
Ribosome Occupancy of the Yeast CPA1 Upstream Open Reading Frame Termination Codon Modulates...
12
Molecular Cell, Vol. 20, 449–460, November 11, 2005, Copyright ª2005 by Elsevier Inc. DOI 10.1016/j.molcel.2005.09.019 Ribosome Occupancy of the Yeast CPA1 Upstream Open Reading Frame Termination Codon Modulates Nonsense-Mediated mRNA Decay Anthony Gaba, 1,4 Allan Jacobson, 2 and Matthew S. Sachs 1,3, * 1 Department of Environmental and Biomolecular Systems Oregon Health and Science University Beaverton, Oregon 97006 2 Department of Molecular Genetics and Microbiology University of Massachusetts Medical School Worcester, Massachusetts 01655 3 Department of Molecular Microbiology and Immunology Oregon Health and Science University Portland, Oregon 97201 Summary Saccharomyces cerevisiae CPA1 mRNA contains an upstream open reading frame (uORF) encoding the ar- ginine attenuator peptide (AAP). Negative transla- tional regulation of CPA1 occurs when the nascent AAP responds to arginine (Arg) by stalling ribosomes at the uORF termination codon. CPA1 expression is also controlled by nonsense-mediated mRNA decay (NMD). Using wild-type and decay-defective strains ex- pressing CPA1-LUC, we determined how this uORF contributes to NMD control. Arg addition to media rap- idly destabilized the CPA1 transcript in wild-type but not upf1D cells. The wild-type uORF exerted transla- tional control and induced NMD of CPA1-LUC; the mutated D13N uORF, which eliminates stalling and reg- ulation, did not. Thus, regulation by NMD was not gov- erned simply by ribosomes encountering the uORF terminator but appeared dependent on the AAP’s ribosome-stalling ability. Improving the D13N uORF initiation context also promoted NMD. Hence, NMD appears to be triggered by increased ribosomal occu- pancy of the uORF termination codon. Introduction Saccharomyces cerevisiae CPA1 encodes the small subunit of arginine-specific carbamoyl phosphate syn- thetase. Genetic analyses showed that CPA1 was neg- atively regulated when growth media contained arginine (Arg) (Thuriaux et al., 1972). Linked cis-domi- nant mutations causing constitutive expression were proposed to be at an operator site (cpa1O) and an un- linked recessive trans-acting mutation to affect a repres- sor (cpaR). Sequencing of cpa1O mutations showed that they did not affect an operator but could alter the translation of a predicted upstream open reading frame (uORF) in the mRNA 5 0 -leader; measurement of enzyme produced by CPA1-lacZ reporters was consistent with the idea that the uORF regulated expression (Werner et al., 1987). A similar uORF is found in Neurospora crassa arg-2, the homolog of CPA1 (Orbach et al., 1990). Pulse-chase experiments, analyses of the distribution of mRNA on polysomes, and characterization of an uORF missense mutation showed that arg-2 is translationally controlled by Arg through its uORF (Freitag et al., 1996; Luo et al., 1995). Cell-free translation experiments showed that the uORF-encoded peptide, named the arginine attenuator peptide (AAP), translationally regulates expression (Gaba et al. [2001] and references therein). The AAP causes Arg-specific stalling of ribosomes at the uORF termination codon, thereby reducing ribosome access to the downstream open reading frame, and the mech- anism by which the AAP affects ribosomes is evolution- arily conserved (Fang et al., 2004). Studies in vivo have demonstrated CPA1 mRNA sen- sitivity to the nonsense-mediated mRNA decay (NMD) pathway (He et al., 2003; Ruiz-Echevarria and Peltz, 2000). In yeast, NMD targets prematurely terminated mRNAs and is dependent on the regulatory factors Upf1p, Nmd2p (Upf2p), and Upf3p (Jacobson and Peltz, 2000). CPA1 mRNA has half-lives of 3 min in wild-type cells and w18 min in an isogenic upf1D strain (Ruiz- Echevarria and Peltz, 2000), providing direct evidence that this mRNA is an NMD substrate. Consistent with NMD control, the cpaR mutation is a UPF1 allele (Mes- senguy et al., 2002). The function of the CPA1 uORF in controlling NMD, and the role of NMD in Arg-regulation, remains unclear. Studies showing CPA1 mRNA sensitivity to NMD did not analyze the role of the uORF or Arg (He et al., 2003; Ruiz- Echevarria and Peltz, 2000). CPA1 may be subject to Arg-specific translational regulation and Arg-indepen- dent NMD control (Messenguy and Dubois, 2000). While it was reported that growth in Arg does not affect the overall level of CPA1 mRNA (Werner et al., 1987), there are indications that Arg reduces the level of a long form of the transcript and that NMD is required for this (Delbecq et al., 2000; Messenguy et al., 2002). Another study that showed that CPA1 mRNA half-life is affected by Arg did not address the roles of NMD and the uORF (Crabeel et al., 1990). Here we show that the CPA1 uORF has a critical role in modulating NMD and derive a model in which the nascent uORF-encoded peptide acts in concert with Arg to reduce CPA1 expression by regulat- ing translation and mRNA susceptibility to NMD. Results The CPA1 uORF Mediates Arg-Specific Regulation of an In Vivo Reporter In cell-free systems, CPA1 uORF-mediated Arg-specific negative regulation of translation requires the nascent uORF-encoded AAP to stall ribosomes at the uORF ter- mination codon in response to Arg (Wang et al., 1999). An AAP missense mutation (D13N) that eliminates stall- ing eliminates regulation of a luciferase (LUC) reporter initiating from a downstream start codon (Wang et al., 1999). To investigate the uORF’s in vivo effects on mRNA stability, CPA1-LUC chimeric reporter transcripts *Correspondence: [email protected] 4 Present address: Department of Pharmacology, University of California, San Diego, La Jolla, California 92093.
Transcript of Ribosome Occupancy of the Yeast CPA1 Upstream Open Reading Frame Termination Codon Modulates...

Molecular Cell, Vol. 20, 449–460, November 11, 2005, Copyright ª2005 by Elsevier Inc. DOI 10.1016/j.molcel.2005.09.019
Ribosome Occupancy of the Yeast CPA1 UpstreamOpen Reading Frame Termination CodonModulates Nonsense-Mediated mRNA Decay
Anthony Gaba,1,4 Allan Jacobson,2
and Matthew S. Sachs1,3,*1Department of Environmental and Biomolecular
SystemsOregon Health and Science UniversityBeaverton, Oregon 970062Department of Molecular Genetics and MicrobiologyUniversity of Massachusetts Medical SchoolWorcester, Massachusetts 016553Department of Molecular Microbiology
and ImmunologyOregon Health and Science UniversityPortland, Oregon 97201
Summary
Saccharomyces cerevisiae CPA1 mRNA contains anupstream open reading frame (uORF) encoding the ar-
ginine attenuator peptide (AAP). Negative transla-tional regulation of CPA1 occurs when the nascent
AAP responds to arginine (Arg) by stalling ribosomesat the uORF termination codon. CPA1 expression is
also controlled by nonsense-mediated mRNA decay(NMD). Using wild-type and decay-defective strains ex-
pressing CPA1-LUC, we determined how this uORFcontributes to NMD control. Arg addition to media rap-
idly destabilized the CPA1 transcript in wild-type butnot upf1D cells. The wild-type uORF exerted transla-
tional control and induced NMD of CPA1-LUC; themutated D13N uORF, which eliminates stalling and reg-
ulation, did not. Thus, regulation by NMD was not gov-erned simply by ribosomes encountering the uORF
terminator but appeared dependent on the AAP’sribosome-stalling ability. Improving the D13N uORF
initiation context also promoted NMD. Hence, NMDappears to be triggered by increased ribosomal occu-
pancy of the uORF termination codon.
Introduction
Saccharomyces cerevisiae CPA1 encodes the smallsubunit of arginine-specific carbamoyl phosphate syn-thetase. Genetic analyses showed that CPA1 was neg-atively regulated when growth media containedarginine (Arg) (Thuriaux et al., 1972). Linked cis-domi-nant mutations causing constitutive expression wereproposed to be at an operator site (cpa1O) and an un-linked recessive trans-acting mutation to affect a repres-sor (cpaR). Sequencing of cpa1O mutations showedthat they did not affect an operator but could alter thetranslation of a predicted upstream open reading frame(uORF) in the mRNA 50-leader; measurement of enzymeproduced by CPA1-lacZ reporters was consistent withthe idea that the uORF regulated expression (Werneret al., 1987).
*Correspondence: [email protected] address: Department of Pharmacology, University of
California, San Diego, La Jolla, California 92093.
A similar uORF is found in Neurospora crassa arg-2,the homolog of CPA1 (Orbach et al., 1990). Pulse-chaseexperiments, analyses of the distribution of mRNA onpolysomes, and characterization of an uORF missensemutation showed that arg-2 is translationally controlledby Arg through its uORF (Freitag et al., 1996; Luo et al.,1995). Cell-free translation experiments showed that theuORF-encoded peptide, named the arginine attenuatorpeptide (AAP), translationally regulates expression(Gaba et al. [2001] and references therein). The AAPcauses Arg-specific stalling of ribosomes at the uORFtermination codon, thereby reducing ribosome accessto the downstream open reading frame, and the mech-anism by which the AAP affects ribosomes is evolution-arily conserved (Fang et al., 2004).
Studies in vivo have demonstrated CPA1 mRNA sen-sitivity to the nonsense-mediated mRNA decay (NMD)pathway (He et al., 2003; Ruiz-Echevarria and Peltz,2000). In yeast, NMD targets prematurely terminatedmRNAs and is dependent on the regulatory factorsUpf1p, Nmd2p (Upf2p), and Upf3p (Jacobson and Peltz,2000). CPA1 mRNA has half-lives of 3 min in wild-typecells and w18 min in an isogenic upf1D strain (Ruiz-Echevarria and Peltz, 2000), providing direct evidencethat this mRNA is an NMD substrate. Consistent withNMD control, the cpaR mutation is a UPF1 allele (Mes-senguy et al., 2002).
The function of the CPA1 uORF in controlling NMD,and the role of NMD in Arg-regulation, remains unclear.Studies showing CPA1 mRNA sensitivity to NMD did notanalyze the role of the uORF or Arg (He et al., 2003; Ruiz-Echevarria and Peltz, 2000). CPA1 may be subject toArg-specific translational regulation and Arg-indepen-dent NMD control (Messenguy and Dubois, 2000). Whileit was reported that growth in Arg does not affect theoverall level of CPA1 mRNA (Werner et al., 1987), thereare indications that Arg reduces the level of a longform of the transcript and that NMD is required for this(Delbecq et al., 2000; Messenguy et al., 2002). Anotherstudy that showed that CPA1 mRNA half-life is affectedby Arg did not address the roles of NMD and the uORF(Crabeel et al., 1990). Here we show that the CPA1 uORFhas a critical role in modulating NMD and derive a modelin which the nascent uORF-encoded peptide acts inconcert with Arg to reduce CPA1 expression by regulat-ing translation and mRNA susceptibility to NMD.
Results
The CPA1 uORF Mediates Arg-Specific Regulation
of an In Vivo ReporterIn cell-free systems, CPA1 uORF-mediated Arg-specificnegative regulation of translation requires the nascentuORF-encoded AAP to stall ribosomes at the uORF ter-mination codon in response to Arg (Wang et al., 1999).An AAP missense mutation (D13N) that eliminates stall-ing eliminates regulation of a luciferase (LUC) reporterinitiating from a downstream start codon (Wang et al.,1999). To investigate the uORF’s in vivo effects onmRNA stability, CPA1-LUC chimeric reporter transcripts
Molecular Cell450
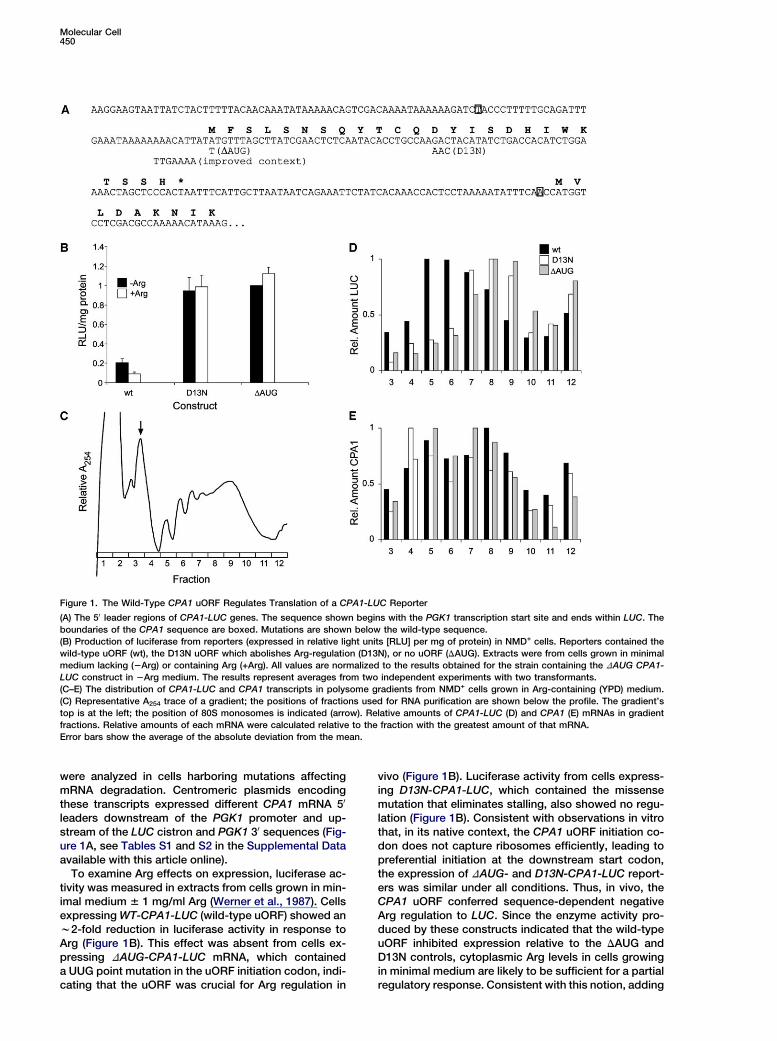
Figure 1. The Wild-Type CPA1 uORF Regulates Translation of a CPA1-LUC Reporter
(A) The 50 leader regions of CPA1-LUC genes. The sequence shown begins with the PGK1 transcription start site and ends within LUC. The
boundaries of the CPA1 sequence are boxed. Mutations are shown below the wild-type sequence.
(B) Production of luciferase from reporters (expressed in relative light units [RLU] per mg of protein) in NMD+ cells. Reporters contained the
wild-type uORF (wt), the D13N uORF which abolishes Arg-regulation (D13N), or no uORF (DAUG). Extracts were from cells grown in minimal
medium lacking (2Arg) or containing Arg (+Arg). All values are normalized to the results obtained for the strain containing the DAUG CPA1-
LUC construct in 2Arg medium. The results represent averages from two independent experiments with two transformants.
(C–E) The distribution of CPA1-LUC and CPA1 transcripts in polysome gradients from NMD+ cells grown in Arg-containing (YPD) medium.
(C) Representative A254 trace of a gradient; the positions of fractions used for RNA purification are shown below the profile. The gradient’s
top is at the left; the position of 80S monosomes is indicated (arrow). Relative amounts of CPA1-LUC (D) and CPA1 (E) mRNAs in gradient
fractions. Relative amounts of each mRNA were calculated relative to the fraction with the greatest amount of that mRNA.
Error bars show the average of the absolute deviation from the mean.
were analyzed in cells harboring mutations affectingmRNA degradation. Centromeric plasmids encodingthese transcripts expressed different CPA1 mRNA 50
leaders downstream of the PGK1 promoter and up-stream of the LUC cistron and PGK1 30 sequences (Fig-ure 1A, see Tables S1 and S2 in the Supplemental Dataavailable with this article online).
To examine Arg effects on expression, luciferase ac-tivity was measured in extracts from cells grown in min-imal medium 6 1 mg/ml Arg (Werner et al., 1987). Cellsexpressing WT-CPA1-LUC (wild-type uORF) showed anw2-fold reduction in luciferase activity in response toArg (Figure 1B). This effect was absent from cells ex-pressing DAUG-CPA1-LUC mRNA, which containeda UUG point mutation in the uORF initiation codon, indi-cating that the uORF was crucial for Arg regulation in
vivo (Figure 1B). Luciferase activity from cells express-ing D13N-CPA1-LUC, which contained the missensemutation that eliminates stalling, also showed no regu-lation (Figure 1B). Consistent with observations in vitrothat, in its native context, the CPA1 uORF initiation co-don does not capture ribosomes efficiently, leading topreferential initiation at the downstream start codon,the expression of DAUG- and D13N-CPA1-LUC report-ers was similar under all conditions. Thus, in vivo, theCPA1 uORF conferred sequence-dependent negativeArg regulation to LUC. Since the enzyme activity pro-duced by these constructs indicated that the wild-typeuORF inhibited expression relative to the DAUG andD13N controls, cytoplasmic Arg levels in cells growingin minimal medium are likely to be sufficient for a partialregulatory response. Consistent with this notion, adding

uORF-Mediated NMD in Yeast451
Figure 2. Effects of the Native-Context CPA1
uORF on Levels of CPA1-LUC Transcripts in
Isogenic NMD+ and nmd2D Strains
(A–C) Cells were grown in YPD and con-
tained CPA1-LUC reporters as described in
Figure 1B. (A) Northern analysis of CPA1-
LUC, CPA1, CYH2, and SCR1 transcripts.
SCR1 was used to normalize lanes for quan-
titative analyses. pCYH2 and mCYH2 denote
CYH2 pre-mRNA and the mature CYH2 tran-
script, respectively. (B and C) The relative
levels of CPA1-LUC (B) and CPA1 (C) tran-
scripts were determined relative to the SCR1
transcript (Experimental Procedures). The
levels of the CPA1-LUC and CPA1 transcripts
in the DAUG CPA1-LUC NMD+ strain were
used to normalize relative transcript levels in
(B) and (C), respectively.
(D–F) Effects of improving the uORF initiation
context on levels of CPA1-LUC transcripts in
NMD+ and nmd2D strains. Cells were grown
in YPD and contained reporters with the
wild-type (wt) or the D13N uORFs in improved
contexts, or no uORF (DAUG). (D) Northern
analysis. Relative levels of CPA1-LUC (E)
and CPA1 (F) transcripts were determined as
described above. Each bar represents the
average of three independent experiments.
Error bars show the average of the absolute
deviation from the mean.
150 mM Arg to extracts promotes a regulatory responsein vitro, and the cytoplasmic Arg concentration is nearthis value (Davis, 1986; Kitamoto et al., 1988). To avoidpotential complications arising from the use of minimalmedium, some of our comparative studies used richmedium (YPD), which contains Arg; the effects of thewild-type uORF appeared maximal in YPD.
In vitro, AAP-mediated ribosome stalling reducesleaky scanning and downstream initiation (Gaba et al.,2001). To test directly whether the WT-CPA1-LUCmRNA was inefficiently translated in vivo, relative to theDAUG- and D13N-CPA1-LUC mRNAs in the presenceof Arg, the polyribosomal distribution of each mRNAwas assessed in cells grown in rich medium. A represen-tative polysome profile and the distribution of the differ-ent LUC transcripts are shown in Figures 1C and 1D. Thenative-context WT-CPA1-LUC mRNA and the endoge-nous CPA1 mRNA (examined in every profile as an inter-nal control) were associated with polysomes that weregenerally smaller than those associated with DAUG-
and D13N-CPA1-LUC transcripts (Figure 1E). Thesedata demonstrate that the latter two reporters weretranslated more efficiently than the wild-type uORF-containing reporter and endogenous CPA1mRNA, indi-cating that the expression differences between thewild-type and D13N constructs are at least partially dueto the wild-type uORF reducing translation downstream.
Translation of the Wild-Type CPA1 uORF InducesNMD of CPA1-LUC Transcripts
Isogenic wild-type and nmd2D cells containing WT-,D13N-, or DAUG-CPA1-LUC were grown in YPD andmRNA steady-state levels were determined. As a control,we examined the level of an endogenous NMD substrate,the CYH2 pre-mRNA (He et al., 1993), and observed thatit was substantially elevated in nmd2D strains (Figure2A), confirming that they were NMD defective. The levelof the mature CYH2 mRNA (not an NMD substrate) wasunaffected in nmd2D cells, but the level of the endoge-nous CPA1 transcript (also examined in every strain)

Molecular Cell452
was elevated approximately 4.5-fold (Figures 2A and2C). These observations confirm earlier studies demon-strating that the CPA1 transcript is controlled by NMD(He et al., 2003; Messenguy et al., 2002; Ruiz-Echevarriaand Peltz, 2000).
The levels of DAUG- and D13N-CPA1-LUC transcriptswere similar in wild-type and nmd2D strains (Figures 2Aand 2B), indicating that these mRNAs were not con-trolled by NMD. However, in the NMD+ strain, WT-CPA1-LUC mRNA was approximately 2-fold reducedcompared to the DAUG- and D13N-CPA1-LUC mRNAs.In the nmd2D strain, the WT-CPA1-LUC mRNA was ap-proximately 3.5-fold more abundant than these others(Figure 2B). Thus, the steady-state abundance of theWT-CPA1-LUC transcript was elevated w7-fold in thenmd2D strain relative to the NMD+ strain. The de-creased level of WT-CPA1-LUC mRNA relative toDAUG- and D13N-CPA1-LUC transcripts in the NMD+
strain, and its increase in the nmd2D strain, indicatedthat the WT-CPA1-LUC transcript, like endogenousCPA1 mRNA, is controlled by NMD.
To verify NMD control of these mRNAs, transcript lev-els were examined in a strain deficient for a secondNMD factor, Upf1p. In both upf1D and nmd2D strains,the steady-state levels of WT-CPA1-LUC and endoge-nous CPA1 mRNAs were similarly elevated relative totheir levels in the NMD+ strain, while the levels ofDAUG- and D13N-CPA1-LUC transcripts were similarin all strains (Figure 3A). The WT-CPA1-LUC and endog-enous CPA1 mRNA levels were thus controlled by NMD.
Arg Induces NMD of CPA1 mRNA
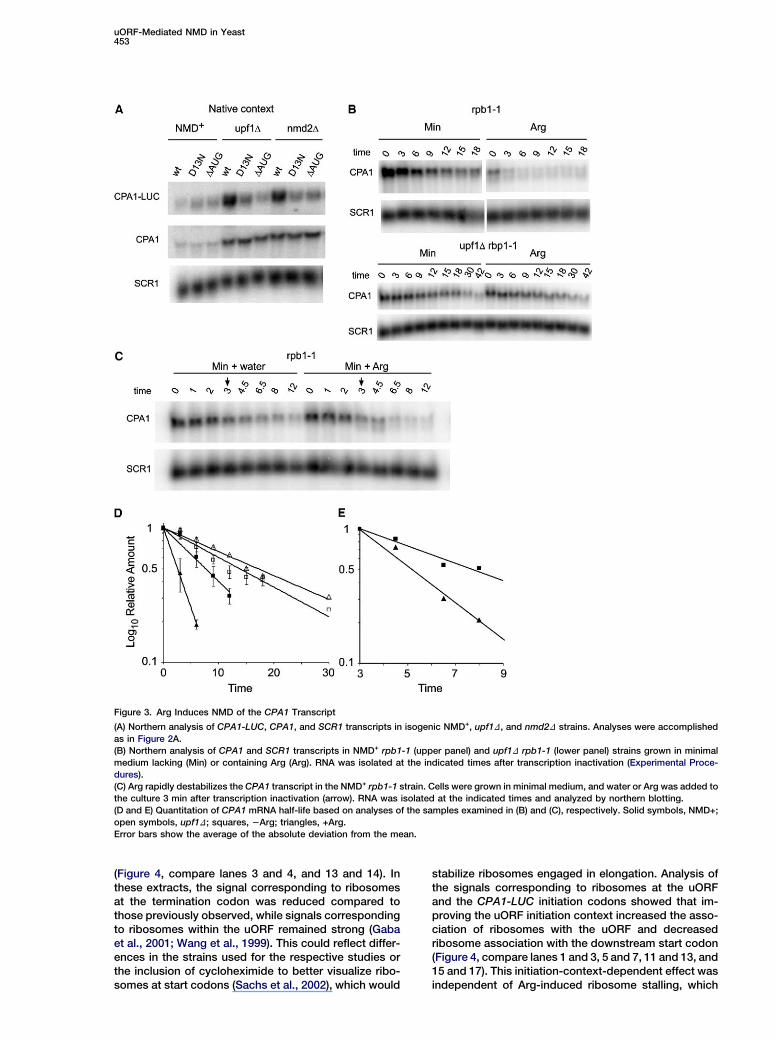
Because the wild-type but not the D13N uORF triggeredNMD (Figures 2A, 2B, and 3A), regulated translation(Figures 1B and 1C), and causes ribosomes to stall atthe uORF termination codon (Gaba et al., 2001; Wanget al., 1999), it appeared that Arg-induced AAP-medi-ated ribosome stalling could be critical for NMD ofCPA1 mRNA. If this were correct, then NMD of CPA1mRNA should be less active in minimal media than inArg-supplemented media. To test this, and to excludepossible secondary effects of disrupting NMD, the sta-bility of the CPA1 transcript was measured in isogenicNMD+ and upf1D strains harboring the rpb1-1 tempera-ture-sensitive allele of RNA polymerase II (Nonet et al.,1987). Cells were grown in minimal medium 6 Arg andCPA1 mRNA decay rates were determined (Figures 3Band 3D). These experiments showed that CPA1 mRNAhalf-lives were w7 and 3 min, respectively, for NMD+
cells grown in media that lacked or contained Arg, while,in upf1D cells, the corresponding half-lives were w14and 17 min, respectively. Published CPA1 decay ratesin YPD medium (Ruiz-Echevarria and Peltz, 2000) weresimilar to those we observed in Arg-containing medium.Consistent with its altered half-life, the level of CPA1mRNA was lower in NMD+ cells grown in Arg beforethe temperature shift (Figure 3B). After inactivation oftranscription, the CPA1 mRNA in NMD+ cells hada more ‘‘doublet’’-like appearance, especially in cellsexposed to Arg. The lower band of this doublet presum-ably corresponds to the smaller transcript postulated tobe an NMD product (Messenguy et al., 2002).
To test directly for an Arg effect on CPA1 mRNA stabil-ity, transcription was inhibited in NMD+ rpb1-1 cells
grown in medium lacking Arg; at 3 min after transcriptioninhibition, the culture was divided and supplementedwith either water or Arg, and samples were taken fornorthern analyses (Figure 3C). The CPA1 mRNA decayrate was not significantly altered by the addition ofwater, but addition of Arg enhanced CPA1 mRNA decaysubstantially (Figures 3C and 3E; data not shown). Thelatter effect was immediate, and the CPA1 mRNA ap-peared as a doublet after Arg addition. Arg did not affectthe stability of the CYH2 mRNA or pre-mRNA (data notshown). These results indicate that Arg rapidly and spe-cifically affects CPA1 mRNA sensitivity to NMD.
Modulation of Ribosome Levels at the CPA1 uORFand Downstream ORF Regulates NMD of CPA1-LUC
TranscriptsThe analyses of Figures 1–3 indicated that AAP-medi-ated ribosome stalling is critical for NMD of WT-CPA1-LUC mRNA. To further assess the role of the uORF,we tested whether modulating ribosome levels onCPA1-LUC mRNAs by means other than AAP-mediatedstalling could also promote NMD. To accomplish this,we improved the uORF initiation context. For theD13N uORF, this change should promote ribosome oc-cupancy to a greater extent than the native context, al-though the intrinsic efficiency of termination shouldremain unchanged. Isogenic wild-type and nmd2D
strains were transformed with plasmids encoding WT-and D13N-CPA1-LUC transcripts that contained uORFsin a more efficient GCN4 uORF1-like initiation context(Figure 1A and Table S1) (Gaba et al., 2001). In NMD+
cells, the levels of both improved-context WT- andD13N-CPA1-LUC transcripts were reduced, approxi-mately 2- and 5-fold, respectively, relative to that ofthe DAUG-CPA1-LUC transcript (Figures 2D and 2E).In the nmd2D strain, the abundance of the improved-context WT- and D13N-CPA1-LUC transcripts waselevated, w6- and 5-fold, respectively, relative to DAUG-CPA1-LUC (Figure 2E). These data show that improved-context WT- and D13N-CPA1-LUC transcripts wereboth NMD substrates, in contrast to what occurs withthe native uORF AUG context, in which the WT- butnot the D13N-CPA1-LUC transcript was a substratefor NMD (Figures 2A, 2B, 2D, and 2E). The response ofthe endogenous CPA1 transcript to a functional NMDpathway was similar in each strain examined, as ex-pected (Figure 2F). These analyses identify two modesfor the induction of NMD by the CPA1 uORF. With a na-tive CPA1 uORF initiation context, ribosome stalling atthe uORF termination codon is required for the uORFto induce NMD of CPA1-LUC mRNA, but when the initi-ation context is improved, ribosome stalling is no longerrequired for the uORF to induce NMD.
Each of these two NMD-inducing modes should in-crease the level of ribosomes translating the uORFand reduce the level initiating translation downstream.The polysomal distribution of reporter transcripts wasconsistent with this (Figure 1D and data not shown).As a further test, we performed toeprinting assays ofthe distribution of ribosomes on CPA1-LUC transcriptsin extracts from wild-type and nmd2D cells. In both ex-tracts, Arg induced ribosome stalling at the wild-typeuORF on native-context (Figure 4, compare lanes 1and 2, and 11 and 12) and improved-context transcripts
uORF-Mediated NMD in Yeast453
Figure 3. Arg Induces NMD of the CPA1 Transcript
(A) Northern analysis of CPA1-LUC, CPA1, and SCR1 transcripts in isogenic NMD+, upf1D, and nmd2D strains. Analyses were accomplished
as in Figure 2A.
(B) Northern analysis of CPA1 and SCR1 transcripts in NMD+ rpb1-1 (upper panel) and upf1D rpb1-1 (lower panel) strains grown in minimal
medium lacking (Min) or containing Arg (Arg). RNA was isolated at the indicated times after transcription inactivation (Experimental Proce-
dures).
(C) Arg rapidly destabilizes the CPA1 transcript in the NMD+ rpb1-1 strain. Cells were grown in minimal medium, and water or Arg was added to
the culture 3 min after transcription inactivation (arrow). RNA was isolated at the indicated times and analyzed by northern blotting.
(D and E) Quantitation of CPA1 mRNA half-life based on analyses of the samples examined in (B) and (C), respectively. Solid symbols, NMD+;
open symbols, upf1D; squares, 2Arg; triangles, +Arg.
Error bars show the average of the absolute deviation from the mean.
(Figure 4, compare lanes 3 and 4, and 13 and 14). Inthese extracts, the signal corresponding to ribosomesat the termination codon was reduced compared tothose previously observed, while signals correspondingto ribosomes within the uORF remained strong (Gabaet al., 2001; Wang et al., 1999). This could reflect differ-ences in the strains used for the respective studies orthe inclusion of cycloheximide to better visualize ribo-somes at start codons (Sachs et al., 2002), which would
stabilize ribosomes engaged in elongation. Analysis ofthe signals corresponding to ribosomes at the uORFand the CPA1-LUC initiation codons showed that im-proving the uORF initiation context increased the asso-ciation of ribosomes with the uORF and decreasedribosome association with the downstream start codon(Figure 4, compare lanes 1 and 3, 5 and 7, 11 and 13, and15 and 17). This initiation-context-dependent effect wasindependent of Arg-induced ribosome stalling, which

Molecular Cell454
Figure 4. Effects of Arg and uORF Initiation Context on Ribosomes at CPA1-LUC ORFs
Translation extracts from NMD+ and nmd2D strains were supplemented with 10 mM Arg (2) or 2000 mM Arg (+) and were programmed with 120
ng of CPA1-LUC transcripts containing the wild-type uORF (wt), the D13N uORF (D13N) in either native or improved (im) initiation contexts, or
no uORF (DAUG). After translation at 25ºC for 10 min, reactions were supplemented with cycloheximide and incubated for an additional 5 min.
Radiolabeled primer ZW4 was used for primer extension analysis and for sequencing the native-context wild-type CPA1-LUC template (the
three left lanes). The nucleotide complementary to the dideoxynucleotide added is indicated below each sequencing lane; the 50 to 30 se-
quence reads from top to bottom (the C0 lane is not shown). The positions of cDNA products corresponding to the mRNA 50 end and
uORF start, uORF termination, and CPA1-LUC start codons are indicated on the right; the uORF termination codon position is boxed in
the sequencing reactions and marked by asterisks in the toeprints.
also reduced levels of ribosomes detected at the down-stream start codon. These data indicate that the twoeffects of uORF sequences that induced NMD in vivo,ribosome stalling at the uORF termination codon andimprovement of the uORF initiation context, caused thesteady-state distribution of ribosomes on CPA1-LUCtranscripts to shift from the LUC ORF to the uORF, andin each case increased the occupancy of the uORF termi-nation codon by ribosomes.
Inactivation of Xrn1p, but Not Ski2p, AltersSteady-State Levels of CPA1 mRNAs
To further characterize CPA1 uORF-mediated modula-tion of mRNA abundance, we determined the effectsof eliminating either Xrn1p, an exoribonuclease requiredfor 50 to 30 mRNA decay (Hsu and Stevens, 1993), orSki2p, a putative RNA helicase, required for 30 to 50 exo-some-mediated mRNA decay (Anderson and Parker,1998). Relative to the isogenic wild-type strain, CPA1and CPA1-LUC transcript levels were w6-fold higherin the xrn1D strain and only slightly elevated (1.2-fold)in the ski2D strain (Figure 5). Consistent with earlier
microarray analyses (He et al., 2003), these results indi-cate that the 50 to 30 decay pathway is the primary reg-ulator of CPA1 and CPA1-LUC transcript levels.Similarly, since the steady-state abundance of the WT-CPA1-LUC transcript was reduced approximately 2-fold relative to the DAUG- and D13N-CPA1-LUC tran-scripts in the wild-type and ski2D strains but not in thexrn1D strain (Figure 5B), uORF-mediated NMD of theWT-CPA1-LUC transcript appeared largely dependenton Xrn1p but not Ski2p.
CPA1 uORF-Mediated Translational Control
Functions Independently of NMDTo investigate the role of the CPA1 uORF on gene ex-pression at the level of translation, we measured lucifer-ase activity and controlled for differences in mRNAlevels by normalizing activity to the level of CPA1-LUCtranscript (expressed as LUC/RNA). NMD+ and nmd2D
strains expressing the DAUG- and native-contextD13N-CPA1-LUC transcripts showed similar LUC/RNAvalues (Figure 6A), indicating that the translation effi-ciency of these reporters, which were not NMD

uORF-Mediated NMD in Yeast455
Figure 5. Relative CPA1-LUC and CPA1 Transcript Levels in Isogenic XRN1 SKI2, xrn1D, and ski2D Strains Expressing Different Native-
Context CPA1-LUC Reporters
Reporters were as described in Figure 1B. (A) Representative northern blots. Strains, reporter constructs, and blotting procedures were as
described for Figure 2. (B and C) The amounts of CPA1-LUC (B) and CPA1 (C) transcripts were determined relative to SCR1 as described
in Figure 2. For the XRN1 SKI2 (wild-type) strain, each bar represents the average of three experiments with one transformant, and two
with another. For xrn1D strains, each bar represents the average of two experiments with one transformant, and one with another. For
ski2D strains, each bar represents the average of two experiments with one transformant, and two with another. All RNA samples were ana-
lyzed in duplicate using two separate blots. Error bars show the average of the absolute deviation from the mean.
substrates, were similar in both strains. In the NMD+
strain, however, native-context WT-CPA1-LUC mRNAyielded LUC/RNA levels that were w4-fold less thanthose obtained from the DAUG- and D13N-CPA1-LUCmRNAs. In the nmd2D strain, the native-context WT-CPA1-LUC mRNA resulted in LUC/RNA levels thatwere w15-fold less than those obtained from DAUG-and D13N-CPA1-LUC mRNAs. Since all mRNAs differedonly by changes in the uORF, these effects cannot beattributed to posttranslational events and most likelyrepresent effects of uORF-mediated translational
control. In the nmd2D strain, the reduced LUC/RNA lev-els observed from native-context WT- but not D13N-CPA1-LUC mRNA relative to those from DAUG-CPA1-LUC mRNA (Figure 6A) indicated that uORF-mediatedtranslational control is separable from a functionalNMD pathway.
When uORF initiation contexts were improved, LUC/RNA levels indicated that the wild-type and D13N uORFswere significantly more inhibitory to LUC synthesis thanwere these uORFs in their native contexts (compare Fig-ures 6A and 6B). When the wild-type uORF was in its
Figure 6. Luciferase Production Normalized
to mRNA Levels
Luciferase activity was measured in extracts
prepared from the cells analyzed in Figures 2
and 5. Levels of luciferase activity were nor-
malized in each experiment relative to that of
the DAUG-CPA1-LUC reporter in the NMD+
strain, as were the levels of RNA. The data
represent the relative level of enzyme activ-
ity/mg protein/RNA (LUC/RNA). (A and B)
CPA1-LUC reporters containing the wild-
type uORF (wt) and the D13N uORF (D13N)
in their native (A) or improved (B) contexts,
or lacking a uORF (DAUG), expressed in
NMD+ and nmd2D strains. The y axis in (B)
is discontinuous because the uORFs in an
improved context strongly inhibited lucifer-
ase expression. (C) Native-context CPA1-
LUC reporters expressed in XRN1 SKI2,
xrn1D, and ski2D strains. Error bars show
the average of the absolute deviation from
the mean.

Molecular Cell456
native or improved context, the LUC/RNA levels in NMD+
cells were 20% and 2%, respectively, of the level ob-served for the DAUG-CPA1-LUC transcript. In NMD+
cells, the D13N uORF in native and improved contextsshowed LUC/RNA levels that were 96% and 12%, re-spectively, of the level from the DAUG-CPA1-LUC. Inthe nmd2D strain, CPA1-LUC mRNA containing thewild-type uORF in native and improved contextsshowed LUC/RNA levels that were 6% and 0.2%, re-spectively, of the level from DAUG-CPA1-LUC, whereasthe D13N-CPA1-LUC mRNA in native and improved con-texts showed LUC/RNA levels that were 80% and 0.6%,respectively, of that fromDAUG-CPA1-LUC.
These results are consistent with a model in which thecontext of the CPA1 uORF initiation codon governs theextent of scanning to the downstream start codon re-gardless of the uORF peptide sequence, and in whichthe wild-type but not the D13N uORF nascent peptideconfers Arg-specific control by blocking scanning ribo-somes in response to Arg. An additional regulatory ef-fect of the CPA1 uORF was also discerned bycomparing mRNA translatability in NMD+ cells to thatin strains defective in NMD. Independent of uORF ef-fects on ribosome stalling, uORF-containing mRNAsthat were normally substrates for NMD but were not de-graded in nmd2D cells were less translatable in the mu-tant than in the NMD+ strain. This phenomenon wasspecific to NMD substrates because it was not detectedwith D13N-CPA1-LUC mRNA harboring the uORF in itsnative context (Figure 6A), nor was it manifested inxrn1D cells (Figure 6C). The reduction in LUC/RNA lev-els observed in xrn1D cells affected all three test tran-scripts uniformly and most likely reflected thedecreased translatability characteristic of the deadeny-lated and decapped transcripts known to accumulate insuch cells (Hsu and Stevens, 1993).
Discussion
Ribosome Stalling at the CPA1 uORF Termination
Codon Controls Gene Expression at Multiple LevelsThe CPA1 uORF-encoded AAP functions in vitro to stallribosomes at the uORF termination codon in responseto Arg, which reduces translation initiation downstream(Gaba et al., 2001; Wang et al., 1999). Although in vivoreporter studies suggested that CPA1 translation isAAP controlled (Werner et al., 1987), biochemical evi-dence for control in vivo is lacking. To this end, we mea-sured the relative levels of the CPA1-LUC andendogenous CPA1 transcripts in polysomes from cellsgrown in Arg-containing medium. Reporter transcriptswith D13N or DAUG uORF mutations fractionated withlarger polysomes than did wild-type uORF-containingreporter and endogenous CPA1 transcripts (Figures1D and 1E), indicating that the wild-type AAP exertstranslational control in vivo, consistent with its abilityto stall ribosomes.
Previous studies indicated that the level and half-lifeof the CPA1 transcript are controlled by NMD (Heet al., 2003; Ruiz-Echevarria and Peltz, 2000), but theuORF role in these effects was unknown. By indepen-dently altering the uORF’s ability to stall ribosomes,the efficiency of uORF translation initiation, and the ac-tivity of specific mRNA decay factors, we elucidated
the coupling of uORF-mediated translational controland mRNA stability. Our results indicate that Arg-in-duced ribosome stalling at the uORF termination codonmodulates NMD. This provides a clear example of multi-faceted control of eukaryotic gene expression in whichthe synthesis of a nascent peptide has strong regulatoryeffects at the levels of both translation and mRNA decay.
Regulated Decay of a Conserved Class of NMD
SubstrateConsidering that, in their native initiation contexts, thewild-type but not the D13N uORF induces NMD (Figures2 and 3A) and that only the wild-type AAP stalls ribo-somes at the uORF termination codon in response toArg (Figure 4) (Gaba et al., 2001; Wang et al., 1999), ourdata indicate that ribosome stalling at the uORF termina-tion codon induces NMD. Consistent with this, the CPA1mRNA, but not CYH2 pre-mRNA, decays more rapidlyin NMD+ cells grown in medium containing Arg than inmedium lacking Arg (Figures 3B and 3D and data notshown). The induction of NMD by Arg does not appearto require newly synthesized CPA1 mRNA because Argaddition 3 min after transcription was inactivated desta-bilizes CPA1 mRNA rapidly (Figure 3C). These resultsare consistent with the idea that nonsense-containingmRNAs are never immune to NMD in yeast (Maderazoet al., 2003), and contrast with results in mammalian cellssuggesting that NMD is limited to early rounds of trans-lation (Ishigaki et al., 2001). These data also show thatNMD can rapidly regulate preexisting cytoplasmic tran-script levels in response to environmental changes.
Although the native-context D13N uORF and DAUGreporter transcripts were relatively insensitive to NMD(Figures 2 and 3), they never reached levels equivalentto those reached by NMD substrate transcripts inupf1D or nmd2D cells (Figure 2). This seemingly surpris-ing result suggests that, in the absence of NMD, thenative-context D13N uORF and DAUG mRNAs weresubstrates for an NMD-independent decay pathwaythat required translation of the downstream LUC ORF,a phenomenon analogous to that seen for MATa1mRNA (Hennigan and Jacobson, 1996). Consistentwith this interpretation, in upf1D or nmd2D cells, theCPA1-LUC mRNAs that are NMD substrates are trans-lated poorly (Figure 6 and data not shown) and are notshunted into this pathway, and all reporter transcriptsexhibited increased levels in Xrn1p-deficient cells (Fig-ure 5). These results additionally suggest that (1) degra-dation of the CPA1 mRNA largely required the 50 to 30
decay pathway, as observed for other yeast NMD sub-strates (He et al., 2003; Ruiz-Echevarria and Peltz,2000), and (2) the pathway used by the D13N uORFand DAUG reporter transcripts also exploited 50 to 30 de-cay. Our data also show that a shorter CPA1 transcriptappears upon Arg addition (Figures 3B and 3C; Messen-guy et al., 2002). The kinetics of appearance and Arg de-pendence of this transcript (Figure 3C) suggest that itmay be a decay intermediate in which a complex involv-ing stalled ribosomes impedes 50 to 30 decay.
Destabilization of Nonsense-Containing mRNAs
Requires Their TranslationThe presence of a premature nonsense codon withina transcript is not sufficient to trigger its degradation.

uORF-Mediated NMD in Yeast457
Destabilization of such mRNAs also requires their trans-lation, a conclusion that follows from observations thatdecay can be antagonized by drugs, mutations, or RNAstructures that interfere with protein synthesis (Bel-grader et al., 1993; Ruiz-Echevarria et al., 1998; Zhanget al., 1997) or by tRNAs that suppress termination (Bel-grader et al., 1993; Gozalbo and Hohmann, 1990; Los-son and Lacroute, 1979). Our studies provide insightinto how translation of an mRNA is involved in controllingits stability through NMD. The native context wild-typeCPA1 uORF efficiently stalls ribosomes and triggersNMD, while the D13N uORF, which stalls ribosomespoorly or not at all, is an inefficient trigger. Improvingthe uORF initiation context renders both the wild-typeand D13N uORFs efficient triggers of NMD. The NMD-inducing effects of the context-improved D13N uORFwere not due to ribosome stalling because the D13NuORF did not mediate ribosome-stalling activity, re-gardless of its initiation context. These data indicatethat the levels of ribosomes at an early stop codoncan be regulated to control NMD (Figure 7). In turn,this implies that early termination of translation on theCPA1 mRNA and/or the subsequent triggering of NMDmust be inefficient, a conclusion consistent with recentstudies showing that premature termination is consider-ably less efficient than normal termination (Amrani et al.,2004). As suggested by the faux-UTR model (Amrani
Figure 7. Model for the Regulation of CPA1 mRNA Decay by AAP-
Mediated Modulation of Ribosome Occupancy of its uORF Termi-
nator
The wild-type CPA1 uORF (solid gray box) in its native initiation
context stalls ribosomes at the termination codon in response to
Arg, inducing NMD of the transcript. The D13N CPA1 uORF (open
box) in its native initiation context does not stall ribosomes in re-
sponse to Arg, and initiation is inefficient. The occupancy of the ter-
mination site by ribosomes for the D13N uORF is therefore low, and
NMD is not induced. Improving the uORF initiation context (black
arrowhead) increases uORF translation and causes NMD of both
wild-type and D13N CPA1 uORF-containing transcripts because
the occupancy of the termination site by ribosomes is high in
both cases.
et al., 2004; Jacobson and Peltz, 2000), inefficient ter-mination might promote NMD by allowing the ribosometo associate with decay-inducing factors routinely pre-cluded from interaction by the events of normaltermination.
The Caenorhabditis elegans gna-2 mRNA is anotherexample of a transcript in which NMD is regulated bythe efficiency of uORF translation (Lee and Schedl,2004), albeit differently. The RNA binding protein GLD-1 protects gna-2 mRNA from NMD by binding to its 50
leader and reducing translation of two uORFs. Interest-ingly, both GLD-1 binding to gna-2 mRNA and AAP-me-diated ribosome stalling on CPA1 mRNA regulatetranslation yet induce opposing effects on NMD.Whereas GLD-1 binding to gna-2 mRNA reducesNMD, AAP-mediated ribosome stalling on the CPA1mRNA induces NMD. A notable similarity in the regula-tion of NMD by these uORFs is that both involvetrans-acting factors (the GLD-1 protein for gna-2 andthe amino acid Arg for CPA1) that act at the level oftranslation to regulate ribosome levels at uORFs.
Although the uORF-containing CPA1 and gna-2mRNAs are NMD substrates, the presence of a uORFdoes not automatically confer such status. Only a por-tion of the cellular uORF-containing mRNAs accumulatein yeast upf1D, nmd2D, or upf3D strains (He et al., 2003;Ruiz-Echevarria and Peltz, 2000), and directed studiesof the uORFs in the YAP1 and GCN4 mRNAs showthat these neither mediate NMD nor destabilize theirmRNAs (Ruiz-Echevarria et al., 1998; Vilela et al.,1998). The inability of the latter uORFs to trigger NMDwas attributed to the presence of flanking stabilizer ele-ments, cis-acting sequences that appeared to counter-act the NMD-inducing effects of early termination. Giventhat premature termination appears to be functionallydistinct from normal termination and that this differenceappears to play a role in promoting NMD (Amrani et al.,2004), such stabilizer elements may affect the qualita-tive nature of the uORF termination event.
Regardless of whether an early termination codon isderived from a uORF, a nonsense mutation, or anothersource, our data indicate that its role as an inducer ofNMD depends on the extent to which it is occupied bythe translation apparatus. A corollary of this conclusionis that events and factors that alter mRNA translatabilityshould also affect NMD. In this regard, NMD in metazo-ans has been thought to be dependent on mRNA asso-ciation with specific components of the exon junctioncomplex (EJC) (Maquat, 2004; Singh and Lykke-Andersen,2003). Since recent studies have shown that these fac-tors can influence the efficiency with which an mRNAis translated (Nott et al., 2004; Wiegand et al., 2003), itis possible that the role of EJC proteins as NMD regula-tors is, at least in part, related to their ability to ensureefficient mRNA translation.
Decreased Translatability of CPA1 mRNA in nmd2D
Strains: A Consequence of Defective Termination?
Reporter genes containing the wild-type uORF alwaysshowed reduced levels of reporter expression per unitof mRNA when compared to those containing theD13N or DAUG mutations (Figure 6), regardless of thehost strain’s capacity to degrade mRNA. The effects ofthe wild-type uORF to reduce downstream translation

Molecular Cell458
were thus independent of subsequent effects on tran-script stability. However, additional translational effectswere observed in our studies. The efficiency of luciferaseproduction from mRNAs shown to be NMD substrateswas selectively reduced in upf1D and nmd2D cells rela-tive to NMD+ cells; the relative translatability of these re-porters in upf1D and nmd2D strains was similar (Figures6A and 6B; data not shown). This effect appeared in-dependent of ribosome stalling because it was alsoobserved with mRNA containing the improved-contextD13N uORF. This indicates that, while mRNA affectedby NMD can be present at an increased level in nmd2
strains, its translatability can nevertheless be impairedrelative to an NMD+ strain. For example, when theCPA1-LUC reporter contained the wild-type uORF inits native initiation context, its mRNA level was 7-foldhigher in the nmd2D strain relative to the NMD+ strain(Figure 2B). However, due to reduced translatability ofthis reporter in the nmd2D strain, the absolute amountof luciferase in nmd2D cells was only approximately2-fold greater than in NMD+ cells (data not shown). This2-fold increase in reporter activity did not correlatewell with the change in mRNA levels but is biologicallysignificant since genetic selection for trans-acting fac-tors that increased expression of CPA1 led to the isola-tion of a upf1 allele (Messenguy et al., 2002; Thuriauxet al., 1972).
The diminished translation efficiency of nonsense-containing mRNAs in upf1D and nmd2D cells might beattributable to overall reduced translation initiation onthem (e. g., the mRNAs are sequestered). Alternatively,events subsequent to termination codon recognitioncan differ markedly in nmd2 cells, and these might con-tribute to the observed reduced translation efficiency. Invivo, several different approaches have shown thatnmd2 cells promote nonsense codon read through(Keeling et al., 2004; Maderazo et al., 2000; Wenget al., 1996a, 1996b) and, in vitro, toeprinting indicatesthat nmd2 extracts eliminate an aberrant toeprint char-acteristic of a premature termination event (Amraniet al., 2004). Therefore, independent of the effects ofArg-induced ribosome stalling, reduced translational ef-ficiency may arise from changes in uORF translation ter-mination in nmd2 mutants that impedes ribosomeaccess to downstream start codons. Such effects maynot, however, occur universally on mRNAs containingearly stop codons (Muhlrad and Parker, 1999).
In conclusion, we show that translational control canserve to specifically target a wild-type mRNA for NMD ina regulated manner. Although the regulation of NMD issuspected to play important biological roles (He et al.,2003; Mendell et al., 2004), the mechanisms responsibleare poorly understood. In mammalian cells, regulatedNMD has been linked to alternative splicing (Greenet al., 2003; Lejeune and Maquat, 2005; Wollertonet al., 2004), and NMD in C. elegans can be regulatedby a mechanism that involves translational repressionat uORFs (Lee and Schedl, 2004). The S. cerevisiaeCPA1 uORF system shows that ribosome stalling ata regulatory uORF controls NMD and indicates that met-abolic control pathways can exploit NMD for their re-sponse to metabolites. The uORF-containing mRNAsthat encode products involved in amino acid homeosta-sis are a conserved class of natural NMD substrate (He
et al., 2003; Mendell et al., 2004). Since the CPA1 uORFencodes an evolutionarily conserved peptide that joinsa growing number of examples of nascent peptidesthat control mRNA degradation (Onouchi et al. [2005]and references therein), our studies raise the possibilitythat a conserved mechanism to regulate NMD involvesuORF-encoded peptides that function to regulate ribo-somes. The impact of regulatory uORFs on gene ex-pression can thus extend beyond their effects ontranslation.
Experimental Procedures
Yeast Strains, Growth Conditions, and General Methods
Standard yeast methods were as described (Burke et al., 2000). The
strains used are listed in Table S2. Strains for steady-state in vivo
analyses were created by transforming the plasmids listed in Table
S1 into isogenic strains HFY1200 (wild-type), HFY870 (upf1D),
HFY1300 (nmd2D), HFY1080 (xrn1D), and HFY1170 (ski2D). Strains
YRP582 and YAH01 contained thermolabile RNA polymerase II
and were used in temperature shift experiments. Translation ex-
tracts were prepared from strains MBS and MBS nmd2D.
Synthetic medium lacking tryptophan was used to select for and
maintain CPA1-LUC expression plasmids. For Arg regulation ex-
periments, cells were grown in appropriately supplemented liquid
minimal medium (Werner et al., 1987) that lacked or contained 1
mg/ml Arg.
Plasmids
The plasmids used are listed in Table S1. All in vivo CPA1-LUC ex-
pression plasmids are derivatives of YCplac22 (Gietz and Sugino,
1988) and contain the PGK1 promoter followed by a firefly luciferase
cistron that had its 50 leader and translation initiation codon re-
placed with BglII-NcoI fragments from pAG101, pAG103, and
pAG105 (Wang et al., 1999) to produce pAG121, pAG122, and
pAG123, respectively. Plasmids pAG101 and pAG103 (Wang et al.,
1999) were used for PCR with primers AG35 (50-TGTTGAAGATC
TACCCTTTTTGCAGATTTGAAATAAAAAAATTGAAAAAT GTTTAGC
TTATCGAAC-30) and AG3 (50-ATCTGACCATGGTTGAAATATTTTTA
GGAGTGGTT-30) to produce 200 bp PCR products that were di-
gested with BglII and NcoI and subcloned to create pAG134 and
pAG135, respectively. Genomic DNA isolated from HFY1200 cells
and primers AG31 (50-CTGGATTCTAGATTCACGCAACCCTTGATT
GG-30) and AG32 (50-TGTTGAAGATCTGACCACCTTTTGCCTCTGG
G-30) were used to PCR amplify a 0.9 kb CPA1 fragment that was di-
gested with BglII and XbaI and subcloned to create plasmid
pAG133. pAG101, pAG103, and pAG105 were used for in vitro anal-
yses (Wang et al., 1999). pAG136 and pAG137 were constructed by
replacing BglII-NcoI fragments from pAG101 and pAG103 with the
respective fragments from pAG134 and pAG135.
RNA Analyses
mRNA decay rates were measured in 100 ml cultures of YRP582 and
YAH01 grown in (appropriately supplemented) minimal media that
lacked or contained 1 mg/ml Arg. At an OD600 of w0.45, cells
were spun down, resuspended in 20 ml of the same medium, re-
turned to a 24ºC shaker incubator for 10 min., and then abruptly ad-
justed to 36ºC by adding an equal volume of medium at 50ºC and
transferring the culture to a shaker at 36ºC. Alternatively, cultures
were grown in minimal medium, shifted to 36ºC with the same me-
dium, and, 3 min after the temperature shift, Arg was added to 1 mg/
ml (or water was added as a control). At the indicated times after
a temperature shift, cells from aliquots (1.9 ml) were collected and
frozen with liquid nitrogen. To measure steady-state mRNA levels,
cultures (40 ml) were grown in YPD to an OD600 of w0.7 and cell ali-
quots (20 ml) were collected. Additional aliquots from these cultures
were harvested for reporter enzyme activity analyses. Cell pellets
were stored at 280ºC.
Total RNA was isolated using hot phenol (Herrick et al., 1990).
RNAs (8 mg) were fractionated on formaldehyde-agarose gels and
analyzed by northern blotting. Relative mRNA levels were deter-
mined by quantitation with a Molecular Dynamics PhosphorImager.

uORF-Mediated NMD in Yeast459
CPA1 and CPA1-LUC blots were normalized for loading using
SCR1, a stable RNA polymerase III transcript. The following DNA
fragments were gel purified and used to generate radiolabeled
probes for blotting: for CPA1, a 0.9 kb BglII-XbaI fragment from
pAG133; for CPA1-LUC, a 1.6 kb EcoRI-SalI fragment from
pDM345 (provided by R. Ganesan), which contains luciferase-
coding and 30-PGK1 sequences; for CYH2, a 0.6 kb EcoRI-HindIII
fragment from pGEM4Z-CYH2 (Herrick et al., 1990); and for SCR1,
a 0.4 kb EcoRI fragment from pCR2.1-SCR1 (provided by R. Gane-
san), in which SCR1 was obtained by PCR with genomic DNA and
primers SCR1-1 and SCR1-2 (He and Jacobson, 2001).
Polysome Analyses
Exponential cultures grown in YPD (125 ml) were harvested at an
A600 of 0.6, and their polysome profiles analyzed on high salt 7%–
47% sucrose gradients (MacKay et al., 2004). Twelve fractions
were collected from each gradient and RNA prepared from each
as follows: first, 400 ml of each fraction was mixed with 0.75 ng of
synthetic capped and polyadenylated sea pansy luciferase RNA
(Wang et al., 1998), which was added as a control for monitoring re-
covery, and precipitated with sodium acetate and ethanol. Precipi-
tated RNAs were purified with the GenElute Mammalian Total RNA
Kit (Sigma), after resuspension in the kit’s lysis buffer + b-mercap-
toethanol, by the manufacturer’s protocol. RNA was ethanol precip-
itated, resuspended in water, and analyzed by northern blotting.
The signals obtained by probing with firefly LUC and CPA1 were
normalized to those from sea pansy LUC.
Luciferase Enzyme Activity Analyses
Duplicate aliquots (3 ml) from cultures were collected by centrifuga-
tion at 4ºC, and the cell pellets were stored at 280ºC. These pellets,
on ice, were resuspended in 100 ml of buffer (0.1 M KPO4 [pH 7.8],
1.0 mM DTT, 1.0 mM PMSF), followed by the addition of 100 ml
acid-washed 0.5 mm glass beads, and cycles of vortexing for 15 s
followed by 30 s on ice, for 225 s. Samples were centrifuged for 5
min at 4ºC; the supernatants were transferred to fresh tubes, frozen
at 280ºC, and thawed for measurement of luciferase activity (Wang
et al., 1998). Relative light units (RLU) were normalized to total pro-
tein concentration (Bradford assay). Samples were assayed in
triplicate.
Cell-Free Translation and Primer Extension Inhibition Analyses
Translation extracts were prepared, and translation and primer ex-
tension inhibition (toeprint) reactions accomplished, as described
(Gaba et al., 2001; Sachs et al., 2002; Wang et al., 1999).
Supplemental Data
Supplemental Data include two tables and Supplemental Refer-
ences and can be found with this article online at http://www.
molecule.org/cgi/content/full/20/3/449/DC1/.
Acknowledgments
This work was supported by grants (award GM47498 to M.S.S. and
award R37 GM27757 to A.J.) from the National Institutes of Health.
A.G. was supported by a Research Supplement for Underrepre-
sented Minorities to GM47498. We are indebted to Robin Ganesan
and Feng He for strains, advice, and technical assistance with initial
experiments and to Vivian McKay and David Morris for assistance
and advice with the polysome analyses.
Received: November 23, 2004
Revised: August 10, 2005
Accepted: September 21, 2005
Published: November 10, 2005
References
Amrani, N., Ganesan, R., Kervestin, S., Mangus, D.A., Ghosh, S.,
and Jacobson, A. (2004). A faux 30-UTR promotes aberrant termina-
tion and triggers nonsense-mediated mRNA decay. Nature 432,
112–118.
Anderson, J.S., and Parker, R.P. (1998). The 30 to 50 degradation of
yeast mRNAs is a general mechanism for mRNA turnover that
requires the SKI2 DEVH box protein and 30 to 50 exonucleases of
the exosome complex. EMBO J. 17, 1497–1506.
Belgrader, P., Cheng, J., and Maquat, L.E. (1993). Evidence to im-
plicate translation by ribosomes in the mechanism by which non-
sense codons reduce the nuclear level of human triosephosphate
isomerase mRNA. Proc. Natl. Acad. Sci. USA 90, 482–486.
Burke, D., Dawson, D., and Stearns, T. (2000). Methods in Yeast
Genetics: A Cold Spring Harbor Laboratory Course Manual (Plain-
view, New York: Cold Spring Harbor Laboratory Press).
Crabeel, M., LaValle, R., and Glansdorff, N. (1990). Arginine-specific
repression in Saccharomyces cerevisiae: kinetic data on ARG1 and
ARG3 mRNA transcription and stability support a transcriptional
control mechanism. Mol. Cell. Biol. 10, 1226–1233.
Davis, R.H. (1986). Compartmental and regulatory mechanisms in
the arginine pathways of Neurospora crassa and Saccharomyces
cerevisiae. Microbiol. Rev. 50, 280–313.
Delbecq, P., Calvo, O., Filipkowski, R.K., Pierard, A., and
Messenguy, F. (2000). Functional analysis of the leader peptide of
the yeast gene CPA1 and heterologous regulation by other fungal
peptides. Curr. Genet. 38, 105–112.
Fang, P., Spevak, C.C., Wu, C., and Sachs, M.S. (2004). A nascent
polypeptide domain that can regulate translation elongation. Proc.
Natl. Acad. Sci. USA 101, 4059–4064.
Freitag, M., Dighde, N., and Sachs, M.S. (1996). A UV-induced
mutation in Neurospora that affects translational regulation in
response to arginine. Genetics 142, 117–127.
Gaba, A., Wang, Z., Krishnamoorthy, T., Hinnebusch, A.G., and
Sachs, M.S. (2001). Physical evidence for distinct mechanisms of
translational control by upstream open reading frames. EMBO J.
20, 6453–6463.
Gietz, R.D., and Sugino, A. (1988). New yeast-Escherichia coli shut-
tle vectors constructed with in vitro mutagenized yeast genes lack-
ing six-base pair restriction sites. Gene 74, 527–534.
Gozalbo, D., and Hohmann, S. (1990). Nonsense suppressors par-
tially revert the decrease of the mRNA level of a nonsense mutant
allele in yeast. Curr. Genet. 17, 77–79.
Green, R.E., Lewis, B.P., Hillman, R.T., Blanchette, M., Lareau, L.F.,
Garnett, A.T., Rio, D.C., and Brenner, S.E. (2003). Widespread pre-
dicted nonsense-mediated mRNA decay of alternatively-spliced
transcripts of human normal and disease genes. Bioinformatics
Suppl. 1 19, i118–i121.
He, F., and Jacobson, A. (2001). Upf1p, Nmd2p, and Upf3p regulate
the decapping and exonucleolytic degradation of both nonsense-
containing mRNAs and wild-type mRNAs. Mol. Cell. Biol. 21,
1515–1530.
He, F., Peltz, S.W., Donahue, J.L., Rosbash, M., and Jacobson, A.
(1993). Stabilization and ribosome association of unspliced pre-
mRNAs in a yeast upf1 – mutant. Proc. Natl. Acad. Sci. USA 90,
7034–7038.
He, F., Li, X., Spatrick, P., Casillo, R., Dong, S., and Jacobson, A.
(2003). Genome-wide analysis of mRNAs regulated by the non-
sense-mediated and 50 to 30 mRNA decay pathways in yeast.
Mol. Cell 12, 1439–1452.
Hennigan, A.N., and Jacobson, A. (1996). Functional mapping of the
translation-dependent instability element of yeast MATalpha1
mRNA. Mol. Cell. Biol. 16, 3833–3843.
Herrick, D., Parker, R., and Jacobson, A. (1990). Identification and
comparison of stable and unstable mRNAs in Saccharomyces cer-
evisiae. Mol. Cell. Biol. 10, 2269–2284.
Hsu, C.L., and Stevens, A. (1993). Yeast cells lacking 50/30 exori-
bonuclease 1 contain mRNA species that are poly(A) deficient
and partially lack the 50 cap structure. Mol. Cell. Biol. 13, 4826–
4835.
Ishigaki, Y., Li, X., Serin, G., and Maquat, L.E. (2001). Evidence for
a pioneer round of mRNA translation: mRNAs subject to non-
sense-mediated decay in mammalian cells are bound by CBP80
and CBP20. Cell 106, 607–617.
Jacobson, A., and Peltz, S.W. (2000). Destabilization of nonsense-
containing transcripts in Saccharomyces cerevisiae. In Transla-
tional Control of Gene Expression, N. Sonenberg, J.W.B. Hershey,

Molecular Cell460
and M.B. Mathews, eds. (Cold Spring Harbor, New York: Cold
Spring Harbor Laboratory Press), pp. 827–847.
Keeling, K.M., Lanier, J., Du, M., Salas-Marco, J., Gao, L., Kaenjak-
Angeletti, A., and Bedwell, D.M. (2004). Leaky termination at prema-
ture stop codons antagonizes nonsense-mediated mRNA decay in
S. cerevisiae. RNA 10, 691–703.
Kitamoto, K., Yoshizawa, K., Ohsumi, Y., and Anraku, Y. (1988). Dy-
namic aspects of vacuolar and cytosolic amino acid pools of Sac-
charomyces cerevisiae. J. Bacteriol. 170, 2683–2686.
Lee, M.H., and Schedl, T. (2004). Translation repression by GLD-1
protects its mRNA targets from nonsense-mediated mRNA decay
in C. elegans. Genes Dev. 18, 1047–1059.
Lejeune, F., and Maquat, L.E. (2005). Mechanistic links between
nonsense-mediated mRNA decay and pre-mRNA splicing in mam-
malian cells. Curr. Opin. Cell Biol. 17, 309–315.
Losson, R., and Lacroute, F. (1979). Interference of nonsense muta-
tions with eukaryotic messenger RNA stability. Proc. Natl. Acad.
Sci. USA 76, 5134–5137.
Luo, Z., Freitag, M., and Sachs, M.S. (1995). Translational regulation
in response to changes in amino acid availability in Neurospora
crassa. Mol. Cell. Biol. 15, 5235–5245.
MacKay, V.L., Li, X., Flory, M.R., Turcott, E., Law, G.L., Serikawa, K.A.,
Xu, X.L., Lee, H., Goodlett, D.R., Aebersold, R., et al. (2004). Gene
expression analyzed by high-resolution state array analysis and
quantitative proteomics: response of yeast to mating pheromone.
Mol. Cell. Proteomics 3, 478–489.
Maderazo, A.B., He, F., Mangus, D.A., and Jacobson, A. (2000).
Upf1p control of nonsense mRNA translation is regulated by
Nmd2p and Upf3p. Mol. Cell. Biol. 20, 4591–4603.
Maderazo, A.B., Belk, J.P., He, F., and Jacobson, A. (2003). Non-
sense-containing mRNAs that accumulate in the absence of a func-
tional nonsense-mediated mRNA decay pathway are destabilized
rapidly upon its restitution. Mol. Cell. Biol. 23, 842–851.
Maquat, L.E. (2004). Nonsense-mediated mRNA decay: splicing,
translation and mRNP dynamics. Nat. Rev. Mol. Cell Biol. 5, 89–99.
Mendell, J.T., Sharifi, N.A., Meyers, J.L., Martinez-Murillo, F., and
Dietz, H.C. (2004). Nonsense surveillance regulates expression of
diverse classes of mammalian transcripts and mutes genomic
noise. Nat. Genet. 36, 1073–1078.
Messenguy, F., and Dubois, E. (2000). Regulation of arginine me-
tabolism in Saccharomyces cerevisiae: a network of specific and
pleiotropic proteins in response to multiple environmental signals.
Food Technol. Biotechnol. 38, 277–285.
Messenguy, F., Vierendeels, F., Pierard, A., and Delbecq, P. (2002).
Role of RNA surveillance proteins Upf1/CpaR, Upf2 and Upf3 in the
translational regulation of yeast CPA1 gene. Curr. Genet. 41, 224–
231.
Muhlrad, D., and Parker, R. (1999). Recognition of yeast mRNAs as
‘‘nonsense containing’’ leads to both inhibition of mRNA translation
and mRNA degradation: implications for the control of mRNA de-
capping. Mol. Biol. Cell 10, 3971–3978.
Nonet, M., Scafe, C., Sexton, J., and Young, R. (1987). Eucaryotic
RNA polymerase conditional mutant that rapidly ceases mRNA
synthesis. Mol. Cell. Biol. 7, 1602–1611.
Nott, A., Le Hir, H., and Moore, M.J. (2004). Splicing enhances
translation in mammalian cells: an additional function of the exon
junction complex. Genes Dev. 18, 210–222.
Onouchi, H., Nagami, Y., Haraguchi, Y., Nakamoto, M.,
Nishimura, Y., Sakurai, R., Nagao, N., Kawasaki, D., Kadokura, Y.,
and Naito, S. (2005). Nascent peptide-mediated translation elonga-
tion arrest coupled with mRNA degradation in the CGS1 gene of
Arabidopsis. Genes Dev. 19, 1799–1810.
Orbach, M.J., Sachs, M.S., and Yanofsky, C. (1990). The Neuros-
pora crassa arg-2 locus: structure and expression of the gene en-
coding the small subunit of arginine-specific carbamoyl
phosphate synthetase. J. Biol. Chem. 265, 10981–10987.
Ruiz-Echevarria, M.J., and Peltz, S.W. (2000). The RNA-binding
protein Pub1 modulates the stability of transcripts containing up-
stream open reading frames. Cell 101, 741–751.
Ruiz-Echevarria, M.J., Gonzalez, C.I., and Peltz, S.W. (1998). Iden-
tifying the right stop: determining how the surveillance complex
recognizes and degrades an aberrant mRNA. EMBO J. 17, 575–589.
Sachs, M.S., Wang, Z., Gaba, A., Fang, P., Belk, J., Ganesan, R.,
Amrani, N., and Jacobson, A. (2002). Toeprint analysis of the posi-
tioning of translational apparatus components at initiation and ter-
mination codons of fungal mRNAs. Methods 26, 105–114.
Singh, G., and Lykke-Andersen, J. (2003). New insights into the for-
mation of active nonsense-mediated decay complexes. Trends
Biochem. Sci. 28, 464–466.
Thuriaux, P., Ramos, F., Pierard, A., Grenson, M., and Wiame, J.M.
(1972). Regulation of carbamoylphosphate synthetase belonging to
the arginine biosynthetic pathway of Saccharomyces cerevisiae. J.
Mol. Biol. 67, 277–287.
Vilela, C., Linz, B., Rodrigues-Pousada, C., and McCarthy, J.E.
(1998). The yeast transcription factor genes YAP1 and YAP2 are
subject to differential control at the levels of both translation and
mRNA stability. Nucleic Acids Res. 26, 1150–1159.
Wang, Z., Fang, P., and Sachs, M.S. (1998). The evolutionarily con-
served eukaryotic arginine attenuator peptide regulates the
movement of ribosomes that have translated it. Mol. Cell. Biol.
18, 7528–7536.
Wang, Z., Gaba, A., and Sachs, M.S. (1999). A highly conserved
mechanism of regulated ribosome stalling mediated by fungal argi-
nine attenuator peptides that appears independent of the charging
status of arginyl-tRNAs. J. Biol. Chem. 274, 37565–37574.
Weng, Y., Czaplinski, K., and Peltz, S.W. (1996a). Genetic and bio-
chemical characterization of mutations in the ATPase and helicase
regions of the Upf1 protein. Mol. Cell. Biol. 16, 5477–5490.
Weng, Y., Czaplinski, K., and Peltz, S.W. (1996b). Identification and
characterization of mutations in the UPF1 gene that affect non-
sense suppression and the formation of the Upf protein complex
but not mRNA turnover. Mol. Cell. Biol. 16, 5491–5506.
Werner, M., Feller, A., Messenguy, F., and Pierard, A. (1987). The
leader peptide of yeast CPA1 is essential for the translational re-
pression of its expression. Cell 49, 805–813.
Wiegand, H.L., Lu, S., and Cullen, B.R. (2003). Exon junction com-
plexes mediate the enhancing effect of splicing on mRNA expres-
sion. Proc. Natl. Acad. Sci. USA 100, 11327–11332.
Wollerton, M.C., Gooding, C., Wagner, E.J., Garcia-Blanco, M.A.,
and Smith, C.W. (2004). Autoregulation of polypyrimidine tract
binding protein by alternative splicing leading to nonsense-
mediated decay. Mol. Cell 13, 91–100.
Zhang, S., Welch, E.M., Hogan, K., Brown, A.H., Peltz, S.W., and
Jacobson, A. (1997). Polysome-associated mRNAs are substrates
for the nonsense-mediated mRNA decay pathway in Saccharomy-
ces cerevisiae. RNA 3, 234–244.




















