
Revisiting Ross and Sims (1971): toward a molecular phylogeny of the Biddulphiaceae and...
16
REVISITING ROSS AND SIMS (1971): TOWARD A MOLECULAR PHYLOGENY OF THE BIDDULPHIACEAE AND EUPODISCACEAE (BACILLARIOPHYCEAE) 1 Matt P. Ashworth, 2 Teofil Nakov Section of Integrative Biology, University of Texas at Austin, Austin, Texas 78712, USA and Edward C. Theriot Texas Natural Science Center, University of Texas at Austin, Austin, Texas 78712, USA The ocellate and pseudocellate diatoms in the Eupodiscaceae and Biddulphiaceae (respectively) are common inhabitants of the marine littoral (and plankton zone) with a rich fossil history making them important components of marine stratigraphic studies and good candidates for molecular dating work. These diatoms are important for un- derstanding the phylogeny of the diatoms as a whole, as molecular phylogenies have blurred the traditional distinction between the pennate and multipolar non-pennate diatoms. However, the convoluted taxonomic history of these groups has the potential to disrupt both stratigraphic and molecular dating studies. Although efforts have been made to examine frustule morphology of several ocellate and pseudocellate diatoms and develop a morphological scheme to define genera, very little work has been done to determine how these groups are interrelated. In this study, we use nuclear and chloroplast molecular markers to construct a phylogeny of a diverse sampling of Eupodiscaceae and Biddulphiaceae taxa. The ocellus-bearing taxa (Eupodiscaceae) are monophyletic, and thus the ocellus may be a useful character in delimiting the Eupodiscaceae, the Biddulphiaceae are polyphyletic and scattered across a number of lineages of multipolar non-pennate diatoms. Hypothesis testing aimed at assessing the likeliness of several morphology based hypotheses against the molecular data highlights uncertainty in both types of data. We present evidence that there are monophyletic genera within both the Biddulphiaceae and Eupodiscaceae, and recommend the taxa within the Odontella mobilensis/sinensis/regia clade be transferred to a new genus: Trieres Ashworth & Theriot. Key index words: Biddulphiaceae; diatoms; Eupodisc- aceae; phylogeny; SEM Sch€ utt (1896) created the subfamily Biddulphioi- deae and defined it as those diatoms with two or more poles, generally elliptical in shape, with poles elevated as bumps or horns. He included in the Biddulphioideae a number of genera which broadly resemble Biddulphia Gray, such as Odontella C. Agardh, Triceratium Ehrenberg, Eucampia Ehrenberg, Isthmia C. Agardh, Hemiaulus Heiberg, Anaulus Ehrenberg, Hemidiscus G.C. Wallich, Eupodiscus J.W. Bailey, Cerataulus Ehrenberg, and Chaetoceros Ehren- berg. Molecular data place these diatoms together with a number of other polar and even radially structured non-pennate diatoms (the Thalassiosi- rales) in a group formally designated the Mediophy- ceae (Medlin and Kaczmarska 2004). Support for Mediophyceae monophyly has been equivocal (Theriot et al. 2009, 2010, 2011) and some authors consider the group to be paraphyletic (Adl et al. 2005). Hendey (1971) lamented the “feeling of apathy” by diatomists toward the diatoms fitting Sch€ utt’s def- inition (Hendey used the term “Biddulphiaceae”), and described the group as “a heterogenous collec- tion of wholly dissimilar units, bound together only by our inability to see clearly taxonomic boundaries providing bases for useful subdivisions.” That same year, Ross and Sims (1971) used the SEM in an attempt to find such boundaries. They argued that there were generally two types of pore fields present in the diatoms they observed: fine pore fields sur- rounded by a solid, unmarked rim (“ocelli”), or apices marked by a gradation of the valve markings to a field of fine pores (“pseudocelli”). Further- more, Ross and Sims grouped taxa observed into general categories based on valve perforation: po- rose valves with pore occlusions that ranged from simple bars (such as in Biddulphiopsis von Stosch and Simonsen or Lampriscus A. Schmidt) to flat or domed vela (such as those in Biddulphia biddulphi- ana [J.W. Smith] C.S. Boyer and Odontella aurita [Lyngbye] C. Agardh, respectively); or loculate, chambered valves (such as Odontella mobiliensis [J.W. Bailey] Grunow or Odontella rhombus [Ehrenberg] K€ utzing). They also identified a number of taxa, which bore pores with domed cribra surrounded by incomplete and irregular chambers, dubbed 1 Received 12 December 2012. Accepted 8 August 2013. 2 Author for correspondence: e-mail [email protected]. Editorial Responsibility: R. Wetherbee (Associate Editor) J. Phycol. 49, 1207–1222 (2013) © 2013 Phycological Society of America DOI: 10.1111/jpy.12131 1207
Transcript of Revisiting Ross and Sims (1971): toward a molecular phylogeny of the Biddulphiaceae and...

REVISITING ROSS AND SIMS (1971): TOWARD A MOLECULAR PHYLOGENY OF THEBIDDULPHIACEAE AND EUPODISCACEAE (BACILLARIOPHYCEAE)1
Matt P. Ashworth,2 Teofil Nakov
Section of Integrative Biology, University of Texas at Austin, Austin, Texas 78712, USA
and Edward C. Theriot
Texas Natural Science Center, University of Texas at Austin, Austin, Texas 78712, USA
The ocellate and pseudocellate diatoms in theEupodiscaceae and Biddulphiaceae (respectively) arecommon inhabitants of the marine littoral (andplankton zone) with a rich fossil history makingthem important components of marine stratigraphicstudies and good candidates for molecular datingwork. These diatoms are important for un-derstanding the phylogeny of the diatoms as awhole, as molecular phylogenies have blurred thetraditional distinction between the pennate andmultipolar non-pennate diatoms. However, theconvoluted taxonomic history of these groups hasthe potential to disrupt both stratigraphic andmolecular dating studies. Although efforts have beenmade to examine frustule morphology of severalocellate and pseudocellate diatoms and develop amorphological scheme to define genera, very littlework has been done to determine how these groupsare interrelated. In this study, we use nuclear andchloroplast molecular markers to construct aphylogeny of a diverse sampling of Eupodiscaceaeand Biddulphiaceae taxa. The ocellus-bearing taxa(Eupodiscaceae) are monophyletic, and thus theocellus may be a useful character in delimiting theEupodiscaceae, the Biddulphiaceae are polyphyleticand scattered across a number of lineages ofmultipolar non-pennate diatoms. Hypothesis testingaimed at assessing the likeliness of severalmorphology based hypotheses against the moleculardata highlights uncertainty in both types of data. Wepresent evidence that there are monophyletic generawithin both the Biddulphiaceae and Eupodiscaceae,and recommend the taxa within the Odontellamobilensis/sinensis/regia clade be transferred to anew genus: Trieres Ashworth & Theriot.
Key index words: Biddulphiaceae; diatoms; Eupodisc-aceae; phylogeny; SEM
Sch€utt (1896) created the subfamily Biddulphioi-deae and defined it as those diatoms with two ormore poles, generally elliptical in shape, with poleselevated as bumps or horns. He included in theBiddulphioideae a number of genera which broadlyresemble Biddulphia Gray, such as Odontella C.Agardh, Triceratium Ehrenberg, Eucampia Ehrenberg,Isthmia C. Agardh, Hemiaulus Heiberg, AnaulusEhrenberg, Hemidiscus G.C. Wallich, Eupodiscus J.W.Bailey, Cerataulus Ehrenberg, and Chaetoceros Ehren-berg. Molecular data place these diatoms togetherwith a number of other polar and even radiallystructured non-pennate diatoms (the Thalassiosi-rales) in a group formally designated the Mediophy-ceae (Medlin and Kaczmarska 2004). Support forMediophyceae monophyly has been equivocal(Theriot et al. 2009, 2010, 2011) and some authorsconsider the group to be paraphyletic (Adl et al.2005).Hendey (1971) lamented the “feeling of apathy”
by diatomists toward the diatoms fitting Sch€utt’s def-inition (Hendey used the term “Biddulphiaceae”),and described the group as “a heterogenous collec-tion of wholly dissimilar units, bound together onlyby our inability to see clearly taxonomic boundariesproviding bases for useful subdivisions.” That sameyear, Ross and Sims (1971) used the SEM in anattempt to find such boundaries. They argued thatthere were generally two types of pore fields presentin the diatoms they observed: fine pore fields sur-rounded by a solid, unmarked rim (“ocelli”), orapices marked by a gradation of the valve markingsto a field of fine pores (“pseudocelli”). Further-more, Ross and Sims grouped taxa observed intogeneral categories based on valve perforation: po-rose valves with pore occlusions that ranged fromsimple bars (such as in Biddulphiopsis von Stoschand Simonsen or Lampriscus A. Schmidt) to flat ordomed vela (such as those in Biddulphia biddulphi-ana [J.W. Smith] C.S. Boyer and Odontella aurita[Lyngbye] C. Agardh, respectively); or loculate,chambered valves (such as Odontella mobiliensis [J.W.Bailey] Grunow or Odontella rhombus [Ehrenberg]K€utzing). They also identified a number of taxa,which bore pores with domed cribra surroundedby incomplete and irregular chambers, dubbed
1Received 12 December 2012. Accepted 8 August 2013.2Author for correspondence: e-mail [email protected] Responsibility: R. Wetherbee (Associate Editor)
J. Phycol. 49, 1207–1222 (2013)© 2013 Phycological Society of AmericaDOI: 10.1111/jpy.12131
1207

“pseudoloculate” valves (Triceratium favus Ehrenbergand T. dictyotum P.A. Sims and R. Ross).
Using the different types of apical pore fields andvalve perforation characters, Ross and Sims createdfour informal groupings: taxa with porose valvesand pseudocelli, taxa with loculate valves andpseudocelli, taxa with porose valves and ocelli andtaxa with loculate valves and ocelli. They suggestedthat the porose/pseudocellate taxa belong in Biddul-phia, though they posited that those taxa withoutinternal ribs might belong in a different genus. Theloculate/pseudocellate taxa were assigned to Trigoni-um Cleve. The ocellate taxa they examined showeda greater degree of variation in valve morphology,however. They divided the loculate/ocellate taxainto two genera: Cerataulus, for taxa which featureda contorted valve (i.e., Cerataulus turgidus [Ehren-berg] Ehrenberg) and Zygoceros Ehrenberg, for taxawithout torsion (i.e., O. mobiliensis, O. rhombus). Theporose/ocellate taxa presented a particularly diffi-cult situation, because of a great deal of morpholog-ical variability beyond these two characters. Thus,Ross and Sims (1971) proposed two options: eitherall porose/ocellate taxa should fall under Odontella(type = O. aurita), or these taxa should be dividedinto a number of genera, including Odontella, thepseudoloculate Triceratium, Amphitetras Ehrenberg,and Amphipentas Ehrenberg.
Other classifications generally followed Ross andSims (1971) in terms of suprageneric groupings,although there was still considerable disagreementabout the relationship (and rank) of these supra-generic groups. Simonsen (1979) created the“Biddulphiineae” which was similar in compositionto Sch€utt’s Biddulphioideae. The Family Biddulphia-ceae was characterized by pseudocelli while theEupodiscaceae was characterized by ocelli. Simon-sen’s (1979) Biddulphiineae was not monophyletic.He placed the Biddulphiaceae in a clade with Chae-toceraceae and Lithodemiaceae and the Eupodisca-ceae were placed as sister to pennates. Round et al.(1990) proposed a similar scheme, but elevatedwhat were essentially Simonsen’s Biddulphiaceaeand Eupodiscaceae to the ordinal level: Biddulphi-ales and Triceratiales, respectively. These wereplaced in the subclass “Biddulphiophycidae.” Niko-laev (1990) split the biddulphioids across threeorders: the Triceratiaceae in the Pyxidiculales, theTrigoniaceae and Eupodiscaceae in the Coscin-odiscales (resembling Sch€utt), and the Auliscaceae,Biddulphiaceae, and Isthmiaceae in the Biddulphi-ales. However, Simonsen’s two-family proposalseems to occur most commonly in the literature.This ambiguity is actually seen in the molecular dataas well—while certain clades of diatoms are well-sup-ported, the relationships between these large clades(roughly analogous to order and family-level) arepoorly resolved (Theriot et al. 2011).
Far less has been written regarding the phyloge-netic relationships within the Eupodiscaceae and
Biddulphiaceae, possibly due to the taxonomicuncertainty discussed previously. Glezer (1979)looked at the phylogeny of the Biddulphiaceae ofextant and fossil forms, but focused primarily onthe areolar structure and the overall frustule shape,asserting that cylindrical forms were ancestral to anellipsoidal lineage (such as Eunotogramma Weisse,Eucampia, Biddulphia) and prismatic lineage (such asTrinacria Heiberg, Amphitetras, or the vaguelydescribed Sheshukovia Glezer). Glezer mentions Rossand Sims’ ocellus, but suggests that it has evolvedseveral times across the diatoms: in Odontella(derived from Biddulphia in her tree diagram), inAmphitetras (derived from Sheshukovia) and inTriceratium (an altogether different lineage notincluded in the tree). In a later paper, Ross andSims (1987) point to the variability in valve outline,even within a species (using Odontella retiformis[A. Mann] von Stosch as an example) as evidenceagainst Glezer’s scheme, however.In a later paper on family and order-level taxon-
omy in the “centric” diatoms, Ross and Sims (1973)state that the defining character for the Eupodisca-ceae is the ocellus, and suggest that this structurewas derived from the “field of smaller areolae onthe elevations that characterizes [the Biddulpia-ceae].” Simonsen (1979) also used the ocellus as akey character of the Eupodiscaceae. They acknowl-edge that the loculate and pseudoloculate valve con-ditions occur outside of the Eupodiscaceae (theyare present on Coscinodiscus Ehrenberg and Stepha-nopyxis [Ehrenberg] Ehrenberg, respectively). How-ever, they suggest it is more likely for loculate valvesto have evolved twice (once in the Eupodiscaceaeand once in the Coscinodiscaceae) than for ocellito have evolved twice.Simonsen (1979) disagreed regarding the impor-
tance of the ocellus, stating that the “true ocellus”could be found in other taxa with no affinity to theEupodiscaceae. As an example, he cites Cymatosiralorenziana Grunow. The Cymatosiraceae do featureelevated, rimmed apical pore fields. However, theytypically possess fewer pores, feature short spikesbetween the pores and are concave in structurerather than convex like the ocelli of Odontella andTriceratium. To reflect these differences, the porefields of the Cymatosiraceae were given the name“ocellulus” (Hasle et al. 1983). Hasle et al. (1983)do not comment on the relationship between theocellulus and the eupodiscoid ocellus except to saythey are similarly structured. The term “ocellus” hasbeen used to describe several different structures,such as the apical pore field in the araphid pennatediatom Bleakeleya Round and is discussed in Lobbanet al. (2011).Part of the ambiguity in previous attempts at clas-
sifying these diatoms may lie in the reliance ofplesiomorphic characters to delimit taxonomy. Thisis a well-documented problem in diatoms; classicalstructural groups such as “centric” and “araphid
1208 MATT P. ASHWORTH ET AL.

pinnate,” defined by lack of apomorphic characters,continue to be used in formal classifications despitetheir non-monophyletic nature (see Williams andKociolek 2007). Some authors continue to argueagainst monophyly as the necessary and sufficientbasis for classification (Medlin 2010). Within thebiddulphioid/eupodiscoid diatoms, all authors whohave written on the subject seem to agree that valveperforation characters are important, but the issuebecomes how these characters are interpreted on aphylogeny. For example, groups of larger poresgrading rapidly into smaller pores (“pseudocellus”)may be homologous within some deeper level of thediatom tree, representing a plesiomorphic conditionrelative to a pore field surrounded by a hyaline ring(“ocellus”). In turn, this may reflect plesiomorphicresemblance to even more derived conditions suchas the ocellulus, representing synapomorphy withinocellate diatoms.
Certainly a helpful and needed approach wouldbe an analysis of a full matrix to resolve questionsof synapomorphy versus plesiomorphy in the mor-phological data. We suggest that a molecularapproach to the phylogeny is a needed “first step”to guide further studies with a full matrix; thematrix would have to include detailed analysis ofthese and other structures from numerous fossiltaxa (not necessarily just those in the Biddulphia-ceae and Eupodiscaceae, since their relationshipwith the rest of the diatom tree is still poorly under-stood) for which there is no DNA data for compari-son anyway. Instead, we will treat the variouscharacters argued as diagnostic for groups (particu-larly the characters of Ross and Sims 1971), as pro-posed synapomorphies and examine the extent towhich there is an agreement between hypothesessuggested by molecular data and these hypothesizedmorphological synapomorphies.
Our molecular data set extends the three-genedata set (nuclear-encoded SSU rDNA, and chloro-plast-encoded rbcL and psbC) of Theriot et al.(2010, 2011), by nearly doubling the number of dia-tom sequences, with a particular increase in Medio-phyceae. We assess the correspondence betweenmolecular data and previously proposed classifi-cation schemes (Ross and Sims 1973, Glezer 1979,Simonsen 1979) or putative diagnostic characters(untested synapomorphies; Ross and Sims 1971) bycomparing the likelihoods of trees constrained toreflect previous hypotheses to the unconstrainedtree. This approach has been used to address ques-tions of classification and character evolution in awide array of taxa including diatoms (e.g., Ruck andTheriot 2011, Theriot et al. 2011), protists (e.g.,Embley et al. 1995, Deschamps et al. 2011), andmetazoans (e.g., Sperling et al. 2009). We recognizethat this approach has its limitations and that acladistic analysis of morphological characters wouldbe a preferred way of testing morphological synapo-morphies or correspondence/conflict between
molecular and morphological data (cf. Losos et al.2012). However, the hypothesis testing performedhere may be our best course of action until anextensive, coded morphological data set is com-piled. We also argue that enough morphologicaland molecular evidence exists to transfer threespecies from Ross and Sims’ concept of Zygoceros,O. mobiliensis, O. regia (M. Schultze) Simonsen, andO. sinensis (Greville) Grunow, to a new genus:Trieres.
MATERIALS AND METHODS
Samples were taken from a variety of benthic and plank-tonic habitats (Table S1 in the Supporting Information).Benthic taxa were isolated from hand-collected macrophytesor scraped directly from hard substrates by SCUBA or snorkeldivers. Planktonic isolates were isolated from collectionsmade by net with a 20 lm mesh size. The remaining sampledmaterial was preserved in a formaldehyde–seawater mix. Allmarine isolates were grown in f/2 medium (Guillard 1975) at27°C (Guam isolates) or at 14°C (northern CA, OR, WA iso-lates) in a Percival model I-36LL incubator under a 12:12light:dark photoperiod under flourescent lights. All other iso-lates were grown at room temperature in a north-facingwindow in ambient light at around 21°C–22°C (TX, FL, HIand Canary Island isolates).
Microscopy preparation. Wild and cultured material wasacid-cleaned with hydrogen peroxide and nitric acid andwashed until pH neutral. Permanent mounts for light micros-copy were made using Naphrax mounting medium.
For SEM preparation, cleaned wild material was filteredand dried onto Millipore 1.2 lm Isopore membrane filterswhile cultured material was dried onto 12 mm round cover-slips. Material was then coated with iridium or platinum/pal-ladium using a Cressington 208 Bench Top Sputter Coaterand observed with a Zeiss SUPRA 40 VP scanning electronmicroscope (Carl Zeiss Microscopy, Thornwood, NY, USA).
All strains used for DNA sequencing have LM and/or SEMphoto vouchers, available from http://www.protistcentral.org/Project/get/project_id/79 or the senior author.
DNA extraction, sequencing, and phylogenetic analysis. Aliquotsof cultured isolates were pelleted in a Sorvall RC-5B refriger-ated superspeed centrifuge for 20 min at 7,649g. DNA wasextracted from the diatom pellets using a Qiagen DNeasyPlant Mini Kit (Qiagen Sciences, Valencia, CA, USA) after45 s of cell disruption using 1.0 mm glass beads in a Mini-Beadbeater (Biospec Products Inc, Bartlesville, OK, USA).PCR amplification and sequencing of small-subunit nuclearrRNA, and the chloroplast-encoded rbcL and psbC markers,uses the primers and follows the protocols of Theriot et al.(2010).
Phylogenetic analyses were conducted using 245 diatomtaxa plus Bolidomonas pacifica L. Guillou & M.-J. Chr�etiennot-Dinet as the outgroup. Genbank accession numbers for thenew sequences presented in this article are listed in Table S1.All extracted DNA were given an accession number local tothe Theriot lab that corresponds to a slide and SEM stub vou-cher (“HK###”). The aligned data matrix is available in Fig-ure S1 in the Supporting Information. Sequence data werepartitioned by gene and by codon position and run in RAx-ML ver. 7.2.8 (Stamatakis 2006) for Maximum Likelihood(ML) analysis using the six-substitution GTR + G + I model.To obtain a most likely tree, we ran 1,024 ML optimizationseach starting from a parsimony tree. Clade support wasobtained from 1,000 nonparametric bootstrap replicates
REVISITING ROSS AND SIMS 1209

using the rapid bootstrap algorithm in RAxML. The tree withthe highest log likelihood score from 1,024 unconstrainedruns with bootstrap proportions is presented in Figure 1 andAppendix S1 in the Supporting Information. All RAxML runswere completed on the RANGER or LONESTAR supercom-puters at the Texas Advanced Computing Center (TACC).Two Bayesian Inference runs were also conducted with MrBa-yes ver. 3.1.2, using the same substitution model and parti-tioning strategy as the ML analyses. Two Markov chain MonteCarlo (MCMC) runs with four chains each (one cold andthree heated) were iterated for 108 generations. The poster-ior distributions of tree topology, branch lengths, and param-eter estimates were sampled every 1,000th iterationproducing a total of 105 samples. The posterior samples ofthe two runs were compared in AWTY (Wilgenbusch et al.2004) for evidence of convergence. The final 103 trees wereused to generate a majority rule consensus tree and obtainposterior probabilities for nodes (plotted to nodes of the MLphylogram in Figs. 1 and S1).
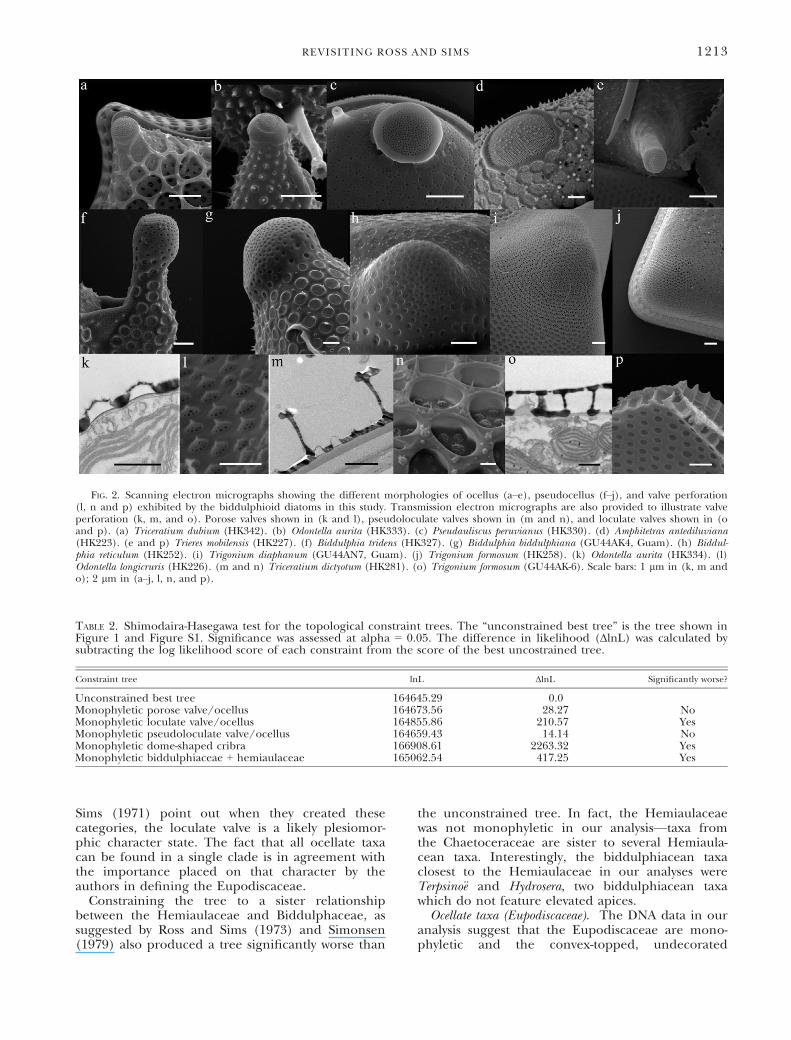
For the constraint analysis, the eupodiscoid and biddulphi-oid taxa used in this analysis were assigned groupings basedon Ross and Sims’ character categories (Table 1). Examplesof these characters can be found on Figure 2: ocelli (a–e),pseudocelli (f–j), porose valves (k and l), pseudoloculate po-rose valves (m and n), and loculate valves (o and p). We usedMesquite ver. 2.74 (Maddison and Maddison 2010) to createconstraint topologies reflecting previous classification hypoth-eses or grouping of taxa based on shared morphological fea-tures (e.g., valve poration and apical process—see Table 1).Using the same procedure as for the unconstrained tree, weperformed ML searches under six topological constraints(Table 2). We then found the best optimization for each ofthe constraint analyses and calculated the per-site log likeli-hoods of all alignment columns given the tree. Using thesefiles, we performed the Shimodaira-Hasegawa test (SH; Shi-modaira and Hasegawa 1999) between the best out of 1,024optimizations for each constraint and the best unconstrainedtree (Figs. 1 and S1).
RESULTS AND DISCUSSION
Phylogenetic analysis. In our combined three-geneanalysis, the Mediophyceae formed a clade (Fig. S1,ML bootstrap = 81%, posterior probability = 1.00)sister to the pennate diatoms (ML bs = 100%,pp = 1.00). The remaining radially symmetricalnon-pennate diatoms formed a basal “grade ofclades,” with clades of the genera LeptocylindrusCleve/Corethron Castracane (ML bs = 85%,pp = 1.00), Proboscia Sundstr€om (ML bs = 100%,pp = 1.00), Stephanopyxis/Paralia Heiberg/Hyalodis-cus Ehrenberg/Endictya Ehrenberg/AulacoseiraThwaites/Melosira C. Agardh (ML bs = 100%,pp = 1.00), Stellarima Hasle & P.A. Sims/Coscinodis-cus Ehrenberg/Palmerina G.R. Hasle plus Aulacodis-cus Ehrenberg/Actinoptychus Ehrenberg/ActinocyclusEhrenberg (ML bs = 100%, pp = 1.00), andRhizosolenia Brightwell/Pseudosolenia B.G. Sun-dstr€om/Guinardia H. Peragallo (ML bs = 100%,pp = 1.00).
Within the Mediophyceae, there are a number ofclades which received strong statistical support inboth Bayesian and ML analyses (Fig. 1). The Thalas-siosirales, Lithodesmidales and Cymatosiraceae areall strongly supported (ML bs = 100%, pp = 1.00).
Relationships between these clades, however, arenot as clear. For example, the internode betweenThalassiosirales plus Eunotogramma sp. and Lithode-smiales has high support in the Bayesian inference(pp = 1.00) but only modest ML support(bs = 77%). The internodes between the Cymatosir-aceae and the “Lithodesmiales/Eunotogramma + Tha-lassiosirales” clade are marked by very low MLsupport (bs = 52%).There are also a number of other mediophycean
clades that are strongly supported but for whichdiagnostic morphological features are not immedi-ately obvious (such as the bilabiate process of theLithodesmiales or the strutted process of the Thalas-siosirales). The two Attheya T. West species, forexample, are sister to the Biddulphia clade with fairlyhigh support values (ML bs = 98%, pp = 1.00). Theother taxa with setae-like apical projections, Chaetoc-eros muelleri Lemmermann and C. peruvianus Bright-well, can be found in a fairly well-supported (MLbs = 98%, pp = 1.00) clade with Acanthoceras H. Ho-nigmann, Urosolenia eriensis (H.L. Smith) F.E. Roundand R.M. Crawford, Terpsino€e musica Ehrenberg,Hydrosera G.C. Wallich, Dactyliosolen blavyanus(H. Peragallo) Hasle, and the polyphyletic Hemiaul-ales (Hemiaulus, Cerataulina Peragallo ex Sch€utt andEucampia in this analysis), though none of the rela-tionships between these taxa are well-supported.The remaining mediophycean taxa in the analysiscan be found in a fairly well-supported clade (MLbs = 86%, pp = 1.00), though relationships withinthe clade are also not well-supported. The “bifacialannulus” group (Medlin et al. 2008) consisting ofArdissonea spp. De Notaris, Climacosphenia elongatumJ.W. Bailey and Toxarium spp. J.W. Bailey is resolvedwithin this clade (ML bs = 100%, pp = 1.00), as arethe monophyletic Lampriscus spp (ML bs = 100%,pp = 1.00). The last clade in this group includesTrigonium formosum (Brightwell) Cleve, Chrysanthemo-discus floriatus A. Mann and Biddulphiopsis spp.,which are paraphyletic with respect to Isthmia min-ima Harvey & Bailey.The ocellus-bearing taxa form a strongly sup-
ported clade (ML bs = 100%, pp = 1.00), with theplanktonic, loculate taxa Trieres sinensis comb. nov,T. regia comb. nov. and T. mobiliensis comb. nov. sisterto the rest of the clade (ML bs = 100%, pp = 1.00).The remaining loculate, ocellate taxa (O. rhomboidesR. Jahn & Kusber, Pseudauliscus peruvianus [Kittonex Pritchard] A. Schmidt, Pleurosira laevis [Ehren-berg] Comp�ere) are more derived in the tree, withO. rhomboides and P. peruvianus forming a clade (MLbs = 100%, pp = 1.00) but little support for Pleuros-ira’s position.The porose, ocellate taxa are also not monophy-
letic. The disk-shaped Mastodiscus radiatus A.K.S.K.Prasad & J.A. Nienow is sister to the rest of the ocel-lates, except for the Trieres clade (ML bs = 100%,pp = 1.00). O. rostrata (Hustedt) Simonsen, Amphi-pentas pentacrinus Ehrenberg, and A. antediluviana
1210 MATT P. ASHWORTH ET AL.
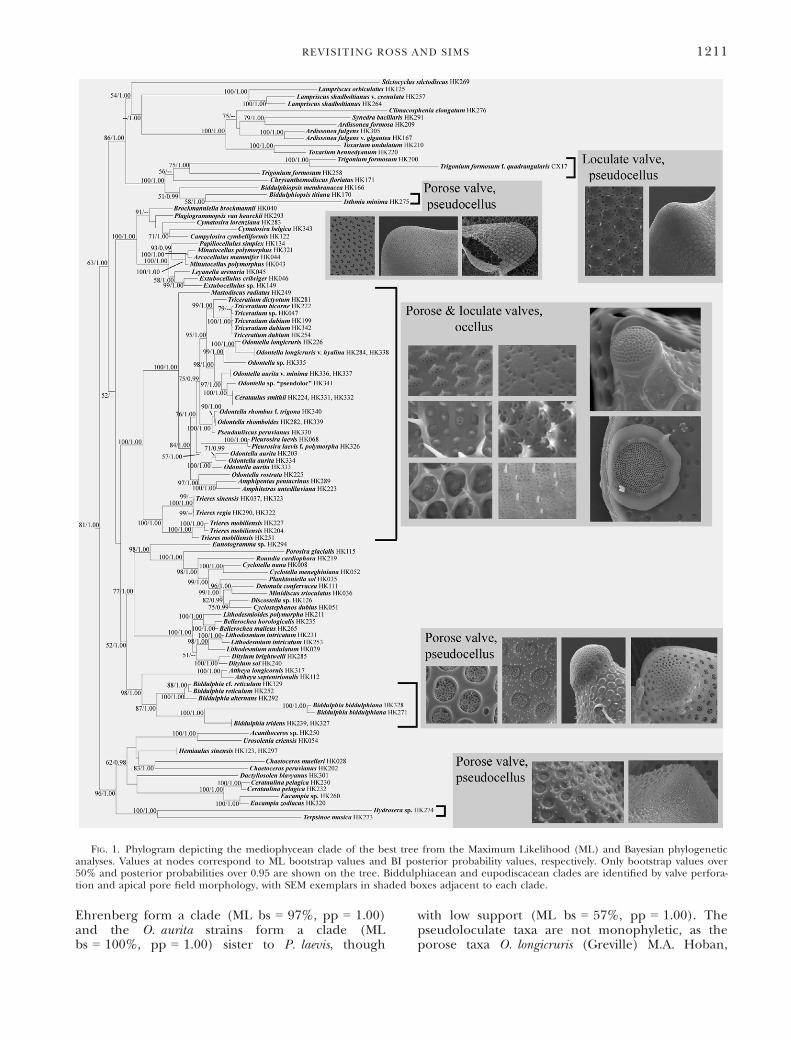
Ehrenberg form a clade (ML bs = 97%, pp = 1.00)and the O. aurita strains form a clade (MLbs = 100%, pp = 1.00) sister to P. laevis, though
with low support (ML bs = 57%, pp = 1.00). Thepseudoloculate taxa are not monophyletic, as theporose taxa O. longicruris (Greville) M.A. Hoban,
FIG. 1. Phylogram depicting the mediophycean clade of the best tree from the Maximum Likelihood (ML) and Bayesian phylogeneticanalyses. Values at nodes correspond to ML bootstrap values and BI posterior probability values, respectively. Only bootstrap values over50% and posterior probabilities over 0.95 are shown on the tree. Biddulphiacean and eupodiscacean clades are identified by valve perfora-tion and apical pore field morphology, with SEM exemplars in shaded boxes adjacent to each clade.
REVISITING ROSS AND SIMS 1211

O. longicruris v. hyalina (J.L.B. Schr€oder) M.A.Hoban and Odontella sp. “ECT3854” are sister to theCerataulus smithii Ralfs ex Pritchard/O. aurita v. min-ima (Grunow in Van Heurck) De Toni/Odontella sp.“pseudoloc” clade (ML bs = 98%, pp = 1.00), Thisgroup is sister to the clade containing T. dictyotum,T. bicorne Cleve, T. dubium Brightwell, and Tricera-tium sp. “CCMP147” (ML bs = 95%, pp = 1.00).Hypothesis testing. The results of the hypothesis
testing of trees for which valve perforation and api-cal pore field type were constrained to be monophy-letic are summarized in Table 2. Constraining the
pseudocellate taxa with porose valves lead to a treewhich was significantly worse than the uncon-strained tree, as the taxa with these characters (ex:Biddulphia, Isthmia, Terpsino€e) were split across threedistinct clades. Constraining the pseudocellate/locu-late valve condition to monophyly was not necessaryas the unconstrained search recovered the mono-phyly of taxa (Trigonium spp.) from this category.Among the constraint trees for the ocellate taxa,
only the tree constraining ocellate taxa with loculatevalves was significantly worse than the unconstrainedtree. This is not particularly surprising; as Ross and
TABLE 1. Biddulphioid taxa used in constraint analyses with assigned morphological character classes. For the “Valve Perfo-ration” character, an extra notation is made for taxa with “pseudoloculate valves”.
Taxon (voucher code HK###) Valve perforation Apical pore field
Biddulphia biddulphiana (HK271) Porose PseudocellusBiddulphia biddulphiana (HK328) Porose PseudocellusBiddulphia tridens (HK239) Porose PseudocellusBiddulphia tridens (HK327) Porose PseudocellusBiddulphia reticulata (HK252) Porose PseudocellusBiddulphia cf reticulata (HK329) Porose PseudocellusBiddulphia alternans (HK292) Porose PseudocellusBiddulphiopsis titiana (HK170) Porose PseudocellusIsthmia minima (HK275) Porose PseudocellusTerpsinoe musica (HK273) Porose PseudocellusHydrosera sp. (HK274) Porose PseudocellusTrigonium formosum (HK200) Loculate PseudocellusTrigonium formosum (HK258) Loculate PseudocellusTrigonium formosum f. quadrangularis (CX17) Loculate PseudocellusCerataulus smithii (HK224) Porose (pseudoloculate) OcellusCerataulus smithii (HK332) Porose (pseudoloculate) OcellusCerataulus smithii (HK331) Porose (pseudoloculate) OcellusPleurosira laevis (HK068) Loculate OcellusPleurosira laevis f. polymorpha (HK326) Loculate OcellusMastodiscus radiatus (HK249) Porose OcellusPseudauliscus peruvianus (HK330) Loculate OcellusOdontella aurita (HK203) Porose OcellusOdontella aurita (HK333) Porose OcellusOdontella aurita (HK334) Porose OcellusOdontella aurita v. minima (HK336) Porose (pseudoloculate) OcellusOdontella aurita v. minima (HK337) Porose (pseudoloculate) OcellusOdontella rostrata (HK225) Porose OcellusOdontella longicruris (HK226) Porose OcellusOdontella longicruris v. hyalina (HK284) Porose OcellusOdontella longicruris v. hyalina (HK338) Porose OcellusOdontella sp. (HK335) Porose OcellusOdontella sp. “pseudoloc” (HK341) Porose (pseudoloculate) OcellusOdontella rhombus f. trigona (HK340) Loculate OcellusOdontella rhomboides (HK339) Loculate OcellusOdontella rhomboides (HK282) Loculate OcellusAmphitetras antediluviana (HK223) Porose OcellusAmphipentas pentacrinus (HK289) Porose OcellusTriceratium bicorne (HK222) Porose (pseudoloculate) OcellusTriceratium dictyotum (HK281) Porose (pseudoloculate) OcellusTriceratium dubium (HK254) Porose (pseudoloculate) OcellusTriceratium dubium (HK342) Porose (pseudoloculate) OcellusTriceratium dubium (HK199) Porose (pseudoloculate) OcellusTriceratium sp. (HK047) Porose (pseudoloculate) OcellusTrieres mobilensis (HK204) Loculate OcellusTrieres mobilensis (HK227) Loculate OcellusTrieres mobilensis (HK251) Loculate OcellusTrieres regia (HK290) Loculate OcellusTrieres regia (HK322) Loculate OcellusTrieres sinensis (HK037) Loculate OcellusTrieres sinensis (HK323) Loculate Ocellus
1212 MATT P. ASHWORTH ET AL.
Sims (1971) point out when they created thesecategories, the loculate valve is a likely plesiomor-phic character state. The fact that all ocellate taxacan be found in a single clade is in agreement withthe importance placed on that character by theauthors in defining the Eupodiscaceae.
Constraining the tree to a sister relationshipbetween the Hemiaulaceae and Biddulphaceae, assuggested by Ross and Sims (1973) and Simonsen(1979) also produced a tree significantly worse than
the unconstrained tree. In fact, the Hemiaulaceaewas not monophyletic in our analysis—taxa fromthe Chaetoceraceae are sister to several Hemiaula-cean taxa. Interestingly, the biddulphiacean taxaclosest to the Hemiaulaceae in our analyses wereTerpsino€e and Hydrosera, two biddulphiacean taxawhich do not feature elevated apices.Ocellate taxa (Eupodiscaceae). The DNA data in our
analysis suggest that the Eupodiscaceae are mono-phyletic and the convex-topped, undecorated
FIG. 2. Scanning electron micrographs showing the different morphologies of ocellus (a–e), pseudocellus (f–j), and valve perforation(l, n and p) exhibited by the biddulphioid diatoms in this study. Transmission electron micrographs are also provided to illustrate valveperforation (k, m, and o). Porose valves shown in (k and l), pseudoloculate valves shown in (m and n), and loculate valves shown in (oand p). (a) Triceratium dubium (HK342). (b) Odontella aurita (HK333). (c) Pseudauliscus peruvianus (HK330). (d) Amphitetras antediluviana(HK223). (e and p) Trieres mobilensis (HK227). (f) Biddulphia tridens (HK327). (g) Biddulphia biddulphiana (GU44AK4, Guam). (h) Biddul-phia reticulum (HK252). (i) Trigonium diaphanum (GU44AN7, Guam). (j) Trigonium formosum (HK258). (k) Odontella aurita (HK334). (l)Odontella longicruris (HK226). (m and n) Triceratium dictyotum (HK281). (o) Trigonium formosum (GU44AK-6). Scale bars: 1 lm in (k, m ando); 2 lm in (a–j, l, n, and p).
TABLE 2. Shimodaira-Hasegawa test for the topological constraint trees. The “unconstrained best tree” is the tree shown inFigure 1 and Figure S1. Significance was assessed at alpha = 0.05. The difference in likelihood (ΔlnL) was calculated bysubtracting the log likelihood score of each constraint from the score of the best uncostrained tree.
Constraint tree �lnL ΔlnL Significantly worse?
Unconstrained best tree �164645.29 0.0Monophyletic porose valve/ocellus �164673.56 �28.27 NoMonophyletic loculate valve/ocellus �164855.86 �210.57 YesMonophyletic pseudoloculate valve/ocellus �164659.43 �14.14 NoMonophyletic dome-shaped cribra �166908.61 �2263.32 YesMonophyletic biddulphiaceae + hemiaulaceae �165062.54 �417.25 Yes
REVISITING ROSS AND SIMS 1213

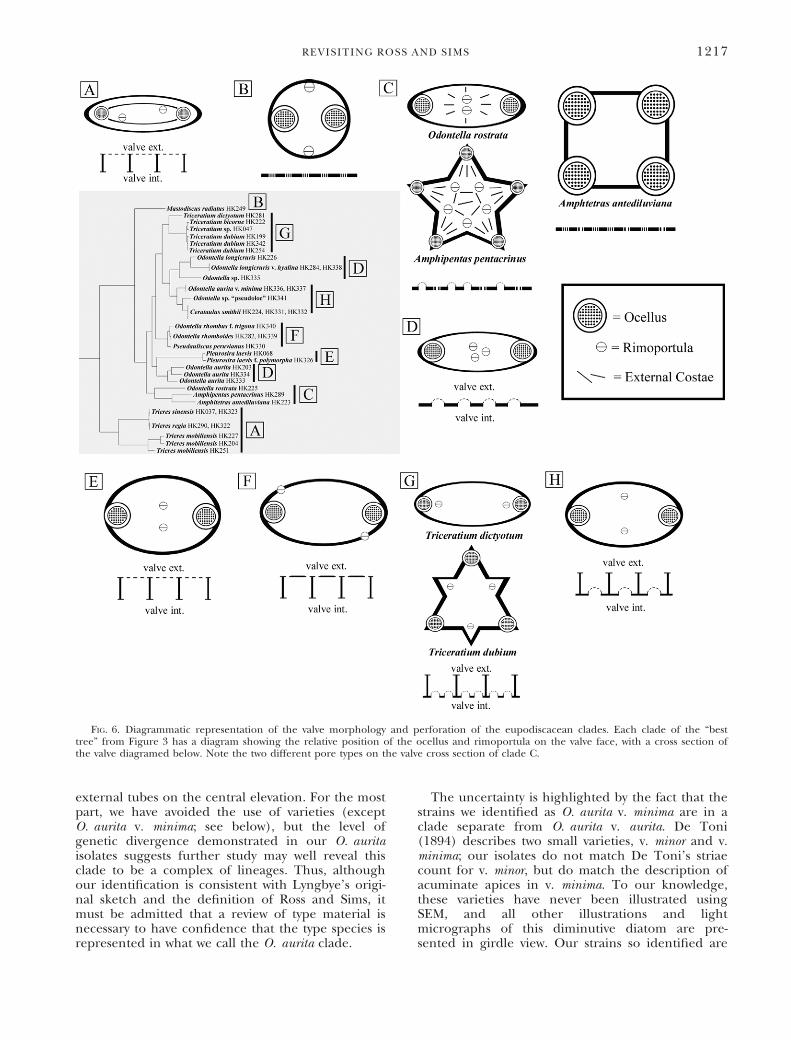
ocellus is likely a synapomorphy of that group, inagreement with the suggestion made by Ross andSims (1973). The relationship of this eupodiscaceanocellus to the cymatosiracean ocellulus is unclear atthis time on the basis of the characteristics of thisfeature alone. Each could be interpreted as inde-pendently derived from pseudocelli or the ocelluluscould be derived from the eupodiscacean ocellus.In combination with our molecular results, the for-mer interpretation is preferred. Although someauthors have referred to the apical pore field ofLampriscus spp. as either a pseudocellus or ocellus,we would suggest that it does not cleanly fit the defi-nition of either. There is indeed a hyaline rim sur-rounding the pores (unlike a pseudocellus), butpore size and density change drastically before thisrim (unlike the eupodiscacean ocellus). In addition,large linking spines are found attached to the pore-field rim (Fig. 3, a and b; Navarro 1981, Navarroand Lobban 2009), whereas eupodiscacean ocellihave structureless rims bordering the pore field.This molecular analysis also suggests that othercharacters proposed by Ross and Sims (1971) arenot entirely characteristic to the Eupodiscaceae; thedeflected valve mantle is absent in the Trieres clade(Fig. 4, a–c) and present in the pseudocellateBiddulphia clade (B. biddulphiana and B. tridens [Eh-renberg] Ehrenberg, Fig. 5, a and b), and the orien-tation of rimoportula (a character also discussed indetail by Simonsen 1979) across the eupodiscaceanclade is highly variable (Fig. 6).
Our results seem to support Ross and Sims’(1971) contention that almost every ocellate medio-phycean diatom with a porose valve is an Odontella,or none are. The taxa with a convex, undecoratedocellus formed a well-supported clade, which
includes the type species of Odontella, O. aurita,which is sister to P. laevis. Other genera makingOdontella paraphyletic include Triceratium, Amphite-tras, Amphipentas, Cerataulus, and Mastodiscus J.W.Bailey. Transferring all these diatoms to Odontellawould result in a monophyletic, but large andhighly diverse genus Odontella, by both morphology(Ross and Sims 1971) and DNA data.The alternative is to break Odontella, as presently
conceived, into smaller genera and to also retainexisting genera which are marked by synapomor-phies. For example, the pseudoloculate taxa formtwo clades—one with multiple pores with dome-shaped cribra per pseudolocular chamber (Fig. 7a)and one with a single pore with domed cribra perchamber (Fig. 7d). Both clades fits Sims and Ross’(1990) definition of the genus Triceratium—bearingpseudoloculate valves and elevated ocelli—but whilethe former clade (multiple pores) contains strainsthat might be considered “classical” Triceratium, thelater clade contains taxa that have never been con-sidered as Triceratium in their taxonomic record.This is likely an artifact of the historical idea thatTriceratium spp. were tri- or multi-angular in shape(Roper 1859, De Toni 1894, Sch€utt 1896), whereasthe single-pore clade strains are bipolar (Fig. 7, eand f) or round (Fig. 7g). At this point we shouldnote that the name attached to one of these strains,Odontella sp. HK341 (Fig. 7e), is merely provisional,and reflects the currently ambiguous nature of thetaxonomy of the genus Odontella.At this point it may be premature to simply sub-
sume all pseudoloculate taxa into Triceratium. Thenon-pseudoloculate O. longicruris (Fig. 8d) strainsform a clade between the two pseudoloculate clades(though constraining the pseudoloculate taxa to asingle clade results in a tree that is not significantlydifferent than the best tree; see Table 2). Morpho-logically, there is little to link O. longicruris to thepseudoloculate clades beyond possessing ocelli,external rimoportula tubes and domed cribra. Evendomed cribra are present in several ocellate clades,such as the O. aurita clade (Figs. 8a and 6d) andthe O. rostrata clade (Figs. 8, c, f; 6c). Constrainingthe domed-cribra taxa to a single clade results in atree that is significantly different from the “best,”unconstrained tree (Table 2).The immediate sister clade to the “domed cribra”
clade is also diagnosable by morphological charac-ters. Psuedauliscus peruvianus (Fig. 4g), O. rhomboides(Fig. 4h), and O. rhombus f. trigona (Fig. 4i), haverimoportulae positioned along the margin of thevalve face in a counter-clockwise rotation (ratherthan equidistant along the margin between ocelli),a condition unique to this group among the ocel-late taxa sampled here. Ross and Sims (1971)suggested that some taxa in this loculate/ocellategroup could be split from the genus Odontella,into Zygoceros (type = Z. rhombus Ehrenberg) andCerataulus, primarily distinguished by valve torsion
FIG. 3. Scanning electron micrographs illustrating the Lampris-cus taxa used in this study, with details of the apical pore fields(b and d). (a) Lampriscus shadboltianus v. crentulata (GU44I-4,Guam). (b) Lampriscus shadboltianus (HK264). (c and d)Lampriscus orbiculatus (HK125). Scale bars: 10 lm (a and c); 2 lm(b and d).
1214 MATT P. ASHWORTH ET AL.
(type = C. turgidus). Hoban (1979) disagreed, sug-gesting this would create a genus that included verydifferent morphologies, such as Z. rhombus and Tri-eres mobiliensis, and instead proposed “Zygocerae” asa section of Odontella. This morphological diversity,however, is mostly in valve outline, which is variableeven within species of the Eupodiscaceae; therefore,we agree with the Ross and Sims (1971) interpreta-tion where O. rhomboides and O. rhombus would bereturned to Zygoceros, with the planktonic loculate/ocellate taxa transferred to Trieres, which is dis-cussed separately below. Defining Zygoceros here,however, is potentially complicated by O. atlantica(Frenguelli) Sar, which has the same loculate valveconstruction as O. rhombus and O. rhomboides, buthas rimoportula immediately adjacent to the ocelli(Sar et al. 2007), rather than submarginally like thelatter taxa (O. rhomboides = Fig. 4h). Commentingon the genus Cerataulus with our current data is alsoproblematic: the genus is only represented byC. smithii in our data set, which is so radically differ-ent from the type C. turgidus in the pseudoloculatevalve (Fig. 7g) and lack of valve torsion that itshould likely be transferred to a different genusaltogether.
O. rostrata (Fig. 8f) shares valve characteristics ofthose taxa with which it is grouped by molecular
data: external ribs on the valve like the multipolarA. pentacrinus and two types of pore occlusions likeboth A. pentacrinus and the multipolar A. antediluvi-ana. (Fig. 8, c and b, respectively). Ross and Sims(1971) previously made a case for separating A. pent-acrinus and A. antediluviana from Triceratium on thebasis of the pseudoloculate valve, which the formertaxa lack. To this we add the molecular evidenceand the second pore occlusion type in the Amphite-tras/Amphipentas clade.Our proposed new genus, Trieres has clockwise
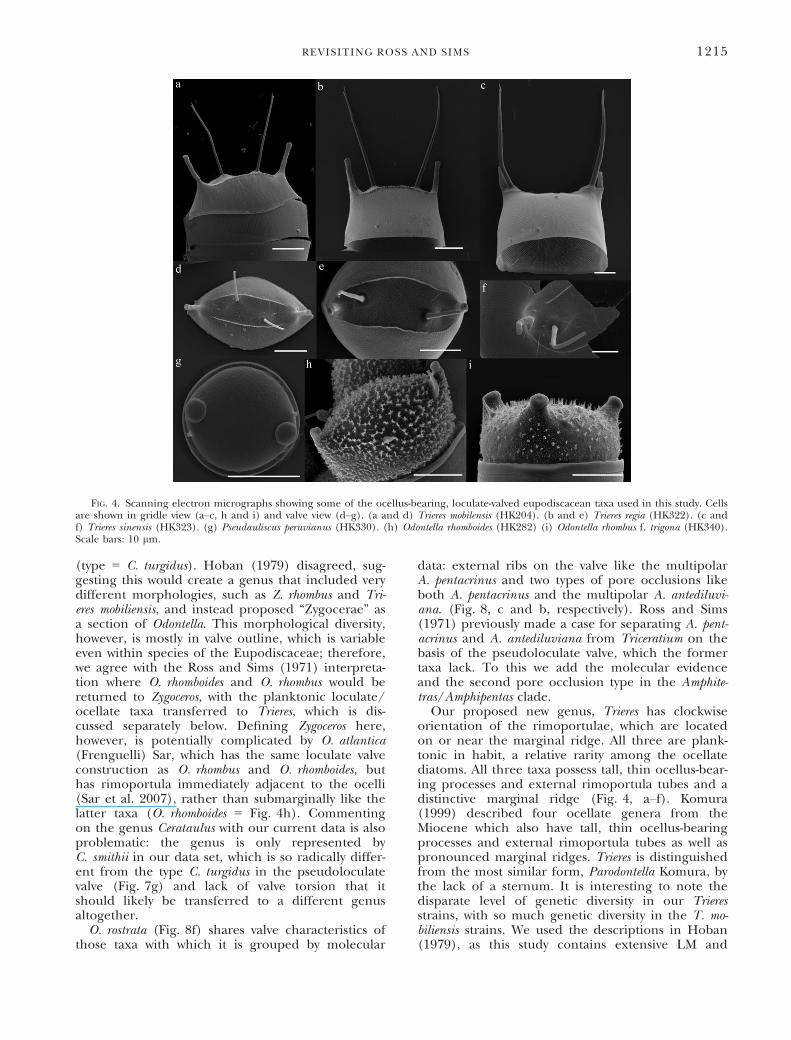
orientation of the rimoportulae, which are locatedon or near the marginal ridge. All three are plank-tonic in habit, a relative rarity among the ocellatediatoms. All three taxa possess tall, thin ocellus-bear-ing processes and external rimoportula tubes and adistinctive marginal ridge (Fig. 4, a–f). Komura(1999) described four ocellate genera from theMiocene which also have tall, thin ocellus-bearingprocesses and external rimoportula tubes as well aspronounced marginal ridges. Trieres is distinguishedfrom the most similar form, Parodontella Komura, bythe lack of a sternum. It is interesting to note thedisparate level of genetic diversity in our Trieresstrains, with so much genetic diversity in the T. mo-biliensis strains. We used the descriptions in Hoban(1979), as this study contains extensive LM and
FIG. 4. Scanning electron micrographs showing some of the ocellus-bearing, loculate-valved eupodiscacean taxa used in this study. Cellsare shown in gridle view (a–c, h and i) and valve view (d–g). (a and d) Trieres mobilensis (HK204). (b and e) Trieres regia (HK322). (c andf) Trieres sinensis (HK323). (g) Pseudauliscus peruvianus (HK330). (h) Odontella rhomboides (HK282) (i) Odontella rhombus f. trigona (HK340).Scale bars: 10 lm.
REVISITING ROSS AND SIMS 1215

SEM documentation of all three species we found,and all our T. mobiliensis isolates fit into Hoban’sdescription of that species. If there are several spe-cies represented in our T. mobiliensis isolates, the dif-ference is not immediately obvious and will requirefurther study. The apparent lack of genetic variationbetween T. sinensis and T. regia is not particularlysurprising; the two taxa share a very similar mor-phology, with the only difference being that therimoportula of T. sinensis are raised on the sameelevations as the ocellus-bearing processes while therimoportula of T. regia are adjacent but not co-occurring on the ocellate elevations (Hoban 1979).
Thus, most of the smaller clades recovered withmolecular data within the ocellate group seem toshare morphological characters (or are at leastunited by overall morphology). We suggest this sup-ports the notion that the ocellate taxa should bereorganized into several genera with a muchrestricted diagnosis of Odontella.
There is one significant exception, however. Thestrains we identified as O. aurita, the generitype,
share the externally deflected cribra morphology(Fig. 8a) of Triceratium and its allies, but the O. auri-ta strains are sister to P. laevis, which has externalcribra and internal foramina. There is also signifi-cant genetic variation among our O. aurita strains,whose identification should be considered provi-sional at this time. This is not a huge surprise, asthis diatom was described and illustrated in such away that many different diatoms fit into the descrip-tion. Lyngbye’s (1819) original sketch showed thefrustules in girdle view only, and the description waslimited to a description of the arrangements of chlo-roplasts and the “eared” state of the cell wall, nodoubt a reference to the ocelli. As a result, manymorphologies have been presented as “Odontellaaurita” throughout the literature (e.g., Peragallo andPeragallo 1898–1908, pl. 98, figs. 3–6, Navarro 1981,fig. 48, Witkowski et al. 2000, pl. 8, figs. 12, 13 andpl. 9, figs. 1–3) and several varieties of the specieshave been described. For this study, we have usedRoss and Sims’ characterization of O. aurita (1971):porose, undecorated valves with two to three
FIG. 5. Scanning electron micrographs showing some of the pseudocellus-bearing, porose-valved biddulphiacean taxa used in thisstudy. External views of the various valves are shown in (a–g), while the internal views can be seen in (h–l). (a and h) Biddulphia biddulphi-ana (HK271). (b and i) Biddulphia tridens (HK239). (c) Isthmia minima (GU44Y-8, Guam). (d) Terpsino€e musica (HK273). (e and j) Biddul-phia alternans (HK292). (f and k) Biddulphia reticulum (HK252). (g and l) Biddulphia cf reticulum (HK329). Scale bars: 20 lm (a); 10 lm(b–d and f–h); 5 lm (e and i–l).
1216 MATT P. ASHWORTH ET AL.
external tubes on the central elevation. For the mostpart, we have avoided the use of varieties (exceptO. aurita v. minima; see below), but the level ofgenetic divergence demonstrated in our O. auritaisolates suggests further study may well reveal thisclade to be a complex of lineages. Thus, althoughour identification is consistent with Lyngbye’s origi-nal sketch and the definition of Ross and Sims, itmust be admitted that a review of type material isnecessary to have confidence that the type species isrepresented in what we call the O. aurita clade.
The uncertainty is highlighted by the fact that thestrains we identified as O. aurita v. minima are in aclade separate from O. aurita v. aurita. De Toni(1894) describes two small varieties, v. minor and v.minima; our isolates do not match De Toni’s striaecount for v. minor, but do match the description ofacuminate apices in v. minima. To our knowledge,these varieties have never been illustrated usingSEM, and all other illustrations and lightmicrographs of this diminutive diatom are pre-sented in girdle view. Our strains so identified are
FIG. 6. Diagrammatic representation of the valve morphology and perforation of the eupodiscacean clades. Each clade of the “besttree” from Figure 3 has a diagram showing the relative position of the ocellus and rimoportula on the valve face, with a cross section ofthe valve diagramed below. Note the two different pore types on the valve cross section of clade C.
REVISITING ROSS AND SIMS 1217
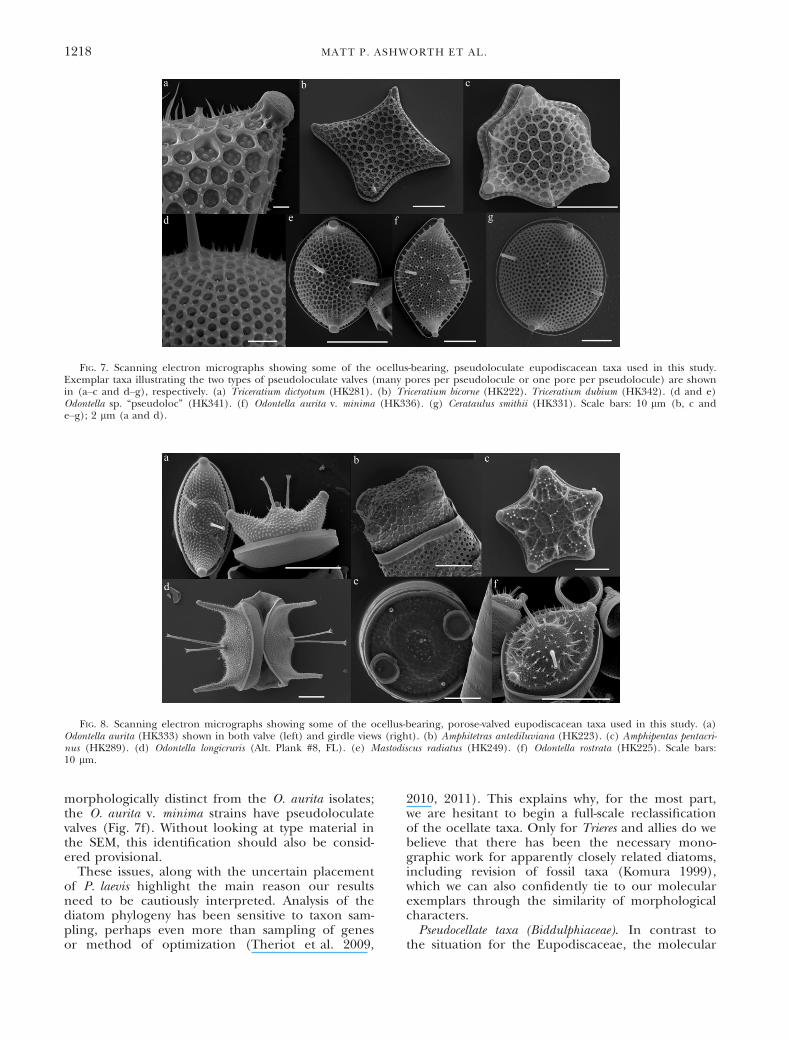
morphologically distinct from the O. aurita isolates;the O. aurita v. minima strains have pseudoloculatevalves (Fig. 7f). Without looking at type material inthe SEM, this identification should also be consid-ered provisional.
These issues, along with the uncertain placementof P. laevis highlight the main reason our resultsneed to be cautiously interpreted. Analysis of thediatom phylogeny has been sensitive to taxon sam-pling, perhaps even more than sampling of genesor method of optimization (Theriot et al. 2009,
2010, 2011). This explains why, for the most part,we are hesitant to begin a full-scale reclassificationof the ocellate taxa. Only for Trieres and allies do webelieve that there has been the necessary mono-graphic work for apparently closely related diatoms,including revision of fossil taxa (Komura 1999),which we can also confidently tie to our molecularexemplars through the similarity of morphologicalcharacters.Pseudocellate taxa (Biddulphiaceae). In contrast to
the situation for the Eupodiscaceae, the molecular
FIG. 7. Scanning electron micrographs showing some of the ocellus-bearing, pseudoloculate eupodiscacean taxa used in this study.Exemplar taxa illustrating the two types of pseudoloculate valves (many pores per pseudolocule or one pore per pseudolocule) are shownin (a–c and d–g), respectively. (a) Triceratium dictyotum (HK281). (b) Triceratium bicorne (HK222). Triceratium dubium (HK342). (d and e)Odontella sp. “pseudoloc” (HK341). (f) Odontella aurita v. minima (HK336). (g) Cerataulus smithii (HK331). Scale bars: 10 lm (b, c ande–g); 2 lm (a and d).
FIG. 8. Scanning electron micrographs showing some of the ocellus-bearing, porose-valved eupodiscacean taxa used in this study. (a)Odontella aurita (HK333) shown in both valve (left) and girdle views (right). (b) Amphitetras antediluviana (HK223). (c) Amphipentas pentacri-nus (HK289). (d) Odontella longicruris (Alt. Plank #8, FL). (e) Mastodiscus radiatus (HK249). (f) Odontella rostrata (HK225). Scale bars:10 lm.
1218 MATT P. ASHWORTH ET AL.

results do not support a monophyletic Biddulphia-ceae. Biddulphia spp. form a clade which is sister toAttheya and several nodes away from the clades bear-ing other taxa often associated with the Biddulphia-ceae such as Trigonium, Eunotogramma, Isthmia, andTerpsino€e, which themselves are scattered across theMediophyceae. Ross and Sims (1971) concept of apseudocellate Biddulphiaceae was defined in part bypossession of a straight valve mantle, but some Bid-dulphia species themselves have a deflected valvemantle characteristic of most Eupodiscaceae (Fig. 5,a and b). Other characters suggested by Ross andSims (1971) were already highly variable within theirBiddulphiaceae, such as internal ribs or ridges,which are present in B. biddulphiana (Fig. 5, a andh), B. alternans (J.W. Bailey) Van Heurck (Fig. 5, eand j) and Terpsino€e musica but absent in B. reticu-lum (Ehrenberg) C.S. Boyer (Fig. 5, f, g, k, and l)and I. minima (Fig. 5c). The internal ribs alsoappear to be missing in B. tridens (Fig. 5, b and i);while there is a hyaline area on the interior of thevalve where the valve face is depressed, this depres-sion does not appear to extend into the interiorspace as it does in the ribs of the aforementionedtaxa.
With so many other mediophycidean lineagesrepresented solely by fossil taxa and unavailable forthis study, it is difficult to draw conclusions aboutthe relationships of the Biddulphiaceae andEupodiscaeae to the rest of the diatom tree at thistime. However, as mentioned previously, severalauthors have proposed a close relationship of theBiddulphiaceae and the Hemiaulaceae (Ross et al.1977, Glezer 1979, Simonsen 1979). This proposalwas based on the fact that Biddulphiaceae andHemiaulaceae share the feature of strongly ele-vated apices on their valves; Hemiaulus has hollowtubes at the apices while Eucampia and Cerataulinafeature costate pore fields at the end of the eleva-tions. Not only do we recover the Biddulphiaceaeas polyphyletic, our analyses also recover the Chae-toceraceae embedded in a paraphyletic Hemiaula-ceae, so a sister group relationship betweenBiddulphiaceae and Hemiaulaceae to the exclusionof Chaetoceraceae and other taxa is rejected bythe topological constraint test using per-site loglikelihoods (Table 2).
Smaller groups of former Biddulphiaceae, how-ever, seem well supported by both molecular andmorphological data. In particular, Ross and Sims’generic concept of Biddulphia holds up very wellwith molecular data. Isolates identified as Biddul-phia in this data set (B. alternans, B. biddulphiana,B. reticulum, and B. tridens) all have pseudocelliand similarly occluded simple pores across thevalve. Internal ribs, another character Ross andSims associated with this genus, were only foundin two of the taxa: the aforementioned B. alternansand B. biddulphiana. The authors mention T. reticu-lum Ehrenberg as likely belonging to this genus
despite the lack of internal ribs, which is sup-ported by the molecular data; this taxon bears themore appropriate name B. reticulum in our analysisand should not be confused with the ocellate/pseudoloculate B. reticulata Roper (see Sims andRoss 1990 for more detail on this taxonomicissue). However, at the moment we cannot suggesta synapomorphy for this taxon based on the pre-sented data.The genus Biddulphiopsis has been associated with
the Biddulphiaceae (Round et al. 1990) basedmostly on its taxonomic history, but Biddulphiopsistitiana (Grunow) von Stosch & Simonsen andB. membranacea (Cleve) von Stosch & Simonsen dif-fer by the presence (B. titiana) or absence (B. mem-branacea) of pseudocelli (von Stosch and Simonsen1984). In our analysis, I. minima is embedded in Bid-dulphiopsis, sharing marginal rimoportulae andpseudocelli with its sister taxon, B. titiana. I. minimais on a long branch in our molecular tree, and alsoseems to be highly derived morphologically, havingautapomorphic characters such as the presence ofonly a single pseudocellus per cell and deep, sac-likedepressions on the valve mantle. The latter charac-ter, however, is unique to I. minima and is notfound in other Isthmia species (Andresen 1995).Both genera require further molecular and morpho-logical analysis.That the freshwater taxa Terpsino€e musica and
Hydrosera sp. form a clade is hardly surprising, con-sidering the undulate rimoportula both taxa shareas an apparent synapomorphy. Both taxa do featureapical pore fields which may not be considered“pseudocelli,” as they are not elevated above thevalve surface (Fig. 5d).The generic concept for Trigonium also held up
strongly with molecular data, although that couldwell be an artifact for poor taxon sampling withonly three strains on the analysis—all of them nomi-nally agreeing with the admittedly vague speciesdiagnosis of T. formosum (Fig. 9) originally providedby Brightwell (described under the name T. striola-tum Ehrenberg in 1853, then renamed T. formosumBrightwell in 1856). Hendey (1971) and Ross andSims (1971) both note that only slight difference inthe valve perforation (density at the center of thevalve and degree of velum convexity) exists betweenthe few Trigonium species each examined and com-pared (T. arcticum Brightwell, T. formosum, andT. graeffeanum [Witt] Hendey in Hendey; T. arcti-cum, T. formosum, and T. montereyi Brightwell in Rossand Sims). Close examination by SEM of the typematerial for Trigonium species should be a prioritymoving forward. In addition, the placement of Trigo-nium in the diatom phylogeny as a whole remainsuncertain. We recover it as sister to Chrysanthemodis-cus (with low statistical support). The latter lacks theloculate areolae and apices of Trigonium, and has acentral pore field instead of pore fields on elevatedapices.
REVISITING ROSS AND SIMS 1219

CONCLUSIONS
In a plea for improved illustration included in sci-entific texts, Wallich (1858) used Triceratium as anexemplar of a genus potentially undone by impro-per documentation of characters. We would arguethat his prophecy is at least partly fulfilled. The phy-logeny of the Biddulphiaceae and Eupodiscaceaeappears to be a case where the use of molecularcharacters does not so much “disagree” with mor-phological characters as they indicate uncertaintypresent in both types of data. On one hand, difficul-ties in interpretation of morphology, may lead toreliance on plesiomorphy to delineate taxa or to dis-count “minutiae” as synapomorphies in their ownright. On the other hand, although much easier tointerpret, molecular data suffer from higher pro-pensity for convergent change and therefore per-haps higher risk to mislead phylogenetic inference.It is evident that more work is needed to resolve theuncertainties uncovered to date or identify conflict-ing inferences that withstand statistical scrutiny (asin lizards: Losos et al. 2012).
Ross and Sims (1971) focused primarily on theapical pore fields (ocellus vs. pseudocellus) as theprimary character to distinguish the Biddulphiaceaeand Eupodiscaceae, and suggested several otherobserved valvar characters to support this distinction.They also suggested (1973) that the pseudocellus
gave rise to the eupodiscoid ocellus, making thepseudocellus plesiomorphic. Our molecular result,however, does not seem to support this interpreta-tion in its entirety, although it supports the hypothe-sis that the eupodiscoid ocellus is synapomorphic.Thus, our analysis does not agree with Glezer’s asser-tion that the eupodiscoid ocellus evolved severaltimes from the pseudocellus (1979).This study is an important step in laying the
groundwork for future systematic studies of theMediophyceae. With the most complete taxonsampling to date, the monophyly of the ocellate Eu-podiscaceae seems well supported. Some groupswithin the Eupodiscaceae appear to be well sup-ported (e.g., Triceratium). Odontella, however, is non-monophyletic. It is scattered among several molecu-lar lineages each of which, in our preliminary analy-ses, are at least phenetically coherent if notsupported by synapomorphy. Likewise the Biddul-phiaceae, formerly diagnosed only by presumedplesiomorphy, appears to be non-monophyletic withgroups scattered across the Mediophyceae. Someclades of former Biddulphiaceae again appear to bephenetically coherent if not diagnosable by synapo-morphy.Despite the new molecular analyses of many medi-
ophytes presented here, there are still many taxawhich need to be sampled. The variety of phyloge-netic placement of the pseudocellate lineages in thisstudy underscores the massive undertaking requiredfor a cladistic, morphological analysis on the fossiland extant Mediophyceae. It will be difficult toselect exemplars, as the “biddulphioid” state actuallyis present on many different branches of the diatomtree. Nevertheless, the agreement of the moleculardata with a synapomorphic, eupodiscacean ocellussuggests that the situation is not as chaotic as itmight seem.Nomenclatural amendments. On the basis of the
conservation of morphological and molecular char-acters discussed above, we recommend the followingtaxa be transferred to the genus Trieres gen. nov.:Trieres M.P. Ashworth & E.C. Theriot, gen. nov.Description: Frustules with large pervalvar distance,
one to two times the transapical distance. Valvesalveolate, with finely perforate vela arrayed in box-like sections and single, circular foramen on thevalve interior. Valves elongate with tall ocelli at api-ces and offset, opposed rimoportula with conspicu-ous external tubes on the valve face. Valve marginedged with a wall-like projection running betweenthe ocelli. Cells planktonic in habit.Etymology: From the Greek “tri�er�es,” a type of
ancient Greek sailing vessel which resembles thevalves of this genus seen in girdle view.Type species: Trieres sinensis (Greville) M.P. Ash-
worth & E.C. Theriot, illustrated in Williams 1988;pl. 25, fig. 3.Trieres mobiliensis (J.W. Bailey) Ashworth and
Theriot comb. nov.
FIG. 9. Scanning electron micrographs showing the pseudocellus-bearing, loculate-valved biddulphiacean taxa used in this study:Trigonium formosum. (a) T. formosum (HK258) shown in valveview. (b) T. formosum (HK258) shown in girdle view. Scale bars:10 lm.
1220 MATT P. ASHWORTH ET AL.

Basionym: Zygoceros mobiliensis Bailey 1851, p. 40,pl. 2: figs 34–35
Homotypic synonyms: Biddulphia mobiliensis (J.W.Bailey) Grunow 1882 (in Van Heurck), pl. 101: figs.4–6
Odontella mobiliensis (J.W. Bailey) Grunow 1884,p. 58
Trieres regia (M. Schultze) M.P. Ashworth & E.C.Theriot comb. nov.
Basionym: Denticella regia Schultze 1859, p. 21,pl. 2: figs. 11, 12
Homotypic synonyms: Biddulphia regia (M. Schultze)Ostenfeld 1908, p. 7, fig. 3
Odontella regia (M. Schultze) Simonsen 1974, p. 27Trieres sinensis (Greville) M.P. Ashworth & E.C.
Theriot comb. nov.Basionym: Biddulphia sinensis Greville 1866, p. 81,
pl. 9: fig. 16Homotypic synonyms: Odontella sinensis (Greville)
Grunow 1884, p. 58
The authors wish to thank the following people for collectionof live material or facilitating the collection of live material:H. Allali, A. Alverson, C. Aumack, M. Brady, T. Frankovich, M.Gruenstaeudl, P. Hargraves, S. Kanno, D. Lam, C. Lobban, E.Ruck, E. Salomaki, M. Schefter, and M. Yu. We wish to thank D.Romanovicz for assistance with electron microscopy and theTexas Advanced Computing Center (TACC) for access to theRANGER and LONESTAR supercomputers. We also wish tothank J. Witkowski and an anonymous reviewer for their help-ful discussions and comments in editing this manuscript. Thisresearch was supported by an NSF AToL grant to ECT (EF0629410) and the Blumberg Fellowship to ECT.
Adl, S. M., Simpson, A. G. B., Farmer, M. A., Andersen, R. A.,Anderson, O. R., Barta, J. R., Bowser, S. S. et al. 2005. Thenew higher level classification of eukaryotes with emphasison the taxonomy of protists. J. Eukaryot. Microbiol. 52:399–451.
Andresen, N. A. 1995. A taxonomic investigation of a little knownspecies of Isthmia. In Kociolek, J. P. & Sullivan, M. J. [Eds.] ACentrury of Diatom Research in North America: A Tribute to theDistinguished Careers of Charles W. Reimer and Ruth Patrick. Koe-ltz Scientific Books USA, Champaign, Illinois, pp. 103–10.
Bailey, J. W. 1851. Microscopical observations made in South Car-olina, Georgia and Florida. Smithson. Contrib. Knowl. 2:1–48.
Brightwell, T. 1853. On the genus Triceratium, with descriptionsand figures of the species. Quart. J. Micr. Sci. 1:245–52.
Brightwell, T. 1856. Further observations on the genus Tricera-tium, with descriptions and figures of new species. Quart.J. Micr. Sci. 4:272–6.
De Toni, G. B. 1894. Sylloge algarum omnium hucusque cognitarum.Vol. II. Bacillarieae; sectio III. Cryptoraphideae. Typis Seminarrii,Patavii, pp. 819–1556.
Deschamps, P., Lara, E., Marande, W., L�opez-Garc�ıa, P., Ekelund,F. & Moreira, D. 2011. Phylogenomic analysis of kinetoplast-ids supports that trypanosomatids arose from within bodo-nids. Mol. Biol. Evol. 28:53–8.
Embley, T. M., Finlay, B. J., Dyal, P. L., Hirt, R. P., Wilkinson, M.& Williams, A. G. 1995. Multiple origins of anaerobic ciliateswith hydrogenosomes within the radiation of aerobic ciliates.Proc. R. Soc. B 262:87–93.
Glezer, Z. I. 1979. Evolution and systematics of the order Biddul-phiales (Bacillariophyta). Paleontol. J. 13:103–11 (translatedfrom Russian).
Greville, R. K. 1866. Descriptions of new and rare diatoms. SeriesXIX. Trans. Microsc. Soc. Lond. 14:77–86.
Guillard, R. R. L. 1975. Culture of phytoplankton for feedingmarine invertebrates. In Smith, W. L. & Chanley, M. H.[Eds.] Culture of marine invertebrate animals. Plenum Press,New York, pp. 26–60.
Hasle, G. R., von Stosch, H. A. & Syvertsen, E. E. 1983. Cymatosir-aceae, a new diatom family. Bacillaria 6:9–156.
Hendey, N. I. 1971. Some marine diatoms from the GalapagosIslands. Nova Hedwigia 22:371–422.
Hoban, M. A. 1979. Morphology, life histories and systematics ofselected genera of biddulphioid diatoms. PhD dissertation,University of Texas at Austin, Austin, Texas.
Komura, S. 1999. Barrel-shaped diatoms from the MioceneNabuto Formation, central Japan. Diatom 15:51–78.
Lobban, C. S., Ashworth, M. P., Arai, Y., Jordan, R. W. & Theriot,E. C. 2011. Marine necklace-chain Fragilariaceae (Bacillario-phyceae) from Guam, including descriptions of Koernerellaand Perideraion, genera nova. Phycol. Res. 59:174–93.
Losos, J. B., Hillis, D. M. & Greene, H. W. 2012. Who speaks witha forked tongue? Science 338:1428–9.
Lyngbye, H. C. 1819. Tentamen Hydrophytologiae Danicae Con-tinens omnia Hydrophyta Cryptogama Daniae, Holsatiae,Faeroae, Islandiae, Groenlandiae hucusque cognita, Systema-tice Disposita, Descripta et iconibus illustrata, Adjectis SimulSpeciebus Norvegicis. Hafniae 248 pp. 70 pls.
Medlin, L. K. 2010. Pursuit of a natural classification of diatoms:an incorrect comparison of published data. Eur. J. Phycol.45:155–66.
Medlin, L. K. & Kaczmarska, I. 2004. Evolution of the diatoms: V.Morphological and cytological support for the major cladesand a taxonomic revision. Phycologia 43:245–70.
Medlin, L. K., Sato, S., Mann, D. G. & Kooistra, W. H. C. F. 2008.Molecular evidence confirms sister relationship of Ardissonea,Climacosphenis and Toxarium with the bipolar centric diatoms(Bacillariophyta, Mediophyceae), and cladistics analyses con-firm that extremely elongated shape has arisen twice in thediatoms. J. Phycol. 44:1340–8.
Navarro, J. N. 1981. A survey of the marine diatoms of PuertoRico II. Suborder Biddulphiineae: families Biddulphiaceae,Lithodesmiaceae and Eupodiscaceae. Bot. Mar. 24:615–30.
Navarro, J. N. & Lobban, C. S. 2009. Freshwater and marine dia-toms from the western Pacific islands of Yap and Guam, withnotes on some diatoms in damselfish territories. Diatom Res.24:123–57.
Nikolaev, V. A. 1990. The system of centric diatoms. In Simola,H. [Ed.] Proceedings of the Tenth International Diatom Sympo-sium, Joensuu, Finland, August 28–September 2, 1988. KoeltzScientific Books, Koenigstein, Germany, pp. 103–10.
Ostenfeld, C.H. 1908. On the immigration of Biddulphia sinensisGrev. and its occurrence in the North Sea during 1903–1907and on its use for the study of the direction and rate of flowof the currents. Meddelelser fra Kommissionen for Danmarks Fisk-eri-Og Havundersøgelser. Serie: Plankton (København) 1:1–44.
Peragallo, H. & Peragallo, M. 1898–1908. Diatome′es marine deFrance et des districts maritimes voisins. M. J. Tempe‘re, Grez-sur-Loing, France, 491 pp. + 137 pl.
Roper, F. C. S. 1859. On the genus Biddulphia and its affinities.Trans. Microsc. Soc. Lond. 7:1–24.
Ross, R. & Sims, P. A. 1971. Generic limits in the Biddulphiaceaeas indicated in the scanning electron microscope. In Hey-wood, V. H. [Ed.] Scanning Electron Microscopy, Systematic andEvolutionary Application. Academic Press, London, pp. 155–77.
Ross, R. & Sims, P. A. 1973. Observations on family and genericlimits in the Centrales. Nova Hedwigia, Beih. 45:97–132.
Ross, R. & Sims, P. A. 1987. Further genera of the Biddulphiaceae(diatoms) with interlocking linking spines. Bull. Br. Mus.(Nat. Hist.) Bot. 16:269–311.
Ross, R., Sims, P. A. & Hasle, G. R. 1977. Observations on somespecies of Hemiauloideae. Nova Hedwigia, Beih. 54:179–203.
Round, F. E., Crawford, R. M. & Mann, D. G. 1990. The Diatoms:Biology and Morphology of the Genera. Cambridge UniversityPress, Cambridge, UK, 747 pp.
Ruck, E. C. & Theriot, E. C. 2011. Origin and evolution of thecanal raphe system in diatoms. Protist 162:723–37.
REVISITING ROSS AND SIMS 1221

Sar, E. A., Sunesen, I., Jahn, R., Kusber, W. H. & Lavigne, A. S.2007. Revision of Odontella atlantica (Frenguelli) Sar comb. etstat. nov. with comparison to two related species, O. rhombus(Ehrenb.) K€utz. and O. rhomboides R. Jahn et Kusber. DiatomRes. 22:341–53.
Schultze, M. 1859. Phenomena of internal movements inDiatomaceae of the North Sea, belonging to the generaCoscinodiscus, Denticella, Rhizosolenia. Quart. J. Micr. Sc. 7:13–21.
Sch€utt, F. 1896. Bacillariales (Diatomeae). In Engler, A. & Prantl,K. [Eds.] Die naturlichen Pflanzenfamilien, Vol. I(1b). Verlagvon Wilhelm Engelmann, Leipzig, pp. 31–150.
Shimodaira, H. & Hasegawa, M. 1999. Multiple comparisons oflog-likelihoods with applications to phylogenetic inference.Mol. Biol. Evol. 16:1114–6.
Simonsen, R. 1974. The diatom plankton of the IndianOcean Expedition of R/V Meteor 1964-5. “Meteor” For-schungsergebnisse. Reihe D: Biologie 19:1–107.
Simonsen, R. 1979. The diatom system: ideas on phylogeny.Bacillaria 2:9–71.
Sims, P. A. & Ross, R. 1990. Triceratium pulvinar and T. unguicula-tum, two confused species. Diatom Res. 5:155–69.
Sperling, E. A., Peterson, K. J. & Pisani, D. 2009. Phylogenetic-sig-nal dissection of nuclear housekeeping genes supports theparaphyly of sponges and the monophyly of Eumetazoa. Mol.Biol. Evol. 26:2261–74.
von Stosch, H. A. & Simonsen, R. 1984. Biddulphiopsis, a newgenus of the Biddulphiaceae. Bacillaria 7:9–36.
Theriot, E. C., Ashworth, M., Ruck, E., Nakov, T. & Jansen, R. K.2010. A preliminary multigene phylogeny of the diatoms(Bacillariophyta): challenges for future research. Plant Ecol.Evol. 143:1–18.
Theriot, E. C., Cannone, J. J., Gutell, R. R. & Alverson, A. J. 2009.The limits of nuclear-encoded SSU rDNA for resolving thediatom phylogeny. Eur. J. Phycol. 44:277–90.
Theriot, E. C., Ruck, E., Ashworth, M., Nakov, T. & Jansen, R. K.2011. Status of the pursuit of the diatom phylogeny. InSeckbach, J. & Kociolek, J. P. [Eds.] The Diatom World.Springer Science+Business Media Springer, New York, NY,pp. 119–42.
Van Heurck, H. 1882. Synopsis des Diatom�ees de Belgique. Jucaju andCie, Anvers.
Wallich, G. C. 1858. On Triceratium and some new allied forms.Quart. J. Micr. Sc. 6:242–53.
Wilgenbusch, J. C., Warren, D. L. & Swofford, D. L. 2004. AWTY:A system for graphical exploration of MCMC convergence inBayesian phylogenetic inference. Available at: http://ceb.csit.fsu.edu/awty.
Williams, D. M. 1988. An illustrated catalogue of the typespecimens in the Greville diatom herbarium. Bull. Br. Mus.(Nat. Hist.) Bot. 18:1–148.
Williams, D. M. & Kociolek, J. P. 2007. Pursuit of a natural classifi-cation of diatoms: history, monophyly and the rejection ofparaphyletic taxa. Eur. J. Phycol. 42:313–9.
Witkowski, A., Lange-Bertalot, H. & Metzelin, D. 2000. DiatomFlora of Marine Coasts I. A.R.G. Gantner Verlag K.G, Ruggell,Germany, 925 pp.
Supporting Information
Additional Supporting Information may befound in the online version of this article at thepublisher’s web site:
Figure S1. Phylogram depicting the best treefrom the Maximum Likelihood and Bayesian phy-logenetic analyses. Nodes with double asterisks(**) denote clades with ≥100% bootstrap supportand ≥1.00 posterior probability, while nodes withsingle asterisks (*) denote clades with ≥75% boot-strap support and ≥0.95 posterior probability. Themediophycean clade is collapsed here for clarityand shown in Figure 1.
Table S1. Taxa used in phylogenetic analysis,with locality, Theriot Lab specimen voucher ID(HK###) and GenBank accession numbers. Taxamarked with asterisks were used in Theriot et al.(2010) but were misidentified in that manuscript(* = “Biddulphia alterans”; ** = “Isthmia enervis”;*** = “Coscinodiscus concinniformis”).
Appendix S1. Aligned DNA sequence data inNEXUS format. Sequences for the 3 genes ofeach taxon are sequential, starting with nuclear-encoded ribosomal small-subunit (bases 1–1,706),chloroplast-encoded rbcL (bases 1,707–3,178) andchloroplast-encoded psbC (bases 3,179–4,238).
1222 MATT P. ASHWORTH ET AL.




















