improve our understandingof Brucella pathogenesis our findings have an important impli-cation in vaccine development. At similar concentrations, theBrucella S vaccine strain (S19) induces higher protection thanthe R vaccine strain (RB51) (46). The lack of protective anti-bodies to the O side chain of the LPS in animals immunizedwith B. abortus RB51 may explain in part why the R straininduces lower protection against infection (49). Herein, wedemonstrated that structural difference between lipid A moietyof LPS from S and R strains are responsible for enhanced cellactivation through TLR4 and cytokine production. Therefore,we suggest that the difference in protection induced by the Sversus R vaccine strains may also be dependent on activationof the innate immune system and proinflammatory cytokineproduction via TLR4.
ACKNOWLEDGMENTS
We thank Douglas T. Golenbock (University of Massachusetts Med-ical School, Worcester, Mass.) and Shizuo Akira (Department of HostDefense, Research Institute for Microbial Diseases, Osaka University,Osaka, Japan) for providing us with the CHO cell lines and the TLR2knockout mice, respectively.
This work was supported by the Conselho Nacional de Desenvolvi-mento Científico e Tecnologico (CNPq), PADCT/CNPq, CBAB/CNPq, FAPEMIG, FIOCRUZ (PAPES III and PDTIS) and National
184 CAMPOS ET AL. INFECT. IMMUN.

Institutes of Health grant RO1 AI48490. M.A.C. and O.B.R. are vis-iting scientists from CNPq/FIOCRUZ, G.M.S.R. is a PostdoctoralFellow from CNPq, and R.T.G., I.C.A., and S.C.O. are ResearchFellows from CNPq. I.C.A. is supported by a grant from the Fundacaode Amparo a Pesquisa do Estado de Sao Paulo (FAPESP).
REFERENCES
1. Abel, B., N. Thieblemont, V. J. F. Quesniaux, N. Brown, J. Mpagi, K. Miyake,F. Bihl, and B. Ryffel. 2002. Toll-like receptor 4 expression is required tocontrol chronic Mycobacterium tuberculosis infection in mice. J. Immunol.169:3155–3162.
2. Aliprantis, A. O., R.-B. Yang, M. R. Mark, S. Suggett, B. Devaux, J. D.Radolf, G. R. Klimpel, P. Godowski, and A. Zychlinsky. 1999. Cell activationand apoptosis by bacterial lipoproteins through Toll-like receptor-2. Science285:736–739.
3. Allen, C. A., L. Garry Adams, and T. A. Ficht. 1998. Transposon-derivedBrucella abortus rough mutants are attenuated and exhibit reduced intracel-lular survival. Infect. Immun. 66:1008–1016.
4. Araya, L. N., and A. J. Winter. 1990. Comparative protection of mice againstvirulent and attenuated strains of Brucella abortus by passive transfer ofimmune T cells and serum. Infect. Immun. 58:254–256.
5. Brightbill, H. D., D. H. Libraty, S. R. Krutzik, R.-B. Yang, J. T. Belisle, J. R.Bleharski, M. Maitland, M. V. Norgard, S. E. Plevy, S. T. Smale, P. J.Brennan, B. R. Bloom, P. J. Godowski, and R. L. Modlin. 1999. Host defensemechanisms triggered by microbial lipoproteins through Toll-like receptors.Science 285:732–736.
6. Brinley Morgan, W. J., and M. J. Corbel. 1990. Brucella infections in manand animals: contagious equine metritis, p. 547–585. In M. T. Parker andL. H. Collier (ed.) Topley and wilson�s principles of bacteriology, virologyand immunology. Edward Arnold, London, UK.
7. Bruna-Romero, O., and A. Rodriguez. 2001. Dendritic cells can initiateprotective immune responses against malaria. Infect. Immun. 69:5173–5176.
8. Camargo, M. M., I. C. Almeida, M. E. S. Pereira, M. A. J. Ferguson, L. R.Travassos, and R. T. Gazzinelli. 1997. GPI-anchored mucin-like glycopro-tein isolated from Trypanosoma cruzi initiate the synthesis of proinflamma-tory cytokines by macrophages. J. Immunol. 158:5890–5901.
9. Campos, M. A. S., I. C. Almeida, O. Takeuchi, et al. 2001. Activation ofToll-like receptor-2 by glycosylphosphatidylinositol anchors from a proto-zoan parasite. J. Immunol. 167:416–423.
10. Delude, R. L., A. Yoshimura, R. R. Ingalls, and D. T. Golenbock. 1998.Construction of a lipopolysaccharide reporter cell line and its use in identi-fying mutants defective in endotoxin, but not TNF-�, signal transduction.J. Immunol. 161:3001–3009.
11. Erridge, C., E. Bennett-Guerrero, and I. R. Poxton. 2002. Structure andfunction of lipopolysaccharides. Microbes Infect. 4:837–851.
12. Folch, J., M. Lees, and G. H. S Stanley. 1957. A simple method for theisolation and purification of total lipids from animal tissues. J. Biol. Chem.226:497–504.
13. Forestier, C., F. Deleuil, N. Lapaque, E. Moreno, and J. Gorvel. 2000.Brucella abortus lipopolysaccharide in murine peritoneal macrophages actsas a down-regulator of T-cell activation. J. Immunol. 165:5202–5210.
14. Galanos, C., O. Luderitz, E. T. Rietschel, O. Westphal, H. Brade, L. Brade,M. Freundenberg, U. Schade, M. Imoto, and H. Yoshimura. 1985. Syntheticand natural E. coli free lipid A express identical entotoxic activities. Eur.J. Biochem. 148:1–5.
15. Golding, B., N. Eller, L. Levy, P. Beining, J. Inman, N. Matthews, D. E. Scott,and H. Golding. 2002. Mucosal immunity in mice immunized with HIV-1peptide conjugated to Brucella abortus. Vaccine 20:1445–1450.
16. Goldstein, J., T. Hoffman, C. Frasch, E. F. Lizzio, P. R. Beining, D. Hoch-stein, Y. L. Lee, R. D. Angus, and B. Golding. 1992. Lipopolysaccharide(LPS) from Brucella abortus is less toxic than that from Escherichia coli,suggesting the possible use of B. abortus and LPS from B. abortus as a carriervaccines. Infect. Immun. 60:1385–1389.
17. Hailman, E., H. S. Lichenstein, M. M. Wurfel, D. S. Miller, D. A. Johnson,M. Kelley, L. A. Busse, M. M. Zukowski, and S. D. Wright. 1994. Lipopoly-saccharide (LPS)-binding protein accelerates the binding of LPS to CD14. J.Exp. Med. 179:269–277.
18. Henneke, P., O. Takeuchi, J. A. Strijp, H-K Guttormsen, J. A. Smith, A. B.Schromm, T. A. Espevik, S. Akira, V. Nizet, D. L. Kasper, and D. T. Golen-bock. 2001. Novel engagement of CD14 and multiple Toll-like receptors bygroup B Streptococci. J. Immunol. 167:7069–7076.
19. Hirschfeld, M., C. J. Kirschning, R. Schwandner, H. Wesche, J. H. Weis,R. M. Wooten, and J. J. Weis. 1999. Cutting edge: inflammatory signaling byBorrelia burgdorferi lipoproteins is mediated by Toll-like receptor 2. J. Im-munol. 163:2382–2386.
20. Hirschfeld, M., J. J. Weis, V. Toshchakov, C. A. Salkowski, M. J. Cody, D. C.Ward, N. Qureshi, S. M. Michalek, and S. N. Vogel. 2001. Signaling bytoll-like receptor 2 and 4 agonists results in differential gene expression inmurine macrophages. Infect. Immun. 69:1477–1482.
21. Hirschfeld, M., Y. Ma, J. H. Weis, S. N. Vogel, and J. J. Weis. 2000. Repu-
rification of lipopolysaccharide eliminates signaling through both human andmurine toll-like receptor 2. J. Immunol. 165:618–622.
22. Hoshino, K., O. Takeuchi, T. Kawai, H. Sanjo, T. Ogawa, Y. Takeda, K.Takeda, and S. Akira. 1999. Toll-like receptor 4 (TLR4)-deficient mice arehyporesponsive to lipopolysaccharide: evidence for TLR4 as the Lps geneproduct. J. Immunol. 162:3749–3752.
23. Jarvis, B. W., and N. Qureshi. 1997. Inhibition of lipopolysaccharide-in-duced transcription factor Sp1 binding by spectrally pure diphosphoryl lipidA from Rhodobacter sphaeroides, protein kinase inhibitor H-8, and dexa-methasone. Infect. Immun. 65:1640–1643.
24. Jarvis, B. W., H. Lichenstein, and N. Qureshi. 1997. Diphosphoryl lipid Afrom Rhodobacter sphaeroides inhibits complexes that form in vitro betweenlipopolysaccharide (LPS)-binding protein, soluble CD14, and spectrally pureLPS. Infect. Immun. 65:3011–3016.
25. Jiang, Q., S. Akashi, K. Miyake, and H. R. Petty. 2000. Lipopolysaccharideinduces physical proximity between CD14 and Toll-like receptor 4 (TLR4)prior to nuclear translocation of NF-�B J. Immunol. 165:3541–3544.
26. Ko, J., A. Gendron-Fitzpatrick, and G. A. Splitter. 2002. Susceptibility ofIFN Regulatory Factor-1 and consensus sequence binding protein-deficentmice to brucellosis. J. Immunol. 168:2433–2440.
27. Kopp, E. B., and R. Medzhitov. 1999. The Toll-receptor family and controlof innate immunity. Curr. Opin. Immunol. 11:13–18.
28. Liautard, J. P., A. Gross, J. Dornand, and S. Kohler. 1996. Interactionsbetween professional phagocytes and Brucella spp. Microbiologia 12:197–206.
29. Lien, E., T. J. Sellati, A. Yoshimura, T. H. Flo, G. Rawadi, R. W. Finberg,J. D. Carroll, T. Espevik, R. R. Ingalls, J. D. Radolf, and D. T. Golenbock.1999. Toll-like receptor 2 functions as a pattern recognition receptor fordiverse bacterial products. J. Biol. Chem. 274:33419–33425.
30. Lien, E., T. K. Means, H. Heine, A. Yoshimura, S. Kusumoto, K. Fukase, M.Fenton, M. Oikawa, N. Qureshi, B. Monks, R. W. Finberg, R. R. Ingalls, andD. T. Golenbock. 2000. Toll-like receptor 4 imparts ligand-specific recogni-tion of bacterial lipopolysaccharide. J. Clin. Investig. 105:497–504.
31. Lorenz, E., D. D. Patel, T. Hartung, and D. A. Schwartz. 2002. Toll-likereceptor 4 (TLR4)-deficient murine macrophage cell line as an in vitro assaysystem to show TLR4-independent signaling of Bacteroides fragilis lipopoly-saccharide. Infect. Immun. 70:4892–4896.
32. Means, T. K., S. Wang, E. Lien, A. Yoshimura, D. T. Golenbock, and M. J.Fenton. 1999. Hum. Toll-like receptors mediate cellular activation by Myco-bacterium tuberculosis. J. Immunol. 163:3920–3927.
33. Medzhitov, R., Jr., and C. A. Janeway. 1997. Innate immunity: the virtues ofa monoclonal system of recognition. Cell 91:295–298.
34. Medzhitov, R., P. Preston-Hurburt, Jr., and C. A. Janeway. 1997. A humanhomologue of the Drosophila Toll protein signals activation of adaptiveimmunity. Nature 388:394–397.
35. Moreno, E., M. W. Pitt, L. M. Jones, G. G. Schurig, and D. T. Berman. 1979.Purification and characterization of smooth and rough lipopolysaccharidesfrom Brucella abortus. J. Bacteriol. 138:361–369.
36. Moriyon, I., and I. Lopez-Goni. 1998. Structure and properties of the outermembranes of Brucella abortus and Brucella melitensis. Int. Microbiol. 1:19–26.
37. Morrison, D. C., and J. L. Ryan. 1987. Endotoxins and disease mechanisms.Annu. Rev. Med. 38:417–432.
38. Murphy, E. A., M. Parent, J. Sathiyaseelan, X. Jiang, and C. L. Baldwin.2001. Immune control of Brucella abortus 2308 infections in BALB/c mice.FEMS Immunol. Med. Microbiol. 32:85–88.
39. Murphy, E. A., J. Sathiyaseelan, M. Parent, B. Zou, and C. L. Baldwin. 2001.Interferon- is crucial for surviving a Brucella abortus infection in bothresistant C57BL/6 and susceptible BALB/c mice. 2001. Immunology 103:511–518.
40. Oliveira, S. C., and G. A. Splitter. 1995. CD8� type 1 CD44hi CD45RBlo Tlymphocytes control intracellular Brucella abortus infection as demonstratedin major histocompatibility complex class-I and class II-deficient mice. Eur.J. Immunol. 25:2551–2557.
41. Perera, P. Y., T. N. Mayadas, O. Takeuchi, S. Akira, M. Zaks-Zilberman,S. M. Goyert, and S. N. Vogel. 2001. CD11b/CD18 acts in concert with CD14and Toll-like receptor (TLR) 4 to elicit full lipopolysaccharide and taxol-inducible gene expression. J. Immunol. 166:574–581.
42. Qureshi, N., J. Hofman, K. Takayama, S. N. Vogel, and D. C. Morrison.1996. Diphosphoryl lipid A from Rhodobacter sphaeroides, a novel lipopoly-saccharide antagonist. In D. C. Morrison and R. L. Ryan (ed.), Noveltherapeutic strategies in the treatment of sepsis. Marcel Dekker, Inc., NewYork, N.Y.
43. Qureshi, N., J. P. Honovich, H. Hara, R. J. Cotter, and K. Takayama. 1988.Location of fatty acids in lipid A obtained from lipopolysaccharide of Rho-dopseudomonas sphaeroides ATCC 17023. J. Biol. Chem. 263:5502–5504.
44. Qureshi, N., K. Takayama, U. Seydel, R. Wang, R. J. Cotter, P. K. Agrawal,C. A. Bush, R. Kurtz, and D. T. Berman. 1994. Structural analysis of the lipidA derived from the lipopolysaccharide of Brucella abortus. J. Endotox. Res.1:137–148.
45. Rock, F. L., G. Hardiman, J. C. Timans, R. A. Kastelein, and J. F. Bazan.
VOL. 72, 2004 TLR4 SIGNALING DURING BRUCELLA INFECTION 185

1998. A family of human receptors structurally related to Drosophila Toll.Proc. Natl. Acad. Sci. 95:588–593.
46. Rosinha, G. M. S., D. A. Freitas, A. Miyoshi, V. Azevedo, E. Campos, S. L.Cravero, O. Rossetti, G. Splitter, and S. C. Oliveira. 2002. Identification andcharacterization of a Brucella abortus ATP-binding cassette transporter ho-molog to Rhizobium meliloti ExsA and its role in virulence and protection inmice. Infect. Immun. 70:5036–5044.
47. Sangari, F. J., and J. Aguero. 1996. Molecular basis of Brucella pathogenic-ity: an update. Microbiologia 12:207–218.
48. Schwandler, R., R. Dziarski, H. Wesche, M. Rothe, and C. J. Kirschning.1999. Peptidoglycan- and lipoteichoic acid-induced cell activation is medi-ated by Toll-like receptor 2. J. Biol. Chem. 274:17406–17409.
49. Stevens, M. G., S. C. Olsen, G. W. Pugh, Jr., and D. Brees. 1995. Comparisonof immune responses and resistance to brucellosis in mice vaccinated withBrucella abortus 19 and RB51. Infect. Immun. 63:264–270.
50. Takeuchi, O., K. Hoshino, and S. Akira. 2000. Cutting edge: TLR2-deficientand MyD88-deficient mice are highly susceptible to Staphylococcus aureusinfection. J. Immunol. 165:5392–5396.
51. Takeuchi, O., T. Kawai, H. Sanjo, N. G. Copeland, D. J. Gilbert, N. A.Jenkins, K. Takeda, and S. Akira. 1999. TLR6: a novel member of anexpanding Toll-like receptor family. Gene 231:59–65.
52. Takeuchi, O., K. Hoshino, T. Kawai, H. Sanjo, H. Takahada, T. Ogawa, K.Takeda, and S. Akira. 1999. Differential roles of TLR2 and TLR4 in recog-nition of gram negative and gram-positive bacterial cell wall components.Immunity 11:443–451.
53. Underhill, D. M., A. H. Ozinsky., M. Adeline, A. Stevens, C. B. Wilson, M.Basseti, and A. Aderem. 1999. The Toll-like receptor 2 is recruited to mac-rophage phagosomes and discriminates between pathogens. Nature 401:811–815.
54. Vasselon, T., and P. A. Detmers. 2002. Toll receptors: a central element ininnate immune responses. Infect. Immun. 70:1033–1041.
55. Visintin, A., A. Mazzoni, J. A. Spitzer, and D. M. Segal. 2001. Secreted MD-2is a large polymeric protein that efficiently confers lipopolysacharide sensi-tivity to Toll-like receptor 4. Proc. Natl. Acad. Sci. 98:12156–12161.
56. Winter, A. J., G. G. Schurig, S. M. Boyle, N. Sriranganathan, J. S. Bevins,F. M. Enright, P. H. Elzer, and J. D. Kope. 1996. Protection of BALB/c miceagainst homologous and heterologous species of Brucella by rough strainvaccines derived from Brucella melitensis and Brucella suis biovar 4. Am. J.Vet. Res. 57:677–683.
57. Yoshimura, A., E. Lien, R. R. Ingalls, E. Tuomanen, R. Dziarski, and D. T.Golenbock. 1999. Cutting edge: recognition of gram-positive bacterial cellwall components by the innate immune system occurs via Toll-like receptor2. J. Immunol. 163:1–5.
58. Young, E. J. 1983. Human brucellosis. Rev. Infect. Dis. 5:821–842.59. Zaitseva, M., H. Golding, J. Manischewitz, D. Webb, and B. Golding. 1996.
Brucella abortus as a potential vaccine candidate: induction of interleukin-12secretion and enhanced B7.1 and B7.2 and intercellular adhesion molecule 1surface expression in elutriated human monocytes stimulated by heat-inac-tivated B. abortus. Infect. Immun. 64:3109–3119.
60. Zaitseva, M., L. R. King, J. Manischewitz, M. Dougan, L. Stevan, H. Gold-ing, and B. Golding. 2001. Human peripheral blood T cells, monocytes, andmacrophages secrete macrophage inflammatory proteins 1a and 1b followingstimulation with heat-inactivated Brucella abortus. Infect. Immun. 69:3817–3826.
61. Zhan, Y., and C. Cheers. 1998. Control of IL-12 and IFN- production inresponse to live and dead bacteria by TNF and other factors. J. Immunol.161:1447–1453.
62. Zinkernagel, R. M. 1974. Restriction by H-2 gene complex of transfer ofcell-mediated immunity to Listeria monocytogenes. Nature 251:230–233.
Editor: J. T. Barbieri
186 CAMPOS ET AL. INFECT. IMMUN.




















