
Resonance Raman spectroscopy of red blood cells using near-infrared laser excitation
13
ORIGINAL PAPER Resonance Raman spectroscopy of red blood cells using near-infrared laser excitation Bayden R. Wood & Peter Caspers & Gerwin J. Puppels & Shveta Pandiancherri & Don McNaughton Received: 17 July 2006 / Revised: 21 September 2006 / Accepted: 22 September 2006 / Published online: 7 December 2006 # Springer-Verlag 2006 Abstract Resonance Raman spectra of oxygenated and deoxygenated functional erythrocytes recorded using 785 nm laser excitation are presented. The high-quality spectra show a mixture of enhanced A 1g ,A 2g ,B 1g ,B 2g ,E u and vinyl modes. The high sensitivity of the Raman system enabled spectra from four oxygenation and deoxygenation cycles to be recorded with only 18 mW of power at the sample over a 60-minute period. This low power prevented photo-/thermal degradation and negated protein denatur- ation leading to heme aggregation. The large database consisting of 210 spectra from the four cycles was analyzed with principal components analysis (PCA). The PC1 loadings plot provided exquisite detail on bands associated with the oxygenated and deoxygenated states. The en- hancement of a band at 567 cm -1 , observed in the spectra of oxygenated cells and the corresponding PC1 loadings plot, was assigned to the Fe–O 2 stretching mode, while a band appearing at 419 cm -1 was assigned to the Fe–O–O bending mode based on previous studies. For deoxygenated cells, the enhancement of B 1g modes at 785 nm excitation is consistent with vibronic coupling between band III and the Soret transition. In the case of oxygenated cells, the enhancement of iron-axial out-of-plane modes and non- totally symmetric modes is consistent with enhancement into the y ,z-polarized transition a 1u p ðÞ! d xz þ O 2 p g centered at 785 nm. The enhancement of non-totally symmetric B 1g modes in oxygenated cells suggests vibronic coupling between band IV and the Soret band. This study provides new insights into the vibrational dynamics, electronic structure and resonant enhancement of heme moieties within functional erythrocytes at near-IR excita- tion wavelengths. Keywords Raman spectroscopy . Red blood cells . Near-infrared excitation . Ligand modes . Vibronic coupling Abbreviations PCA principal components analysis PC principal component RERS resonance-enhanced Raman scattering Hb hemoglobin RBCs red blood cells r.p.m revolutions per minute CCD charged coupled device NIR near-infrared HPRM high-performance Raman module Introduction Resonance Raman spectroscopy is a proven tool for elucidating structural information from hemes and other metalloporphyrins [1]. The high symmetry and chromo- phoric structure of hemes results in strong resonance- enhanced Raman scattering (RERS) upon excitation at a number of different wavelengths within electronic absorp- Anal Bioanal Chem (2007) 387:1691–1703 DOI 10.1007/s00216-006-0881-8 Part of this work was first presented at SPEC 2006, Shedding New Light on Disease: Optical Diagnosis for the New Millennium, held at Heidelberg, Germany, on 20–24 May 2006. B. R. Wood (*) : S. Pandiancherri : D. McNaughton Centre for Biospectroscopy and School of Chemistry, Monash University, Clayton, Victoria 3800, Australia e-mail: [email protected] P. Caspers : G. J. Puppels Center for Optical Diagnostics and Therapy, Erasmus MC, University Medical Center, 3000 DR Rotterdam, The Netherlands
Transcript of Resonance Raman spectroscopy of red blood cells using near-infrared laser excitation

ORIGINAL PAPER
Resonance Raman spectroscopy of red blood cells usingnear-infrared laser excitation
Bayden R. Wood & Peter Caspers & Gerwin J. Puppels &
Shveta Pandiancherri & Don McNaughton
Received: 17 July 2006 /Revised: 21 September 2006 /Accepted: 22 September 2006 / Published online: 7 December 2006# Springer-Verlag 2006
Abstract Resonance Raman spectra of oxygenated anddeoxygenated functional erythrocytes recorded using785 nm laser excitation are presented. The high-qualityspectra show a mixture of enhanced A1g, A2g, B1g, B2g, Eu
and vinyl modes. The high sensitivity of the Raman systemenabled spectra from four oxygenation and deoxygenationcycles to be recorded with only 18 mW of power at thesample over a 60-minute period. This low power preventedphoto-/thermal degradation and negated protein denatur-ation leading to heme aggregation. The large databaseconsisting of 210 spectra from the four cycles was analyzedwith principal components analysis (PCA). The PC1loadings plot provided exquisite detail on bands associatedwith the oxygenated and deoxygenated states. The en-hancement of a band at 567 cm!1, observed in the spectraof oxygenated cells and the corresponding PC1 loadingsplot, was assigned to the Fe–O2 stretching mode, while aband appearing at 419 cm!1 was assigned to the Fe–O–Obending mode based on previous studies. For deoxygenatedcells, the enhancement of B1g modes at 785 nm excitation isconsistent with vibronic coupling between band III and theSoret transition. In the case of oxygenated cells, the
enhancement of iron-axial out-of-plane modes and non-totally symmetric modes is consistent with enhancementinto the y,z-polarized transition a1u p! " ! dxz # O2 pg
! "
centered at 785 nm. The enhancement of non-totallysymmetric B1g modes in oxygenated cells suggests vibroniccoupling between band IV and the Soret band. This studyprovides new insights into the vibrational dynamics,electronic structure and resonant enhancement of hememoieties within functional erythrocytes at near-IR excita-tion wavelengths.
Keywords Raman spectroscopy . Red blood cells .
Near-infrared excitation . Ligand modes . Vibronic coupling
AbbreviationsPCA principal components analysisPC principal componentRERS resonance-enhanced Raman scatteringHb hemoglobinRBCs red blood cellsr.p.m revolutions per minuteCCD charged coupled deviceNIR near-infraredHPRM high-performance Raman module
Introduction
Resonance Raman spectroscopy is a proven tool forelucidating structural information from hemes and othermetalloporphyrins [1]. The high symmetry and chromo-phoric structure of hemes results in strong resonance-enhanced Raman scattering (RERS) upon excitation at anumber of different wavelengths within electronic absorp-
Anal Bioanal Chem (2007) 387:1691–1703DOI 10.1007/s00216-006-0881-8
Part of this work was first presented at SPEC 2006, Shedding NewLight on Disease: Optical Diagnosis for the New Millennium, held atHeidelberg, Germany, on 20–24 May 2006.
B. R. Wood (*) : S. Pandiancherri :D. McNaughtonCentre for Biospectroscopy and School of Chemistry,Monash University,Clayton, Victoria 3800, Australiae-mail: [email protected]
P. Caspers :G. J. PuppelsCenter for Optical Diagnostics and Therapy, Erasmus MC,University Medical Center,3000 DR Rotterdam, The Netherlands

tion bands. With the advent of the Raman microscope,investigators have been able to probe heme sites and otherchromophores within single functional human blood cellsincluding lymphocytes [2, 3], granulocytes [4–7] anderythrocytes [8–14]. Previously we reported the occurrenceof strong Raman scattering in red blood cells compared tothat observed in single crystals and concentrated solutionswhen using excitation at 633 nm, and clear spectraldifferences were observed between oxygenated and deox-ygenated cells [8, 9, 11–13, 15]. Most notable werechanges in the “core size” or “spin state” marker bandregion (1650–1500 cm!1) between the two states. Otherdifferences were observed in the methine deformationregion between 1250–1200 cm!1. More recently, it wasdemonstrated that exposure of red cells to prolonged laserpower and/or high temperatures (>42 °C) resulted in ageneral increase in signal-to-noise, along with the enhance-ment of a number of bands [8]. Based on Ramantemperature and power studies along with UV/visiblemeasurements, it was reasoned that the increase in thesignal-to-noise ratio along with a shift in the Soret bandobserved in the electronic spectrum resulted from theformation of heme aggregates in red blood cells in responseto protein denaturation [8]. A series of bands appearing at1398, 1366, 1248 and 972 cm!1 were designated hemeaggregation marker bands and the 1248 cm!1 band wasused to image heme aggregation in cells exposed toprolonged laser exposure [8] and also to monitor hemestacking in single functional sickle cells [14]. In anotherstudy, spectra of red cells orientated horizontally andvertically with respect to the plane of the Petri dish wererecorded using both parallel- and perpendicular-scatteredlight [9]. The results indicate a possible ordering of hememoieties within the cell that is not prevalent in Hb solution[9]. Based on these results, and the fact that only poor-quality spectra can be obtained from single crystals andhighly concentrated solutions of hemoglobin (up to 17 mM),it was postulated that the enhancement observed in the redblood cell at 633 nm excitation could be due to excitonicinteractions occurring throughout the aggregated hemenetwork in the highly concentrated environment of the redcell at high temperatures and/or after prolonged laserexposure [8]. While it has been possible to record spectraat 488, 514, 568 and 633 nm for the system [12], there arefew examples of spectra recorded at near-IR excitationwavelengths, possibly due to the lack of sensitivity of CCDdetectors in this region of the electromagnetic spectrum.Recently Xun-ling et al. [16] reported a spectrum of a singleoxygenated red blood cell obtained using 782 nm excitationand observed bands associated with the protein in addition tothose associated with the porphyrin macrocycle.
In this paper we report high-quality spectra recorded at785 nm from erythrocytes undergoing a series of oxygen-
ation and deoxygenation cycles using a River Diagnostics(Rotterdam, The Netherlands) spectrometer system. Basedon loading plots obtained using principal componentsanalysis (PCA) and previous studies, we assign markerbands for oxygenation, deoxygenation and aggregation.Spectra of RBCs are also compared with spectra of singlecells recorded at excitation wavelengths (488, 514, 568,633 and 782 nm using a Renishaw (Wotton-under-Edge,UK) microscope system. We report the enhancement ofnon-totally symmetric modes in deoxygenated red cells anda combination of out-of-plane iron axial modes and non-totally symmetric modes in oxygenated cells, and explainour results in terms of electronic structure and vibroniccoupling theory.
Experimental procedures
Preparation of blood samples
Ten milliliters of blood was extracted from a healthyvolunteer and placed in a lithium-heparinized anticoagulanttube. The blood was centrifuged (3500 r.p.m. for12 minutes) to form a buffy coat and 100 !L red bloodcells were carefully removed from the hematocrit andplaced in 3 ml of phosphate-buffered saline, which servedas the stock solution. In the first experiment, 10 !L ofpacked cells were added to a 6-cm-diameter aluminumsputter-coated Petri dish that was further coated in 0.01%poly-L-lysine to affix the cells to the bottom of the Petridish. The cells were allowed to settle for ten minutes priorto measurement. In a second experiment, 100 !L from thestock suspension was placed into the centre of an O-ringthat had been affixed to a CaF2 coverslip (250 !m thick)using nail polish. The coverslip was placed onto the quartzwindow (0.8 mm thick) of the spectrometer with a drop ofwater placed in-between the quartz window and coverslipso as to provide surface tension to hold the coverslip inposition (Fig. 1). The actual working distance is 1.5 mm,with the optical path (from the objective, through theimmersion oil and the window) refractive index-matched.The high spatial resolution of the set-up avoids any signalinfluence from the immersion oil. The fused silica signalwas subtracted from the spectra, and no influence of thisband was observed in either the spectra or the PCA loadingplots.
Oxygenation/deoxygenation experiments
In the first experiment, spectra were recorded in theoxygenated and deoxygenated states by flowing drynitrogen gas in a purpose built flow-through cell, aspreviously described [8].
1692 Anal Bioanal Chem (2007) 387:1691–1703

In a second experiment, cells were initially deoxygenat-ed with nitrogen gas using a liquid squeeze bottle filledwith liquid nitrogen (Fig. 2) with a number of spaces placedbetween the N2 flow outlet and the cell suspension toprevent freezing. At frame 25, the N2 gas was removed andthe cells were re-equilibrated with atmospheric oxygen. Atframe 50, the N2 flow was again applied, and it wasremoved at frame 112. From 112 to 149 the cells wereexposed to atmospheric oxygen; from frame 149 to 175 thecells were once again deoxygenated. From 175 to 185 thecells were reoxygenated; they were finally deoxygenatedfrom 185 to 207. Consequently the cells underwent fourdeoxygenation/oxygenation cycles, with a spectrum ac-quired within every ten-second frame.
Raman instrumentation
Raman spectra of single viable erythrocytes were recordedon a Renishaw 2000 system (equipped with a Peltier cooledCCD detector) using a 782 nm excitation line from a diodelaser and a modified BH2-UMA Olympus (Tokyo, Japan)optical microscope with a Zeiss (Oberkochen, Ger-many)!60 water immersion objective. Power at the sample
was "2–3 mW for a 1–2 !m laser spot size. Spectra wererecorded between 1800 and 200 cm!1 at a resolution of "1–2 cm!1. Ten scans were accumulated for each spectrum, andthe laser exposure for each scan was ten seconds.
Spectra of functional erythrocytes were also recorded ona River Diagnostics Model 3510 Skin CompositionAnalyzer system; this is a confocal Raman microspectrom-eter that is fully optimized to perform rapid in vivomeasurements on the human skin. The measurement stageof the Model 3510 is a dedicated NIR-optimized invertedmicroscope. It receives laser light of 785 nm from a Model25000 High Performance Raman Module (HPRM), which isthen focused into the sample by means of a NIR-optimized0.85 NAmicroscope objective. Scattered light is collected bythe same objective and imaged onto the same HPRM-pinhole, through which laser light is emitted into themeasurement stage. The effective spatial depth resolutionof the system is 4.5±0.5 !m (full width at half maximum foran infinitesimal thin planar source). The Model 2500 HPRMhas a wavelength- and polarization-independent signalthroughput (pinhole to detector) of "80% and is equippedwith a NIR-optimized back-illuminated CCD camera. Theaccuracy with which wavenumber shifts can be measureddepends on the accuracy and reproducibility of the calibra-tion of the wavenumber axis. It is largely independent of thespectral resolution. The calibration accuracy and reproduc-ibility of the model 3510 permits the detection of peak shiftsthat are much smaller than 0.1 cm!1, which is well below thespectral resolution of 5 cm!1.
Because of the high signal throughput, the choice ofexcitation wavelength and the inverted microscope config-uration, the system proved to be well-suited to Ramanmeasurements on erythrocytes.
Data analysis
Spectra recorded from the oxygen/deoxygenation experi-ment were processed using PCA. In this study PCA hasbeen used to model the data and to determine the spectralchanges associated with oxygenated and deoxygenatedcells. PCA enables the decomposition of original data intoa model consisting of a signal part and a noise part. Theanalysis is oriented toward modeling a variance–covariancestructure of a data matrix from which the eigenvalues,corresponding to principal components, are extracted. Eachprincipal component (PC) is a linear combination of the nindependent wavenumber variables x1 . . . xn. So, forexample:
PC1 $ a1x1 # % % % # anxn
The first PC accounts for the greatest variance, and socorresponds to the largest eigenvalue. The second PC isorthogonal to the first, with each successive PC being both
LN
N2
M
S
Fig. 2 Apparatus used inoxygenation/deoxygenationexperiments showing a squeezebottle filled with liquid nitrogen.LN, liquid nitrogen; N2, nitrogengas; M, microscope objective; S,blood sample
fused silica window
RBC's deposited here
oil immersionobjective
O-ring
CaF coverslip2
Fig. 1 Experimental set-up for recording Raman spectra of intact cellsusing the River Diagnostics spectrometer
Anal Bioanal Chem (2007) 387:1691–1703 1693

orthogonal to all those preceding, and accounting for adecreasing proportion of the variance. The smallest PCs areusually interpreted as noise components. One can use toolssuch as score and loading plots to present the datadecomposed by PCA. The scores indicate which objectsare responsible for most of the variance in the data. Eachscore represents a whole spectrum plotted in one, two orthree spatial dimensions. A loading plot is used to assigncoefficients to relevant PCs from an x,y-variable data set.Loadings express relations between variables (i.e., wave-number values), and enable one to tell which variables aredominating or influencing the model and how they areassociated with one another. A data matrix consisting of210 spectra with 1600 wavenumber values (2000–400 cm!1) was constructed. The spectra were transformedby performing a multiplicative scatter correction (MSC)and then baseline-corrected using a linear function acrossthe two spectral endpoints using Unscrambler (CAMO,Oslo, Norway). The data was mean-centered and a fullcross-validation principal components analysis (PCA) wasperformed using six PCs.
Results
Figure 3 depicts spectra recorded from single functional redblood cells in the oxygenated and deoxygenated state using
the Renishaw 2000 system with the microscope attachmentand 782 nm excitation. Detailed band assignments, sym-metry terms and local coordinates for bands observed in the785 nm excitation spectrum of oxygenated red blood cellsare detailed in Table 1. The spectra show a mixture ofbands from the porphyrin macrocycle and the proteincomponents. The proteinaceous bands include the amide Iat 1650 cm!1, the CH2/CH3 deformation modes fromprimarily amino acid side chains at 1446 cm!1, and thephenylalanine band at 1003 cm!1. We adopt the D4h
notation system by convention for assignment of the hemebands because this is a closer approximation to theidealized symmetry of the heme group in the oxygenatedferric low-spin state. The spectrum recorded using 782 nmexcitation in oxygenated cells shows a mixture of bandsfrom all symmetry groups, including A1g (!4, !6), B1g (!10,!11, !13, !18), A2g (!19, !20, !21, !22), B2g (!29, !30) and Eu
(!37, !38, !45). The spectra show a number of features thatcharacterize the two different states, which are consistentwith earlier studies on red blood cells [8, 9, 11–13, 15, 17]and hemoglobin solutions [1, 18–20]. In particular, theband at 1637 cm!1 assigned to the B1g mode !10 appears inthe spectra of oxygenated cells but not in deoxygenatedcells. The 1620 cm!1 band is the most intense inoxygenated cells, while the 1545 cm!1 band is the mostintense in deoxygenated cells. In oxygenated cells, thebands at 1372 cm!1 and 1399 cm!1 are more pronounced
Fig. 3 Averaged spectra ofoxygenated and deoxygenatedsingle red blood cells inphosphate-buffered saline,recorded using the Renishawmicroscope system with a waterimmersion objective (2–3 mWof power from laser at sample,ten seconds of exposure, oneaccumulation, four pixelsbinned)
1694 Anal Bioanal Chem (2007) 387:1691–1703

compared to deoxygenated cells, while the 1224 cm!1 bandshifts to 1211 cm!1. While it is possible to record a singlespectrum of a single cell in both the oxygenated anddeoxygenated state with the Renishaw microscope config-uration at 782 nm, it is not possible to record multiplespectra through a series of cycles with this configuration.Multiple spectra recorded on a single cell results in photo-and/or thermal degradation, as previously reported for633 nm excitation [8]. By utilizing the sensitivity of theRiver Diagnostics Model 3510 Skin Composition Analyzersystem with the inverted confocal microscope system andcoverslip arrangement, as depicted in Fig. 2, we were ableto record cells oxygenating and deoxygenating over four
cycles. In this configuration power at a single cell isreduced because the Brownian motion of cells underneaththe coverslip stops the same single cell from being targeted.Moreover, the high sensitivity of the instrument requiresonly minimal power to achieve high-quality spectra in rapidtime. Initial experiments with this system were aimed atdetermining the power required to obtain high-qualityspectra without causing photo- and/or heat damage to thecells. In a recent study [8], it was demonstrated thatprolonged laser power and/or an increase in temperaturecauses the protein to denature and the hemes to stacktogether, giving rise to the enhancement of a number ofbands in the Raman spectra. In particular, one band at
Table 1 Band positions, assignments, symmetry terms and local coordinates for oxygenated and deoxygenated cells recorded using 785 nmexcitation
Oxy 785 nm Deoxy 785 nm Assignment a Symmetry b Local coordinateb
1653 (m) 1650 (m) Amide I !(C=O)+!(C!N)+"(NH2)1639 (s) absent !10 B1g !(C"Cm)asym1620 (s) 1620 (s) !(C=C)vinyl !(Ca=Cb)1604 (s) 1605 (s) !(C=C)vinyl !(Ca=Cb)1582 (m) 1583 (m) !37 Eu !(C"Cm)asym1566 (vs) 1563 (s) !19 B1g !(C#C#)1547 (s) 1548 (vs) !11 B1g !(C#C#)1526 (sh) 1526 (m) !38 Eu !(pyr breathing)1448 (m) 1446 (m) "(CH2/CH3) "(CH2/CH3)1430 (sh) 1430 (sh) !28 B2g !(C"Cm)sym1397 (w) 1397 (w) !20 A2g !(pyr quater-ring)1384 (w) 1384 (vw) !12 B1g !(pyr half-ring)sym1371 (w) 1356 (w) !4 A1g !(pyr half-ring)sym1337 (m) 1336 (m) !41 Eu !(pyr half-ring)sym1322 (m) 1325 (m) ?1306 (m) 1305 (m) !21 B1g "(CmH)1248* 1248* !42 Eu "(CmH)1226 (s) 1223 (m) !13 or !42 B1g or Eu "(CmH)1212 (m) 1213 (s) !5+!18 A1g+B1g "(CmH)1174 (m) 1170 (m) !30 B2g !(pyr half-ring)asym1156 (w) 1156 (w) !44 Eu !(pyr half-ring)asym1127 1128 !5 B1g !(C#–methyl)1082 1085 "(=CbH2)4 "(=CbH2)41074 1072 "(=CbH2)4 "(=CbH2)41047 !(O=O) !(O=O)1031 1031 "(=CbH2)4 "(=CbH2)41003 1003 phenylanaline993 995 !45 Eu !(C#C1)asym974 974 $(CaH=)827 827 $10 B1u $(CmH)789 789 !6 A1g #(pyr breathing)754 754 !15 B1g #(pyr breathing)676 673 !7 A1g "(pyr deform) sym
622 622 ?567 !(Fe–O2) A1g !(Fe–O2)419 "(Fe–O–O) "(Fe–O–O)
aAssignments are based mainly on labeling scheme originally devised by Abe et al. [25] for octaethylporphyrinato-Ni(II);b Local coordinates based mainly on studies by Hu et al. [24] for myoglobin
Anal Bioanal Chem (2007) 387:1691–1703 1695

1248 cm!1 was assigned as a major marker band for proteindenaturation and heme aggregation [8]. Figure 4 depictsmultiple spectra recorded on one spot using 24 mW of laserpower in the 1250–1200 cm!1 region. As more spectra areaccumulated, there is a marked increase in bands at 1248,1337, 1371 and 1398 cm!1, along with a general increase inthe baseline profile in the 1200–1400 cm!1 region, which isconsistent with a recent study using 633 nm excitation [8]and is indicative of protein denaturation resulting in hemeaggregation within the cells. A number of bands also showa decrease in relative intensity compared to the amide Iband after prolonged laser exposure, including bands at1639, 1620, 1547, 1566, 1547, 1212, 1003 and 755 cm!1.The reduction in the band at 1639 cm!1, which is a markerfor oxygenation, indicates that prolonged laser exposureresults in photooxidation. A power series study demon-strated that 18 mW of power at the sample was optimal forobtaining a high signal-to-noise ratio and for minimizingthe formation of heme aggregates. Consequently this powerwas used for all subsequent measurements.
Figure 5a and b shows a series of spectra recordedduring a four cycles of oxygenation and deoxygenation forboth the 1650–1500 cm!1 and the 1250–1200 cm!1
spectral windows. Each frame represents the time taken torecord a spectrum ("10 s). The methine deformation regionis particularly sensitive to changes in oxygen concentrationwithin the cells. As the cells become oxygenated, anincrease in the 1225 cm!1 band is observed along with aconcomitant decrease in the 1212 cm!1 band and vice versawhen the cells are deoxygenated. A number of changes are
also observed in the 1650–1500 cm!1 region; most notableare the increases in the 1639, 1620 and 1565 cm!1 bandswhen the cells become oxygenated, while the 1548 and1526 cm!1 bands appear to increase in the deoxygenatedstate. Figure 6 depicts representative spectra taken at frames1, 25 and 50 showing the deoxygenated, partially or mixedoxygenated and deoxygenated cells and oxygenated cells,respectively. The spectra for the fully oxygenated anddeoxygenated cells (frames 1 and 50) are very similar tothose recorded with the Renishaw microscope system forcells deoxygenated with dry N2 gas, indicating that there isno evidence of freezing or dehydration upon using the“squeeze bottle” and coverslip approach. The band assignedto !4, which appears at 1371 cm!1 in oxygenated RBCs, isshifted to 1356 cm!1 in the deoxygenated cells, consistentwith the results of Yamomoto et al. [21], who applied441.6 nm excitation (near the Soret band) to heme solutionsand reported this band shifting from 1356 cm!1 for reduced(ferrous) heme proteins to 1378–1370 cm!1 for oxidized(ferric) proteins. The fact that both the 1639 cm!1 and the1371 cm!1 bands have totally disappeared after nitrogen-purging indicates that the cell is totally deoxygenated. Themiddle spectrum exhibits both bands, although at a much-reduced intensity. The band at 1225 cm!1, assigned to theC–H methine in-plane bending vibration, loses intensity asthe cells become deoxygenated, whilst the band at1212 cm!1 appears to gain intensity. It is important to notethat the 1248 cm!1 band does not increase in intensity asmore scans are accumulated, indicating that the hemes arenot aggregating and that the protein is not denatured. The
Fig. 4 Spectra recorded for redblood cells on the same spotusing 24 mW of power at thesample and 785 nm excitation,showing the 400–2000 cm!1
region. Bands that increase inresponse to continuous high-power laser exposure are high-lighted with arrows
1696 Anal Bioanal Chem (2007) 387:1691–1703

band at 1547 cm!1 assigned to !11 is dramatically enhancedrelative to the vinyl C=C peak at 1620 cm!1 in thedeoxygenated cells compared to their oxygenated counter-parts. The 1639 cm!1 band is observed in the oxygenatedcells and the middle spectrum (which may contain amixture of oxygenated and deoxygenated cells and/orpartially oxygenated cells), but not in the fully deoxygen-ated cells, consistent with earlier studies using visibleexcitation wavelengths on hemoglobin [18, 19] andfunctional erythrocytes [8, 11, 13]. Figure 7 depicts a seriesof spectra in the low wavenumber region (400–700 cm!1)recorded for every frame taken upon moving from thedeoxygenated state to the oxygenated state. The spectrumof the oxygenated cell shows a relatively strong bandappearing at 567 cm!1, which is assigned to the !(Fe–O2)mode based on an early study by Brunner [22], whoidentified this band in hemoglobin solution using isotopicsubstitution and 488 nm excitation. Another band thatappears at "419 cm!1 in the spectra of oxygenated cells butnot in those of deoxygenated cells correlates well with thepreviously assigned "(Fe–O–O) bending mode [23, 24]. Aslight shift in the band assigned to !7 from 676 cm!1 in thespectra of oxygenated cells to 673 cm!1 in deoxygenatedcells is apparent.
PCA was applied to gain further insight into the spectralchanges that characterize the oxygenation/deoxygenationcycle of functional erythrocytes. The explained varianceplot for all six PCs for all 210 spectra (constituting four
deoxygenation/oxygenation cycles) showed that 45% ofvariance is explained in the first PC, 22% in the second,10% in the third, and the rest in the remaining PCs.Figure 8a shows the PC1 versus PC2 scores plot for all 210spectra recorded through four deoxygenation/oxygenationcycles. It is apparent from the scores plot that there isconsiderable variation in the degree of oxygenationbetween measurements. However, a clear trend is observedin that the positive scores along PC1 are mainly associatedwith individual spectra of deoxygenated cells, while thenegative scores along PC1 correspond mainly to the spectraof oxygenated cells. The corresponding PC1 loadings plot(Fig. 8b) provides important insights into the spectral bandsassociated with the oxygenated and deoxygenated statesand demonstrates the power of this approach for a largedata set. Bands that are correlated with oxygenated cellsinclude 1639, 1620, 1585, 1565, 1399, 1375, 1342 and1225 cm!1. Some of these bands, namely those at 1639,1620, 1565 and 1225 cm!1, are easily identifiable in thespectra of oxygenated cells presented in Fig. 3 and Fig. 6,but the others are not as obvious. It is interesting to note thecorrelation of the 567 and 419 cm!1 bands with theoxygenated cells. The 567 cm!1 band, as mentioned above,is assigned to the !(Fe–O2) stretching mode based onisotopic substitution studies, while the band at 419 cm!1 isassigned to the "(Fe–O–O) [23, 24]. Bands associated withdeoxygenated cells include those at 1605, 1545, 1523,1356, 1212, 1114 and possibly 789 and 749 cm!1. Once
Fig. 5a–b a Spectra recorded for four deoxygenation/oxygenationcycles showing the 1500–1650 cm!1 region using 785 nm excitation.The arrows highlight bands that are particularly sensitive to the change
from the deoxygenated to the oxygenated state. b The same experimentas described in Fig. 4a, except that the methine deformation region isshown
Anal Bioanal Chem (2007) 387:1691–1703 1697
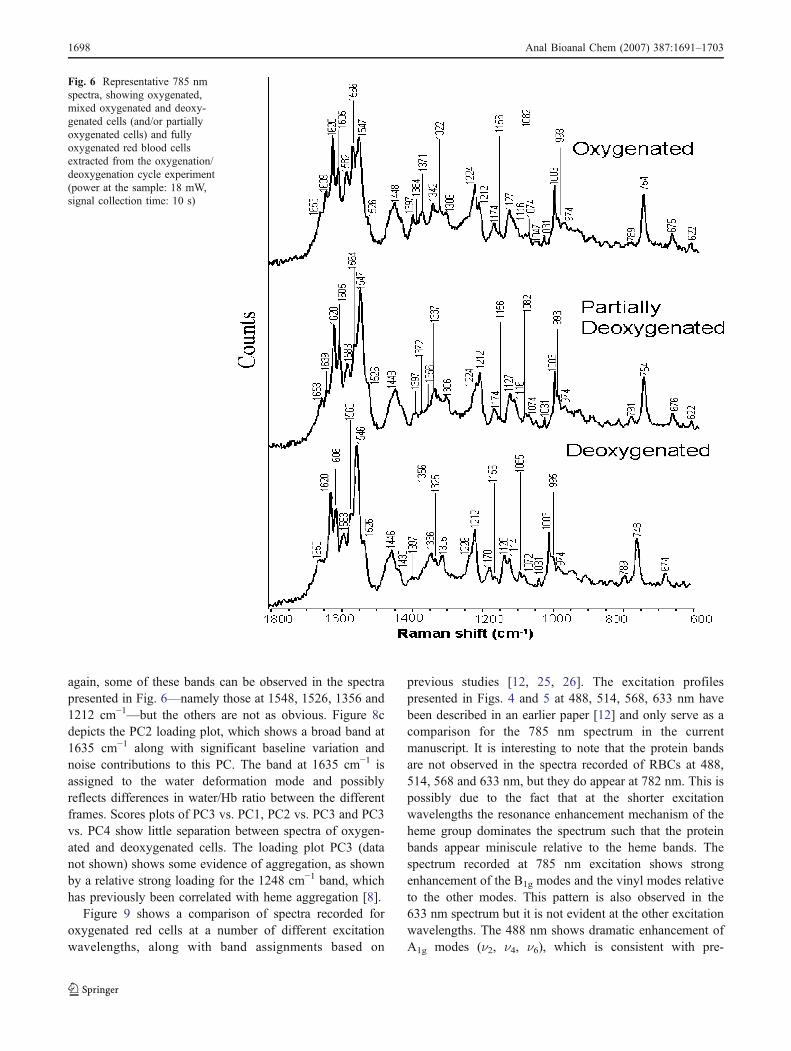
again, some of these bands can be observed in the spectrapresented in Fig. 6—namely those at 1548, 1526, 1356 and1212 cm!1—but the others are not as obvious. Figure 8cdepicts the PC2 loading plot, which shows a broad band at1635 cm!1 along with significant baseline variation andnoise contributions to this PC. The band at 1635 cm!1 isassigned to the water deformation mode and possiblyreflects differences in water/Hb ratio between the differentframes. Scores plots of PC3 vs. PC1, PC2 vs. PC3 and PC3vs. PC4 show little separation between spectra of oxygen-ated and deoxygenated cells. The loading plot PC3 (datanot shown) shows some evidence of aggregation, as shownby a relative strong loading for the 1248 cm!1 band, whichhas previously been correlated with heme aggregation [8].
Figure 9 shows a comparison of spectra recorded foroxygenated red cells at a number of different excitationwavelengths, along with band assignments based on
previous studies [12, 25, 26]. The excitation profilespresented in Figs. 4 and 5 at 488, 514, 568, 633 nm havebeen described in an earlier paper [12] and only serve as acomparison for the 785 nm spectrum in the currentmanuscript. It is interesting to note that the protein bandsare not observed in the spectra recorded of RBCs at 488,514, 568 and 633 nm, but they do appear at 782 nm. This ispossibly due to the fact that at the shorter excitationwavelengths the resonance enhancement mechanism of theheme group dominates the spectrum such that the proteinbands appear miniscule relative to the heme bands. Thespectrum recorded at 785 nm excitation shows strongenhancement of the B1g modes and the vinyl modes relativeto the other modes. This pattern is also observed in the633 nm spectrum but it is not evident at the other excitationwavelengths. The 488 nm shows dramatic enhancement ofA1g modes (!2, !4, !6), which is consistent with pre-
Fig. 6 Representative 785 nmspectra, showing oxygenated,mixed oxygenated and deoxy-genated cells (and/or partiallyoxygenated cells) and fullyoxygenated red blood cellsextracted from the oxygenation/deoxygenation cycle experiment(power at the sample: 18 mW,signal collection time: 10 s)
1698 Anal Bioanal Chem (2007) 387:1691–1703

resonance enhancement from the Soret band where aFranck–Condon (Albrecht A-type) scattering mechanismdominates. Excitation at 568 nm in resonance with the " (orQ0) electronic transition of oxyhemoglobin gives rise to theenhancement of anomalously polarized A2g modes (!19,!20, !21, !22) consistent with vibronic coupling theoryinvolving an Albrecht B-type scattering mechanism. The514 nm excitation spectrum appears to have a mixture ofA1g and A2g modes enhanced relative to the other excitationwavelengths investigated, which indicates pre-resonanceenhancement from both the Soret band and # (or Qv)electronic transitions.
Figure 10 depicts spectra recorded for deoxygenatedcells at various excitation wavelengths, including 785 nm.The 785 nm spectrum is quite different to the other spectra,especially in the 1600–1500 cm!1 region. The 488, 514,568 and 633 nm spectra show the characteristic high-spinstate (S $ 5
2) profile with three major bands at approxi-mately 1545, 1581 and 1608 cm!1. The 785 nm spectrumexhibits additional bands in this region, namely 1526, 1563,1620 cm!1 and the amide I band at 1650 cm!1. Thesebands, along with the band at 1608 cm!1, also appear in thespectra of oxygenated cells, although the 1526 cm!1 ismuch stronger in the deoxygenated state. Both the 1620 and1608 cm!1 bands are assigned to vinyl modes of the hememoiety [17]. It is interesting to note the position of !4,which appears at 1356 cm!1 when excited with 785 nmexcitation, consistent with the Yamamoto et al. [21]oxidation state marker band hypothesis for ferrous hemes.However, as previously reported [12], this hypothesisappears to break down for excitation at 633 nm, wherethe position of this band is observed at 1365 cm!1. At
excitation wavelengths of 514 and 488 nm, !4 appears inthe range consistent with the Yamamoto et al. [21]hypothesis, while at 568 nm excitation the band appearsat 1359 cm!1, which is at the top end of this range.
Discussion
Band assignment
High-quality spectra of red blood cells have been recordedin both the oxygenated and the deoxygenated states forsingle cells at 782 nm using a Renishaw Raman microscopesystem and for a series of oxygenation/deoxygenationcycles at 785 nm using a River Diagnostics Model 3510Skin Composition Analyzer system. The spectra exhibitnumerous bands associated with the porphyrin macrocycleand proteinaceous component of the Hb molecule. Byreducing the power to 18 mW at the sample, hemedenaturation leading to heme aggregation, which is charac-terized by the appearance of the aggregation state markerband at 1248 cm!1 as identified in a recent study [8], wasminimized. This enabled us to record four deoxygenation/oxygenation cycles over a 35-minute period using the RiverDiagnostics system. Spectra associated with oxygenated,deoxygenated and partially deoxygenated cells (or amixture of oxygenated and deoxygenated cells) wereidentified. The partially deoxygenated cells (or mixture ofoxygenated and deoxygenated cells) exhibit features con-sistent with both oxygenation states. In particular, !4appeared at both 1372 cm!1 and 1356 cm!1, consistentwith the iron atoms in both the ferric and ferrous oxidationstates. Another feature that is often used to distinguishoxygenated heme from deoxygenated heme is the shiftingof !10 from "1639 cm!1 in oxygenated heme to"1610 cm!1 in deoxygenated heme. We clearly observe!10 in the oxygenated cells; however, we do not see thisband at all in the deoxygenated cells. The band at1606 cm!1, which appears in deoxygenated cells, cannotbe !10 because it also appears in oxygenated cells. Based onan earlier study [17], and the fact that both the 1620 cm!1
and 1606 cm!1 bands appear in the spectra of oxygenatedand deoxygenated cells, we assign them to the vinyl modes.Further supporting this assignment is the enhancement ofmany Eu modes. Vinyl groups are known to remove thedegeneracy of Eu modes, thereby inducing their Ramanactivity [27]. Analysis of the PC1 loading plot shows thatthe vinyl modes appear more enhanced in the oxygenatedcells compared to the deoxygenated cells. The band at567 cm!1 is assigned to the !(Fe–O2) stretching mode,based on an early study by Brunner [22], who identified asimilar band appearing using 488 nm excitation at567 cm!1 in oxygenated hemoglobin that shifted upon
Fig. 7 A series of spectra in the low wavenumber region (400–700 cm!1), recorded for every frame taken when going from thedeoxygenated state to the oxygenated state
Anal Bioanal Chem (2007) 387:1691–1703 1699

18O2 isotopic substitution, while a band appearing at419 cm!1 was assigned to the "(Fe–O–O) bending mode[23, 24]. This paper provides the first evidence that thesebands appear in the spectra of hemoglobin within functionalerythrocytes. The analysis of four deoxygenation/oxygena-tion cycles under the same conditions provided a largestatistical database for multivariate analysis. Analysis of thePCA PC1 loading plot provided exquisite spectral detail for
the bands associated with the oxygenated and deoxygenatedcells and showed a number of other bands that are notclearly resolved in the averaged spectra. Bands associatedwith positive loadings on the PC1 loading plot wereassociated with deoxygenated cells, while those associatedwith negative loadings were indicative of oxygenated cells.The bands at 567 cm!1 and 419 cm!1 assigned to the ligandmodes dominated the loadings below 600 cm!1.
-0.15
-0.10
-0.05
0
0.05
0.10
0.15
0.20
400 600 800 1000 1200 1400 1600 1800 2000
16391620
15851565
1399
1375
1342
1225
1212
1002755677569419
1605
1545
1523
13561114993789
749
Loa
ding
s
1037
b
a
-0.04
-0.02
0
0.02
0.04
0.06
400 600 800 1000 1 1400 1600 1800 2
Loa
ding
s
PC2
Wavenumber (cm-1)
Wavenumber (cm-1)11200 2000
c
Fig. 8a–c a PCA PC1 vs. PC2 scores plot for spectra from the four cycles of deoxygenation/oxygenation. b Corresponding PC1 loading plot.c PC2 loading plot
1700 Anal Bioanal Chem (2007) 387:1691–1703

Enhancement mechanism
Franzen et al. [28] reported a complete resonanceexcitation profile from 740–780 nm and demonstratedthat non-totally symmetric modes of B1g symmetry areenhanced in the vicinity of a very weak charge transferband centered at 758 nm and designated band III [29].This band is observed in deoxyhemoglobin and deoxy-myoglobin and was assigned by Eaton et al. [29] to thea2u p! " ! dyz transition. This band is more than a 1000times less intense than the Soret band [28], is quite narrow,and models that fit the line shape are reported to exhibitonly weak coupling to a low-frequency vibrational mode[30]. Based on symmetry arguments, Franzen et al. [28]explained the enhancement by adopting a vibroniccoupling model, and not by a Frank–Condon mechanism.In this model, the charge transfer transition is vibronicallycoupled to the Soret band through a low-frequency
vibrational mode of B1g symmetry, giving rise to enhancedB1g modes. In the case of red blood cells in both theoxygenated and the deoxygenated states, a mixture ofmodes including A1g, A2g, B1g, B2g and Eu are allobserved to be relatively enhanced to varying degrees. Itis interesting to note that in the case of red cells excited at785 nm, we observe the enhancement of a band at567 cm!1 assigned to the ! (Fe–O2). The enhancementof the axial ligand–iron bond depends mainly on the azz0
tensor element, the enhancement of which requiresresonance with a z-polarized (out-of-plane) electronictransition [20]. A charge-transfer transition designated byEaton et al.[29] as band IV a1u p! " ! dxz#O2 pg
! "is
centered at 780 nm in oxygenated hemoglobin. This bandhas been reported to have a z-polarized component [29],which could explain the enhancement of the out-of-planeligand mode !(Fe–O2) mode at 567 cm!1 when irradiatingwith 785 nm excitation. The fact that the vinyl modes are
Fig. 9 A comparison of spectrarecorded for oxygenated cellsusing 785, 633, 568, 514 and488 nm laser excitation lines,showing the major band assign-ments in the 1800–600 cm!1
region
Anal Bioanal Chem (2007) 387:1691–1703 1701

also more enhanced in the oxygenated cells versus thedeoxygenated cells is an indicator that these modes alsohave significant out-of-plane character, which would beexplained by enhancement into an electronic charge-transfer transition with a z-polarized component. It wouldappear that enhancement of bands in deoxygenated cellsresults from the vibronic coupling of band III via the Soretband, as suggested by Franzen et al. [28] and indicated bythe strong enhancement of B1g modes. The inclusion ofenhanced B1g modes, ligand modes and out-of-planemodes observed in spectra of oxygenated cells may alsobe explained by a vibronic coupling mechanism, in thiscase involving a y,z-polarized charge-transfer transitionknown as band IV a1u p! " ! dxz#O2 pg
! "and coupled to
the Soret band via a low-frequency B1g mode in a similarmanner to that described by Franzen et al. [28] for band
III. A full resonance Raman profile in the 700–800 nmrange would need to be performed on intact oxygenatedand deoxygenated cells before a more definitive statementcould be made about the enhancement mechanism inoperation in functional erythrocytes.
Conclusion
High-quality spectra of functional cells were recorded in theoxygenated and deoxygenated states for the first time using785 nm excitation. The high sensitivity of the RiverDiagnostics system enabled spectra to be recorded fromfour cycles of oxygenation and deoxygenation using18 mW of power at the sample, thus minimizing photo-and thermal degradation that can result in protein denatur-
Fig. 10 A comparison of spec-tra recorded for deoxygenatedcells using 785, 633, 568, 514and 488 nm laser excitationlines
1702 Anal Bioanal Chem (2007) 387:1691–1703

ation and ultimately heme aggregation. The spectra of bothoxygenated and deoxygenated cells at 785 nm excitationshow a mixture of enhanced vibrational modes includingthe A1g, A2g, B1g B2g modes along with the vinyl modes. Inthe case of oxygenated cells, the enhancement is mostlikely due to enhancing into the y,z-polarized a1u p! " !dxz#O2 pg
! "transition known as band IV, as shown by the
enhancement of iron–ligand modes at 567 and 419 cm!1,which were evident in both the oxygenated spectra and thePC1 loadings plot. These modes are normally only enhancedin the vicinity of z-polarized transitions. The enhancementof non-totally symmetric B1g modes in oxygenated cellswould appear to involve vibronic coupling between bandIVA and the Soret band in a similar way to that describedby Franzen et al. [28] for band III and the Soret band fordeoxygenated myoglobin. In the case of deoxygenatedcells, the enhancement possibly results from enhancing intothe band III region, which is vibronically coupled to theSoret band, as shown by the strong enhancement of B1g
modes [28]. This study indicates that 785 nm excitation canbe used as a nondestructive molecular probe that haspotential as an analytical and diagnostic tool for a variety oferythrocyte disorders. The technique may also haveapplications in monitoring heme perturbation in responseto in vivo drug therapy within functional erythrocytes.
Acknowledgment This work is funded by an Australian ResearchCouncil Discovery Grant. Dr. Wood!s work is supported by anAustralian Synchrotron Program Fellowship Grant and a MonashUniversity Synchrotron Fellowship Grant.
References
1. Spiro TG (ed) (1988) Biological applications of Raman spectros-copy. Wiley, New York
2. Puppels GJ, Olminkhof JHF, Segers-Nolten GMJ, Otto C, Mul deFFM, Greve J (1991) Exp Cell Res 195:361–367
3. Puppels GJ, Garritsen GMJ, Kummer JA, Greve J (1993)Cytometry 14:251–256
4. Van Manen H-J, Kraan YM, Roos D, Otto C (2004) J Phys ChemB 108:18762–18771
5. Puppels GJ, Garritsen GMJ, Segers-Nolten GMJ, De Mul FFM,Greve J (1991) Biophys J 60:436–446
6. Salmaso BLN, Puppels GJ, Caspers PJ, Floris R, Greve J (1994)Biophys J 67:36–446
7. Otto C, Sijisema NM, Greve J (1998) Eur Biophys J 271:582–5898. Wood BR, Hammer L, Davis L, McNaughton D (2004) J Biomed
Opt 10:140059. Wood BR, Hammer L, McNaughton D (2005) Vib Spectrosc
78:71–7810. Wood BR, Langford S, Cooke BM, Glenister FK, Lim J, Duriska
M, McNaughton D (2003) FEBS Lett 554:247–25211. Wood BR, McNaughton D (2002) Biopolymers (Biospectroscopy)
67:259–26212. Wood BR, McNaughton D (2002) J Raman Spectrosc 33:517–52313. Wood BR, Tait B, McNaughton D (2001) Biochim Biophys Acta
1539:58–7014. McNaughton D, Lim J, Langford S, Collie J, Wood BR (2005)
Proc SPIE 5651:52–6015. Wood BR, McNaughton D (2006) In: O’Malley PD (ed) New
developments in sickle cell disease. Nova, New York, pp 63–11916. Yan X-l, Dong R-X, Wang Q-G (2004) Spectrosc Spect Anal
24:576–57817. Hoey S, Brown DH, McConnell AA, Smith WE, Marabani M,
Sturrock RD (1988) J Inorg Biochem 34:189–19918. Brunner H, Mayer A, Sussner H (1972) J Mol Biol 70:153–15619. Spiro TG, Streakas TC (1973) J Am Chem Soc 96:338–34520. Spiro TG (1975) Biochim Biophys Acta 416:169–18921. Yamamoto T, Palmer G (1973) J Biol Chem 248:5211–521322. Brunner H (1974) Naturwissenschaften 61:129–13023. Jeyarajah S, Proniewicz LM, Bronder H, Kincaid JR (1994) J Biol
Chem 269:31047–3105024. Hu S, Kincaid JR (1991) J Am Chem Soc 113:7189–719425. Abe M, KitagawaT, Kyogoku K (1978) J Chem Phys 69:4526–453426. Hu S, Smith KM, Spiro TG (1996) J Am Chem Soc 118:12638–
1264627. Choi S, Spiro TG, Langry KC, Smith KM (1982) J Am Chem Soc
104:433728. Franzen S, Wallace-Williams SE, Shreve AP (2001) J Am Chem
Soc 124:7146–715529. Eaton WA, Hanson LK, Stephens PJ, Sutherland JC, Dunn JBR
(1978) J Am Chem Soc 100:4991–500330. Srajer V, Champion PM (1991) Biochemistry 30:7390–7402
Anal Bioanal Chem (2007) 387:1691–1703 1703




















