
Review Article
14
ENDOCRINE PRACTICE Vol 19 No. 3 May/June 2013 497 Review Article Elena Barengolts, MD Submitted for publication August 21, 2012 Accepted for publication December 12, 2012 From the Section of Endocrinology, Department of Medicine, University of Illinois Medical Center and Jesse Brown VA Medical Center, Chicago, Illinois. Address correspondence to Dr. Elena Barengolts, Department of Medicine, Section of Endocrinology, MC 640, 1819 West Polk Street, Chicago, IL 60612. E-mail: [email protected]. Published as a Rapid Electronic Article in Press at http://www.endocrine practice.org on January 21, 2013. DOI:10.4158/EP12263.RA To purchase reprints of this article, please visit: www.aace.com/reprints. Copyright © 2013 AACE. ABSTRACT Objective: To review the role of human large bowel microbacteria (microbiota) in the glucose homeostasis, to address vitamin D (VD) and prebiotics interactions with microbiota, and to summarize recent randomized clinical trials (RCTs) of VD and prebiotics supplementation in pre- diabetes (PreDM) and type 2 diabetes mellitus (T2DM). Methods: Primary literature was reviewed in the fol- lowing areas: composition and activity of human micro- biota associated with PreDM and T2DM, interactions between microbiota and glucose homeostasis, the interac- tion of microbiota with VD/prebiotics, and RCTs of VD/ prebiotics in subjects with PreDM or T2DM. Results: The human microbiota is comprised of 100 trillion bacteria with an aggregate genome that is 150-fold larger than the human genome. Data from the animal mod- els and human studies reveal that an “obesogenic” diet results into the initial event of microbiota transformation from symbiosis to dysbiosis. The microbial antigens, such as Gram(-) bacteria and lipopolysaccharide (LPS), trans- locate to the host interior and trigger increased energy harvesting and Toll-like receptor (TLR) activation with VXEVHTXHQW LQÁDPPDWRU\ SDWKZD\V VLJQDOLQJ 7KH ´GRXEOH hit” of steatosis (ectopic fat accumulation) and “–itis” LQÁDPPDWLRQ DQG FRQWULEXWLRQ RI ´FRULVNVµ HJ YLWDPLQ ' GHÀFLHQF\ >9''@ DUH UHTXLUHG WR DFWLYDWH PROHFXODU signaling, including impaired insulin signaling and secre- tion, that ends with T2DM and associated diseases. Dietary changes (e.g., prebiotics, VD supplementation) may ame- liorate this process if initiated prior to the process becom- ing irreversible. Conclusion: Emerging evidence suggests an impor- tant role of microbiota in glucose homeostasis. VD sup- plementation and prebiotics may be useful in managing PreDM and T2DM. (Endocr Pract. 2013;19:497-510) Abbreviations: 1,25D = 1,25-dixydroxyvitamin D; ApoE = apolipopro- tein E; GutM = gut (large bowel) microbiota; HOMA = homeostasis model of assessment; LPS = lipopoly- saccharide; PreDM = prediabetes; RCT = randomized clinical trial; TLR = toll-like receptors; TNF-_ = tumor necrosis factor-_; VD = vitamin D; VDD = vitamin D GHÀFLHQF\ VDR = vitamin D receptor INTRODUCTION The process of deterioration of normal glucose toler- ance into prediabetes (PreDM) and type 2 diabetes melli- tus (T2DM) involves the contribution of “nature,” such as LQWHUQDO QRQPRGLÀDEOH IDFWRUV JHQHWLF EDFNJURXQG DJH DQG JHQGHU DQG ´QXUWXUHµ ZKLFK DUH H[WHUQDO PRGLÀDEOH factors. Among the external risk factors, the western diet KLJK IDW REHVLW\SURQH DQG YLWDPLQ ' 9' GHÀFLHQF\ (VDD) are important (1,2). Emerging evidence suggests WKDW DQRWKHU LPSRUWDQW H[WHUQDO LQÁXHQFH RQ WKH SDWKR- physiology of glucose homeostasis may be the large bowel (gut) microbial community (microbiota) (3-10). Over the past 10 years, there has been an explosion of knowl- edge; the number of published papers; PubMed articles with the key word “microbiota” increased exponentially from 121 (2002) to 2,234 (2012), and those with the key
Transcript of Review Article

ENDOCRINE PRACTICE Vol 19 No. 3 May/June 2013 497
Review Article
Elena Barengolts, MD
Submitted for publication August 21, 2012
Accepted for publication December 12, 2012
From the Section of Endocrinology, Department of Medicine, University
of Illinois Medical Center and Jesse Brown VA Medical Center, Chicago,
Illinois.
Address correspondence to Dr. Elena Barengolts, Department of Medicine,
Section of Endocrinology, MC 640, 1819 West Polk Street, Chicago, IL 60612.
E-mail: [email protected].
Published as a Rapid Electronic Article in Press at http://www.endocrine
practice.org on January 21, 2013. DOI:10.4158/EP12263.RA
To purchase reprints of this article, please visit: www.aace.com/reprints.
Copyright © 2013 AACE.
ABSTRACT
Objective: To review the role of human large bowel
microbacteria (microbiota) in the glucose homeostasis, to
address vitamin D (VD) and prebiotics interactions with
microbiota, and to summarize recent randomized clinical
trials (RCTs) of VD and prebiotics supplementation in pre-
diabetes (PreDM) and type 2 diabetes mellitus (T2DM).
Methods: Primary literature was reviewed in the fol-
lowing areas: composition and activity of human micro-
biota associated with PreDM and T2DM, interactions
between microbiota and glucose homeostasis, the interac-
tion of microbiota with VD/prebiotics, and RCTs of VD/
prebiotics in subjects with PreDM or T2DM.
Results: The human microbiota is comprised of 100
trillion bacteria with an aggregate genome that is 150-fold
larger than the human genome. Data from the animal mod-
els and human studies reveal that an “obesogenic” diet
results into the initial event of microbiota transformation
from symbiosis to dysbiosis. The microbial antigens, such
as Gram(-) bacteria and lipopolysaccharide (LPS), trans-
locate to the host interior and trigger increased energy
harvesting and Toll-like receptor (TLR) activation with
VXEVHTXHQW�LQÁDPPDWRU\�SDWKZD\V�VLJQDOLQJ��7KH�´GRXEOH�
hit” of steatosis (ectopic fat accumulation) and “–itis”
�LQÁDPPDWLRQ��DQG�FRQWULEXWLRQ�RI�´FRULVNVµ��H�J���YLWDPLQ�'� GHÀFLHQF\� >9''@�� DUH� UHTXLUHG� WR� DFWLYDWH� PROHFXODU�signaling, including impaired insulin signaling and secre-
tion, that ends with T2DM and associated diseases. Dietary
changes (e.g., prebiotics, VD supplementation) may ame-
liorate this process if initiated prior to the process becom-
ing irreversible.
Conclusion: Emerging evidence suggests an impor-
tant role of microbiota in glucose homeostasis. VD sup-
plementation and prebiotics may be useful in managing
PreDM and T2DM. (Endocr Pract. 2013;19:497-510)
Abbreviations:1,25D = 1,25-dixydroxyvitamin D; ApoE = apolipopro-
tein E; GutM = gut (large bowel) microbiota; HOMA = homeostasis model of assessment; LPS = lipopoly-
saccharide; PreDM = prediabetes; RCT = randomized
clinical trial; TLR = toll-like receptors; TNF-_�= tumor
necrosis factor-_; VD = vitamin D; VDD = vitamin D
GHÀFLHQF\��VDR = vitamin D receptor
INTRODUCTION
The process of deterioration of normal glucose toler-
ance into prediabetes (PreDM) and type 2 diabetes melli-
tus (T2DM) involves the contribution of “nature,” such as
LQWHUQDO��QRQPRGLÀDEOH� IDFWRUV� �JHQHWLF�EDFNJURXQG��DJH��DQG�JHQGHU��DQG�´QXUWXUH�µ�ZKLFK�DUH�H[WHUQDO��PRGLÀDEOH�factors. Among the external risk factors, the western diet
�KLJK� IDW�� REHVLW\�SURQH�� DQG� YLWDPLQ�'� �9'�� GHÀFLHQF\�(VDD) are important (1,2). Emerging evidence suggests
WKDW� DQRWKHU� LPSRUWDQW� H[WHUQDO� LQÁXHQFH� RQ� WKH� SDWKR-
physiology of glucose homeostasis may be the large bowel
(gut) microbial community (microbiota) (3-10). Over
the past 10 years, there has been an explosion of knowl-
edge; the number of published papers; PubMed articles
with the key word “microbiota” increased exponentially
from 121 (2002) to 2,234 (2012), and those with the key

498
word “human microbiota” grew from 74 (2002) to 1,324
(2012). The complete composition and function of the gut
microbiota (GutM) is still unknown, but it is proposed to
FRQWULEXWH�WR�WKH�ORZ�JUDGH�LQÁDPPDWLRQ�WKDW�SOD\V�D�VXE-
stantial role in the dysregulation of normal glucose toler-
ance into PreDM and T2DM (5-10). The complexity of the
SURFHVV�QRWZLWKVWDQGLQJ��WKHUH�PD\�EH�VLJQLÀFDQW�LQWHUDF-tions between GutM, dietary components (such as prebiot-
ics and VD), and glucose homeostasis suggesting that diet
enrichment or supplementation with prebiotics and VD
may improve glucose homeostasis.
This article will review the role of GutM in the patho-
genesis of prediabetes and T2DM, address how VD and
prebiotics interact with GutM, and summarize the recent
randomized clinical trials (RCTs) of VD and prebiotics
supplementation in PreDM and T2DM.
Gut Microbiota Composition and Function In Prediabetes and Type 2 Diabetes Mellitus
Recent studies demonstrate that the diverse microor-
ganisms that colonize the body at birth live on mucosal sur-
faces and in the lumen of the human large bowel and exist
LQ�EHQHÀFLDO�V\PELRWLF�UHODWLRQVKLSV�ZLWK�WKHLU�KRVW��������%DFWHULD�PDNH�XS�PRVW�RI�WKH�ÁRUD�DQG�XS�WR�����RI�WKH�dry mass of the feces (7). The GutM is comprised of 100
trillion bacteria (10-fold the number of cells in the human
body), and the genomes of these bacteria (microbiome)
have 150 times more genes than the human genome (3,4).
Multiple molecular methods, including quantitative poly-
merase chain reaction PCR, barcoded pyrosequencing, and
SK\ORJHQHWLF�PLFURDUUD\V�RI���6�U51$�KHOSHG�WR�JHQHUDWH�D�FRPSUHKHQVLYH�PLFURELDO�FRPPXQLW\�SURÀOH��7KLV�SURÀOH�VKRZV�WKDW�PRUH�WKDQ�����RI�PLFURELRWD�DUH�DQDHUREHV��DQG�����FRQWDLQ�VHYHUDO�IDPLOLHV��SK\OD��LQFOXGLQJ�Firmicutes
������� Bacteroidetes� ������� Proteobacteria� ������ DQG�Actinobacteria� ����� ���������7KH� KXPDQ� KRVW� SURYLGHV� D�nutrient-rich habitat and impacts GutM survival by dietary
composition (5,9,10) and the use of prebiotics (11-13), pro-
biotics (13,14), and antibiotics (14). Reciprocally, GutM
HQ]\PH� DFWLYLWLHV� DUH� LPSRUWDQW� IRU� FDUFLQRJHQ� GHWR[LÀ-
cation and facilitating nutrient absorption, metabolizing
YLWDPLQV��GUXJV��DQG�HQGRJHQRXV�KRUPRQHV����������������suggesting that GutM may be a major player in human dis-
HDVH��LQIHFWLRQ��LQÁDPPDWLRQ��DOOHUJ\��FDQFHU��REHVLW\��DQG�diabetes) and possibly longevity (3-5,7-17).
There are multiple convincing lines of evidence of
GutM involvement in glucose homeostasis (Table 1) (9,18-
32). The data from in vitro experiments and in vivo animal
models (knockout, germ-free, and humanized gnotobi-
otic mice) helped to elucidate the mechanisms of glucose
homeostasis regulation by GutM (Tables 1 and 2) (33-44)
and reciprocal changes produced by the western “obeso-
genic” diet that alter the nutrients available to GutM and
result in detrimental qualitative and quantitative dysbio-
sis of the GutM ecosystem with preferential increase in
Gram(-) bacteria (5,9,10,18,21,22).
Particularly revealing in elucidating the contributions
of GutM to human biology and pathobiology was studying
WKH�KXPDQL]HG�JQRWRELRWLF�PRXVH�PRGHO�� D�ZHOO�GHÀQHG��representative animal model of the human gut ecosystem
created by transplanting human fecal microbial commu-
nities into wild-type germ-free mice (10). In this model,
switching from a low-fat, polysaccharide-rich “healthy”
plant diet to a high-fat, high-sugar “obesogenic” diet
shifted GutM ecosystem, metabolic pathways, and gene
expression within a single day, and after 2 weeks the “obe-
sogenic” diet-fed mice almost doubled their body fat (10),
suggesting that “obesogenic GutM” had increased capacity
to harvest energy (9,10). Furthermore, this trait was trans-
missible by GutM: the germ-free mice colonized with an
“obesogenic GutM” doubled their body fat compared to
mice colonized with a “lean GutM” despite consuming the
exact same diet (9,10). Similarly, transplantation of GutM
from mice with metabolic syndrome and insulin resistance
to the wild-type germ-free mice resulted in the transmis-
sion of insulin resistance to the recipient mice (8).
GutM dysbiosis resulted in increased production of
lipopolysaccharide (LPS), a molecule from the outer mem-
brane of Gram(-) bacteria that disrupted mucosal immunity,
breeched the mucosal barrier, acquired access to the inte-
ULRU�RI�WKH�KRVW��DQG�DFWLYDWHG�LQÁDPPDWLRQ�DQG�V\VWHPLF�immunity (Tables 1 and 2). In a mouse model, LPS was
LGHQWLÀHG� DV� D� FDXVDWLYH� WULJJHULQJ� IDFWRU�RI� WKH�RQVHW� RI�obesity, insulin resistance, and diabetes (5). In this model,
the usual diet increased or decreased circulating LPS during
the fed or fasted state, respectively. In contrast, a high-fat
diet chronically increased LPS by 2 to 3 times, a threshold
WKDW�ZDV�GHÀQHG�E\�WKH�DXWKRUV�DV�PHWDEROLF�HQGRWR[HPLD�(for comparison, sepsis and other infections would increase
circulating LPS by 10 to 50 times). Importantly, a high-fat
diet increased the proportion of an LPS-generating GutM,
as well as endogenous LPS production proportionally to
the amount of dietary fat and enhanced the absorption of
orally administered LPS (5). When metabolic endotoxemia
was induced through continuous subcutaneous LPS infu-
sion, fasted glycemia and insulinemia and whole-body,
liver, and adipose tissue weight gain increased to a similar
extent as in high-fat-diet mice. Both endogenous and exog-
enous LPS caused insulin resistance acting via CD14 (a
protein from the Cluster of Differentiation group that acts
as a co-receptor for LPS along with TLR-4, LPS binding
SURWHLQ�� DQG�0'�� SURWHLQ��� 0DFURSKDJH� LQÀOWUDWLRQ� ZDV�higher in adipose tissue of mice fed a high-fat diet and in
mice infused with LPS, and this effect was not observed
in the absence of CD14 (5). The authors suggested that
/36�PLJKW�EH�D�QHZO\�LGHQWLÀHG�LQÁDPPDWRU\�IDFWRU�IURP�

499
GutM, which upon binding to its receptor serves as a vec-
WRU�IRU�WKH�WULJJHULQJ�DQ�´LQÁDPPDWLRQ�LQVXOLQ�UHVLVWDQFH�diabetes” cascade induced by high-fat feeding.
In healthy humans, circulating LPS was associated
with both prevalent and incident T2DM (45) and with fat
DQG�HQHUJ\�LQWDNHV�������DQG�FLUFXODWLQJ�&'���FRUUHODWHG�with insulin resistance (homeostasis model of assessment
>+20$@�,5��� YLVFHUDO� DGLSRVLW\�� DQG�PDUNHUV� RI� LQÁDP-
mation and endothelial dysfunction (47). In patients with
T2DM, genetic polymorphism of CD14 correlated with
insulin sensitivity index (ISI) and endothelial dysfunction
������+HDOWK\� VXEMHFWV� KDG� KLJKHU� IDVWLQJ�/36� DIWHU� FRQ-
suming a fat-enriched meal compared to a control meal (5).
Recently, intravenous LPS infusion to healthy subjects was
used to establish a human model of endotoxemia and sub-
FOLQLFDO�LQÁDPPDWLRQ�������,Q�D�GRXEOH�EOLQG��SODFHER�FRQ-
trolled, random sequence crossover study, low-dose LPS
infusion caused a rapid and transient increase of plasma
WXPRU� QHFURVLV� IDFWRU� �71)��_ (25-fold) and interleukin
�,/���������IROG��IROORZHG�E\�PRGHVW�LQÁDPPDWLRQ�LQ�DGL-pose tissue manifested by induction of mediators of insulin
UHFHSWRU� VLJQDOLQJ� �,/����71)�_, monocyte chemoattrac-
WDQW�SURWHLQ�>0&3��@��IUDFWDONLQH��DQG�VXSSUHVVRU�RI�F\WR-
NLQH�VLJQDOLQJ� >62&6@���DQG� �����0RVW� LPSRUWDQWO\��/36�FDXVHG�VLJQLÀFDQW�LQGXFWLRQ�RI�V\VWHPLF�LQVXOLQ�UHVLVWDQFH�measured by intravenous glucose tolerance (IVGTT, insu-
OLQ�VHQVLWLYLW\�GHFOLQHG�E\������ZLWKRXW�HYLGHQFH�RI�SDQ-
FUHDWLF�EHWD�FHOO�G\VIXQFWLRQ��QR�FKDQJH�LQ�WKH�ÀUVW�SKDVH�RI� WKH� DFXWH� LQVXOLQ� UHVSRQVH� >$,5J@� LQGH[��� &RQVLVWHQW�ZLWK� WKH� ,9*77� GDWD�� LQVXOLQ� UHVLVWDQFH� E\� +20$�,5�LQFUHDVH� E\� ���� ZKLOH� +20$�%� ZDV� XQFKDQJHG� ������Bariatric surgery in morbidly obese patients with and with-
RXW�GLDEHWHV�UHVXOWHG�LQ�VLJQLÀFDQW�FKDQJHV�LQ�*XW0�DIWHU���DQG���PRQWKV�DQG�OHG�WR�PHWDEROLF�SURÀOH�LPSURYHPHQWV��including blood glucose, glycated hemoglobin A1C, insu-
OLQHPLD��+20$�,5��OHSWLQ��DQG�DGLSRQHFWLQ�������,Q�WKHVH�
Table 1Human Studies of the Major Interactions Between the Gut Microbiota
and Glucose Metabolism: Data from Normal Weight, Obese, and Diabetic Patients
Population Major changes in human microbiota composition and activity
+HDOWK\��QRUPDO�weight
Increased calorie intake from 2400 to 3400 kcal/d lead to increased absorption ~150 kcal resulting
���LQWR�LQFUHDVHG�)LUPLFXWHV��a�����DQG�GHFUHDVHG�%DFWHURLGHWHV��a���������Decreased calorie intake from 3400 to 2400 kcal/d lead to decreased absorption ~150 kcal resulting
���LQWR�LQFUHDVHG�%DFWHURLGHWHV��a�����DQG�GHFUHDVHG�)LUPLFXWHV��a���������,QFUHDVHG�LQÁDPPDWRU\�PDUNHUV��,/����71)�_, MCP-1, CX3CL1) in adipose tissue after
LPS IV (19)
/36��FRUUHODWHG�ZLWK�,16�����Obese 'HFUHDVHG�RYHUDOO�*XW0�GLYHUVLW\��GHFUHDVHG�)LUPLFXWHV�DQG�%LÀGREDFWHULD� variable Bacteroidetes
���DQG�$UFKDHD��PHWKDQRJHQV��LQ�2%�YV��&21�����������'HFUHDVHG�%LÀGREDFWHULD�LQ�QHRQDWHV�RI�2:�PRWKHUV and associated with obesity later in
childhood (24)
Increased Bacteroides and Proteobacteria and decreased Firmicutes with wt loss diet (24,25)
���RU�EDULDWULF�VXUJHU\�����*XW0�SURÀOH�FRUUHODWHG�ZLWK�,16�VHQVLWLYLW\�����
LPS: increased LPS after a high fat meal (28); LPS predicted by fat and energy intakes (18);
���/36�FRUUHODWHG�SRVLWLYHO\�ZLWK�,16������DQG�7*�������DQG�GLG�QRW�FRUUHODWH�ZLWK�%*�����������RU�+'/�����
T2DM 'HFUHDVHG�RYHUDOO�*XW0�GLYHUVLW\��GHFUHDVHG�)LUPLFXWHV�DQG�%LÀGREDFWHULD��LQFUHDVHG�%DFWHURLGHV����LQ�7�'0�YV��&21��������*XW0�SURÀOH�FRUUHODWHG�ZLWK�%*������DQG�LQÁDPPDWRU\�PDUNHUV�����
/36��LQFUHDVHG�/36�LQ�7�'0�YV��2%�VH[���DQG�DJH�PDWFKHG�&21�������/36�FRUUHODWHG�SRVLWLYHO\����ZLWK�,16���������DQG�7*�QHJDWLYHO\ ZLWK�+'/�������DQG�GLG�QRW�FRUUHODWH�ZLWK�%*��������
$EEUHYLDWLRQV��%*� �EORRG�JOXFRVH��&21� �FRQWUROV��&;�&/�� �IUDFWDONLQH��,16� �FLUFXODWLQJ�LQVXOLQ��,9� �LQWUDYHQRXV�LQIXVLRQ��,/��� �LQWHUOHXNLQ����/36� �FLUFXODWLQJ�OLSRSRO\VDFFKDULGH��0&3��� �PRQRF\WH�FKHPRWDFWLF�SURWHLQ����2%� �REHVH��2:� �RYHU-weight; T2DM = type 2 diabetes mellitus; 7*� �WULJO\FHULGHV��71)�_�= tumor necrosis factor-_; wt = weight.
500
patients, Faecalibacterium prausnitzii, a conserved and
dominant species of the healthy GutM, was directly linked
WR�WKH�UHGXFWLRQ�LQ�ORZ�JUDGH�LQÁDPPDWLRQ�VWDWH�LQGHSHQ-
dently of calorie intake and was negatively associated with
+20$�,5� ������ )LQDOO\�� WUDQVSODQWDWLRQ� RI� *XW0� IURP�lean healthy donors to recipients with metabolic syndrome
improved insulin sensitivity of recipients (median rate of
JOXFRVH�GLVDSSHDUDQFH�LQFUHDVHG�IURP������WR������+mol/
kg/min; P<.05) and increased abundance of butyrate-pro-
ducing GutM (48).
The investigation of the molecular mechanisms of the
effects of LPS on pancreatic beta-cell function showed
that LPS inhibited insulin gene expression and decreased
glucose-induced insulin secretion (39). These deleterious
effects of LPS were mediated through Toll-like receptor
�7/5����DQG�YLD�QXFOHDU�IDFWRU�NDSSD�%��1)�gB) signaling
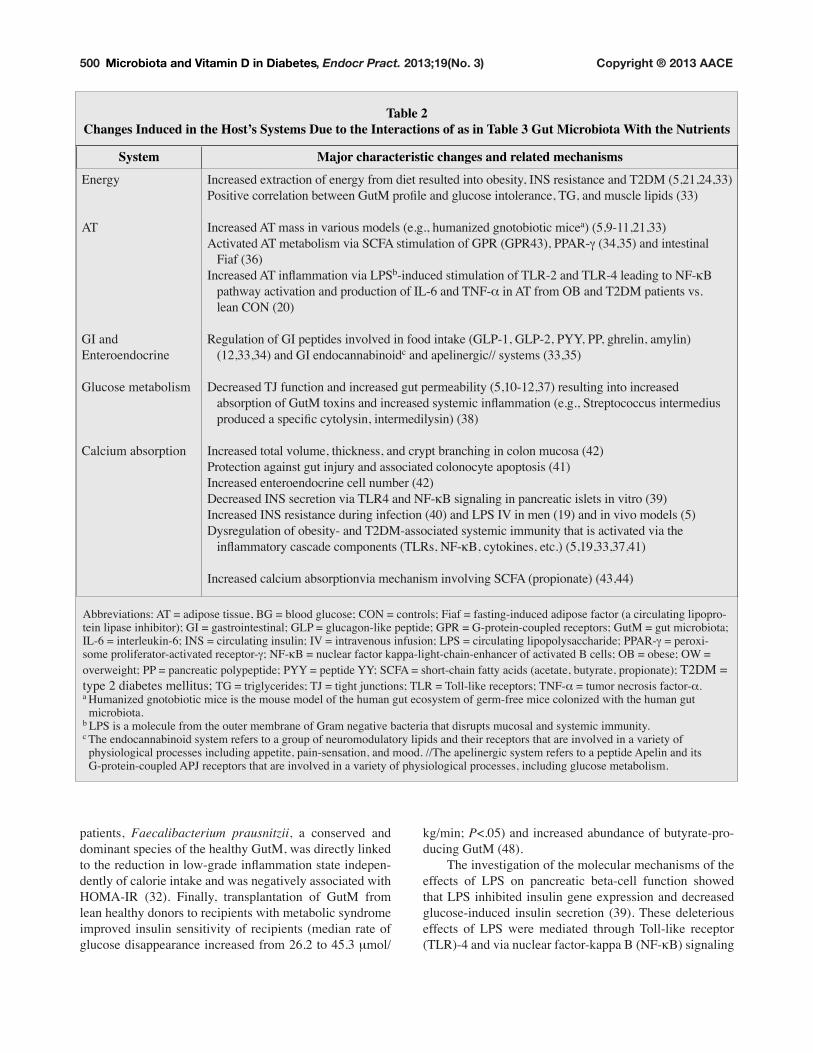
Table 2Changes Induced in the Host’s Systems Due to the Interactions of as in Table 3 Gut Microbiota With the Nutrients
System Major characteristic changes and related mechanismsEnergy
AT
GI and
Enteroendocrine
Glucose metabolism
Calcium absorption
,QFUHDVHG�H[WUDFWLRQ�RI�HQHUJ\�IURP�GLHW�UHVXOWHG�LQWR�REHVLW\��,16�UHVLVWDQFH�DQG�7�'0�������������3RVLWLYH�FRUUHODWLRQ�EHWZHHQ�*XW0�SURÀOH�DQG�JOXFRVH�LQWROHUDQFH��7*��DQG�PXVFOH�OLSLGV�����
Increased AT mass in various models (e.g., humanized gnotobiotic micea) (5,9-11,21,33)
Activated AT metabolism via SCFA stimulation of GPR (GPR43), PPAR-a (34,35) and intestinal
���)LDI������,QFUHDVHG�$7�LQÁDPPDWLRQ�YLD�/36b�LQGXFHG�VWLPXODWLRQ�RI�7/5���DQG�7/5���OHDGLQJ�WR�1)�gB
���SDWKZD\�DFWLYDWLRQ�DQG�SURGXFWLRQ�RI�,/���DQG�71)�_ in AT from OB and T2DM patients vs.
���OHDQ�&21������
Regulation of GI peptides involved in food intake (GLP-1, GLP-2, PYY, PP, ghrelin, amylin)
(12,33,34) and GI endocannabinoidc and apelinergic// systems (33,35)
Decreased TJ function and increased gut permeability (5,10-12,37) resulting into increased
���DEVRUSWLRQ�RI�*XW0�WR[LQV�DQG�LQFUHDVHG�V\VWHPLF�LQÁDPPDWLRQ��H�J���6WUHSWRFRFFXV�LQWHUPHGLXV����SURGXFHG�D�VSHFLÀF�F\WRO\VLQ��LQWHUPHGLO\VLQ������
Increased total volume, thickness, and crypt branching in colon mucosa (42)
Protection against gut injury and associated colonocyte apoptosis (41)
Increased enteroendocrine cell number (42)
'HFUHDVHG�,16�VHFUHWLRQ�YLD�7/5��DQG�1)�gB signaling in pancreatic islets in vitro (39)
,QFUHDVHG�,16�UHVLVWDQFH�GXULQJ�LQIHFWLRQ������DQG�/36�,9�LQ�PHQ������DQG�LQ�YLYR�PRGHOV�����Dysregulation of obesity- and T2DM-associated systemic immunity that is activated via the
���LQÁDPPDWRU\�FDVFDGH�FRPSRQHQWV��7/5V��1)�gB, cytokines, etc.) (5,19,33,37,41)
Increased calcium absorptionvia mechanism involving SCFA (propionate) (43,44)
$EEUHYLDWLRQV��$7� �DGLSRVH�WLVVXH��%*� �EORRG�JOXFRVH��&21� �FRQWUROV��)LDI� �IDVWLQJ�LQGXFHG�DGLSRVH�IDFWRU��D�FLUFXODWLQJ�OLSRSUR-
tein lipase inhibitor); GI = gastrointestinal; GLP = glucagon-like peptide; GPR = G-protein-coupled receptors; GutM = gut microbiota;
,/��� �LQWHUOHXNLQ����,16� �FLUFXODWLQJ�LQVXOLQ��,9� �LQWUDYHQRXV�LQIXVLRQ��/36� �FLUFXODWLQJ�OLSRSRO\VDFFKDULGH��33$5�a = peroxi-
some proliferator-activated receptor-a��1)�gB = nuclear factor kappa-light-chain-enhancer of activated B cells; OB = obese; OW =
overweight; PP = pancreatic polypeptide; PYY = peptide YY; SCFA = short-chain fatty acids (acetate, butyrate, propionate); T2DM =
type 2 diabetes mellitus; 7*� �WULJO\FHULGHV��7-� �WLJKW�MXQFWLRQV��7/5� �7ROO�OLNH�UHFHSWRUV��71)�_ = tumor necrosis factor-_. a +XPDQL]HG�JQRWRELRWLF�PLFH�LV�WKH�PRXVH�PRGHO�RI�WKH�KXPDQ�JXW�HFRV\VWHP�RI�JHUP�IUHH�PLFH�FRORQL]HG�ZLWK�WKH�KXPDQ�JXW� microbiota. b LPS is a molecule from the outer membrane of Gram negative bacteria that disrupts mucosal and systemic immunity. c The endocannabinoid system refers to a group of neuromodulatory lipids and their receptors that are involved in a variety of
physiological processes including appetite, pain-sensation, and mood. //The apelinergic system refers to a peptide Apelin and its
G-protein-coupled APJ receptors that are involved in a variety of physiological processes, including glucose metabolism.

501
in pancreatic islets, suggesting a novel mechanism by
which GutM might affect pancreatic beta-cell function
(39). All these data support but do not prove the emerging
view that GutM might play a causal role in pathogenesis of
insulin resistance, prediabetes, and diabetes.
One of the links in the microbiota-glycemia interac-
tion is the enteroendocrine system and the related hor-
mones, including glucagon like peptide-1 and -2 (GLP-1,
GLP-2) and peptide YY (PYY) secreted by the intestinal
L-cells; ghrelin, secreted by the stomach; amylin and pan-
creatic polypeptide (PP) secreted by the pancreas; and adi-
ponectin and leptin secreted by adipose tissue (Table 2).
These hormones are well known modulators of appetite,
food intake, and glucose and fat metabolism and some of
them (e.g., GLP-2) are involved in regulating gut barrier
function through the expression of tight junction proteins
(49).
Various models were utilized to establish the role of
enteroendocrine system in the microbiota-glycemia con-
nection through manipulation of GutM, including geneti-
cally obese and high-fat diet-induced obese and diabetic
mice and fermentable carbohydrate-enriched (prebiotic)
diets. In the study of genetically obese and high-fat diet-
induced obese and diabetic mice, both obesity and pre-
biotic supplementation caused changes in GutM and the
enteroendocrine system (33). The researchers used multiple
molecular methods to generate comprehensive microbial
FRPPXQLW\� SURÀOHV� DQG� VKRZHG� WKDW� SUHELRWLF� LQWHUYHQ-
tion resulted in a decrease of Firmicutes and an increase
of Bacteroidetes phyla and a change of 102 distinct taxa,
��� RI�ZKLFK� GLVSOD\HG� D� !���IROG� FKDQJH� LQ� DEXQGDQFH��and these changes ameliorated obesity-induced gut per-
meability and metabolic endotoxemia (33). Interestingly,
VRPH� VSHFLÀF� EDFWHULD� ZHUH� LGHQWLÀHG� DV� SRVVLEO\� SOD\-
ing particularly important roles. For example, there was
a very strong correlation between gut permeability and
the abundance of Streptococcus intermedius, known to
SURGXFH� D� VSHFLÀF� F\WRO\VLQ� �LQWHUPHGLO\VLQ�� ����� WKDW�alters intestinal cell tight-junction architecture (33). Also,
Akkermansia muciniphila (A. muciniphila), which is asso-
ciated with healthy colon mucosa, strongly and positively
correlated with the enteroendocrine L-cell number (33).
Of note, A. muciniphila inversely correlated with body
weight in humans, increasing after gastric bypass-induced
weight loss (27). Moreover, prebiotic-induced change in
mice GutM was accompanied by increased colon L-cell
QXPEHU� �a������HQKDQFHG� LQWHVWLQDO�SURJOXFDJRQ�P51$�H[SUHVVLRQ� �a��������� LQFUHDVHG� SODVPD� */3��� OHYHOV��a���������� DQG� LPSURYHG� OHSWLQ� VHQVLWLYLW\� �a���������Prebiotic use counteracted high-fat-induced body weight
gain and fat mass development and resulted in improved
glucose tolerance, decreased plasma triglycerides and mus-
cle lipid content (total, triglycerides, and phospholipids),
and reduced expression of oxidative stress (measured by
1$'3+�R[LGDVH��DQG�LQÁDPPDWRU\��,/���P51$��PDUNHUV�LQ�WKH�FRORQ�������,Q�WKH�VDPH�PRXVH�PRGHO��VLPLODU�EHQHÀ-
FLDO�FKDQJHV�LQ�*XW0��VSHFLÀFDOO\�DQ�LQFUHDVH�LQ�ELÀGREDF-teria and a decrease in lactobacilli) were accompanied by
improved gut barrier integrity, improved metabolic endo-
WR[HPLD� DQG�JOXFRVH� WROHUDQFH�� DQG�GHFUHDVHG� LQÁDPPD-tion and weight (49). Multiple enteroendocrine and other
hormones were studied (GLP-1, PYY, PP, amylin, leptin,
JKUHOLQ��ZLWK�VLJQLÀFDQW�FKDQJHV�REVHUYHG�LQ�*/3����3<<��DQG� OHSWLQ��$QDO\VLV�RI� WKH�FRORQ�JHQH�H[SUHVVLRQ�SURÀOH�showed up-regulation of PYY, peroxisome proliferator-
activated receptor (PPAR)-a, TLR-4, and endocannabinoid
system receptor CB2 genes. The authors postulated that the
observed effects could be related to modulation of GutM,
VXFK�DV�KLJKHU�FRQWHQW�RI�ELÀGREDFWHULD�LQ�WKH�FDHFXP������ A randomized, double-blind, parallel, placebo-con-
trolled trial of fermentable carbohydrates (prebiotic) in
healthy humans showed a threefold increase in gut micro-
bial fermentation, increased satiety, and decreased hunger
and energy intake, whereas postprandial plasma glucose
responses decreased after the standardized meal. These
changes were accompanied by higher serum GLP-1 and
PYY concentrations (12). In overweight and obese adults,
prebiotic supplementation resulted in lower circulating
ghrelin, higher PYY, and improved insulin sensitivity in
the intervention group vs. control (51). The mechanism of
enteroendocrine system action might involve sensing the
presence of bacterial antigens (including LPS) through
TLR and neutralizing intestinal bacteria by releasing che-
mokines and defensins as demonstrated with in vitro and in
vivo models, and by inducing contraction of the muscular
tunica, favoring the emptying of the distal small intestine
(52).
The other systems important in obesity-dysglycemia-
dyslipidemia and implicated in the crosstalk between
GutM and metabolism are the endocannabinoid and ape-
linergic systems, with the former involving a group of
neuromodulatory lipids and their receptors, and the latter
involving a peptide apelin and its G-protein-coupled APJ
receptors (Table 2) (12,27,33-35,49-51). In vitro and in
vivo studies using endocannabinoid system agonists and
antagonists showed that this system linked microbiota with
adipogenesis and glucose metabolism by modulating gut
SHUPHDELOLW\��WLJKW�MXQFWLRQ�SURWHLQV��H[SUHVVLRQ�RI�LQÁDP-
matory markers, markers of adipocyte differentiation, and
adipocyte size and number and via their effects on colon
and adipose tissue cannabinoid receptors, with LPS acting
as a master switch (52). Although causality remains to be
proven, these experiments highlighted the essential role
of the enteroendocrine and endocannabinoid systems in
PLFURELRWD�UHODWHG� PRGXODWLRQ� RI� V\VWHPLF� LQÁDPPDWLRQ�and glucose metabolism.

502
Vitamin D Interaction With Gut Microbiota and Vitamin D Use in Randomized Clinical Trials for
Improving Glucose Homeostasis In humans, active 1,25-dixydroxyvitamin D (1,25D)
can be produced and function in endocrine, paracrine and
autocrine manner. VD system directly or indirectly regu-
ODWHV�DERXW����RI�PRXVH�DQG�KXPDQ�JHQRPH�������LQFOXG-
ing genes involved in glucose homeostasis. The role of
VD in glucose homeostasis is one of the well-established
nonclassical roles of this multipotential vitamin (2). All the
components of the VD system are present in the mucosal
cells lining the large bowel (colonocytes), including VD
receptor (VDR), VD response elements (VDREs), and the
enzymes involved in synthesis and catabolism of active
����'��������� VD interactions with GutM may become yet another
nonclassical function of VD (Table 3) (57-70). There are
several pathways for possible VD and GutM relationships
in the large bowel. VD may contribute to acquisition and
PDLQWHQDQFH� RI� V\PELRWLF� JXW� PLFURÁRUD�� %RWK� 9'� DQG�GutM regulate calcium absorption, cellular tight junctions,
mucosal and systemic immunity, and colonocyte activity
(Tables 2 and 3).
VD-regulated intestinal calcium absorption, the clas-
sical action of VD, occurs predominantly in duodenum
and jejunum and involves active intracellular and passive
paracellular absorption. Similar mechanisms are involved
LQ�DERXW�����RI�FDOFLXP�DEVRUSWLRQ�WKDW�RFFXUV�LQ�WKH�ODUJH�ERZHO� ��������� DQG� WKLV� SURFHVV� LV� HQKDQFHG� E\� *XW0�produced propionic acid (43,44). Calcium plays crucial
role in all physiological functions, and it is possible that
VD and GutM act synergistically to maintain calcium bal-
ance. The tight junctions between the intestinal cells play
an important role in paracellular calcium absorption and
are also an integral component of mucosal immunity. Both
VD and GutM are involved in the regulation of the tight
junctions that establish mucosal barriers of defense against
intestinal pathogens (Tables 2 and 3). These intestinal
pathogens may have developed ways of penetrating the
mucosal barrier to exploit the host’s physiological path-
ways of nutrient absorption (e.g., calcium) as a crucial step
in gaining access to the host’s internal resources. Both VD
and GutM play a role in the regulation and signaling path-
ways of all 3 components (mucosal, innate, and adaptive)
of the immune system (Tables 2 and 3) and act to develop
and maintain self-tolerance by down regulating the adap-
tive immune responses while enhancing the protective
innate immune responses. Similarly, both VD and GutM
have trophic effects on colonocytes (Tables 2 and 3) that
have shown in vitro, in animal models, and in human stud-
LHV�RI� LQÁDPPDWRU\�ERZHO�GLVHDVH�DQG�FRORUHFWDO�FDQFHU��LQFOXGLQJ�JHQHWLF�GDWD��������� A few studies suggested possible interdependence
of microbiota and VD and its role in glycemia. In human
monocytic cells, LPS stimulated secretion of 1,25D via
upregulation of 1_-hydroxylase, a transcription factor
(STAT1_), a coactivator (p300), and chromatin remod-
eling (77). Similarly, LPS activated 1_-hydroxylase in
the kidney (78) and in endothelial cells (79). In a human
colon cancer cells, butyrate (a metabolic product of GutM)
VLJQLÀFDQWO\� LQFUHDVHG�9'5� QXPEHU� DQG� DFWLYLW\� ������YV��FRQWURO���%RWK�EXW\UDWH�������YV��FRQWURO��DQG�����'�������YV��FRQWURO��VLJQLÀFDQWO\�VWLPXODWHG�GLIIHUHQWLDWLRQ�of cells, whereas combined treatment with butyrate and
����'� UHVXOWHG� LQ� D� V\QHUJLVWLF� DPSOLÀFDWLRQ� RI� FHOO� GLI-IHUHQWLDWLRQ� ������� YV�� FRQWURO��� 7KH� UHVSRQVLYHQHVV� WR�LPS of monocytes modulated by 1,25D differed between
T2DM patients and controls (80,81). The direct evidence
of the interaction between GutM, VD, and glucose metabo-
OLVP�FDPH�IURP�DSROLSRSURWHLQ�(�GHÀFLHQW�PLFH��$SR(–/–),
a widely used model of obesity with accelerated athero-
sclerosis. A recent work showed that ApoE–/– mice had
impaired glucose tolerance and insulin signaling; increased
LQWHVWLQDO� SHUPHDELOLW\�� LQÁDPPDWRU\� PHGLDWRUV� �71)�_,
5$17(6��LQWUDFHOOXODU�DGKHVLRQ�PROHFXOH�>,&$0@����DQG�PDFURSKDJH�LQÁDPPDWRU\�SURWHLQ�>0,3@��_ in the colon,
liver, and adipose tissue), and scavenger cells among circu-
ODWLQJ�PDFURSKDJHV��DV�ZHOO�DV�D�VLJQLÀFDQW�LQFUHDVH�LQ�WKH�intestinal expression of VDR. The probiotic bacterial com-
ELQDWLRQ� HIÀFLHQWO\� SUHYHQWHG� LQVXOLQ� UHVLVWDQFH� LQ� WKHVH�ApoE–/– mice, likely acting via transactivation of PPAR-a, Farnesoid-X-receptors (FXR), and VDR (82).
The VDD models highlight importance of VD in the
GutM-glycemia relationship. In the VDD rats, oral VD
VXSSOHPHQWDWLRQ�LPSURYHG�PHWDEROLF�HIÀFLHQF\��OLNHO\�DFW-ing via modulation of GutM; body mass was maintained
but there were increases in food intake (1.8 times), cae-
cum size (1.4-times), short-chain fatty acid (SCFA) pro-
duction, and metabolic rate (83). In a diet-induced VDD
mouse model, 27 genes were up- or downregulated (more
than twofold), and VDD mice had elevated levels (50-
fold) of bacteria in the colonic tissue, suggesting that VDD
dysregulated colonic antimicrobial activity and impaired
GutM ecosystem (57). The activated genes were involved
in containment of enteric bacteria (an angiogenin-4 gene
coding for an antimicrobial protein) and in glucose homeo-
stasis. For example, the carbonyl reductase 3 (CBR3) gene
was downregulated threefold in VDD mice, and genetic
variation in CBR3 conferred risk of T2DM and insulin
resistance, most likely via regulation of adipogenesis (84).
Some other genes included apolipoprotein C-II (a modu-
lator of cardiovascular disease in T2DM) (85); TF3, an
adaptive-response gene induced by various stress signals
relevant to T2DM and involved in insulin gene transcrip-
WLRQ�������DQG�SOHLRPRUSKLF�DGHQRPD�JHQH�OLNH����3ODJO����which is involved in insulin secretion and may be impor-
tant in beta-cell exhaustion in insulin-resistant states (87).
� $OWKRXJK�VXJJHVWLYH��WKH�ÀQGLQJV�REWDLQHG�LQ�WKHVH�LQ�vitro and in vivo animal models cannot be translated into
human clinical settings, and further studies are needed
503
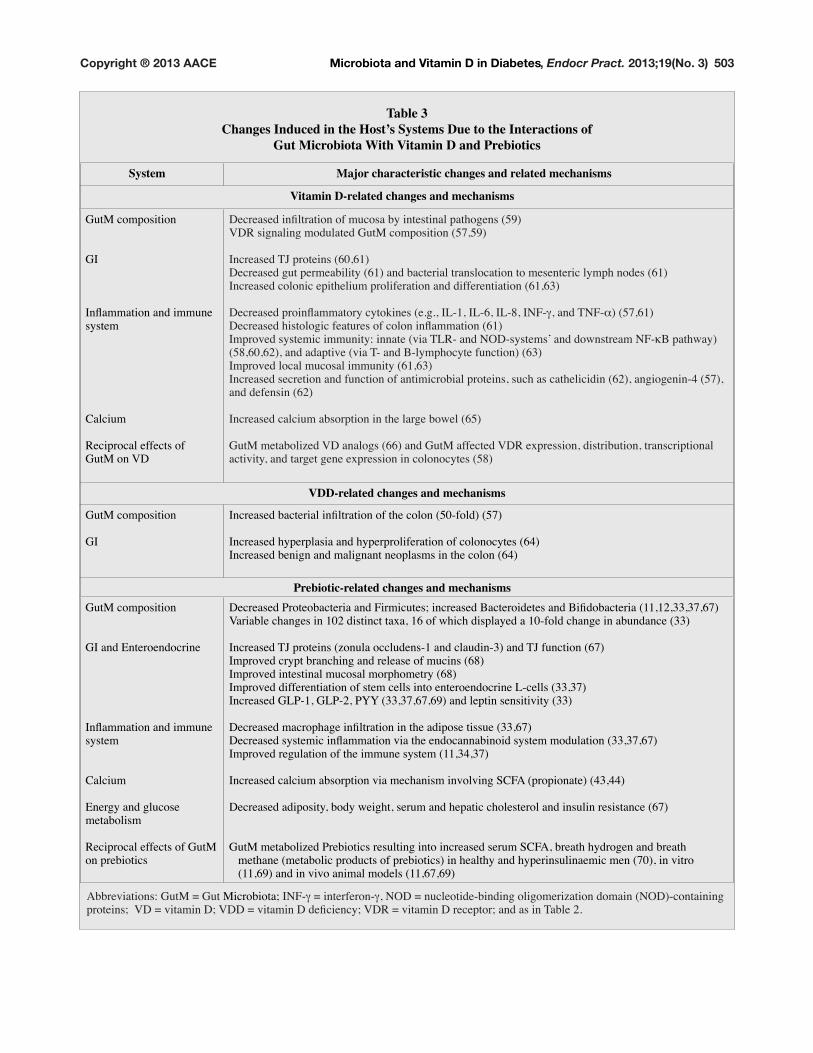
Table 3Changes Induced in the Host’s Systems Due to the Interactions of
Gut Microbiota With Vitamin D and Prebiotics
System Major characteristic changes and related mechanisms
Vitamin D-related changes and mechanisms
GutM composition
GI
,QÁDPPDWLRQ�DQG�LPPXQH�system
Calcium
Reciprocal effects of
GutM on VD
'HFUHDVHG�LQÀOWUDWLRQ�RI�PXFRVD�E\�LQWHVWLQDO�SDWKRJHQV������VDR signaling modulated GutM composition (57,59)
,QFUHDVHG�7-�SURWHLQV��������'HFUHDVHG�JXW�SHUPHDELOLW\������DQG�EDFWHULDO�WUDQVORFDWLRQ�WR�PHVHQWHULF�O\PSK�QRGHV������,QFUHDVHG�FRORQLF�HSLWKHOLXP�SUROLIHUDWLRQ�DQG�GLIIHUHQWLDWLRQ���������
'HFUHDVHG�SURLQÁDPPDWRU\�F\WRNLQHV��H�J���,/����,/����,/����,1)�a��DQG�71)�_����������'HFUHDVHG�KLVWRORJLF�IHDWXUHV�RI�FRORQ�LQÁDPPDWLRQ������,PSURYHG�V\VWHPLF�LPPXQLW\��LQQDWH��YLD�7/5��DQG�12'�V\VWHPV·�DQG�GRZQVWUHDP�1)�gB pathway)
������������DQG�DGDSWLYH��YLD�7��DQG�%�O\PSKRF\WH�IXQFWLRQ�������,PSURYHG�ORFDO�PXFRVDO�LPPXQLW\��������,QFUHDVHG�VHFUHWLRQ�DQG�IXQFWLRQ�RI�DQWLPLFURELDO�SURWHLQV��VXFK�DV�FDWKHOLFLGLQ�������DQJLRJHQLQ���������DQG�GHIHQVLQ������
,QFUHDVHG�FDOFLXP�DEVRUSWLRQ�LQ�WKH�ODUJH�ERZHO�����
*XW0�PHWDEROL]HG�9'�DQDORJV������DQG�*XW0�DIIHFWHG�9'5�H[SUHVVLRQ��GLVWULEXWLRQ��WUDQVFULSWLRQDO�activity, and target gene expression in colonocytes (58)
VDD-related changes and mechanisms
GutM composition
GI
,QFUHDVHG�EDFWHULDO�LQÀOWUDWLRQ�RI�WKH�FRORQ�����IROG������
,QFUHDVHG�K\SHUSODVLD�DQG�K\SHUSUROLIHUDWLRQ�RI�FRORQRF\WHV�����,QFUHDVHG�EHQLJQ�DQG�PDOLJQDQW�QHRSODVPV�LQ�WKH�FRORQ�����
Prebiotic-related changes and mechanismsGutM composition
GI and Enteroendocrine
,QÁDPPDWLRQ�DQG�LPPXQH�system
Calcium
Energy and glucose
metabolism
Reciprocal effects of GutM
on prebiotics
'HFUHDVHG�3URWHREDFWHULD�DQG�)LUPLFXWHV��LQFUHDVHG�%DFWHURLGHWHV�DQG�%LÀGREDFWHULD�����������������9DULDEOH�FKDQJHV�LQ�����GLVWLQFW�WD[D�����RI�ZKLFK�GLVSOD\HG�D����IROG�FKDQJH�LQ�DEXQGDQFH�����
,QFUHDVHG�7-�SURWHLQV��]RQXOD�RFFOXGHQV���DQG�FODXGLQ����DQG�7-�IXQFWLRQ�����,PSURYHG�FU\SW�EUDQFKLQJ�DQG�UHOHDVH�RI�PXFLQV�����,PSURYHG�LQWHVWLQDO�PXFRVDO�PRUSKRPHWU\�����Improved differentiation of stem cells into enteroendocrine L-cells (33,37)
,QFUHDVHG�*/3����*/3����3<<���������������DQG�OHSWLQ�VHQVLWLYLW\�����
'HFUHDVHG�PDFURSKDJH�LQÀOWUDWLRQ�LQ�WKH�DGLSRVH�WLVVXH���������'HFUHDVHG�V\VWHPLF�LQÁDPPDWLRQ�YLD�WKH�HQGRFDQQDELQRLG�V\VWHP�PRGXODWLRQ�����������Improved regulation of the immune system (11,34,37)
Increased calcium absorption via mechanism involving SCFA (propionate) (43,44)
'HFUHDVHG�DGLSRVLW\��ERG\�ZHLJKW��VHUXP�DQG�KHSDWLF�FKROHVWHURO�DQG�LQVXOLQ�UHVLVWDQFH�����
GutM metabolized Prebiotics resulting into increased serum SCFA, breath hydrogen and breath
methane (metabolic products of prebiotics) in healthy and hyperinsulinaemic men (70), in vitro
�����������DQG�LQ�YLYR�DQLPDO�PRGHOV�����������
Abbreviations: GutM = Gut Microbiota; ,1)�a = interferon-a��12'� �QXFOHRWLGH�ELQGLQJ�ROLJRPHUL]DWLRQ�GRPDLQ��12'��FRQWDLQLQJ�SURWHLQV���9'� �YLWDPLQ�'��9''� �YLWDPLQ�'�GHÀFLHQF\��9'5� �YLWDPLQ�'�UHFHSWRU��DQG�DV�LQ�7DEOH���

504
to elucidate causal relationships among GutM, glucose
metabolism, and VD. The available data suggest that VDD
may act as a predisposing or/and precipitating factor that
contributes (with other mechanisms) to destabilization
of GutM, decreased colon cell antimicrobial activity, and
increased mucosal permeability. These events may allow
the intact bacteria and/or bioactive bacterial products (e.g.,
LPS) to translocate from the intestinal lumen to the body’s
interior and induce (after activation of multiple complex
SDWKZD\V�� ORZ�JUDGH� LQÁDPPDWLRQ��ZKLFK� LV�D� VXJJHVWHG�mechanism linking GutM with metabolic disorders, includ-
ing prediabetes and T2DM.
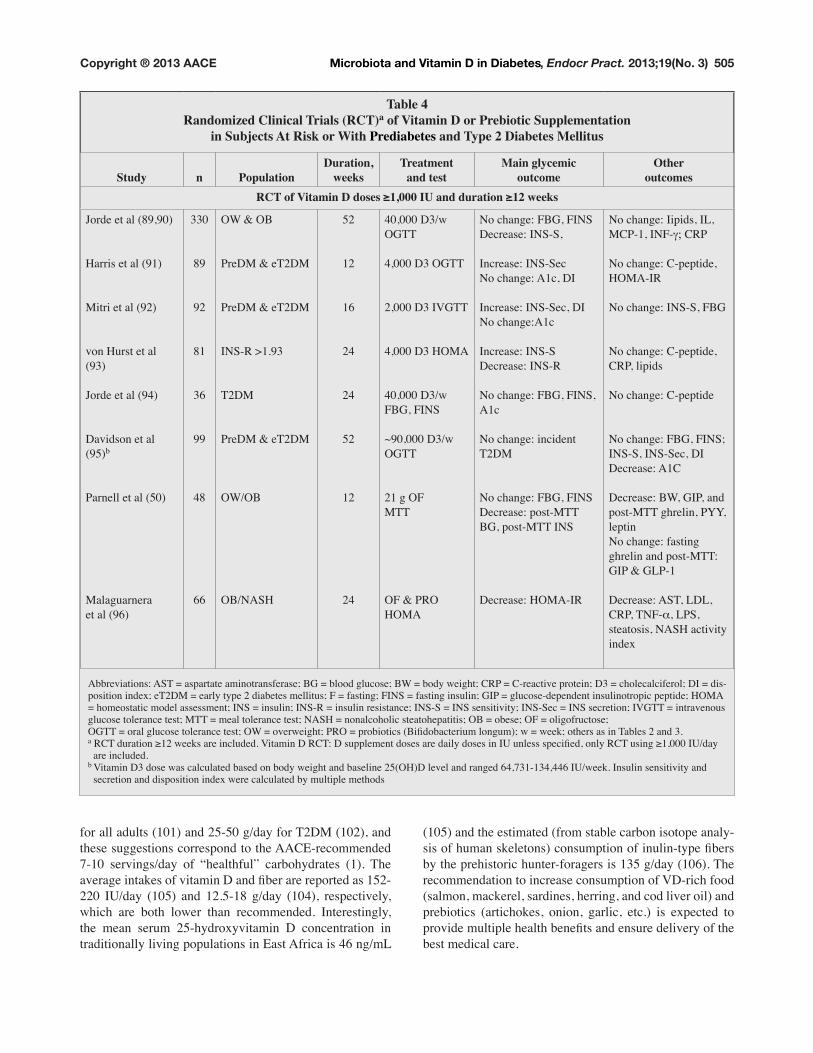
Several RCTs in subjects with prediabetes and T2DM
are underway to prove causality of VD levels in these con-
ditions (90). The results from a few published clinical tri-
als are controversial (Table 4) (89-95) and do not clearly
HVWDEOLVK�WKH�EHQHÀWV�RI�9'�VXSSOHPHQWDWLRQ�IRU�LPSURY-
ing glucose homeostasis, proving the sentiment of Thomas
+HQU\�+X[OH\��ZKR�VDLG��´7KH�JUHDW�WUDJHG\�RI�VFLHQFH�³�the slaying of a beautiful hypothesis by an ugly fact.”
Prebiotics Interaction With Gut Microbiota and Prebiotic Use in Randomized Clinical Trials for
Improving Glucose Homeostasis Prebiotics (11) are nondigestible fermentable food
LQJUHGLHQWV�WKDW�DUH�DEOH�WR�FKDQJH�WKH�JXW�PLFURÁRUD�FRP-
SRVLWLRQ�DQG�RU� DFWLYLW\� LQ� D�ZD\� WKDW� LV�EHQHÀFLDO� WR� WKH�KRVW��7DEOHV���DQG�������������3UHELRWLFV�DUH�QRW�GLJHVWHG�and reach the large bowel where they are fermented by
GutM to produce SCFAs (acetate, propionate, and butyr-
ate), L-lactate, CO2, hydrogen, methane, and other metabo-
lites that regulate downstream metabolic processes in mul-
WLSOH�ZD\V��7KH�EHQHÀFLDO�SURSHUWLHV�RI�SUHELRWLFV�LQFOXGH�their contribution to reducing constipation and weight loss,
improving levels of glucose and lipids, and exerting an anti-
carcinogenic effect (11). The most studied prebiotics are
complex carbohydrates (i.e., fructans and arabinoxylans).
Fructans are polymers of fructose molecules (97),
composed of linear chains of fructose units linked by `
glycosidic bonds and typically terminating in a glucose
unit. There are short- chain (oligofructose) and long-chain
(polyfructose, i.e., inulin and levan) fructans, and they
are typically found in the roots and serve as energy reser-
voirs for many plants instead of starch. An arabinoxylan
is a hemicellulose, a copolymer of two pentose sugars,
arabinose and xylose. It is found in cell walls of plants,
LQFOXGLQJ�FHUHDO�JUDLQ�DQG�FKLHÁ\�VHUYHV�D�VWUXFWXUDO�UROH�(97). Fructans belong to soluble and arabinoxylans belong
WR�VROXEOH�DQG�LQVROXEOH�GLHWDU\�ÀEHUV��%RWK�IUXFWDQV�DQG�arabinoxylans change GutM composition in that they
SURPRWH�WKH�SUROLIHUDWLRQ�RI�EHQHÀFLDO�%LÀGREDFWHULD�DQG�Lactobacilli (Table 3).
The examples of raw natural food particularly rich in
prebiotics (percent content by weight) include chicory root
�a������-HUXVDOHP�DUWLFKRNH��a������EDUOH\��������JDUOLF�
�a������ RQLRQ� �a������ JOREH� DUWLFKRNH� �a����� U\H� EUDQ�RU� JUDLQ� �a�����ZKHDW� EUDQ� �a����� DQG� DVSDUDJXV� ������DQG�H[DPSOHV�RI�FRRNHG�IRRG�DUH�FKRFRODWH������DQG�ZKLWH�EUHDG��a������������ Since Roberfroid introduced the concept of prebiotics
in 1995 (11), in vitro and in vivo models have produced
convincing evidence of the profound effects of prebiot-
ics on energy and glucose homeostasis (Tables 3 and 4).
Some of the mechanisms of these effects remain to be fur-
WKHU�FODULÀHG�EXW�LQFOXGH�WKH�FKDQJH�RI�*XW0�FRPSRVLWLRQ�(e.g., decreased Firmicutes and increased Bacteroidetes),
increased levels of satietogenic gut peptides, decreased
V\VWHPLF� LQÁDPPDWLRQ�� DQG� LPSURYHG� JOXFRVH� WROHUDQFH�(Table 3). These data prompted RCTs that produced prom-
ising but variable results (Table 4) that require further
FRQÀUPDWLRQ�
CONCLUSION
The role of GutM in pathogenesis of prediabetes and
diabetes, as well as the contribution of VD and prebiot-
ics to this process are inadequately understood. Available
data suggest that an “obesogenic” diet leads to the initial
event of microbiota transformation from symbiosis to
dysbiosis. Microbial antigens, such as Gram(-) bacteria
and LPS, translocate to the host interior and trigger TLR
complex activation with subsequent macrophage tissue
LQÀOWUDWLRQ� DQG� LQÁDPPDWRU\� SDWKZD\V� VLJQDOLQJ�� 7KH�“double hit” of steatosis (ectopic fat accumulation) and
´²LWLVµ��LQÁDPPDWLRQ��DQG�FRQWULEXWLRQ�RI�´FR�ULVNVµ��H�J���VDD) are required to activate molecular signaling, includ-
ing impaired insulin signaling and secretion that ends with
diabetes and diseases associated with T2DM (Fig. 1). The
awareness that VDD may play a role only as a contrib-
uting factor helps to understand the failure of recent ran-
domized trials of VD supplementation in prediabetes and
indicates that the search for an appropriate intervention or
intervention combination (e.g., VD and prebiotics) and an
appropriate time window (prior to the process becoming
irreversible) should continue.
The complexity of multidirectional effects and mecha-
nisms of diabetes pathogenesis remains to be elucidated.
In the interim, the current professional society guide-
OLQHV�FDQ�EH�XVHG�IRU�UHFRPPHQGLQJ�9'�DQG�GLHWDU\�ÀEHU�daily intake. For VD, the Institute of Medicine (IOM),
the Endocrine Society, and the American Association of
Clinical Endocrinologists (AACE) Recommended Daily
$OORZDQFHV��5'$V��DUH�����DQG�����,8�GD\�IRU�DJHV�������\HDUV�DQG�����\HDUV��UHVSHFWLYHO\��������,8�GD\�LV�UHFRP-
mended for obese adults, while raising the blood level of
25-hydroxyvitamin D above 30 ng/mL may require 1,500-
������ ,8�GD\� IRU� DOO� DGXOWV� DQG�������� ,8�GD\� IRU� REHVH�DGXOWV��������)RU�ÀEHU��HQGRUVHG�E\�WKH�,20��WKH�$PHULFDQ�Diabetes Association, and other groups) the Recommended
Adequate Intake (RAI) is 25-38 g/day (14 g/1,000 kcal/day)
505
for all adults (101) and 25-50 g/day for T2DM (102), and
these suggestions correspond to the AACE-recommended
7-10 servings/day of “healthful” carbohydrates (1). The
DYHUDJH�LQWDNHV�RI�YLWDPLQ�'�DQG�ÀEHU�DUH�UHSRUWHG�DV��������� ,8�GD\� ������ DQG� �������� J�GD\� ������� UHVSHFWLYHO\��which are both lower than recommended. Interestingly,
the mean serum 25-hydroxyvitamin D concentration in
WUDGLWLRQDOO\�OLYLQJ�SRSXODWLRQV�LQ�(DVW�$IULFD�LV����QJ�P/�
(105) and the estimated (from stable carbon isotope analy-
VLV�RI�KXPDQ�VNHOHWRQV��FRQVXPSWLRQ�RI�LQXOLQ�W\SH�ÀEHUV�E\�WKH�SUHKLVWRULF�KXQWHU�IRUDJHUV�LV�����J�GD\��������7KH�recommendation to increase consumption of VD-rich food
(salmon, mackerel, sardines, herring, and cod liver oil) and
prebiotics (artichokes, onion, garlic, etc.) is expected to
SURYLGH�PXOWLSOH�KHDOWK�EHQHÀWV�DQG�HQVXUH�GHOLYHU\�RI�WKH�best medical care.
Table 4Randomized Clinical Trials (RCT)a of Vitamin D or Prebiotic Supplementation
in Subjects At Risk or With Prediabetes and Type 2 Diabetes Mellitus
Study n PopulationDuration,
weeksTreatment
and testMain glycemic
outcomeOther
outcomes5&7�RI�9LWDPLQ�'�GRVHV��������,8�DQG�GXUDWLRQ�����ZHHNV
Jorde et al (89,90)
+DUULV�HW�DO�����
Mitri et al (92)
YRQ�+XUVW�HW�DO�(93)
Jorde et al (94)
Davidson et al
(95)b
Parnell et al (50)
Malaguarnera
et al ����
330
89
92
81
��
99
48
��
OW & OB
PreDM & eT2DM
PreDM & eT2DM
,16�5�!�����
T2DM
PreDM & eT2DM
OW/OB
2%�1$6+
52
12
��
24
24
52
12
24
40,000 D3/w
OGTT
4,000 D3 OGTT
2,000 D3 IVGTT
������'��+20$
40,000 D3/w
)%*��),16
~90,000 D3/w
OGTT
21 g OF
MTT
OF & PRO
+20$
1R�FKDQJH��)%*��),16
'HFUHDVH��,16�6��
,QFUHDVH��,16�6HF�1R�FKDQJH��$�F��',
,QFUHDVH��,16�6HF��',�1R�FKDQJH�$�F
,QFUHDVH��,16�6'HFUHDVH��,16�5
1R�FKDQJH��)%*��),16��A1c
1R�FKDQJH��LQFLGHQW�T2DM
1R�FKDQJH��)%*��),16Decrease: post-MTT
%*��SRVW�077�,16�
'HFUHDVH��+20$�,5
1R�FKDQJH��,LSLGV��,/��0&3����,1)�a� CRP
1R�FKDQJH��&�SHSWLGH��+20$�,5
1R�FKDQJH��,16�6��)%*
1R�FKDQJH��&�SHSWLGH��CRP, lipids
1R�FKDQJH��&�SHSWLGH
1R�FKDQJH��)%*��),16��,16�6��,16�6HF��',Decrease: A1C
Decrease: BW, GIP, and
post-MTT ghrelin, PYY,
leptin
1R�FKDQJH��IDVWLQJ�ghrelin and post-MTT:
GIP & GLP-1
Decrease: AST, LDL,
&53��71)�_, LPS,
VWHDWRVLV��1$6+�DFWLYLW\�index
Abbreviations: AST = aspartate aminotransferase; BG = blood glucose; BW = body weight; CRP = C-reactive protein; D3 = cholecalciferol; DI = dis-
SRVLWLRQ�LQGH[��H7�'0� �HDUO\�W\SH���GLDEHWHV�PHOOLWXV��)� �IDVWLQJ��),16� �IDVWLQJ�LQVXOLQ��*,3� �JOXFRVH�GHSHQGHQW�LQVXOLQRWURSLF�SHSWLGH��+20$� �KRPHRVWDWLF�PRGHO�DVVHVVPHQW��,16� �LQVXOLQ��,16�5� �LQVXOLQ�UHVLVWDQFH��,16�6� �,16�VHQVLWLYLW\��,16�6HF� �,16�VHFUHWLRQ��,9*77� �LQWUDYHQRXV�JOXFRVH�WROHUDQFH�WHVW��077� �PHDO�WROHUDQFH�WHVW��1$6+� �QRQDOFRKROLF�VWHDWRKHSDWLWLV��2%� �REHVH��2)� �ROLJRIUXFWRVH��2*77� �RUDO�JOXFRVH�WROHUDQFH�WHVW��2:� �RYHUZHLJKW��352� �SURELRWLFV��%LÀGREDFWHULXP�ORQJXP���Z� �ZHHN��RWKHUV�DV�LQ�7DEOHV���DQG���a�5&7�GXUDWLRQ�����ZHHNV�DUH�LQFOXGHG��9LWDPLQ�'�5&7��'�VXSSOHPHQW�GRVHV�DUH�GDLO\�GRVHV�LQ�,8�XQOHVV�VSHFLÀHG��RQO\�5&7�XVLQJ��������,8�GD\� are included. b 9LWDPLQ�'��GRVH�ZDV�FDOFXODWHG�EDVHG�RQ�ERG\�ZHLJKW�DQG�EDVHOLQH����2+�'�OHYHO�DQG�UDQJHG����������������,8�ZHHN��,QVXOLQ�VHQVLWLYLW\�DQG� secretion and disposition index were calculated by multiple methods

506
Fig. 1. The microbiota and T2DM and related diseases. The “obesogenic” diet results into detrimental change
in microbiota, the initial event of pathologic metabolic processes. The “double hit” of steatosis (ectopic fat
DFFXPXODWLRQ��DQG�´²LWLVµ��LQÁDPPDWLRQ��DQG�´FRULVNVµ��H�J���9'�GHÀFLHQF\��DUH�UHTXLUHG�IRU�WKH�SURJUHV-VLRQ�WR�WKH�GLVHDVH��7�'0��&9'��DQG�1$6+��'LHWDU\�FKDQJHV��H�J���SUHELRWLFV��9'�VXSSOHPHQWDWLRQ��PD\�prevent or ameliorate the process if started at the appropriate time prior to the process becoming irrevers-
ible. The ovals represent the “states,” rectangles represent the “processes,” and double lines highlight the
RXWFRPHV��$EEUHYLDWLRQV��&9'� �FDUGLRYDVFXODU�GLVHDVH��/36� �OLSRSRO\VDFFKDULGH��1$6+� �QRQ�DOFRKROLF�steatohepatitis; TLR = Toll-like receptor; VD = vitamin D; VDR = vitamin D receptor.

507
DISCLOSURE
The author has no multiplicity of interest to disclose.
REFERENCES
1. Handelsman Y, Mechanick JI, Blonde L, et al. American
Association of Clinical Endocrinologists Medical
Guidelines for clinical practice for developing a diabetes
mellitus comprehensive care plan. Endocr Pract. 2011;17
(Suppl 2):1-53.
2. Barengolts E. Vitamin D role and use in prediabetes.
Endocr Pract������������������� ��� 1,+� +XPDQ� 0LFURELRWD� 5RDGPDS� 3URMHFW��$YDLODEOH� DW��
http://nihroadmap.nih.gov/hmp/. Accessed December 12,
2012.
4. Human Microbiome Project Consortium. Structure,
function and diversity of the healthy human microbiome.
Nature.������������������ 5. Cani PD, Amar J, Iglesias MA, et al. Metabolic endo-
toxemia initiates obesity and insulin resistance. Diabetes. ������������������
� �� Defronzo RA. Banting Lecture. From the triumvirate to the
ominous octet: a new paradigm for the treatment of type 2
diabetes mellitus. Diabetes. 2009;58:773-795.
7. Guarner F, Malagelada JR.�*XW�ÁRUD� LQ�KHDOWK�DQG�GLV-ease. Lancet.�������������������
8. Vijay-Kumar M, Aitken JD, Carvalho FA, et al. Metabolic syndrome and altered gut microbiota in mice
lacking Toll-like receptor 5. Science. 2010;328:228-231.
9. Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial
ecology: human gut microbes associated with obesity.
Nature.��������������������� 10. Turnbaugh PJ, Ridaura VK, Faith JJ, Rey FE, Knight
R, Gordon JI. The effect of diet on the human gut micro-
biome: a metagenomic analysis in humanized gnotobiotic
mice. Sci Transl Med. ��������UD��� 11. Roberfroid M, Gibson GR, Hoyles L, et al. Prebiotic
HIIHFWV�� PHWDEROLF� DQG� KHDOWK� EHQHÀWV�� Br J Nutr. ���������6XSSO����6��6���
12. Cani PD, Lecourt E, Dewulf EM, et al. Gut microbiota
fermentation of prebiotics increases satietogenic and incre-
tin gut peptide production with consequences for appetite
sensation and glucose response after a meal. Am J Clin Nutr.�������������������
13. Worthley DL, Le Leu RK, Whitehall VL, et al. A human,
double-blind, placebo-controlled, crossover trial of prebi-
otic, probiotic, and synbiotic supplementation: effects on
OXPLQDO��LQÁDPPDWRU\��HSLJHQHWLF��DQG�HSLWKHOLDO�ELRPDUN-
ers of colorectal cancer. Am J Clin Nutr.����������������� 14. Cremonini F, Di Caro S, Nista EC, et al. Meta-analysis: the
effect of probiotic administration on antibiotic-associated
diarrhoea. Aliment Pharmacol Ther.������������������� 15. Hill MJ.�,QWHVWLQDO�ÁRUD�DQG�HQGRJHQRXV�YLWDPLQ�V\QWKHVLV��
Eur J Cancer Prev.��������6XSSO����6���6��� ���� Manning BW, Cerniglia CE, Federle TW.
Biotransformation of 1-nitropyrene to 1-aminopyrene and
1�IRUP\O���DPLQRS\UHQH� E\� WKH� KXPDQ� LQWHVWLQDO� PLFUR-
biota. J Toxicol Environ Health.����������������� 17. Lawler DF, Larson BT, Ballam JM, et al. Diet restric-
tion and ageing in the dog: major observations over two
decades. Br J Nutr. 2008;99:793-805.
18. Jumpertz R, Le DS, Turnbaugh PJ, et al. Energy-balance
studies reveal associations between gut microbes, caloric
load, and nutrient absorption in humans. Am J Clin Nutr. ���������������
19. Mehta NN, Heffron SP, Patel PN, et al. A human model
RI� LQÁDPPDWRU\� FDUGLR�PHWDEROLF� G\VIXQFWLRQ�� D� GRXEOH�blind placebo-controlled crossover trial. J Transl Med. 2012; 10:124.
20. Creely SJ, McTernan PG, Kusminski CM, et al. Lipopolysaccharide activates an innate immune system
response in human adipose tissue in obesity and type 2 diabe-
tes. Am J Physiol Endocrinol Metab. 2007;292:E740-E747.
21. Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut micro-
biome with increased capacity for energy harvest. Nature. �������������������
22. Turnbaugh PJ, Hamady M, Yatsunenko T, et al. A core
gut microbiome in obese and lean twins. Nature. 2009;
457:480-484.
23. Mai V, McCrary QM, Sinha R, Glei M. Associations
between dietary habits and body mass index with gut
microbiota composition and fecal water genotoxicity: an
observational study in African American and Caucasian
American volunteers. Nutr J. 2009;8:49.
24. Kalliomäki M, Collado MC, Salminen S, Isolauri E. Early differences in fecal microbiota composition in chil-
dren may predict overweight. Am J Clin Nutr. 2008;87:
534-538.
25. Nadal I, Santacruz A, Marcos A, et al. Shifts in clos-
tridia, bacteroides and immunoglobulin-coating fecal bac-
teria associated with weight loss in obese adolescents. Int J Obes. (Lond.) �����������������
��� Zhang H, DiBaise JK, Zuccolo A, et al. +XPDQ� JXW�microbiota in obesity and after gastric bypass. Proc Natl Acad Sci. U S A �������������������
27. Serino M, Fernández-Real JM, Fuentes EG, et al. The
JXW�PLFURELRWD�SURÀOH� LV�DVVRFLDWHG�ZLWK� LQVXOLQ�DFWLRQ� LQ�humans. Acta Diabetol.������-XQ�����>(SXE�DKHDG�RI�SULQW@�
28. Erridge C, Attina T, Spickett CM, Webb DJ. A high-fat
meal induces low-grade endotoxemia: evidence of a novel
PHFKDQLVP�RI�SRVWSUDQGLDO�LQÁDPPDWLRQ��Am J Clin Nutr. �������������������
29. Al-Attas OS, Al-Daghri NM, Al-Rubeaan K, et al. Changes in endotoxin levels in T2DM subjects on anti-dia-
betic therapies. Cardiovasc Diabetol. 2009;8:20. 30. Larsen N, Vogensen FK, van den Berg FW, et al. Gut
microbiota in human adults with type 2 diabetes differs
from non-diabetic adults. PLoS One. 2010;5:e9085.
31. Wu X, Ma C, Han L, et al. Molecular characterisation of
the faecal microbiota in patients with type II diabetes. Curr Microbiol. ��������������
32. Furet JP, Kong LC, Tap J, et al. Differential adaptation of
human gut microbiota to bariatric surgery-induced weight
ORVV�� OLQNV� ZLWK� PHWDEROLF� DQG� ORZ�JUDGH� LQÁDPPDWLRQ�markers. Diabetes. 2010;59:3049-3057.
33. Everard A, Lazarevic V, Derrien M, et al. Responses of
gut microbiota and glucose and lipid metabolism to pre-
biotics in genetic obese and diet-induced leptin-resistant
mice. Diabetes.������������������� 34. Le Poul E, Loison C, Struyf S, et al. Functional charac-
terization of human receptors for short chain fatty acids
and their role in polymorphonuclear cell activation. J Biol Chem. 2003;278:25481-25489.
35. Samuel BS, Shaito A, Motoike T, et al. Effects of the
gut microbiota on host adiposity are modulated by the
short-chain fatty-acid binding G protein-coupled receptor,
Gpr41. Proc Natl Acad Sci U S A.���������������������� ��� Bäckhed F, Ding H, Wang T, et al. The gut microbiota
as an environmental factor that regulates fat storage. Proc Natl Acad Sci U S A 2004;101:15718-15723.

508
37. Cani PD, Possemiers S, Van de Wiele T, et al. Changes
LQ�� JXW� PLFURELRWD� FRQWURO� LQÁDPPDWLRQ� LQ� REHVH� PLFH�through a mechanism involving GLP-2-driven improve-
ment of gut permeability. Gut. 2009;58:1091-1103.
38. Macey MG, Whiley RA, Miller L, Nagamune H. Effect
RQ� SRO\PRUSKRQXFOHDU� FHOO� IXQFWLRQ� RI� D� KXPDQ�VSHFLÀF�cytotoxin, intermedilysin, expressed by Streptococcus
intermedius. Infect Immun.������������������� 39. Amyot J, Semache M, Ferdaoussi M, Fontés G, Poitout
V. Lipopolysaccharides impair insulin gene expression in
isolated islets of Langerhans via Toll-Like Receptor-4 and
1)�gB signalling. PLoS One.��������H������� 40. Virkamäki A, Puhakainen I, Koivisto VA, Vuorinen-
Markkola H, Yki-Järvinen H. Mechanisms of hepatic
and peripheral insulin resistance during acute infections in
humans. J Clin Endocrinol Metab.������������������ 41. Rakoff-Nahoum S, Paglino J, Eslami-Varzaneh F,
Edberg S, Medzhitov R. Recognition of commensal
PLFURÁRUD� E\� WROO�OLNH� UHFHSWRUV� LV� UHTXLUHG� IRU� LQWHVWLQDO�homeostasis. Cell. 2004;118:229-241.
42. Uribe A, Alam M, Johansson O, Midtvedt T, Theodorsson E.�0LFURÁRUD�PRGXODWHV� HQGRFULQH�FHOOV� LQ�the gastrointestinal mucosa of the rat. Gastroenterology. �������������������
43. Hara H, Suzuki T, Kasai T, Aoyama Y, Ohta A. Ingestion
of guar. J Nutr. 1999;129:39-45.
44.� $EUDPV�6$��*ULIÀQ�,-��+DZWKRUQH�.0��HW�DO� A combi-
nation of prebiotic short- and long-chain inulin-type fruc-
tans enhances calcium absorption. Am J Clin Nutr. 2005;82:
��������� 45. Pussinen PJ, Havulinna AS, Lehto M, Sundvall J,
Salomaa V. Endotoxemia is associated with an increased
risk of incident diabetes. Diabetes Care. 2011;34:392-397.
��� Amar J, Burcelin R, Ruidavets JB, et al. Energy intake
is associated with endotoxemia in apparently healthy men.
Am J Clin Nutr. 2008;87:1219-1223.
47. Fernández-Real JM, Broch M, Richart C, Vendrell J, López-Bermejo A, Ricart W. CD14 monocyte receptor,
LQYROYHG�LQ�WKH�LQÁDPPDWRU\�FDVFDGH��DQG�LQVXOLQ�VHQVLWLY-
ity. J Clin Endocrin Metab. 2003;88:1780-1784.
48. Vrieze A, Van Nood E, Holleman F, et al. Transfer of
intestinal microbiota from lean donors increases insu-
lin sensitivity in individuals with metabolic syndrome.
Gastroenterology.������������������H�� 49. Neyrinck AM, Van Hée VF, Piront N, et al. Wheat-
derived arabinoxylan oligosaccharides with prebiotic effect
increase satietogenic gut peptides and reduce metabolic
endotoxemia in diet-induced obese mice. Nutr. Diabetes. 2012;2:e28.
50. Parnell JA, Reimer RA. Weight loss during oligofructose
supplementation is associated with decreased ghrelin and
increased peptide YY in overweight and obese adults. Am J Clin Nutr. 2009;89:1751-1759.
51. Palazzo M, Balsari A, Rossini A, et al. Activation of entero-
endocrine cells via TLRs induces hormone, chemokine,
and defensin secretion. J Immunol.�������������������� 52. Muccioli GG, Naslain D, Bäckhed F, et al. The endocan-
nabinoid system links gut microbiota to adipogenesis. Mol Syst Biol. �����������
53. Bouillon R, Carmeliet G, Verlinden L, et al. Vitamin D
and human health: lessons from vitamin D receptor null
mice. Endocr Rev.������������������ 54. Kallay E, Pietschmann P, Toyokuni S, et al.
Characterization of a vitamin D receptor knockout mouse
DV�D�PRGHO�RI�FRORUHFWDO�K\SHUSUROLIHUDWLRQ�DQG�'1$�GDP-
age. Carcinogenesis. 2001;22:1429-1435.
55. Xue Y, Fleet JC. Intestinal vitamin D receptor is required
for normal calcium and bone metabolism in mice.
Gastroenterology.��������������������� ��� Lechner D, Kallay E and Cross HS. 1alpha,25-dihy-
droxyvitamin D3 downregulates CYP27B1 and induces
CYP24A1 in colon cells. Mol Cell Endocrinol������������������
57. Lagishetty V, Misharin AV, Liu NQ, et al. 9LWDPLQ�'�GHÀ-
ciency in mice impairs colonic antibacterial activity and pre-
disposes to colitis. Endocrinology. 2010;151:2423-2432.
58. Wu S, Liao AP, Xia Y, et al. Vitamin D receptor negatively
UHJXODWHV�EDFWHULDO�VWLPXODWHG�1)�NDSSD%�DFWLYLW\�LQ�LQWHV-tine. Am J Pathol.������������������
59. Gudmundsson GH, Bergman P, Andersson J, Raqib R, Agerberth B. Battle and balance at mucosal surfaces
-the story of Shigella and antimicrobial peptides. Biochem Biophys Res Commun.������������������
� ��� Kong J, Zhang Z, Musch MW, et al. 1RYHO� UROH�RI� WKH�vitamin D receptor in maintaining the integrity of the intes-
tinal mucosal barrier. Am J Physiol Gastrointest Liver Physiol.����������*����*����
� ��� Zhao H, Zhang H, Wu H, et al. Protective role of
�����2+���YLWDPLQ�'�� LQ� WKH�PXFRVDO� LQMXU\� DQG� HSLWKH-lial barrier disruption in DSS-induced acute colitis in mice.
BMC Gastroenterol. 2012;12:57.
� ��� Wang TT, Dabbas B, Laperriere D, et al. Direct and indi-
UHFW�LQGXFWLRQ�E\������GLK\GUR[\YLWDPLQ�'��RI�WKH�12'��CARD15-defensin beta2 innate immune pathway defective
in Crohn disease. J Biol Chem. 2010;285:2227-2231.
� ��� Yu S, Bruce D, Froicu M, Weaver V, Cantorna MT. Failure of T cell homing, reduced CD4/CD8alphaalpha
LQWUDHSLWKHOLDO�O\PSKRF\WHV��DQG�LQÁDPPDWLRQ�LQ�WKH�JXW�RI�vitamin D receptor KO mice. Proc Natl Acad Sci U S A. 2008;105:20834-20839.
� ��� Newmark HL, Yang K, Kurihara N, Fan K, Augenlicht LH, Lipkin M. Western-style diet induced colonic tumors
DQG�WKHLU�PRGXODWLRQ�E\�FDOFLXP�DQG�YLWDPLQ�'�LQ�&��%O���mice: a preclinical model for human sporadic colon cancer.
Carcinogenesis. 2009;30:88-92.
��� Favus MJ.�)DFWRUV�WKDW�LQÁXHQFH�DEVRUSWLRQ�DQG�VHFUHWLRQ�of calcium in the small intestine and colon. Am J Physiol. 1985;248:G147-G157.
� ��� Goff JP, Koszewski NJ, Haynes JS, Horst RL. Targeted
delivery of vitamin D to the colon using `-glucuronides of
YLWDPLQ�'��WKHUDSHXWLF�HIIHFWV�LQ�D�PXULQH�PRGHO�RI�LQÁDP-
matory bowel disease. Am J Physiol Gastrointest Liver Physiol.����������*��������
� ��� Neyrinck AM, Possemiers S, Druart C, et al. Prebiotic
HIIHFWV�RI�ZKHDW�DUDELQR[\ODQ�UHODWHG�WR�WKH�LQFUHDVH�LQ�ELÀ-
dobacteria, Roseburia and Bacteroides/Prevotella in diet-
induced obese mice. PLoS One.��������H������� ��� McCullogh JS, Ratcliffe B, Mandir N, Carr KE,
Goodlad RA.� 'LHWDU\� ÀEUH� DQG� LQWHVWLQDO� PLFURÁRUD��effects on intestinal morphometry and crypt branching.
Gut.������������������ ��� Pyra KA, Saha DC, Reimer RA. 3UHELRWLF�ÀEHU�LQFUHDVHV�
hepatic acetyl CoA carboxylase phosphorylation and sup-
presses glucose-dependent insulinotropic polypeptide
secretion more effectively when used with metformin in
obese rats. J Nutr. 2012;142:213-220.
70. Fernandes J, Vogt J, Wolever TM. Inulin increases
short-term markers for colonic fermentation similarly in
healthy and hyperinsulinaemic humans. Eur J Clin Nutr. ������������������
71. Kelly D, King T, Aminov R. Importance of microbial
colonization of the gut in early life to the development of
immunity. Mutat Res.����������������

509
72. Round JL, Mazmanian SK. The gut microbiota shapes
intestinal immune responses during health and disease. Nat Rev Immunol. 2009;9:313-323.
73. Eckmann L. Sensor molecules in intestinal innate immu-
nity against bacterial infections. Curr Opin Gastroenterol. ����������������
74. Keita AV, Salim SY, Jiang T, et al. Increased uptake of
non-pathogenic E. coli via the follicle-associated epithe-
lium in longstanding ileal Crohn’s disease. J Pathol. 2008;
215:135-144.
75. Eloranta JJ, Wenger C, Mwinyi J, et al. Association of
a common vitamin D-binding protein polymorphism with
LQÁDPPDWRU\� ERZHO� GLVHDVH�� Pharmacogenet Genomics. ����������������
� ��� Bai YH, Lu H, Hong D, Lin CC, Yu Z, Chen BC. Vitamin
D receptor gene polymorphisms and colorectal cancer
risk: a systematic meta-analysis. World J Gastroenterol. ������������������
77. Overbergh L, Stoffels K, Waer M, Verstuyf A, Bouillon R, Mathieu C. Immune regulation of 25-hydroxyvitamin
'��DOSKD�K\GUR[\ODVH� LQ� KXPDQ� PRQRF\WLF� 7+3�� FHOOV��mechanisms of interferon-gamma-mediated induction. J Clin Endocrinol Metab.��������������������
78. Costa EM, Feldman D. Modulation of 1,25-dihydroxyvi-
tamin D3 receptor binding and action by sodium butyrate
in cultured pig kidney cells (LLC-PK1). J Bone Miner Res. 1987;2:151-159.
79. Zehnder D, Bland R, Chana RS, et al. Synthesis of
1,25-dihydroxyvitamin D(3) by human endothelial cells
LV�UHJXODWHG�E\�LQÁDPPDWRU\�F\WRNLQHV��D�QRYHO�DXWRFULQH�determinant of vascular cell adhesion. J Am Soc Nephrol. ����������������
80. Du T, Zhou ZG, You S, et al. Modulation of monocyte
hyperresponsiveness to TLR ligands by 1,25-dihydroxy-
vitamin D3 from LADA and T2DM. Diabetes Res Clin Pract. 2009;83:208-214.
81. Yang M, Shen Z, Chen D, et al. (IIHFWV�RI�������2+����D (3) on the expressions of vitamin D receptor, STAT5
and cytoskeletal rearrangement in human monocytes
incubated with sera from type 2 diabetes patients and
diabetic nephropathy patients with uremia. ,QÁDPP� 5HV� ����������������
82. Mencarelli A, Cipriani S, Renga B, et al. VSL#3 resets
LQVXOLQ� VLJQDOLQJ� DQG� SURWHFWV� DJDLQVW�1$6+� DQG� DWKHUR-
sclerosis in a model of genetic dyslipidemia and intestinal
LQÁDPPDWLRQ��PLoS One. 2012;7:e45425.
83. Yahav S, Buffenstein R. Cholecalciferol supplementation
alters gut function and improves digestibility in an under-
JURXQG�LQKDELWDQW��WKH�QDNHG�PROH�UDW��+HWHURFHSKDOXV�JOD-ber), when fed on a carrot diet. Br J Nutr.�����������������
84. Chang YC, Liu PH, Tsai YC, et al. Genetic variation in
the carbonyl reductase 3 gene confers risk of type 2 diabe-
tes and insulin resistance: a potential regulator of adipogen-
esis. J Mol Med (Berl). 2012;90:847-858.
85. Roselli della Rovere G, Lapolla A, Sartore G, et al. Plasma lipoproteins, apoproteins and cardiovascular dis-
ease in type 2 diabetic patients. A nine-year follow-up
study. Nutr Metab Cardiovasc Dis.���������������
��� Zmuda EJ, Qi L, Zhu MX, Mirmira RG, Montminy MR, Hai T. The roles of ATF3, an adaptive-response gene,
in high-fat-diet-induced diabetes and pancreatic beta-cell
dysfunction. Mol Endocrinol. 2010;24:1423-1433.
87. Anderson AA, Helmering J, Juan T, et al. Pancreatic islet
H[SUHVVLRQ� SURÀOLQJ� LQ� GLDEHWHV�SURQH� &��%/.6�-� PLFH�reveals transcriptional differences contributed by DBA
ORFL��LQFOXGLQJ�3ODJO��DQG�1QW��Pathogenetics. 2009;2:1.
88. U.S. National Institutes of Health. http://clinicaltrials.
JRY�FW��UHVXOWV"WHUP YLWDPLQ�'�$1'�GLDEHWHV Accessed
December 12, 2012.
89. Jorde R, Sneve M, Torjesen P, Figenschau Y.� 1R�improvement in cardiovascular risk factors in overweight
and obese subjects after supplementation with vitamin D3
for 1 year. J Intern Med.������������������� 90. Jorde R, Sneve M, Torjesen PA, Figenschau Y,
Gøransson LG, Omdal R.� 1R� HIIHFW� RI� VXSSOHPHQWD-tion with cholecalciferol on cytokines and markers of
LQÁDPPDWLRQ�LQ�RYHUZHLJKW�DQG�REHVH�VXEMHFWV��Cytokine. 2010;50:175-180.
91. Harris SS, Pittas AG, Palermo NJ. A randomized, pla-
cebo-controlled trial of vitamin D supplementation to
improve glycaemia in overweight and obese African
Americans. Diabetes Obes Metab. 2012;14:789-794.
92. Mitri J, Dawson-Hughes B, Hu FB, Pittas AG. Effects
of vitamin D and calcium supplementation on pancreatic
` cell function, insulin sensitivity, and glycemia in adults
at high risk of diabetes: the Calcium and Vitamin D for
Diabetes Mellitus (CaDDM) randomized controlled trial.
Am J Clin Nutr.������������������ 93. von Hurst PR, Stonehouse W, Coad J. Vitamin D supple-
mentation reduces insulin resistance in South Asian women
OLYLQJ�LQ�1HZ�=HDODQG�ZKR�DUH�LQVXOLQ�UHVLVWDQW�DQG�YLWDPLQ�'�GHÀFLHQW� �� D� UDQGRPLVHG��SODFHER�FRQWUROOHG� WULDO��Br J Nutr. 2010;103:549-555.
94. Jorde R, Figenschau Y. Supplementation with cholecal-
ciferol does not improve glycaemic control in diabetic sub-
jects with normal serum 25-hydroxyvitamin D levels. Eur J Nutr. 2009;48:349-354.
95. Davidson MB, Duran P, Lee ML, Friedman TC.�+LJK�dose vitamin D supplementation in people with prediabetes
and hypovitaminosis D. Diabetes Care.������������������ ��� Malaguarnera M, Vacante M, Antic T, et al.
%LÀGREDFWHULXP� ORQJXP� ZLWK� IUXFWR�ROLJRVDFFKDULGHV� LQ�patients with non alcoholic steatohepatitis. Dig Dis Sci. 2012;57:545-553.
97. Raninen K, Lappi J, Mykkänen H, Poutanen K. Dietary
ÀEHU�W\SH�UHÁHFWV�SK\VLRORJLFDO�IXQFWLRQDOLW\��FRPSDULVRQ�RI� JUDLQ� ÀEHU�� LQXOLQ�� DQG� SRO\GH[WURVH��Nutr Rev. 2011;
�������� 98. Shepherd SJ, Gibson PR. Fructose malabsorption and
symptoms of irritable bowel syndrome: guidelines for
effective dietary management. J Am Diet Assoc.���������������������
99. Prosky L, Hoebregs H. Methods to determine food inulin
and oligofructose. J Nutr. 1999;129:1418S-1423S.

510
100. Holick MF, Binkley NC, Bischoff-Ferrari HA, et al. (YDOXDWLRQ�� WUHDWPHQW�� DQG� SUHYHQWLRQ� RI� YLWDPLQ�'� GHÀ-
ciency: an Endocrine Society clinical practice guideline. J Clin Endocrinol Metab.��������������������
101. Franz MJ, Bantle JP, Beebe CA, et al. Evidence-based
nutrition principles and recommendations for the treat-
ment and prevention of diabetes and related complications.
Diabetes Care.���������6������102. Anderson JW, Randles KM, Kendall CW, Jenkins DJ.
&DUERK\GUDWH� DQG� ÀEHU� UHFRPPHQGDWLRQV� IRU� LQGLYLGXDOV�with diabetes: a quantitative assessment and meta-analysis
of the evidence. J Am Coll Nutr. 2004;23:5-17.
103. Hill KM, Jonnalagadda SS, Albertson AM, Joshi NA, Weaver CM. Top food sources contributing to vitamin
D intake and the association of ready-to-eat cereal and
breakfast consumption habits to vitamin D intake in
&DQDGLDQV� DQG� 8QLWHG� 6WDWHV� $PHULFDQV�� J Food Sci. ��������+����+�����
104. King DE, Mainous AG III, Lambourne CA. Trends in
GLHWDU\�ÀEHU�LQWDNH�LQ�WKH�8QLWHG�6WDWHV�������������J Acad Nutr Diet.������������������
105. Luxwolda MF, Kuipers RS, Kema IP, Dijck-Brouwer DA, Muskiet FA. Traditionally living populations in East
Africa have a mean serum 25-hydroxyvitamin D concen-
tration of 115 nmol/l. Br J Nutr.������������������������� Leach JD, Sobolik KD.�+LJK�GLHWDU\� LQWDNH�RI�SUHELRWLF�
inulin-type fructans in the prehistoric Chihuahuan Desert.
Br J Nutr.�������������������.




















