
Antología musical de la Catedral de Jaca en el siglo XVIII (Huesca, 2002)
Upload
independentCategory
view
3download
0
BIOLOGICAL SOIL CRUST BIOWEATHERING 1651
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
Earth Surface Processes and Landforms
Earth Surf. Process. Landforms 29, 1651–1661 (2004)Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/esp.1118
ROLE OF BIOLOGICAL SOIL CRUST COVER IN BIOWEATHERING
AND PROTECTION OF SANDSTONES IN A SEMI-ARID LANDSCAPE
(TORROLLONES DE GABARDA, HUESCA, SPAIN)
VIRGINIA SOUZA-EGIPSY1*, JACEK WIERZCHOS2, CARLOS SANCHO3, ANCHEL BELMONTE3
AND CARMEN ASCASO1
1 Centro Ciencias Medioambientales, CSIC, Serrano 115 dpdo, Madrid 28006, Spain2 Servei de Microscópia Electrònica, Universitat de Lleida, Rovira Roure 44, Lleida 25196, Catalunya, Spain
3 Dpto. Ciencias de la Tierra (Geomorfología), Facultad de Ciencias, Universidad de Zaragoza, Pedro Cerbuna 12,
Zaragoza 50009, Spain
Received 19 March 2003; Revised 18 March 2004; Accepted 20 April 2004
ABSTRACT
Sandstone structural landscapes in the semi-arid Torrollones de Gabarda area (Province of Huesca, NE Spain) are oftencovered by a well developed biological soil crust of lichens, mosses and cyanobacteria and black coatings on verticalsurfaces. By using scanning electron microscopy with backscattered detector imaging, the biological soil crust studiedevidenced high activity in the sandstone–crust interface. Processes such as physical disintegration, etching and dwelling aswell as biomineralization by calcium oxalate and fixation of mineral particles by extracellular polymeric substances wereobserved. On the horizontal sandstone surfaces these processes may cause the occurrence of gnammas and the developmentof a protective coating that favours intense flaking when the crust is disturbed. On the sandstone cliffs, columnar and tafoniweathering development is clearly guided by the protective action of the biological soil crust. These qualitative observationsare important to develop methodologies to address their quantitative importance in geomorphological processes in semi-aridlandscapes. Copyright © 2004 John Wiley & Sons, Ltd.
KEY WORDS: biological soil crust; micro-weathering; lichen; cyanobacteria; fungi; moss
INTRODUCTION
There is no doubt that biological soil crusts play an important role as agents of weathering on natural rock
surfaces in dry environments (Cooke et al., 1993; Chen et al., 2000). Despite their importance and the research
works carried out in arid and semi-arid zones (Schipull, 1978; Danin and Garty, 1983; Wessels and Schoemann,
1988; Wessels and Büdel, 1995; Mellor et al., 1997; Paradise, 1997; Robinson and Williams, 2000), we are still
far from a general knowledge of the relationships between biological soil crusts, weathering processes and
resulting landforms (Viles and Pentecost, 1994). The application of more specialized analytical techniques as
well as the work of multidisciplinary scientific teams can help us to understand the effect of biological soil
coatings in rock weathering processes (Adamo and Violante, 2000). Many studies concerned with the long-term
stability of architectural materials have contributed greatly to defining communities of organisms and the dam-
age they cause to natural and synthetic materials (see reviews in Warsteid and Braams, 2000; Sterflinger, 2000).
Geomorphologists deal with the relief features of the Earth and seek a genetic interpretation of them. In this
regard geomicrobiologists should make careful observations of the phenomena from the macroscopic to the
microscopic scale, bringing together the knowledge of biologists, chemists, geologists and physicists, in order
to understand the biological impact on the alteration of natural rock.
Detailed geomorphological field recognition of the area studied (Sancho and Belmonte, 2000) indicates an
evident relationship between microweathering features (columnar weathering, gnammas, tafoni and flaking) and
biological soil crusts. Thus it is of geomorphological interest to understand the role of biological soil crust
activity in the nucleation and development of these micromorphologies. However, we have little information
* Correspondence to: V. Souza-Egipsy, Centro de Astrobiología CSIC-INTA, Ctr. Torrejón a Ajalvir Km4, Torrejón de Ardoz 28850,Madrid, Spain. E-mail: [email protected]

1652 V. SOUZA-EGIPSY ET AL.
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
about the processes controlling this relationship. Recent reviews in the literature about biological soil crusts
(Belnap and Lange, 2001) still record some apparently contradictory information on the role of biological soil
crusts in ecosystem processes. We think that some of these contradictory results can be explained by careful
microscale investigations of the biological soil crust–sandstone interface.
SITE DESCRIPTION
The study area is located in the Torrollones de Gabarda included in the Monegros region (Huesca Province,
Central Ebro Basin, NE Spain) (see Figure 1). The altitude in the area of study ranges from 200 to 350 m.
Climate is semi-arid–Mediterranean with a strong seasonal contrast (Ascaso and Cuadrat, 1981; Del Valle,
1996). The mean annual temperature is 13·5 °C and the annual evapotranspiration ranges between 700 and
800 mm. The mean annual precipitation is 445 mm with about 70 rainy days per year. Fog and dew are present
during 27 and 45 days per year, respectively. From a geological point of view, the bedrock in the study area
is made up by fluvial sandstones and clays corresponding to the Miocene Sariñena Formation (García et al.,
1991). Optical microscopy analysis has shown that sandstones are heterometric, fine- to medium-grained
lithoarenites with intact framework and intergranular contacts. The sandstone’s porosity is 17 per cent, and it
contains fragments of shale (40 per cent: illite, kaolinite and chlorite in order of decreasing abundance), lime-
stone (22 per cent), quartz (29 per cent) and feldspars (9 per cent). Scarce calcite cement and clay matrix
(muscovite and biotite) were observed. Mineralogical composition was supported by X-ray analysis. Quaternary
fluvial incision has carved the mesas and towers in sandstones to form a spectacular semi-arid landscape (Fig-
ure 2). As a consequence, horizontal as well as vertical sandstone surfaces are exposed and are affected by
intense weathering processes. Numerous tafoni and alveoli on the sandstone cliffs and gnammas on the horizontal
surfaces have developed (Sancho and Belmonte, 2000). A continuous biological soil crust covers exposed
Figure 1. Map of the studied area

BIOLOGICAL SOIL CRUST BIOWEATHERING 1653
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
horizontal and vertical surfaces. From a macroscopic point of view two different situations were recognized
during the field work: (1) widespread biological soil crusts, made up mostly by blackish crust, lichens and
mosses, cover horizontal surfaces, and (2) black rock coatings consisting of fungi and cyanolichens run down
the side of the sandstone bedrock cliffs.
MATERIAL AND METHOD
Pieces of the sandstone from horizontal and vertical surfaces bearing biological soil crust were hand-cut using
a chisel. Horizontal surfaces were covered practically 100 per cent, whereas the vertical surfaces were covered
from 50 to 70 per cent.
Three main components of the biological soil crust were identified in the laboratory: a green algal crustose
lichen, Diploschistes actinostomus (Ach.) Zahlbr.; a cyanolichen Collema flaccidum (Ach.) Ach.; and a moss,
Grimmia orbicularis Bruch. In the laboratory, five selected transversal sections of the biological soil crust were
studied: three with examples of the main components from the horizontal surfaces plus two black undifferentiated
biological soil crusts, one from a horizontal surface and one from a vertical surface. After selection they were
processed following conventional transmission electron microscopy procedures for the preparation of lichen
material (De los Rios and Ascaso, 2001). This includes fixation with 2·5 per cent glutaraldehyde in buffer
phosphate 0·1 M, a post-fixation with 1 per cent osmium tetroxide, and a dehydration stage involving application
of saturated uranyl acetate solution in 70 per cent ethanol. Samples were embedded in LR-White (London Resin)
high-viscosity resin to improve visualization of ultrastructural details. The transversal surface of the biological
soil crust–rock interface was finely polished and then coated with evaporated carbon. Detailed information on
this preparation technique and visualization method has been reported in previous studies (Ascaso and Wierzchos,
1994; Wierzchos and Ascaso, 1994). Samples were examined by scanning electron microscopy with backscattered
electron imaging (SEM-BSE) using a Zeiss 960 DSM SEM microscope equipped with a BSE detector and an
energy dispersive X-ray (EDS) Link ISIS (Oxford) microanalytical system.
RESULTS
Horizontal sandstone surfaces
Horizontal sandstone surfaces are well covered by biological soil crust. The crust on these surfaces is formed
mostly by lichens and mosses as shown in Figure 3. Locally, some microweathering features such as rock-basins
Figure 2. General view of the studied area

1654 V. SOUZA-EGIPSY ET AL.
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
Figure 3. General aspect of the horizontal surfaces with gnamma formation
or gnammas have been observed. In addition, flaking affecting sandstone surfaces is frequent when trampling
disturbs the biological soil crust.
The scanning electron micrographs presented in this study were taken using the detection of the backscattered
electron (BSE) signal which is strongly dependent on the atomic number of the target (Robinson and Nickel, 1979;
Wierzchos et al., 1995). When using BSE in examining a flat rock specimen the compositional information domin-
ates the topographical information allowing for easy distinction of the resin in the pores (black in Figure 4A)
from the different soil mineral particles and the biological material (different contrast, see Figure 4A).
D. actinostomus thalli (UC, upper cortex; AL, algal layer and M, medulla; Figure 4A) form a continuous crust
over this type of substratum. The lichen medulla revealed a great amount of microdivided mineral fragments and
in the interface between the thallus and the substratum it was possible to observe how potassium-rich laminated
minerals were actively disintegrated by the growth of hyphae (asterisks in Figure 4A). The organic material can
be readily distinguished as a result of the greater absorbance of the heavy metals used for fixing and staining
during the preparation procedure. The black areas on Figure 4B are empty spaces filled with resin. Between the
bright mineral fragments, there are filaments with less contrast that correspond to hyphae. Some of them are cut
in longitudinal section (lower arrow in Figure 4B) and others in transverse section (upper arrow in Figure 4B).
The hyphae (arrows in Figure 4B) produce an intense disintegration on the potassium-rich mineral fragments.
Through the formation of calcium oxalate crystals at the medulla and upper cortex, hyphae of D. actinostomus
thalli developed an important biomineralization process. Figure 4C shows one of the areas (UC, upper cortex;
AL, algal layer and M, medulla) in which digital mapping of silica (Figure 4D) and calcium has been obtained
(Figure 4E). The calcium distribution indicates that the whitish colour is related to the presence of calcium
oxalate crystals in the lichen (L in Figure 5).
C. flaccidum is one of the typical jelly-like cyanolichens in the blackish biological soil crust (asterisk in
Figure 5). The thallus of these cyanolichens is homiomerous without a clear upper cortex or medulla differentia-
tion, and forms a discontinuous crust over the surface (cf in Figure 6A). Note the difference in particle size under
the thalli with respect to the green algal lichens. This type of lichen thallus does not hold mineral fragments
inside. However, some mineral grains have adhered to the surface of the thallus (arrows in Figure 6A).
In between these two types of macroscopic thalli there was also a black biological soil crust of micro-
organisms without clear patterns of growth. These crusts, which fill the voids between the main thalli, have been
formed by lichen thalli in their initial stages of growth (pl in Figure 6B) in which it was possible to distinguish
the photosynthetic and the mycobiont part of the symbiosis. The mycobiont has been shown to be the main
participant in the weathering of the surface. The physico-chemical action of hyphae has been observed in the

BIOLOGICAL SOIL CRUST BIOWEATHERING 1655
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
Figure 4. SEM-BSE images of Diploschistes actinostomus growing on a horizontal surface. (A) The thallus: UC, upper cortex; Al, algallayer; M, medulla. The zone marked by the square has been extended in (B). The asterisks indicate potassium-rich rock fragments that areactively fragmented by the action of the lichen. Scale bar = 500 µm. (B) The growth of hyphae in the rock fragment causes disintegration.The lower arrow indicates a longitudinal section and the upper arrow a cross-section of the hyphae between the mineral particles. Scale bar= 20 µm. (C) Image of the zone of the lichen thallus in which digital mapping has been made. The thallus of D. actinostomus producesa superficial crust (UC) and accumulates a great amount of calcium oxalate crystals between medullary hyphae (M), leaving the algal layer
free of minerals (AL). (D) Map of distribution of silica (Si). (E) Map of the distribution of calcium (Ca)
Figure 5. Detail of the biological soil crust formed by lichens (L) and mosses (M). Asterisk indicates the area occupied by the blackishundifferentiated crust

1656 V. SOUZA-EGIPSY ET AL.
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
Figure 6. (A) SEM-BSE image of Collema flaccidum on a horizontal surface (cf). The arrows show mineral particles adhering to the surfaceof the thallus but they did not appear in the interior. Scale bar = 1 mm. (B) Aspect of the black crust over the substratum between the lichenthalli. The arrows indicate the aspect of minerals in contact with micro-organisms (pt). Scale bar = 50 µm. (C) SEM-BSE image of Grimmia
orbicularis growing over a horizontal surface. The moss cushion grows on a great amount of microdivided minerals that were retainedbetween the organic parts (asterisks). The arrows indicate the weathered surface of different minerals in contact with the rhizoids. Scale bar= 200 µm. (D) Between the leaves (L) there were large amounts of mineral fragments. The brighter zones such as leaves (L) and the apex
of growth (a) indicate a greater metabolic activity than the darkest zones in the stem (s). Scale bar = 100 µm
formation of etched surfaces (white arrow in Figure 6B) and disintegration (black arrows in Figure 6B) of
substrate minerals.
In the case of moss cushions such as G. orbicularis (M in Figure 5) the disintegrating action in the substrate
was observed in the laminated potassium-rich mineral fragments (k) at a higher level than in the quartz (q) grains
(arrows in Figure 6C). These cushions contain (asterisks in Figure 6C) great quantities of mineral fragments.
This phenomenon occurs from the zones in contact with the substratum (bottom left corner in Figure 6C) to the
surface between the leaves (L in Figure 6D). The zones with high metabolic activity, such as the apex of the
stem and leaves, present a brighter aspect in the BSE signal due to the osmium and uranyl acetate stain applied
during the sample preparation.
After careful observation of the horizontal surfaces with a black crust it was possible to describe different
micro-organism weathering processes. Figure 7A shows how the calcite fragments (Ca in Figure 7A) colonized
by euendolithic hyphae were developing channels (arrow in Figure 7A). Between the mineral particles it was
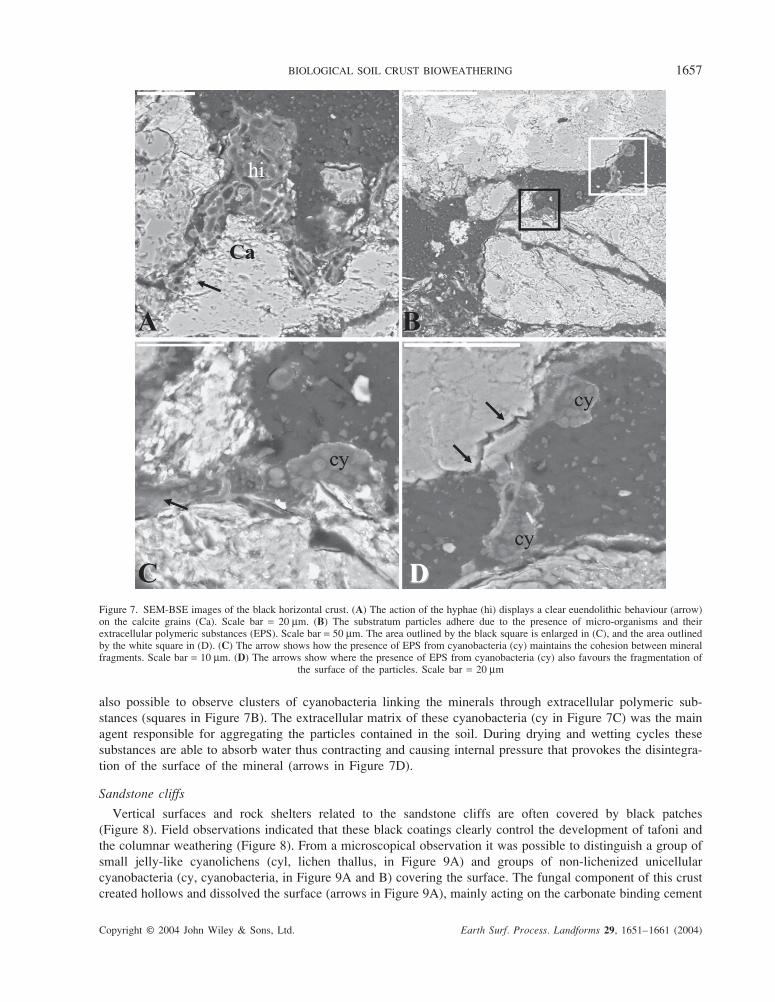
BIOLOGICAL SOIL CRUST BIOWEATHERING 1657
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
also possible to observe clusters of cyanobacteria linking the minerals through extracellular polymeric sub-
stances (squares in Figure 7B). The extracellular matrix of these cyanobacteria (cy in Figure 7C) was the main
agent responsible for aggregating the particles contained in the soil. During drying and wetting cycles these
substances are able to absorb water thus contracting and causing internal pressure that provokes the disintegra-
tion of the surface of the mineral (arrows in Figure 7D).
Sandstone cliffs
Vertical surfaces and rock shelters related to the sandstone cliffs are often covered by black patches
(Figure 8). Field observations indicated that these black coatings clearly control the development of tafoni and
the columnar weathering (Figure 8). From a microscopical observation it was possible to distinguish a group of
small jelly-like cyanolichens (cyl, lichen thallus, in Figure 9A) and groups of non-lichenized unicellular
cyanobacteria (cy, cyanobacteria, in Figure 9A and B) covering the surface. The fungal component of this crust
created hollows and dissolved the surface (arrows in Figure 9A), mainly acting on the carbonate binding cement
Figure 7. SEM-BSE images of the black horizontal crust. (A) The action of the hyphae (hi) displays a clear euendolithic behaviour (arrow)on the calcite grains (Ca). Scale bar = 20 µm. (B) The substratum particles adhere due to the presence of micro-organisms and theirextracellular polymeric substances (EPS). Scale bar = 50 µm. The area outlined by the black square is enlarged in (C), and the area outlinedby the white square in (D). (C) The arrow shows how the presence of EPS from cyanobacteria (cy) maintains the cohesion between mineralfragments. Scale bar = 10 µm. (D) The arrows show where the presence of EPS from cyanobacteria (cy) also favours the fragmentation of
the surface of the particles. Scale bar = 20 µm

1658 V. SOUZA-EGIPSY ET AL.
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
Figure 8. General view of the black coatings on vertical surfaces
Table I. Elemental composition (%) of the laminated minerals in contact with the superficialhyphae in Figure 9B
Mg Al Si K Ca Ti Fe
Biotite (external) 3 6·76 16·44 2·71 4·32 0·78 9·86Biotite (central) 4 7·92 16·02 3·80 2·43 0·99 12·30Muscovite 3·19 12·08 11·93 – 1·35 0·13 17·13
(Ca, calcite, in Figure 9A). Black hyphae were part of the biological soil crust and in Figure 9B appeared to
be attacking the crystal surface of a laminated mineral. The physical disruption of the lamination increased the
area of the reactive surface and exposed fresh material, thus increasing the chemical and physical weathering
processes (arrows in Figure 9B). A microanalytical study of this crystal showed that it was not homogeneous; the
central part, with higher contrast than the external part, was muscovite whereas the right side corner was a biotite
(Table I). The digital mapping of the zone inside the square in Figure 9B shows how the siliceous structure
(arrow in Figure 9C) had lost part of the potassium present in the rest of the biotite (arrow in Figure 9D).
DISCUSSION
Examination of biological soil crust ultrastructure can greatly assist in the understanding of its distribution,
abundance and function on microweathering and finally in its influence in the landscape geomorphology. We
propose an appropiate technique for microscale investigations that has proved to be useful in the study of
physical and chemical differences in the lichen species that alter some features of the surfaces (Souza-Egipsy
et al., 2002a,b).
In the case of a biological soil crust on horizontal surfaces, gnammas or rock-basins are the most frequent
weathering micromorphology. Two actions by biological soil crust can be differentiated in the gnamma forma-
tion: first, the etching associated with chemical dissolution from the lichen thallus; and second, disintegration
processes related to hyphae penetration and thallus expansion and contraction during wetting and drying cycles.
Both favour a differential weathering affecting the sandstone surface and the beginning of the nucleation of
gnammas. Schipull (1978), under similar environmental and lithological conditions in Colorado Plateau (Utah,
USA), reported that together with the geochemical dissolution the action of lichen and mosses in the formation
of gnammas or microdepressions must be taken into account. In addition, Robinson and Williams (2000)
described circular microdepressions on sandstone surfaces in Morocco and discussed the processes by which

BIOLOGICAL SOIL CRUST BIOWEATHERING 1659
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
Figure 9. SEM-BSE images of biological soil crust growing over a northern exposure vertical wall. (A) The organic compound of thecyanolichens (cyl) covered the surface homogeneously and under the surface the calcite cement (Ca) was deeply dissolved (arrows). Thereare groups of non-lichenized cyanobacteria (cy). Scale bar = 100 µm. (B) The action of hyphae on laminar minerals as biotite (Bi) andmuscovite (Mu) was fundamentally physical; the arrows point to parts of the mineral that were altered. There are non-lichenized cyanobacteria(cy). Scale bar = 50 µm. (C) Silica digital mapping (Si). (D) Potassium digital mapping. Note the different distribution of potassium in
contact with the hyphae (arrow); there is a loss of potasium from the biotite surface
lichens attack sandstone. With our approach we have in situ visualization of the mechanisms implicated in the
weathering processes.
The internal structure of crustose green algal lichen and cyanolichens in the horizontal biological soil crust
reduces the possibility of soil loss by water or wind erosion with different mechanisms. Airborne silt and clay
particles are trapped by sticky extracellular polymeric substances (EPS) around cyanolichens, non-lichenized
cyanobacteria and fungi, whereas crustose lichens hold fine particles around the hyphae from the medulla and
penetrate the substrate a few micrometres under the surface. The amount of soil stabilization depends on the type
of lichens that colonize the substrate.
The external morphology of the biological soil crust determines the times of generation of ponding and runoff
of the surfaces (Alexander and Calvo, 1990) but the differences between species are determined by their internal
structure. The surfaces covered by green algal lichens such as Diploschistes have a protective layer of calcium
oxalate crystals that shield the surface of the substrate from external environmental factors such as wind and splash
erosion from rain events. However, the cyanolichens and free-living coccoid cyanobacteria have EPS. This sheath

1660 V. SOUZA-EGIPSY ET AL.
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
material sticks to the surfaces of soil particles, forming a web of fibres covering the soil. In this way, loose soil
particles are protected, and otherwise unstable and highly erosion-prone surfaces become resistant to both wind and
water erosion. The EPS are able to hold greater amounts of water, decreasing the capacity for ponding and runoff
generation, whereas Diploschistes surfaces have been described as quite impervious (Souza-Egipsy et al., 2002a,b).
Despite the different ways that sediment coheres, these two types of components are highly susceptible to soil-
surface disturbance such as trampling by hooves or feet, or driving of off-road vehicles, especially in soils with
low aggregate stability (Watts, 1998). Cyanobacteria, lichens and mosses are brittle when dry, and crush easily
when subjected to compressional or shear forces by activities such as trampling or vehicular traffic, or wind
erosion (Williams et al., 1994). Thus their protective capability is also very fragile and intense flaking is
observed in sandstone surfaces where the biological soil crust is disturbed.
In our samples the black coatings on vertical surfaces are formed by an irregular biological soil crust of fungi
and cyanobacteria, as in previous studies (Krumbein and Jens, 1981). Areas with visible black coatings were
more resistant to sediment erosion than adjacent areas without obvious microbial colonization, producing the
columnar aspect in the vertical surfaces of the tafoni. Field observations indicate that biological soil crusts
covering vertical surfaces (sandstone cliff and shelters) are clearly controlled by water running down. Several
authors have noted the influence of the rock varnish crust on the case-hardening processes affecting the outer
sandrock surfaces that control the development of the tafoni in arid regions (Cooke et al., 1993; Viles and
Pentecost, 1994; Mellor et al., 1997) and columnar weathering morphologies (Howard and Selby, 1994). In
addition, others studies (Mustoe, 1983; Ariño et al., 1995) show the protective role of the biological soil crust
and rock varnish on the underlying rock from environmental factors.
Other important components observed in the biological soil crust were moss cushions. While bryophytes do
not exactly form surface crusts in the field area studied, they do cover the sandstone regolith. It has long been
noted that mosses prefer north-facing slopes where water availability is highest (Alexander and Calvo, 1990) and
that they help to stabilize airborne dust (Danin and Ganor, 1991). Previous studies have focused attention on
archaeological sites with bryological flora (Alteri and Ricci, 1997), but as far as we know this is the first study
where the weathering action of the rhizoids has been seen in situ on a natural substrate. These cushions have
a geomorphological role as the mechanical component of the rhizoid network help to stabilize and hold soil
particles in place. To quantify these effect it will necessary to measure the amount of particles held between the
moss leaves. The protective effect on the substratum of the rhizinae and leaves network is more powerful than
the destructive effect of the bioweathering processes.
CONCLUSIONS
Scanning electron microscopy with an EDS microanalytical system in fixed and contrasted samples provided a
detailed characterization of the bioweathering and protective effects of biological soil crusts. The resulting
insight would not be achieved using other optical and conventional scanning microscopy techniques.
To evaluate the influence of biological soil crust on the microgeomorphology of a landscape it is necessary to
identify the importance of different components. Three main factors are important to characterize the relation of
each component on the microgeomorphology: the distribution, the protective role and the bioweathering effect.
The external morphology and internal structure of biological soil crust are very different from physical crusts
(Belnap, 2001). Microbial crusts can differ greatly in apperance, species composition and ecological function.
Futhermore, the internal features of the different biological soil crusts can determine how susceptible the
substrate is to bioweathering, and to water and wind erosion.
ACKNOWLEDGEMENTS
The authors wish to thank Dr I. Martinez and Dr G. Aragon, Department of Plant Biology, UCM, Madrid, for
their collaboration in taxonomical determination, and F. Pinto for helpful assistance in SEM. We also wish to
thank K. H. Nealson and J. Yu, Department of Earth Sciences, USC, for their assistance and support. This study
was supported by funds from the DGCYT (BOS2000-1121) and a grant from the Monegros Centro de Desarrollo
(FEOGA), Spain.

BIOLOGICAL SOIL CRUST BIOWEATHERING 1661
Copyright © 2004 John Wiley & Sons, Ltd. Earth Surf. Process. Landforms 29, 1651–1661 (2004)
REFERENCES
Alexander RW, Calvo A. 1990. The influence of lichens on slope processes in some Spanish badlands. In Vegetation and Erosion, ThornesJB (ed.). John Wiley: Chichester; 385–398.
Altieri A, Ricci S. 1997. Calcium uptake in mosses and its role in stone biodeterioration. International Biodeterioration & Biodegradation
40: 201–204.Adamo P, Violante P. 2000. Weathering of rocks and neogenesis of minerals associated with lichen activity. Applied Clay Science 16: 229–
256.Ariño X, Ortega-Calvo JJ, Gomez-Bolea A, Saiz-Jimenez C. 1995. Lichen colonization of the Roman pavement at Baelo Claudia (Cadiz,
Spain): biodeterioration vs. bioprotection. The Science of the Total Environment 167: 353–363.Ascaso A, Cuadrat JM. 1981. El clima. In Geografía de Aragón, Tomo I, Higueras A (eds). Guara Editorial: Zaragoza; 93–139.Ascaso C, Wierzchos J. 1994. Structural aspects of the lichen-rock interface using backscattered electron imaging. Botanica Acta 107: 251–
256.Belnap J. 2001. Comparative structure of physical and biological soil crusts. In Biological Soil Crusts: Structure, Function, and Manage-
ment, Belnap J, Lange OL (eds). Ecological Studies 150. Springer-Verlag: Berlin-Heidelberg; 177–180.Belnap J, Lange OL. 2001. Biological Soil Crusts: Structure, Function and Management. Ecological Studies 150. Springer-Verlag: Berlin-
Heidelberg.Chen J, Blume HP, Beyer L. 2000. Weathering of rocks induced by lichen colonization-a review. Catena 39: 121–146.Cooke RU, Waren A, Goudie AS. 1993. Desert Geomorphology. UCL Press: London.Danin A, Ganor E. 1991. Trapping of airborne dust by mosses in the Negev Desert, Israel. Earth Surface Processes and Landforms 16: 153–
162.Danin A, Garty J. 1983. Distribution of cyanobacteria and lichens on hillsides of the Negev Highlands and their impact on biogenic
weathering. Zeitschrift für Geomorphologie 27: 423–444.De los Ríos A, Ascaso C. 2001. Preparative techniques for Transmission Electron microscopy and Confocal Laser Scanning Microscopy
of lichens. In Protocols in Lichenology. Culturing, Biochemistry, Ecophysiology and Use in Biomonitoring, Kranner I, Beckett RP, VarmaAK (eds). Springer Lab Manual. Springer-Verlag: New York; 87–117.
Del Valle J. 1996. El clima del Prepirineo Central y Occidental aragonés y sus Somontanos. Serie Investigación 5. Consejo de Protecciónde la Naturaleza de Aragón: Aragón.
García JM, Sanz J, Samso JM. 1991. Mapa Geológico de España 1:50·000. Grañén 324. Instituto Tecnológico Geominero de España.Howard AD, Selby MJ. 1994. Rock slopes. In Geomorphology of Desert Environments, Abrahams A, Parsons AJ (eds). Chapman & Hall:
London; 135–172.Krumbein WE, Jens K. 1981. Biogenic rock varnishes of the Negev Desert (Israel) an ecological study of iron and manganese transformation
by cyanobacteria and fungi. Oecologia 50: 25–38.Mellor A, Short J, Kirkby J. 1997. Tafoni in the El Chorro area, Andalucia, Southern Spain. Earth Surface Processes and Landforms 22:
817–833.Mustoe GE. 1983. Cavernous weathering in the Capitol Reef Desert, Utah. Earth Surface Processes and Landforms 8: 517–526.Paradise TR. 1997. Disparate sandstone weathering beneath lichens, Red Mountain, Arizona. Geografiska Annaler 79A: 177–184.Robinson BM, Nickel EH. 1979. A useful new technique for mineralogy: the backscattered-electron low vacuum mode of SEM operation.
American Mineralogist 64: 1322–1328.Robinson DA, Williams RBG. 2000. Accelerated weathering of a sandstone in the High Atlas Mountains of Morocco by an epilithic lichen.
Zeitschrift für Geomorphologie 44: 513–528.Sancho Marcén C, Belmonte Ribas A. 2000. Bases geológicas, geomorfológicas, paisajísticas y arqueológicas para el aprovechamiento
cultural de La Plana de Mobache y los Torrollones de Gabarda (Monegros, Huesca). Serie Investigación 24. Consejo de Protección dela Naturaleza de Aragón. Monegros Centro de Desarrollo: Zaragoza.
Schipull K. 1978. Waterpockets (Opferkessel) in sandsteinen des zentralen Colorado-Plateaus. Zeitschrift für Geomorphologie 22: 426–438.
Souza-Egipsy V, Ascaso C, Wierzchos J. 2002a. Ultrastructural and biogeochemical features of biological soil crusts and their implicationsin a semi-arid habitat. Geomicrobiology Journal 19: 567–580.
Souza-Egipsy V, Ascaso C, Sancho LG. 2002b. Water distribution within terricolous lichens revealed by scanning electron microscopy andits relevance in soil crust ecology. Mycological Research 106(11): 1367–1374.
Sterflinger K. 2000. Fungi as geologic agents. Geomicrobiology Journal 17: 97–124.Viles H, Pentecost A. 1994. Problems in assessing the weathering action of lichens with an example of epiliths on sandstone. In Rock
Weathering and Landforms Evolution, Robinson DA, Williams RGB (eds). Wiley: New York; 99–116.Warsteid T, Braams J. 2000. Biodeterioration of stone: a review. International Biodeterioration & Biodegradation 46(4): 343–368.Watts S. 1998. Short-term influence of tank tracks on vegetation and microphytic crusts in shrubsteppe habitat. Environmental Management
22(4): 611–616.Wessels DCJ, Büdel B. 1995. Epilithic and cryptoendolithic cyanobacteria of Clarens sandstone cliffs in the Golden Gate Highlands National
Park, South Africa. Botanica Acta 108: 220–226.Wessels DCJ, Schoemann B. 1988. Mechanism and rate of weathering of Clarens Sanstone by an endolithic lichen. South African Journal
of Science 84: 274–277.Wierzchos J, Ascaso C. 1994. Application of backscattered electron imaging to the study of the lichen–rock interface. Journal of Microscopy
175: 54–59.Wierzchos J, García González MT, Ascaso C. 1995. Advantages of application of the backscattered electron image in the determination of
soil structure and soil constituents. International Agrophysics 9: 41–47.Williams JD, Dobrowolski JP, West NE, Gillete DA. 1994. Microphytic crust influence on wind erosion. Transations of the American
Society of Agricultural Engineers 38: 131–137.
Copyright © 2022 FDOKUMEN




















