
Effects of Right Ventricular Ejection Fraction on Outcomes in Chronic Systolic Heart Failure

Role of bacteriophage SPP1 tail spike protein gp21 on hostcell receptor binding and trigger of phage DNA ejectionmmi_7931 1..15
Inês Vinga,1,2§ Catarina Baptista,1,2§ Isabelle Auzat,3
Isabelle Petipas,3 Rudi Lurz,4 Paulo Tavares,3
Mário A. Santos5 and Carlos São-José1,2*1Centro de Patogénese Molecular, Faculdade deFarmácia da Universidade de Lisboa, Av. Prof. GamaPinto, 1649-003 Lisboa, Portugal.2Instituto de Medicina Molecular, Av. Prof. Egas Moniz,Ed. Egas Moniz, 1649-028 Lisboa, Portugal.3Laboratoire de Virologie Moléculaire et Structurale,Centre de Recherche de Gif, CNRS UPR 3296 and IFR115, 91198 Gif-sur-Yvette, France.4Max-Planck Institut für Molekulare Genetik, Ihnestraße63–73, D-14195 Berlin, Germany.5Center for Biodiversity, Functional and IntegrativeGenomics (BioFig) and Departamento de BiologiaVegetal, Faculdade de Ciências de Lisboa, Ed. ICAT,1749-016 Lisboa, Portugal.
Summary
Bacteriophages recognize and bind specific recep-tors to infect suitable hosts. Bacteriophage SPP1targets at least two receptors of the Bacillus subtiliscell envelope, the glucosylated wall teichoic acidsand the membrane protein YueB. Here, we identify akey virion protein for YueB binding and for the triggerof DNA ejection. Extracts from B. subtilis-infectedcells applied to a YueB affinity matrix led to prefer-ential capturing of gp21 from SPP1. To assess thesignificance of this interaction, we isolated mutantphages specifically affected in YueB binding. Themutants exhibited a very low inactivation rate and astrong defect to eject DNA when challenged withYueB. The phenotype correlated with presence of asingle amino acid substitution in the gp21 carboxylterminus, defining a region involved in YueB bind-ing. Immunoelectron microscopy located the gp21N-terminus in the SPP1 cap and probably in the adja-cent tail spike region whereas the gp21 C-terminuswas mapped further down in the spike structure.Antibodies against this part of gp21 interfered with
the interaction of YueB with SPP1 and triggered DNAejection. The gp21 C-terminal region thus plays acentral role in two early key events that commit thevirus to deliver its genome into host cells.
Introduction
The virus particle of the vast majority of studied bacterioph-ages is composed of an icosahedral capsid attached to atail structure, defining the order Caudovirales. The capsid,or phage head, shields a double-stranded DNA moleculewhile the tail is responsible for specific adsorption/bindingto host surface receptors and for efficient delivery of thephage genome across the bacterial cell envelope. Theseare the key events that initiate viral infection of the host cell.There are multiple solutions for phages to adsorb to theirhosts but at least one structural phage protein, known asreceptor-binding protein (RBP) has to be in contact with, atleast, one bacterial surface receptor (Vinga et al., 2006).Early studies have readily highlighted that phage adsorp-tion could involve a reversible and an irreversible interac-tion with bacterial receptors, the later committing phages toinfection (Adams, 1959).
For some phages infecting Gram-negative bacteria,such as l, T4, T5, T7 and P22, the nature and function oftail RBPs has been fairly characterized at the molecularlevel (for a review see Vinga et al., 2006). Knowledge onRBPs of phages targeting Gram-positive bacteria is morelimited. Exceptions are the recent structural and functionalstudies on RBPs of lactococcal phages (Ricagno et al.,2006; Bebeacua et al., 2010; Sciara et al., 2010; andreferences therein) and the well-characterized gp12 tail‘appendages’ of Bacillus subtilis phage j29 (Xiang et al.,2009 and references therein). A few genetic and biochemi-cal studies have identified putative RBP genes in strepto-coccal, lactobacilli and staphylococcal phages (Duplessisand Moineau, 2001; Ravin et al., 2002; Kaneko et al.,2009). RBPs are typically components of long and shortfibres attached to a contractile tail (myoviruses), of spikesor fibres attached to a short non-contractile tail (podovi-ruses), or of baseplates, fibres, spikes or single straightfibres attached to a long non-contractile tail (siphoviruses).However, details of the RBP–receptor interaction wereonly documented for a few phages like l, T5, P22 andBPP-1 (Wang et al., 2000; Böhm et al., 2001; Vinga et al.,
Accepted 18 November, 2011. *For correspondence. [email protected]; Tel. (+351) 217946420; Fax (+351) 217986055.§These authors contributed equally to the work.
Molecular Microbiology (2011) � doi:10.1111/j.1365-2958.2011.07931.x
© 2011 Blackwell Publishing Ltd

2006; Andres et al., 2010; Dai et al., 2010 and referencestherein).
Bacillus subtilis phage SPP1 is the best-characterizedsiphovirus infecting Gram-positive bacteria (Alonso et al.,2006). The SPP1 tail has been characterized by electronmicroscopy and X-ray crystallography allowing theassignment of most of its components in the tail structure(Plisson et al., 2007; Veesler et al., 2010a, Goulet et al.,2011; Fig. 1). The tail helical tube is composed of themajor tail proteins gp17.1 and gp17.1*. These proteinsshare a common amino-terminus, but the latter has anextension of ~10 kDa resulting from a programmed trans-lational frameshift (Auzat et al., 2008). The tape measureprotein gp18 occupies the tail interior. The tail tube isclosed by a cap to which the tail spike is attached. The capmain component is a hexameric ring of gp19.1, a proteinrecently coined as the distal tail protein Dit of SPP1(Veesler et al., 2010a; Goulet et al., 2011). The majorcomponent of the spike is predicted to be a gp21 trimer(Plisson et al., 2007; Goulet et al., 2011). The protein(s)composing the distal end of the tail spike rod (Fig. 1) arepresently unknown although it was hypothesized thatproducts of the five genes that follow gene 21 in thegenome (22, 23, 23.1, 24 and 24.1) could be part of it orparticipate on its assembly (Plisson et al., 2007; Veesleret al., 2010b,c)
We have recently shown that phage SPP1 interactsspecifically with glucosylated teichoic acids of theB. subtilis cell wall for the reversible step of adsorption(Baptista et al., 2008). This interaction is required for therapid and efficient recognition of a second bacterial recep-tor, the membrane protein YueB. YueB mediates SPP1irreversible adsorption and triggers phage DNA ejection(São-José et al., 2004; 2006). Both the interaction ofSPP1 with its receptors and DNA transfer to the B. subtiliscytoplasm occur preferentially at the host cell poles duringinfection (Jakutytè et al., 2011). The purified extracellular
region of YueB interacts with the SPP1 tail extremity distalfrom the capsid. It binds most probably to the tail capand/or to the tail spike that are main components of thetail adsorption apparatus (Fig. 1; São-José et al., 2006;Plisson et al., 2007). Here we have identified gp21 as acritical component of the SPP1 particle for receptorbinding and for the trigger of DNA ejection. These func-tions were assigned to a region of the protein whosetopology in the phage tail spike is also established.
Results
Capture of SPP1 tail proteins by a YueB affinity matrixand their identification
We used an affinity matrix containing the SPP1 receptorectodomain (YueB780) to capture SPP1 tail RBPs fromextracts of infected cells. Extracts were prepared fromB. subtilis infected with two SPP1 mutants blocked eitherin capsid or in tail tube assembly (SPP1sus31 andSPP1sus45 respectively). Non-permissive infections withSPP1sus31 that is defective in synthesis of the majorcapsid protein gp13 lead to the accumulation of assembledtails in the host cytoplasm (Becker et al., 1997). Infectionwith SPP1sus45, which is impaired in the major tail pro-teins gp17.1 and gp17.1* (Auzat et al., 2008), leads to theaccumulation of DNA-filled capsids and possibly of adsorp-tion apparatus complexes if tail assembly initiates at thisstructure as found for phage lambda (Katsura and Kühl,1975).
The purified ectodomain YueB780 with a hexahistidine atits carboxyl terminus was immobilized in a Ni-NTA agarosebeads column. Extracts of non-infected bacteria (control)or infected with the SPP1 sus mutants under non-permissive conditions were first depleted with free Ni-NTAbeads and then loaded into the YueB780-bound matrix.After extensive washing, YueB780 and its associated pro-
Fig. 1. Schematic representation of relevant SPP1 tail genes and location of their products in the tail structure. Gene organization on theSPP1 genome (Alonso et al., 1997) is shown in the lower part of the scheme. Genes and respective proteins whose location in the phagestructure is known or predicted have the same filling pattern (upper scheme). The black gradient denotes the unassigned carboxyl terminus ofgp21. Note the localization of the gp21 amino-terminus in the tail cap–spike interface observed in this work and which agrees with recentstructural data (Goulet et al., 2011). Genes 17.5 and 17.5* code for nonstructural proteins assisting tail assembly (Plisson et al., 2007). Thefunction of the proteins encoded by the rightmost array of 5 genes (22 to 24.1) remains unclear although it was hypothesized that at leastsome might be components of the unassigned tail spike distal end (in white in the upper scheme).
2 I. Vinga et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology
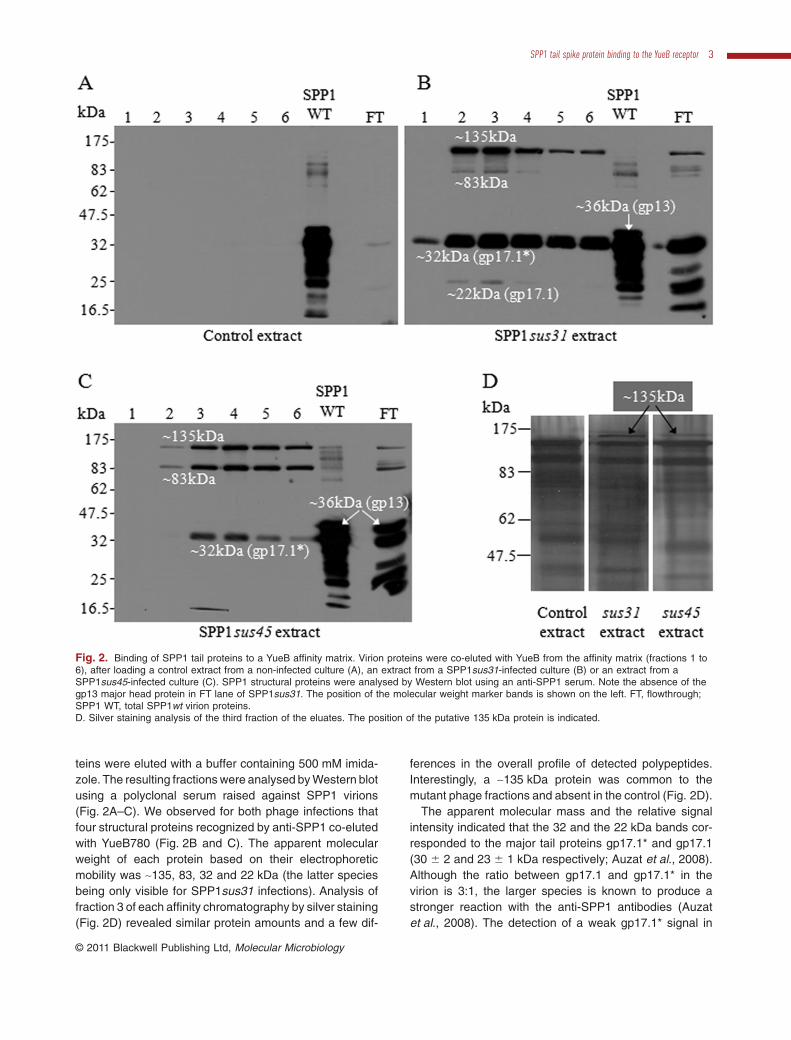
teins were eluted with a buffer containing 500 mM imida-zole. The resulting fractions were analysed by Western blotusing a polyclonal serum raised against SPP1 virions(Fig. 2A–C). We observed for both phage infections thatfour structural proteins recognized by anti-SPP1 co-elutedwith YueB780 (Fig. 2B and C). The apparent molecularweight of each protein based on their electrophoreticmobility was ~135, 83, 32 and 22 kDa (the latter speciesbeing only visible for SPP1sus31 infections). Analysis offraction 3 of each affinity chromatography by silver staining(Fig. 2D) revealed similar protein amounts and a few dif-
ferences in the overall profile of detected polypeptides.Interestingly, a ~135 kDa protein was common to themutant phage fractions and absent in the control (Fig. 2D).
The apparent molecular mass and the relative signalintensity indicated that the 32 and the 22 kDa bands cor-responded to the major tail proteins gp17.1* and gp17.1(30 � 2 and 23 � 1 kDa respectively; Auzat et al., 2008).Although the ratio between gp17.1 and gp17.1* in thevirion is 3:1, the larger species is known to produce astronger reaction with the anti-SPP1 antibodies (Auzatet al., 2008). The detection of a weak gp17.1* signal in
Fig. 2. Binding of SPP1 tail proteins to a YueB affinity matrix. Virion proteins were co-eluted with YueB from the affinity matrix (fractions 1 to6), after loading a control extract from a non-infected culture (A), an extract from a SPP1sus31-infected culture (B) or an extract from aSPP1sus45-infected culture (C). SPP1 structural proteins were analysed by Western blot using an anti-SPP1 serum. Note the absence of thegp13 major head protein in FT lane of SPP1sus31. The position of the molecular weight marker bands is shown on the left. FT, flowthrough;SPP1 WT, total SPP1wt virion proteins.D. Silver staining analysis of the third fraction of the eluates. The position of the putative 135 kDa protein is indicated.
SPP1 tail spike protein binding to the YueB receptor 3
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

SPP1sus45 fractions can be explained by a small per-centage of revertants and/or by the presence of inputvirions, which are genotypically defective for 17.1/17.1*but structurally wild type, in the extract produced fromSPP1sus45 infection.
The estimated mass of the high molecular weight proteindetected in the eluted fractions of both mutant phages(~135 kDa; Fig. 2B and C) was compatible with that of thetail structural components gp18 and gp21. They havetheoretical masses of ~110 kDa and ~123.5 kDa respec-tively. The mass of gp21 takes into account the new gp21amino acid sequence numbering used from hereafter thatstarts at the leucine residue that precedes the amino-terminus sequence SKNIXI determined experimentally forthe gp21 present in SPP1 particles (data not shown). Thisleucine codon occupies the positions 16134 to 16136 ofthe SPP1 nucleotide sequence with access code X97918.The 83 kDa polypeptide detected in Fig. 2B and C is ofunknown origin. Its relative ratio to other phage proteinsis significantly higher in extracts of cells infected withSPP1sus45 that does not assemble tail tubes (Fig. 2C).This observation suggests that it is a truncated product ofa structural protein recognized by anti-SPP1 antibodiesthat accumulates when tail assembly is blocked.
The relative ratio between SPP1 proteins in each indi-vidual fraction captured by affinity to YueB shows a majorenrichment of the 135 kDa species when compared withthe ratio found in viral particles (Fig. 2B and C). Thisenrichment is observed even in conditions that completetail assembly occurs (SPP1sus31 infection; Fig. 2B)showing that most of the 135 kDa protein retained inthe affinity column is not associated to intact tails. The135 kDa species is thus stable in infected cells, accumu-lating in excess amounts for the tail assembly reaction.
The clear identification of a 135 kDa protein in silverstained gels that is absent in non-infected cells (Fig. 2D)prompted us to carry out a preparative experiment to obtainsufficient material for its identification by mass spectrom-etry. The corresponding band was recovered from the gel,digested with trypsin and the tryptic fragments submitted toMALDI-TOF mass spectrometry. Data analyses with theMascot tool (Perkins et al., 1999) against the ‘All Taxa’database identified gp21 of SPP1 as the first prokaryoticprotein hit. Gp21 was also the first hit when the search wasrestricted to the ‘Viruses’ database with a score slightlybelow the cut-off for a significant match [score of 62; scoresgreater than 72 were considered significant (P < 0.05)] anda 28 % coverage of the gp21 sequence. This poor signal isattributed to the low amount of material present in the135 kDa band. However, the observations that gp21 is theonly SPP1 protein identified in the mass spectrometryexperiment and the compatibility of its mass with theelectrophoretic mobility of the 135 kDa protein clearly hintthat gp21 is the species enriched in the YueB780 affinity
column. Presence of YueB in the 135 kDa band was alsodetected when the search was made against the ‘Bacillussubtilis’ database. This was an expected result as YueB780was co-eluted from the column matrix together with theextract retained proteins and because the 87.3 kDa recep-tor subunit has an abnormal electroporetic mobility inSDS-PAGE gels behaving like a globular ~120 kDa protein(São-José et al., 2006).
Isolation of SPP1 mutants specifically affected inirreversible adsorption
The results presented above suggested that the putativeSPP1 tail spike protein gp21, or a structure containing it,binds to the receptor YueB. To further investigate thisinteraction we developed a strategy to isolate SPP1mutants specifically affected in irreversible binding toYueB, followed by identification of the mutated genesresponsible for the deficient adsorption phenotype.
We were able to separate the two steps of SPP1 adsorp-tion by constructing B. subtilis mutants that express onlyone of the two receptors involved in the process (Baptistaet al., 2008). Strain CSJ3.gtaC bears non-glucosylatedWTA (impaired reversible adsorption) and overproducesYueB upon IPTG induction. The increased surface concen-tration of YueB in this strain (Jakutytè et al., 2011) wasshown to bypass the requirement for reversible interac-tion, allowing SPP1 irreversible adsorption rates inliquid medium similar to those observed for the wild-typeB. subtilis (Baptista et al., 2008).
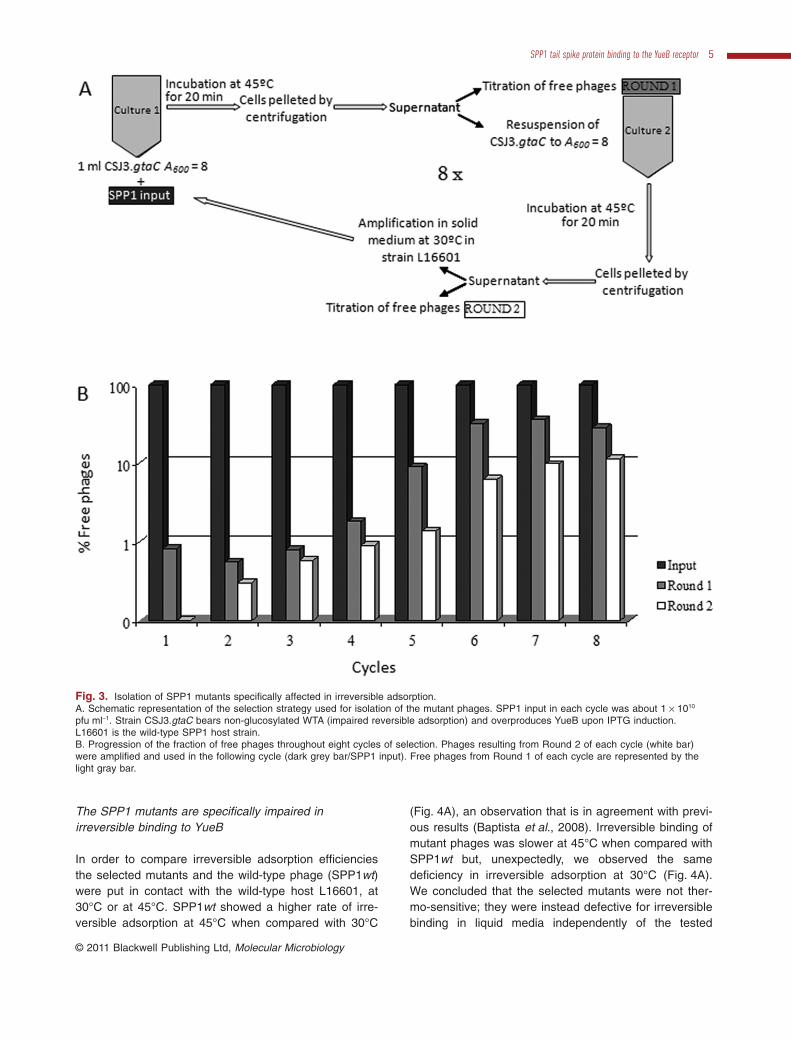
Because irreversible binding is essential for SPP1 infec-tion (São-José et al., 2004) we delineated a strategy toisolate thermo-sensitive mutants, i.e. phages deficient forirreversible adsorption at 45°C in liquid medium, but whichwere able to form plaques in solid medium at 30°C (seeExperimental procedures and Fig. 3A). We performedseveral cycles where induced cultures of strain CSJ3.gtaCwere incubated at 45°C with SPP1 during a period of timethat would bind more than 99% of the wild-type phageinput. Each cycle corresponded to two rounds of contactbetween SPP1 and strain CSJ3.gtaC in a way that thesecond round was performed with virions that failed toadsorb in the first round.After each cycle, free phages werepropagated in solid media at 30°C in the wild-type B. sub-tilis L16601, recovered and challenged again with strainCSJ3.gtaC at 45°C in the following cycle. The percentageof free phages increased throughout the cycles and afterthe 8th cycle we obtained an SPP1 lysate enriched inmutant phages with impaired binding to YueB (Fig. 3B).Several isolated plaques from this lysate were picked,propagated and confirmed for the deficient adsorptionphenotype. Three mutant isolates with reduced irreversibleadsorption were selected for further studies (SPP1mut6,SPP1mut7, and SPP1mut9).
4 I. Vinga et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology
The SPP1 mutants are specifically impaired inirreversible binding to YueB
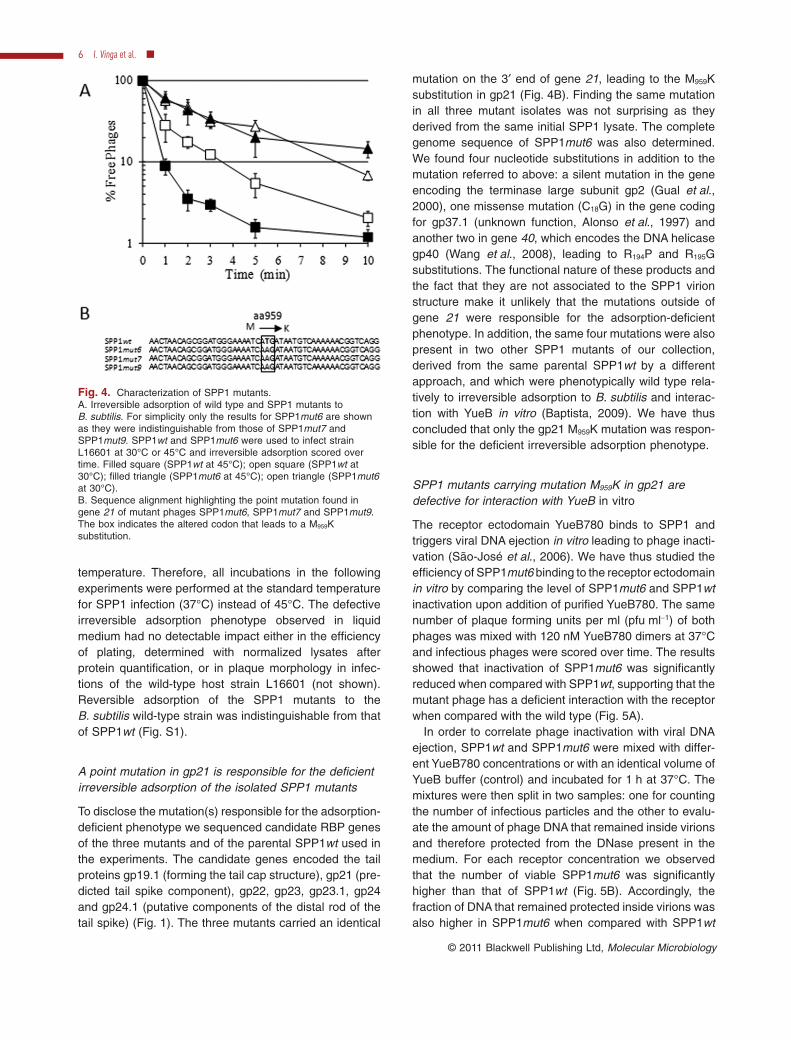
In order to compare irreversible adsorption efficienciesthe selected mutants and the wild-type phage (SPP1wt)were put in contact with the wild-type host L16601, at30°C or at 45°C. SPP1wt showed a higher rate of irre-versible adsorption at 45°C when compared with 30°C
(Fig. 4A), an observation that is in agreement with previ-ous results (Baptista et al., 2008). Irreversible binding ofmutant phages was slower at 45°C when compared withSPP1wt but, unexpectedly, we observed the samedeficiency in irreversible adsorption at 30°C (Fig. 4A).We concluded that the selected mutants were not ther-mo-sensitive; they were instead defective for irreversiblebinding in liquid media independently of the tested
Fig. 3. Isolation of SPP1 mutants specifically affected in irreversible adsorption.A. Schematic representation of the selection strategy used for isolation of the mutant phages. SPP1 input in each cycle was about 1 ¥ 1010
pfu ml-1. Strain CSJ3.gtaC bears non-glucosylated WTA (impaired reversible adsorption) and overproduces YueB upon IPTG induction.L16601 is the wild-type SPP1 host strain.B. Progression of the fraction of free phages throughout eight cycles of selection. Phages resulting from Round 2 of each cycle (white bar)were amplified and used in the following cycle (dark grey bar/SPP1 input). Free phages from Round 1 of each cycle are represented by thelight gray bar.
SPP1 tail spike protein binding to the YueB receptor 5
© 2011 Blackwell Publishing Ltd, Molecular Microbiology
temperature. Therefore, all incubations in the followingexperiments were performed at the standard temperaturefor SPP1 infection (37°C) instead of 45°C. The defectiveirreversible adsorption phenotype observed in liquidmedium had no detectable impact either in the efficiencyof plating, determined with normalized lysates afterprotein quantification, or in plaque morphology in infec-tions of the wild-type host strain L16601 (not shown).Reversible adsorption of the SPP1 mutants to theB. subtilis wild-type strain was indistinguishable from thatof SPP1wt (Fig. S1).
A point mutation in gp21 is responsible for the deficientirreversible adsorption of the isolated SPP1 mutants
To disclose the mutation(s) responsible for the adsorption-deficient phenotype we sequenced candidate RBP genesof the three mutants and of the parental SPP1wt used inthe experiments. The candidate genes encoded the tailproteins gp19.1 (forming the tail cap structure), gp21 (pre-dicted tail spike component), gp22, gp23, gp23.1, gp24and gp24.1 (putative components of the distal rod of thetail spike) (Fig. 1). The three mutants carried an identical
mutation on the 3′ end of gene 21, leading to the M959Ksubstitution in gp21 (Fig. 4B). Finding the same mutationin all three mutant isolates was not surprising as theyderived from the same initial SPP1 lysate. The completegenome sequence of SPP1mut6 was also determined.We found four nucleotide substitutions in addition to themutation referred to above: a silent mutation in the geneencoding the terminase large subunit gp2 (Gual et al.,2000), one missense mutation (C18G) in the gene codingfor gp37.1 (unknown function, Alonso et al., 1997) andanother two in gene 40, which encodes the DNA helicasegp40 (Wang et al., 2008), leading to R194P and R195Gsubstitutions. The functional nature of these products andthe fact that they are not associated to the SPP1 virionstructure make it unlikely that the mutations outside ofgene 21 were responsible for the adsorption-deficientphenotype. In addition, the same four mutations were alsopresent in two other SPP1 mutants of our collection,derived from the same parental SPP1wt by a differentapproach, and which were phenotypically wild type rela-tively to irreversible adsorption to B. subtilis and interac-tion with YueB in vitro (Baptista, 2009). We have thusconcluded that only the gp21 M959K mutation was respon-sible for the deficient irreversible adsorption phenotype.
SPP1 mutants carrying mutation M959K in gp21 aredefective for interaction with YueB in vitro
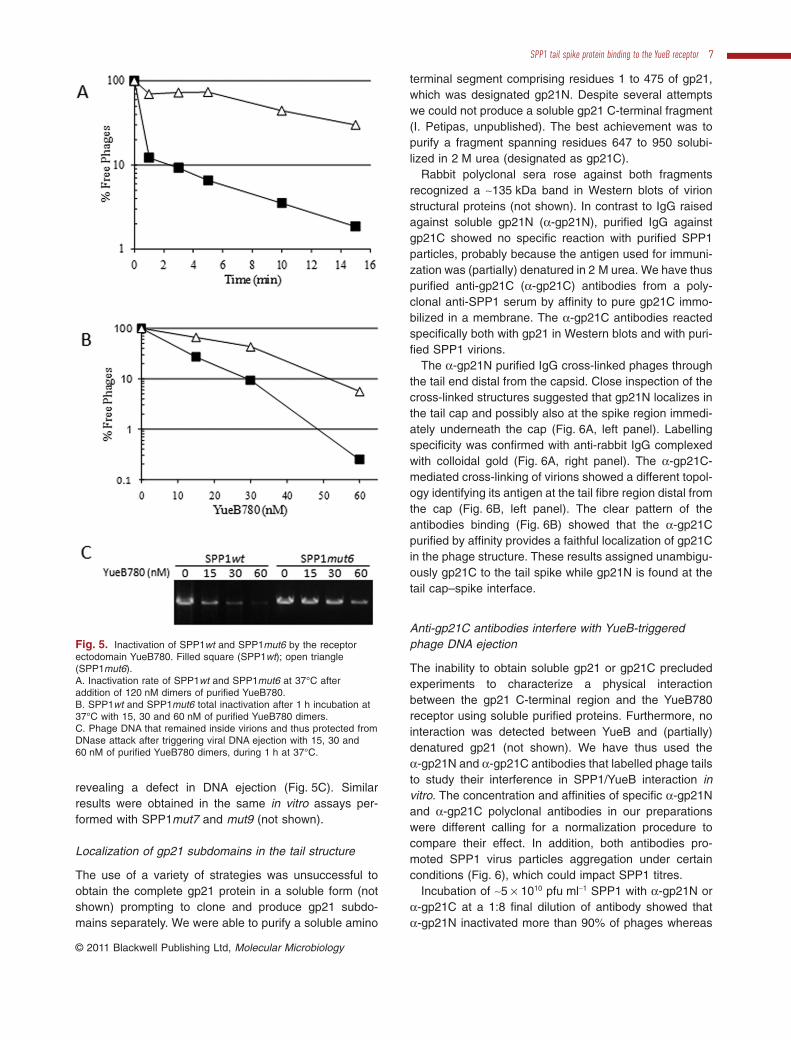
The receptor ectodomain YueB780 binds to SPP1 andtriggers viral DNA ejection in vitro leading to phage inacti-vation (São-José et al., 2006). We have thus studied theefficiency of SPP1mut6 binding to the receptor ectodomainin vitro by comparing the level of SPP1mut6 and SPP1wtinactivation upon addition of purified YueB780. The samenumber of plaque forming units per ml (pfu ml-1) of bothphages was mixed with 120 nM YueB780 dimers at 37°Cand infectious phages were scored over time. The resultsshowed that inactivation of SPP1mut6 was significantlyreduced when compared with SPP1wt, supporting that themutant phage has a deficient interaction with the receptorwhen compared with the wild type (Fig. 5A).
In order to correlate phage inactivation with viral DNAejection, SPP1wt and SPP1mut6 were mixed with differ-ent YueB780 concentrations or with an identical volume ofYueB buffer (control) and incubated for 1 h at 37°C. Themixtures were then split in two samples: one for countingthe number of infectious particles and the other to evalu-ate the amount of phage DNA that remained inside virionsand therefore protected from the DNase present in themedium. For each receptor concentration we observedthat the number of viable SPP1mut6 was significantlyhigher than that of SPP1wt (Fig. 5B). Accordingly, thefraction of DNA that remained protected inside virions wasalso higher in SPP1mut6 when compared with SPP1wt
Fig. 4. Characterization of SPP1 mutants.A. Irreversible adsorption of wild type and SPP1 mutants toB. subtilis. For simplicity only the results for SPP1mut6 are shownas they were indistinguishable from those of SPP1mut7 andSPP1mut9. SPP1wt and SPP1mut6 were used to infect strainL16601 at 30°C or 45°C and irreversible adsorption scored overtime. Filled square (SPP1wt at 45°C); open square (SPP1wt at30°C); filled triangle (SPP1mut6 at 45°C); open triangle (SPP1mut6at 30°C).B. Sequence alignment highlighting the point mutation found ingene 21 of mutant phages SPP1mut6, SPP1mut7 and SPP1mut9.The box indicates the altered codon that leads to a M959Ksubstitution.
6 I. Vinga et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology
revealing a defect in DNA ejection (Fig. 5C). Similarresults were obtained in the same in vitro assays per-formed with SPP1mut7 and mut9 (not shown).
Localization of gp21 subdomains in the tail structure
The use of a variety of strategies was unsuccessful toobtain the complete gp21 protein in a soluble form (notshown) prompting to clone and produce gp21 subdo-mains separately. We were able to purify a soluble amino
terminal segment comprising residues 1 to 475 of gp21,which was designated gp21N. Despite several attemptswe could not produce a soluble gp21 C-terminal fragment(I. Petipas, unpublished). The best achievement was topurify a fragment spanning residues 647 to 950 solubi-lized in 2 M urea (designated as gp21C).
Rabbit polyclonal sera rose against both fragmentsrecognized a ~135 kDa band in Western blots of virionstructural proteins (not shown). In contrast to IgG raisedagainst soluble gp21N (a-gp21N), purified IgG againstgp21C showed no specific reaction with purified SPP1particles, probably because the antigen used for immuni-zation was (partially) denatured in 2 M urea. We have thuspurified anti-gp21C (a-gp21C) antibodies from a poly-clonal anti-SPP1 serum by affinity to pure gp21C immo-bilized in a membrane. The a-gp21C antibodies reactedspecifically both with gp21 in Western blots and with puri-fied SPP1 virions.
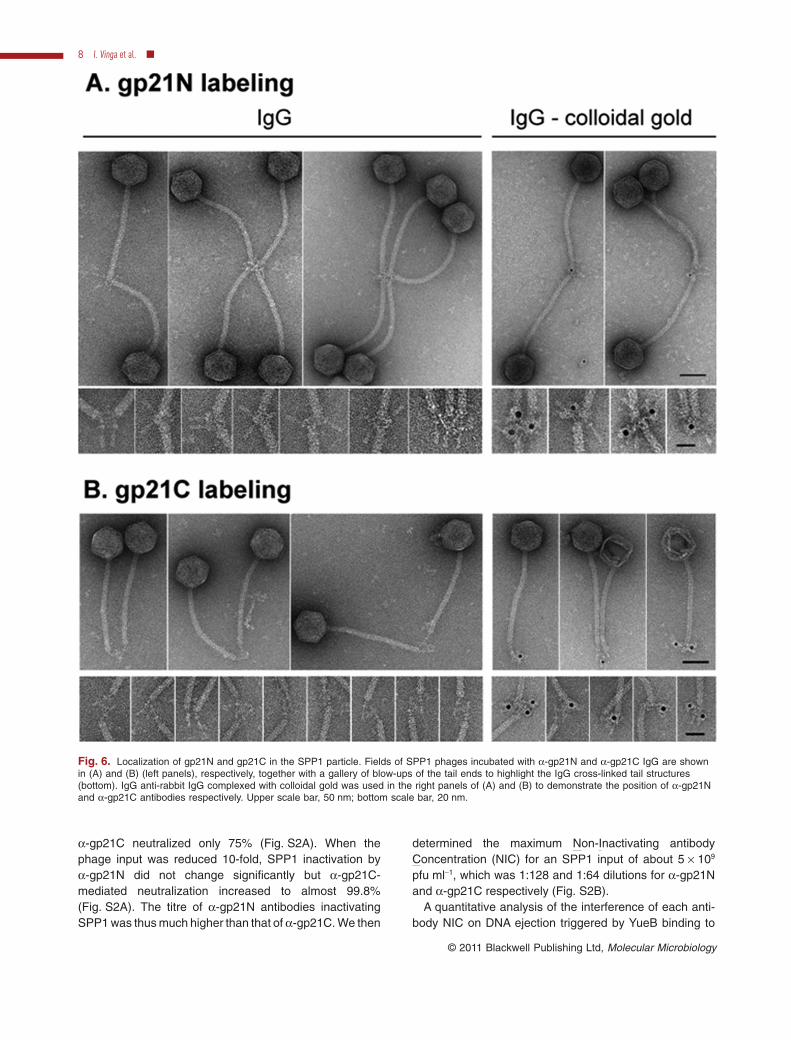
The a-gp21N purified IgG cross-linked phages throughthe tail end distal from the capsid. Close inspection of thecross-linked structures suggested that gp21N localizes inthe tail cap and possibly also at the spike region immedi-ately underneath the cap (Fig. 6A, left panel). Labellingspecificity was confirmed with anti-rabbit IgG complexedwith colloidal gold (Fig. 6A, right panel). The a-gp21C-mediated cross-linking of virions showed a different topol-ogy identifying its antigen at the tail fibre region distal fromthe cap (Fig. 6B, left panel). The clear pattern of theantibodies binding (Fig. 6B) showed that the a-gp21Cpurified by affinity provides a faithful localization of gp21Cin the phage structure. These results assigned unambigu-ously gp21C to the tail spike while gp21N is found at thetail cap–spike interface.
Anti-gp21C antibodies interfere with YueB-triggeredphage DNA ejection
The inability to obtain soluble gp21 or gp21C precludedexperiments to characterize a physical interactionbetween the gp21 C-terminal region and the YueB780receptor using soluble purified proteins. Furthermore, nointeraction was detected between YueB and (partially)denatured gp21 (not shown). We have thus used thea-gp21N and a-gp21C antibodies that labelled phage tailsto study their interference in SPP1/YueB interaction invitro. The concentration and affinities of specific a-gp21Nand a-gp21C polyclonal antibodies in our preparationswere different calling for a normalization procedure tocompare their effect. In addition, both antibodies pro-moted SPP1 virus particles aggregation under certainconditions (Fig. 6), which could impact SPP1 titres.
Incubation of ~5 ¥ 1010 pfu ml-1 SPP1 with a-gp21N ora-gp21C at a 1:8 final dilution of antibody showed thata-gp21N inactivated more than 90% of phages whereas
Fig. 5. Inactivation of SPP1wt and SPP1mut6 by the receptorectodomain YueB780. Filled square (SPP1wt); open triangle(SPP1mut6).A. Inactivation rate of SPP1wt and SPP1mut6 at 37°C afteraddition of 120 nM dimers of purified YueB780.B. SPP1wt and SPP1mut6 total inactivation after 1 h incubation at37°C with 15, 30 and 60 nM of purified YueB780 dimers.C. Phage DNA that remained inside virions and thus protected fromDNase attack after triggering viral DNA ejection with 15, 30 and60 nM of purified YueB780 dimers, during 1 h at 37°C.
SPP1 tail spike protein binding to the YueB receptor 7
© 2011 Blackwell Publishing Ltd, Molecular Microbiology
a-gp21C neutralized only 75% (Fig. S2A). When thephage input was reduced 10-fold, SPP1 inactivation bya-gp21N did not change significantly but a-gp21C-mediated neutralization increased to almost 99.8%(Fig. S2A). The titre of a-gp21N antibodies inactivatingSPP1 was thus much higher than that of a-gp21C. We then
determined the maximum Non-Inactivating antibodyConcentration (NIC) for an SPP1 input of about 5 ¥ 109
pfu ml-1, which was 1:128 and 1:64 dilutions for a-gp21Nand a-gp21C respectively (Fig. S2B).
A quantitative analysis of the interference of each anti-body NIC on DNA ejection triggered by YueB binding to
Fig. 6. Localization of gp21N and gp21C in the SPP1 particle. Fields of SPP1 phages incubated with a-gp21N and a-gp21C IgG are shownin (A) and (B) (left panels), respectively, together with a gallery of blow-ups of the tail ends to highlight the IgG cross-linked tail structures(bottom). IgG anti-rabbit IgG complexed with colloidal gold was used in the right panels of (A) and (B) to demonstrate the position of a-gp21Nand a-gp21C antibodies respectively. Upper scale bar, 50 nm; bottom scale bar, 20 nm.
8 I. Vinga et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology
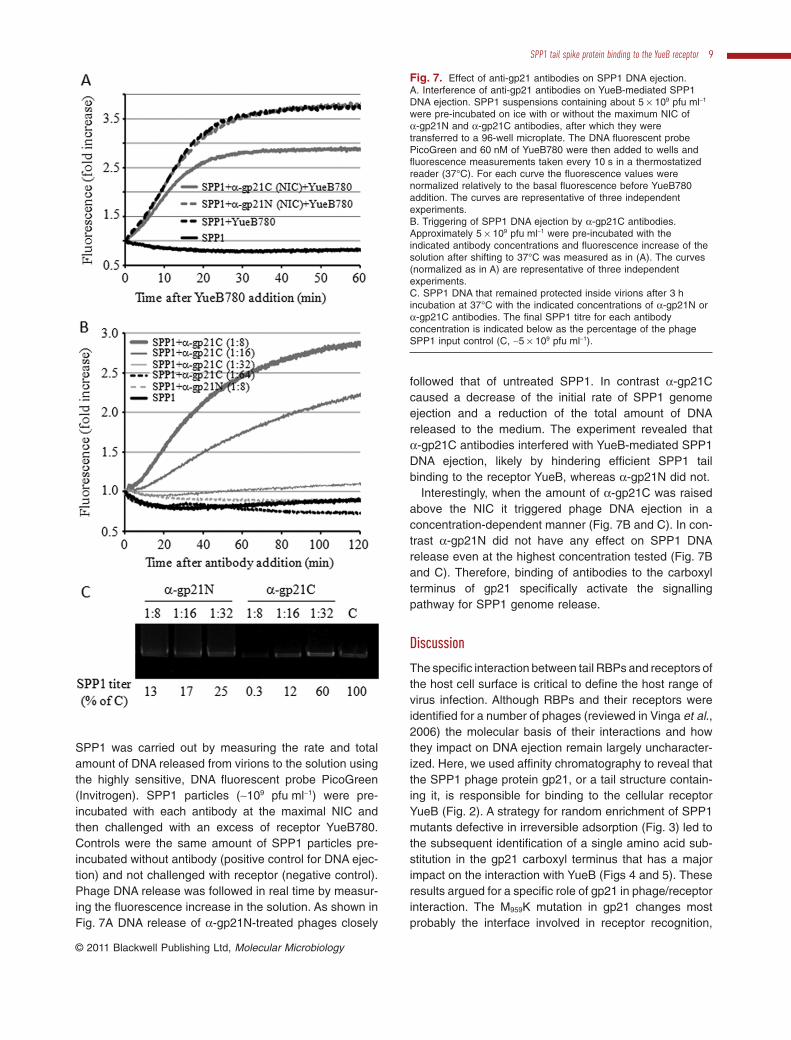
SPP1 was carried out by measuring the rate and totalamount of DNA released from virions to the solution usingthe highly sensitive, DNA fluorescent probe PicoGreen(Invitrogen). SPP1 particles (~109 pfu ml-1) were pre-incubated with each antibody at the maximal NIC andthen challenged with an excess of receptor YueB780.Controls were the same amount of SPP1 particles pre-incubated without antibody (positive control for DNA ejec-tion) and not challenged with receptor (negative control).Phage DNA release was followed in real time by measur-ing the fluorescence increase in the solution. As shown inFig. 7A DNA release of a-gp21N-treated phages closely
followed that of untreated SPP1. In contrast a-gp21Ccaused a decrease of the initial rate of SPP1 genomeejection and a reduction of the total amount of DNAreleased to the medium. The experiment revealed thata-gp21C antibodies interfered with YueB-mediated SPP1DNA ejection, likely by hindering efficient SPP1 tailbinding to the receptor YueB, whereas a-gp21N did not.
Interestingly, when the amount of a-gp21C was raisedabove the NIC it triggered phage DNA ejection in aconcentration-dependent manner (Fig. 7B and C). In con-trast a-gp21N did not have any effect on SPP1 DNArelease even at the highest concentration tested (Fig. 7Band C). Therefore, binding of antibodies to the carboxylterminus of gp21 specifically activate the signallingpathway for SPP1 genome release.
Discussion
The specific interaction between tail RBPs and receptors ofthe host cell surface is critical to define the host range ofvirus infection. Although RBPs and their receptors wereidentified for a number of phages (reviewed in Vinga et al.,2006) the molecular basis of their interactions and howthey impact on DNA ejection remain largely uncharacter-ized. Here, we used affinity chromatography to reveal thatthe SPP1 phage protein gp21, or a tail structure contain-ing it, is responsible for binding to the cellular receptorYueB (Fig. 2). A strategy for random enrichment of SPP1mutants defective in irreversible adsorption (Fig. 3) led tothe subsequent identification of a single amino acid sub-stitution in the gp21 carboxyl terminus that has a majorimpact on the interaction with YueB (Figs 4 and 5). Theseresults argued for a specific role of gp21 in phage/receptorinteraction. The M959K mutation in gp21 changes mostprobably the interface involved in receptor recognition,
Fig. 7. Effect of anti-gp21 antibodies on SPP1 DNA ejection.A. Interference of anti-gp21 antibodies on YueB-mediated SPP1DNA ejection. SPP1 suspensions containing about 5 ¥ 109 pfu ml-1
were pre-incubated on ice with or without the maximum NIC ofa-gp21N and a-gp21C antibodies, after which they weretransferred to a 96-well microplate. The DNA fluorescent probePicoGreen and 60 nM of YueB780 were then added to wells andfluorescence measurements taken every 10 s in a thermostatizedreader (37°C). For each curve the fluorescence values werenormalized relatively to the basal fluorescence before YueB780addition. The curves are representative of three independentexperiments.B. Triggering of SPP1 DNA ejection by a-gp21C antibodies.Approximately 5 ¥ 109 pfu ml-1 were pre-incubated with theindicated antibody concentrations and fluorescence increase of thesolution after shifting to 37°C was measured as in (A). The curves(normalized as in A) are representative of three independentexperiments.C. SPP1 DNA that remained protected inside virions after 3 hincubation at 37°C with the indicated concentrations of a-gp21N ora-gp21C antibodies. The final SPP1 titre for each antibodyconcentration is indicated below as the percentage of the phageSPP1 input control (C, ~5 ¥ 109 pfu ml-1).
SPP1 tail spike protein binding to the YueB receptor 9
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

decreasing the SPP1 tail spike affinity for YueB. The muta-tion could also affect the interaction of gp21 with anunknown component of the SPP1 tail spike involved inbinding to YueB. However, the strong interference ofa-gp21C in YueB780-mediated triggering of SPP1 DNAejection (Fig. 7A) demonstrates a central role of gp21C inthe cross-talk with YueB. Taken together with the observa-tion that the gp21 carboxyl terminus is not shielded by otherproteins for antibody labelling, these results support adirect interaction between gp21 and YueB. The finding thata-gp21C specifically triggers SPP1 DNA ejection (Fig. 7Band C), mimicking the effect of YueB binding to the tailspike, unravels a new function for the gp21 carboxyl termi-nus as an initiator of the cascade of structural rearrange-ments leading to viral DNA ejection.
No obvious similarity between the gp21 C-terminalregion and other proteins was detected (Veesler et al.,2010a). In contrast, its amino-terminus has homology tonumerous tail proteins from phages infecting Gram-positive bacteria. Their coding genes follow in most phagegenomes the ones encoding gp19.1-like polypeptides,which compose the tail distal end (Dit proteins) and whichform hexameric rings in the cap structure (Sciara et al.,2010; Veesler et al., 2010a; Goulet et al., 2011). Theobservation that such sequence relationship is found alsoin phages whose RBPs are attached to a baseplate sug-gests that they all share a common structural organizationin which the amino-terminus of gp21-like proteins binds tothe tail cap. Recently, the cryo-EM structure of a complexformed by the cap protein gp19.1 and the N-terminus ofgp21 (gp211–552) was determined (Goulet et al., 2011). Itsdocking into the EM maps of the SPP1 tail showed thatthe SPP1 cap is formed by a hexamer of gp19.1 and atrimer of the gp21 amino-terminus. This is consistent withthe observed topology of the gp21 amino-terminus at theregion covering the transition from the tail cap to thebeginning of the tail spike, as shown by immunoelectronmicroscopy (Fig. 6A). We have now assigned the gp21carboxyl terminus position to the SPP1 tail spike structure(Fig. 6B).
Although the N-terminal region of gp21-like proteinsparticipates in the composition of the tail cap structure inseveral phages, positioning of the RBP function in thetail adsorption apparatus is more diverse, determiningdistinct strategies of recognition and binding to the host,which are specific to the phage system (Sciara et al.,2008). The role of the gp21 carboxyl terminus in bindingto YueB that is shown here for SPP1 was also observedin case of coliphage l whose gpJ last 249 residuesinteract with the maltoporin LamB (Wang et al., 2000).Both phages have a related organization of their hostadsorption apparatus featuring a long tail spike attachedto the tail tube cap (Fig. 1). No other obvious structuresfor binding to the host are present even though the origi-
nal l isolate featured thin tail fibres lost early in the labo-ratory history of this phage (Hendrix and Duda, 1992).The emerging picture is that the major tail spike compo-nent of both phages is a long polypeptide (gp21 has1108 residues and gpJ 1132 residues) whose carboxylterminus is responsible for receptor binding. This activityis a necessary requirement to trigger DNA ejection fromthe phage particle (Figs 5C and 7). In contrast, receptorrecognition by phage T5 is mediated by protein pb5 thatbelongs to a conical structure that caps the tail helicaltube, whereas the single straight fibre pb2 serves asconduit for T5 DNA transfer into Escherichia coli cells(Letellier et al., 2004).
The data presented here together with available knowl-edge allow proposing a comprehensive model for theevents leading to genome ejection from the SPP1 particle(Fig. 8). The probable direct interaction of YueB with thecarboxyl terminus of gp21 exposed to the exterior of thetail spike (step 1 in Fig. 8) leads to destabilization of thespike structure and initiates a signal causing a switch inthe gp21 amino-terminus structure that opens at the tailcap (step 2) (this work; Goulet et al., 2011). The signal isthen transmitted across the cap, probably through thegp19.1 hexamer, and through the tail tube structure tar-geting the head-to-tail connector (step 2) (Plisson et al.,2007). Opening of the connector (Lhuillier et al., 2009)leads to release of DNA through the tail tube that shall bepreceded by exit of the tape measure protein that occu-pied the tail lumen (steps 3 and 4) (Plisson et al., 2007).The studies reported here provide a first contribution tounderstand the critical cross-talk between the receptorand the SPP1 phage structure that initiates the wholeprocess.
Experimental procedures
Bacterial strains, bacteriophages and growth conditions
Escherichia coli and B. subtilis were grown in Luria–Bertani(LB) medium (Sambrook and Russell, 2001) unless statedotherwise. SPP1 phages were multiplied as previouslydescribed (São-José et al., 2004). B. subtilis strain L16601(wild-type strain, Margot and Karamata, 1996) was used asindicator and propagation host for SPP1wt (Riva et al., 1968)and SPP1mut phages. SPP1mut phages were propagated insolid medium at 30°C. SPP1 phages were purified by isopyc-nic centrifugation in step caesium chloride (CsCl) gradients(Sambrook and Russell, 2001). Lysates of mutant phagesSPP1sus31 (TAG stop codon replacing codon Gln31 in gene13, Becker et al., 1997; Dröge et al., 2000) and SPP1sus45(TAA stop codon replacing codon Gln133 in gene 17.1, Auzatet al., 2008) were multiplied in the suppressor B. subtilis strainHA101B (Okubo and Yanagida, 1968) as described (Drögeand Tavares, 2000). The percentage of SPP1sus revertantswas controlled by simultaneous titration in strain HA101B andin the non-permissive host L16601 (less than 1% in the finallysates). All phages were maintained and diluted in TBT buffer
10 I. Vinga et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

(100 mM Tris-Cl, pH 7.5, 100 mM NaCl, 10 mM MgCl2).B. subtilis strain CSJ3.gtaC (grown in the presence of0.5 mg ml-1 erythromycin, 5 mg ml-1 chloramphenicol and1 mM MgCl2, Baptista et al., 2008) produces non-glucosylatedWTA (impaired in SPP1 reversible adsorption) and overpro-duces YueB upon IPTG induction (1 mM). B. subtilis strainCSJ1 is a yueB mutant (São-José et al., 2004) that onlyadsorbs SPP1 reversibly (Baptista et al., 2008). The E. colistrain used for production of YueB780 (CG65, São-José et al.,2004) was grown at 30°C before protein induction and at 37°Cafter induction in LB supplemented with 100 mg ml-1 of ampi-cillin and 40 mg ml-1 of kanamycin.
Cloning of gene 21 segments
Gene 21 DNA fragments encoding gp211–475 (gp21N) andgp21647–950 (gp21C) encompass the nucleotide positions16125–17558 and 18072–18983, respectively, of the SPP1genome sequence deposited in GenBank under X97918.Here although the gp21 amino acid co-ordinates incorporatethe newly assigned N-terminus of the protein present in SPP1virions (see text). These fragments were amplified by poly-merase chain reaction from SPP1 DNA using oligonucle-otides pairs GCAGGCAGGATCCGTGAGAAGCTTGTCA/CGCACTGCAGTTATTTCCCTTCAAAAATTATG and GGCAGGATCCAGTTCATTGAAAG/GGCACTGCAGTTACCACCCGTTAGCGAAATG respectively. PCR fragments weredigested with BamHI and PstI and cloned into pRSETa (Invit-
rogen) cleaved with the same enzymes. Both resulting plas-mids (pIP5 and pIP9) code for a fusion protein containing ahexahistidine sequence followed by a 26 amino acids longlinker at the N-terminus extremity.
Purification of gp21N and gp21C for antibody production
Escherichia coli JS218 freshly transformed with plasmid pIP5was grown in LB broth containing 100 mg ml-1 ampicillin to acell density of 106 cfu ml-1 and then induced with 2 mM IPTGfor about 3 h. Pelleted bacteria were resuspended in lysisbuffer (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.1% TritonX-100, 10 mM MgSO4) and sonicated on ice with 1 min bursts(Vibra Cell). The crude extract was centrifuged at 4°C twicefor 20 min at 27 000 g. Imidazole was then added to a finalconcentration of 20 mM and the recombinant gp211–475 puri-fied by metal-chelating chromatography using HiTrap column(GE Healthcare) according to the supplier conditions (20 mMNaH2PO4 pH 7.4, NaCl 500 mM). Protein was washed withsteps of increasing imidazole concentrations, eluted at250 mM, concentrated and further purified by gel filtration ona Superdex 200 column (GE Healthcare) in 100 mM NaCl,20 mM Tris-HCl, pH 7.5.
Escherichia coli BL21(DE3) freshly transformed withplasmid pIP9 was grown in LB broth containing 100 mg ml-1
ampicillin to a cell density of 106 cfu ml-1 and then induced with2 mM IPTG for about 3 h at 25°C. Pelleted bacteria wereresuspended in lysis buffer (50 mM Tris-HCl pH 7.5, 150 mM
Fig. 8. Model for the sequential steps involved in SPP1 binding to its receptor YueB780 and subsequent viral genome ejection in vitro. Themodel integrates the data from the present work and that of São-José et al. (2006; 2007), Plisson et al. 2007, Lhuillier et al. 2009 and Gouletet al. 2011. SPP1 binds to the receptor through the C-terminal region of the gp21 trimer subunits composing the tail spike (1). The number ofYueB780 receptors per phage particle required for productive interaction is unknown (?). This interaction initiates a cascade of conformationalchanges at the gp21 carboxyl terminus causing disassembly of the tail spike organization and opening of the cap region formed by theamino-terminus of gp21 (Goulet et al., 2011) (2). The open channel extends through the gp19.1 hexamer in the tail cap to the tail inner tailtube. The conformational changes at the tail cap generate a signal transmitted through the tail tube, via structural rearrangements of the majortail tube subunits, to the capsid connector (Plisson et al., 2007). Opening of this structure (Lhuillier et al., 2009) releases the highly pressurizedDNA present inside the capsid (São-José et al., 2007) to expel the tail tape measure protein (TMP) followed by the phage DNA (3, 4). Thefate of the tape measure proteins that occupied the tail tube lumen is unknown (??).
SPP1 tail spike protein binding to the YueB receptor 11
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

NaCl, 0.1% Triton X-100, 10 mM MgSO4) with EDTA-freeproteases inhibitors (Roche) and sonicated 10 times on icewith 1 min bursts (Vibra Cell). The crude extract was centrifu-gated at 4°C for 20 min at 27 000 g. The pellet was resus-pended in buffer containing 6 M guanidinium hydrochloridewith protease inhibitors (Roche), sonicated twice for 1 min andincubated at room temperature for 2.5 h. Two centrifugationruns were performed at 4°C for 20 min at 27 000 g. Gp21647–950
was purified by metal-chelating chromatography using anHiTrap column (GE Healthcare) equilibrated with buffer A(20 mM NaH2PO4 pH 7.4, NaCl 500 mM, 8 M urea). Thecolumn was washed with increasing steps of imidazole con-centration in buffer A and eluted between 100 and 250 mMimidazole. The urea concentration was finally reduced to 2 Mby rapid dilution with nickel buffer.
Rabbit polyclonal antisera production against both purifiedproteins and IgG purification from sera were carried outaccording to standard protocols (Harlow and Lane, 1988).
Affinity purification of anti-gp21C antibodies from ananti-SPP1 polyclonal serum
Purified gp21C polypeptides in 2 M urea were run in pre-parative 16% SDS-polyacrylamide gels and membrane-immobilized by blotting transfer to Nitrocellulose PROTRANmembranes (Whatman). Anti-gp21C antibodies were purifiedby affinity from an anti-SPP1 serum as described by Orlovaet al. (2003). Their specificity to gp21 was confirmed byWestern blot and immunoEM.
Production of SPP1sus total protein extracts frominfected cells
A 200 ml culture of B. subtilis strain L16601 was supple-mented with 10 mM CaCl2 when it reached a density of ~108
cfu ml-1 (A600 = 0.8) and infected with SPP1sus31 orSPP1sus45 at an input multiplicity of 5. After an incubationperiod of 10 min at 37°C with shaking, the infection mixtureswere centrifuged at room temperature in a pre-warmed rotor.The cellular pellets were resuspended in 1/10 volume ofpre-warmed LB medium and incubated in the same conditionsuntil 1 h post infection. To complete cell lysis, 100 mg ml-1
lysozyme, 300 mM NaCl, 50 mM imidazole, 10 mg ml-1 DNaseand EDTA-free Anti-Protease Cocktail (Roche AppliedScience) were added to the lysates, which were then incu-bated on ice for 10 min. After 3 ¥ 1 min sonication the crudeextracts were cleared by centrifugation.
Binding assay of SPP1sus proteins to a YueB780affinity matrix
Extracts of B. subtilis infected with SPP1sus mutants (seeabove) were depleted from proteins that bind non-specificallyto Ni-NTA agarose beads (Qiagen) by adding 5% glycerol and500 ml of beads suspension that had been pre-washed withLB medium. The extracts were left at 4°C on a rotating wheelfor 1 h to saturate Ni-NTA beads. Beads were then removedby centrifugation and the depleted SPP1sus extracts wereapplied to a YueB affinity column.
To prepare the YueB780 affinity matrix, 1 ml of Ni-NTAbeads was washed with ejection buffer (100 mM Tris-Cl,pH 7.5, 300 mM NaCl, 10 mM MgCl2), resuspended in 1 mlof the same buffer supplemented with 50 mM imidazoleand 117 mg of purified His6-tagged YueB780 (670 pmol ofdimers), and incubated on a rotating wheel at 4°C for 1 h.The rationale for the used amount of YueB780 was thefollowing. In the conditions of infection we had 108 cfu ml-1
in a total volume of 200 ml. In a SPP1 wild-type infectionthese cells would release an average of 200 pfu perinfected cell, corresponding to a total of approximately4 ¥ 1012 pfu. These correspond to 6.6 pmol of SPP1. São-José et al. (2006) showed that inactivation of 99% of aSPP1 suspension required receptor/phage molar ratio of~100:1. Therefore, we bound 670 pmol of YueB780 dimersto Ni-NTA beads to guarantee this ratio in our bindingassays. The YueB780 affinity matrix thus prepared waspoured into an Econo-Pac Mini column (Bio-Rad) coupledto a peristaltic pump and washed with two column volumesof ejection buffer supplemented with 50 mM imidazole.Depleted extracts of bacteria infected with SPP1susmutants (see above) were passed twice through the matrix(an aliquot of the flowthrough was kept for analysis). Thematrix was washed with four column volumes of ejectionbuffer supplemented with 50 mM imidazole. Complexes ofYueB780 with affinity bound proteins were eluted by gravityflow with 1.2 ml of ejection buffer supplemented with500 mM imidazole. The eluate was fractionated in 200 mlsamples. For protein analysis, 20 ml of each fraction wasused. The affinity purification procedure was scaled-up for apreparative affinity chromatography in a FPLC system toobtain enough material for mass spectrometry.
Protein analysis by SDS-PAGE and Western blotting
Protein samples were analysed by electrophoresis on 8–10%SDS-PAGE gels and visualized by Coomassie Blue or Silverstaining. Protein blotting onto 0.45 mm nitrocellulose mem-branes (Bio-Rad) was carried out with a Trans-blot Cell TankTransfer system (Bio-Rad). Immuno-detection of YueB780was performed with an anti-YueB780 rabbit polyclonal serumdiluted 1:30 000. Immuno-detection of SPP1 virion proteinswas performed with an anti-SPP1 rabbit polyclonal serumdiluted 1:10 000. Antigen/antibody complexes were detectedeither with the Chemioluminescence Western Blotting Kit(Roche Applied Science) or with ECL (ECL Western BlottingDetection Reagents, GE Healthcare Life Science) and a sec-ondary antibody (Anti-Rabbit IgG, Sigma), according to themanufacturer’s instructions.
MALDI TOF mass spectrometry analysis
Proteins from selected fractions of the SPP1sus31-YueB780binding assay were pooled, concentrated and separated in a10% SDS-PAGE. A ~135 kDa band (Fig. 2D) was recoveredand submitted to complete digestion with trypsin followed bymatrix-assisted laser desorption/ionization-time of flight massspectrometry analysis (MALDI-TOF MS). Peptide mappinganalysis was performed using the Mascot tool (Perkins et al.,1999).
12 I. Vinga et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

Isolation of SPP1 mutants specifically affected inirreversible adsorption
The strategy used to isolate SPP1 mutants deficient inirreversible adsorption is schematized in Fig. 3A and wasadapted from a method previously described to select SPP1mutants specifically impaired in reversible adsorption (Bap-tista, 2009). A CSJ3.gtaC induced culture of 10 ml was grownto exponential phase (A600 = 0.8, about 108 cfu ml-1), centri-fuged and the cellular pellet recovered in 1 ml LB supple-mented with 15 ml MnCl2 (Culture 1). After an equilibrationperiod of 10 min at 45°C, 1 ¥ 1010 pfu of SPP1 were addedand the mixture further incubated for 20 min. A control with nobacteria was treated equally to determine phage input. Afterthis period, cells were pelleted by centrifugation and thesupernatant used to titrate free SPP1 (Round 1) and to resus-pend the pellet of a second 10 ml culture as above (Culture2). After a second period of 20 min incubation at 45°C cellswere pelleted by centrifugation and the supernatant recov-ered for phage titration (Round 2) and for propagation of freephages in solid media at 30°C, using L16601 as host strain.The amplified phages were used to initiate a new cycle of tworounds (SPP1 input = 1 ¥ 1010 pfu ml-1). A total of eight cycleswas performed to obtain an enriched lysate of SPP1 deficientfor irreversible adsorption at 45°C (Fig. 3B).
Measurement of SPP1 reversible and irreversibleadsorption
The efficiency of reversible or irreversible adsorption ofSPP1wt and mutant derivatives was determined as describedpreviously (Baptista et al., 2008) using exponentially growingB. subtilis (A600 = 0.8) and a phage input of 1 ¥ 107 pfu ml-1.
Genome sequencing of SPP1 mutant phages
DNA of SPP1mut phages for sequencing was obtained from250 ml samples of CsCl-purified lysates (~1 ¥ 1012 pfu ml-1) asfollows. Virions were disrupted by treatment with 0.5% SDS,20 mM EDTA pH = 8 and 50 mg ml-1 proteinase K during 1 hat 65°C. After this period proteins were precipitated by adding550 mM KCl (30 min incubation on ice) and removed bycentrifugation. The supernatant containing phage DNA wasextracted twice with a mixture of phenol/chloroform/isoamylalcohol (25:24:1), once with chloroform/isoamyl alcohol(24:1) and finally extensively dialysed against 5 mM Tris-HClpH 7.5. Complete phage genome sequence to detect theposition of point mutations was made using Illumina tech-nology in a HiSeq 2000 sequencer at GATC (Konstanz,Germany). The nucleotide sequence of both strands of over-lapping PCR products covering genes 19.1, 21, 22, 23, 23.1,24 and 24.1 was also determined (service purchased toMacrogen, Seoul, Korea). Oligonucleotide sequences areavailable upon request.
Analysis of the SPP1/YueB interaction in vitro: phageinactivation and DNase protection assays
CsCl-purified SPP1wt and SPP1mut6 were diluted to about5 ¥ 1010 pfu ml-1 in ejection buffer and equilibrated 10 min at
37°C. At time zero, 120 nM of pure YueB780 dimers wereadded and phage inactivation followed over time by plating.The His6-tagged ectodomain YueB780 was overproduced inE. coli and purified as described (São-José et al., 2006). Inanother experiment the same amount of phages was incu-bated with different concentrations of YueB780 (15, 30 and60 nM) in the presence of 1.25 units ml-1 of Benzonase (MerckKGaA). After 1 h of incubation at 37°C, half of the mixtureswere used for phage titration and the other half deproteinizedas described (São-José et al., 2007). Still in another assaySPP1wt was pre-incubated with different sera (1:100 finaldilution) (Fig. 7) for 2h at 37°C before addition of 60 nMYueB780. The reaction was incubated for 1h at 37°C followedby deproteinization as above. The phage titre was determinedafter pretreatment with antibodies and after incubation withYueB780. A sample of the material resulting from these twoexperiments (DNA that remained protected inside virions) wasanalysed in a 0.5% agarose gel (the equivalent to ~ 2 ¥ 108
phage particles of each ejection reaction was loaded). Tocontrol phage input, samples with no antibody pretreatmentand/or with no added receptor were equally processed.
Effect of anti-gp21 antibodies on SPP1 DNA ejection
To study the interference of anti-gp21 antibodies on YueB780-triggered SPP1 DNA ejection CsCl-purified SPP1wt suspen-sions with ~5 ¥ 109 pfu ml-1 were pre-incubated on ice withthe maximum, non-inactivating concentration of a-gp21N ora-gp21C antibodies (1:128 and 1:64 final dilution, respec-tively) in 45 ml TBT buffer, during 1 h. After this period themixtures were transferred to a 96-well, flat bottom, blackmicroplate (Corning) and supplemented with 5 ml of a 24-fold-concentrated, PicoGreen solution in TBT buffer (PicoGreenstock solution was 400-fold concentrated, Invitrogen). Theplates were inserted in an Infinite 200 reader (TECAN) and thefluorescence readings stabilized for 10 min at 37°C. Finally,5 ml of a YueB780 solution at 660 nM was added and fluores-cence measurements taken at 10 s intervals. The ability of theindicated anti-gp21 antibody concentrations to trigger SPP1DNA ejection was performed as above except that noYueB780 was added to samples.
The fluorescence measurement conditions were: excitationwavelength, 485 nm; emission wavelength, 535 nm; excita-tion bandwidth, 9 nm; emission bandwidth, 20 nm; number offlashes, 25; integration time, 20 ms; lag time, 0 ms; settle time,0 ms.
Immunoelectron microscopy
For gp21N labelling about 1 ¥ 1010 pfu of SPP1 in 50 ml TBTwere incubated with 10 ml of a-gp21N IgG overnight at roomtemperature. The samples were diluted by adding 120 ml TBTbuffer and centrifuged in a Beckman Airfuge at 30 psi(repeated 2x) to remove unbound IgG antibodies. Final pelletwas resuspended in about 20 ml TBT.
For gp21C labelling about 1 ¥ 1010 pfu of SPP1 in 40 mlTBT were incubated with 5 ml of affinity purified a-gp21C IgGfor 3 h at room temperature. Due to the high specificity ofa-gp21C IgG the samples required no further purification.
Aliquots of 2 ml from both samples were used for negativestaining with uranyl acetate (Steven et al., 1988). For gold
SPP1 tail spike protein binding to the YueB receptor 13
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

labelling 1 ml of goat anti-rabbit IgG complexed with 5 nmcolloidal gold (BritishBioCell, batch 10624) was added to thesamples. After incubation overnight at 4°C the gold labelledsamples were negatively stained as above.
Acknowledgements
We are most thankful to Dr Jean-Pierre Le Caer (ICSN,Gif-sur-Yvette, France) for carrying out the mass spectrom-etry analysis and for useful discussions. Paulette Decot-tignies (IBBMC, Orsay, France) is acknowledged for proteinamino-terminus sequencing. Sandrine Brasilès (VMS, Gif-sur-Yvette) kindly provided purified YueB780. We areindebted to João Nascimento for helpful discussions duringthe design of the method to isolate SPP1 mutants. This workhas been supported through Grant PTDC/BIA-MIC/66412/2006 (to C.S.-J.) from Fundação para a Ciência e a Tecno-logia (FCT, MCTES, Portugal) and Grant BLAN07-1_191968from Agence Nationale de la Recherche (ANR, France). I.V.and C.B. work has been supported through post-doc fellow-ships SFRH/BPD/34892/2007 and SFRH/BPD/63392/2009,respectively (FCT, MCTES, Portugal).
References
Adams, M.H. (1959) Bacteriophages. New York, NY: WileyInterscience.
Alonso, J.C., Lüder, G., Stiege, A.C., Chai, S., Weise, F., andTrautner, T.A. (1997) The complete nucleotide sequenceand functional organization of Bacillus subtilis bacterioph-age SPP1. Gene 204: 201–212.
Alonso, J.C., Tavares, P., Lurz, R., and Trautner, T.A. (2006)Bacteriophage SPP1. In The Bacteriophages. Calendar, R.(ed.). New York, NY: Oxford University Press, pp. 331–349.
Andres, D., Hanke, C., Baxa, U., Seul, A., Barbirz, S., andSeckler, R. (2010) Tailspike interactions with lipopolysac-charide effect DNA ejection from phage P22 particles invitro. J Biol Chem 285: 36768–36775.
Auzat, I., Dröge, A., Weise, F., Lurz, R., and Tavares, P.(2008) Origin and function of the two major tail proteins ofbacteriophage SPP1. Mol Microbiol 70: 557–569.
Baptista, C. (2009) Recognition of Bacillus subtilis receptorsby bacteriophage SPP1: a genetic and biochemicalapproach. Ph.D thesis, Faulty of Sciences, University ofLisbon [WWW document]. URL http://hdl.handle.net/10451/1638
Baptista, C., Santos, M.A., and São-José, C. (2008) PhageSPP1 reversible adsorption to Bacillus subtilis cell wallteichoic acids accelerates virus recognition of membranereceptor YueB. J Bacteriol 190: 4989–4996.
Bebeacua, C., Bron, P., Lai, L., Vegge, C.S., Brøndsted, L.,Spinelli, S., et al. (2010) Structure and molecular assign-ment of lactococcal phage TP901-1 baseplate. J Biol Chem285: 39079–39086.
Becker, B., de la Fuente, N., Gassel, M., Günther, D.,Tavares, P., Lurz, R., et al. (1997) Head morphogenesisgenes of the Bacillus subtilis bacteriophage SPP1. J MolBiol 268: 822–839.
Böhm, J., Lambert, O., Frangakis, A.S., Letellier, L.,Baumeister, W., and Rigaud, J.L. (2001) FhuA-mediated
phage genome transfer into liposomes: a cryo-electrontomography study. Curr Biol 11: 1168–1175.
Dai, W., Hodes, A., Hui, W.H., Gingery, M., Miller, J.F., andZhou, Z.H. (2010) Three-dimensional structure of tropism-switching Bordetella bacteriophage. Proc Natl Acad SciUSA 107: 4347–4352.
Dröge, A., and Tavares, P. (2000) In vitro packaging of DNAof the Bacillus subtilis bacteriophage SPP1. J Mol Biol 296:103–115.
Dröge, A., Santos, M.A., Stiege, A.C., Alonso, J.C., Lurz, R.,Trautner, T.A., and Tavares, P. (2000) Shape and DNApackaging activity of bacteriophage SPP1 procapsid:protein components and interactions during assembly.J Mol Biol 296: 117–132.
Duplessis, M., and Moineau, S. (2001) Identification of agenetic determinant responsible for host specificity inStreptococcus thermophilus bacteriophages. Mol Microbiol41: 325–336.
Goulet, A., Lai-Kee-Him, J., Veesler, D., Auzat, I., Robin, G.,Shepherd, D.A., et al. (2011) The opening of the SPP1bacteriophage tail, a prevalent mechanism in Gram-positive-infecting siphophages. J Biol Chem 286: 25397–25405.
Gual, A., Camacho, A.G., and Alonso, J.C. (2000) Functionalanalysis of the terminase large subunit, G2P, of Bacillussubtilis bacteriophage SPP1. J Biol Chem 275: 35311–35319.
Harlow, E., and Lane, D. (1988) Antibodies. A LaboratoryManual, Cold Spring Harbor, NY: Cold Spring Harbor Labo-ratory Press.
Hendrix, R.W., and Duda, R.L. (1992) Bacteriophage lambdaPaPa: not the mother of all lambda phages. Science 258:1145–1148.
Jakutytè, L., Baptista, C., São-José, C., Daugelavicius, R.,Carballido-López, R., and Tavares, P. (2011) Bacterioph-age infection in rod-shaped gram-positive bacteria: evi-dence for a preferential polar route for phage SPP1 entry inBacillus subtilis. J Bacteriol 193: 4893–4903.
Kaneko, J., Narita-Yamada, S., Wakabayashi, Y., and Kamio,Y. (2009) Identification of ORF636 in phage phiSLTcarrying Panton-Valentine leukocidin genes, acting as anadhesion protein for a poly(glycerophosphate) chain oflipoteichoic acid on the cell surface of Staphylococcusaureus. J Bacteriol 191: 4674–4680.
Katsura, I., and Kühl, P.W. (1975) Morphogenesis of the tail ofbacteriophage lambda. III. Morphogenetic pathway. J MolBiol 91: 257–273.
Letellier, L., Boulanger, P., Plançon, L., Jacquot, P., and San-tamaria, M. (2004) Main features on tailed phage, hostrecognition and DNA uptake. Front Biosci 9: 1228–1339.
Lhuillier, S., Gallopin, M., Gilquin, B., Brasilès, S., Lancelot,N., Letellier, G., et al. (2009) Structure of bacteriophageSPP1 head-to-tail connection reveals mechanism for viralDNA gating. Proc Natl Acad Sci USA 106: 8507–8512.
Margot, P., and Karamata, D. (1996) The wprA gene ofBacillus subtilis 168, expressed during exponential growth,encodes a cell-wall-associated protease. Microbiology142: 3437–3444.
Okubo, S., and Yanagida, T. (1968) Isolation of a suppressormutant in Bacillus subtilis. J Bacteriol 95: 1187–1188.
Orlova, E.V., Gowen, B., Dröge, A., Stiege, A., Weise, F.,
14 I. Vinga et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology

Lurz, R., et al. (2003) Structure of a viral DNA gatekeeperat 10 Å resolution by cryo-electron microscopy. EMBO J22: 1255–1262.
Perkins, D.N., Pappin, D.J., Creasy, D.M., and Cottrell, J.S.(1999) Probability-based protein identification by searchingsequence databases using mass spectrometry data. Elec-trophoresis 20: 3551–3567.
Plisson, C., White, H.E., Auzat, I., Zafarani, A., São-José, C.,Lhuillier, S., et al. (2007) Structure of bacteriophage SPP1tail reveals trigger for DNA ejection. EMBO J 26: 3720–3728.
Ravin, V., Räisänen, L., and Alatossava, T. (2002) A con-served C-terminal region in Gp71 of the small isometric-head phage LL-H and ORF474 of the prolate-head phageJCL1032 is implicated in specificity of adsorption of phageto its host, Lactobacillus delbrueckii. J Bacteriol 184: 2455–2459.
Ricagno, S., Campanacci, V., Blangy, S., Spinelli, S., Trem-blay, D., Moineau, S., et al. (2006) Crystal structure of thereceptor-binding protein head domain from Lactococcuslactis phage bIL170. J Virol 80: 9331–9335.
Riva, S., Polsinelli, M., and Falaschi, A. (1968) A new phageof Bacillus subtilis with infectious DNA having separablestrands. J Mol Biol 35: 347–356.
Sambrook, J., and Russell, D.W. (2001) Molecular Cloning: ALaboratory Manual, 3rd edn. Cold Spring Harbor, NY: ColdSpring Harbor Laboratory Press.
São-José, C., Baptista, C., and Santos, M.A. (2004) Bacillussubtilis operon encoding a membrane receptor for bacte-riophage SPP1. J Bacteriol 186: 8337–8346.
São-José, C., Lhuillier, S., Lurz, R., Melki, R., Lepault, J.,Santos, M.A., and Tavares, P. (2006) The ectodomain of theviral receptor YueB forms a fiber that triggers ejectionof bacteriophage SPP1 DNA. J Biol Chem 281: 11464–11470.
São-José, C., de Frutos, M., Raspaud, E., Santos, M.A., andTavares, P. (2007) Pressure built by DNA packing insidevirions: enough to drive DNA ejection in vitro, largely insuf-ficient for delivery into the bacterial cytoplasm. J Mol Biol374: 346–355.
Sciara, G., Blangy, S., Siponen, M., Mc Grath, S., van Sin-deren, D., Tegoni, M., et al. (2008) A topological model ofthe baseplate of lactococcal phage Tuc2009. J Biol Chem283: 2716–2723.
Sciara, G., Bebeacua, C., Bron, P., Tremblay, D., Ortiz-Lombardia, M., Lichière, J., et al. (2010) Structure of lac-
tococcal phage p2 baseplate and its mechanism ofactivation. Proc Natl Acad Sci USA 107: 6852–6857.
Steven, A.C., Trus, B.L., Maizel, J.V., Unser, M., Parry, D.A.D., Wall, J.S., et al. (1988) Molecular substructure of a viralreceptor-recognition protein. The gp17 tail-fiber of bacte-riophage T7. J Mol Biol 200: 351–365.
Veesler, D., Robin, G., Lichière, J., Auzat, I., Tavares, P.,Bron, P., et al. (2010a) Crystal structure of bacteriophageSPP1 distal tail protein (gp19.1): a baseplate hub paradigmin gram-positive infecting phages. J Biol Chem 285:36666–36673.
Veesler, D., Blangy, S., Spinelli, S., Tavares, P., Campanacci,V., and Cambillau, C. (2010b) Crystal structure of Bacillussubtilis SPP1 phage gp22 shares fold similarity with adomain of lactococcal phage p2 RBP. Protein Sci 19:1439–1443.
Veesler, D., Blangy, S., Lichière, J., Ortiz-Lombarda, M.,Tavares, P., Campanacci, V., and Cambillau, C. (2010c)Crystal structure of Bacillus subtilis SPP1 phage gp23.1, aputative chaperone. Protein Sci 19: 1812–1816.
Vinga, I., São-José, C., Tavares, P., and Santos, M.A. (2006)Bacteriophage entry in the host cell. In Modern Bacterioph-age Biology and Biotechnology. Wegrzyn, G. (ed.). Kerala:Research Signpost, pp. 165–203.
Wang, G., Klein, M.G., Tokonzaba, E., Zhang, Y., Holden,L.G., and Chen, X.S. (2008) The structure of a DnaB-familyreplicative helicase and its interactions with primase. NatStruct Mol Biol 15: 94–100.
Wang, J., Hofnung, M., and Charbit, A. (2000) The C-terminalportion of the tail fiber protein of bacteriophage lambda isresponsible for binding to LamB, its receptor at the surfaceof Escherichia coli K-12. J Bacteriol 182: 508–512.
Xiang, Y., Leiman, P.G., Li, L., Grimes, S., Anderson, D.L.,and Rossmann, M.G. (2009) Crystallographic insights intothe autocatalytic assembly mechanism of a bacteriophagetail spike. Mol Cell 34: 375–386.
Supporting information
Additional supporting information may be found in the onlineversion of this article.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for the article.
SPP1 tail spike protein binding to the YueB receptor 15
© 2011 Blackwell Publishing Ltd, Molecular Microbiology
Copyright © 2022 FDOKUMEN




















