
Nutrient relationships in shallow water in an African Lake, Lake Naivasha

IN SITU MEASUREMENT OF DAPHNIA LONGISPINA GRAZING ONALGAE AND BACTERIA IN A HIGH MOUNTAIN LAKE (LAKE
PAIONE SUPERIORE, NORTHERN ITALY) USING FLUORESCENTLYLABELED CELLS
NICOLETTA RICCARDIIstituto per lo Studio degli Ecosistemi – CNR, Largo Tonolli 50, 28922 Verbania Pallanza, Italy
(∗ author for correspondence, e-mail: [email protected], fax: +39 0323 556513)
(Accepted 18 March 2001)
Abstract. Grazing impact of Daphnia longispina on phytoplankton and bacteria in Lake PaioneSuperiore (Northern Italy) was evaluated using fluorescently labeled cells in short-term in situ exper-iments. Structure and size distribution of Daphnia were studied weekly from its appearance in Julyto its decline in September. Relative importance of algae and bacteria in the diet varied over time,as did consequently the relative impact of Daphnia on the phytoplankton and bacteria populations.Biomass of bacteria in the diet of Daphnia varied from 1 to 42%, and was highest at the beginningof population development when young individuals dominated. The percent biomass of algae in thediet increased with population development, ranging from 58 to 99%. Daphnia population consumedper hour 0.04–1.8% of bacteria and 0.3–17.6% of phytoplankton abundances.
Keywords: algae and bacteria, Daphnia longispina, fluorescence labeling, high mountain lake, in-gestion rates, population grazing, zooplankton abundance
1. Introduction
High mountain lakes are often regarded as ‘laboratories’ for acquiring basic in-formation on the flux of energy through the ecosystem; their oligotrophy and ex-treme physical conditions combine to set limits to the development of communities,resulting in extremely simplified biological communities with relatively simpletrophic links among the different compartments.
Limnological studies in recent years on the oligotrophic Lake Paione Superi-ore have revealed a very simply structured plankton community where the keyorganisms are bacteria, flagellates and Daphnia (Callieri et al., 1999). Daphnia canattain unexpectedly dense populations comprising well-fed organisms, visibly richin lipids and able to produce more than one generation/year of parthenogenetic fe-males at high densities (Cammarano and Manca, 1997; Manca and Comoli, 1999).Simultaneously with the increase of Daphnia density during late summer, a declinein phytoplankton density (Pugnetti and Bettinetti, 1999; Manca and Comoli, 1999)and a decrease in the carbon fraction immobilized in the microbial loop and phyto-plankton were observed (Callieri et al., 1999). This prompted the hypothesis that
Water, Air, and Soil Pollution: Focus 2: 343–357, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.

344 N. RICCARDI
Daphnia exploits both flagellate and bacteria populations to sustain its populationdevelopment.
The food sources actually exploited by Daphnia in LPS (Lake Paione Superi-ore), and their relative importance in its diet, were the questions behind this study,which was aimed at evaluating the grazing impact of Daphnia on LPS phytoplank-ton and bacterioplankton.
1.1. STUDY SITE
Lake Paione Superiore is a small glacial cirque lake (surface area 0.014 km2;max depth 11.7 m; volume 69 × 106 m3) lying on acidic bedrocks (gneiss) at analtitude of 2269 m in the Bognanco Valley (Pennine Alps, Italy). The lake is oligo-trophic (SRP < 5 µg L−1) with a mean pH of 5.82 and a mean alkalinity and Ca2+concentration of 7 and 42 µeq L−1, respectively (The MOLAR Water ChemistryGroup, 1999). The ice-free period generally lasts from May–June to November.Autotrophic picoplankton and picocyanobacteria are negligible, heterotrophic pi-coplankton (bacteria) are at their maximum density (corresponding to about 60 µgC L−1) in August–September (Callieri and Bertoni, 1999). The mean biomass ofheterotrophic nanoflagellates, which are mainly represented by <8 µm cells, rangesfrom 3 to 5 µg C L−1; mean ciliate biomass is about 2 µg C L−1 (Callieri etal., 1999). Phytoplankton association comprises relatively few species, mainly ofnanoplanktonic unicellular algae, the only exception being the colonial Dinobryonsertularia (Pugnetti and Bettinetti, 1999). The most important species as regardsbiomass and density are Chromulina sp., Dinobryon sertularia, Mallomonas alve-olata and Gymnodinium sp. (Pugnetti and Bettinetti, 1999; Callieri et al., 1999).Zooplankton association is also made up of a limited number of species, amongwhich Cyclops abyssorum tatricus and Daphnia longispina dominate during theice-free season, with rotifers (mainly Keratella quadrata) dominating during theice-cover period (Cammarano and Manca, 1997; Manca and Comoli, 1999). Arcto-diaptomus alpinus, present in the lake before the 1950s (Tonolli, 1949), has dis-appeared, probably because of lake acidification, and has not been found in recentyears. Daphnia longispina generally appears in August and peaks in September,reaching densities of about 10 ind L−1, exceptionally even higher (i.e. 20 ind L−1,Manca and Comoli, 1999). The ability of Daphnia to reach relatively high densitieseven at the extremely low average food concentrations in the lake is favored by theabsence of fish and invertebrate predators other than Cyclops.
2. Material and Methods
2.1. ZOOPLANKTON SAMPLING AND MEASUREMENTS
Zooplankton was collected about weekly from the end of July to the end of Septem-ber 1999. Semi-quantitative samples were collected by duplicate vertical net

DAPHNIA LONGISPINA IN AN ACIDIC LAKE 345
(126 µm mesh) hauls from the bottom to the surface at the point of maximumdepth. Two samples corresponding to about 600 L filtered water were collected oneach date: one sample was fixed in 4% formaldehyde for microscope countings,while the other sample was used for sorting Daphnia specimens for in situ grazingexperiments. The fixed zooplankton sample was used for microscope countingsand measurements. Total length (excluding terminal spine and furcal setae) andmaximum width were measured by an ocular micrometer on at least 200 indi-viduals of the most abundant species (Daphnia longispina and Cyclops abyssorumtatricus); in few specimen samples the total number of individuals present wasmeasured. The individual dry weight of Cyclops copepodites and adults was calcu-lated from length/weight regression equations (McCauley, 1984); that of Daphniawas measured after drying at 105 ◦C for 24 hr. Carbon content was estimated fromdry weight, taking carbon = 0.5 d.w.
2.2. In situ GRAZING EXPERIMENTS
In situ grazing experiments were performed on five dates (27 July, 11, 17 and31 August, and 8 September), i.e. from the beginning of the Daphnia popula-tion increase through its maximum development until the beginning of its decline.Feeding rates were determined by the use of fluorescently labeled algae (FLA)and bacteria (FLB) as markers. This method was chosen mainly for three reasons:(a) the experiments can be done in a short time, which avoids the problem of longincubations, inconvenient in poorly accessible high mountain lakes; (b) bacteriaand algae ingestion rates can be determined at the same time in the same treatment;and (c) the method offers some advantages over radiotracer techniques, such asthe possibility of adapting incubation time to gut retention time and inspectinganimals for labeled cells absorption on external body surfaces (e.g. Wiedner andVareschi, 1995). A deciding advantage of this method for us was the possibilityof comparing Daphnia grazing with flagellate grazing, measured using the sametechnique (Corno, 2000).
Laboratory cultures of cocci and rods (mean diameters 1.13 and 0.68 µm, meanvolume 0.52 µm3) and Chlamydomonas reinhardi (mean diameters 7.7 and 6.3 µm;mean volume 160 µm3) were chosen as tracers because they are in the middle of thesize range of the natural bacteria and phytoplankton populations in LPS (biovolumeof bacteria: mean = 0.49 µm3, range = 0.35–0.70 µm3; biovolume of algae: mean =525 µm3, median = 165 µm3, range 8–2870 µm3). Algae and bacteria were stainedwith 5(4,6-dichlorotriazin-2-yl) amino-fluorescein (DTAF) (Sherr et al., 1987).
Experiments were carried out only on adult Daphnia specimens, to reduce anyvariability which might be attributable to variations in ingestion rates in anim-als of different sizes (e.g. Lampert, 1987). Daphnia density in each experimentalbottle corresponded to 20 ind L−1, which is the maximum density observed duringprevious years at the maximum population peak (Manca and Comoli, 1999).

346 N. RICCARDI
Four polycarbonate bottles (1.5-L) were filled with 500 mL of epilimnetic water(filtered through a 126 µm Nylon mesh to remove zooplankton) and ten activelyswimming Daphnia were carefully added to each of them. Two additional bottles,one without and one with daphnids, were kept as controls for phytoplankton andbacteria countings. Immediately after the addition of FLB and FLA, a 10 mL sub-sample was taken from the control bottles to estimate the concentration of tracerparticles and natural bacteria and phytoplankton. The amount of FLB and FLAadded to each experiment was calculated from previous countings of natural phyto-and bacterioplankton to obtain a final tracer concentration of 25% of the total num-ber of cells. Due to variations in natural phytoplankton and bacteria densities, thefinal concentration of FLA in the experiments ranged from 6 to 41%, and that ofFLB from 8 to 23%. All the treatments and the control were incubated in the epi-limnion (temperature 11.7–15.5 ◦C) for 5 to 20 min. Incubation times were chosenon the basis of previous laboratory tests and according to reported gut retentiontimes for cladocerans (Haney, 1973; Geller, 1975; Cauchie et al., in press). After5, 10, 15 and 20 min the daphnids were gently collected by filtration on a 25 µmmesh, narcotized in carbonated water to prevent egestion of gut contents, rinsedwith sterile Milli-Q water and preserved in 1% formaldehyde. A water samplefor bacteria and phytoplankton countings was collected after 20 min from boththe control bottles. In the laboratory, after microscopic observation to confirm theremoval of uningested FLA and FLB from the animals’ body surface, the daphnidsof each treatment were sonicated in 10 mL sterile water at 50 W 3 times for 15 secwith a Branson sonifier-cell disrupter. One mL of this suspension was filtered on0.2 µm gold stained Nucleopore filters and bacteria stained with DAPI; 1 mL ofthe same suspension was filtered on 2 µm gold stained Nucleopore filters for FLAenumeration. Random strips on each 0.2 µm filter were counted in epifluorescenceat 1250× until about 400 FLB and 400 natural bacteria were counted; followingthe same procedure, FLA were counted on 2 µm filters at 400×.
Uptake rates (algae-FLA ind−1 hr−1 and bacteria-FLB ind−1 hr−1) after 5, 10,15 and 20 min were calculated from the number of labeled cells ingested, multi-plied by the ratio of (algal or bacteria cells). (FLA or FLB)−1. Uptake rates werealso evaluated from the difference between the numbers of FLA and FLB at the be-ginning and after 20 min in the control treatments with animals, to confirm the dataresulting from direct counting of ingested particles. The two estimates coincidedquite well for FLA, while the differences in FLB concentration before and afterincubation were undetectable, falling in the range of statistical variation. Ingestionrates were calculated as the initial slope of the uptake vs the time curve (Figure 1).The selectivity coefficient was calculated as the ratio between the clearance rate ofone item and that of the most preferred food item (Knisely and Geller, 1986).
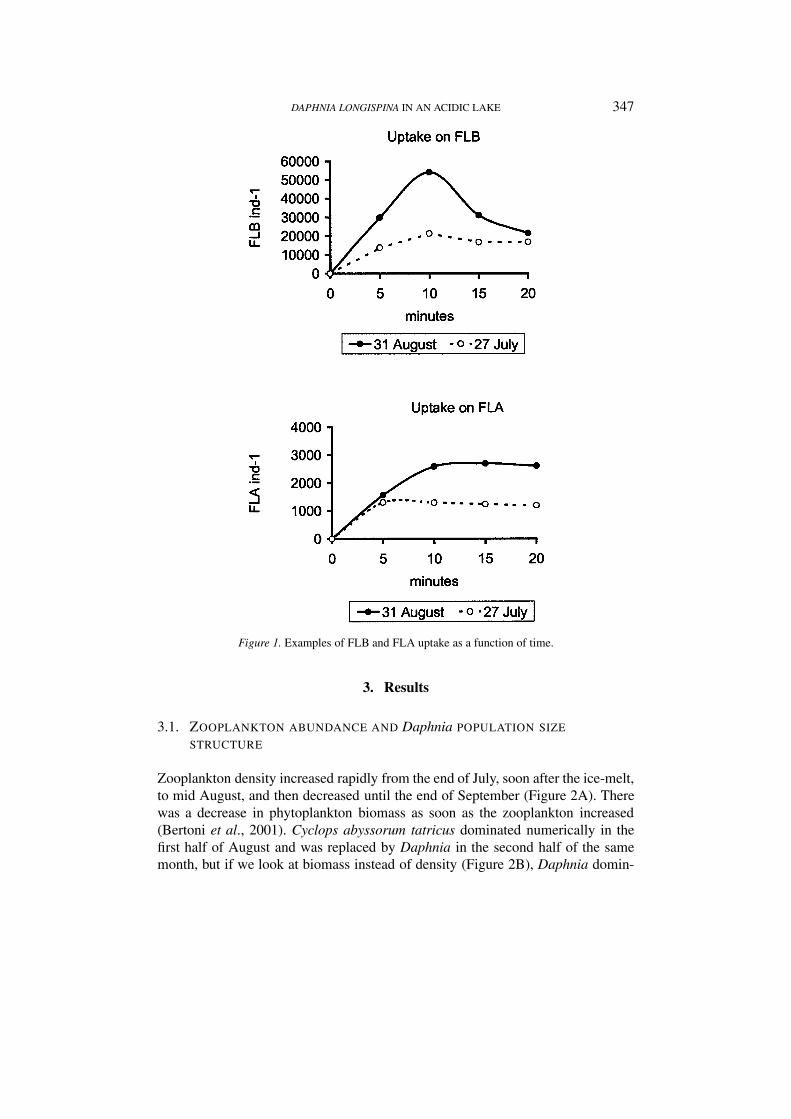
DAPHNIA LONGISPINA IN AN ACIDIC LAKE 347
Figure 1. Examples of FLB and FLA uptake as a function of time.
3. Results
3.1. ZOOPLANKTON ABUNDANCE AND Daphnia POPULATION SIZE
STRUCTURE
Zooplankton density increased rapidly from the end of July, soon after the ice-melt,to mid August, and then decreased until the end of September (Figure 2A). Therewas a decrease in phytoplankton biomass as soon as the zooplankton increased(Bertoni et al., 2001). Cyclops abyssorum tatricus dominated numerically in thefirst half of August and was replaced by Daphnia in the second half of the samemonth, but if we look at biomass instead of density (Figure 2B), Daphnia domin-

348 N. RICCARDI
Figure 2. (A) Density of the different zooplankton taxa and biovolume of phytoplankton in surface(–1 m) water. (B) Biomass (µg C L−1) of Cyclops abyssorum tatricus and Daphnia longispina.(C) Population structure of Cyclops abyssorum tatricus. (D) Population structure of Daphnialongispina, percentage of ovigerous females and mean clutch size.
ated over the whole period. In fact, the Cyclops abyssorum population was madeup of a high number of nauplia compared to a relatively low number of copepoditesand rare adults (Figure 2C). At the end of July, when Daphnia appeared, the pop-ulation mainly comprised small females (Figure 3). As the body size of the cohortincreased, the smallest size fraction of the population was almost completely madeup of males which appeared in the first half of August and increased in number untilthe beginning of September. The percentage of ovigerous females out of the totalnumber of females ranged from 1.7 to 8.2 (Figure 2D), with the highest value beingfound one week before the population peak on 31 August. The mean clutch sizewas generally low, except on 4 August when the first generation of parthenogeneticfemales, which had appeared at the end of July, matured. The lowest percentage ofovigerous females was observed on this date, but up to 15 eggs were counted in the
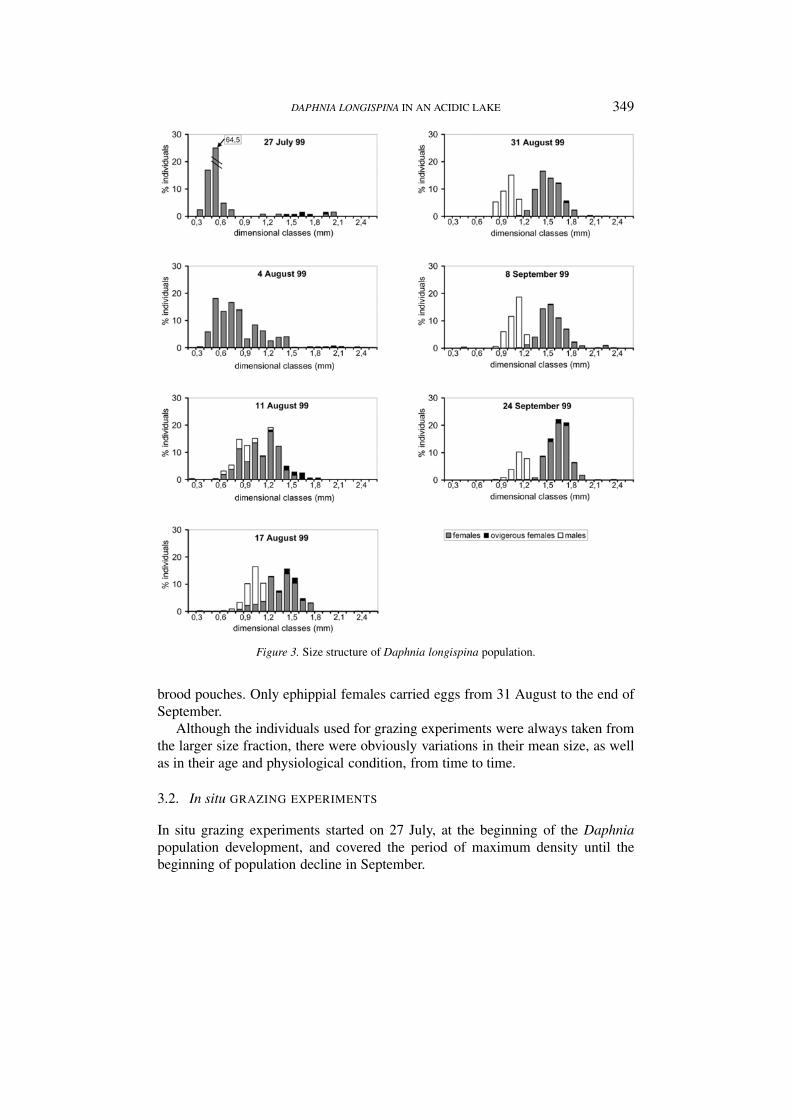
DAPHNIA LONGISPINA IN AN ACIDIC LAKE 349
Figure 3. Size structure of Daphnia longispina population.
brood pouches. Only ephippial females carried eggs from 31 August to the end ofSeptember.
Although the individuals used for grazing experiments were always taken fromthe larger size fraction, there were obviously variations in their mean size, as wellas in their age and physiological condition, from time to time.
3.2. In situ GRAZING EXPERIMENTS
In situ grazing experiments started on 27 July, at the beginning of the Daphniapopulation development, and covered the period of maximum density until thebeginning of population decline in September.

350 N. RICCARDI
In all the experiments the number of ingested fluorescently labeled cells perindividual increased to its maximum after 5 to 15 min, when it reached a plateau,after which it remained more or less constant over time or decreased (Figure 1).This suggests that an equilibrium was reached between ingestion and digestion oregestion/defecation rates. The initial slope of the curve of uptake vs time representsthe ingestion rate before the beginning of particle digestion or defecation, while theslope of the curve after the inflection point is determined by the difference betweenthe rate of ingestion and that of digestion/defecation. The initial slope of the curveprovides an estimate of ingestion rates not affected by the loss of tracer throughdefecation.
Ingestion and clearance rates of both bacteria and algae varied widely over time,but algae were always cleared at a consistently higher rate than bacteria (Table I).The marked preference for algae is evident from selectivity coefficients, the valuesof which ranged from 0.04 to 0.20 (Table I). The lowest value, recorded on 8September, was the result of an increase in the feeding rate on algae rather thana decline in the feeding rate on bacteria (Figure 4A, Table I).
Bacteria and algae were ingested at the highest rate during the phase of Daphniapopulation increase before mid August; later on, the ingestion rate of bacteriawas reduced by about 7 fold and that of algae 3 fold. Consequently, the rate oftotal biomass ingestion (algae and bacteria) appeared to be considerably higherwhen the population abundance was rapidly increasing and more biomass wasavailable (Figure 4A). Algae ingestion rates were in good agreement with phyto-plankton abundance in surface waters (Figure 4B), while bacteria ingestion rateswere related neither to bacteria availability nor to algae ingestion rates.
The total biomass consumed by one Daphnia in one hour ranged from 0.5 to2.8 µg C, or 2 to 10% of its own biomass.
Our results suggest that the Daphnia population could consume from 0.04 to1.8% of bacteria abundance and 0.3 to 17.6% of phytoplankton abundance per hour(Figure 5). These values are obviously an overestimation of the actual impact ofDaphnia population on phyto- and bacterioplankton, being based on the ingestionrates of adult individuals. Since juveniles were abundant only on 27 July, the riskof overestimation should be highest on this date.
4. Discussion and Conclusions
4.1. METHODOLOGICAL CONSIDERATIONS
The method used for the experiments was relatively simple and had the advantageof estimating grazing rates within a time shorter than the gut passage time of theanimals, thus avoiding the loss of labeled particles through defecation. In fact, bymeasuring uptake over several incubation times it was possible to highlight thebeginning of the digestion or egestion/defecation process. Another source of po-

DAPHNIA LONGISPINA IN AN ACIDIC LAKE 351
TAB
LE
I
Tem
pera
ture
ofth
esu
rfac
ew
ater
inL
ake
Paio
neS
uper
iore
,ind
ivid
uali
nges
tion
rate
and
clea
ranc
era
teon
bact
eria
and
alga
e,an
dse
lect
ivit
yco
effi
cien
t
Tem
pB
acte
ria
inge
stio
nra
teA
lgae
inge
stio
nra
teC
lear
ance
rate
Sel
ectiv
ity
◦ CC
ell
106
µm
3µ
gC
Cel
l10
6µ
m3
µg
C(a
lgae
)(b
acte
ria)
coef
fici
ent
(ind
−1hr
−1)
(ind
−1hr
−1)
(mL
ind−
1hr
−1)
27Ju
ly15
.533
1824
62.
390.
3145
1923
5812
.37
2.47
329
.83
3.14
0.11
11A
ugus
t13
.949
6360
12.
070.
3171
3352
82.
830.
565
21.0
94.
140.
20
17A
ugus
t11
.781
2834
1.09
0.06
4219
388
2.33
0.46
64.
470.
600.
13
31A
ugus
t12
.496
7482
0.40
0.06
1064
540
5.70
1.14
016
.92
1.03
0.06
08S
epte
mbe
r12
.270
1139
0.30
0.04
2456
384
5.84
1.16
818
.70
0.67
0.04
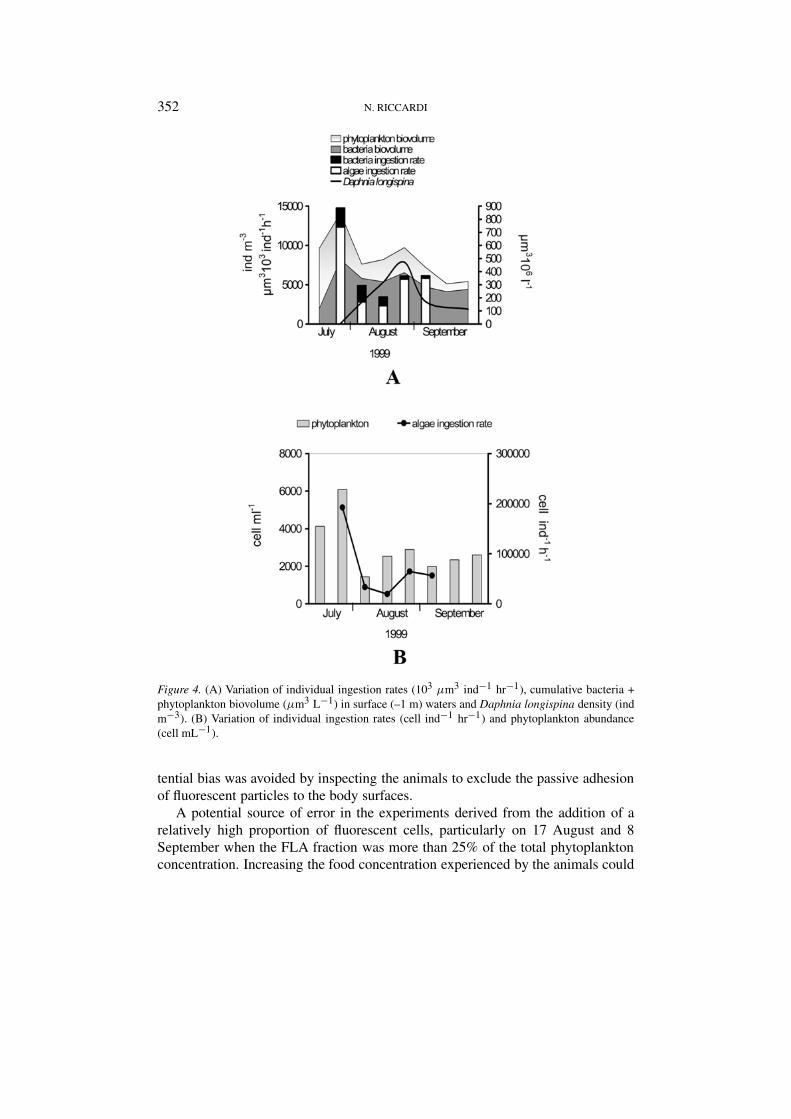
352 N. RICCARDI
Figure 4. (A) Variation of individual ingestion rates (103 µm3 ind−1 hr−1), cumulative bacteria +phytoplankton biovolume (µm3 L−1) in surface (–1 m) waters and Daphnia longispina density (indm−3). (B) Variation of individual ingestion rates (cell ind−1 hr−1) and phytoplankton abundance(cell mL−1).
tential bias was avoided by inspecting the animals to exclude the passive adhesionof fluorescent particles to the body surfaces.
A potential source of error in the experiments derived from the addition of arelatively high proportion of fluorescent cells, particularly on 17 August and 8September when the FLA fraction was more than 25% of the total phytoplanktonconcentration. Increasing the food concentration experienced by the animals could

DAPHNIA LONGISPINA IN AN ACIDIC LAKE 353
Figure 5. Percent consumption of bacteria and algae abundance by Daphnia longispina population.
cause a reduction in their filtering activity and lead to an underestimation of theactual clearance rate. However, since feeding activity is also controlled by gutfullness, and because the ingestion rates were estimated on incubation times prob-ably shorter than gut filling time, the effect of any deviation from the ideal tracerassumption may still be negligible. There is also the fact that Daphnia regulates itsfeeding activity through physiological mechanisms of adaptation to environmentalfood conditions (e.g. Lampert, 1987, 1994). In our experiments (and particularlythose on 17 August and 8 September), the fact that the animals were not pre-adapted to increased food levels might have caused an error in the estimate of in-gestion rates by affecting the initial slope of the functional response (e.g. Lampert,1994).
However, our results may have been affected by other uncertainties, some ofthem common to all short term experiments based on the uptake of labeled particles(e.g Haney, 1985; Wiedner and Vareschi, 1995).
As Daphnia seems to select food particles mainly by size (e.g. Gophen andGeller, 1984), we tried to minimize the potential bias involved in this selective feed-ing by using FLA of a size close to the optimal range for D. longispina (Gliwicz,1977) and in the middle of the size range of phytoplankton in LPS.
4.2. FEEDING RATES
Ingestion and filtering rates of algae in our study (Table I) are of the same orderof magnitude or even higher than the high ingestion rates measured by Holm et al.(1983) for D. pulex fed Aphanizomenon flos-aquae (12 000–50 000 cell ind−1 hr−1)and by Ravera and Scotto (1999) for mixed zooplankton communities (up to 12 400cell ind−1 hr−1). A number of factors, such as species, body size, nutritional history,food quality and quantity, temperature, directly or indirectly influence grazing andclearance rates (e.g. Lampert, 1987). According to Callieri et al. (1999), the Daph-nia population in LPS is probably not limited by food availability, since it starts

354 N. RICCARDI
to grow when the microorganism carbon concentration is above the threshold foodconcentration for the growth of large-bodied species (Gliwicz, 1990). However,there is a marked discrepancy between the threshold limits reported by differentauthors (e.g. Duncan et al., 1985; Gliwicz, 1990; Lampert, 1994) which can beexplained, according to Lampert (1994), by differences in the nutritional history ofthe animals. Very low threshold limits can be the result of preadaptation to low-food levels due to phenotypic responses, such as filter screen enlargement andchanges in gut capacity and digestion, which lead to more efficient food uptake(Lampert, 1994). If we accept the threshold limits given by Lampert (1994), theconclusion must be that Daphnia was limited by food availability; on the otherhand, the development of a numerically large population is a clear suggestion thatindividuals adapted to low-food levels. Whatever physiological mechanism wasinvolved, this adaptation could provide an explanation for the remarkably highingestion rates of algae.
Again, the variability deriving from the use of different methodologies must betaken into account in any comparison of grazing rates reported by different au-thors. According to Lampert (1987), only short-term estimates not affected by lossthrough defecation can measure true ingestion rates, while long-term experimentscannot measure a real ingestion rate, yielding a value closer to an assimilation rate.It follows that higher ingestion and clearance rates should result from short-termthan from long-term experiments.
Grazing rates on bacteria (Table I) were near the upper limit of the range ofliterature values (reviewed by Jürgens, 1994) and were always one order of mag-nitude lower than grazing rates on algae. As already observed in many cases (e.g.Porter et al., 1983; Kankaala, 1988; Jürgens et al., 1994), Daphnia feeds less effi-ciently on bacteria than on algae. It has been suggested that bacteria serve only as asupplementary food for zooplankton (Porter, 1984), and only in certain situations,for example when the availability of algal food is limited, can they became thedominant food source (e.g. Børsheim and Olsen, 1984; Kankaala, 1988; Jürgens,1994). In our study, bacteria represented a relatively small fraction of the diet ofDaphnia compared to algae, but their importance increased to 42% of the totalingested biomass when phytoplankton availability was reduced in August. Sincethe P-concentration of bacteria is generally high compared to that of algae (e.g.Vadstein et al., 1988) bacteria can represent an important source of P for filterfeed-ing zooplankton (Hessen and Andresen, 1990). Since P-limitation seems to cause adecrease in the nutritional value of algae for zooplankton (e.g. Gulati and DeMott,1997), it may be that bacteria consumption in an oligotrophic lake like LPS canhelp to compensate for any possible nutritional deficiency in Daphnia’s diet.
The variations over time in the total biomass ingested by an individual probablyreflected both the physiological condition of the individuals and food availabil-ity. The highest carbon uptake was measured in young adults on 27 July, whenpopulation density was lowest. As population density increased and individualsage, a decrease in food uptake probably resulted from the increased competition

DAPHNIA LONGISPINA IN AN ACIDIC LAKE 355
and the reduced energy requirements associated with lower fecundity. A negativecorrelation between feeding rates and the density of a Daphnia population has beenobserved by many authors (e.g. Peters and Downing, 1984; Helgen, 1987), as hasa variation in feeding rates with the physiological condition of the animals (e.g.Kankaala, 1988; Lair, 1991)
The grazing pressure exerted by the Daphnia population on algae was highand proportional to Daphnia abundance; in contrast, the pressure exerted on bac-teria mostly reflected individual requirements and reached its maximum when thefast-growing population experienced the most severe phytoplankton shortage (11August).
According to previous studies, the structure of the whole microbial loop seemsto be controlled by Daphnia during summer (Callieri et al., 1999). The results ofour study suggest that there is a definite reciprocal control between phytoplanktonproduction and Daphnia grazing activity. It is certainly likely that, in the absenceof predators, Daphnia population growth is controlled only by food availability andthe climatic cycle. Bacteria provide a supplementary food item, exploited largelywhen the supply of phytoplankton declines.
According to Cammarano and Manca (1997), the ability of Daphnia not onlyto survive, but to reach a relatively high population density in LPS is surprising, aspH, Chl a and total phosphorus mean concentrations suggest that the lake shouldbe assigned to category 2 in the classification of Vihnalek et al. (1994). Lakes inthis category are in fact characterized by the disappearance of zooplankton, whichhas been attributed to starvation induced by a pH-driven ultraoligotrophication.To explain these apparently contradictory results, Cammarano and Manca (1997)hypothesized that LPS is not a truly oligotrophic lake (Guilizzoni et al., 1996),and suggested that the wide spatial and temporal variability of the standing stockbiomass of both zooplankton and phytoplankton should be investigated to givemore reliable estimates of the lake trophy and the trophic interactions among thedifferent components of the food web. The results of our study highlighted theability of Daphnia to feed on algae at a very high rate, suggesting the possibility ofa very efficient adaptation to low-food availability. If this hypothesis is confirmedby further investigations, it could offer a contribution to our understanding of thesurvival of Daphnia in this lake.
Acknowledgements
I am grateful to Andrea Mussi, Gianluca Corno, Stefania Martinella, Roberta Pisciaand Mario Contesini for help in the field and in the laboratory. Special thanks toCristiana Callieri and Roberto Bertoni, who gave me the opportunity to carry outthis research, for their help with the experimental work, for their critical commentson the manuscript, and for making their unpublished data available. I am alsograteful to Jaroslav Vrba and Miroslav Macek for providing cultured bacteria and

356 N. RICCARDI
for suggestions about the labeling technique, to Oscar Ravera for comments on anearlier draft of the manuscript and to Sandra Spence for revising the English. I amvery indebted to two anonymous referees who contributed to the amelioration ofthe manuscript with their constructive criticism.
References
Bertoni, R., Callieri, C. and Corno, G.: 2001, ‘The mixotrophic flagellates as key organisms fromDOC to Daphnia in a oligotrophic alpine lake’, Verh. Int. Verein. Limnol. 28, (in press).
Børsheim, K. Y. and Olsen, Y.: 1984, ‘Grazing activities by Daphnia pulex on natural populations ofbacteria and algae’, Verh. Int. Verein. Limnol. 22, 644–648.
Callieri, C. and Bertoni, R.: 1999, ‘Organic Carbon and Microbial Food Web Assemblages in anOligotrophic Alpine Lake’, in V. Stras̆krabová, C. Callieri and J. Fott (eds), Pelagic Food Web inMountain Lakes, MOuntain LAkes Research Program, J. Limnol. 58, pp. 136–143.
Callieri, C., Pugnetti, A. and Manca, M.: 1999, ‘Carbon Partitioning in the Food Web of a HighMountain Lake: From Bacteria to Zooplankton’, in V. Stras̆krabová, C. Callieri and J. Fott(eds), Pelagic Food Web in Mountain Lakes, MOuntain LAkes Research Program, J. Limnol.58, pp. 144–151.
Cammarano, P. and Manca, M.: 1997, ‘Studies on zooplankton in two acidified high mountain lakesin the Alps’, Hydrobiologia 356, 97–109.
Cauchie, H. M., Deguée, N., Joaquim-Justo, C., Thomé, J. P. and Thys, I.: 2000, ‘A note on the use offluorescently labelled algae for the determination of gut passage time in Bosmina and Daphnia’,Verh. Int. Verein. Limnol. 27, (in press).
Corno, G.: 2000, ‘Ruolo dei flagellati mixotrofi nel microbial loop di un lago alpino oligotrofo (LagoPaione Superiore, Alpi Centrali)’, Thesis, University of Milan, 150 pp.
Duncan, A., Lampert, W. and Rocha, O.: 1985, ‘Carbon weight on lenght regressions of Daphnia sp.grown at threshold food concentrations’, Verh. Int. Verein. Limnol. 22, 3109–3115.
Geller, W.: 1975, ‘Die Nahrungsaufnahme von Daphnia pulex in Abhänginkeit von der Futterkonzen-tration, der Temperature, der Korpergrösse und dem Hungerzustand der Tiere’, Arch. Hydrobiol.Suppl. 48, 47–107.
Gliwicz, Z. M.: 1977, ‘Food size selection and seasonal succession of filter feeding zooplankton inan eutrophic lake’, Ekol. Pol. 25, 179–225.
Gliwicz, Z. M.: 1990, ‘Food thresholds and body size in cladocerans’, Nature 343, 638–640.Gophen, M. and Geller, W.: 1984, ‘Filter mesh size and food particle uptake by Daphnia’, Oecologia
64, 408–412.Guilizzoni, P., Marchetto, A., Lami, A., Cameron, N. G., Appleby, P. G., Rose, N. L., Schnell, Ø.
A., Belis, C. A., Giorgis, A. and Guzzi, L.: 1996, ‘The environmental history of a mountainlake (Lago Paione Superiore, Central Alps, Italy) for the last ca. 100 years: A multidisciplinarypaleolimnological study’, J. Paleolimnol. 15, 245–264.
Gulati, R. D. and DeMott, W. R.: 1997, ‘The role of food quality for zooplankton: Remarks on thestate-of-the-art, perspectives and priorities’, Freshw. Biol. 38, 753–768.
Haney, J. F.: 1973, ‘An in situ examination of the grazing activities of natural zooplanktoncommunities’, Arch. Hydrobiol. 72, 87–132.
Haney, J. F.: 1985, ‘Regulation of cladoceran filtering rates in nature by body size, food concentra-tion, and diel feeding patterns’, Limnol. Oceanogr. 30, 397–411.
Helgen, J. C.: 1987, ‘Feeding rate inhibition in crowded Daphnia pulex’, Hydrobiologia 154, 113–119.
Hessen, D. O. and Andersen, T.: 1990, ‘Bacteria as a source of phosphorus for zooplankton’,Hydrobiologia 206, 217–223.

DAPHNIA LONGISPINA IN AN ACIDIC LAKE 357
Holm, N. P., Ganf, G. G. and Shapiro, J.: 1983, ‘Feeding and assimilation rate of Daphnia pulex fedAphanizomenon flos-aquae’, Limnol. Oceanogr. 28, 677–687.
Jürgens, K.: 1994, ‘The impact of Daphnia on microbial food webs – A review’, Mar. Microb. FoodWebs 8, 295–324.
Jürgens, K., Gasol, J. M., Massana, R. and Pedros-Alió, C.: 1994, ‘Control of heterotrophic bacteriaand protozoans by Daphnia pulex in the epilimnion of Lake Cisó’, Arch. Hydrobiol. 131, 55–78.
Kankaala, P.: 1988, ‘The relative importance of algae and bacteria as food for Daphnia longispina(Cladocera) in a polyhumic lake’, Freshw. Biol. 19, 285–196.
Knisely, K. and Geller, W.: 1986, ‘Selective feeding of four zooplankton species on natural lakephytoplankton’, Oecologia 69, 86–94.
Lair, N.: 1991, ‘Grazing and assimilation rates of natural populations of planktonic cladocerans in aeutrophic lake’, Hydrobiologia 215, 51–61.
Lampert, W.: 1987, ‘Feeding and Nutrition in Daphnia’, in R. H. Peters and R. de Bernardi (eds),Daphnia, Mem. Ist. Ital. Idrobiol., Vol. 45, pp. 143–192.
Lampert, W.: 1994, ‘Phenotypic plasticity of the filter screens in Daphnia: Adaptation to a low-foodenvironment’, Limnol. Oceanogr. 39, 997–1006.
Manca, M. and Comoli, P.: 1999, ‘Studies on Zooplankton of Lago Paione Superiore’, in V.Stras̆krabová, C. Callieri and J. Fott (eds), Pelagic Food Web in Mountain Lakes, MOuntainLAkes Research Program. J. Limnol. 58, pp. 131–135.
McCauley, E.: 1984, ‘The Estimation of the Abundance and Biomass of Zooplankton in Samples’,in J. A. Downing and F. H. Rigler (eds), A Manual on Methods for Assessment of SecondaryProductivity in Freshwaters, Blackwell Scientific Publications, pp. 228–265.
Peters, R. H. and Downing, J. A.: 1984, ‘Empirical analysis of zooplankton filtering and feedingrates’, Limnol. Oceanogr. 29, 763–784.
Porter, K. G.: 1984, ‘Natural Bacteria as Food Resources for Zooplankton’, in M. J. Klug and C.A. Reddy (eds), Current Perspectives in Microbial Ecology, American Society for Microbiology,pp. 340–354.
Porter, K. G., Feig, Y. S. and Vetter, E. F.: 1983, ‘Morphology, flow regimes, and filtering rates ofDaphnia, Ceriodaphnia, and Bosmina fed natural bacteria’, Oecologia 58, 153–163.
Pugnetti, A. and Bettinetti, R.: 1999, ‘Biomass and Species Structure of the Phytoplankton of anHigh Mountain Lake (Lake Paione Superiore, Central Alps, Italy)’, in V. Stras̆krabová, C. Callieriand J. Fott (eds), Pelagic Food Web in Mountain Lakes, MOuntain LAkes Research Program. J.Limnol. 58, pp. 127–130.
Ravera, O. and Scotto, S.: 1999, ‘A sampler-incubator for studying zooplankton grazing andphytoplankton production’, J. Limnol. 58, 49–57.
Sherr, B. F., Sherr, E. B. and Fallon, R. D.: 1987, ‘Use of monodispersed, fluorescently labelledbacteria to estimate in situ protozoan bacterivory’, Appl. Environ. Microbiol. 53, 958–965.
The MOLAR Water Chemistry Group: 1999, ‘The MOLAR Project: Atmospheric Deposition andLake Water Chemistry’, in V. Stras̆krabová, C. Callieri and J. Fott (eds), Pelagic Food Web inMountain Lakes, MOuntain LAkes Research Program. J. Limnol. 58, pp. 88–106.
Tonolli, V.: 1949, ‘Gli alti laghi della Val Bognanco. Part II’, Mem. Ist. Ital. Idrobiol. 5, 39–93.Vadstein, O., Jensen, A., Olsen, Y. and Reinertsen, H.: 1988, ‘Growth and phosphorus status of
limnetic phytoplankton and bacteria’, Limnol. Oceanogr. 33, 489–503.Vihnálek, V., Fott, J. and Kopácek, J.: 1994, ‘Chlorophyll-phosphorus relationship in acidified lakes
of the High Tatra Mountains (Slovakia)’, Hydrobiologia 274, 171–177.Wiedner, C. and Vareschi, E.: 1995, ‘Evaluation of a fluorescent microparticle technique for
measuring filtering rates of Daphnia’, Hydrobiologia 302, 89–96.
Copyright © 2022 FDOKUMEN








![The critical importance of defined media conditions in Daphnia magna nanotoxicity studies [Open Access]](https://static.fdokumen.com/doc/165x107/6313f21dc32ab5e46f0ca6d1/the-critical-importance-of-defined-media-conditions-in-daphnia-magna-nanotoxicity.jpg)











