
Reciprocal Chromosome Translocation, rcp(7;17)(q26;q11), in a Boar Giving Reduced Litter Size and...
11
Hereditas 122: 257-267 (1995) Reciprocal chromosome translocation, rcp( 7;17)( q26;q? l), in a boar giving reduced litter size and increased rate of piglets dying in the early life D. A. F. VILLAGOMEZ', I. GUSTAVSSON', L. JONSSON' and L. PLOEN3 ' Department of Animal Breeding and Genetics, Department of Pathology, and Department of Anatomy and Histology, Swedish University of Agricultural Sciences, Uppsala, Sweden ViLLAGoMEz, D. A. F., GUSTAVSSON, I., JONSSON, L. and PLOEN, L. 1995. Reciprocal chromosome translocation, rcp(7;17)(q26;ql I), in a boar giving reduced litter size and increased rate of piglets dying in the early life. - Heredifas 122: 257-267. Lund, Sweden. ISSN 0018-0661. Received February 15, 1995. Accepted June 22, 1995 Cytogenetic investigations in a boar causing a 41 YO reduction in litter size and also producing piglets that died soon after birth revealed the presence of a reciprocal chromosome translocation, rcp(7;17)(qZ6;ql I). The translocation resulted in one extremely small telocentric chromosome marker. Synaptonemal complex analysis of spread spermatocytes by electron microscopy revealed an unusual behaviour of the transloca- tion. This formed not only different types of quadrivalents (78.1 % of the cells), similar to those previously found in boars heterozygous for reciprocal exchanges, but also completely or incompletely paired trivalent configurations, plus univalent (21.9 %). Association between the sex bivalent and the translocation configuration was observed (18.7 %), but testicular histology was normal. Furthermore, the boar with the translocation was found to produce tertiary monosomy and trisomy in some of the liveborn piglets. Some of the tertiary monosomic offspring, which died in the early extra-uterine life, demonstrated ventricular septa1 defect and cleft palate. Ingemar Gustavsson, Department of Animal Breeding and Genetics, The Sweedish University of Agriciiltural Sciencess, Box 7023, S-750 07 Uppsala, Sweden Reciprocal translocations are often observed in boars responsible for reduced litter size ( GUSTAVS- SON 1990). The reduction in litter size is due to the production of gametes that are chromosomally unbalanced. These gametes have a normal appear- ance and fertilization capability, but the embryos, being chromosomally abnormal, usually die early, most often before implantation (e.g., KING et al. 1981). However, there have been rare cases in which perinatal and postnatal death have been observed (WKESSON and HENRICSON 1972; GUS- TAVSSON and JONSSON 1992). Earlier descriptions of the meiotic pairing be- haviour of reciprocal translocations in boars have shown a regular synaptic pattern with the forma- tion of completely and incompletely paired quadri- valents ( GUSTAVSSON et al. 1988; GABRIEL-ROBEZ et al. 1988; JAAFAR et al. 1989). Reciprocal trans- locations are often associated with meiotic arrest in man and mouse (CHANDLEY 1982; FOREJT 1982). The reciprocal translocation, rcp( 2;4)(pl7;ql I), in pig (GUSTAVSSON et al. unpubl.) seemed to be the very first indication that a similar phenomenon may occur in pigs. In the present work we describe a case of recip- rocal translocation in a boar producing not only reduced litter size but also malformed piglets. Material and methods The proband and his offspring A Hampshire boar, 19 months old, was reported to produce 41 YO reduced litter size, estimated on 13 litters, yet had a normal semen picture. The num- ber of repeat breedings was 3. According to the farmer, 9 YO of the liveborn piglets looked sickly and died during the first few days. Blood samples were taken from the boar, from 2 piglets that died during the first days of life and from 13 piglets aged 27 to 50 days. Skin samples were taken from another 3 piglets that died during the first days of life. Tissue culture and chromosome bandings Peripheral blood cultures were set up in the con- ventional way with RPMI 1640 including 20%
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Reciprocal Chromosome Translocation, rcp(7;17)(q26;q11), in a Boar Giving Reduced Litter Size and...

Hereditas 122: 257-267 (1995)
Reciprocal chromosome translocation, rcp( 7;17)( q26;q? l), in a boar giving reduced litter size and increased rate of piglets dying in the early life D. A. F. VILLAGOMEZ', I. GUSTAVSSON', L. JONSSON' and L. PLOEN3
' Department of Animal Breeding and Genetics, Department of Pathology, and Department of Anatomy and Histology, Swedish University of Agricultural Sciences, Uppsala, Sweden
ViLLAGoMEz, D. A. F., GUSTAVSSON, I., JONSSON, L. and PLOEN, L. 1995. Reciprocal chromosome translocation, rcp(7;17)(q26;ql I) , in a boar giving reduced litter size and increased rate of piglets dying in the early life. - Heredifas 122: 257-267. Lund, Sweden. ISSN 0018-0661. Received February 15, 1995. Accepted June 22, 1995
Cytogenetic investigations in a boar causing a 41 YO reduction in litter size and also producing piglets that died soon after birth revealed the presence of a reciprocal chromosome translocation, rcp(7;17)(qZ6;ql I). The translocation resulted in one extremely small telocentric chromosome marker. Synaptonemal complex analysis of spread spermatocytes by electron microscopy revealed an unusual behaviour of the transloca- tion. This formed not only different types of quadrivalents (78.1 % of the cells), similar to those previously found in boars heterozygous for reciprocal exchanges, but also completely or incompletely paired trivalent configurations, plus univalent (21.9 %). Association between the sex bivalent and the translocation configuration was observed (18.7 %), but testicular histology was normal. Furthermore, the boar with the translocation was found to produce tertiary monosomy and trisomy in some of the liveborn piglets. Some of the tertiary monosomic offspring, which died in the early extra-uterine life, demonstrated ventricular septa1 defect and cleft palate.
Ingemar Gustavsson, Department of Animal Breeding and Genetics, The Sweedish University of Agriciiltural Sciencess, Box 7023, S-750 07 Uppsala, Sweden
Reciprocal translocations are often observed in boars responsible for reduced litter size ( GUSTAVS- SON 1990). The reduction in litter size is due to the production of gametes that are chromosomally unbalanced. These gametes have a normal appear- ance and fertilization capability, but the embryos, being chromosomally abnormal, usually die early, most often before implantation (e.g., KING et al. 1981). However, there have been rare cases in which perinatal and postnatal death have been observed (WKESSON and HENRICSON 1972; GUS- TAVSSON and JONSSON 1992).
Earlier descriptions of the meiotic pairing be- haviour of reciprocal translocations in boars have shown a regular synaptic pattern with the forma- tion of completely and incompletely paired quadri- valents ( GUSTAVSSON et al. 1988; GABRIEL-ROBEZ et al. 1988; JAAFAR et al. 1989). Reciprocal trans- locations are often associated with meiotic arrest in man and mouse (CHANDLEY 1982; FOREJT 1982). The reciprocal translocation, rcp( 2;4)( pl7;ql I ) , in pig (GUSTAVSSON et al. unpubl.) seemed to be the very first indication that a similar phenomenon may occur in pigs.
In the present work we describe a case of recip- rocal translocation in a boar producing not only reduced litter size but also malformed piglets.
Material and methods The proband and his offspring
A Hampshire boar, 19 months old, was reported to produce 41 YO reduced litter size, estimated on 13 litters, yet had a normal semen picture. The num- ber of repeat breedings was 3. According to the farmer, 9 YO of the liveborn piglets looked sickly and died during the first few days. Blood samples were taken from the boar, from 2 piglets that died during the first days of life and from 13 piglets aged 27 to 50 days. Skin samples were taken from another 3 piglets that died during the first days of life.
Tissue culture and chromosome bandings
Peripheral blood cultures were set up in the con- ventional way with RPMI 1640 including 20%

258 D. A. F. VILLAGOMEZ ET AL. Hereditas 122 (1995)
foetal calf serum as medium and with pokeweed as mitogen. On day 3 the cultures were harvested after treatment withcolcemid for 30 min, 0.075 M KCI for 20 min and fixation with glacial acetic acid plus methanol (1 + 3). Cell cultures to be used for RBA banding (see below) were treated with 5-bromo- deoxyuridine (200 pg/ml) for 7 h before harvest.
Metaphase spreads of chromosomes were stained according to the GTG- (SEABRIGHT 1971), the CBG- (SUMNER 1972), the QFQ- (CASPERSSON et al. 1971), and RBA- (DUTRILLAUX et al. 1973) banding techniques. Chromosomes were also stained conventionally with Giemsa.
Synaptonemal complex analysis
The analysis was carried out according to the surface spreading technique (COUNCE and MEYER 1973) with some modifications. Small pieces of both testes were carefully minced with tweezers and applied on drops of 0.2M sucrose on slides which had been precoated with a film of Necoloidine (BDH Chem- icals Ltd, Poole, England). After the sucrose drops had dried, the preparations were fixed with 4 % paraformaldehyde in 0.1 M sucrose containing 0.03 % of sodium dodecyl sulphate (SDS, Sigma) and stained with a solution of 1 part of 4 % aqueous solution of phosphotungstic acid (PTA) plus 3 parts of 95 % ethanol. The film with the cells was floated off on water and 100-mesh copper grids were applied on cells earlier marked out under phase contrast microscopy. The cells were studied with a Philips EM 210.
Studies on diakinesis-MI
Conventional meiotic studies were carried out ac- cording to KING (1981), adapted from PATHAK et al. (1976), and the chromosomes were stained with Giemsa or banded according to the CBG ( SUMNER 1972) or the THA (DUTRILLAUX 1973) technique.
Patho -anatomical investigations
Chromosomally unbalanced offspring plus offspring dying in early extra-uterine life, but not cytogenet- ically investigated, were X-rayed and studied patho- anatomically. Malformations were photographed.
Testicular histology
Pieces of both testes were fixed with 3 YO glutaralde- hyde in 0.067 M cacodylate buffer (pH 7.2), post-
fixed in osmium tetroxide, and embedded in Epon. Semithin sections of approximately 1 pm were stained with buffered toluidine blue.
Results Cytogenetics
Karyotype of the boar
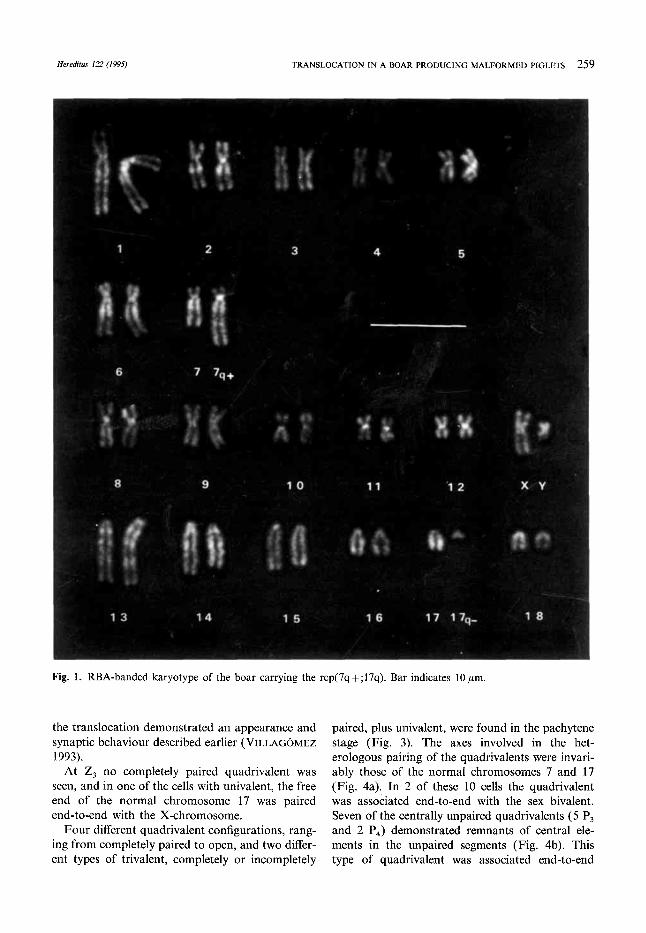
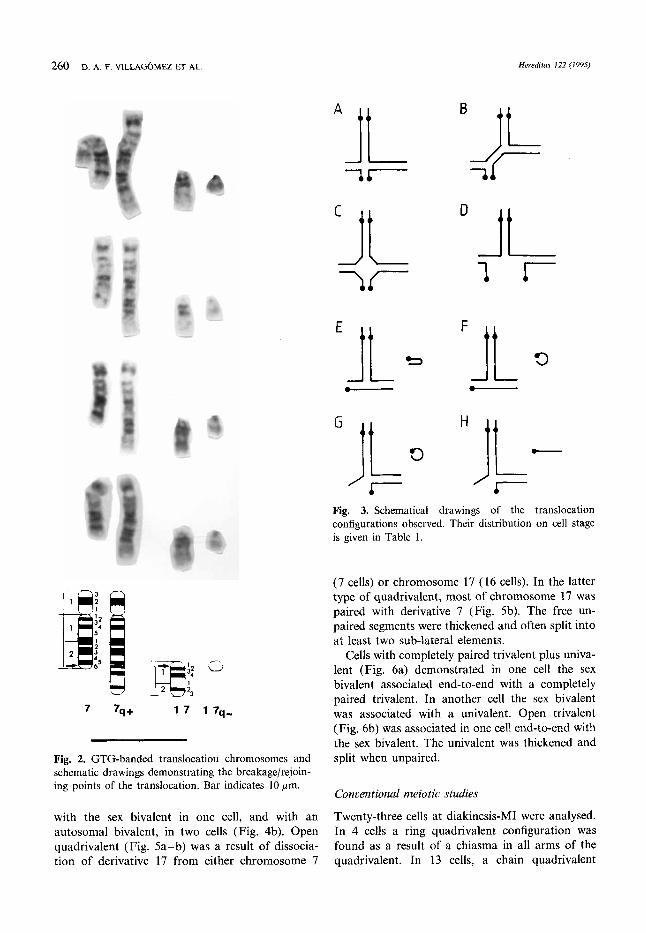
The most remarkable feature of the conventionally Giemsa-stained metaphases was an unusually small chromosome in the karyotype consisting of 38 chromosomes. On closer examination it could also be observed that there was only one normally sized chromosome 7; the second one had a size similar to chromosome 6. Additionally, only three small one- armed chromosomes of normal size could be found. After banding techniques (Fig. 1 and 2) it was quite clear that there had been an exchange of material between chromosomes 7 and 17. Most of chromosome 17, except the centromeric segment, had been translocated to chromosome 7, which had lost the most terminal segment. Apparently the breakages in both chromosomes had occurred in R-positive bands. According to the COMMITTEE
DOMESTIC PIG ( 1988) the translocation could be designated rcp(7;17)(q26;qll).
FOR THE STANDARDIZED U R Y O T Y P E OF THE
Synaptonemal complex analysis
Altogether 64 spermatocytes were investigated. Seven (10.9 YO) were at zygotene and 57 (89 %) at pachytene. They are substaged in Table 1 accord- ing to the nomenclature proposed by VILLAGOMEZ (1993). Zygotene was subdivided into three stages: early (Z,), mid-(Z,), and late (Z3), and pachytene, into three main stages: early, mid-, and late, each of which had two substages (P, - P,, P, - P4, and P, - P,, respectively). In a few cells the grid bars partly covered the nuclei. Nevertheless, the translo- cation configuration could be identified and the substage distinguished. Fifty of the 64 cells (78.2 YO) demonstrated a quadrivalent, while the remaining 14 cells (21.8 Yn) had a trivalent plus univalent. The appearance of the quadrivalent varied, as did the appearance of trivalent plus univalent (Fig. 3). The distribution of cells accord- ing to configuration is demonstrated in Table 1. In 12 cells (18.7 %) the translocation products were paired or associated with the sex bivalent. The sex chromosomes and the autosomes not involved in
Hereditas 122 (1995) TRANSLOCATION IN A BOAR PRODUCING MALFORMED PIGLETS 259
Fig. 1. RBA-banded karyotype of the boar carrying the rcp(7q+;17q). Bar indicates 10pm.
the translocation demonstrated an appearance and synaptic behaviour described earlier ( VILLAG~MEZ 1993).
At Z, no completely paired quadrivalent was seen, and in one of the cells with univalent, the free end of the normal chromosome 17 was paired end-to-end with the X-chromosome.
Four different quadrivalent configurations, rang- ing from completely paired to open, and two differ- ent types of trivalent, completely or incompletely
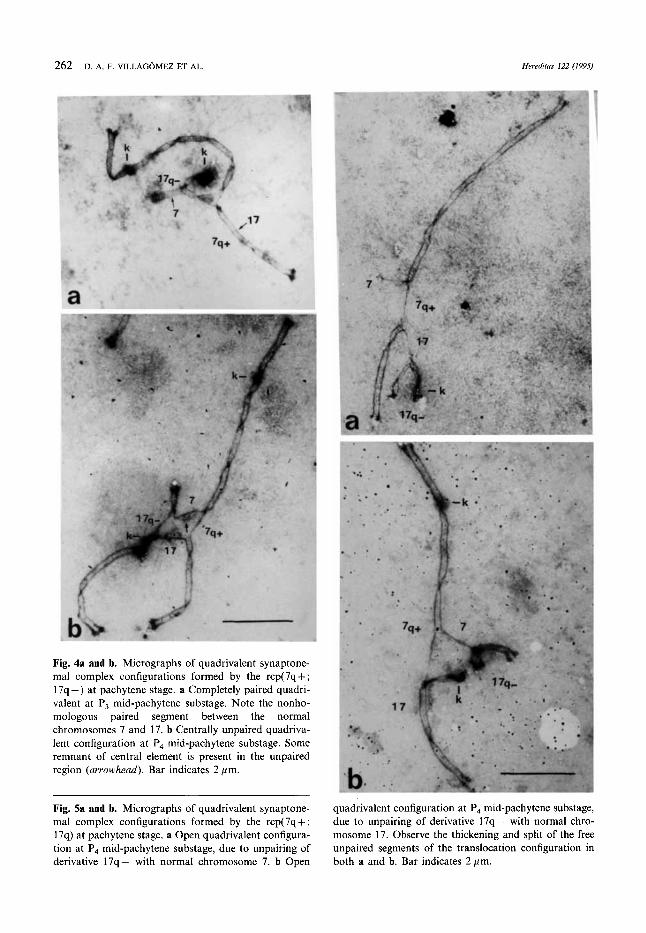
paired, plus univalent, were found in the pachytene stage (Fig. 3). The axes involved in the het- erologous pairing of the quadrivalents were invari- ably those of the normal chromosomes 7 and 17 (Fig. 4a). In 2 of these 10 cells the quadrivalent was associated end-to-end with the sex bivalent. Seven of the centrally unpaired quadrivalents (5 P, and 2 P4) demonstrated remnants of central ele- ments in the unpaired segments (Fig. 4b). This type of quadrivalent was associated end-to-end
260 D. A. F. VILLAG6MEZ ET AL.
*
Hereditas 122 (199s)
s P 9 7q +
m: 1 7
Fig. 2. GTG-banded translocation chromosomes and schematic drawings demonstrating the breakage/rejoin- ing points of the translocation. Bar indicates 10 pm.
with the sex bivalent in one cell, and with an autosomal bivalent, in two cells (Fig. 4b). Open quadrivalent (Fig. 5a-b) was a result of dissocia- tion of derivative 17 from either chromosome 7
Fig. 3. Schematical drawings of the translocation configurations observed. Their distribution on cell stage is given in Table 1.
(7 cells) or chromosome 17 (16 cells). In the latter type of quadrivalent, most of chromosome 17 was paired with derivative 7 (Fig. 5b). The free un- paired segments were thickened and often split into at least two sub-lateral elements.
Cells with completely paired trivalent plus univa- lent (Fig. 6a) demonstrated in one cell the sex bivalent associated end-to-end with a completely paired trivalent. In another cell the sex bivalent was associated with a univalent. Open trivalent (Fig. 6b) was associated in one cell end-to-end with the sex bivalent. The univalent was thickened and split when unpaired.
Conventional meiotic studies
Twenty-three cells at diakinesis-MI were analysed. In 4 cells a ring quadrivalent configuration was found as a result of a chiasma in all arms of the quadrivalent. In 13 cells, a chain quadrivalent
Hereditas 122 (1995) TRANSLOCATION I N A BOAR PRODUCING MALFORMED PIGLETS 261
Table 1. Distribution of spermatocytes according to stage and translocation configuration of the rcp(7;17)(q26;qI 1) (Letter within parenthesis indicates the translocation configuration schematically given in Fig. 3)
Cell No. of Quadrivalent morphology Trivalent plus univalent morphology stage cells
Zygotene
Early Pachytene
(A) ( B) ( C ) ( D) ( E) ( F) ( G) (H)
z3 7 0 0 1 2 0 0 I 3*
PI 2 0 0 0 2 1 0 0 0 p2 2 1 0 0 0 0 0 0 0
p3 30 0 3 7 14 0 3 2 I p4 16 0 4 5 4 I 2 0 0
p5 6 0 2 1 3 0 0 0 0 P6 1 0 1 0 0 0 0 0 0
Totdl 64 1 10 14 25 2 5 3 4
Mid-Pachytene
Late Pachytene
* In one cell the univalent could not be identified A = Completely homologously paired B = Completely paired including heterosynapsis C = Centrally unpaired D = Open E = Completely paired trivalent and self-paired univalent F = Completely paired trivalent and end-to-end paired univalent G = Open trivalent and end-to-end paired univalent H = Open trivalent and rod-shaped univalent
configuration was observed, which was presumably due to failure of chiasma formation at the proxi- mal segment of chromosome pair 17. A chain trivalent configuration plus a univalent (derivative 17) was observed in 6 cells.
Testicular histology of the boar
The histological investigations of the testes demon- strated a normal picture.
Karyotypes of offspring dying in early extra-uterine life
Altogether 5 piglets (4 males, 1 female) from two litters (litter size 5 and 6, respectively) were investi- gated cytogenetically. Four of them (3 males, 1 female) demonstrated 37,XX or XY, - 7, - 17, + der( 7),rcp( 7; 17)(q26;qll) (Fig. 7), and 1 (male) 39,XY,+der( 17), rcp(7;17)(q26;qll) (Fig. 8).
Karyotypes of offspring surviving early extra-uterine lije
Of 13 piglets (7 males, 6 females) surviving the early period of life (age 27-50 days), 5 demon-
strated the normal karyotype, 7 the balanced translocation karyotype, and 1 the unbalanced karyotype 39, XY,+der( 17),rcp(7;17)(q26;qll). The cytogenetics of the latter boar is more com- pletely described in another paper ( VILLAGOMEZ et al. 1995).
Pathology
Piglets with the karyotype 37,XX or X Y - 7,- 17, + der(7), rcp(7; I 7)(q26;q 1 1)
The piglets had body weights ranging between 949 and 1607 g (average 1170 8). X-ray invest- igation excluded the existence of skeletal mal- formations. One male had an apparently nor- mal appearance but the remaining 3 all had a ventricular septa1 defect (Fig. 9a), varying in di- ameter ( 1 mm, 3-4mm and. 3-4mm, respec- tively). Two of these also had a cleft palate (Fig. 9b). Furthermore, one of the latter was very oedematous, having a dilated renal pelvis, ure- thral atresia bilaterally, persisting (23 cm) ureter from the renal hilus (hydronephros), and diluted uracus.
262 D. A. F. VILLAG~MEZ ET AL Heredifas 122 (1995)
Fig. 4a and b. Micrographs of quadrivalent synaptone- ma1 complex configurations formed by the rcp(7q+; 17q -) at pachytene stage. a Completely paired quadri- valent at P, mid-pachytene substage. Note the nonho- mologous paired segment between the normal chromosomes 7 and 17. b Centrally unpaired quadriva- lent configuration at P4 mid-pachytene substage. Some remnant of central element is present in the unpaired region (arrowhead). Bar indicates 2 pm.
Fig. 5a and b. Micrographs of quadrivalent synaptone- ma1 complex configurations formed by the rcp(7q+; 17q) at pachytene stage. a Open quadrivalent configura- tion at P, mid-pachytene substage, due to unpairing of derivative 17q- with normal chromosome 7. b Open
quadrivalent configuration at P4 mid-pachytene substage, due to unpairing of derivative 17q- with normal chro- mosome 17. Observe the thickening and split of the free unpaired segments of the translocation configuration in both a and b. Bar indicates 2 pm.
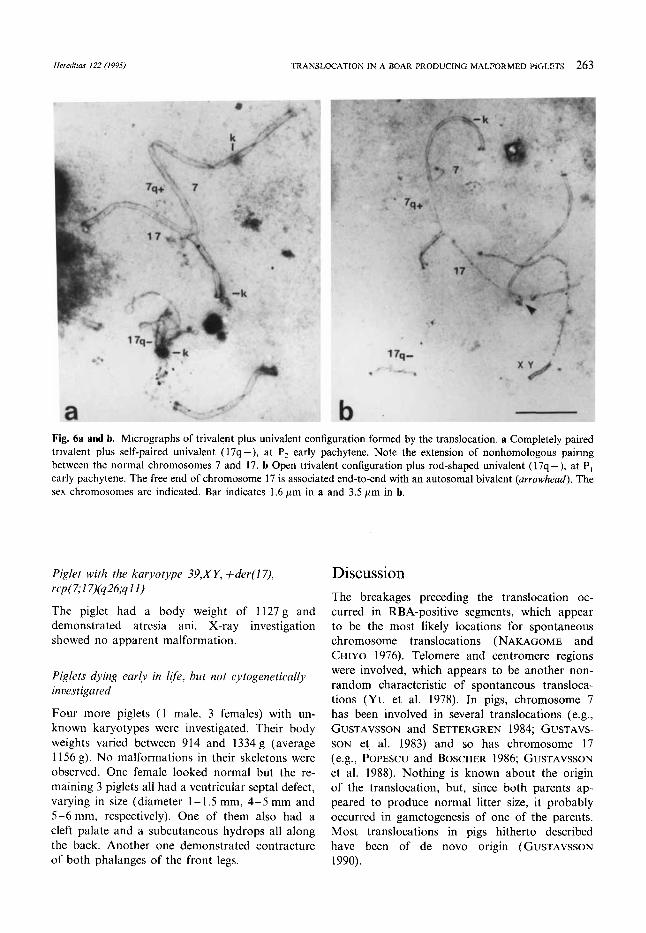
Heredtias I22 (1995) TRANSLOCATION IN A BOAR PRODUCING MALFORMED PIGLETS 263
Fig. 6a and b. Micrographs of trivalent plus univalent configuration formed by the translocation. a Completely paired trivalent plus self-paired univalent (17q -), at P, early pachytene. Note the extension of nonhomologous pairing between the normal chromosomes 7 and 17. b Open trivalent configuration plus rod-shaped univalent (17q-), at P, early pachytene. The free end of chromosome 17 is associated end-to-end with an autosomal bivalent (arrowhead). The sex chromosomes are indicated. Bar indicates 1.6 pm in a and 3.5 pm in b.
Piglet with the karyotype 39,XY, +der(l7), rcp(7; l7)(q26;q I I )
The piglet had a body weight of 1127g and demonstrated atresia ani. X-ray investigation showed no apparent malformation.
Piglets dying early in life, but not cytogenetically investigated
Four more piglets ( 1 male, 3 females) with un- known karyotypes were investigated. Their body weights varied between 914 and 1334g (average 1156 8). N o malformations in their skeletons were observed. One female looked normal but the re- maining 3 piglets all had a ventricular septa1 defect, varying in size (diameter 1 - 1.5 mm, 4-5 mm and 5-6mm, respectively). One of them also had a cleft palate and a subcutaneous hydrops all along the back. Another one demonstrated contracture of both phalanges of the front legs.
Discussion The breakages preceding the translocation oc- curred in RBA-positive segments, which appear to be the most likely locations for spontaneous chromosome translocations ( NAKAGOME and CHIYO 1976). Telomere and centromere regions were involved, which appears to be another non- random characteristic of spontaneous transloca- tions (Yu et al. 1978). In pigs, chromosome 7 has been involved in several translocations (e.g., GUSTAVSSON and SETTERGREN 1984; GUSTAVS- SON et al. 1983) and so has chromosome 17 (e.g., POPESCU and BOSCHER 1986; GUSTAVSSON et al. 1988). Nothing is known about the origin of the translocation, but, since both parents ap- peared to produce normal litter size, it probably occurred in gametogenesis of one of the parents. Most translocations in pigs hitherto described have been of de novo origin (GUSTAVSSON 1990).
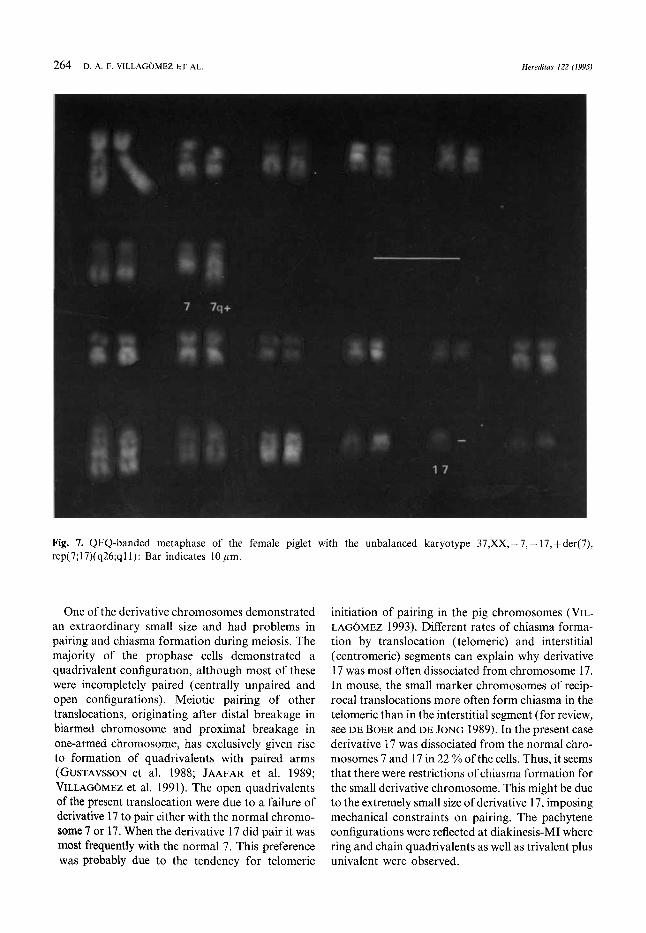
264 I). A. F. VILLAGOMEZ ET AL Hereditas 122 (1995)
Fig. 7. QFQ-banded metaphase of the female piglet with the unbalanced karyotype 37,XX, -7,- 17, +der(7), rcp(7;17)(q26;qlI): Bar indicates 10 pm
One of the derivative chromosomes demonstrated an extraordinary small size and had problems in pairing and chiasma formation during meiosis. The majority of the prophase cells demonstrated a quadrivalent configuration, although most of these were incompletely paired (centrally unpaired and open configurations). Meiotic pairing of other translocations, originating after distal breakage in biarmed chromosome and proximal breakage in one-armed chromosome, has exclusively given rise to formation of quadrivalents with paired arms (GIJSTAVSSON et al. 1988; JAAFAR et al. 1989; VILLAG~MEZ et al. 1991). The open quadrivalents of the present translocation were due to a failure of derivative 17 to pair either with the normal chromo- some 7 or 17. When the derivative 17 did pair it was most frequently with the normal 7. This preference was probably due to the tendency for telomeric
initiation of pairing in the pig chromosomes (VIL- LAGOMEZ 1993). Different rates of chiasma forma- tion by translocation (telomeric) and interstitial (centromeric) segments can explain why derivative 17 was most often dissociated from chromosome 17. In mouse, the small marker chromosomes of recip- rocal translocations more often form chiasma in the telomeric than in the interstitial segment (for review, see DE BOER and DE JONG 1989). In the present case derivative 17 was dissociated from the normal chro- mosomes 7 and 17 in 22 % ofthe cells. Thus, it seems that there were restrictions of chiasma formation for the small derivative chromosome. This might be due to the extremely small size of derivative 17, imposing mechanical constraints on pairing. The pachytene configurations were reflected at diakinesis-MI where ring and chain quadrivalents as well as trivalent plus univalent were observed.

Hereditas 122 (1995) TRANSLOCATION IN A BOAR PRODUCING MALFORMED PIGLETS 265
Fig. 8. QFQ-banded metaphase of a male piglet having the unbalanced karyotype 39,XY, +der( 17),rcp(7;17) (q26;qll). Bar indicates 10 pm.
it was interesting to note that the free unpaired derivative 17 was thickened and more intensely stained than were paired segments. Thickening seems to be a general feature of asynapsed chro- mosomal segments (e.g., SPEED 1984; HULTEN et al. 1987). Therefore we would like to argue that absence of thickening in the centrally unpaired configurations, which sometimes showed remnants of central elements, represent previously com- pletely paired quadrivalents. However, it is possi- ble that centrally asynapsed segments differentiate less than those located terminally. In favour of the former argument, centrally unpaired quadrivalents occurred exclusively at mid and late pachytene.
The pairing and chiasma formation problems of the small derivative chromosome observable in sev- eral stages of meiosis apparently caused the occur- rence of the large number of chromosomally unbalanced piglets. Autosomally unbalanced pigs are still only occasionally described in the litera- ture. When they do occur it is most often in a
mosaic condition. Two females, one pure trisomic and the other mosaic for an undefined centric fragment, were sired by two boars with the chro- mosome constitutions 37,XY - 18/38, XY/39,XY + 18 and 37,XY - 18/38,XY, respectively (VOGT et al. 1974). Recently a rcp(14;15)(q29;q24) was found to give an increased rate of stillborn piglets due to the production of chromosomally unbal- anced stillborns ( GUSTAVSSON and J ~ N S S O N 1992). By patho-anatomical investigation, those piglets demonstrated the same malformations - cleft palate and ventricular septal defect - as ob- served in the present work. Malformations are generally rare in pigs (e.g., SELBY et al. 1971) and it is therefore quite conceivable that the cleft palate and ventricular septal defect observed were associ- ated with the chromosomally unbalanced kary- otype. The same malformations are common in humans with different types of unbalanced chro- mosomal aberrations ( SCHINZEL 1984). Remaining malformations, including atresia ani, occurred in
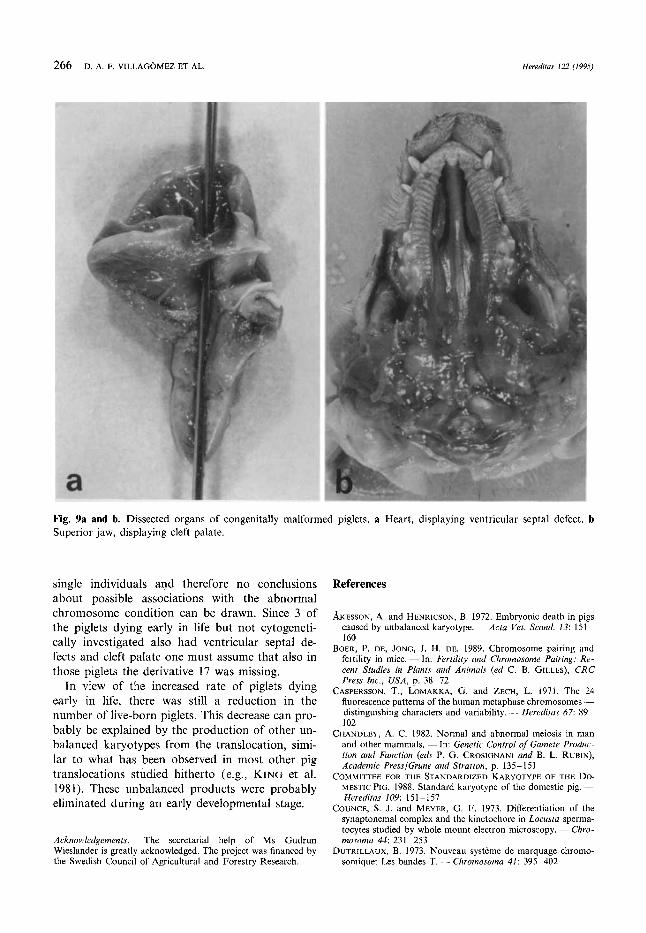
266 D. A. F. VILLAG~MEZ ET AL. Hereditas 122 (1995)
Fig. 9a and b. Dissected organs of congenitally malformed piglets. a Heart, displaying ventricular septal defect. b Superior jaw, displaying cleft palate.
single individuals and therefore no conclusions about possible associations with the abnormal chromosome condition can be drawn. Since 3 of the piglets dying early in life but not cytogeneti- cally investigated also had ventricular septal de- fects and cleft palate one must assume that also in those piglets the derivative 17 was missing.
In view of the increased rate of piglets dying early in life, there was still a reduction in the number of live-born piglets. This decrease can pro- bably be explained by the production of other un- balanced karyotypes from the translocation, simi- lar to what has been observed in most other pig translocations studied hitherto (e.g., KING et al. 1981). These unbalanced products were probably eliminated during an early developmental stage.
Acknowledgements. ~ The secretarial help of Ms Gudrun Wieslander is greatly acknowledged. The project was financed by the Swedish Council of Agricultural and Forestry Research.
References
AKESSON, A. and HENRICSON, B. 1972. Embryonic death in pigs caused by unbalanced karyotype. - Acfa Vet. Scand. 13: 151 - 160
BOER, P. DE, JONG, J. H. DE. 1989. Chromosome pairing and fertility in mice. ~ In: Fertility and Chromosome Pairing: Re- cent Studies in Plants and Animals (ed C. B. GILLES), C R C Press fnc., USA, p. 38-72
CASPERSSON, T., LOMAKKA, G. and ZECH, L. 1971. The 24 fluorescence patterns of the human metaphase chromosomes distinguishing characters and variability. -- Hereditas 67: 89- 102
CHANDLEY, A. C. 1982. Normal and abnormal meiosis in man and other mammals. - In: Genetic Control of Gamete Produc- tion and Function (eds P. G. CROSIGNANI and B. L. RUBIN), Academic PresslGrune and Stratton, p. 135- 151
COMMITTEE FOR THE STANDARDIZED KARYOTYPE OF THE Do- MESTIC PIG. 1988. Standard karyotype of the domestic pig. ~
Hereditas 109: 151-157 COUNCE, S. J. and MEYER, G. F. 1973. Differentiation of the
synaptonemal complex and the kinetochore in Locusta sperma- tocytes studied by whole mount electron microscopy. ~ Chro- mosoma 44: 231-253
DUTRILLAUX, B. 1973. Nouveau systeme de marquage chromo- soniique: Les bandes T. -- Chromosoma 41: 395-402

Hereditus 122 (1995) TRANSLOCATION I N A BOAR PRODUCING MALFORMED PIGLETS 261
DUTRILLAUX, B., LAURENT, C. , COUTURIER J . and LEIRUNE, J . 1973. Coloration des chromosomes humains par I’acridine or- ange apres traitement par le 5 bromodeoxyuridine. ~ C.R. Acad. Sci. (Paris) 276: 3179-3181
FOREJT, J . 1982. X Y involvement in male sterility caused by autosome translocations -- a hypothesis. ~ In: Genetic Control of Gamete Production and Function (eds P. G. CROSIGNANI and B. L. RUBIN), Academic PresslGrune and Stratton, p. 135-151
GARRIEL-ROBEZ, O., JAAFAR, H., RATOMPONIRINA, C . , BOSCHER, J., BONNEAU, J . , POPESCU, C . P. and RUMPLRR, Y. 1988. Heterosynapsis in a heterozygous fertile boar carrier of a 3;7 translocation. ~ Chromosoma 9 7 26-32
GUSTAVSSON, 1. 1990. Chromosomes in the pig. - In: Domestic Animal Cytogenetics. Advances in Veterinary Science and Com- parative Medicine, vol. 34 (ed R. A. MCFEELY), Academic Press Inc., USA, p. 73-107
GUSTAVSSON, I . and JONSSON, L. 1992. Stillborns, partially monosomic and partially trisomic, in the offspring of a boar carrying a translocation: rcp( 14;15)(q29;q24). - Hereditas 1 1 7 31-37
GUSTAVSSON, I. and SETTERGREN, I. 1984. Reciprocal chromo- some translocation with transfer of centromeric heterochro- matin in the domestic pig karyotype. - Hereditas 100: 1-5
GUSTAVSSON, I., SETTERGREN, I. and KING, A. 1983. Occurrence of two different reciprocal translocations in the same litter of domestic pigs. Hereditas 99: 257-267
GUSTAVSSON, I., S W I T O ~ ~ S K I , M., LARSSON, K., PLOEN, L. and HOJER, K. 1988. Chromosome banding studies and synaptone- inal complex analysis of four reciprocal translocations in the domestic pig. - - Hereditas 109: 169-184
H~JLTBN, M., SAADALLAH, N . and BATANIAN, J . 1987. Meiotic chromosome pairing in the human male: experience from sur- face spread synaptonemal complexes. ~ In: Chromosomes To- day, vol. 9 (eds A. STAHL, J. M. LUCIANI and A. M. VAGNER- CAPODANO), p. 218-229
JAAFAR, H., GABRIEL-ROBEZ, O., RATOMPONIRINA, C., BOSCHER, J., BONNRAU, J . , POPESCU, C. P. and RUMPLER, Y. 1989. Analysis of synaptonemal complexes in two fertile het- erozygous boars, both carriers of a reciprocal translocation involving an acrocentric chromosome. -~ Cytogmet . Cell Genet. SO: 20 225
KING, W. A. 1981. Meiotic behaviour of a rcp(13q-;14q+) translocation in heterozygous pigs. ~ Hereditas 94: 235-240
KING, W. A,, GUSTAVSSON, I . , POPESCU, C . P. and LINARES, T. 1981. Gametic products transmitted by rep( 134-;14q+) translocation heterozygous pigs, and resulting embryonic loss. ~ Hereditas 95 239%246
NAKAGOME, Y. and CHIVO, H. 1976. Nonrandom distribution of exchange points in patients with structural rearrange- ments. -- Am. J . Hum. Genet. 28: 31 -41
PATHAK, S., Hsu, T. C. and MARKVONG, A. 1976. Pachytene mapping of the male Chinese hamster. ~~ Cytogenet. Cell Genet. 1 7 1-8
POPESCU, C . P. and BOSCHER, J . 1986. A new reciprocal translocation in a hypoprolific boar. ~ GPn4t. Sd. Evol. 18: 123-130
SCHINZEL, A. 1984. Catalogue of Unbalanced Chromosome Aberrations in Man. de Gupter, Berlin, Ncw York
SEABRIGHT, M. 1971. A rapid banding technique for human chromosomes. - Lancet (2): 971 -972
SRLHY, L. A,, MARIENFHLI), C . J . , HEIDLAGE, W., WRIGHT, H. T. and YOUNG, V. E. 1971. Evaluation of a method to estimate the prevalence of congenital malformations in swine, using a mailed questionnaire. ~ Cornell Vet. 41: 203-213
SPEED, R. M. 1984. Meiotic configurations in female trisomy 21 foetuses. - Hum. Genel. 66: 176- 180
SUMNER, A. T. 1972. A simple technique for demonstrating centromeric heterochromatin. Exp. CeN Res. 75: 304 306
VILLAGOMEZ, D. A. F. 1993. Zygotene-pachytene substaging and synaptonemal complex karyotyping of boar spermato- cytes. ~ Hereditas 118: 87-99
VILLAGOMEZ, D. A. F., GUSTAVSSON, I. and PLOEN, L. 1991. Synaptonemal complex analysis of reciprocal chromosome translocations in the domestic pig. ~ GtnPt. SPl. Evol. 23: 217s-221s
Synaptonemal complex analysis in a boar with tertiary trisomy, product of a rcp(7;17)(q26;qlI) translocation. ~
Hereditas 122: 269-277 VOGT, D. W., ARAKAKI, D. T. and BROOKS, C. C. 1974. Re-
duced litter size associated with aneuploid cell lines in a pair of full-brother Duroc boars. ~ Am. J . Vet. Res. 3 5 1 1 27 I I30
Yu, C. W., BORGAONKAR, D. S. and BOLLING, D. R. 1978. Breakpoints in human chromosomes. ~ Hum. Hered. 28: 210- 225
VILLAGOMEZ, D. A. F., GUSTAVSSON, I. and PLOEN, L. 1995.




















