
Relationship between enzymatic activity loss and post-translational protein modification in aging
11
Arch. Gerontol. Geriatr., 12 (1991) 187-197 © 1991 Elsevier Science Publishers B.V. 0167-4943/91/$03.50 AGG00395 187 Relationship between enzymatic activity loss and post-translational protein modification in aging * A. Machado, A. Ayala, E. Gordillo, E. Revilla and C. Santa Maria Departamento de Bioquimica, Bromatologia y Toxicologia, Universidad de Sevilla, 41012 Sevilla, Spain (Received 1 October 1990; accepted 13 December 1990) Summary During aging, there is a decrease in the activity of many enzymes. The mechanism causing the loss of activity is still not well understood in most cases. We have studied the decrease in the activities of the malic, 6-phosphogluconate dehydrogenase and superoxide dismutase enzymes. The old malic enzyme is about 36% less active than the young enzyme and the old 6-phosphogluconic dehydrogenase enzyme is about 26% less active than the young enzyme. In this paper, some chemical properties of these enzymes are studied. Diethyl pyrocarbonate measurements indicate that the old malic enzyme has 1 histidine residue less than the young malic enzyme. Moreover, the treatment of the young malic enzyme with ascorbate for 15 rain produces the loss of 36% of enzymatic activity and the loss of 1.2 histidine residues. 2,4,6-trinitrobenzenesulfonic acid measurements indicate that the old 6-phosphogluconate dehydrogenase enzyme has 11 lysine residues less than the young 6-phosphogluconate dehydrogenase enzyme. The proteolysis with trypsin produces more peptides in the young 6-phosphogluconate dehydrogenase enzyme than in the old one. However, similar numbers of peptides were produced when endoproteinase Arg-C was used in both enzymes, young and old 6-phosphogluconate dehydrogenase. Moreover, the treatment of young 6-phosphogluconate dehydrogenase enzyme with ascorbate for 15 min produces the loss of 8 lysine residues. These results suggest that during aging the modification of histidine residue could be involved in the loss of malic enzyme activity, and the modification of lysine residues could be involved in the loss of 6-phosphogluconate dehydrogenase activity. These results could also suggest that the modification of histidine and lysine residues during aging could be produced by oxidation. This could be a general process in aging, with an increase in the oxidation of many proteins. The relevance of this process in the aging effects must be related to the kind of proteins that are susceptible of oxidation and that this oxidation affects their enzymatic or biological function. We have also studied other enzymes one of which is the superoxide dismutase enzyme involved in the protection against oxidative damage. Our results are similar to those described for malic enzyme. In the latter case, the failure to measure one of the histidines in the Cu/Zn SOD is due to a chemical modification, probably caused by oxidation of the residue. * Paper presented and discussed in the program of the International Symposium 'Fritz Verz~r Project - 2000', held from 30 September to 3 October 1990 at Ancona, Italy. Correspondence to: Dr. A. Machado, Departamento de Bioquimica, Bromatologia y Toxicologia, Universidad de Sevilla, 41012 Sevilla, Spain.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Relationship between enzymatic activity loss and post-translational protein modification in aging

Arch. Gerontol. Geriatr., 12 (1991) 187-197 © 1991 Elsevier Science Publishers B.V. 0167-4943/91/$03.50
A G G 0 0 3 9 5
187
Relationship between enzymatic activity loss and post-translational protein modification in aging *
A. Machado, A. Ayala, E. Gordillo, E. Revilla and C. Santa Maria Departamento de Bioquimica, Bromatologia y Toxicologia, Universidad de Sevilla, 41012 Sevilla, Spain
(Received 1 October 1990; accepted 13 December 1990)
Summary
During aging, there is a decrease in the activity of many enzymes. The mechanism causing the loss of activity is still not well understood in most cases. We have studied the decrease in the activities of the malic, 6-phosphogluconate dehydrogenase and superoxide dismutase enzymes. The old malic enzyme is about 36% less active than the young enzyme and the old 6-phosphogluconic dehydrogenase enzyme is about 26% less active than the young enzyme. In this paper, some chemical properties of these enzymes are studied. Diethyl pyrocarbonate measurements indicate that the old malic enzyme has 1 histidine residue less than the young malic enzyme. Moreover, the treatment of the young malic enzyme with ascorbate for 15 rain produces the loss of 36% of enzymatic activity and the loss of 1.2 histidine residues. 2,4,6-trinitrobenzenesulfonic acid measurements indicate that the old 6-phosphogluconate dehydrogenase enzyme has 11 lysine residues less than the young 6-phosphogluconate dehydrogenase enzyme. The proteolysis with trypsin produces more peptides in the young 6-phosphogluconate dehydrogenase enzyme than in the old one. However, similar numbers of peptides were produced when endoproteinase Arg-C was used in both enzymes, young and old 6-phosphogluconate dehydrogenase. Moreover, the treatment of young 6-phosphogluconate dehydrogenase enzyme with ascorbate for 15 min produces the loss of 8 lysine residues. These results suggest that during aging the modification of histidine residue could be involved in the loss of malic enzyme activity, and the modification of lysine residues could be involved in the loss of 6-phosphogluconate dehydrogenase activity. These results could also suggest that the modification of histidine and lysine residues during aging could be produced by oxidation. This could be a general process in aging, with an increase in the oxidation of many proteins. The relevance of this process in the aging effects must be related to the kind of proteins that are susceptible of oxidation and that this oxidation affects their enzymatic or biological function. We have also studied other enzymes one of which is the superoxide dismutase enzyme involved in the protection against oxidative damage. Our results are similar to those described for malic enzyme. In the latter case, the failure to measure one of the histidines in the C u / Z n SOD is due to a chemical modification, probably caused by oxidation of the residue.
* Paper presented and discussed in the program of the International Symposium 'Fri tz Verz~r Project - 2000', held from 30 September to 3 October 1990 at Ancona, Italy.
Correspondence to: Dr. A. Machado, Departamento de Bioquimica, Bromatologia y Toxicologia, Universidad de Sevilla, 41012 Sevilla, Spain.

188
Aging; Malic enzyme, 6-phosphogluconate dehydrogenase; Superoxide dismutase; Histidine modifica- tion; Lysine modification; Protein modification
Introduction
The alteration of a large number of enzymes with age has been well documented in many organisms (Reiss and Gershon, 1976; Reznick and Gershon, 1979; Sharma and Rothstein, 1980; Rothstein, 1982; Machado et al., 1983). It has been found that in many enzymes there was a significant reduction in specific activity per unit of enzyme antigen and changes in heat sensitivity as a function of age (Reznick and Gershon, 1979). However, no changes were found in molecular weights, kinetic parameters, or electrophoretic mobility of active enzyme (Reiss and Gershon, 1976; Goren et al., 1977). The mechanism causing the loss of activity is still not well understood. We have studied this phenomenon assuming that one explanation could be a post-translational modification.
The study was carried out on the enzymes: malic enzyme (Gordillo et al., 1988), 6-phosphogluconic dehydrogenase (Gordillo et al., 1989) and superoxide dismutase (Revilla et al., 1990).
Malic enzyme
Previous studies carried out in our laboratory (Vitorica et al., 1981) showed that malic enzyme has a lower activity in old tissue homogenates of rat liver than in young ones.
Immunoprecipitat ion can give a good estimation of the amount of im- munoprecipitable malic enzymes. The loss in enzyme activity after incubation with different amounts of antiserum is proportional to the amount of antiserum added.
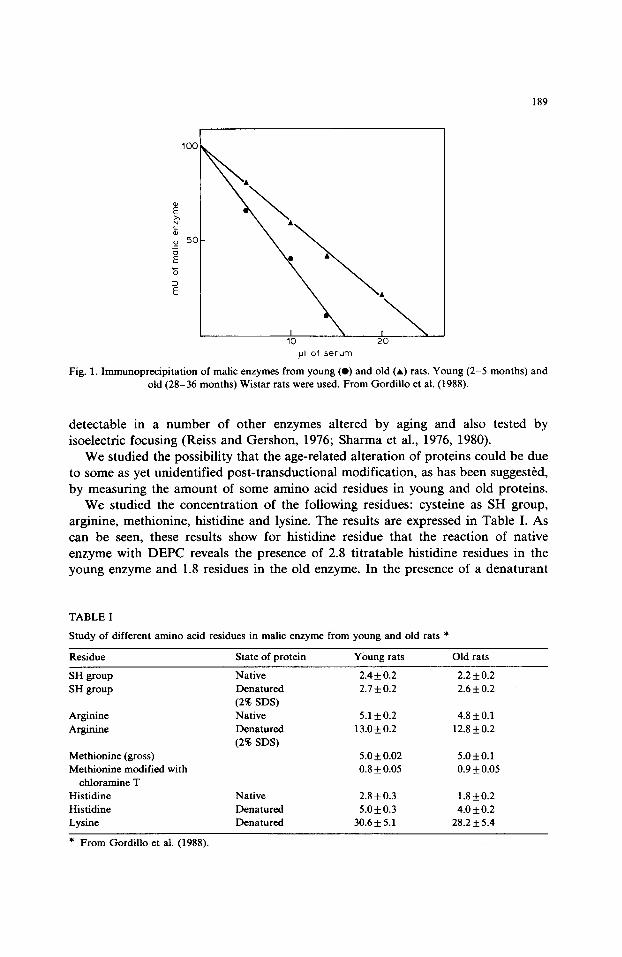
In this experiment, the results of which are shown in Fig. 1, it was found that the amount of antibody needed to inactivate 50% of the activity of tissue extract was different according to age of extract. We needed 36% more antibody to inactivate the same number of milliunits of malic enzyme from old liver extract than from young liver extract. This suggests either that in old liver extract there is more malic enzyme protein available to produce the immunoprecipitable reaction or that the old enzyme has 36% less activity than the young one. Thus, the indication is that old liver extract contains a less active form of malic enzyme or it contains a mixture of fully active and inactive malic enzyme molecules. Furthermore, this suggests that malic enzyme is altered and accumulated with age. This is not an abnormal behaviour for the enzyme during aging.
No changes were found in some properties of malic enzyme from young and old animals, for example, molecular weight, subunit number, isoelectric point, K m for oxalacetate and N A D P H and thermostability. This suggests that the differences in activity observed in young and old malic enzymes are not the result of covalent changes such as phosphorylation, acylation or deamination. Changes were not
189
10o . o \ \
\ \ 10 2 0
jJI o l s e r u m
Fig. 1. Immunoprecipitation of malic enzymes from young (e) and old (A) rats. Young (2-5 months) and old (28-36 months) Wistar rats were used. From Gordillo et al. (1988).
detectable in a number of other enzymes al tered by aging and also tested by
isoelectric focusing (Reiss and Gershon, 1976; Sharma et al., 1976, 1980).
We studied the possibil i ty that the age-related a l terat ion of proteins could be due
to some as yet unident i f ied pos t - t ransduct ional modif icat ion, as has been suggested,
by measur ing the a m o u n t of some amino acid residues in young and old proteins.
We studied the concent ra t ion of the fol lowing residues: cysteine as SH group,
arginine, methionine, hist idine and lysine. The results are expressed in Table I. As
can be seen, these results show for hist idine residue that the react ion of nat ive
enzyme with D E P C reveals the presence of 2.8 t i t ra table hist idine residues in the
young enzyme and 1.8 residues in the old enzyme. In the presence of a dena turan t
TABLE I
Study of different amino acid residues in malic enzyme from young and old rats *
Residue State of protein Young rats Old rats
SH group Native 2.4 + 0.2 2.2 + 0.2 SH group Denatured 2.7 + 0.2 2.6 + 0.2
(2% SDS) Arginine Native 5.1 + 0.2 4.8 + 0.1 Arginine Denatured 13.0 + 0.2 12.8 + 0.2
(2% SDS) Methionine (gross) 5.0 + 0.02 5.0 + 0.1 Methionine modified with 0.8 + 0.05 0.9 + 0.05
chloramine T Histidine Native 2.8 + 0.3 1.8 + 0.2 Histidine Denatured 5.0 + 0.3 4.0 + 0.2 Lysine Denatured 30.6 + 5.1 28.2 + 5.4
* From Gordillo et al. (1988).
190
100
75
"~ 5O
25
. ['-.. I 1
Groups modified per molecule
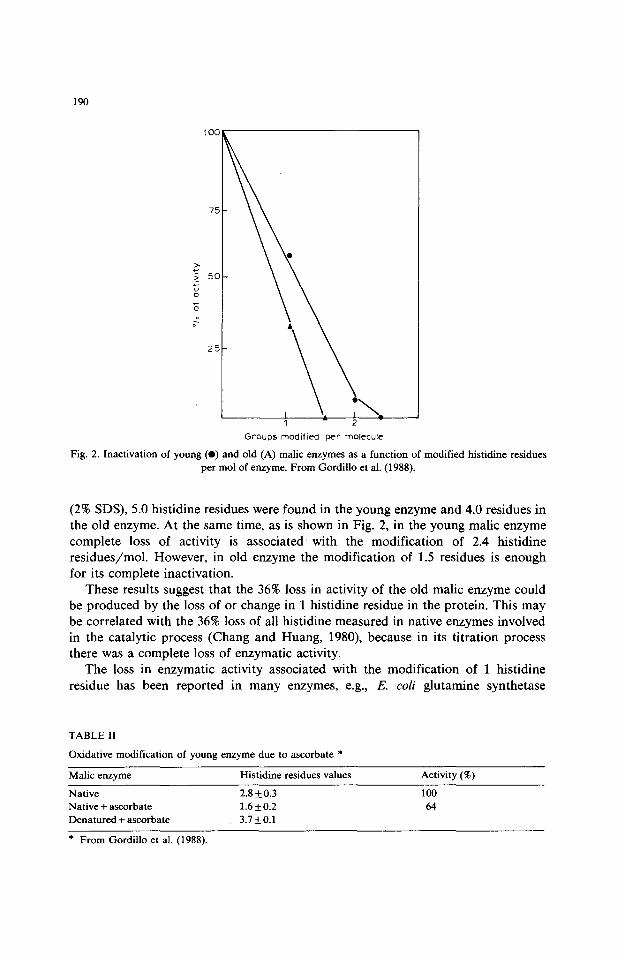
Fig. 2. Inactivation of young (e) and old (A) malic enzymes as a function of modified histidine residues per mol of enzyme. From Gordillo et al. (1988).
(2% SDS), 5.0 histidine residues were found in the young enzyme and 4.0 residues in the old enzyme. At the same time, as is shown in Fig. 2, in the young malic enzyme complete loss of activity is associated with the modification of 2.4 histidine residues/mol. However, in old enzyme the modification of 1.5 residues is enough for its complete inactivation.
These results suggest that the 36% loss in activity of the old malic enzyme could be produced by the loss of or change in 1 histidine residue in the protein. This may be correlated with the 36% loss of all histidine measured in native enzymes involved in the catalytic process (Chang and Huang, 1980), because in its titration process there was a complete loss of enzymatic activity.
The loss in enzymatic activity associated with the modification of 1 histidine residue has been reported in many enzymes, e.g., E. coli glutamine synthetase
TABLE II
Oxidative modification of young enzyme due to ascorbate *
Malic enzyme Histidine residues values Activity (%)
Native 2.8 5:0.3 100 Native + ascorbate 1.6 + 0.2 64 Denatured + ascorbate 3.7 5:0.1
* From Gordillo et al. (1988).
191
(Levine, 1983), phosphoglycerate kinase (Fucci et al., 1983) and mammalian super- oxide dismutase (Bray et al., 1974; Hodgson and Fridovich, 1975). In these cases, the change is produced by oxidative inactivation that is accompanied by the loss of 1 or more histidine residues (Bray et al., 1974; Hodgson and Fridovich, 1975; Fucci et al., 1983; Levine, 1983).
We have also studied the behaviour of the young malic enzyme with ascorbate. The results are shown in Table II. The addition of 15 mM ascorbate produced a loss of 36% of the malic enzyme activity and the loss of 1.2 histidine residues measured with DPEC after 15 min. This suggests that the loss in enzymatic activity found in old malic enzyme could be produced by the oxidation of 1 histidine residue of the native enzyme.
6-Phosphogluconic dehydrogenase
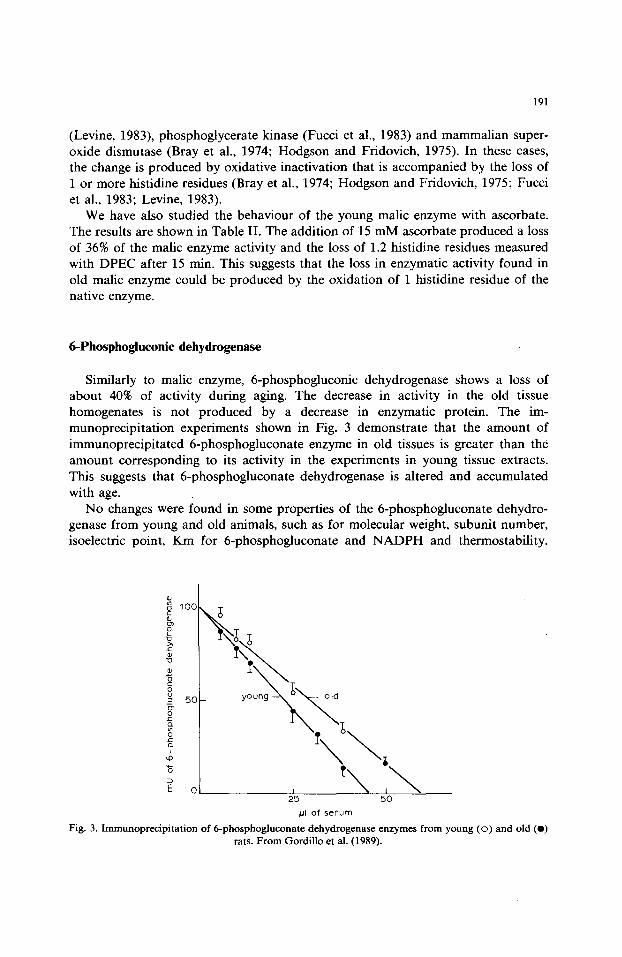
Similarly to malic enzyme, 6-phosphogluconic dehydrogenase shows a loss of about 40% of activity during aging. The decrease in activity in the old tissue homogenates is not produced by a decrease in enzymatic protein. The im- munoprecipitation experiments shown in Fig. 3 demonstrate that the amount of immunoprecipitated 6-phosphogluconate enzyme in old tissues is greater than the amount corresponding to its activity in the experiments in young tissue extracts. This suggests that 6-phosphogluconate dehydrogenase is altered and accumulated with age.
No changes were found in some properties of the 6-phosphogluconate dehydro- genase from young and old animals, such as for molecular weight, subunit number, isoelectric point, Km for 6-phosphogluconate and NADPH and thermostability.
o
{
8 ~ .c o .
"5
E
100
50
T
y o u n g - - ~ - old
' \ , \ 25 50
ul ol s@rum
Fig. 3. Immunoprecipitat ion of 6-phosphogluconate dehydrogenase enzymes from young (o ) and old (e) rats. From Gordillo et al. (1989).

192
TABLE III
Study of different amino acid residues in 6-phosphogluconate dehydrogenase from young and old rats *
Residue State of protein Young rats Old rats
SH group Native 7.4 + 0.5 7.5 + 0.4 SH group Denatured 9.7 + 0.5 9.3 + 0.5 Arginine Native 8.1 + 0.2 7.9 + 0.2 Arginine Denatured 15.6 __. 0.2 15.7 + 0.2 Methionine (gross) 10.0 + 0.05 10.0 + 0.1 Methionine modified with 0.8 + 0.05 0.9 + 0.05
chloramine T Histidine Native 4.0 + 0.1 3.6 + 0.2 Histidine Denatured 7.4 + 0.2 7.2 + 0.2 Lysine Denatured 39.0 + 2.8 28.2 + 5.3 * *
* From Gordillo et al. (1989). ** P <0.02 when compared with corresponding values from young enzyme.
This suggests that the differences in activity observed in young and old enzymes are not the results of covalent changes such as phosphorylat ion, acylation or deamina- tion. Similar results were found for malic enzyme.
As with malic enzyme, the modificat ions of some amino acid residues in young and old 6-phosphogluconate dehydrogenase enzymes have been studied. The results are summarized in Table III . As can be seen, in old enzyme there was a loss or change of 11 lysine res idues /mol of protein, al though no change was found in the amoun t of other amino acids studied, including histidine residues.
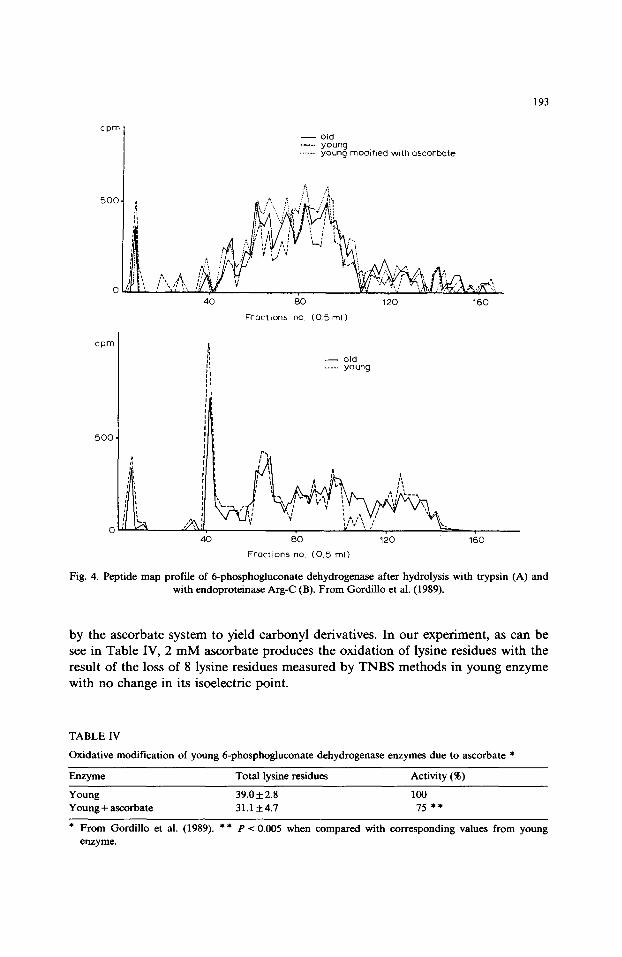
The modifications of lysine have also been confirmed by use of trypsin which splits peptide bonds of protein in the arginine and lysine residues. The peptide maps of young and old enzyme were different, as can be see in Fig. 4A, The old enzyme has fewer peptides than the young one.
However, when endoproteinase Arg-C, which cuts the protein only on arginine residues, was used, the peptide maps of both enzymes were similar (Fig. 4B). These results confirm that the change found with trypsin was due to the presence of some lysine residues in the old enzyme.
In contrast to the case of histidine (Bray et al., 1974; Hodgson and Fridovich, 1975; Fucci et al., 1983; Levine 1983; Gordil lo et al., 1988), the loss in enzymatic activity associated with the modificat ion of lysine has not been reported. However, the S tadtman group has suggested that the presence of inactive or less active enzymes during aging may be due to the oxidation of histidine or lysine residues (Stadtman, 1986). The 6-phosphogluconate dehydrogenase enzyme could be one example of oxidation in lysine residues.
We have studied the behaviour of the young enzyme when it is incubated with ascorbate products that produce oxidation of histidine in some enzymes (Levine 1983; Gordil lo et al., 1988). The oxidative capaci ty of ascorbate is produced by radical forms (Lewin, 1976; Winterbourn, 1981). Furthermore, studies with amino acid homopolymers show that histidine, lysine and other amino acids are oxidized
193
c p m
5 0 0 .
o l d ---- y o u n g .......... y o u n g m o d i f i e d w i t h a s c o r b e t e
, / ; :.'4 , ' ~.. ~ ~ " / " I
,' y[ ,,"-, ,/ ~"-~
4 0 8 0 1 2 0 1 6 0
F r a c t i o n s no. ( O b m l )
cpm - - o l d . . . . . y o u n g
5 0 0 -
" r ,L - j~ i l
, ;, ; ;,, t A , I
, ... i ,~, ~--...
; I ' ~ - - . , , ' , "
o ; ,,7~' 4 0 8 0 1 2 0 1 6 0
F r a c t i o n s no. ( 0 . 5 m l )
Fig. 4. Peptide map profile of 6-phosphogluconate dehydrogenase after hydrolysis with trypsin (A) and with endoproteinase Arg-C (B). From Gordillo et al. (1989).
by the ascorbate system to yield carbonyl derivatives. In our experiment, as can be see in Table IV, 2 m M ascorbate produces the oxidat ion of lysine residues with the
result of the loss of 8 lysine residues measured by TNBS methods in young enzyme
with no change in its isoelectric point .
TABLE IV
Oxidative modification of young 6-phosphogluconate dehydrogenase enzymes due to ascorbate *
Enzyme Total lysine residues Activity (~)
Young 39.0 + 2.8 100 Young+ ascorbate 31.14-4.7 75 * *
* From GordiUo et al. (1989). * * P < 0.005 when compared with corresponding values from young enzyme.
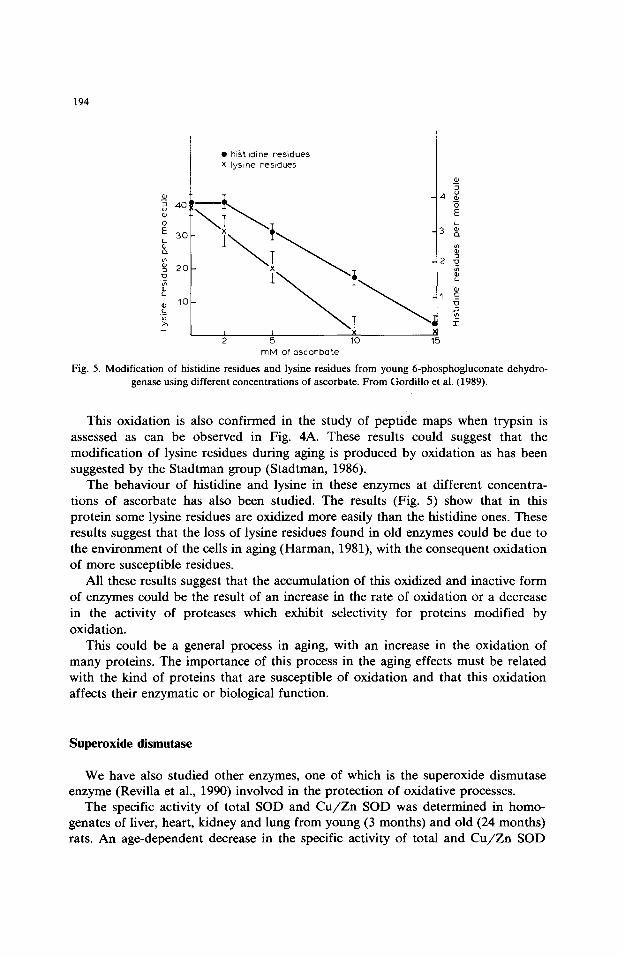
194
• hist idine residues x lysine residues
dJ
-5 40 i- "6
o E 30 3 ~
I:l
~2o 2~
2 T I I x 2 5 10 15
mM of oscorba te
Fig. 5. Modification of histidine residues and lysine residues from young 6-phosphogluconate dehydro- genase using different concentrations of ascorbate. From Gordillo et al. (1989).
This oxidation is also confirmed in the study of peptide maps when trypsin is assessed as can be observed in Fig. 4A. These results could suggest that the modification of lysine residues during aging is produced by oxidation as has been suggested by the Stadtman group (Stadtman, 1986).
The behaviour of histidine and lysine in these enzymes at different concentra- tions of ascorbate has also been studied. The results (Fig. 5) show that in this protein some lysine residues are oxidized more easily than the histidine ones. These results suggest that the loss of lysine residues found in old enzymes could be due to the environment of the cells in aging (Harman, 1981), with the consequent oxidation of more susceptible residues.
All these results suggest that the accumulation of this oxidized and inactive form of enzymes could be the result of an increase in the rate of oxidation or a decrease in the activity of proteases which exhibit selectivity for proteins modified by oxidation.
This could be a general process in aging, with an increase in the oxidation of many proteins. The importance of this process in the aging effects must be related with the kind of proteins that are susceptible of oxidation and that this oxidation affects their enzymatic or biological function.
Superoxide dismutase
We have also studied other enzymes, one of which is the superoxide dismutase enzyme (Revilla et al., 1990) involved in the protection of oxidative processes.
The specific activity of total SOD and C u / Z n SOD was determined in homo- genates of liver, heart, kidney and lung from young (3 months) and old (24 months) rats. An age-dependent decrease in the specific activity of total and C u / Z n SOD

195
from liver, kidney and heart was observed, whereas no such decrease occurred in the enzyme from lung.
The amount of C u / Z n SOD enzyme in liver, heart, kidney and lung homogenates from young and old rats was measured by inmunoprecipitation with an antiserum prepared against purified young rat liver SOD. In each case, increasing amounts of the antiserum were added to aliquots of enzyme activity. There was a higher amount of immunoprecipitable protein in liver, heart and kidney of the old rat, but no change was found in lung. In fact, this latter tissue showed no variation in activity during aging. These results suggest that there is an accumulation of altered protein in those organs where a decrease in the SOD activity occurs during aging.
This decrease in enzymatic activity associated with no decrease in the amount of protein in hver of aged rats indicates that there is an inactivation of C u / Z n SOD during aging. It is currently assumed that the decrease in activity during aging is due to a post-translational modification of enzyme. To detect this possible alteration, the quantification of various amino acid residues in C u / Z n SOD protein of young and old rats was determined by immunoaffinity using purified protein. We have used liver enzyme because this is the organ that exhibits maximal activity.
The results show that there is no difference between the concentrations of cysteine, arginine and lysine in the SOD of young and old rats. However, a statistically significant difference was obtained between the histidine residues; the old protein has less histidine than the young and this residue is accessible in the native protein. These results are similar to those described for malic enzyme (Gordillo et al., 1988) and suggest that, as in the latter case, the failure to measure one of the histidines in the C u / Z n SOD is due to a chemical modification, probably caused by oxidation of the residue. This is supported by the finding that the C u / Z n SOD is inactivated by oxidation in vitro (Fucci et al., 1983) and that this inactiva- tion is accompanied by the loss of one or more histidine residues (Bray et al., 1974; Hodgson and Fridovich, 1975).
This result could be very important because the reduction in SOD activity as a function of age could result in an impaired protection against the toxic effects of radicals such as the superoxide anion, and thus might lead to severe cellular damage. The superoxide free radical has been suggested as an important agent implicated in the aging process.
Stadtman has shown that several enzymes are inactivated by oxidation in vitro (Fucci et al., 1983). Moreover, some of these enzymes lose activity during aging, as found for malic enzyme, 6 -P GDH and C u / Z n SOD. These old enzymes show either one fewer measured histidine (malic enzyme and SOD) or several fewer lysine residues. The same results, i.e., the loss of histidine or lysine, are shown in the young enzyme when inactivated by oxidation in vitro. Some data point to the possibility that the amino acid residues modified in vitro by oxidation and during the aging process are the same in the sequence, i.e., they occupy identical position in the protein structure.
The above discussion suggests that there is a great similarity between the inactivation process by oxidation in vitro and the inactivation produced during aging. The loss of amino acid residues found in old enzymes could be produced by

196
an increase in the oxidative environment of cells in aging (Harman, 1981), with the consequent oxidation of the more susceptible residues.
In summary, this work suggests that during aging there is oxidative damage to the protein in which the oxidative modification of some amino acid residues (histidine and/or lysine) takes place. In this process, there could be a loss of biological activity in some of these proteins, depending on which amino acid is oxidized. The importance of this process will depend on the physiological relevance of the protein involved in it.
Acknowledgement
This work was supported by grants from the" DGICYT PM89-0173.
Discussion of the lecture
Ivy: This is probably a very naive question: in my lecture I talked about the decrease of thiol-pro- teinase activities, and in my understanding, the thiol groups are the most sensitive to oxidative damage. You have shown that thiol groups have not been damaged in your particular enzymes. Does it mean that I should reverse my hypothesis?
Machado: The determinant factor is the accessibility of the given groups for the oxidative agents. In the case of the enzymes studied by us, the lysine residues exposed on the surface of the molecules are especially accessible, whereas SH groups may be present in a more hidden, protected position in the given protein molecules.
Floyd: I have a comment on that. Stadtman has made a number of experiments on these topics, and your and his results agree very well, as far as I know. He has found that the site specificity of metal binding near to the active site of the molecule has also an important role in the damaging processes. Could you confirm these results of Dr. Stadtman?
Machado: I am sorry, we did not analyze the role of metal binding sites.
Cutler: I want to make a general comment on the idea that oxidatively damaged proteins may be accumulated in the cells, leading to a decrease of their functional activity. I believe that the most important targets of the oxidative damage could be the regulatory proteins, which control some regulatory genes, etc. The damage of these regulatory proteins may have more consequences than the damage of enzymes not involved directly in gene regulation. This probably may explain better the importance of altered physiological functions during aging.
Machado: You may be right, but for me the observed changes are important, too, since they may alter the specific structure of the enzyme molecules to different extents. Some molecules are sensitive for the damage of one single amino acid, others tolerate better even more extensive damage, in terms of their chemical characteristics. It remains a fact that the altered enzyme molecules are of lower activity.
Zs.-Nagy: I think it should be stressed by experimental gerontologists that it is not sufficient any more to measure just enzyme activities per mg of total protein basis, etc, as usual. It would be desirable to measure the number of enzyme molecules present in the system (e.g., by inmuno-reactive methods), and then express the specific activity of a given enzyme per molecule. I want to stress that David Gershon has shown already 20 years ago that the immunologlcally reactive amount of certain enzymes increases with age in various tissues, whereas the total enzyme activities decreased. For me it was always surprising that people usually do not pay attention to these facts. If we have a two-fold increase in the number of enzyme molecules present, and an unchanged enzyme activity per mg of total protein, we just

197
mask a 50% drop of the specific activity of the given enzyme molecules. I realize that such an approach requires much more work, but this cannot be the limiting factor for the introduction of much more reliable methods for measuring enzyme activities.
References
Bray, R.C., Cockle, S.A., Fielden, E.M., Roberts, P.B., Rotilio, G. and Calabrese, L. (1974): Detection and inactivation of superoxide dismutase by hydrogen peroxide. Biochem. J., 139, 43-48.
Chang, G.G. and Huang, T.M. (1980): Involvement of tyrosyl residues of the substrate binding of pigeon liver malic enzyme. Biochim. Biophys. Acta, 611,217-226.
Fucci, L., Oliver, C.N., Coon, M.J. and Stadman, R. (1983): Inactivation of key metabolic enzymes by mixed-function oxidation reactions: Possible implication in protein turnover and ageing. Proc. Natl. Acad. Sci. U.S.A., 80, 1521-1525.
Gordillo, E., Ayala, A., F-Lobato, M., Bautista, J. and Machado, A. (1988): Possible involvement of histidine residues in the loss of enzymatic activity of rat liver malic enzyme during aging. J. Biol. Chem., 263, 8053-8057.
Gordillo, E., Ayala, A., Bautista, J. and Machado, A. (1989): Implication of lysine residues in the loss of enzymatic activity in rat liver 6-phosphogluconate dehydrogenase found in aging. J. Biol. Chem., 264, 17014-17019.
Goren, P., Reznick, A., Reiss, U. and Gershon, D. (1977): Isoelectric properties of nematede aldolase and liver superoxide dismutase from young and old animals. FEBS Lett., 84, 83-86.
Harman, D. (1981): The ageing process. Proc. Natl. Acad. Sci. U.S.A., 78, 7124-7128. Hodgson, E.K. and Fridovich, I. (1975): The interaction of bovine erythrocyte superoxide dismutase with
hydrogen peroxide: inactivation of the enzyme. Biochemistry, 14, 5294-5299. Levine, R.L. (1983): Oxidative modification of glutamine synthetase. J. Biol. Chem., 258, 11828-11833. Lewin, S. (1976): Vitamin C: its Molecular Biology and Medical Potential. Academic Press, Orlando, FL. Machado, A., Satrustegui, J., Andres, A., Vitorica, J., Cano, J. and Fabregat, I. (1983): Age-related
changes in enzymes activities linked to energy metabolism. In: Cardiology and Aging, pp. 65-80. Editor: D. Platt. F.K. Schattauer Verlag, New York.
Reiss, U. and Gershon, D. (1976): Rat liver superoxide dismutase. Eur. J. Biochem., 63, 617-623. Revilla, E., Santa Maria, C., Ayala, A. and Machado A. (1990): Changes in histidine residues of Cu/Zn
superoxide dismutase during ageing (in preparation). Reznick, A.Z. and Gershon, D. (1979): The effect of age on the protein degradation system in the
nematode Turbatrix aceti. Mech. Ageing Dev., 11,403-415. Rothstein, M. (1982): Biochemical Approaches to Aging, pp. 213-255. Academic Press, Orlando, FL. Sharma, H.K., Gupta, S.K. and Rothstein, M. (1976): Age-related alterations of enolase in the free living
nematode Turbatrix aceti. Arch. Biochem. Biophys., 174, 324-332. Sharma, H.K. and Rothstein, M. (1980): Altered enolase in aged Turbatrix aceti results from conforma-
tion changes in the enzyme. Proc. Natl. Acad. Sci. U.S.A., 77, 5865-5868. Sharma, H.K., Prasanna, H.R. and Rothstein, M. (1980): Altered phosphoglycerate kinase in aged rats. J.
Biol. Chem., 255, 5043-5050. Stadtman, E.R. (1986): Oxidation of proteins by mixed-function oxidation system: implication in protein
turnover. Aging and neutrophil function. Trends Biochem. Sci., 11, 11-12. Vitorica, J., Satrustegui, J. and Machado, A. (1981): Changes in activities of key lipogenic and
gluconeogenic enzymes in the aged rat liver. Enzyme, 26, 144-152. Winterbourn, C.C. (1981): Hydroxyl radical production in body fluids. Biochem. J., 198, 125-131.




















