
Repression of Protein Kinase C and Stimulation of Cyclic AMP Response Elements by Fumonisin, a...
6
1995;55:1655-1659. Cancer Res Chongxi Huang, Martin Dickman, Gail Henderson, et al. Which Is a Carcinogen Response Elements by Fumonisin, a Fungal Encoded Toxin Repression of Protein Kinase C and Stimulation of Cyclic AMP Updated version http://cancerres.aacrjournals.org/content/55/8/1655 Access the most recent version of this article at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications Research. on October 20, 2014. © 1995 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on October 20, 2014. © 1995 American Association for Cancer cancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Repression of Protein Kinase C and Stimulation of Cyclic AMP Response Elements by Fumonisin, a...

1995;55:1655-1659. Cancer Res Chongxi Huang, Martin Dickman, Gail Henderson, et al. Which Is a CarcinogenResponse Elements by Fumonisin, a Fungal Encoded Toxin Repression of Protein Kinase C and Stimulation of Cyclic AMP
Updated version
http://cancerres.aacrjournals.org/content/55/8/1655
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on October 20, 2014. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on October 20, 2014. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from

[CANCER RESEARCH 55, 1655-1659, April 15, 1995]
Advances in Brief
Repression of Protein Kinase C and Stimulation of Cyclic AMP Response Elementsby Fumonisin, a Fungal Encoded Toxin Which Is a Carcinogen1
Chongxi Huang, Martin Dickman, Gail Henderson, and Clinton Jones2
Department of Plant Pathology [C. H., M. D.}, Center for Biotechnology IG. H., C. ].], and Department of Veterinary and Biomedicai Sciences, Universitv of Nebraska. Lincoln.Nebraska 68583
Abstract
Fusarium monili/orme (FM) is a major fungal pathogen of corn and isinvolved with stalk rot disease. FM is widely spread throughout the world,including the United States. Most strains of FM produce several nij co-
toxins, the most prominent of which is called fumonisin. Recent epidemiológica! studies indicated that ingestion of fumonisin correlates with ahigher incidence of esophageal cancer in Southern and Northern Africaand China. Furthermore, fumonisin causes a neurodegenerative disease inhorses, induces hepatic cancer in rats, and induces pulmonary edema inswine. Considering that high levels of fumonisin have been detected inhealthy and diseased corn grown in the United States, fumonisin may posea health threat to humans and livestock animals. Structurally, fumonisinresembles sphingolipids which are present in the membranes of animaland plant cells. At the present time, very little is known concerning themechanism by which fumonisin elicits its carcinogenic effect. Our studiesindicate that fumonisin represses expression of protein kinase C andAP-1-dependent transcription. In contrast, fumonisin stimulated a simple
promoter containing a single cyclic AMP response element. Since fumonisin did not alter protein kinase A activity, it appears that cyclic AMPresponse element activation was independent of protein kinase A. It ishypothesized that the ability of fumonisin to alter signal transductionpathways plays a role in carcinogenesis.
Introduction
FM3 is a fungal pathogen of corn and is found in all parts of the
world, including the United States. FM is a causative agent of stalk rotdisease, an economically important disease of corn (1-4). Moststrains of this fungus produce a variety of mycotoxins, the major classof which are called fumonisin. Fumonisin is comprised of at least fourclosely related structural analogues, fumonisin Bt_4, which resemblesphingolipids (5). Only B¡and B2 are of toxicological significancebecause B3 and B4 plus the two A toxins are present at very lowconcentrations on plants. Furthermore, B3, B4, A,, and A2 toxinsexhibited no cancer-initiating activity when tested at concentrationswhere B, and B2 were active (6). If rats are fed a diet that contains 50mg/Kg fumonisin Bu primary hepatocellular carcinoma occurs in twoof three of the animals (6-9). Fumonisin also increases the incidenceof esophageal cancer in Africa and China (10), induces equine leu-koencephalomalacia or hepatotoxicity (11), and porcine pulmonaryedema (12). Fumonisin can be detected on healthy plants (13, 14),suggesting that the seriousness of fumonisin in the food chain is
Received 1/18/95; accepted 3/3/95.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1This study was supported by a grant from the Institute of Agriculture and Natural
Resources (University of Nebraska-Lincoln) and the Center for Biotechnology.2 To whom requests for reprints should be addressed, at Department of Veterinary and
Biomedicai Sciences, University of Nebraska Lincoln, P.O. Box 830905, Lincoln, NE66583-0905.
3 The abbreviations used are: FM, Fusarium moniliforme; PKC, protein kinase C;
DAG, diacylglycerol; PKA, protein kinase A; CRE, cAMP response element; CREB,CRE-binding proteins; CMF, calcium/magnesium-free; CAT, chloramphenicol acetyl-transferase; PBS-T, PBS tween.
underestimated. For example, fumonisin is present in substantialamounts, up to 2.7 parts/million, in 74% of U.S. corn-based foodproducts. Taken together, these observations suggest that fumonisinmay be a health risk to humans.
Structurally, fumonisin resembles sphinganine, which is an intermediate in the biosynthesis of the sphingosine backbone of morecomplex sphingolipids such as ceramides, sphingomyelin, cerebro-sides, gangliosides, and sulfatides (Fig. L4; reviewed in Ref. 15).Several studies have demonstrated that the addition of fumonisin tocultured cells or to the diet of rats inhibits sphingolipid biosynthesis(15-17). An important site of inhibition is the reaction catalyzed bysphingosine A'-acetyltransferase (ceramide synthase). Sphingolipidshave been implicated as playing a role in cell contact, growth, anddifferentiation. Since fumonisin bears a remarkable structural similarity to sphingosine (Fig. 1A; Ref. 17) and sphingosine is a potentinhibitor of PKC in vitro (18), it is possible that fumonisin alters PKCactivity. Sphingomyelin has also been hypothesized to initiate a signaltransduction cycle, with ceramide being generated as a second messenger (19). Although the effecters of this cycle have not beenidentified, they may include ceramide-activated protein kinases (20)or phosphatases (21).
PKC, a serine/threonine kinase, is central to one of the majormammalian signal transduction pathways and is activated upon external stimulation of cells by various ligands including hormones,neurotransmittors, and growth factors (22). These external signalsinduce the hydrolysis of membrane-associated inositol phospholipidsgenerating two second messengers, inositol triphosphate and DAG, anendogenous activator of PKC. Phorbol esters, which are capable oftumor promotion, can substitute for DAG in enzyme activation, butunlike DAG, phorbol esters are not rapidly metabolized and thus areuseful for in vitro analysis (23). PKC in mammals is comprised of atleast 12 distinct polypeptides or isoforms (reviewed in Ref. 24). PKCplays a role in differentiation, cell growth, and transcription (25, 26),and there is growing evidence that specific PKC isoforms orchestratethese processes. This hypothesis is supported by the finding thatmarked tissue differences exist among the PKC isotypes. Moreover,the subcellular distribution of PKC ¡soformschanges dramaticallywhen the cell is activated, and specific PKC isoforms have substrateselectivity. Physiologically, activation of PKC is regulated by DAG,which leads to a rapid translocation from the cytosol to the membrane.Since PKC can be found in the nucleus (27), PKC can directlyphosphorylate nuclear proteins. A primary target of PKC-dependentsignal transduction is a as-acting transcriptional motif (AP-1; reviewed in Refs. 24 and 28). A family of transcription factors, including the gene products of the proto-oncogenes Fos and Jun, specificallybind to the AP-1 site, and their activation occurs by a PKC-dependentmechanism. Thus, alterations in PKC activity would have a profoundeffect on cellular processes.
Two major signal transduction pathways are: (a) PKC dependent(reviewed in Ref. 24); and (b) PKA dependent (reviewed in Refs.29-32). PKA-dependent signal transduction is a cAMP-dependent
1655
Research. on October 20, 2014. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from

FUMONISIN REPRESSION OF PKC AND CAMP STIMULATION
process. The cAMP molecule binds to a regulatory subunit of PKA,releasing the regulatory subunit, and PKA is activated. PKA stimulates the transcription of genes which contain a CRE by phosphoryl-ating CREB. Although CREB is the major activator of CRE-depend-
ent transcription, other transcription factors can bind and regulate,positively or negatively, CRE-dependent transcription. Another level
of complexity of CRE dependent transcription is found with CREBwhich can dimerize with members of the ATF family and jun familymembers. Although PKA and PKC are independent signal transduc-tion pathways, there is evidence that "cross talk" occurs between the
two pathways, implying that if one pathway is induced or repressed,the other may be affected.
We hypothesized that fumonisin regulates PKC activity becausefumonisin is structurally similar to a potent inhibitor of PKC, sphin-
gosine (17, 18). To test this hypothesis, the effect of fumonisin onPKC activity was tested in a monkey kidney cell line (CV-1). Fumonisin inhibited PKC activity in a dose-dependent fashion when cells
were treated for 3 or 16 h. Western blotting analysis demonstrated thatseveral specific isoforms of PKC were diminished after fumonisintreatment. In transient transfection assays, fumonisin inhibited AP-1-
dependent transcription, confirming the conclusion that fumonisininhibits PKC activity. However, fumonisin stimulated CRE-dependent transcription in a PKA-independent manner. These results dem
onstrated that fumonisin B, alters signal transduction pathways inCV-1 cells.
Materials and Methods
Fumonisin. Two sources of fumonisin B, were used in these studies:Sigma (>98% pure); or preparations obtained from Dr. R. D. Plattner (USDA,PeorÃa,IL; >90% pure). Identical results were obtained with both preparations.Fumonisin was dissolved in CMF PBS [80 HIMNa2HPO4, 20 mM NaH2PO4,and 100 mM NaCl (pH 7.5)] and stored at 4°Cin the dark.
Cells and Media. CV-1 cells (African green monkey kidney cells) wereobtained from Robert Su (NIH, Bethesda, MD). Cells were grown in Earle's
modified Eagle's medium supplemented with 5% fetal bovine serum. All
media contained penicillin (10 units/ml) and streptomycin (100 mg/ml).Measurement of CAT Activity in Transfected Cells. Cells were trans-
fected with 10 /xg of the designated DNA using CaPO4 precipitation asdescribed previously (33-37). A DMSO or glycerol shock was not usedbecause this treatment altered 12-O-tetradecanoylphorbol-13-acetate response
in CAT assays (33, 34). Forty h posttransfection, CAT activity was measuredas described previously (35-37). Amounts of radioactivity associated with the
various forms of chloramphenicol were determined using a Phosphorimager(Molecular Dynamics, Sunnyvale, CA). Plasmids pBLcat4, 5x col/cat TRE,and CREcat were obtained from Dr. Peter Angel (Kernforschungszentrum
Karlsruhe, Karlsruhe, Germany). A summary of these constructs is presentedin Fig. 4. All plasmids were extracted from bacteria by alkaline lysis followed
by 2 CsCl gradients (38).Measurement of PKC and PKA Activity. PKC activity and PKA activity
was conducted as described previously (39).Western Blot Analysis. CV-1 cells, normal or fumonisin treated, were
washed in CMF-PBS and scraped with a rubber policeman. Cells were pelleted
by centrifuging (2000 RPM for 10 min). The cell pellet was resuspended inCMF-PBS. Total proteins were extracted from cells, and 50 fig protein were
electrophoresed in 8% SDS polyacrylamide gels. Protein concentrations weremeasured by the method of Bradford (40). Proteins were electrophoreticallytransferred to nitrocellulose (Hybond ECL; Amersham) according to the manufacturer's recommendations. Filters were blocked with PBS-T for l h at room
temperature with shaking. Filters were incubated with a mixture of PKCantibodies (a, B, and y) for l h at room temperature in PBS-T. Followingwashing, a horseradish peroxidase-labeled secondary antibody was added, and
the membrane was incubated for l h at room temperature. The filter wassubsequently washed with PBS-T. Bands were visualized by chemilumines-
cence using ECL reagents (Amersham).
Results
Repression of PKC Activity by Fumonisin. Since fumonisinstructurally resembles a potent PKC inhibitor, sphingosine (Refs. 17and 18; Fig. IA), the effect fumonisin has on PKC activity wasmeasured in CV-1 cells. When CV-1 cells were treated for 3 h with 5/LIMfumonisin, PKC activity decreased approximately 2-fold relative
to controls (Fig. IB). Repression of PKC activity was also evidentafter cells were treated with 1 or 5 JUMfumonisin for 16 h. Repressionwas dose dependent since 0.01 JAMhad a negligible effect and 0.1 /XMonly inhibited 15% after 3 h treatment. In contrast, PKA levels werenot affected by any fumonisin concentration used in these studies.These studies indicated that fumonisin inhibited PKC activity in CV-1
cells after only 3 h treatment and repression required at least l /J.Mfumonisin.
The effect fumonisin has on steady-state levels of PKC was as
sessed by Western blot analysis. A panel of antibodies directed againstthe major isoforms of PKC (a, ß,and y) were used for these studies.A Mr 120,000 protein was consistently repressed after CV-1 cells
were treated with fumonisin (1 or 5 JLIM)for 3 or 16 h (Fig. 2). Inaddition, two proteins migrating with an apparent molecular weight of70,000 and 83,000 were repressed when cells were treated with 1 JIMfumonisin for 3 h. However, levels of a protein migrating with anapparent molecular weight of 65,000 was not altered by fumonisin.Since there are at least 12 different isoforms of PKC in mammals(reviewed in Ref. 24), isotype-specific antibodies were used to iden
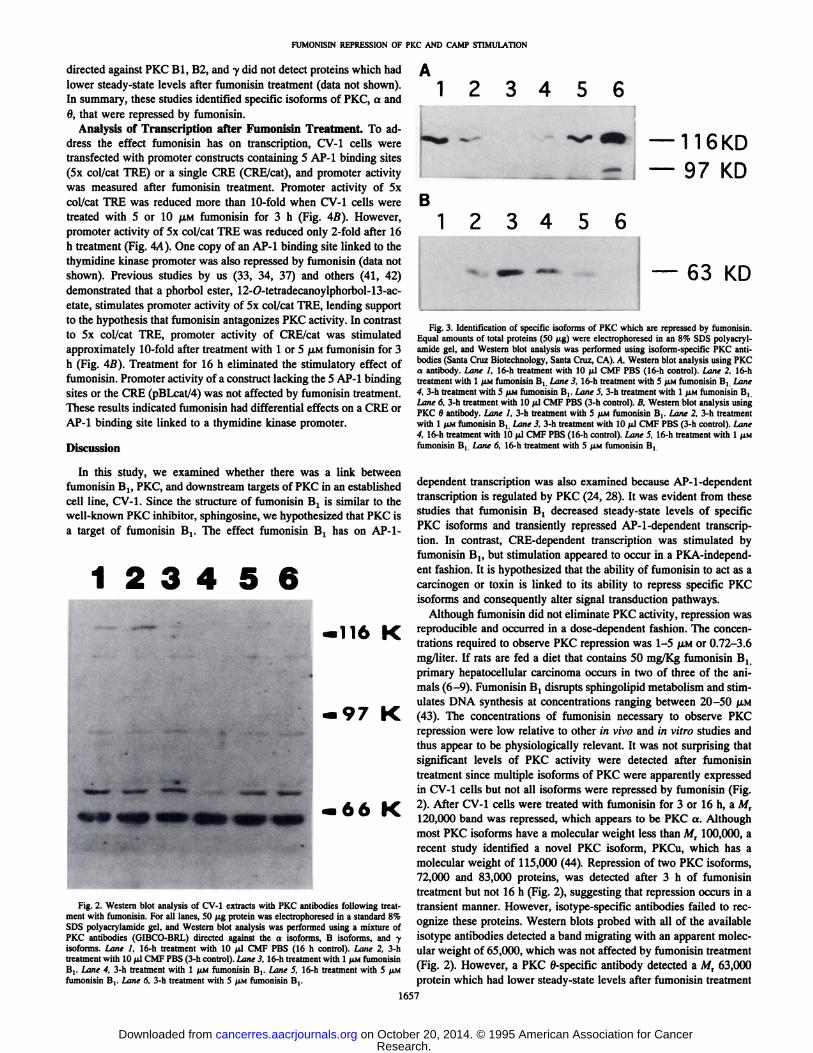
tify which PKC isoforms were repressed by fumonisin. A PKC aantibody recognized a Mr 120,000 protein which was repressed byfumonisin (Fig. 3A). A PKC 6 antibody recognized a M, 63,000protein which was repressed by fumonisin (Fig. 3B). Antibodies
A.Sphingosine
II COOH0
B. PKC act. PKflact.
CnF PBS Controls 100±i I00±7
0.01 uH Fuionlsln 81 (3 H) 95±6 ND
0.1 u!)Fuionlaln Bl (3 H) 80±7 HD
I un Fusioni!in 91 (3 H) 1V»6 96*6
S un Fuionlsln Bl (3 H) 13*5 95±7
IO u«Fuionlsln Bl (3 H) 37*8 ND
I un FusionistiBl (I6 H) 5I.«7 94±6
5 un FuionlsinBl (I6 H) 32±8 91±5
Fig. 1. Measurement of PKC and PKA activity in CV-1 cells following treatment withfumonisin. A, structure of fumonisin and sphingosine. B. measurement of protein kinaseactivity. CV-1 cells were treated with the indicated concentration of fumonisin (JIM) for
3 or 16 h. As a control, cells were treated with CMF PBS. Total protein was extracted fromhomogenized cells, and equivalent amounts of protein (50 ¿ig)were used in a standardPKC or PKA assay as described in "Materials and Methods." Results are an average of
three independent experiments.
1656
Research. on October 20, 2014. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from
FUMONISIN REPRESSION OF PKC AND CAMP STIMULATION
directed against PKC Bl, B2, and y did not detect proteins which hadlower steady-state levels after fumonisin treatment (data not shown).
In summary, these studies identified specific isoforms of PKC, a and6, that were repressed by fumonisin.
Analysis of Transcription after Fumonisin Treatment. To address the effect fumonisin has on transcription, CV-1 cells weretransfected with promoter constructs containing 5 AP-1 binding sites
(5x col/cat TRE) or a single CRE (CRE/cat), and promoter activitywas measured after fumonisin treatment. Promoter activity of 5xcol/cat TRE was reduced more than 10-fold when CV-1 cells were
treated with 5 or 10 /AMfumonisin for 3 h (Fig. 4ß).However,promoter activity of 5x col/cat TRE was reduced only 2-fold after 16h treatment (Fig. 4/4). One copy of an AP-1 binding site linked to the
thymidine kinase promoter was also repressed by fumonisin (data notshown). Previous studies by us (33, 34, 37) and others (41, 42)demonstrated that a phorbol ester, 12-O-tetradecanoylphorbol-13-ac-
etate, stimulates promoter activity of 5x col/cat TRE, lending supportto the hypothesis that fumonisin antagonizes PKC activity. In contrastto 5x col/cat TRE, promoter activity of CRE/cat was stimulatedapproximately 10-fold after treatment with 1 or 5 /AMfumonisin for 3
h (Fig. 4ß).Treatment for 16 h eliminated the stimulatory effect offumonisin. Promoter activity of a construct lacking the 5 AP-1 binding
sites or the CRE (pBLcat/4) was not affected by fumonisin treatment.These results indicated fumonisin had differential effects on a CRE orAP-1 binding site linked to a thymidine kinase promoter.
Discussion
In this study, we examined whether there was a link betweenfumonisin B,, PKC, and downstream targets of PKC in an establishedcell line, CV-1. Since the structure of fumonisin B, is similar to thewell-known PKC inhibitor, sphingosine, we hypothesized that PKC isa target of fumonisin B^ The effect fumonisin B[ has on AP-1-
123456
123456
-116 K
-97 K
-66 K
Fig. 2. Western blot analysis of CV-1 extracts with PKC antibodies following treatment with fumonisin. For all lanes, 50 /ig protein was electrophoresed in a standard 8%SDS polyacrylamide gel, and Western blot analysis was performed using a mixture ofPKC antibodies (GIBCO-BRL) directed against the a isoforms, B isoforms, and -y¡soforms.Lane I. 16-h treatment with 10 jxl CMF PBS (16 h control). Lane 2. 3-htreatment with 10 /il CMF PBS (3-h control). Lane 3, 16-h treatment with l /IM fumonisinB|. Lane 4, 3-h treatment with l /IM fumonisin B,. Lane 5, 16-h treatment with 5 /ÌMfumonisin B]. Lane 6, 3-h treatment with 5 /ÌMfumonisin B,.
116KD97 KD
B123456
— 63 KD
Fig. 3. Identification of specific isoforms of PKC which are repressed by fumonisin.Equal amounts of total proteins (50 /ig) were electrophoresed in an 8% SDS polyacryl-amide gel, and Western blot analysis was performed using isoform-specific PKC anti
bodies (Santa Cruz Biotechnology, Santa Cruz, CA). A, Western blot analysis using PKCa antibody. Lane I, 16-h treatment with 10 /il CMF PBS (16-h control). Lane 2. 16-htreatment with l UM fumonisin B, Lane 3, 16-h treatment with 5 /ÌMfumonisin B, Lane4, 3-h treatment with 5 /AMfumonisin Bt. Lane 5, 3-h treatment with l /IM fumonisin BtLane 6, 3-h treatment with 10 u.1CMF PBS (3-h control). B. Western blot analysis usingPKC 6 antibody. Lane 1, 3-h treatment with 5 ¡IMfumonisin B,. Lane 2, 3-h treatmentwith 1 /ÌMfumonisin B, Lane 3, 3-h treatment with 10 /il CMF PBS (3-h conlrol). Lane4, 16-h treatment with 10 /il CMF PBS (16-h control). Lane 5. 16-h treatment with 1 /ÌMfumonisin Bt Lane 6. 16-h treatment with 5 /IM fumonisin Bj
dependent transcription was also examined because AP-1-dependent
transcription is regulated by PKC (24, 28). It was evident from thesestudies that fumonisin B, decreased steady-state levels of specificPKC isoforms and transiently repressed AP-1-dependent transcription. In contrast, CRE-dependent transcription was stimulated byfumonisin Bu but stimulation appeared to occur in a PKA-independ-
ent fashion. It is hypothesized that the ability of fumonisin to act as acarcinogen or toxin is linked to its ability to repress specific PKCisoforms and consequently alter signal transduction pathways.
Although fumonisin did not eliminate PKC activity, repression wasreproducible and occurred in a dose-dependent fashion. The concentrations required to observe PKC repression was 1-5 /AMor 0.72-3.6
mg/liter. If rats are fed a diet that contains 50 mg/Kg fumonisin B,primary hepatocellular carcinoma occurs in two of three of the animals (6-9). Fumonisin B, disrupts sphingolipid metabolism and stimulates DNA synthesis at concentrations ranging between 20-50 /AM
(43). The concentrations of fumonisin necessary to observe PKCrepression were low relative to other in vivo and in vitro studies andthus appear to be physiologically relevant. It was not surprising thatsignificant levels of PKC activity were detected after fumonisintreatment since multiple isoforms of PKC were apparently expressedin CV-1 cells but not all isoforms were repressed by fumonisin (Fig.2). After CV-1 cells were treated with fumonisin for 3 or 16 h, a M,
120,000 band was repressed, which appears to be PKC a. Althoughmost PKC isoforms have a molecular weight less than MT 100,000, arecent study identified a novel PKC isoform, PKCu, which has amolecular weight of 115,000 (44). Repression of two PKC isoforms,72,000 and 83,000 proteins, was detected after 3 h of fumonisintreatment but not 16 h (Fig. 2), suggesting that repression occurs in atransient manner. However, isotype-specific antibodies failed to rec
ognize these proteins. Western blots probed with all of the availableisotype antibodies detected a band migrating with an apparent molecular weight of 65,000, which was not affected by fumonisin treatment(Fig. 2). However, a PKC 6-specific antibody detected a Mr 63,000protein which had lower steady-state levels after fumonisin treatment
1657
Research. on October 20, 2014. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from

FUMONISIN REPRESSION OF PKC AND CAMP STIMULATION
A. B.pBLcat/4 Sx col/cat TRE CRE/cat pBLcat/4 Sx col /cat TRE CRE/cat
Fumonisin (uM) 0
Ac-CM(%)
C.
pBLcat4
Hlndlll/ Bamm
Hlndlll BamHI
I5XTHE!5X col/cat TRE ^lIpFiPl HSV-l TK I CAT _,
T6AOTCA~TRE~
CRE/cat
Fig. 4. The effect of fumonisin treatment on transcription in transient transfection assays. A CAT construct containing the herpes simplex virus 1 (HSV-1) thymidine kinase promoter(pBLcat4), pBLcat4 containing 5 AP-1 binding sites (5x col/cat TRE), or pBLcat4 containing a single consensus CRE (CRE/cat) was transfected into CV-1 cells (10 (ig DNA/60 mmdish; Ref. 60). A, some cultures were treated with fumonisin for 16 h beginning at 24 h posttransfection. B, some cultures were treated with fumonisin for 3 h beginning at 37 hposttransfection. Fumonisin was dissolved in CMF PBS to give a final concentration of 5 rrtM.After diluting the stock solution in CMF PBS, 10 fj.1was added to 60-mm dishes to obtaina final concentration of 1, 5, or 10 (IM. Those samples with no fumonisin (0) were given 10 fil CMF PBS. At 40 h posttransfection, CAT enzymatic activity was measured as describedpreviously (35—37).The amount of acetylated chloramphenicol (ac-CM) was measured with a Phosphorimager (Molecular Dynamics). The values given are representative of six independentexperiments. C, diagram of plasmids used in this study. The consensus TRE was from Angel et al. (42), and the consensus CRE was from Lee and Mason (29) and Roesler et ai (31).
(Fig. 3ß).It ¡sknown that multiple PKC isoforms migrate with anapparent molecular weight between 60,000 and 70,000 (reviewed inRef. 24). Thus, it is likely that more than one isoform, with molecularweights between 60,000 and 65,000, was detected when Western blotswere probed with a mixture of PKC antibodies.
There is mounting evidence that specific isoforms of PKC playdistinct roles in regulating cellular proliferation or differentiation(reviewed in Refs. 24 and 45). In K562 erythroleukemia cells, expression of PKC B2 correlates with proliferative capacity (26). Incontrast, expression of PKC a correlates with induction ofmegakaryocytic differentiation and cessation of growth. PKC is alsointimately involved in the normal processes and survival of neurons(reviewed in Ref. 45). Expression of c-jun and c-fos is stimulated after
cells are treated with growth stimulating factors: serum, epidermalgrowth factor, transforming growth factor a, or platelet-derivedgrowth factor (41, 46-48). Nerve growth factor also induces c-jun andc-fos in PC12 cells, but this treatment arrests growth and induces neuro
nal differentiation (49). It is likely that, in some tissue, induction of PKCand the AP-1 complex leads to differentiation, whereas in other cell types,
it stimulates proliferation. Since fumonisin represses PKC activity andspecific isoforms of PKC (Figs. 2-4), it is tempting to speculate that
repression of PKC isoforms is necessary for its carcinogenic potential.This hypothesis is supported by the fact that fumonisin B, is not a potentcarcinogen in a standard Ames test (8) but can induce proliferation ofSwiss 3T3 cells in conjunction with insulin (43).
AP-1-dependent transcription is regulated by numerous diverse
mechanisms involving the jun and fos family of transcription factors(reviewed in Refs. 24 and 28). Stimulation of PKC results in thedephosphorylation of specific residues proximal to the DNA binding
domain of c-jun. Specific PKC isoforms (a, B,, y, but not e) phos-phorylate glycogen synthase kinase-3B, thus preventing glycogensynthase kinase-3B from phosphorylating the DNA binding domain ofc-jun (50). Conversely, phosphorylation of specific amino acids at theNH2 terminus of c-jun by a distant relative of MAP kinase, JNK1,activates c-jun (51, 52). At least two proteins which bind c-jun repressc-jun DNA binding and activation of AP-1 dependent transcription:IP-1 (53) and Jif-1 (54). In addition to fos, it is clear that CREB,specific members of the ATF family, and NFKB bind to c-jun andregulate AP-1 transcription (reviewed in Refs. 29-32, 55, 56). Two
other members of the jun family, JunB and JunD, can also dimerizewith c-jun and alter AP-1-dependent transcription (57, 58). The ability
of fumonisin to inhibit PKC activity would, therefore, have a negativeeffect on AP-1-dependent transcription. However, repression of API-dependent transcription might not be limited to PKC repression.
The observation that fumonisin activated a CRE linked to a thymidine kinase promoter in transient transfection assays but had no effecton PKA activity (Figs. 1 and 4) suggested that transcriptional activation was independent of PKA. Although CREB is the major activatorof CRE-dependent transcription and is primarily activated by PKA,other transcription factors can positively regulate CRE-dependenttranscription (reviewed in Refs. 29-32). For example, members of the
ATF family can bind and activate a CRE. CREB can also dimerizewith members of the ATF family and jun family members. Since amember of the Jun family (JunB) activates a CRE but repressestranscription of genes which carry AP-1 binding sites (57, 58), it is
possible that fumonisin activates junB. The finding that calcium/calmodulin-dependent protein kinases have the potential to phospho-
rylate and activate CREB (59) also suggested that fumonisin may
1658
Research. on October 20, 2014. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from

FUMONISIN REPRESSION OF PKC AND CAMP STIMULATION
activate other protein kinases which positively regulate CRE-depend-
ent transcription. Experiments designed to determine how fumonisinactivates a CRE but represses AP-1 are currently in progress.
Acknowledgments
We thank Mike Nelson for intellectual stimulation.
References
1. Dickman, M. B., and Partridge. J. E. Use of molecular markers for monitoring fungiinvolved in stalk rot of corn. Theor. Appi. Genet., 77: 535-539, 1989.
2. Dickman, M. B., and Leslie, J. F. Complementation of nnu mutant of Gibberellafujikaroi (Fusarium moniUforme) by the regulatory nil-2 gene of Neurospora crossa.Mol. Gen. Genet., 2^5: 458-462, 1992.
3. Leslie, J. F., and Dickman, M. B. Fate of DNA encoding hygromycin resistance aftermeiosis in transformed strains of Fusarium moniliforme. Appi. Environ. Microbio!..57: 1423-1429. 1991.
4. Van, K. Y., Dickman, M. B., Xu, J. R., and Leslie, J. F. Sensitivity of field strains ofFusarium moniliforme to benomyl and hygromycin B. Mycologia, 85: 206-213,1993.
5. Thiel, P. G., Marasas, W. F. O., Sydenham. E. W., Shepard, G., and Gelderbloom.W. C. A. The implications of naturally occurring levels of fumonisins in corn andanimal health. Mycopathologia, 117: 3-9, 1992.
6. Gelderbloom, W. C. A., Semple, E., Marasas, W. F. 0., and Farber, E. The cancerinitiating potential of the fumonisin-B mycotoxins. Carcinogenesis (Lond.), 13:433-437, 1992.
7. Gelderbloom, W. C. A., Jaskiewicz, K., Marasas, W. F. O., Thiel, P. G., Horak, R. M.,Vleggaar. R., and Kriek, N. P. J. Fumonisins: novel mycotoxins with cancer promoting activity produced by Fusarium moniliforme. Appi. Environ. Microbio!., 54:1806-1811, 1988.
8. Gelderbloom, W. C. A., Kriek, N. P. J.. Marasas, W. F. 0., and Theil, P. G. Toxicityand carcinogenicity of the Fiisarium moniliforme metabolite, fumonisin Bl. in rats.Carcinogenesis (Lond.), 12: 1247-1251. 1991.
9. Jaskiewicz, K., van Rensburg, S. J., Marasas, W. F. O., and Gelderbloom. W. C. A.Carcinogenicity of Fusarium moniliforme culture media in rats. J. Nati. Cancer Inst.,78: 321-325, 1987.
10. Marasas. W. F. O. Mycotoxicological investigations on corn produced in esophagealcancer areas in Transkei. In: C. J. Pfeiffer (ed.). Cancer of the Esophagus. Vol. 1, pp.29-40. Boca Raton, FL: CRC Press, Inc., 1982.
11. Marasas, W. F. O., Kellerman, T. S., Gelderbloom, W. C. A., Coetzer, J. A. W., Theil,P. G., and van der Lugt, J. J. Leukoencephalomalacia in a horse induced by fumonisinBj isolated from Fusarium moniliforme. Onderstepoort J. Vet. Res., 55: 197-203,1988.
12. Harrison, L. R., Colvin, B., Greene, J. T.. Newman, L. E., and Cole, J. R. Pulmonaryedema and hydrothorax in swine produced by fumonisin Bh a toxic metabolite ofFusarium moniliforme. J. Vet. Diag. Invest., 2: 217-221, 1990.
13. Nelson, P. E., Desjardins, A. E.. and Plattner. R. D. Fumonisins. mycotoxins producedby Fusarium species: biology, chemistry, and significance. Annu. Rev. Phytopathol.,31: 233-252, 1993.
14. Sydenman, E. W., Shepard, G. S., Thiel, P. G., Marasas. W. F. O., and Stockenstrom,S. Fumonisin contamination of commercial corn-based human foodstuffs. J. Agrie.Food Chem., 39: 2014-2018, 1991.
15. Merrill, A. H. M.. Wang, E., Gilchrist, D. G., and Riley. R. T. Fumonisins and otherinhibitors of de novo sphingolipid biosynthesis. Adv. Lipid Res., 26: 215-234, 1993.
16. Norred, W. P., Wang, E., Yoo, H., Riley, R. T.. and Merrill. A. H., Jr. In vitro toxicologyof fumonisins and the mechanistic implications. Mycopathologia, 117: 73-78. 1992.
17. Wang, E., Norred, W. P., Bacon, C. W., Riley, R. T., and Merrill, A. H., Jr. Inhibitionof sphingolipid biosynthesis by fumonisins. J. Biol. Chcm., 266: 14486-14490. 1991.
18. Bell, R. M.. and Burns, J. E. Lipid activation of protein kinase C. J. Biol. Chem., 266:4661-4664, 1989.
19. Hannun, Y. A., Loomis, C. R., Merrill, A. H., and Bell, R. M. Sphingosine inhibitionof protein kinase C activity and phorbol dibutyrate binding in vitro and in humanplatelets. J. Biol. Chem., 267: 12604-12609, 1986.
20. Mathias, S., Dressler, K. A., and Kolesnick. R. N. Characterization of a ceramide-activated protein kinase: stimulation by tumor necrosis factor a. Proc. Nati. Acad. Sci.USA, fifi: 10009-10013, 1991.
21. Dobrowsky, R. T., and Hannun. Y. A. Ceramide stimulates a cytosolic proteinphosphatase. J. Biol. Chem., 267: 5048-5051, 1992.
22. Nizhizuka, Y. Intracellular signaling by hydrolysis of phospholipids and activation byprotein kinase C. Science (Washington DC), 25Ä:607-614, 1992.
23. Castagna, M., Takai, Y., Kaibuchi, K., Sano, K.. Kikkawa. U.. Nishizuka, Y. Directactivation of calcium-activated, phospholipid-dependent protein kinase by tumor-promoting phorbol esters. J. Biol. Chem., 257: 7847-7851, 1982.
24. Dekker, L. V., and Parker, P. J. Protein kinase C: a question of specificity. TrendsBiol. Sci., 19: 73-77, 1994.
25. Hato, A., Akita, Y., Suzuki, K., and Ohno, S. Functional divergence of protein kinaseC (PKC) family members. J. Biol. Chem., 268: 9122-9129, 1993.
26. Murray, N. R.. Baumgardner. G. P., Burns, D. J., and Fields, A. P. Protein kinase Cisotypes in human erythroleukemia (K562) cell proliferation and differentiation.J. Biol. Chem., 26«:15847-15853, 1993.
27. Gallo, A., Benusiglio, E., Bonopace, I. M., Feliciello, A., Cassano, S., Garbi, C.,Musti, A. M., Gottesman, M. E., and Avedimento. E. V. v-ras and protein kinase Cdedifferentiate thyroid cells by down regulating nuclear cAMP-dependent proteinkinase A. Genes Dev., 6: 1621-1630, 1992.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
Ransone. L. J.. and Vcrma. I. M. Nuclear proto-oncogenes Fos and Jun. Annu. Rev.Cell Biol., 6: 539-557, 1990.
Lee, K. A. W., and Masson. N. Transcriptional regulation by CREB and its relatives.Biochem. Biophys. Acta. II74: 221-233, 1993.Meyer, T. E.. and Habener. J. F. Cyclic adenosine 3',5'-monophosphate response
element binding protein (CREB) and related transcription-activating deoxyribonu-cleic acid binding proteins. Endocr. Rev., 14: 269-290, 1993.
Roesler, W. J.. Vanderbark. G. R., and Hanson. R. W. Cyclic AMP and the inductionof eucaryotic gene transcription. J. Biol. Chem., 26: 9063-9(166, 1988.Taylor, S. S., Buechler, J. A., and Yonemoto. W. cAMP-dependent protein kinase:framework for a diverse family of regulatory enzymes. Annu. Rev. Biochem., 59:971-1005, 1990.Garret, L. R., Jones, C.. and Cuchens. M. A. Pristane-induced gene activation. Chem.Biol. Interact., fi: 119-130, 1992.Garret. L. R., Ackland-Berglund, C. E., Jones, C. J., and Cuchens. M. A. Differential
effects of TPA and pristane on gene expression and transformation in mouse epidermal cells. Exp. Cell Res., 205: 416-421, 1993.Jones. C. The minimal transforming fragment of HSV-2 can function as a complextranscriptional promoter. Virology, 169: 346-353. 1989.Jones C., Delhon. G., Bratanich. A., and Rock, D. Analysis of the transcriptionalpromoter which regulates the latency-related transcript of bovine herpesvirus 1.J. Virol.. 64: 1164-1170, 1990.Jones, C., Zhu, F.. and Dhanwada, K. An intergenic element of HSV-2 gene encodingthe large subunit of ribonucleotide reducÃase can function as a distal activatingelement in human keratinocytcs. DNA Cell Biol., 12: 127-137, 1993.Sambrook, J.. Fritsch, E. F., and Maniatis, T. Molecular Cloning: A LaboratoryManual. Cold Spring Harbor. NY: Cold Spring Harbor Laboratory. 1989.Banbura. M., Ackland-Berglund, C.. Lee, S-H.. Hamernik. D., and Jones. C. Analysisof transcriptional activation of a cyclic AMP response element by 2,6,10.14-tctram-cthylpcntadecane (pristane) in JB6 mouse epidermal cells. Mol. Carcinog., 11:204-214. 1994.Bradford. M. M. A rapid and sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein-dye binding. Anal. Biochem..72: 248-264, 1976.Angel, P.. Hattori, K., Smeal, T., and Karin, M. The c-jun protooncogene is positivelyregulated by its product. Jun/AP-1. Cell, 55: 875-885, 1988.
Angel, P.. Imagawa. M.. Chiù.R.. Stein. B., Imbra. R. J., Rahmsdorf. H. J., Jonal, C.,Herrlich, P.. and Karin. M. Phorbol ester-inducible genes contain a common cis elementrecognized by a TPA modulated rnin.v-acting factor. Cell. 49: 729-739, 1987.Schroeder. J. J.. Crane, H. M., Xia, J.. Liotta, D. C., and Merrill, A. H., Jr. Disruptionof sphingolipid metabolism and stimulation of DNA synthesis of fumonisin B,.J. Biol. Chcm., 269: 3475-3481. 1994.Johannes. F-J., Presile, J., Eis, S., Oberhagemann, P.. and Pfi/enmaier, K. PKCu
is a novel, atypical member of the protein kinase C family. J. Biol. Chem., 269:6140-6148, 1994.
Tanaka, C., and Nishizuka, Y. The protein kinase C family for neural signaling. Annu.Rev. Neurosci., 17: 551-567. 1994.Brenner, D. A., O'Hara, M., Angel, P., Chojker, M., and Karin, M. Prolonged
activation of jun and collagenase genes by tumor necrosis factor a. Nature (Lond.),3.17: 661-663, 1989.
Greenberg, M. E.. and Ziff. E. B. Stimulation of 3T3 cells induces transcription of thec-fos proto-oncogene. Nature (Lond.), 311: 433-437. 1984.
Ryder. K.. and Nathans. D. A gene activated by growth factors is related to theoncogene v-jun. Proc. Nati. Acad. Sci. USA, 85: 8464-8467, 1988.
Darteli, D. P.. Shcng, M., Lau. L. F., and Greenberg, M. E. Growth factors andmembrane depolarization activate distinct programs of early response gene expression: dissociation of fos and jun induction. Genes Dev., 3: 304-313, 1989.Goode, N.. Hughes, K., Woodgett, J. R., and Parker, P. J. Differential regulation ofglycogen synthase kinase-3B by protein kinase C isotypes. J. Biol. Chem., 267:16878-16882, 1992.Hibi. M.. Lin. A.. Smcal, T.. Minden. A., and Karin, M. Identification of anoncoprotein- and UV-responsivc protein kinase that binds and potentiates Ihe c-Junactivation domain. Genes Dev., 7: 22135-22148, 1993.Derijard. B., Hibi, M.. Wu, I-H., Barrett, T., Su, B., Deng, T., Karin, M., Davis, R. J.JNKl: a protein kinase stimulated by UV light and Ha-Ras that binds and phospho-rylaled the c-Jun activation domain. Cell, 76: 1025-1037, 1994.Auwerx, J.. and Sassone-Corsi. P. IP-1: a dominant inhibitor of fos/jun whose activityis modulated by phosphorylation. Cell, 64: 983-993, 1992.Monteclaro, F. S.. and Vogt, P. K. A Jun-binding protein related to a putative tumorsuppressor. Proc. Nail. Acad. Sci. USA, 90: 6726-6730. 1993.Hagmeyer, B. M.. Konig, H., Herr, !.. Offringa, R.. Zantema. A., van der Eh, A. J.,Herrlich. P.. and Angel. P. Adenovirus negatively and positively modulates transcription of AP-1 dependent genes by dimer-specific regulation of the DNA binding andtransactivation activities of Jun. EMBO J., 12: 3559-3572, 1993.
Stein, B.. Baldwin, A. S., Baldwin, A. S., Jr.. Ballard, D. W., Greene, W. C, Angel.P.. and Herrlich. P. Cross coupling of the NF-KB p65 and Fos/Jun transcription factorsproduces potentiated biological function. EMBO J.. 13: 3879-3890, 1994.Chiù.R.. Angel. P.. and Karin. M. Jun-B differs in its biological properties from, andis a negative regulator of, c-Jun. Cell, 59: 979-986, 1989.Hsu, J-C., Crcssman, D. E.. and Traub, R. Promoter-specific frtw.y-activation andinhibition mediated by JunB. Cancer Res., 53: 3789-3794, 1993.Dash, P. K., Karl, K. A., Cólicos, M. A., Prywes, R., and Kandel, E. R. cAMPresponse element-binding protein is activated by Ca1 '/calmodulin as well as cAMP-
dependent protein kinase. Proc. Nati. Acad. Sci. USA. fifi: 506I-5065, 1991.Graham. F.. and van der Eh, A. A new technique for the assay of infeclivity of humanadenovirus 5 DNA. Virology. 52: 456-457, 1973.
1659
Research. on October 20, 2014. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from




















