
Repeated exposure to stress across the childhood-adolescent period alters rats' anxiety- and...
13
Repeated Exposure to Stress Across the Childhood–Adolescent Period Alters Rats’ Anxiety- and Depression-Like Behaviors in Adulthood: The Importance of Stressor Type and Gender Joanna Pohl, Mary C. Olmstead, Katherine E. Wynne-Edwards, Kate Harkness, and Janet L. Menard Queen’s University This research tests the hypothesis that specific forms of adversity in early life map onto behavioral signs analogous to depression versus anxiety in later life. Male and female rats were exposed to either severe sporadic stress or chronic mild stress during the childhood–adolescent period, and their behavior was tested in adulthood. Males in the severe sporadic stress group showed exaggerated anxiety-related behaviors, as indicated by increases in shock-probe burying and escape-like responses (jumps) from the open arms of the elevated plus-maze. Females exposed to severe sporadic stress displayed no change in burying behavior but did display increases in escape behavior. These same females also exhibited behaviors analogous to depression that manifested as decreased sucrose consumption. The chronic mild stress regime produced effects only in females, including reduced burying, decreased sucrose consump- tion, and an exaggerated corticosterone response to cold-water immersion stress. Findings reiterate the importance of early life experience to the development of adult psychopathologies and emphasize the need to consider both the type of early experience and gender differences in these analyses. Keywords: early life adversity, animal models of anxiety, animal models of depression, anhedonia, stress Early life experiences can have a profound impact on the health and psychological functioning of humans (McEwen, 2003). Most notably, childhood abuse and neglect are associated with the development of anxiety disorders and major depression in adult- hood (Heim & Nemeroff, 2001; Luecken & Lemery, 2004; Weiss, Longhurst, & Mazure, 1999). Depression and the anxiety disorders share many common symptoms, including irritability, sleep and appetite disruption, and low energy, as well as disruptions in social and cognitive functioning. However, depression and anxiety are distinguished by two core symptoms. In particular, the anxiety disorders are distinguished from depression by chronic somatic hyperarousal, which is not necessarily precipitated by explicit threatening stimuli (Clark, Beck, & Stewart, 1990). The main distinguishing feature of depression, in contrast, is anhedonia, or a decreased interest or pleasure in nearly all activities (American Psychiatric Association, 2000). Given the heterogeneity of syndromal expression associated with childhood abuse and neglect, a number of researchers have attempted to find preferential relations between specific types of adversity and specific syndromes. In particular, researchers have predicted that childhood physical (i.e., nonsexual) and sexual abuse, with their clear themes of danger, are preferentially predic- tive of later anxiety disorders, whereas childhood neglect, which involves themes of loss, is preferentially predictive of depression. Physical and sexual abuse in childhood have been preferentially associated with anxiety alone (Gibb, Butler, & Beck, 2003) and with anxiety comorbid with depression (Harkness & Wildes, 2002; Levitan, Rector, Sheldon, & Goering, 2003). In contrast, childhood neglect and other forms of emotional abuse have been preferen- tially related to depression (Hankin, 2005; Harkness & Wildes, 2002). However, other studies have failed to find any evidence for specificity (e.g., Brown & Harris, 1993). Human studies are limited in their ability to examine specificity in the relations between childhood adversity and adult psychopa- thology because different forms of maltreatment often co-occur in the same individual. Furthermore, because most human studies are retrospective and correlational, it is difficult to establish causal relations between specific childhood experiences and adult psy- chopathology. Animal studies can address some of these limita- tions if animal models can be developed that have face, predictive, and theoretical validity (Willner, 1986). Abuse is characterized by danger (i.e., immediate physical threat) and temporal unpredict- ability; modeling this situation in rats would involve exposing animals to sporadic (randomly delivered) stressors that, although relatively brief, are highly noxious. In contrast, childhood neglect is a more pervasive state in which the provision of basic necessi- ties, such as food, bathing, household cleanliness, and supervision, is chaotic and limited (Cowen, 1999). Although not physically threatening in an immediate sense, neglect is nonetheless stressful when taken over the long term (De Bellis, 2005; Hildyard & Wolfe, 2002). This experience could be modeled in animals by Joanna Pohl, Mary C. Olmstead, Kate Harkness, and Janet L. Menard, Department of Psychology, Queen’s University, Kingston, Ontario, Can- ada; Katherine E. Wynne-Edwards, Department of Biology, Queen’s Uni- versity, Kingston, Ontario, Canada. This research was supported by an Advisory Research Council grant from Queen’s University and by Natural Sciences and Engineering Re- search Council grants. Correspondence concerning this article should be addressed to Janet L. Menard, Department of Psychology, Queen’s University, Humphrey Hall, 62 Arch Street, Kingston, Ontario K7L 3N6, Canada. E-mail: [email protected] Behavioral Neuroscience Copyright 2007 by the American Psychological Association 2007, Vol. 121, No. 3, 462– 474 0735-7044/07/$12.00 DOI: 10.1037/0735-7044.121.3.462 462
Transcript of Repeated exposure to stress across the childhood-adolescent period alters rats' anxiety- and...

Repeated Exposure to Stress Across the Childhood–Adolescent PeriodAlters Rats’ Anxiety- and Depression-Like Behaviors in Adulthood: The
Importance of Stressor Type and Gender
Joanna Pohl, Mary C. Olmstead, Katherine E. Wynne-Edwards, Kate Harkness, and Janet L. MenardQueen’s University
This research tests the hypothesis that specific forms of adversity in early life map onto behavioral signsanalogous to depression versus anxiety in later life. Male and female rats were exposed to either severesporadic stress or chronic mild stress during the childhood–adolescent period, and their behavior wastested in adulthood. Males in the severe sporadic stress group showed exaggerated anxiety-relatedbehaviors, as indicated by increases in shock-probe burying and escape-like responses (jumps) from theopen arms of the elevated plus-maze. Females exposed to severe sporadic stress displayed no change inburying behavior but did display increases in escape behavior. These same females also exhibitedbehaviors analogous to depression that manifested as decreased sucrose consumption. The chronic mildstress regime produced effects only in females, including reduced burying, decreased sucrose consump-tion, and an exaggerated corticosterone response to cold-water immersion stress. Findings reiterate theimportance of early life experience to the development of adult psychopathologies and emphasize theneed to consider both the type of early experience and gender differences in these analyses.
Keywords: early life adversity, animal models of anxiety, animal models of depression, anhedonia, stress
Early life experiences can have a profound impact on the healthand psychological functioning of humans (McEwen, 2003). Mostnotably, childhood abuse and neglect are associated with thedevelopment of anxiety disorders and major depression in adult-hood (Heim & Nemeroff, 2001; Luecken & Lemery, 2004; Weiss,Longhurst, & Mazure, 1999). Depression and the anxiety disordersshare many common symptoms, including irritability, sleep andappetite disruption, and low energy, as well as disruptions in socialand cognitive functioning. However, depression and anxiety aredistinguished by two core symptoms. In particular, the anxietydisorders are distinguished from depression by chronic somatichyperarousal, which is not necessarily precipitated by explicitthreatening stimuli (Clark, Beck, & Stewart, 1990). The maindistinguishing feature of depression, in contrast, is anhedonia, or adecreased interest or pleasure in nearly all activities (AmericanPsychiatric Association, 2000).
Given the heterogeneity of syndromal expression associatedwith childhood abuse and neglect, a number of researchers haveattempted to find preferential relations between specific types ofadversity and specific syndromes. In particular, researchers have
predicted that childhood physical (i.e., nonsexual) and sexualabuse, with their clear themes of danger, are preferentially predic-tive of later anxiety disorders, whereas childhood neglect, whichinvolves themes of loss, is preferentially predictive of depression.Physical and sexual abuse in childhood have been preferentiallyassociated with anxiety alone (Gibb, Butler, & Beck, 2003) andwith anxiety comorbid with depression (Harkness & Wildes, 2002;Levitan, Rector, Sheldon, & Goering, 2003). In contrast, childhoodneglect and other forms of emotional abuse have been preferen-tially related to depression (Hankin, 2005; Harkness & Wildes,2002). However, other studies have failed to find any evidence forspecificity (e.g., Brown & Harris, 1993).
Human studies are limited in their ability to examine specificityin the relations between childhood adversity and adult psychopa-thology because different forms of maltreatment often co-occur inthe same individual. Furthermore, because most human studies areretrospective and correlational, it is difficult to establish causalrelations between specific childhood experiences and adult psy-chopathology. Animal studies can address some of these limita-tions if animal models can be developed that have face, predictive,and theoretical validity (Willner, 1986). Abuse is characterized bydanger (i.e., immediate physical threat) and temporal unpredict-ability; modeling this situation in rats would involve exposinganimals to sporadic (randomly delivered) stressors that, althoughrelatively brief, are highly noxious. In contrast, childhood neglectis a more pervasive state in which the provision of basic necessi-ties, such as food, bathing, household cleanliness, and supervision,is chaotic and limited (Cowen, 1999). Although not physicallythreatening in an immediate sense, neglect is nonetheless stressfulwhen taken over the long term (De Bellis, 2005; Hildyard &Wolfe, 2002). This experience could be modeled in animals by
Joanna Pohl, Mary C. Olmstead, Kate Harkness, and Janet L. Menard,Department of Psychology, Queen’s University, Kingston, Ontario, Can-ada; Katherine E. Wynne-Edwards, Department of Biology, Queen’s Uni-versity, Kingston, Ontario, Canada.
This research was supported by an Advisory Research Council grantfrom Queen’s University and by Natural Sciences and Engineering Re-search Council grants.
Correspondence concerning this article should be addressed to Janet L.Menard, Department of Psychology, Queen’s University, Humphrey Hall,62 Arch Street, Kingston, Ontario K7L 3N6, Canada. E-mail:[email protected]
Behavioral Neuroscience Copyright 2007 by the American Psychological Association2007, Vol. 121, No. 3, 462–474 0735-7044/07/$12.00 DOI: 10.1037/0735-7044.121.3.462
462

exposing them to a series of mildly aversive stimuli on a more orless continuous basis.
The present study tested the hypothesis that specific early ad-versity modeled on abuse versus neglect predicts specific behav-ioral signs analogous to anxiety versus depression in adult rats.Abuse was modeled by presenting brief but severe sporadic stres-sors (restraint–water immersion, inescapable foot shock), whereasneglect was modeled by exposing a different group of rats to aseries of chronic mild stressors (e.g., wet bedding, cage tilt, fooddeprivation, and so forth). The stress paradigms were presentedfrom weaning on Postnatal Day (PD) 23 through to PD 51. Byconservative estimates, adolescence in the rat occurs from approx-imately PD 28–PD 42 (Spear, 2000), although some developmen-tal processes specific to adolescence persist through PD 55 (Ojeda& Urbanski, 1994; Spear, 2000; Spear & Brake, 1983). Stressors inour study were applied from preadolescence (childhood) throughto late adolescence, as broadly defined. We chose this develop-mental range because in humans, childhood abuse and neglecthave their strongest effect on adult psychopathology when theypersist from early and late childhood through adolescence (Thorn-berry, Ireland, & Smith, 2001). As adults, anxiety was assessedusing the elevated plus-maze and the shock-probe defensive bury-ing task, two well-validated animal models of anxiety (Pellow,Chopin, File, & Briley, 1985; Treit, 1990). Anhedonia was mod-eled by testing for sucrose consumption (Papp, Willner, & Muscat,1991). We also used the social transmission of food preferencetask (Galef & Wigmore, 1983) to assess possible memory deficits,as this is a feature of both anxiety and depression in humans(Airaksinen, Larsson, & Forsell, 2005; Airaksinen, Larsson, Lund-berg, & Forsell, 2004; Landro, Stiles, & Sletvold, 2001). Finally,we assessed hypothalamic–pituitary–adrenal axis activity by mea-suring glucocorticoid responses to a stressful stimulus (cold-waterimmersion). Both male and female animals were tested, allowingfor an examination of gender differences in the effects of early lifeadversity on anxiety and depression. On the basis of clinicalfindings, we hypothesized that exposure to severe sporadic stressacross rearing (i.e., from weaning through late adolescence) wouldlead to anxiety-like signs, whereas chronic mild stress during thatperiod would induce depressive-like signs. We also predicted thatthe effects of both rearing conditions would be more prominent infemales than in males (Piccinelli & Wilkinson, 2000; Weiss et al.,1999).
Method
Subjects
The subjects were Long–Evans rats (45 males and 45 females),obtained from an in-house breeding facility at Queen’s University,Kingston, Ontario, Canada. The rats were weaned on PD 21 andindividually housed in standard polycarbonate cages (59.0 �38.5 � 20.0 cm), containing aspen wood chips for bedding. Thecolony was maintained on a 12-hr light–dark cycle, with lights onat 0700. Food and water were provided ad libitum, except whereotherwise noted in the behavioral procedures. Experimental pro-cedures were conducted in accordance with the Canadian Councilon Animal Care and were approved by the Queen’s UniversityAnimal Care Committee.
Rearing Conditions
Beginning on PD 23, each group was exposed to their respectiverearing condition for 28 days (i.e., until PD 51). The chronic mildstress group was exposed to a series of stressors, as outlined inTable 1. The severe sporadic stress group was exposed to twostressors per week presented at random intervals. One severestressor involved water immersion and restraint stress; that is, ratswere immersed, to the level of the syphoid process, in 24 °C waterfor a period of 45 min while being restrained in a cylindrical tube11 cm in diameter. The second severe stressor consisted of threefoot shocks (0.6 mA for a period of 3 s) delivered randomlythroughout a 5-min session through the grid floor of a chamber(29 � 22 � 20 cm). The control group was handled twice a weekto mimic the physical handling associated with stressor applica-tions in the chronic mild stress and severe sporadic stress groups.Stressors ended at roughly PD 51, and the rats were then leftundisturbed (except for weekly cage cleaning) for a 3-week no-stress recovery period. After this recovery period, at roughly PD72, all rats underwent a series of anxiety and depression screeningtests. The chronology of testing is presented in Table 2.
Elevated Plus-Maze
Elevated plus-maze testing occurred between 1400 and 1700,during the light photoperiod. The elevated plus-maze consisted ofa plus-shaped maze made of urethane-sealed wood with two op-posing arms (open and closed), raised 50 cm above the floor withdimensions of 50 � 10 cm and 50 � 10 � 40 cm, respectively.Testing was performed in a room illuminated by a red light. Priorto testing, rats were handled in the room containing the maze for5 min on 6 consecutive days. In our hands, handling prior to theplus-maze test yields control levels of open-arm activity (30%–35%) that are ideal for detecting potential treatment effects ineither direction (i.e., increases or decreases). On the 7th day, eachrat was individually placed at the center of the maze facing an openarm and allowed to explore the entire maze for 5 min. All testingwas videotaped and later coded offline. An arm entry was definedas all four paws in a given arm. The percentage of entries madeinto the open arms and percentage time spent in the open arms (i.e.,
Table 1Chronic Mild Stress Schedule
Day Stressor
Sunday Stroboscopic light (11 a.m.–4 p.m.)Soiled bedding (5 p.m.–8 a.m.)
Monday 40° cage tilt (11 a.m.–5 p.m.)Overnight illumination (5 p.m.–8 a.m.)
Tuesday White noise (1 p.m.–3 p.m.)Wednesday Group housing (12 p.m.–2 p.m.)Thursday Stroboscopic light (1 p.m.–5 p.m.)
Overnight illumination (5 p.m.–9 a.m.)Water deprivation (removal of water bottle;
7 p.m.–11 a.m.)Friday Empty water bottle provided (11 a.m.–12 p.m.)
Group housing (1 p.m.–5 p.m.)
Note. The sequence and time at which each of the stressors started variedacross weeks, but the duration of each stressor remained constant through-out the rearing period.
463CHILDHOOD–ADOLESCENT STRESS EFFECTS IN ADULT RATS

100 � open / [open � closed]) were used as indices of anxiety.The number of entries rats made into the closed arms of the mazeand the total number of entries made into any arm were measuredas indices of general locomotor activity. Because highly reactiverats are known to jump off the maze (Kalynchuk, Pinel, Treit, &Kippin, 1997), we also recorded jumps (i.e., the rat leans and peersover the edge of an open arm and then proceeds head first off themaze) and falls (i.e., the rat rears, turns, or is startled on an openarm, loses its balance, and leaves the open arm either sideways orrump first). If a rat jumped or fell off one of the open arms, it wasquickly picked up and returned to the center of the maze. The mazewas thoroughly cleaned with water after each rat was tested.
Shock-Probe Defensive Burying
Shock-probe testing occurred between 1600 and 1900. Theshock-probe burying test consisted of a Plexiglas test chamber(40 � 30 � 40 cm) with approximately 5 cm of bedding (cat litter)spread evenly on the floor of the test chamber. An electrifiedwire-wrapped Plexiglas probe (6.0 � 0.5 � 0.5 cm) was insertedthrough a small hole (centered 2 cm above the bedding material).The electrical current was distributed through two copper wireswrapped around the probe. Using a 2,000-V shock source, theintensity of the shock was set at 2 mA. During the habituationtrials, the rats were brought into the testing room and individuallyplaced in the test chamber (without the shock-probe present) for 15min on each of 4 consecutive days. Twenty-four hours after the lasthabituation trial, the shock-probe was secured 6 cm into thechamber and the rats were individually placed into the test cham-ber. The rat received a brief 2-mA shock whenever its snout or pawtouched the electrified probe. The 15-min test duration beganimmediately after the rat’s first shock. After each test, feces wereremoved and bedding material was smoothed out to the original 5cm thickness.
The behavior of each rat was videotaped using a digital videosystem for later analysis of the following behaviors, as definedpreviously (Treit, 1990): (a) total duration of time spent burying(i.e., pushing bedding material toward the probe using rapid,forward thrusts of the forelimb), (b) total duration of generallocomotor activity (i.e., forward or backward locomotion not as-sociated with burying), (c) number of stretch–attend postures (i.e.,the rat stretches toward the shock-probe in an elongated fashionand then quickly retracts to its original body position), and (d) theduration of probe investigation (e.g., sniffing directed toward theprobe with the snout being within 5 cm of the probe). Stretch–attend postures and probe investigation constitute risk assessmentand are thought to provide a means for rodents to gather informa-
tion about a threatening stimulus (Blanchard & Blanchard, 1989).Finally, we recorded the total number of probe-contact-inducedshocks received, and the rats’ behavioral response to each shockwas scored using a 4-point scale: flinch involving only the forepawor head (1); whole body flinch, without ambulation away from theshock-probe (2); whole body flinch and/or jumping with ambula-tion away from the shock-probe (3); and whole body flinch and/orjumping followed by running to the opposite side of the testchamber (4). A mean shock reactivity score was derived for eachrat by summing the individual shock reactivity scores and dividingthis total by the total number of shocks.
Blood Collection and Corticosterone Radioimmunoassay
Whole blood was collected between 0900 and 1200, with sam-pling times counterbalanced across treatment groups (males andfemales were sampled on separate days). Individual samples tookless than 3 min to collect. This sampling time is appropriate forobtaining baseline measures of corticosterone (e.g., Pecoraro et al.,2006). Rats were gently restrained by hand and whole blood wascollected, without anesthesia, from the saphenous vein into hepa-rinized microhematocrit tubes (2 � 75 �l) and centrifuged at5000 � g for 5 min to separate the red blood cells from the plasma.Plasma was stored in 200-�l Eppendorf tubes at –20 °C untilquantified by radioimmunoassay (ICN [125I] double-antibody forrats and mice; MP Biomedicals, LLC, Orangeburg, NY). Sug-gested methods were followed exactly. All samples were quanti-fied in duplicate (10 �l plasma) over five assay runs, with allplasma samples from any individual in the same assay run. Intra-assay variability (quadruplicate determinations in each assay run)was 7.48% at 81 ng/ml and 6.60% at 670 ng/ml. However, inter-assay variability was unacceptably high (43.6%) at the lowerconcentration (compared with 13.5% at the higher concentration).As a result, statistical comparisons between individuals usedwithin-individual relative concentrations (initial sample � 100%)rather than absolute concentrations.
Following the initial blood collection, rats were subjected to acold-water immersion test for 15 min such that they were im-mersed individually in an opaque plastic tub (37 � 28 � 30 cm)containing water at a temperature of 15 °C and at a depth of 12 cm(males) or 10 cm (females). The depth of water was kept highenough for the rats to be submerged to the syphoid process. Asecond blood sample was collected immediately following thewater immersion. Rats were then returned to their home cage andplaced under a heat lamp for a period of 45 min, after which a thirdblood sample was collected. The initial blood sample measuredbaseline corticosterone levels, whereas the second and third sam-ples served as measures of stress reactivity and recovery, respec-tively.
Social Transmission of Food Preference
Testing occurred between 1000 and 1600. Before testing, all ratswere food deprived for 16 hr and then exposed to unflavoredground chow for 1 hr; all rats consumed the ground chow duringthis period. Testing began 24 hr later by presenting a demonstratorrat (i.e., not one of the rats in the experiment) with ground rat chowflavored with either 2% unsweetened cocoa (Master Choice Pre-mium Cocoa) or 1% cinnamon (Club House ground cinnamon) for
Table 2Testing Schedule
Procedure Age
Weaning 3 weeksGroup treatment 3–7 weeksElevated plus-maze 10–13 weeksShock-probe burying 14–17 weeksCorticosterone stress reactivity 22–23 weeksSocial transmission of food preference 25–28 weeksSucrose consumption 45 weeks
464 POHL ET AL.

1 hr. A pilot experiment was conducted to verify that rats did nothave a natural preference for either one of the flavors used in theexperiment. Demonstrators were required to consume at least 1 gof chow to be included in the experiment. Same-sex demonstratorand experimental rats were then placed in a square open-fieldarena (100 � 100 � 50 cm) constructed of clear Plexiglas con-taining bedding at a depth of 3 cm for 15 min. Immediatelyfollowing the interaction period, the experimental rat was broughtback to the colony room and presented with two food dishes, oneflavored with cocoa and the other flavored with cinnamon, for 1 hr.Social transmission of food preference is manifested as increasedconsumption of the demonstrator-paired flavored chow.
Sucrose Consumption
All tests were performed between 1000 and 1400. The sucroseconsumption test was conducted in the colony room in opaquecages (45 � 23 � 15 cm) containing bedding at a depth of 2 cm.During the habituation trial, rats were presented with a sucrosesolution (10% w/v; 0.29 M) in a standard water bottle for 1 hr.Over the next 2 days, rats were presented with a bottle containingeither 0% or 7% (0.20 M) sucrose for 1 hr (counterbalanced). Theamount of sucrose solution consumed (ml) over the 1-hr exposureperiod was measured for each of the three concentrations (0%, 7%,and 10%).
Data Analysis
Body weight was recorded at ages 22, 28, and 45 weeks, anddata from each sampling time point were analyzed separately using2 � 3 factor analyses of variance (ANOVAs), with sex (female,male) and rearing condition (control, chronic mild stress, severesporadic stress) as between-subjects factors. Data from the ele-vated plus-maze and shock-probe burying tests were analyzedusing two separate 2 � 3 factor ANOVAs, with sex and rearingcondition as between-subjects measures. Differences in the pro-portion of rats that jumped from the open arms of the plus-mazewere analyzed using a chi-square test. Social transmission of foodpreference data were analyzed using a 2 � 2 � 3 factor ANOVA,with chow familiarity (demonstrator paired vs. novel), sex, andrearing condition as between-subjects factors. The sucrose con-sumption data were analyzed using a 2 � 3 ANOVA, with sex andrearing condition as between-subjects factors. Because the 10%sucrose concentration yielded data that violated the assumption ofhomogeneity of variance, a separate analysis was conducted foreach of the three different sucrose concentrations (0%, 7%, and10%), and the data obtained using 10% sucrose was transformed(log10) prior to analysis. Corticosterone levels at the three sam-
pling points (baseline, stress reactivity, and recovery) were con-verted to percentage baseline values, which were used to determinearea under the curve (AUC) with respect to the initial sample.AUCs of the female rats were analyzed using ANOVA, withrearing group as a between-subjects factor. Because of technicalproblems, corticosterone data from the male rats were not availablefor analysis.
For all analyses, significant main effects of rearing conditionwere followed by post hoc analysis using the least significantdifference (LSD) test. Significant interactions were further ana-lyzed using one-way ANOVAs, separate for each sex, andfollow-up pairwise comparisons (LSD test). For all analyses, thealpha level was set at p � .05.
Results
Body Weight Analysis
There was a significant main effect of sex on body weight, suchthat males weighed more than females at each of the three sam-pling points: 22 weeks, F(1, 83) � 533.49, p � .01; 28 weeks, F(1,81) � 670.31, p � .01; and 45 weeks, F(1, 42) � 226.47, p � .01.There were no main effects of rearing or Sex � Rearing interac-tions at any sampling point (all ps � .05). A summary of bodyweight at the three sampling points is shown in Table 3.
Elevated Plus-Maze
Data from the elevated plus-maze test are provided in Table 4.There were no main effects of sex, rearing, or Sex � Rearinginteractions on either the percentage of open-arm entries (all ps �0.5) or the absolute number of open-arm entries (all ps � .05; seeTable 4). However, there was a main effect of rearing condition,F(2, 84) � 5.58, p � .01, on the percentage of time rats spent onthe open arms of the maze (see Figure 1A). Subsequent pairwisecomparisons (LSD test) confirmed that rats exposed to severesporadic stress across the childhood–adolescent period spent agreater percentage of time on the open arms than did rats in theno-stress control ( p � .01) or chronic mild stress ( p � .01)conditions. This increase in open-arm time was not secondary togeneral increases in locomotor activity. In fact, rats in the severesporadic stress group made fewer closed-arm entries relative toboth of the other groups (see data in Table 4), a pattern confirmedsignificant by ANOVA, F(2, 84) � 4.32, p � .05, and subsequentpairwise comparisons (both ps � .01).
Only 2 rats (both controls) lost their balance and fell sidewaysfrom an open arm. Thus, the number of falls was not analyzed.However, as can be seen in Figure 1B, rats in the severe sporadic
Table 3Mean (� SEM) Body Weight (in Grams) of Rats at 22, 28, and 45 Weeks of Age
Age
Females Males
No-stresscontrols
Chronic mildstress
Severesporadic stress
No-stresscontrols
Chronic mildstress
Severesproadic stress
22 weeks 313.33 (6.82) 309.13 (8.96) 322.00 (7.56) 536.73 (16.82) 559.64 (15.36) 521.67 (12.67)28 weeks 328.27 (7.34) 332.20 (8.90) 324.93 (8.48) 605.13 (16.88) 597.43 (15.69) 594.47 (14.81)45 weeks 378.13 (19.13) 402.00 (15.41) 378.88 (13.76) 695.63 (31.48) 682.75 (26.52) 658.00 (30.14)
465CHILDHOOD–ADOLESCENT STRESS EFFECTS IN ADULT RATS
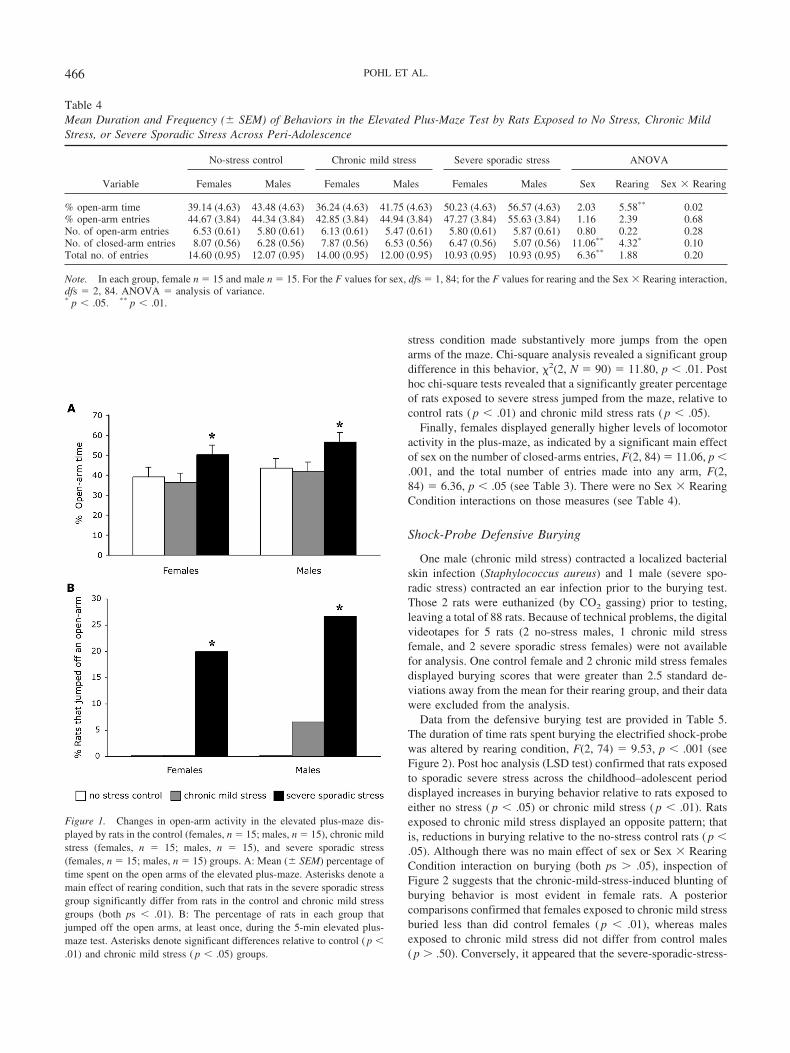
stress condition made substantively more jumps from the openarms of the maze. Chi-square analysis revealed a significant groupdifference in this behavior, �2(2, N � 90) � 11.80, p � .01. Posthoc chi-square tests revealed that a significantly greater percentageof rats exposed to severe stress jumped from the maze, relative tocontrol rats ( p � .01) and chronic mild stress rats ( p � .05).
Finally, females displayed generally higher levels of locomotoractivity in the plus-maze, as indicated by a significant main effectof sex on the number of closed-arms entries, F(2, 84) � 11.06, p �.001, and the total number of entries made into any arm, F(2,84) � 6.36, p � .05 (see Table 3). There were no Sex � RearingCondition interactions on those measures (see Table 4).
Shock-Probe Defensive Burying
One male (chronic mild stress) contracted a localized bacterialskin infection (Staphylococcus aureus) and 1 male (severe spo-radic stress) contracted an ear infection prior to the burying test.Those 2 rats were euthanized (by CO2 gassing) prior to testing,leaving a total of 88 rats. Because of technical problems, the digitalvideotapes for 5 rats (2 no-stress males, 1 chronic mild stressfemale, and 2 severe sporadic stress females) were not availablefor analysis. One control female and 2 chronic mild stress femalesdisplayed burying scores that were greater than 2.5 standard de-viations away from the mean for their rearing group, and their datawere excluded from the analysis.
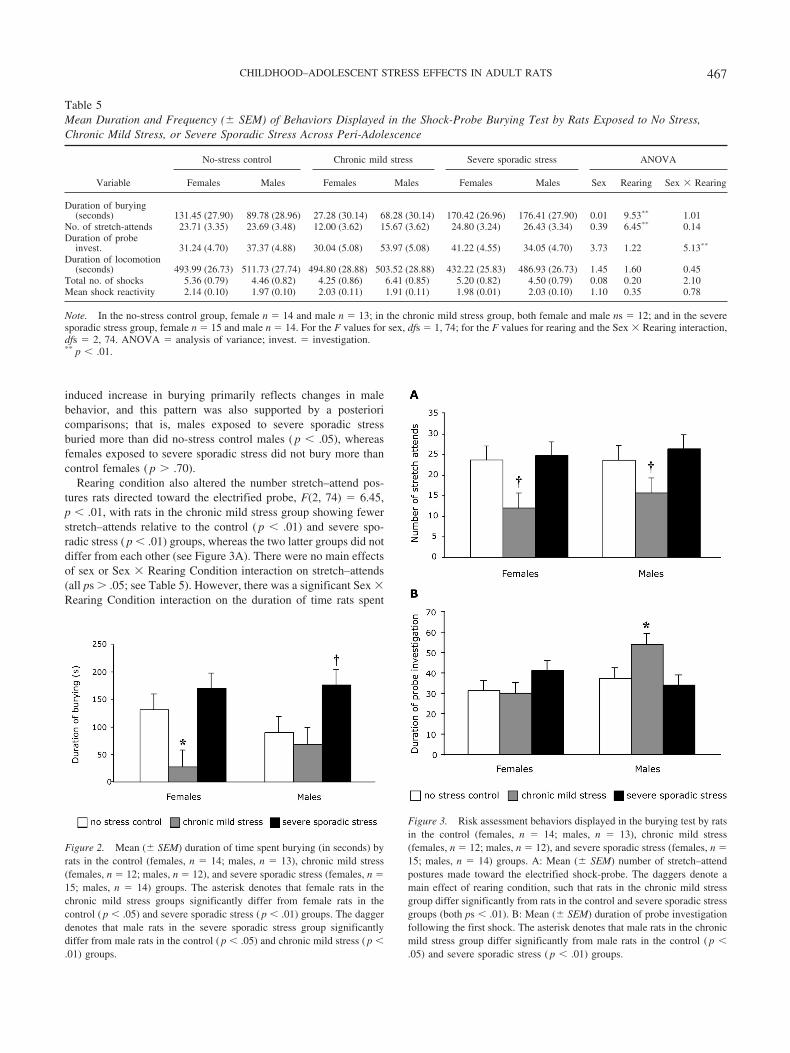
Data from the defensive burying test are provided in Table 5.The duration of time rats spent burying the electrified shock-probewas altered by rearing condition, F(2, 74) � 9.53, p � .001 (seeFigure 2). Post hoc analysis (LSD test) confirmed that rats exposedto sporadic severe stress across the childhood–adolescent perioddisplayed increases in burying behavior relative to rats exposed toeither no stress ( p � .05) or chronic mild stress ( p � .01). Ratsexposed to chronic mild stress displayed an opposite pattern; thatis, reductions in burying relative to the no-stress control rats ( p �.05). Although there was no main effect of sex or Sex � RearingCondition interaction on burying (both ps � .05), inspection ofFigure 2 suggests that the chronic-mild-stress-induced blunting ofburying behavior is most evident in female rats. A posteriorcomparisons confirmed that females exposed to chronic mild stressburied less than did control females ( p � .01), whereas malesexposed to chronic mild stress did not differ from control males( p � .50). Conversely, it appeared that the severe-sporadic-stress-
Figure 1. Changes in open-arm activity in the elevated plus-maze dis-played by rats in the control (females, n � 15; males, n � 15), chronic mildstress (females, n � 15; males, n � 15), and severe sporadic stress(females, n � 15; males, n � 15) groups. A: Mean (� SEM) percentage oftime spent on the open arms of the elevated plus-maze. Asterisks denote amain effect of rearing condition, such that rats in the severe sporadic stressgroup significantly differ from rats in the control and chronic mild stressgroups (both ps � .01). B: The percentage of rats in each group thatjumped off the open arms, at least once, during the 5-min elevated plus-maze test. Asterisks denote significant differences relative to control ( p �.01) and chronic mild stress ( p � .05) groups.
Table 4Mean Duration and Frequency (� SEM) of Behaviors in the Elevated Plus-Maze Test by Rats Exposed to No Stress, Chronic MildStress, or Severe Sporadic Stress Across Peri-Adolescence
Variable
No-stress control Chronic mild stress Severe sporadic stress ANOVA
Females Males Females Males Females Males Sex Rearing Sex � Rearing
% open-arm time 39.14 (4.63) 43.48 (4.63) 36.24 (4.63) 41.75 (4.63) 50.23 (4.63) 56.57 (4.63) 2.03 5.58** 0.02% open-arm entries 44.67 (3.84) 44.34 (3.84) 42.85 (3.84) 44.94 (3.84) 47.27 (3.84) 55.63 (3.84) 1.16 2.39 0.68No. of open-arm entries 6.53 (0.61) 5.80 (0.61) 6.13 (0.61) 5.47 (0.61) 5.80 (0.61) 5.87 (0.61) 0.80 0.22 0.28No. of closed-arm entries 8.07 (0.56) 6.28 (0.56) 7.87 (0.56) 6.53 (0.56) 6.47 (0.56) 5.07 (0.56) 11.06** 4.32* 0.10Total no. of entries 14.60 (0.95) 12.07 (0.95) 14.00 (0.95) 12.00 (0.95) 10.93 (0.95) 10.93 (0.95) 6.36** 1.88 0.20
Note. In each group, female n � 15 and male n � 15. For the F values for sex, dfs � 1, 84; for the F values for rearing and the Sex � Rearing interaction,dfs � 2, 84. ANOVA � analysis of variance.* p � .05. ** p � .01.
466 POHL ET AL.
induced increase in burying primarily reflects changes in malebehavior, and this pattern was also supported by a posterioricomparisons; that is, males exposed to severe sporadic stressburied more than did no-stress control males ( p � .05), whereasfemales exposed to severe sporadic stress did not bury more thancontrol females ( p � .70).
Rearing condition also altered the number stretch–attend pos-tures rats directed toward the electrified probe, F(2, 74) � 6.45,p � .01, with rats in the chronic mild stress group showing fewerstretch–attends relative to the control ( p � .01) and severe spo-radic stress ( p � .01) groups, whereas the two latter groups did notdiffer from each other (see Figure 3A). There were no main effectsof sex or Sex � Rearing Condition interaction on stretch–attends(all ps � .05; see Table 5). However, there was a significant Sex �Rearing Condition interaction on the duration of time rats spent
Figure 2. Mean (� SEM) duration of time spent burying (in seconds) byrats in the control (females, n � 14; males, n � 13), chronic mild stress(females, n � 12; males, n � 12), and severe sporadic stress (females, n �15; males, n � 14) groups. The asterisk denotes that female rats in thechronic mild stress groups significantly differ from female rats in thecontrol ( p � .05) and severe sporadic stress ( p � .01) groups. The daggerdenotes that male rats in the severe sporadic stress group significantlydiffer from male rats in the control ( p � .05) and chronic mild stress ( p �.01) groups.
Figure 3. Risk assessment behaviors displayed in the burying test by ratsin the control (females, n � 14; males, n � 13), chronic mild stress(females, n � 12; males, n � 12), and severe sporadic stress (females, n �15; males, n � 14) groups. A: Mean (� SEM) number of stretch–attendpostures made toward the electrified shock-probe. The daggers denote amain effect of rearing condition, such that rats in the chronic mild stressgroup differ significantly from rats in the control and severe sporadic stressgroups (both ps � .01). B: Mean (� SEM) duration of probe investigationfollowing the first shock. The asterisk denotes that male rats in the chronicmild stress group differ significantly from male rats in the control ( p �.05) and severe sporadic stress ( p � .01) groups.
Table 5Mean Duration and Frequency (� SEM) of Behaviors Displayed in the Shock-Probe Burying Test by Rats Exposed to No Stress,Chronic Mild Stress, or Severe Sporadic Stress Across Peri-Adolescence
Variable
No-stress control Chronic mild stress Severe sporadic stress ANOVA
Females Males Females Males Females Males Sex Rearing Sex � Rearing
Duration of burying(seconds) 131.45 (27.90) 89.78 (28.96) 27.28 (30.14) 68.28 (30.14) 170.42 (26.96) 176.41 (27.90) 0.01 9.53** 1.01
No. of stretch-attends 23.71 (3.35) 23.69 (3.48) 12.00 (3.62) 15.67 (3.62) 24.80 (3.24) 26.43 (3.34) 0.39 6.45** 0.14Duration of probe
invest. 31.24 (4.70) 37.37 (4.88) 30.04 (5.08) 53.97 (5.08) 41.22 (4.55) 34.05 (4.70) 3.73 1.22 5.13**
Duration of locomotion(seconds) 493.99 (26.73) 511.73 (27.74) 494.80 (28.88) 503.52 (28.88) 432.22 (25.83) 486.93 (26.73) 1.45 1.60 0.45
Total no. of shocks 5.36 (0.79) 4.46 (0.82) 4.25 (0.86) 6.41 (0.85) 5.20 (0.82) 4.50 (0.79) 0.08 0.20 2.10Mean shock reactivity 2.14 (0.10) 1.97 (0.10) 2.03 (0.11) 1.91 (0.11) 1.98 (0.01) 2.03 (0.10) 1.10 0.35 0.78
Note. In the no-stress control group, female n � 14 and male n � 13; in the chronic mild stress group, both female and male ns � 12; and in the severesporadic stress group, female n � 15 and male n � 14. For the F values for sex, dfs � 1, 74; for the F values for rearing and the Sex � Rearing interaction,dfs � 2, 74. ANOVA � analysis of variance; invest. � investigation.** p � .01.
467CHILDHOOD–ADOLESCENT STRESS EFFECTS IN ADULT RATS
exploring the shock-probe, F(2, 74) � 5.13, p � .01 (see Figure3B). One-way ANOVAs revealed that the duration of probe ex-ploration varied according to rearing condition in males, F(2,36) � 3.62, p � .01, but only tended to alter this response infemales, F( 2, 38) � 2.32, p � .06. Post hoc analysis of the maledata revealed that male rats in the chronic mild stress conditionspent more time engaged in shock-probe sniffing than did ratsin the no-stress and severe sporadic stress conditions (both ps �.05).
It is important to note that there were no main effects of sex orrearing condition or Sex � Rearing Condition interactions on theduration of time spent in general locomotor activity, total numberof shocks received, and the mean shock reactivity scores (seeTable 5).
Social Transmission of Food Preference
One female rat from the severe sporadic stress group developedbreathing difficulties and was euthanized (by CO2 gassing) prior tothe social transmission of food preference test. Further, 5 malesand 5 females from the control group along with 3 additional malesand 4 females (not part of the experimental colony) were used inpilot experiment to determine whether there was a natural prefer-ence for one of the two flavors of chow (cinnamon vs. cocoa) usedin the experiment. Rats used in this pilot study were not used in thepreference test. In addition, data from any rat that spilled thecontents of the food bowls containing the chow was not includedin the analysis. Thus, data from 73 rats (35 females and 38 males)were included in the analysis.
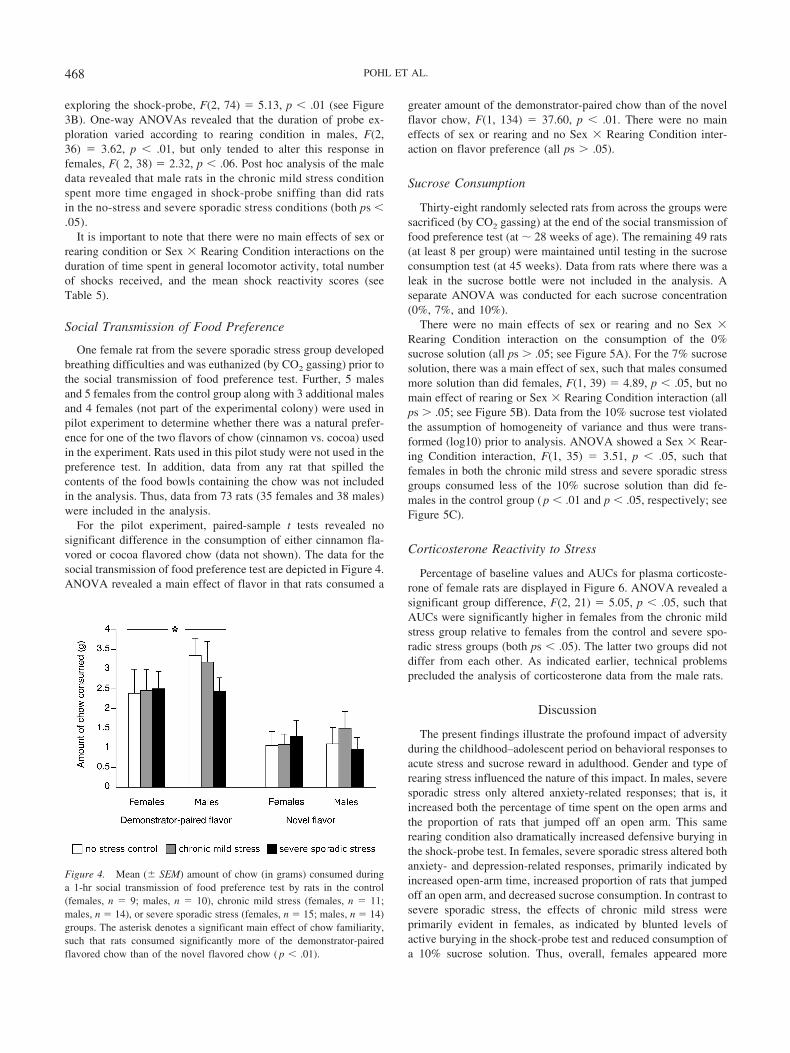
For the pilot experiment, paired-sample t tests revealed nosignificant difference in the consumption of either cinnamon fla-vored or cocoa flavored chow (data not shown). The data for thesocial transmission of food preference test are depicted in Figure 4.ANOVA revealed a main effect of flavor in that rats consumed a
greater amount of the demonstrator-paired chow than of the novelflavor chow, F(1, 134) � 37.60, p � .01. There were no maineffects of sex or rearing and no Sex � Rearing Condition inter-action on flavor preference (all ps � .05).
Sucrose Consumption
Thirty-eight randomly selected rats from across the groups weresacrificed (by CO2 gassing) at the end of the social transmission offood preference test (at 28 weeks of age). The remaining 49 rats(at least 8 per group) were maintained until testing in the sucroseconsumption test (at 45 weeks). Data from rats where there was aleak in the sucrose bottle were not included in the analysis. Aseparate ANOVA was conducted for each sucrose concentration(0%, 7%, and 10%).
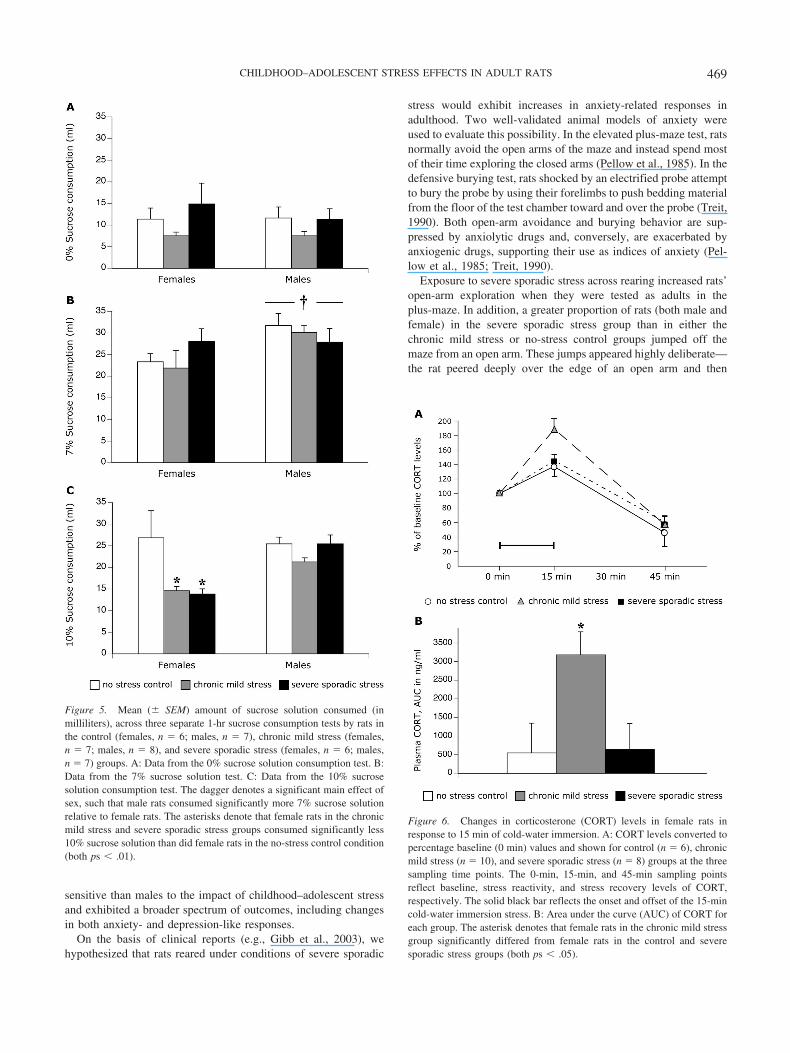
There were no main effects of sex or rearing and no Sex �Rearing Condition interaction on the consumption of the 0%sucrose solution (all ps � .05; see Figure 5A). For the 7% sucrosesolution, there was a main effect of sex, such that males consumedmore solution than did females, F(1, 39) � 4.89, p � .05, but nomain effect of rearing or Sex � Rearing Condition interaction (allps � .05; see Figure 5B). Data from the 10% sucrose test violatedthe assumption of homogeneity of variance and thus were trans-formed (log10) prior to analysis. ANOVA showed a Sex � Rear-ing Condition interaction, F(1, 35) � 3.51, p � .05, such thatfemales in both the chronic mild stress and severe sporadic stressgroups consumed less of the 10% sucrose solution than did fe-males in the control group ( p � .01 and p � .05, respectively; seeFigure 5C).
Corticosterone Reactivity to Stress
Percentage of baseline values and AUCs for plasma corticoste-rone of female rats are displayed in Figure 6. ANOVA revealed asignificant group difference, F(2, 21) � 5.05, p � .05, such thatAUCs were significantly higher in females from the chronic mildstress group relative to females from the control and severe spo-radic stress groups (both ps � .05). The latter two groups did notdiffer from each other. As indicated earlier, technical problemsprecluded the analysis of corticosterone data from the male rats.
Discussion
The present findings illustrate the profound impact of adversityduring the childhood–adolescent period on behavioral responses toacute stress and sucrose reward in adulthood. Gender and type ofrearing stress influenced the nature of this impact. In males, severesporadic stress only altered anxiety-related responses; that is, itincreased both the percentage of time spent on the open arms andthe proportion of rats that jumped off an open arm. This samerearing condition also dramatically increased defensive burying inthe shock-probe test. In females, severe sporadic stress altered bothanxiety- and depression-related responses, primarily indicated byincreased open-arm time, increased proportion of rats that jumpedoff an open arm, and decreased sucrose consumption. In contrast tosevere sporadic stress, the effects of chronic mild stress wereprimarily evident in females, as indicated by blunted levels ofactive burying in the shock-probe test and reduced consumption ofa 10% sucrose solution. Thus, overall, females appeared more
Figure 4. Mean (� SEM) amount of chow (in grams) consumed duringa 1-hr social transmission of food preference test by rats in the control(females, n � 9; males, n � 10), chronic mild stress (females, n � 11;males, n � 14), or severe sporadic stress (females, n � 15; males, n � 14)groups. The asterisk denotes a significant main effect of chow familiarity,such that rats consumed significantly more of the demonstrator-pairedflavored chow than of the novel flavored chow ( p � .01).
468 POHL ET AL.
sensitive than males to the impact of childhood–adolescent stressand exhibited a broader spectrum of outcomes, including changesin both anxiety- and depression-like responses.
On the basis of clinical reports (e.g., Gibb et al., 2003), wehypothesized that rats reared under conditions of severe sporadic
stress would exhibit increases in anxiety-related responses inadulthood. Two well-validated animal models of anxiety wereused to evaluate this possibility. In the elevated plus-maze test, ratsnormally avoid the open arms of the maze and instead spend mostof their time exploring the closed arms (Pellow et al., 1985). In thedefensive burying test, rats shocked by an electrified probe attemptto bury the probe by using their forelimbs to push bedding materialfrom the floor of the test chamber toward and over the probe (Treit,1990). Both open-arm avoidance and burying behavior are sup-pressed by anxiolytic drugs and, conversely, are exacerbated byanxiogenic drugs, supporting their use as indices of anxiety (Pel-low et al., 1985; Treit, 1990).
Exposure to severe sporadic stress across rearing increased rats’open-arm exploration when they were tested as adults in theplus-maze. In addition, a greater proportion of rats (both male andfemale) in the severe sporadic stress group than in either thechronic mild stress or no-stress control groups jumped off themaze from an open arm. These jumps appeared highly deliberate—the rat peered deeply over the edge of an open arm and then
Figure 5. Mean (� SEM) amount of sucrose solution consumed (inmilliliters), across three separate 1-hr sucrose consumption tests by rats inthe control (females, n � 6; males, n � 7), chronic mild stress (females,n � 7; males, n � 8), and severe sporadic stress (females, n � 6; males,n � 7) groups. A: Data from the 0% sucrose solution consumption test. B:Data from the 7% sucrose solution test. C: Data from the 10% sucrosesolution consumption test. The dagger denotes a significant main effect ofsex, such that male rats consumed significantly more 7% sucrose solutionrelative to female rats. The asterisks denote that female rats in the chronicmild stress and severe sporadic stress groups consumed significantly less10% sucrose solution than did female rats in the no-stress control condition(both ps � .01).
Figure 6. Changes in corticosterone (CORT) levels in female rats inresponse to 15 min of cold-water immersion. A: CORT levels converted topercentage baseline (0 min) values and shown for control (n � 6), chronicmild stress (n � 10), and severe sporadic stress (n � 8) groups at the threesampling time points. The 0-min, 15-min, and 45-min sampling pointsreflect baseline, stress reactivity, and stress recovery levels of CORT,respectively. The solid black bar reflects the onset and offset of the 15-mincold-water immersion stress. B: Area under the curve (AUC) of CORT foreach group. The asterisk denotes that female rats in the chronic mild stressgroup significantly differed from female rats in the control and severesporadic stress groups (both ps � .05).
469CHILDHOOD–ADOLESCENT STRESS EFFECTS IN ADULT RATS

dropped or leapt head first from the maze. Jumping was easilydistinguished from falling. Two control rats lost their balance (e.g.,while rearing) and fell sideways from the maze after attempting toprevent their fall with a righting response. Struggling or rightingattempts were never seen with jumping behavior.
One intriguing explanation for this pattern (i.e., increases inopen-arm exploration concomitant with increases in the proportionof rats that jumped off the maze) is that exposure to severesporadic stress across development shifted rats’ coping strategy inthe plus-maze from a passive (withdrawal–avoidance) to a moreactive (exploration–escape) coping strategy (for a review of pas-sive and active coping styles, see Koolhaas et al., 1999). A relatedpossibility is that relatively low to moderate levels of fear mightmotivate rats to withdraw to the closed arms as a means to avoidthe more aversive open arms. In contrast, high levels of fear mightmotivate rats to search for an escape route from the maze (Kalyn-chuk et al., 1997). In our study, severe sporadic stress rats oftentraveled either midway or to the far end of an open arm, where theystopped and visually scanned the regions beyond the edge of thearm, using back and forth movements of the head with the upperbody and/or head extended forward. Such scanning behavior oftenpreceded jumping. Although not quantified, higher levels of scan-ning might account for the increase in open-arm time displayed bythe severe sporadic stress rats. Notably, our unexpected findingsare not without precedence. Long-term amygdala-kindled rats dis-play increases in open-arm exploration and open-arm jumps in theplus-maze, concomitant with heightened fear responses in otheranimal models of anxiety (Kalynchuk, 2000; Kalynchuk et al.,1997). Also, prior exposure to either severe or chronic intermittentstressors during adulthood increases rats’ open-arm explorationwhile exacerbating fear-related responding in other tests(D’Aquila, Brain, & Willner, 1994; Renard, Suarez, Levin, &Rivarola, 2005; Zhukov & Vinogradova, 1994).
Consistent with the idea that severe sporadic stress promotes anactive, perhaps more fearful, defense profile, rats reared in thatcondition displayed dramatic increases in shock-probe buryingbehavior relative both to controls and to rats exposed to chronicmild stress. This increase in burying was selective in that it was notaccompanied by changes in either the number of shocks received,mean shock reactivity, or general locomotor activity. Further, aposterior analysis of the data suggested that the effect of severesporadic stress on burying was primarily evident in males. Becausethis sex difference was only supported after the fact, it needs to beconsidered with caution. This said, in accordance with prior re-ports, we saw no evidence of sex-related differences in buryingbehavior in our control rats (Maggio & Harder, 1983; Sluyter et al.,1996; Treit, Terlecki, & Pinel, 1980). Few studies have examinedthe possible influence of estrous cycling on burying behavior. Onefound that proestrus, estrus, and diestrus females bury the shock-probe at levels that are indistinguishable from those of males(Frye, Petralia, & Rhodes, 2000). By contrast, there is someevidence that females in metestrus (diestrus I) bury more thanmales (Fernandez-Guasti & Picazo, 1990). Thus, we cannot pre-clude the possibility that the hormonal status of the female rats didnot contribute to the behavioral profiles we observed in this or anyof our tests.
Exposure to chronic mild stress during the childhood–adolescent period produced a completely opposite profile to that ofsevere sporadic stress; that is, chronic mild stress dramatically
suppressed burying relative to both the control and the severesporadic stress condition. Of interest, a posterior analysis sug-gested that the effects of chronic mild stress on burying wereprimarily apparent in female rats. The possibility that females aremore sensitive than males to chronic mild stress during thechildhood–adolescent period needs to be further substantiated. Atany rate, the blunted levels of burying we observed could not beaccounted for by changes in pain sensitivity because chronic-mild-stress-exposed rats did not differ from the other groups in terms oftheir shock-probe avoidance or mean shock reactivity. Chronicmild stress also suppressed risk assessment responses. However,although both males and females made fewer stretch–attend pos-tures toward the shock-probe, male rats displayed a compensatoryresponse (i.e., significant increases in probe investigation). Thislatter finding is consistent with the finding that males in thechronic mild stress group buried the probe as much as males in thecontrol group.
It could be argued that exposure to chronic mild stress through-out the childhood–adolescent developmental period exerted pro-tective actions, at least in females, ultimately reducing their sen-sitivity to stress in later life. However, this interpretation isinconsistent with the stress-induced increases in corticosterone weobserved in females from the chronic mild stress group. Othershave argued that, under certain conditions, lower levels of buryingmight, in fact, reflect a maladaptive response. Thus, Wistar–Kyoto(WKY) rats display blunted levels of burying in the shock-probetest despite being hypersensitive to stress in other paradigms (Pare,1992, 1994). The WKY strain is widely regarded as a geneticmodel of depression in that WKY rats develop stress-inducedulcers more readily than other strains and display exaggeratedhypothalamic–pituitary–adrenal axis responses to acute stress, re-duced sensitivity to reward, and more depression-like behaviorwhen tested in either the forced swim test or inescapable shock–learned helplessness paradigm (De la Garza, 2005; Morilak et al.,2005). In the context of this broader profile, it has been suggestedthat the lower levels of burying displayed by WKY rats reflecttheir predisposition to respond to environmental threats with highlevels of passivity and blunted behavioral responsivity (Morilak etal., 2005; Pare, 1992, 1994; Rittenhouse, Lopez-Rubalcava, Stan-wood, & Lucki, 2002). Clearly, further studies are needed todetermine whether exposure to chronic mild stress during thechildhood–adolescent period gives rise to a predominantlydepressive-like profile in female rats. However, the findings thusfar are consistent with this possibility in that female rats in thechronic mild stress group displayed a profile of reduced buryingbehavior, exaggerated corticosterone responses to stress, and re-duced consumption of a 10% sucrose solution (discussed furtherbelow).
We hypothesized that exposure to chronic mild stress through-out childhood and adolescence would lead to the development ofdepression-like responses in adulthood and that this effect wouldbe greater in females. In accordance with our hypothesis, female(but not male) rats in the chronic mild stress group consumed lessof the 10% sucrose solution relative to female control rats. Thesame effect was also observed in female (but not male) rats in thesevere sporadic stress group. These reductions in sucrose con-sumption were not secondary to neophobia, as there were no groupdifferences in the latency to approach the bottle spout (data notshown). Similarly, we did not observe group differences in the
470 POHL ET AL.

consumption of 0% sucrose (i.e., water) and body weight. Thus,the decreases in sucrose consumption are suggestive of an anhe-donic state rather than of general decreases in fluid consumption orfluid requirements. We did not observe group differences in theconsumption of a 7% sucrose solution. This likely reflects differ-ences in palatability between the 7% and 10% solutions. A sucroseconcentration of 7% was chosen on the basis of previous work inour laboratory and others (Montgomery & Suri, 1996; Montgom-ery & Willner, 1988; Vollmayr et al., 2004) showing that a sucroseconcentration of 7% offers the greatest hedonic value. Thus, a highhedonic value of the 7% sucrose might have masked the effects ofrearing stress on sensitivity to reward.
We also tested our rats in the social transmission of foodpreference test. Although not traditionally viewed as measuringanxiety-like or depression-like responses, we included this test inour battery because human anxiety disorders and clinical depres-sion are often accompanied by short-term memory deficits(Airaksinen et al., 2004, 2005; Landro et al., 2001). We saw noevidence of memory deficits in the current study. That is, regard-less of rearing group, all rats displayed a preference for the chowflavor that had been paired with the demonstrator rat. Rats’ abilityto acquire social transmission of food preference is hippocampaldependent (Eichenbaum, 1998; Ergorul & Eichenbaum, 2006), andhippocampal-dependant memory is disrupted following chronicstress in adulthood (Hui, Guang-Yu, Chong-Tao, Quan, & Xiao-Hu, 2005; Luine, Villegas, Martinez, & McEwen, 1994; McEwen,2005; Sousa, Lukoyanov, Madeira, Almeida, & Paula-Barbosa,2000). However, the memory impairments associated with adultchronic stress paradigms are transitory and are no longer apparentafter 21 days of no-stress recovery (Luine et al., 1994; Sousa et al.,2000). By contrast, exposing developing rats to chronic variablestress from PD 28 through to PD 56 has been shown to result inmemory deficits in a Morris water maze task that were still evident3 weeks following the end of the stress regimen (Isgor, Kabbaj,Akil, & Watson, 2004). Thus, we cannot discount that our ratsmight have had memory deficits in the period closely following theend of our stressors. In contrast, it is noteworthy that we foundeffects on anxiety- and depression-related responses long after theend of our rearing stress regimens (e.g., at least 7–10 weeks in thecase of the burying test and 38 weeks in the case of the sucroseconsumption test). These remarkably long-lasting effects stand insharp contrast to the relatively transient impact of chronic stressorsapplied in adulthood (e.g., Grippo, Beltz, & Johnson, 2003; Luineet al., 1994; Sousa et al., 2000; Willner, Sampson, Papp, Phillips,& Muscat, 1991).
Stress-related disruptions in the estrous cycle might account, atleast in part, for the generally higher sensitivity to acute stress weobserved in female (relative to male) rats exposed to stress fromweaning through to late adolescence–early adulthood. Chronicallyexposing female rats to stress in adulthood reliably disrupts estrouscycling, indexed primarily by longer, less frequent cycles (Baker,Kentner, Konkle, Santa-Maria Barbagallo, & Bielajew, 2006;Dalla et al., 2005; Gonzalez, Rodrıguez Echandıa, Cabrera, &Foscolo, 1994; Grippo et al., 2005; Konkle et al., 2003; RodrıguezEchandıa, Gonzalez, Cabrera, & Fracchia, 1988) and increasedtime in diestrus (Baker et al., 2006; Gonzalez et al., 1994; Rodrı-guez Echandıa et al., 1988). Recent evidence supports the possi-bility that similar disruptions in estrous cycling might have oc-curred in our study. Specifically, young female rats were exposed
to once-daily saline-injection stress from PD 21 through to PD 60(Raap, Morin, Medici, & Smith, 2000) and their postpubertalestrous phase status was monitored. Compared with uninjectedcontrol females, the saline-injected rats displayed no differences inpuberty onset (PD 36) and age at first ovulation (PD 37). Remark-ably, however, injection stress did reduce the proportion of regular4–5 day estrous cycles and substantively increased the number ofdays spent in diestrus between PD 41 to PD 60. These differenceswere transient and disappeared once injections ended on PD 60(Raap et al., 2000). By contrast, exposure to relatively moreintense stressors, as used in the current study, might producelonger lasting disruptions in estrous cycling. In support, althoughmany females exposed to chronic mild stress as adults eventuallydisplayed normal cycling, others showed little recovery even aftera 6-week stress-free period (Baker et al., 2006). Furthermore,although our stress manipulations ended on PD 51, our rats wereintermittently exposed to testing-related stressors through to theend of the study (at age 45 weeks).
Evidence from preclinical studies suggests that stress-inducedincreases in the length of diestrus might translate into exacerbatedbehavioral responses to acute (and potentially chronic) stress.Anxiety-related responses are generally higher in diestrus femalesthan in estrus and proestrus females (Dıaz-Veliz, Alarcon, Espi-noza, Dussaubat, & Mora, 1997; Fernandez-Guasti & Picazo,1990, 1992; Frye et al., 2000; Frye & Rhodes, 2006; Marcondes,Miguel, Melo, & Spadari-Bratfisch, 2001; Mora, Dussaubat, &Dıaz-Veliz, 1996), and, perhaps to a lesser extent, than in male rats(Dıaz-Veliz et al., 1997; Fernandez-Guasti & Picazo, 1990).Diestrus females also seem to be more vulnerable than estrusfemales to the induction of depression-like responses (Jenkins,Williams, Kramer, Davis, & Petty, 2001; Marvan et al., 1996,1997). As seen with the anxiety tests, male vulnerability to acquiredepression-like behavior in some (but not all) animal analogues ofdepression seems to fall midway between that of diestrus andestrus females (Jenkins et al., 2001). Clearly it would be prematureon the basis of behavioral data alone to conclude that at least someof our results reflect disruptions to estrous cycling. However, thepossibility is no less compelling, especially when considered inlight of the substantive overlap with affective disorders in womenand hormone fluctuations during menstrual-related events (Amer-ican Psychiatric Association, 2000).
To the best of our knowledge, the current study is the firstcomprehensive examination of whether stress across thechildhood–adolescent period influences affective responses ofmale and female rats in later life. Our finding that childhood–adolescent stress regimens produce long-lasting changes in emo-tionality echoes the relatively permanent alterations in sensitivityto stress that are known to follow stress during the prenatal(Maccari et al., 2003; Owen, Andrews, & Matthews, 2005) orpostnatal, that is, within the first 3 weeks of life (de Kloet, Sibug,Helmerhorst, & Schmidt, 2005; Gutman & Nemeroff, 2002;Holmes et al., 2005), period. Developmental processes are stillongoing during the childhood–adolescent period, including exten-sive synaptic overexpression followed by synaptic pruning(Andersen, 2003), as well as continued development of stress-responsive neural circuits (Choi, Weisberg, & Kellogg, 1997; Isgoret al., 2004). Throughout this period, the organism might incor-porate information into a relatively more stable form than wouldoccur in adulthood (Andersen, 2003; Nemeroff, 2004).
471CHILDHOOD–ADOLESCENT STRESS EFFECTS IN ADULT RATS

In summary, we attempted to mimic early adversity in the formof neglect and abuse using regimens of chronic mild stress andsevere sporadic stress, respectively. We acknowledge the inherentdifficulty of trying to model human experiences in the rat. None-theless, our stress regimens have a certain degree of face validity.In the chronic mild stress regime, varying stressors were present ona relatively continuous basis, similar to the enduring, chaoticfeatures of childhood neglect. In contrast, the severe sporadicstress condition involved randomly delivered brief but noxiousstressors, thus mimicking the random, periodic administration ofhighly noxious events that characterizes childhood physical abuse(Belsky, 1993). A positive indicator that our stress regimens couldbe useful frameworks for modeling childhood abuse and neglectresides in the general concordance between our findings and thoseobserved in preliminary studies in humans. As such, the presentfindings lend important validity to the hypothesis that childhoodneglect is preferentially associated with later depression, whereaschildhood abuse is preferentially associated with later anxiety(Gibb et al., 2003; Harkness & Wildes, 2002). In our study,chronic mild stress across the childhood–adolescent period led todepression-like responses in adulthood, whereas severe sporadicstress was associated with increases in both anxiety- anddepression-related behaviors. Further, the depression-like re-sponses were observed in the female, but not male, rats, whereasthe anxiety-like responses were observed in rats of both sexes.Depression in the clinic is twice as common in women as in men,and this sex difference emerges in late childhood (age 10–14years; American Psychiatric Association, 2000). Of specific rele-vance to the present findings, prominent theories for the genderdifference in depression point to higher stress sensitivity in womenas a primary mechanism (e.g., Cyranowski, Frank, Young, &Shear, 2000). Anxiety and most anxiety disorders are also morecommon in girls and women (Mackinaw-Koons & Vasey, 2000).However, the cause of the sex difference in anxiety remains largelyundetermined and may relate primarily to other, non-stress-relatedetiological factors (Craske, 2003). Depression is the second lead-ing cause of disability worldwide, and the direct and indirect costsof depression and anxiety total in the billions of dollars (Murray &Lopez, 1996). Understanding the fine-grained etiological precur-sors to depression versus anxiety may help refine existing treat-ments, with the ultimate goal of preventing these devastatingconditions. At a minimum, the current findings underscore theimportance of considering gender and stressor type when examin-ing the impact of prior chronic stress on behavioral responding. Ofmore importance, they raise the possibility that chronic stressthroughout the childhood–adolescent period leads to relativelylong-lasting changes in emotionality.
References
Airaksinen, E., Larsson, M., & Forsell, Y. (2005). Neuropsychologicalfunctions in anxiety disorders in population-based samples: Evidence ofepisodic memory dysfunction. Journal of Psychiatric Research, 39,207–214.
Airaksinen, E., Larsson, M., Lundberg, I., & Forsell, Y. (2004). Cognitivefunctions in depressive disorders: Evidence from a population-basedstudy. Psychological Medicine, 34, 83–91.
American Psychiatric Association. (2000). Diagnostic and statistical man-ual of mental disorders (4th ed., text rev.). Washington, DC: Author.
Andersen, S. L. (2003). Trajectories of brain development: Point of vul-
nerability or window of opportunity? Neuroscience & BiobehavioralReviews, 27, 3–18.
Baker, S. L., Kentner, A. C., Konkle, A. T. M., Santa-Maria Barbagallo, L.,& Bielajew, C. (2006). Behavioral and physiological effects of chronicmild stress in female rats. Physiology & Behavior, 87, 314–322.
Belsky, J. (1993). Etiology of child maltreatment: A developmental–ecological analysis. Psychological Bulletin, 114, 413–434.
Blanchard, R. J., & Blanchard, D. C. (1989). Attack and defense in rodentsas ethoexperimental models for the study of emotion. Progress inNeuropsychopharmacology and Biological Psychiatry, 13, 3–14.
Brown, G. W., & Harris, T. O. (1993). Aetiology of anxiety and depressivedisorders in an inner-city population. 1. Early adversity. PsychologicalMedicine, 23, 143–154.
Choi, S., Weisberg, S. N., & Kellogg, C. K. (1997). Control of endogenousnorepinephrine release in the hypothalamus of male rats changes overadolescent development. Brain Research Developmental Brain Re-search, 98, 134–141.
Clark, D. A., Beck, A. T., & Stewart, B. (1990). Cognitive specificity andpositive–negative affectivity: Complementary or contradictory views onanxiety and depression? Journal of Abnormal Psychology, 99, 148–155.
Cowen, P. S. (1999). Child neglect: Injuries of omission. Pediatric Nurs-ing, 95, 401–418.
Craske, M. G. (2003). Origins of phobias and anxiety disorders: Why morewomen than men? New York: Elsevier.
Cyranowski, J. M., Frank, E., Young, E., & Shear, M. K. (2000). Adoles-cent onset of the gender difference in lifetime rates of major depression:A theoretical model. Archives of General Psychiatry, 57, 21–27.
Dalla, C., Antoniou, K., Drossopoulou, G., Xagoraris, X., Kokras, K.,Sfikakis, A., & Papapdopoulou-Daifoti, Z. (2005). Chronic mild stressimpact: Are females more vulnerable? Neuroscience, 135, 703–714.
D’Aquila, P. S., Brain, P., & Willner, P. (1994). Effects of chronic mildstress on performance in behavioural tests relevant to anxiety anddepression. Physiology & Behavior, 56, 861–867.
De Bellis, M. D. (2005). The psychobiology of neglect. Child Maltreat-ment, 10, 150–172.
de Kloet, E. R., Sibug, R. M., Helmerhorst, F. M., & Schmidt, M. (2005).Stress, genes and the mechanism of programming the brain for later life.Neuroscience & Biobehavioral Reviews, 29, 271–281.
De la Garza, R., II. (2005). Wistar–Kyoto rats exhibit reduced sucrosepellet reinforcement behavior and intravenous nicotine self-administration. Pharmacology Biochemistry and Behavior, 82, 330–337.
Dıaz-Veliz, G., Alarcon, T., Espinoza, C., Dussaubat, N., & Mora, S.(1997). Ketanserin and anxiety levels: Influence of gender, estrous cycle,ovariectomy and ovarian hormones in female rats. Pharmacology Bio-chemistry and Behavior, 58, 637–642.
Eichenbaum, H. (1998). Using olfaction to study memory. Annals of theNew York Academy of Sciences, 855, 657–669.
Ergorul, C., & Eichenbaum, H. (2006). Essential role of the hippocampalformation in rapid learning of higher-order sequential associations. Jour-nal of Neuroscience, 26, 4111–4117.
Fernandez-Guasti, A., & Picazo, O. (1990). The actions of diazepam andserotonergic anxiolytics vary according to the gender and the estrouscycle phase. Pharmacology, Biochemistry and Behavior, 37, 77–81.
Fernandez-Guasti, A., & Picazo, O. (1992). Changes in burying behaviorduring the estrous cycle: Effect of estrogen and progesterone. Psycho-neuroendocrinology, 17, 681–689.
Frye, C. A., Petralia, S. M., & Rhodes, M. E. (2000). Estrous cycle and sexdifferences in performance on anxiety tasks coincide with increases inhippocampal progesterone and 3alpha,5alpha-THP. Pharmacology Bio-chemistry and Behavior, 67, 587–596.
Frye, C. A., & Rhodes, M. E. (2006). Progestin concentrations are in-creased following paced mating in midbrain, hippocampus, diencepha-
472 POHL ET AL.

lon, and cortex of rats in behavioral estrus, but only in midbrain ofdiestrous rats. Neuroedocrinology, 83, 336–347.
Galef, B. G., & Wigmore, S. W. (1983). Transfer of information concern-ing distant foods: A laboratory investigation of the “information center”hypothesis. Animal Behaviour, 31, 748–758.
Gibb, B. E., Butler, A. C., & Beck, J. S. (2003). Childhood abuse,depression, and anxiety in adult psychiatric outpatients. Depression andAnxiety, 17, 226–228.
Gonzalez, A. S., Rodrıguez Echandıa, E. L., Cabrera, R., & Foscolo, M. R.(1994). Neonatal chronic stress induces subsensitivity to chronic stressin adult rats: II. Effects on estrous cycle in females. Physiology &Behavior, 56, 591–595.
Grippo, A. J., Beltz, T. G., & Johnson, A. K. (2003). Behavioral andcardiovascular changes in the chronic mild stress model of depression.Physiology & Behavior, 78, 703–710.
Grippo, A. J., Sullivan, N. R., Damjanoska, K. J., Crane, J. W., Carrasco,G. A., Shi, J., et al. (2005). Chronic mild stress induces behavioral andphysiological changes, and may alter serotonin 1A receptor function, inmale and cycling female rats. Psychopharmacology, 179, 769–780.
Gutman, D. A., & Nemeroff, C. B. (2002). Neurobiology of early lifestress: Rodent studies. Seminars in Clinical Neuropsychiatry, 7, 89–95.
Hankin, B. L. (2005). Childhood maltreatment and psychopathology: Pro-spective tests of attachment, cognitive vulnerability, and stress as me-diating processes. Cognitive Therapy and Research, 29, 645–671.
Harkness, K. L., & Wildes, J. E. (2002). Childhood adversity and anxietyversus dysthymia co-morbidity in major depression. Physiological Med-icine, 32, 1239–1249.
Heim, C., & Nemeroff, C. B. (2001). The role of childhood trauma in theneurobiology of mood and anxiety disorders: Preclinical and clinicalstudies. Biological Psychiatry, 49, 1023–1039.
Hildyard, K. L., & Wolfe, D. A. (2002). Child neglect: Developmentalissues and outcomes. Child Abuse and Neglect, 26, 679–695.
Holmes, A., le Guisquet, A. M., Vogel, E., Millstein, R. A., Leman, S., &Belzung, C. (2005). Early life genetic, epigenetic and environmentalfactors shaping emotionality in rodents. Neuroscience & BiobehavioralReviews, 29, 1335–1346.
Hui, Z., Guang-Yu, M., Chong-Tao, X., Quan, Y., & Xiao-Hu, X. (2005).Phenytoin reverses the chronic stress-induced impairment of memoryconsolidation for water maze training and depression of LTP in rathippocampal CA1 region, but does not affect motor activity. CognitiveBrain Research, 24, 380–385.
Isgor, C., Kabbaj, M., Akil, H., & Watson, S. J. (2004). Delayed effects ofchronic variable stress during peripubertal–juvenile period on hippocam-pal morphology and on cognitive and stress axis functions in rats.Hippocampus, 14, 636–648.
Jenkins, J. A., Williams, P., Kramer, G. L., Davis, L. L., & Petty, F. (2001).The influence of gender and the estrous cycle on learned helplessness inthe rat. Biological Psychology, 58, 147–158.
Kalynchuk, L. E. (2000). Long-term amygdala kindling in rats as a modelfor the study of interictal emotionality in temporal lobe epilepsy. Neu-roscience & Biobehavioral Reviews, 24, 691–704.
Kalynchuk, L. E., Pinel, J. P., Treit, D., & Kippin, T. E. (1997). Changesin emotional behavior produced by long-term amygdala kindling in rats.Biological Psychiatry, 41, 438–451.
Konkle, A. T. M., Baker, S. L., Kentner, A. C., Santa-Maria Barbagallo, L.,Merali, Z., & Bielajew, C. (2003). Evaluation of the effects of chronicmild stressors on hedonic and physiological responses: Sex and straincompared. Brain Research, 992, 227–238.
Koolhaas, J. M., Korte, S. M., De Boer, S. F., Van Der Vegt, B. J., VanReenen, C. G., Hopster, H., et al. (1999). Coping styles in animals:Current status in behavior and stress-physiology. Neuroscience & Biobe-havioral Reviews, 23, 925–935.
Landro, N. I., Stiles, T. C., & Sletvold, H. (2001). Neuropsychological
function in nonpsychotic unipolar major depression. Neuropsychiatry,Neuropsychology, and Behavioral Neurology, 14, 233–240.
Levitan, R. D., Rector, N. A., Sheldon, T., & Goering, P. (2003). Child-hood adversities associated with major depression and/or anxiety disor-ders in a community sample of Ontario: Issues of co-morbidity andspecificity. Depression and Anxiety, 17, 34–42.
Luecken, L. J., & Lemery, K. S. (2004). Early caregiving and physiologicalstress responses. Clinical Psychology Review, 24, 171–191.
Luine, V., Villegas, M., Martinez, C., & McEwen, B. S. (1994). Repeatedstress causes reversible impairments of spatial memory performance.Brain Research, 639, 167–170.
Maccari, S., Darnaudery, M., Morley-Fletcher, S., Zuena, A. R., Cinque,C., & Van Reeth, O. (2003). Prenatal stress and long-term consequences:Implications of glucocorticoid hormones. Neuroscience & Biobehav-ioral Reviews, 27, 119–127.
Mackinaw-Koons, B., & Vasey, M. W. (2000). Considering sex differencesin anxiety and its disorders across the life span: A construct-validationapproach. Applied and Preventive Psychology, 9, 191–209.
Maggio, J. C., & Harder, D. B. (1983). Genotype and environment inter-actively determine the magnitude, directionality, and abolition of defen-sive burying in mice. Animal Learning & Behavior, 11, 162–172.
Marcondes, F. K., Miguel, K. J., Melo, L. L., & Spadari-Bratfisch, R. C.(2001). Estrous cycle influences the response of female rats in theelevated plus-maze test. Physiology & Behavior, 74, 435–440.
Marvan, M. L., Chavez-Chavez, L., & Santana, S. (1996). Clomipraminemodifies fluctuations of forced swimming immobility in different phasesof the rat estrous cycle. Archives of Medical Research, 27, 83–87.
Marvan, M. L., Santana, S., Chavez, L. C., & Bertran, M. (1997). Ines-capable shocks accentuate fluctuations of forced swimming immobilityin different phases of the rat estrous cycle. Archives of Medical Re-search, 28, 369–372.
McEwen, B. S. (2003). Early life influences on life-long patterns ofbehavior and health. Mental Retardation and Developmental DisabilitiesResearch Reviews, 9, 149–154.
McEwen, B. S. (2005). Glucocorticoids, depression, and mood disorders:Structural remodeling in the brain. Metabolism: Clinical and Experi-mental, 54(5, Suppl. 1), 20–23.
Montgomery, A. M. J., & Suri, A. (1996). Potentiation of the effects ofraclopride on sucrose consumption by the 5-HT2 antagonist ritanserin.Psychopharmacology, 123, 98–102.
Montgomery, A. M. J., & Willner, P. (1988). Fenfluramine disrupts thebehavioural satiety sequence in rats. Psychopharmacology, 94, 397–401.
Mora, S., Dussaubat, N., & Dıaz-Veliz, G. (1996). Effects of the estrouscycle and ovarian hormones on behavioural indices of anxiety in femalerats. Psychoneuroendocrinology, 21, 609–620.
Morilak, D. A., Barrera, G., Echevarria, D. J., Garcia, A. S., Hernandez, A.,Ma, S., & Petre, C. O. (2005). Role of brain norepinephrine in thebehavioral response to stress. Progress in Neuro-Psychopharmacology& Biological Psychiatry, 29, 1214–1224.
Murray, C. J. L., & Lopez, A. C. (1996). Global burden of disease andinjury. Boston: Harvard School of Public Health.
Nemeroff, C. B. (2004). Neurobiological consequences of childhoodtrauma. Journal of Clinical Psychiatry, 65, 18–28.
Ojeda, S. R., & Urbanski, H. F. (1994). Puberty in the rat. In E. Knobil &J. D. Neil (Eds.), The physiology of reproduction (2nd ed., pp. 363–409).New York: Raven.
Owen, D., Andrews, M. H., & Matthews, S. G. (2005). Maternal adversity,glucocorticoids and programming of neuroendocrine function and be-haviour. Neuroscience & Biobehavioral Reviews, 29, 209–226.
Papp, M., Willner, P., & Muscat, R. (1991). An animal model of anhedoniaattenuation of sucrose consumption and place preference conditioningby chronic unpredictable mild stress. Psychopharmacology (Berlin),104, 255–259.
473CHILDHOOD–ADOLESCENT STRESS EFFECTS IN ADULT RATS

Pare, W. P. (1992). The performance of WKY rats on three tests ofemotional behavior. Physiology & Behavior, 51, 1051–1056.
Pare, W. P. (1994). Open field, learned helplessness, conditioned defensiveburying, and forced-swim tests in WKY rats. Physiology & Behavior,55, 433–439.
Pecoraro, N., Dallman, M. F., Warne, J. P., Ginsberg, A. B., Laugero,K. D., la Fleur, S. E., et al. (2006). From Malthus to motive: How theHPA axis engineers the phenotype, yoking needs to wants. Progress inNeurobiology, 79, 247–340.
Pellow, S., Chopin, P., File, S. E., & Briley, M. (1985). Validation ofopen:closed arm entries in an elevated plus-maze as a measure of anxietyin the rat. Journal of Neuroscience Methods, 14, 149–167.
Piccinelli, M., & Wilkinson, G. (2000). Gender differences in depression.Critical review. British Journal of Psychiatry, 177, 486–492.
Raap, D. K., Morin, B., Medici, C. N., & Smith, R. F. (2000). Adolescentcocaine and injection stress effects on the estrous cycle. Physiology &Behavior, 70, 417–424.
Renard, G. M., Suarez, M. M., Levin, G. M., & Rivarola, M. A. (2005).Sex differences in rats: Effects of chronic stress on sympathetic systemand anxiety. Physiology & Behavior, 85, 363–369.
Rittenhouse, P. A., Lopez-Rubalcava, C., Stanwood, G. D., & Lucki, I.(2002). Amplified behavioral and endocrine responses to forced swimstress in the Wistar–Kyoto rat. Psychoneuroendocrinology, 27, 303–318.
Rodrıguez Echandıa, E. L., Gonzalez, A. S., Cabrera, R., & Fracchia, L. N.(1988). A further analysis of behavioral and endocrine effects of unpre-dictable chronic stress. Physiology & Behavior, 43, 789–795.
Sluyter, F., Korte, S. M., Bohus, B., & Van Oortmerssen, G. A. (1996).Behavioral stress response of genetically selected aggressive and non-aggressive wild house mice in the shock-probe/defensive burying test.Pharmacology Biochemistry and Behavior, 54, 113–116.
Sousa, N., Lukoyanov, N. V., Madeira, M. D., Almeida, O. F., & Paula-Barbosa, M. M. (2000). Reorganization of the morphology of hippocam-pal neurites and synapses after stress-induced damage correlates withbehavioral improvement. Neuroscience, 97, 253–266.
Spear, L. P. (2000). The adolescent brain and age-related behaviouralmanifestations. Neuroscience & Biobehavioral Reviews, 24, 417–463.
Spear, L. P., & Brake, S. C. (1983). Periadolescence: Age dependentbehavior and psychopharmacological responsivity in rats. Developmen-tal Psychobiology, 16, 83–109.
Thornberry, T. P., Ireland, T. O., & Smith, C. A. (2001). The importanceof timing: The varying impact of childhood and adolescent maltreatmenton multiple problem outcomes. Development and Psychopathology, 13,957–979.
Treit, D. (1990). A comparison of anxiolytic and nonanxiolytic agents inthe shock-probe/burying test for anxiolytics. Pharmacology Biochemis-try and Behavior, 36, 203–205.
Treit, D., Terlecki, L. J., & Pinel, J. P. J. (1980). Conditioned defensiveburying: Organismic variables. Bulletin of the Psychonomic Society, 16,451–454.
Vollmayr, B., Bachteler, D., Vengeliene, V., Gass, P., Spanagel, R., &Henn, F. (2004). Rats with congenital learned helplessness respond lessto sucrose but show no deficits in activity or learning. Behavioural BrainResearch, 150, 217–221.
Weiss, E. L., Longhurst, J. G., & Mazure, C. M. (1999). Childhood sexualabuse as a risk factor for depression in women: Psychosocial andneurobiological correlates. American Journal of Psychiatry, 156, 816–828.
Willner, P. (1986). Validation criteria for animal models of human mentaldisorders: Learned helplessness as a paradigm case. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 10, 677–690.
Willner, P., Sampson, D., Papp, M., Phillips, G., & Muscat, R. (1991).Animal models of anhedonia. In P. Soubrie (Ed.), Anxiety, depression,and mania. Animal models of psychiatric disorders (pp. 71–99). Basel,Switzerland: Karger.
Zhukov, D. A., & Vinogradova, K. P. (1994). Inescapable shock inducesthe opposite changes of the plus-maze test behavior in rats with diver-gent coping strategy. Physiology & Behavior, 56, 1075–1079.
Received October 9, 2006Revision received February 23, 2007
Accepted February 26, 2007 �
474 POHL ET AL.




















