
Repeated exposure to Δ 9-tetrahydrocannabinol reduces prefrontal cortical dopamine metabolism in...
12
Repeated exposure to D 9 -tetrahydrocannabinol alters heroin-induced locomotor sensitisation and Fos-immunoreactivity M.E. Singh a , I.S. McGregor b , P.E. Mallet a, * a School of Psychology, University of New England, Armidale, NSW 2351, Australia b School of Psychology, Sydney University, Sydney, NSW 2006, Australia Received 16 December 2004; received in revised form 13 July 2005; accepted 14 July 2005 Abstract The present study examined the effect of chronic exposure to D 9 -tetrahydrocannabinol (THC) on heroin-induced locomotor sensitisation and Fos-immunoreactivity (Fos-IR). Adult male albino Wistar rats (n Z 60) were injected intraperitoneally (i.p.) 21 times with vehicle, 0.05, 0.5, or 5.0 mg/kg THC (once every 48 h for 41 days). Locomotor activity was assessed for 180 min on pre-exposure days 1, 21, and 41. Following a 2-week washout period, rats were divided into five equal groups (n Z 12) and injected subcutaneously (s.c.) with vehicle or heroin (0.5 mg/kg). Locomotor activity was recorded for 240 min. In drug-naı¨ve rats, heroin significantly increased locomotor activity. THC pre-exposure further increased heroin-induced locomotion. After an interval of 2 weeks, rats pre-exposed to vehicle and 5.0 mg/kg THC in the first part of the experiment were randomly assigned to one of four treatment groups (n Z 6) and injected s.c. with vehicle or 0.5 mg/kg heroin and perfused 2 h later. Fos-IR was examined in several brain regions. Acute heroin increased Fos-IR in drug-naı¨ve rats in the caudate-putamen (CPu; central, medial and dorsomedial regions), nucleus accumbens (NAC; core and shell regions), bed nucleus of the stria terminalis (BNST), lateral septum, central nucleus of the amygdala (CEA), periaqueductal grey (PAG; dorsolateral, dorsomedial, and lateral), and the EdingereWestphal nucleus. Pre-exposure to THC significantly increased heroin-induced Fos-IR in the dorsomedial CPu and the NAC (core). Conversely, THC pre-exposure reduced heroin-induced Fos-IR in the BNST, CEA, and the PAG (dorsolateral and lateral). The present study demonstrates that THC pre-exposure increases the locomotor stimulating effects of heroin and provides new evidence for the neural correlates that may underlie cannabinoid and opioid cross-sensitisation. Ó 2005 Elsevier Ltd. All rights reserved. Keywords: Cannabinoids; Heroin; Drug addiction; Locomotor activity; Cannabinoid receptor CB1; Reward; Opiate addiction; Rats 1. Introduction Cannabis is among the most widely abused illicit drug in the world with more than a third of the adult population having tried it (Hall and Solowij, 1998). A recent Australian survey suggests that approximately 62% of males and 56% of females in the 20e29-year-old age bracket have used cannabis at least once, while 16% of males and 8.6% of females have used it in the last week (AIWH, 2002). When considering only young adults in the 20e24-year age group, the percentage of cannabis use is almost double (Donnelly and Hall, 1994). Recent debate has focused attention on the possibility that prior cannabis use may enhance the vulnerability of individuals to harder drug of abuse such as heroin. Epidemiological data has provided credence * Corresponding author. Tel.: C61 2 6773 3725; fax: C61 2 6773 3820. E-mail address: [email protected] (P.E. Mallet). 0028-3908/$ - see front matter Ó 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.neuropharm.2005.07.008 Neuropharmacology 49 (2005) 1189e1200 www.elsevier.com/locate/neuropharm
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Repeated exposure to Δ 9-tetrahydrocannabinol reduces prefrontal cortical dopamine metabolism in...

Neuropharmacology 49 (2005) 1189e1200
www.elsevier.com/locate/neuropharm
Repeated exposure to D9-tetrahydrocannabinol altersheroin-induced locomotor sensitisation
and Fos-immunoreactivity
M.E. Singh a, I.S. McGregor b, P.E. Mallet a,*
a School of Psychology, University of New England, Armidale, NSW 2351, Australiab School of Psychology, Sydney University, Sydney, NSW 2006, Australia
Received 16 December 2004; received in revised form 13 July 2005; accepted 14 July 2005
Abstract
The present study examined the effect of chronic exposure to D9-tetrahydrocannabinol (THC) on heroin-induced locomotorsensitisation and Fos-immunoreactivity (Fos-IR). Adult male albino Wistar rats (nZ 60) were injected intraperitoneally (i.p.)21 times with vehicle, 0.05, 0.5, or 5.0 mg/kg THC (once every 48 h for 41 days). Locomotor activity was assessed for 180 min on
pre-exposure days 1, 21, and 41. Following a 2-week washout period, rats were divided into five equal groups (nZ 12) and injectedsubcutaneously (s.c.) with vehicle or heroin (0.5 mg/kg). Locomotor activity was recorded for 240 min. In drug-naı̈ve rats, heroinsignificantly increased locomotor activity. THC pre-exposure further increased heroin-induced locomotion. After an interval of
2 weeks, rats pre-exposed to vehicle and 5.0 mg/kg THC in the first part of the experiment were randomly assigned to one of fourtreatment groups (n Z 6) and injected s.c. with vehicle or 0.5 mg/kg heroin and perfused 2 h later. Fos-IR was examined in severalbrain regions. Acute heroin increased Fos-IR in drug-naı̈ve rats in the caudate-putamen (CPu; central, medial and dorsomedial
regions), nucleus accumbens (NAC; core and shell regions), bed nucleus of the stria terminalis (BNST), lateral septum, centralnucleus of the amygdala (CEA), periaqueductal grey (PAG; dorsolateral, dorsomedial, and lateral), and the EdingereWestphalnucleus. Pre-exposure to THC significantly increased heroin-induced Fos-IR in the dorsomedial CPu and the NAC (core).Conversely, THC pre-exposure reduced heroin-induced Fos-IR in the BNST, CEA, and the PAG (dorsolateral and lateral). The
present study demonstrates that THC pre-exposure increases the locomotor stimulating effects of heroin and provides new evidencefor the neural correlates that may underlie cannabinoid and opioid cross-sensitisation.� 2005 Elsevier Ltd. All rights reserved.
Keywords: Cannabinoids; Heroin; Drug addiction; Locomotor activity; Cannabinoid receptor CB1; Reward; Opiate addiction; Rats
1. Introduction
Cannabis is among the most widely abused illicit drugin the world with more than a third of the adultpopulation having tried it (Hall and Solowij, 1998). Arecent Australian survey suggests that approximately
* Corresponding author. Tel.: C61 2 6773 3725; fax: C61 2 6773
3820.
E-mail address: [email protected] (P.E. Mallet).
0028-3908/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.neuropharm.2005.07.008
62% of males and 56% of females in the 20e29-year-oldage bracket have used cannabis at least once, while 16%of males and 8.6% of females have used it in the lastweek (AIWH, 2002). When considering only youngadults in the 20e24-year age group, the percentage ofcannabis use is almost double (Donnelly and Hall,1994). Recent debate has focused attention on thepossibility that prior cannabis use may enhance thevulnerability of individuals to harder drug of abuse suchas heroin. Epidemiological data has provided credence

1190 M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
to the hypothesis that cannabis may act as a ‘gateway’ toother drugs (Hall and Solowij, 1998; Miller et al., 1990).However, the conclusions drawn from human studiesare correlational in nature and thus cannot establisha causal relationship between cannabis use and sub-sequent transition to other harder drugs. Thus, there isa need for laboratory studies using animal models ofaddiction to provide non-correlational evidence for thepossible gateway effect of cannabis.
Studies using adult rats have provided evidence thatcannabinoid pre-exposure enhances the locomotoractivating effects of drugs such as opiates as measuredby the cross-sensitisation paradigm. Cross-sensitisationis an interesting phenomenon whereby pre-exposure toone drug of abuse increases sensitivity to the activatingor rewarding properties of the other (Cunningham et al.,1997; Lett, 1989). Using this model, cross-sensitisationbetween opioids and D9-tetrahydrocannabinol (THC)dthe main psychoactive constituent in cannabis d andother synthetic cannabinoid receptor agonists such asWIN 55, 212-2 and CP 55,940 have been demonstrated.For example, pre-exposure to THC or WIN 55, 212-2enhances the locomotor activity stimulating effects ofheroin (Lamarque et al., 2001; Pontieri et al., 1997,2001). Furthermore, our group has recently shownenhanced morphine-induced locomotor activity andself-administration in rats pre-exposed to CP 55,940(Norwood et al., 2003). In addition to locomotoractivity, a recent study has shown that pre-exposure toTHC cross-sensitises to other behavioural effects ofopiates such as sniffing and gnawing (Cadoni et al.,2001). These behavioural studies suggest that chroniccannabinoid exposure may lead to long-lasting neuralchanges in brain regions involved in opiate addiction.However, the neural mechanisms involved in mediatingcross-sensitisation between cannabinoids and opioidsare not well understood. Hence, the primary purpose ofthe present study was to examine whether cross-sensitisation between cannabinoid and opioids is detect-able at the level of gene expression.
Fos-IR as a dependent measure represents a usefultest of alterations in brain function following drugadministration (Arnold et al., 1998). For example, long-term exposure to morphine alters the Fos responses tococaine and MDMA (ecstasy) (Erdtmann-Vourliotiset al., 2000). Given that an interaction exists between thecannabinoid and opioid systems (for reviews seeNavarro et al., 2001; Tanda and Goldberg, 2003), itcould be suggested that pre-exposure to THC maylead to permanent changes in opioid-induced geneexpression.
In order to validate the experimental protocol and thedoses used, we began the present study by examiningwhether chronic THC exposure cross-sensitises to thelocomotor activating effect of heroin using a wide rangeof THC doses. Consistent with previous studies, it was
predicted that heroin-induced locomotor activity wouldbe significantly enhanced in THC pre-exposed comparedto drug-naı̈ve rats. Following the behavioural testing,we examined the effect of chronic THC exposure onheroin-induced Fos-IR. Examination of alterations inheroin-induced Fos-IR in THC pre-exposed rats wasexpected to provide new insights into the mechanismsunderlying the functional interactions between theseneurotransmitter systems.
2. Methods
2.1. Subjects
Experimentally naı̈ve male albino Wistar rats ap-proximately 8 weeks of age at the start of the experimentwere housed in opaque plastic cages 4e6 per cage ona reverse 12 : 12 h light/dark cycle (lights off at 08:00 h)with free access to standard laboratory chow (Barastoc,Ridley AgriProducts, Australia) and tap water. Exper-imental testing was conducted during the dark cyclebetween 08:30 and 16:00 h. Animals were treated inaccordance with the National Institutes of Health Guidefor the Care and Use of Laboratory Animals (NIHPublications No. 8023, revised 1978) and the AustralianCode of Practice for the Care and Use of Animals forScientific Purposes. This study was approved by theUniversity of New England Animal Ethics Committee.
2.2. Drugs
D9-Tetrahydrocannabinol (THC; AGAL, Pymble,NSW, Australia), available as a 2.0 mg THC/ml ethanolsolution, was first mixed with a few drops of TWEEN-80(polyoxyethylene sorbitan monooleate, ICN Biomedi-cals, Seven Hills, NSW, Australia). The suspension wasstirred continuously under a stream of nitrogen gas untilall ethanol was evaporated. Physiological saline was thenadded and the solution was stirred until the TWEEN-80/THC suspension was well dissolved. The final vehiclesolution contained 15 ml TWEEN-80 per 2 ml saline.THC was administered intraperitoneally (i.p.) at dosesof 0.05, 0.5, and 5.0 mg/kg in a volume of 1 ml/kg.
Heroin (API/AMED, Australia) was dissolved ina vehicle containing 0.05 ml acetic acid (Sigma-Aldrich,Australia) in 2 ml of physiological saline (0.9%). Heroinwas administered subcutaneously (s.c.) at a dose of0.5 mg/kg in a volume of 1 ml/kg.
2.3. Apparatus
Eight identical dimly lit (13 lux) rectangular Perspexlocomotor activity chambers (280 mm ! 230 mm !300 mm high) were used. The walls were constructedof clear Perspex and aluminium, and the floors were

1191M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
made of galvanised wire mesh (1 cm2). A passiveinfrared detector (Quantum passive infrared motionsensors, NESS Security Products, Australia, part no.890-087-2) mounted to the ceiling of each box was usedto quantify locomotor activity using custom designedsoftware. A 10 mF capacitor located near LK2 of thesensor’s printed circuit board was replaced with a 0.1 mFcapacitor serving to alter the sensor alarm period from5 s to approximately 50 ms. Activity chambers werelocated inside sound attenuating boxes (800 mm !360 mm ! 650 mm high) and fitted with a fan thatprovided ventilation and masking noise.
2.4. Procedure
2.4.1. Effects of THC pre-exposure on behaviouralsensitisation to heroin
Rats (n Z 60) approximately 8 weeks old werehandled for 1 min per day for 6 consecutive days beforethe start of the experiment. Next, rats received 21injections (i.p.) of vehicle (VEH; TWEEN-80 in saline;n Z 24), 0.05 mg/kg THC (0.05THC; n Z 12), 0.5 mg/kg THC (0.5THC; n Z 12), or 5.0 mg/kg THC (5.0THC; n Z 12) every 48 h for a total of 41 days (Fig. 1).The effect of THC on locomotor activity was measuredat the beginning (day 1), in the middle (day 21), and atthe end (day 41) of chronic THC treatment. During thistime rats were injected with vehicle or THC and placedin a locomotor activity chamber for 180 min.
Following a 2-week washout period (day 56), all ratswere injected with the vehicle for heroin (acetic acid insaline, s.c.) and placed in the apparatus for 240 min inorder to habituate the rats to the apparatus and todetermine the baseline rate of locomotor activity. A testlength of 240 min was chosen as our previous studyusing the opioid receptor agonist morphine foundmaximal stimulation of locomotor activity at thisinterval (Norwood et al., 2003). Rats were then dividedinto five equal groups (n Z 12 per group) as follows:vehicle/vehicle (VEH/VEH), vehicle/0.5 mg/kgheroin (VEH/HER), 0.05 mg/kg THC/0.5 mg/kgheroin (0.05THC/HER), 0.5 mg/kg THC/0.5 mg/kg heroin (0.5THC/HER), 5.0 mg/kg THC/0.5 mg/
kg heroin (5.0THC/HER). Rats were injected 48 hlater (day 58) with vehicle or heroin (0.5 mg/kg, s.c.) andplaced in the apparatus for 240 min. A dose of 0.5 mg/kg heroin was chosen because our recent work revealedthat this dose induced hyperactivity, whereas a higherdose (i.e., 2.0 mg/kg) suppressed locomotor activity inrats (Singh et al., in press).
2.4.2. Effects of THC pre-exposure on heroin-inducedFos-IR
After an interval of 2 weeks (day 72), rats in theVEH/HER and 5.0THC/HER groups above wereused in a second experiment examining Fos-IR. Theserats were reassigned to one of four treatment groups(n Z 6) according to a standard 2 ! 2 design. Rats inthe VEH/HER group above were divided into twoseparate groups: VEH/VEH and VEH/HER, whilethe rats in the 5.0THC/HER group above weredivided into two separate groups: THC/VEH andTHC/HER.
Next, all rats were habituated to the testing apparatusand injection procedure for 4 consecutive habituationdays in order to minimise novelty-induced Fos-IR. Oneach of the habituation days, rats were injected s.c. withvehicle. Rats were then placed into the locomotoractivity boxes for 120 min. Treatments were staggeredat 10-min intervals to allow sufficient time betweenconsecutive perfusions. Each home cage contained atleast one rat from each treatment group. The protocolfor the test day was similar to the habituation days withthe exception of drug treatment and perfusion. On thetest day, rats were injected with either vehicle or 0.5 mg/kg heroin (s.c.) and placed into the locomotor activityboxes for 120 min and were then perfused.
Rats were deeply anaesthetised with 120 mg/kgsodium pentobarbital (i.p.) and perfused transcardiallywith 100 ml phosphate-buffered saline (PBS, 0.1 M, pH7.2) followed by 150 ml 4% paraformaldehyde in PBS.Brains were then extracted and placed in 4% para-formaldehyde at 4 �C for 24 h, transferred to 15%sucrose in a phosphate buffer solution (PB, 0.1 M, pH7.2) at 4 �C for 24 h, and finally placed in 30% sucrosesolution in PB at 4 �C for 48 h. Whole brains were then
1 21 41
2-week washout
56 58 72 76
Heroin or Vehthen perfusion
Day
THC or VEH every 48 h
Veh(Hab)
Hab for c-fosstudy begins
2-week washout
Heroinprobe
Locomotionmeasured
Locomotionmeasured
Locomotionmeasured
Locomotionmeasured
Locomotionmeasured
Fig. 1. Schematic overview of the experimental design. THC, D9-tetrahydrocannabinol; Veh, vehicle; Hab, habituation.

1192 M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
sectioned coronally at 40 mm using a cryostat andsections were collected in PB.
Free-floating sections were given two consecutive 30-min washes in PB, a 30-min wash in 0.9% hydrogenperoxide in 50% ethanol, and a 30-min wash in 3%normal horse serum in PB. Sections were then incubatedfor 72 h at 4 �C in the primary c-fos antibody (s52,Santa Cruz Biotechnology, Santa Cruz, CA; rabbitpolyclonal, specific for the amino acid terminus of c-Fosp62, non cross-reactive with FosB, Fra-1 or Fra-2)diluted in 1:2000 in phosphate-buffered horse serum(PBH, 0.1% bovine serum albumin, 0.2% Triton X-100,2% normal horse serum in PB). Next, sections werewashed for 30 min in PB and incubated for 60 min inbiotinylated anti-rabbit IgG (Vector Laboratories,Burlingame, CA) diluted 1:500 in PBH. They were thenwashed in PB for 30 min, and subsequently incubatedfor 60 min in ExtrAvidinehorseradish peroxidase (SigmaeAldrich, Castle Hill, NSW) diluted 1:1000 in PBH. Afterthree further 30-min washes in PB, horseradish perox-idase activity was visualised with the nickel diamino-benzidine and glucose oxidase reaction with nickelenhancement as described by Shu et al. (1988). Thereaction was terminated 10 min later by washing in PB.Sections were mounted onto gelatine-coated slides,dehydrated, Histolene-cleared, and coverslipped.
A total of 15 brain regions or sub-regions wereexamined using an Olympus CH-2 light microscope at200! magnification. The atlas of Paxinos and Watson(1998) was used to identify brain regions, shown inFig. 2. A 10 ! 10 square graticule was positioned overeach structure and the number of labelled cells withinthe graticule, which covered a 500 ! 500 mm area, wascounted manually. Where the structure was larger thanthe graticule, a standardised region within the structurewas counted. Only round and oval nuclei that werecompletely black were counted. Because manual count-ing can be subject to experimenter bias, microscopeslides were labelled such that the person counting thelabelled cells did not know which group the ratsbelonged to.
Images of Fos-IR cells were captured using a NikonDigital Sight camera system attached to a Nikon Eclipse55i microscope. Post-production image processing andlayout was conducted using Deneba Canvas 9.0, andincluded reduction of colour to greyscale, and stand-ardisation of brightness and contrast.
2.5. Statistical analysis
2.5.1. Effects of THC pre-exposure on behaviouralsensitisation to heroin
Locomotor activity occurring during each of the 601-min measurement intervals was summed to producea single activity score for each 60-min interval. THC-induced locomotor activity data on days 1, 21, and 41
were grouped into three independent 60-min bins (at 60,120, and 180 min) and analysed using separate group bytime mixed-design ANOVAs with repeated measures onthe second factor. Where significant main effects werefound, groups were compared using planned unpairedt-tests with Bonferroni adjustments for multiple com-parisons. The three contrasts of interest were: (1)VEH/VEH vs. 0.05THC/HER, (2) VEH/VEHvs. 0.5THC/HER and (3) VEH/VEH vs.5.0THC/HER. Where a significant interaction wasfound, a separate one-way ANOVA was used tocompare groups at each measurement interval.
Locomotor activity during the habituation phase(day 56) and the heroin probe phase (day 58) weregrouped into eight independent 30-min bins (at 30, 60,90, 120, 150, 180, 210, and 240 min) and analysed usinga group by time mixed-design ANOVA with repeatedmeasures on the second factor. Where significant maineffects were found, groups were compared using plannedunpaired t-tests with Bonferroni adjustments for multi-ple comparisons. The four contrasts of interest were:(1) VEH/VEH vs. VEH/HER, (2) VEH/HER vs.0.05THC/HER, (3) VEH/HER vs. 0.5THC/HER,and (4) VEH/HER vs. 5.0THC/HER. Wherea significant interaction was found, a separate one-wayANOVA was used to compare groups at each timeinterval.
2.5.2. Effects of THC pre-exposure on heroin-inducedFos-IR
Locomotor activity data from the perfusion day weregrouped into four 30-min bins and were analysed usinga mixed-design (dose by time) ANOVA with repeatedmeasures on the second factor. Group differences inlocomotor activity were compared by conducting a one-way ANOVA at each 30-min interval. Groups were thencompared using post hoc Tukey tests where significantmain effects were found.
The number of labelled cells across groups wascompared using one-way ANOVAs. Planned contrastswere used to compare groups where significant ANOVAmain effects were observed. The five contrasts of interest(a Z 0.05) were: (1) VEH/VEH vs. THC/VEH,(2) VEH/VEH vs. VEH/HER, (3) VEH/VEH vs.THC/HER, (4) THC/HER vs. THC/VEH, and(5) THC/HER vs. VEH/HER. A two-wayANOVA was also conducted for each brain region todetermine whether the combined effect of the drugs wassignificantly higher or lower than would be expectedby the sum of the individual drug effects. The first factorwas the presence or absence of THC and the second wasthe presence or absence of heroin. Because the in-dividual main effects provided no additional relevantinformation only the interaction term was reported.
Where the ANOVA assumptions were violated,randomisation tests of scores were conducted using
1193M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
12
4
5
7
+1.00
8
6
3
-0.26
9
10
-2.80
-5.30
14
-6.72
12 1113
-5.60
15
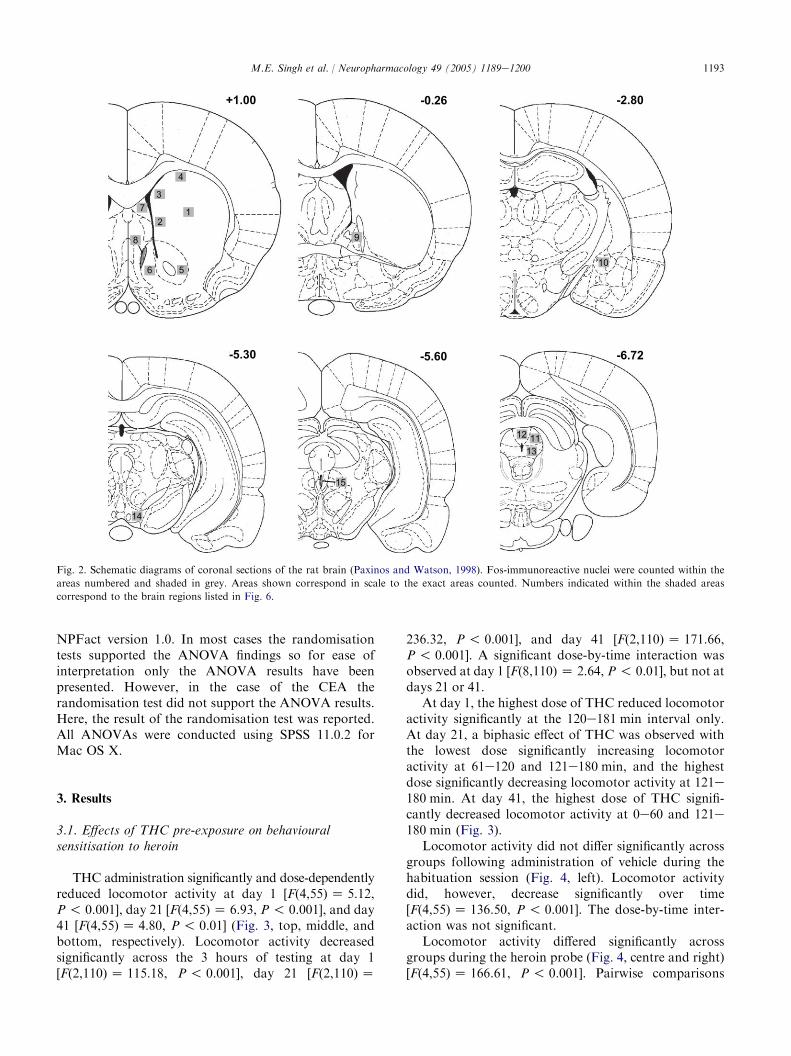
Fig. 2. Schematic diagrams of coronal sections of the rat brain (Paxinos and Watson, 1998). Fos-immunoreactive nuclei were counted within the
areas numbered and shaded in grey. Areas shown correspond in scale to the exact areas counted. Numbers indicated within the shaded areas
correspond to the brain regions listed in Fig. 6.
NPFact version 1.0. In most cases the randomisationtests supported the ANOVA findings so for ease ofinterpretation only the ANOVA results have beenpresented. However, in the case of the CEA therandomisation test did not support the ANOVA results.Here, the result of the randomisation test was reported.All ANOVAs were conducted using SPSS 11.0.2 forMac OS X.
3. Results
3.1. Effects of THC pre-exposure on behaviouralsensitisation to heroin
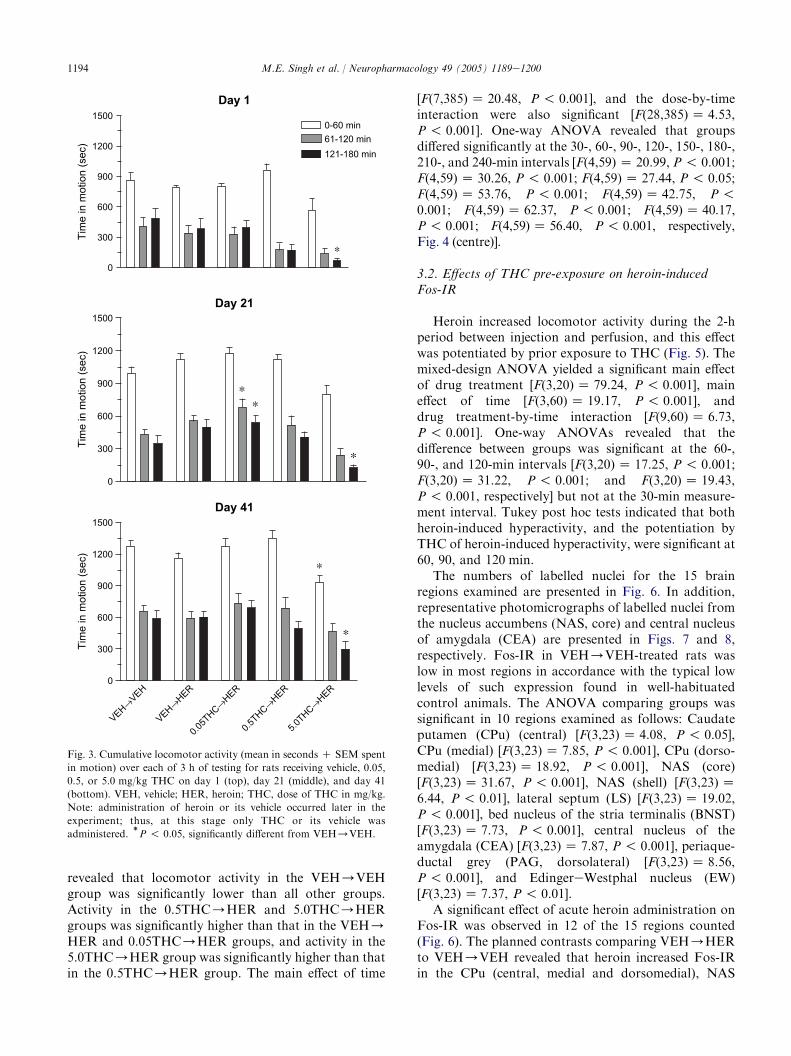
THC administration significantly and dose-dependentlyreduced locomotor activity at day 1 [F(4,55) Z 5.12,P ! 0.001], day 21 [F(4,55)Z 6.93, P ! 0.001], and day41 [F(4,55)Z 4.80, P ! 0.01] (Fig. 3, top, middle, andbottom, respectively). Locomotor activity decreasedsignificantly across the 3 hours of testing at day 1[F(2,110)Z 115.18, P ! 0.001], day 21 [F(2,110)Z
236.32, P ! 0.001], and day 41 [F(2,110) Z 171.66,P ! 0.001]. A significant dose-by-time interaction wasobserved at day 1 [F(8,110) Z 2.64, P ! 0.01], but not atdays 21 or 41.
At day 1, the highest dose of THC reduced locomotoractivity significantly at the 120e181 min interval only.At day 21, a biphasic effect of THC was observed withthe lowest dose significantly increasing locomotoractivity at 61e120 and 121e180 min, and the highestdose significantly decreasing locomotor activity at 121e180 min. At day 41, the highest dose of THC signifi-cantly decreased locomotor activity at 0e60 and 121e180 min (Fig. 3).
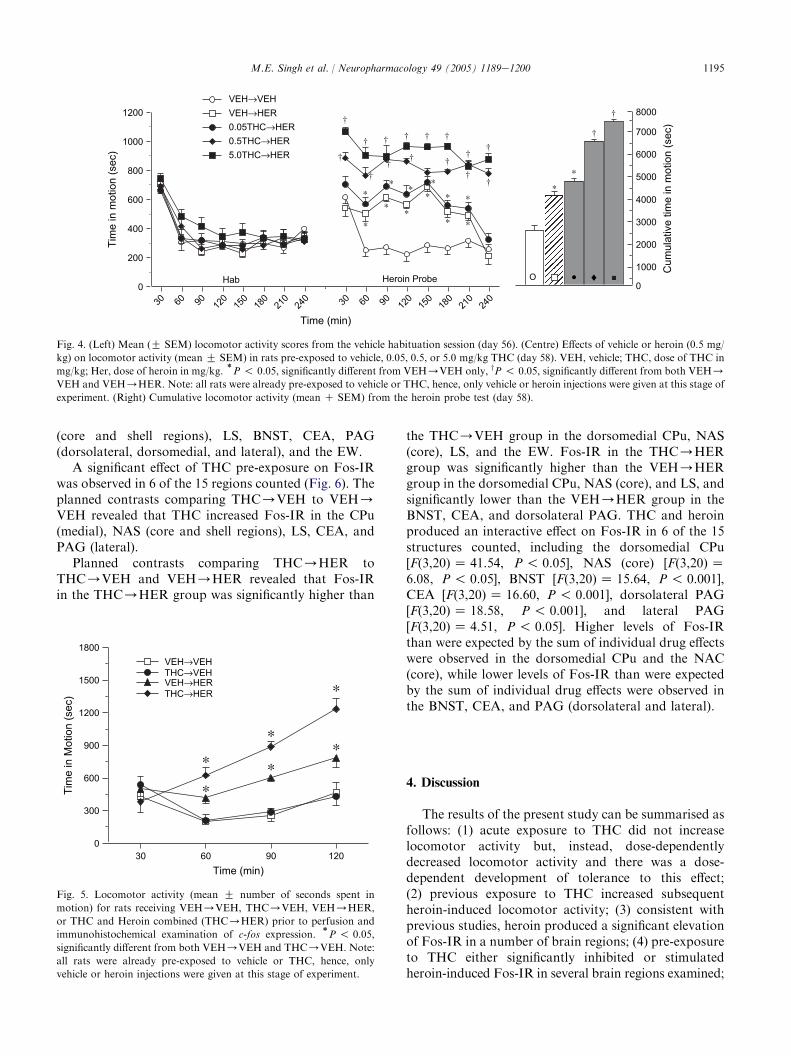
Locomotor activity did not differ significantly acrossgroups following administration of vehicle during thehabituation session (Fig. 4, left). Locomotor activitydid, however, decrease significantly over time[F(4,55)Z 136.50, P ! 0.001]. The dose-by-time inter-action was not significant.
Locomotor activity differed significantly acrossgroups during the heroin probe (Fig. 4, centre and right)[F(4,55)Z 166.61, P ! 0.001]. Pairwise comparisons
1194 M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
revealed that locomotor activity in the VEH/VEHgroup was significantly lower than all other groups.Activity in the 0.5THC/HER and 5.0THC/HERgroups was significantly higher than that in the VEH/HER and 0.05THC/HER groups, and activity in the5.0THC/HER group was significantly higher than thatin the 0.5THC/HER group. The main effect of time
0
300
600
900
1200
15000-60 min61-120 min121-180 min
*
0
300
600
900
1200
1500
**
*
VEH→VEH
VEH→HER
0.05T
HC→HER
0.5THC→
HER
5.0THC→
HER0
300
600
900
1200
1500
*
*
Tim
e in
mot
ion
(sec
)Ti
me
in m
otio
n (s
ec)
Tim
e in
mot
ion
(sec
)
Day 1
Day 21
Day 41
Fig. 3. Cumulative locomotor activity (mean in seconds C SEM spent
in motion) over each of 3 h of testing for rats receiving vehicle, 0.05,
0.5, or 5.0 mg/kg THC on day 1 (top), day 21 (middle), and day 41
(bottom). VEH, vehicle; HER, heroin; THC, dose of THC in mg/kg.
Note: administration of heroin or its vehicle occurred later in the
experiment; thus, at this stage only THC or its vehicle was
administered.)P ! 0.05, significantly different from VEH/VEH.
[F(7,385) Z 20.48, P ! 0.001], and the dose-by-timeinteraction were also significant [F(28,385)Z 4.53,P ! 0.001]. One-way ANOVA revealed that groupsdiffered significantly at the 30-, 60-, 90-, 120-, 150-, 180-,210-, and 240-min intervals [F(4,59) Z 20.99, P ! 0.001;F(4,59) Z 30.26, P ! 0.001; F(4,59)Z 27.44, P ! 0.05;F(4,59) Z 53.76, P ! 0.001; F(4,59)Z 42.75, P !0.001; F(4,59) Z 62.37, P! 0.001; F(4,59)Z 40.17,P! 0.001; F(4,59)Z 56.40, P! 0.001, respectively,Fig. 4 (centre)].
3.2. Effects of THC pre-exposure on heroin-inducedFos-IR
Heroin increased locomotor activity during the 2-hperiod between injection and perfusion, and this effectwas potentiated by prior exposure to THC (Fig. 5). Themixed-design ANOVA yielded a significant main effectof drug treatment [F(3,20)Z 79.24, P ! 0.001], maineffect of time [F(3,60) Z 19.17, P ! 0.001], anddrug treatment-by-time interaction [F(9,60)Z 6.73,P ! 0.001]. One-way ANOVAs revealed that thedifference between groups was significant at the 60-,90-, and 120-min intervals [F(3,20)Z 17.25, P ! 0.001;F(3,20) Z 31.22, P ! 0.001; and F(3,20)Z 19.43,P ! 0.001, respectively] but not at the 30-min measure-ment interval. Tukey post hoc tests indicated that bothheroin-induced hyperactivity, and the potentiation byTHC of heroin-induced hyperactivity, were significant at60, 90, and 120 min.
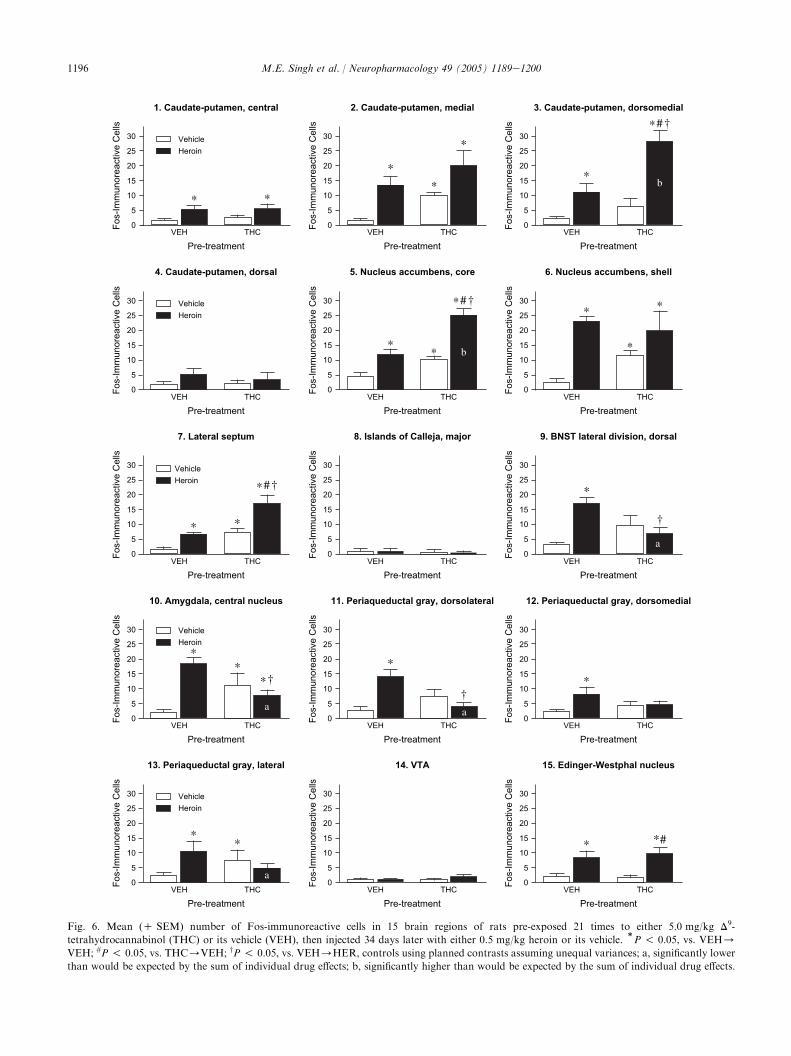
The numbers of labelled nuclei for the 15 brainregions examined are presented in Fig. 6. In addition,representative photomicrographs of labelled nuclei fromthe nucleus accumbens (NAS, core) and central nucleusof amygdala (CEA) are presented in Figs. 7 and 8,respectively. Fos-IR in VEH/VEH-treated rats waslow in most regions in accordance with the typical lowlevels of such expression found in well-habituatedcontrol animals. The ANOVA comparing groups wassignificant in 10 regions examined as follows: Caudateputamen (CPu) (central) [F(3,23) Z 4.08, P ! 0.05],CPu (medial) [F(3,23)Z 7.85, P ! 0.001], CPu (dorso-medial) [F(3,23) Z 18.92, P ! 0.001], NAS (core)[F(3,23) Z 31.67, P ! 0.001], NAS (shell) [F(3,23) Z6.44, P ! 0.01], lateral septum (LS) [F(3,23)Z 19.02,P ! 0.001], bed nucleus of the stria terminalis (BNST)[F(3,23) Z 7.73, P ! 0.001], central nucleus of theamygdala (CEA) [F(3,23)Z 7.87, P ! 0.001], periaque-ductal grey (PAG, dorsolateral) [F(3,23)Z 8.56,P ! 0.001], and EdingereWestphal nucleus (EW)[F(3,23) Z 7.37, P ! 0.01].
A significant effect of acute heroin administration onFos-IR was observed in 12 of the 15 regions counted(Fig. 6). The planned contrasts comparing VEH/HERto VEH/VEH revealed that heroin increased Fos-IRin the CPu (central, medial and dorsomedial), NAS
1195M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
0
1000
2000
3000
4000
5000
6000
7000
8000
Cum
ulat
ive
time
in m
otio
n (s
ec)
30 60 90 120
150
180
210
240 30 60 90 12
015
018
021
024
00
200
400
600
800
1000
1200Ti
me
in m
otio
n (s
ec)
Time (min)
VEH→HER0.05THC→HER0.5THC→HER5.0THC→HER
Hab Heroin Probe
VEH→VEH
*
*
*
†
†
† † † †
†
†
††
††
†††
†
†
*
*
*
*
* *
*
*
*
♦•O
*
*
Fig. 4. (Left) Mean (G SEM) locomotor activity scores from the vehicle habituation session (day 56). (Centre) Effects of vehicle or heroin (0.5 mg/
kg) on locomotor activity (mean G SEM) in rats pre-exposed to vehicle, 0.05, 0.5, or 5.0 mg/kg THC (day 58). VEH, vehicle; THC, dose of THC in
mg/kg; Her, dose of heroin in mg/kg.)P ! 0.05, significantly different from VEH/VEH only, yP ! 0.05, significantly different from both VEH/
VEH and VEH/HER. Note: all rats were already pre-exposed to vehicle or THC, hence, only vehicle or heroin injections were given at this stage of
experiment. (Right) Cumulative locomotor activity (mean C SEM) from the heroin probe test (day 58).
(core and shell regions), LS, BNST, CEA, PAG(dorsolateral, dorsomedial, and lateral), and the EW.
A significant effect of THC pre-exposure on Fos-IRwas observed in 6 of the 15 regions counted (Fig. 6). Theplanned contrasts comparing THC/VEH to VEH/VEH revealed that THC increased Fos-IR in the CPu(medial), NAS (core and shell regions), LS, CEA, andPAG (lateral).
Planned contrasts comparing THC/HER toTHC/VEH and VEH/HER revealed that Fos-IRin the THC/HER group was significantly higher than
30 60 90 1200
300
600
900
1200
1500
1800
Tim
e in
Mot
ion
(sec
)
Time (min)
*
VEH→VEHTHC→VEH
*
*
*
*
*
VEH→HERTHC→HER
Fig. 5. Locomotor activity (mean G number of seconds spent in
motion) for rats receiving VEH/VEH, THC/VEH, VEH/HER,
or THC and Heroin combined (THC/HER) prior to perfusion and
immunohistochemical examination of c-fos expression.)P ! 0.05,
significantly different from both VEH/VEH and THC/VEH. Note:
all rats were already pre-exposed to vehicle or THC, hence, only
vehicle or heroin injections were given at this stage of experiment.
the THC/VEH group in the dorsomedial CPu, NAS(core), LS, and the EW. Fos-IR in the THC/HERgroup was significantly higher than the VEH/HERgroup in the dorsomedial CPu, NAS (core), and LS, andsignificantly lower than the VEH/HER group in theBNST, CEA, and dorsolateral PAG. THC and heroinproduced an interactive effect on Fos-IR in 6 of the 15structures counted, including the dorsomedial CPu[F(3,20)Z 41.54, P ! 0.05], NAS (core) [F(3,20) Z6.08, P ! 0.05], BNST [F(3,20)Z 15.64, P ! 0.001],CEA [F(3,20)Z 16.60, P ! 0.001], dorsolateral PAG[F(3,20)Z 18.58, P ! 0.001], and lateral PAG[F(3,20)Z 4.51, P ! 0.05]. Higher levels of Fos-IRthan were expected by the sum of individual drug effectswere observed in the dorsomedial CPu and the NAC(core), while lower levels of Fos-IR than were expectedby the sum of individual drug effects were observed inthe BNST, CEA, and PAG (dorsolateral and lateral).
4. Discussion
The results of the present study can be summarised asfollows: (1) acute exposure to THC did not increaselocomotor activity but, instead, dose-dependentlydecreased locomotor activity and there was a dose-dependent development of tolerance to this effect;(2) previous exposure to THC increased subsequentheroin-induced locomotor activity; (3) consistent withprevious studies, heroin produced a significant elevationof Fos-IR in a number of brain regions; (4) pre-exposureto THC either significantly inhibited or stimulatedheroin-induced Fos-IR in several brain regions examined;
1196 M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
1. Caudate-putamen, central 2. Caudate-putamen, medial 3. Caudate-putamen, dorsomedial
VEH THC0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
VehicleHeroin
VehicleHeroin
VehicleHeroin
VehicleHeroin
VehicleHeroin
**
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
4. Caudate-putamen, dorsal 5. Nucleus accumbens, core 6. Nucleus accumbens, shell
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
*
*
*
*
#†*
#†*
#†*
††
†
*
b
** b
7. Lateral septum 8. Islands of Calleja, major 9. BNST lateral division, dorsal
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
*
*
*
* *
10. Amygdala, central nucleus 11. Periaqueductal gray, dorsolateral 12. Periaqueductal gray, dorsomedial
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
13. Periaqueductal gray, lateral 14. VTA 15. Edinger-Westphal nucleus
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
VEH THC
Pre-treatment
Fos-
Imm
unor
eact
ive
Cel
ls
*
a
**
a
*
a
*
**
a
*#*
Fig. 6. Mean (C SEM) number of Fos-immunoreactive cells in 15 brain regions of rats pre-exposed 21 times to either 5.0 mg/kg D9-
tetrahydrocannabinol (THC) or its vehicle (VEH), then injected 34 days later with either 0.5 mg/kg heroin or its vehicle.)P ! 0.05, vs. VEH/
VEH; #P ! 0.05, vs. THC/VEH; yP ! 0.05, vs. VEH/HER, controls using planned contrasts assuming unequal variances; a, significantly lower
than would be expected by the sum of individual drug effects; b, significantly higher than would be expected by the sum of individual drug effects.

1197M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
Fig. 7. Photomicrograph showing Fos-immunoreactive neurons in the nucleus accumbens (core) in representative sections from rats treated with
VEH/VEH (A), THC/VEH (B), VEH/HER (C), or THC and heroin combined (THC/HER) (D). Scale bar Z 250 mm. The square indicates
the region in which labelled cells were quantified.
(5) interestingly, THC pre-exposure increased basal Fos-IR in several regions.
One finding of this study was that THC pre-exposuredose-dependently increased locomotor activity in re-sponse to a single injection of heroin. The presentfinding supports a previous study showing that exposureto THC enhances locomotor cross-sensitivity to heroin(Lamarque et al., 2001). Furthermore, the result of thisstudy complements a recent finding from our groupshowing cross-sensitisation between the cannabinoidreceptor agonist CP 55,940 and morphine (Norwoodet al., 2003). The results of this study provide furtherevidence that pre-exposure to cannabinoids cross-sensitises to the locomotor activating effects of opiates.
It is important to note that the results obtained heredo not suggest that long-term cannabinoid exposurenecessarily enhances the rewarding properties ofopioids. Although a recent study by our group hasreported increased morphine self-administration behav-iour in rats exposed to CP 55,940 (Norwood et al.,2003), the increase in lever pressing observed in CP55,940 pre-exposed rats was not specific to the activelever; that is, inactive lever presses were also increased,suggesting a general behavioural activation to morphinein rats that were pre-exposed to CP 55,940 (Norwoodet al., 2003). More recently, Solinas et al. (2004)reported that THC exposure in adult rats increases the
frequency of self-administered heroin injections withoutincreasing the reinforcing efficacy of heroin. Specifically,the maximum ratio at which rats would continue torespond to heroin injections did not differ between ratspre-exposed to THC and rats pre-exposed to vehicle.However, under continuous reinforcement conditions(fixed-ratio one), rats pre-exposed to THC showedhigher levels of heroin intake with shorter post-injectionpauses. Taken together, these studies suggest that long-term cannabinoid exposure may have produced changesin the pharmacological profile of heroin withoutnecessarily altering its rewarding properties.
The primary aim of this study was to examine theneural correlates underlying cannabinoideopioid cross-sensitisation. The results revealed that induction of Fos-IR differed according to the rat’s drug history. There areseveral findings worth noting in this regard. First, heroinadministration to drug-naı̈ve rats induced significantFos-IR in the CPu (central, medial and dorsomedial),NAS (core and shell regions), LS, BNST, CEA, PAG,and EW. The increase in heroin-induced Fos-IRobserved in the NAC (shell) and CPu are in goodagreement with recent experiments (Curran et al., 1996;D’Este et al., 2002). To the best of our knowledge, theincreases in Fos-IR following acute heroin administra-tion observed in the NAC (core), BNST, CEA, PAG,and EW are novel findings. It should be noted, however,

1198 M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
Fig. 8. Photomicrograph showing Fos-immunoreactive neurons in the central nucleus of the amygdala (CEA) in representative sections from rats
treated with VEH/VEH (A), THC/VEH (B), VEH/HER (C), or THC and heroin combined (THC/HER) (D). Scale bar Z 250 mm. The
square indicates the region in which labelled cells were quantified.
that previous studies using the m-opioid receptor agonistmorphine have found similar results to those obtained inthe present study using heroin (Grabus et al., 2004;Warty et al., 2002).
Second, acute heroin administration to rats withTHC exposure increased Fos-IR in the dorsomedial partof the CPu, while a non-interactive additive effect wasobserved in the LS. The CPu and the LS play animportant role in motor integration, and concurrentlesions of the dorsomedial CPu and the LS reducemorphine-induced hyperlocomotion (Stevens et al.,1986). Furthermore, a significant increase in Fos-IRwas also observed in the NAC (core). Anatomical datasuggest that the dorsomedial CPu receives strong inputsfrom the NAC (core) (Brown et al., 1998) and theseregions are involved in mediating stereotypic activity inresponse to sensitisation to drugs of abuse (Cadoniet al., 2001; D’Este et al., 2002). These regions alsocontain DA terminal fields and denervation of the DAterminal fields reduces locomotion (Fink and Smith,1980). Hence, this novel finding in the dorsomedial CPu,LS, and the NAC (core) could represent a marker forthe intense heroin-induced locomotor hyperactivityobserved during the behavioural phase and just priorto perfusion in rats pre-exposed to THC (Figs. 4 and 5).
Third, the present study showed for the first time thatTHC pre-exposure significantly reduces acute heroin-
induced Fos-IR in the BNST, CEA, and the PAG(dorsolateral and lateral). The BNST and CEA areanatomically, neurochemically, and cytoarchitecturallyrelated (Alheid et al., 1995). Studies suggest that thesestructures play an important role in mediating thereinforcing effects of drugs of abuse (Caine et al., 1995;Di Chiara et al., 1999; Olmstead and Franklin, 1997a,b). The BNST and CEA are richly innervated bydopaminergic neurons (Freedman and Cassell, 1994;Shammah-Lagnado et al., 2001) and stimulating theseregions releases dopamine (DA) (Freedman and Cassell,1994; Shammah-Lagnado et al., 2001). THC exposuremay therefore inhibit activation of DA neurons in theBNST and CEA leading to an increase in the sensitisingproperties of addictive substances as a means tocompensate for this deficit. Indeed, chronic THCexposure has been shown to alter DA sensitivity (Wuand French, 2000). This hypothesis agrees well with thecross-sensitivity observed in THC pre-exposed ratsfollowing heroin administration (Fig. 4).
Evidence suggests a role for the PAG in mediatinglocomotor sensitisation and place preference. Forexample, morphine administered into the PAG producesconditioned place preference (Olmstead and Franklin,1997b), while lesions of the PAG reduces morphine-induced place preference (Olmstead and Franklin,1997a). Furthermore, cocaine-induced locomotor

1199M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
sensitisation is believed to be mediated via the PAG(Miczek et al., 1999). As both cannabinoid (Herkenhamet al., 1991) and opioid (Vaughan et al., 2003) receptorsare localised in the PAG, this region may be a novelneural substrate mediating the cannabinoid-opioidcross-sensitivity observed in Fig. 4.
A remarkable and surprising finding was the wide-spread stimulation of basal Fos-IR in rats followingTHC exposure. This increase was observed in the CPu(medial), NAC (core and shell regions), LS, CEA, andPAG (lateral). It is important to note that there wasa long delay lasting several weeks between drugexposure and perfusion. It is not clear at present whyFos-IR was observed so long after cessation of drugexposure; however, these results provide evidence thatbrain function continues to be altered weeks aftercannabinoid exposure has ceased. This long-lastingchange in basal Fos-IR by THC should, however, beconsidered with caution as it may simply reflecta conditioned drug effect. That is, the increase in basalFos-IR by THC may be the result of the development ofa conditioned response to one or more drug-related cues(e.g. handling, injection, weighing). The administrationof vehicle to these rats after a long period of abstinencefrom THC may therefore have induced a motivationalstate previously associated with THC administration.Animals were perfused 2 h following vehicle adminis-tration, and after being placed in the locomotor activitychambers. Therefore, Fos production associated witha conditioned state would have had sufficient time todevelop. In support of this hypothesis, a relatedexperiment recently conducted in our laboratory (sub-mitted) also examined the residual effects of chroniccannabinoid exposure on basal Fos-IR, but because wewere not interested in the acute effects of a second drugon Fos-IR, locomotor activity was not measured, andanimals were not injected with vehicle prior toperfusion. Under these conditions we did not observeany residual cannabinoid-induced changes in basal Fos-IR. It also follows that the alteration in heroin-inducedFos-IR by prior THC exposure may also be attribut-able, at least in part, to a conditioned drug effect.
It is noteworthy that rats had been exposed to a singleinjection of heroin 2 weeks prior to the beginning of theFos study (i.e. for the cross-sensitisation study), and thismay have subsequently influenced either the locomotordata, the number of labelled cells, or both in the Fosstudy. However, we do not believe this to be the case. Aprevious study by our group examining the effect ofa single heroin injection on Fos-IR in rats perinatallyexposed to THC showed similar Fos expression to thatobserved in the current study (Singh et al., in press).
In summary, the present study provides furtherevidence that THC pre-exposure increases the locomo-tor stimulating effects of heroin. Results further revealedthat pre-exposure to THC significantly increased heroin-
induced Fos-IR in the dorsomedial CPu and the NAC(core), but reduced heroin-induced Fos-IR in the BNST,CEA, and the PAG (dorsolateral and lateral). Thesefindings provide new evidence for the neural correlatesthat may underlie cannabinoid and opioid cross-sensitisation.
Acknowledgements
This study was supported by the University of NewEngland and by a grant from the Australian ResearchCouncil to I.S.M. and P.E.M. M.E.S.’s studies weresupported by an Australian Postgraduate Award. Theauthors would also like to thank June Young, MelanieO’Shea and Aaron Verty for technical assistance.
References
AIWH, 2002. Statistics on Drug Use in Australia 2002. Australian
Institute for Health and Welfare, Canberra.
Alheid, G., De Olmos, J.E., Beltramino, C.A., 1995. Amygdala and
extended amygdala. In: Paxinos, G. (Ed.), The Rat Nervous
System. Academic Press, New York, pp. 495e578.Arnold, J.C., Topple, A.N., Hunt, G.E., McGregor, I.S., 1998. Effects
of pre-exposure and co-administration of the cannabinoid receptor
agonist CP 55,940 on behavioural sensitization to cocaine.
European Journal of Pharmacology 354, 9e16.Brown, L.L., Smith, D.M., Goldbloom, L.M., 1998. Organizing
principles of cortical integration in the rat neostriatum: Corticos-
triate map of the body surface is an ordered lattice of curved
laminae and radial points. Journal of Comparative Neurology 392,
468e488.
Cadoni, C., Pisanu, A., Solinas, M., Acquas, E., Di Chiara, G., 2001.
Behavioural sensitization after repeated exposure to D9-tetrahy-
drocannabinol and cross-sensitization with morphine. Psychophar-
macology (Berlin) 158, 259e266.
Caine, S.B., Heinrichs, S.C., Coffin, V.L., Koob, G.F., 1995. Effects of
the dopamine D-1 antagonist SCH 23390 microinjected into the
accumbens, amygdala or striatum on cocaine self-administration in
the rat. Brain Research 692, 47e56.
Cunningham, S.T., Finn, M., Kelley, A.E., 1997. Sensitization of the
locomotor response to psychostimulants after repeated opiate
exposure: Role of the nucleus accumbens. Neuropsychopharma-
cology 16, 147e155.
Curran, E.J., Akil, H., Watson, S.J., 1996. Psychomotor stimulant-
and opiate-induced c-fos mRNA expression patterns in the rat
forebrain: comparisons between acute drug treatment and a drug
challenge in sensitized animals. Neurochemical Research 21, 1425e
1435.
D’Este, L., Scontrini, A., Casini, A., Pontieri, F.E., Renda, T.G., 2002.
Heroin sensitization as mapped by c-Fos immunoreactivity in the
rat striatum. Brain Research 933, 144e149.
Di Chiara, G., Tanda, G., Bassareo, V., Pontieri, F., Acquas, E.,
Fenu, S., Cadoni, C., Carboni, E., 1999. Drug addiction as
a disorder of associative learning. Role of nucleus accumbens shell/
extended amygdala dopamine. Annals of the New York Academy
of Sciences 877, 461e485.
Donnelly, N., Hall, W., 1994. Patterns of Cannabis Use in Australia.
Australian National Drug Strategy Monograph Series No. 27.
Erdtmann-Vourliotis, M., Mayer, P., Riechert, U., Hollt, V., 2000.
Prior experience of morphine application alters the c-Fos response

1200 M.E. Singh et al. / Neuropharmacology 49 (2005) 1189e1200
to MDMA (‘ecstasy’) and cocaine in the rat striatum. Molecular
Brain Research 77, 55e64.
Fink, J.S., Smith, G.P., 1980. Mesolimbicocortical dopamine terminal
fields are necessary for normal locomotor and investigatory
exploration in rats. Brain Research 199, 359e384.
Freedman, L.J., Cassell, M.D., 1994. Distribution of dopaminergic
fibers in the central division of the extended amygdala of the rat.
Brain Research 633, 243e252.Grabus, S.D., Glowa, J.R., Riley, A.L., 2004. Morphine- and cocaine-
induced c-Fos levels in Lewis and Fischer rat strains. Brain
Research 998, 20e28.
Hall, W., Solowij, N., 1998. Adverse effects of cannabis. Lancet 352,
1611e1616.
Herkenham, M., Lynn, A.B., Johnson, M.R., Melvin, L.S.,
de Costa, B.R., Rice, K.C., 1991. Characterization and
localization of cannabiniod receptors in rat brain: A quantitative
in vitro autoradiographic study. Journal of Neuroscience 11,
563e583.
Lamarque, S., Taghzouti, K., Simon, H., 2001. Chronic treatment with
D9-tetrahydrocannabinol enhances the locomotor response to
amphetamine and heroin. Implications for vulnerability to drug
addiction. Neuropharmacology 41, 118e129.
Lett, B.T., 1989. Repeated exposures intensify rather than diminish the
rewarding effects of amphetamine, morphine, and cocaine.
Psychopharmacology (Berlin) 98, 357e362.
Miczek, K.A., Nikulina, E.M., Kream, R.M., Carter, G., Espejo, E.F.,
1999. Behavioral sensitization to cocaine after a brief social defeat
stress: c-fos expression in the PAG. Psychopharmacology 141,
225e234.
Miller, N.S., Klahr, A.L., Gold, M.S., Sweeney, K., Cocores, J.A.,
Sweeney, D.R., 1990. Cannabis diagnosis of patients receiving
treatment for cocaine dependence. Journal of Substance Abuse 2,
107e111.
Navarro, M., Carrera, M.R., Fratta, W., Valverde, O., Cossu, G.,
Fattore, L., Chowen, J.A., Gomez, R., del Arco, I.,
Villanua, M.A., Maldonado, R., Koob, G.F., de Fonseca, F.R.,
2001. Functional interaction between opioid and cannabinoid
receptors in drug self-administration. Journal of Neuroscience 21,
5344e5350.
Norwood, C.S., Cornish, J.L., Mallet, P.E., McGregor, I.S., 2003. Pre-
exposure to the cannabinoid receptor agonist CP 55940 enhances
morphine behavioural sensitization and alters morphine self-
administration in Lewis rats. European Journal of Pharmacology
465, 105e114.
Olmstead, M.C., Franklin, K.B., 1997a. The development of
a conditioned place preference to morphine: effects of lesions of
various CNS sites. Behavioural Neuroscience 111, 1313e1323.
Olmstead, M.C., Franklin, K.B., 1997b. The development of a condi-
tioned place preference to morphine: Effects of microinjections into
various CNS sites. Behavioural Neuroscience 111, 1324e1334.
Paxinos, G., Watson, C., 1998. The Rat Brain in Stereotaxic
Coordinates. Academic Press, New York.
Pontieri, F.E., Calo, L., Di Grezia, R., Orzi, F., Passarelli, F., 1997.
Functional correlates of heroin sensitization in the rat brain.
European Journal of Pharmacology 335, 133e137.Pontieri, F.E., Monnazzi, P., Scontrini, A., Buttarelli, F.R.,
Patacchioli, F.R., 2001. Behavioral sensitization to heroin by
cannabinoid pretreatment in the rat. European Journal of
Pharmacology 421, R1eR3.
Shammah-Lagnado, S.J., Alheid, G.F., Heimer, L., 2001. Striatal and
central extended amygdala parts of the interstitial nucleus of the
posterior limb of the anterior commissure: evidence from tract-
tracing techniques in the rat. Journal of Comparative Neurology
439, 104e126.
Shu, S.Y., Ju, G., Fan, L.Z., 1988. The glucose oxidase-DAB-nickel
method in peroxidase histochemistry of the nervous system.
Neuroscience Letters 85, 169e171.
Singh, M.E., McGregor, I.S., Mallet, P.E. Perinatal exposure to D9-
tetrahydrocannabinol alters heroin-induced place conditioning
and Fos-immunoreactivity. Neuropsychopharmacology, in press.
doi:10.1038/sj.npp.1300770.
Solinas, M., Panlilio, L.V., Goldberg, S.R., 2004. Exposure to D9-
tetrahydrocannabinol (THC) increases subsequent heroin taking
but not heroin’s reinforcing efficacy: A self-administration study in
rats. Neuropsychopharmacology 29, 1301e1311.
Stevens, K.E., Mickley, G.A., McDermott, L.J., 1986. Brain areas
involved in production of morphine-induced locomotor hyperac-
tivity of the C57B1/6J mouse. Pharmacology Biochemistry and
Behavior 24, 1739e1747.
Tanda, G., Goldberg, S.R., 2003. Cannabinoids: Reward, dependence,
and underlying neurochemical mechanisms d a review of recent
preclinical data. Psychopharmacology (Berlin) 169, 115e134.
Vaughan, C.W., Bagley, E.E., Drew, G.M., Schuller, A., Pintar, J.E.,
Hack, S.P., Christie, M.J., 2003. Cellular actions of opioids on
periaqueductal grey neurons from C57B16/J mice and mutant
mice lacking MOR-1. British Journal of Pharmacology 139,
362e367.
Warty, N.A., Singh, M.E., McGregor, I.S., Mallet, P.E., 2002.
Cannabinoid receptor antagonist modulation of morphine-induced
Fos expression in feeding-related brain regions. 32nd Annual
Meeting of the Society for Neuroscience, Orlando, Florida, USA.
Wu, X., French, E.D., 2000. Effects of chronic D9-tetrahydrocannab-
inol on rat midbrain dopamine neurons: an electrophysiological
assessment. Neuropharmacology 39, 391e398.




















